
Abstract
The primary reason for the failure of malaria vector control across endemic regions is the widespread insecticide resistance observed in Anopheles vectors. The most dominant African vectors of malaria parasites are Anopheles gambiae and Anopheles funestus mosquitoes. These species often exhibit divergent behaviours and adaptive changes underscoring the importance of deploying active and effective measures in their control. Unlike An. gambiae, An. funestus mosquitoes are poorly studied in Benin Republic. However, recent reports indicated that An. funestus can adapt and colonize various ecological niches owing to its resistance against insecticides and adaptation to changing breeding habitats. Unfortunately, scientific investigations on the contribution of An. funestus to malaria transmission, their susceptibility to insecticide and resistance mechanism developed are currently insufficient for the design of better control strategies. In an attempt to gather valuable information on An. funestus, the present review examines the progress made on this malaria vector species in Benin Republic and highlights future research perspectives on insecticide resistance profiles and related mechanisms, as well as new potential control strategies against An. funestus. Literature analysis revealed that An. funestus is distributed all over the country, although present in low density compared to other dominant malaria vectors. Interestingly, An. funestus is being found in abundance during the dry seasons, suggesting an adaptation to desiccation. Among the An. funestus group, only An. funestus sensu stricto (s.s.) and Anopheles leesoni were found in the country with An. funestus s.s. being the most abundant species. Furthermore, An. funestus s.s. is the only one species in the group contributing to malaria transmission and have adapted biting times that allow them to bite at dawn. In addition, across the country, An. funestus were found resistant to pyrethroid insecticides used for bed nets impregnation and also resistant to bendiocarb which is currently being introduced in indoor residual spraying formulation in malaria endemic regions. All these findings highlight the challenges faced in controlling this malaria vector. Therefore, advancing the knowledge of vectorial competence of An. funestus, understanding the dynamics of insecticide resistance in this malaria vector, and exploring alternative vector control measures, are critical for sustainable malaria control efforts in Benin Republic.
Similar content being viewed by others

Background
Malaria continues to burden communities living in endemic areas of Africa. In these countries, intense efforts are being made to bring malaria under control to move forward its elimination. In Benin, despite number of efforts to control malaria, the disease remains largely prevalent across the country [1]. Since 2011, the National Malaria Control Programme (NMCP) on behalf of the Ministry of Health, in collaboration with international organizations and partners, has implemented various malaria control strategies to reduce the burden of the disease [2]. Some of the key malaria control interventions in the country include mass distribution of insecticide-treated nets (ITNs) every 3 years, indoor residual spraying (IRS) and improved diagnosis and treatment by the deployment of anti-malarial drug-based interventions [2]. These efforts targeting either the parasites or vectors have resulted in a significant increase in the usage and ownership of impregnated bed nets, with rates reaching up to 85% [1, 3]. As results, this has contributed to a substantial reduction in malaria-induced morbidity and mortality [1]. Health authorities in Benin have also utilized behavioural change communication (BCC) strategies to raise awareness about malaria prevention and control measures, encouraging communities to adopt positive behaviours to protect themselves from malaria [4]. It has been recognized that vector control is the main malaria control measure that has significantly contributed to the decline in malaria burden through the use of ITNs and IRS [5] targeting mosquito vectors.
Among the malaria-transmitting vectors, Anopheles funestus is one of the primary species in sub-Saharan Africa, including Benin [6]. The An. funestus group comprises nine species distributed across sub-Saharan Africa including An. funestus sensu stricto (s.s.), Anopheles vaneedeni, Anopheles leesoni, Anopheles parensis, Anopheles rivulorum, Anopheles fuscivenosus, Anopheles brucei, Anopheles aruni, and Anopheles confusus [7, 8]. Only An. funestus s.s. is known to be involved in malaria transmission [7, 8]. All other species, except for An. rivulorum, appear not to be associated with a human host-seeking tendency. Indeed, some reports have demonstrated An. rivulorum contribution in malaria transmission [9, 10]. It is well known that effective vector control strategies heavily rely on better understanding the abundance dynamics of the malaria-transmitting vectors, such as An. funestus, and the factors influencing their ability to transmit the disease. Despite the significant implication of An. funestus s.s. in malaria transmission, few studies have been conducted in Benin on this malaria vector [11, 12]. The existing studies have primarily focused on investigating only its resistance status to the insecticides frequently used in public health [11, 12]. This highlights the necessity to gather comprehensive information which will significantly contribute to this vector control in Benin. Indeed, several research gaps remain in (i) understanding An. funestus vectorial competence (identification of transmission hotspots, influencing factors, vector-parasite interactions); (ii) advancing insecticide resistance monitoring and management (resistance mechanisms, its spread and dynamics); and (iii) identifying potential alternative vector control strategies (biological control, genetic control, integrated vector management).
So far in Benin, malaria vector control measures are regardless of individual members in Anopheles species. Therefore, controlling specifically An. funestus poses several challenges, such as (i) behavioural change in biting activity (females mosquitoes have a tendency to bite in the evening or before bedtime when people might not be protected by ITNs) [13, 14]; (ii) breeding habitat adaptability (larvae can adapt and thrive in various breeding habitats) [15, 16]; and (iii) insecticide resistance (An. funestus has shown a remarkable ability to develop resistance to commonly used insecticides, such as pyrethroids) [17, 18]. Addressing these challenges requires a multifaceted approach, including the development of new vector control strategies, and continued surveillance for insecticide resistance.
To help establish effective resistance management strategies, it is important to better understand the distribution of An. funestus, its insecticide resistance profiles with underlying mechanisms and its impact on effectiveness of control interventions and malaria transmission [19]. In this review, the primary aim was to comprehensively evaluate the prevailing distribution and vectorial competence of An. funestus in Benin. Additionally, the insecticide resistance patterns exhibited by this mosquito species are analysed and future perspectives for vector control strategies are discussed. This will provide valuable insights that can inform and the elaboration of evidence-based policy and the implementation of more effective vector control interventions in Benin Republic.
Literature search
A literature search was performed using the search terms (Anopheles funestus; An. funestus; Benin Republic; insecticide resistance) with the Boolean operator (AND) as follow: “Anopheles funestus and Benin Republic”, “An. funestus and Benin Republic” and “Insecticide resistance in An. funestus and Benin Republic”. The search was limited to publications written in English and in French, and it was done using the most popular search platform Google, Google Scholar and PubMed databases. In addition, Google and Google Scholar automatically index most information from the academic web. All papers that reported collection of at least one An. funestus (based on morphological identification) and from the year 2007, where insecticide resistance monitoring in An. funestus in Benin Republic started, were considered. Finally, twenty-one (21) published studies reporting work undertaken between 2007 and 2019 in Benin were selected for this review.
Distribution and density of Anopheles funestus populations
Most of the available data on An. funestus in Benin were recorded when conducting a survey on Anopheles gambiae sensu lato (s.l.). Since 2007. There has been a scarcity of entomological research available on An. funestus in Benin (Additional file 1).
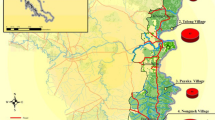
Despite having a relatively lower population density compared to An. gambiae s.l., An. funestus is widely distributed in Benin. It has been found in the four climatic regions of the country, namely North Sudanese, wet Sudanese, Sub-Sudanese and Sub-equatorial as described by Djouaka et al. [20] (Fig. 1). Both North Sudanese and wet Sudanese climatic regions are characterised by a long dry and a short rainy season. Large water bodies are found in North Sudanese climatic region where temperatures are the highest particularly during the dry season [20]. However, the west part of the North Sudanese region is dominated by hills of up to 800 m of altitude and several small water bodies. The vegetation is partially made of wet savanna with the lowest temperature in the country, and rainfall ranging from 1200 to 1300 mm per year [20]. The sub-Sudanese climatic region covers the center of the country and part of the South. This region is less hilly and the vegetation is of wet savanna. Annual rainfall is between 900 and 1200 mm [15].

Distribution of An. funestus sites in Benin
The sub-Equatorial climatic region covers the country’s southern part and extends up to the coastal areas. The relative humidity is high and can up to 95%, temperatures are relatively low ranging from 25 to 31 °C, and the vegetation is a mosaic of coastal wetlands, forests, and savannah [15].
Anopheles funestus was found in sympatry with An. gambiae and more often in the country’s western part and Savannah regions, as shown in Fig. 1. The relatively high occurrence of this species in the western regions of Benin could be attributed to the relatively low temperatures, humidity associated with the hilly landscape, and the presence of streams covered with vegetation [21].
In the eastern part, very little population of An. funestus was found, certainly due to the low rainfall and high dryness. The low occurrence of An. funestus population in this part of the country may also be attributed to the sampling period or the presence of a few permanent freshwater bodies covered with vegetation [22]. However, an extensive An. funestus surveys are required in this part of the country to confirm the observed lower occurrence.
During dry seasons within the study areas, An. funestus was found more abundant with a peak density during the dry-to-rainy transition period [20, 23]. This is probably due to their larval breeding habitats’ stability and adaptation to desiccation [24]. Moiroux and colleagues [25] have reported that in Ouidah-Kpomasse-Tori Bossito district located in Southern Benin, An. funestus becomes the predominant Anopheles mosquitoes during the dry-hot season. They get more aggressive during that period with a high density around localities bordering large water bodies such as the Toho Lake. Surveys carried out in Kpome (southern Benin) and Tanongou (northern Benin) showed a relatively high density of An. funestus (up to 3 mosquitoes/room) during the dry-to-rainy transition period [20]. High density of An. funestus was reported in other southern villages with 0.29 to 13.48 mosquitoes per village [6, 12]. The presence of An. funestus was also reported in others localities across most of departments in Benin, but at low density [23, 26,27,28,29,30,31,32]. Nevertheless, the increased An. funestus density in localities during the dry season contributes to the residual malaria transmission in Benin [20]. Indeed, during the dry season, mosquitoes’ aggressivity, especially malaria vectors, becomes very low [25]. In regards to this situation, vector control tools, such as ITNs are used less by inhabitants living in endemic regions and could impede the achievement of the World Health Organization (WHO) ultimate goal, which was to eliminate malaria in some countries by 2030 [33].
Furthermore, in Lokohoue and Tokoli villages located in OKT district (sub-Equatorial climatic regions), a human could receive 2.1 to 18.73 bites of An. funestus per night [34]. However, An. funestus was found to be relatively less aggressive in both rainy and dry seasons with 0.35 b/h/n (bite/human/night) and 0.84 b/h/n respectively in Copargo in wet-Sudanese climatic zone [23]. In Kérou and Djougou districts (wet-Sudanese climatic zone), a human could receive 0.66 and 6.55 bites of An. funestus mosquitoes per night, respectively [35, 36]. The lowest aggressivity observed in these regions may be due to the sampling period, rainy seasons (not much suitable for An. funestus development) and the rarity of stable larval habitats during the same period. Considering the information above and for a sustainable malaria control, it becomes important to redeploy vector control tools in the dry season because this season seems not to be a target period to the NMCPs for implementing these strategies.
Contribution of Anopheles funestus to malaria transmission
Most members of An. funestus group are zoophilic (preference to take blood meals on animals), except An. funestus s.s. [37], which is a the primary vector for Plasmodium species [11, 20, 37,38,39,40,41]. Although, few studies have investigated the distribution of An. funestus members [11], PCR-based species identification of wild-caught An. funestus in various localities across Benin revealed two main sibling species, including An. funestus s.s. (the most abundant; up to 99%) and An. leesoni (less than 10%) [6, 20]. These two sibling species were mainly caught indoors confirming the endophilic (tendance to inhabit/rest indoors) behaviour of An. funestus circulating in Benin [40]. Anopheles funestus, when exhibiting indoor biting behaviour, increases the likelihood of direct contact with humans, typically during the night when they are not yet under the bed nets. This increases the risk of transmitting the malaria parasite. Furthermore, An. funestus may exhibit seasonal variations in biting behaviour, with an increased tendency to bite indoors during certain periods [13]. Such information can help to implement suitable vector control strategies during the times when transmission risk is high. Indoor biting behaviour of An. funestus should, therefore, be taking into account when developing novel strategies to control malaria vectors.
Plasmodium infection rate has been reported at a very high level in An. funestus s.s. in Benin [11, 20]. This rate ranged from 2.64 to 15.78% in mosquitoes from Pahou and Gakpé near the Toho Lake in OKT district in coastal area of Benin (Additional file 1). Also, a high Plasmodium infection rate has been reported in the southern inland region of Benin at Toffo and Lokossa with values of 13.33 to 18.51%, respectively [20].
Overall, high Plasmodium infection rates of An. funestus s.s. observed in some villages in the West part of Benin and its related high anthropophily (preference to take blood meals on human), confirm the role of this vector species in malaria transmission, exceeding in some cases, An. gambiae s.l. (Additional file 1). A relatively high Plasmodium infection rate of An. funestus s.s. collected indoors during the dry season was reported in Toffo district, where this specie competes with Anopheles coluzzii, one of the primary malaria vectors in southern Benin [11]. The infection rates recorded in both An. funestus s.s. and An. coluzzii during the same period may explain the high malaria transmission and incidence during the dry seasons in Toffo [11]. The same trend was observed in Copargo in the Donga department, where a high infection rate was also reported in An. funestus, despite the high abundance of An. gambiae s.l. [23]. The former (26.08%) exhibited three times higher sporozoite rate than the later (8.49%) [23]. In Tanongou neighborhood located in the Atacora department, An. funestus s.s. was also relatively highly infected with Plasmodium (5.62%) [20] (Additional file 1).
Considered as a main indicator to estimate the overall contribution of Anopheles mosquitoes to malaria transmission, the Entomological Inoculation Rate (EIR) is the product of sporozoite rate and human biting rate over a defined time and space [42, 43]. The EIR of both An. gambiae s.l. and An. funestus s.s. varied significantly according to the season. In fact, during the rainy season, 75 infective bites per person were attributed to An. gambiae s.l. while An. funestus s.s. ensured only 18.75 infective bites per human. In contrast, during the dry season, An. funestus s.s. accounted for 37.5 infective bites per human while An. gambiae s.s. resulted in 28.12 infective bites per human in Northern Benin [23]. A relatively similar infective bites per human per 100 nights has been reported in An. funestus s.s. (0.67 infective bites) and An. gambiae s.l. (1.38 infective bites) in southern Benin [6].
The ability of An. leesoni to transmit Plasmodium remains unexplored in Benin. None of the studies included in this review has described the presence of Plasmodium species in An. leesoni, suggesting that it may not be involved yet in malaria transmission in Benin [6, 20, 40] and highlighting that it has no public health significance in Benin [20]. In addition, An. leesoni has demonstrated a very high zoophilic behavior and has been found in sympatry with An. funestus s.s. in Tanguiéta, northwestern Benin [20]. Only one An. leesoni was found in the coastal area at Tokoli-Vidjinangnimon village in OKT district [6]. However, recent studies in Cameroon reported the presence of An. leesoni infected with Plasmodium spp. [38, 39] suggesting that, extended studies on An. funestus group in Benin might provide more valuable information regarding the implication of An. leesoni in malaria transmission in the country.
Other members of An. funestus group have not been found in Benin. However, certain members of this group have been reported to relatively contribute to malaria transmission in Nigeria and Tanzania [10, 40, 44]. Although, An. rivulorum was found with an anthropophilic rate (an important factor in vectorial capacity) of 40% in the southern region of Nigeria, its implication in malaria transmission in this country was not yet elucidated [40]. Further, specimen of An. rivulorum was positive for P. falciparum found in Tanzania [10, 44]. Anopheles vaneedeni, which can easily harbor Plasmodium parasites under laboratory conditions, is either exophilic or anthrophilic [9]. Another member of An. funestus group, An. parensis, was reported not susceptible to malaria parasite infection [40, 45, 46]. However, recent study reported 1.6% of Plasmodium infection rate in An. parensis in Mozambique [47].
Besides, the contribution of An. funestus to malaria transmission in the urban environment remains scanty in Benin while such studies in other African countries have shown the implication of An. funestus in malaria parasite transmission in the city of Yaoundé in Cameroon [38]. This instigates to further investigate the eco-epidemiological characteristics and behavioral traits of An. funestus mosquito in urban settings, in order to map malaria risk and burden and to improve current vector control strategies.
Insecticide susceptibility profile
Until 2010, no data on insecticide susceptibility profile and mechanisms of insecticide resistance of An. funestus were published in Benin. From 2011, characterization of insecticide resistance in An. funestus populations have been reported [12]. Four class of insecticides were monitored: 4% DDT and 4% dieldrin (Organochlorines), 0.75% permethrin and 0.05% deltamethrin (Pyrethroids), 0.1% Bendiocarb (Carbamate) and 5% malathion (Organophosphate). An. funestus susceptibility to insecticides was assessed in only 5 locations across the country [11, 12, 20, 34, 48] (Additional file 2), indicating that there is a need to further investigate resistance status in this malaria vector to better characterize areas where high resistance levels prevail nationwide.
Pyrethroid resistance in this An. funestus seems to be spread across the country as its counterpart An. gambiae s.l. In addition, An. funestus collected in 2007 and 2008 was found to be fully susceptible to a diagnostic dose of deltamethrin in Tokoli and Lokohoué (Additional file 2) according to Moiroux et al. [34]. In contrast, An. funestus collected between 2009 and 2011 in Pahou near Tokoli and Lokohoué villages, were resistant to diagnostic doses of deltamethrin and permethrin with mortality rates of 66.4% and 88.8%, respectively [12]. Elsewhere, high resistance has been reported in An. funestus collected in 2014 in Kpome village, with mortalities rates of 13% and 46.5% for permethrin and deltamethrin respectively [11]. In addition, even when An. funestus from Kpome was exposed for 90 min, it was still resistant to diagnostic dose of permethrin (51.62% mortality) [11]. This constitutes a serious threat for the effectiveness of pyrethroid-based interventions.
In 2017, the same trend was observed in An. funestus from Kpome where resistance to diagnostic dose of permethrin and deltamethrin was recorded with mortality rates of 14.84% and 44.15%, respectively [48]. This resistance profiles suggest an increase in the overall level of pyrethroid resistance in southern Benin within 6 years (from 2011 to 2017) and might be attributed to the increased ITNs coverage across Benin and additional selection factors such as pesticides use in agriculture [49, 50].
Similarly, permethrin resistance was also observed in 2014 in Doukonta, located in Lokossa district in southern Benin with a mortality rate of 11% [20]. The presence of resistant An. funestus in inland areas of Kpome and Doukonta compared to coastal localities (e.g. Pahou) [20] may be associated with gene flow among An. funestus in south of Benin [11].
A comparative analysis of An. funestus collected from Doukonta and Tanongou located respectively in southern and northern transect of Benin, reveals contrasting resistance patterns in An. funestus, marked by a full susceptibility to permethrin in Tanongou compared to a resistance in Doukonta (Additional file 2).
In addition to the resistance to pyrethroids, An. funestus from Benin are also resistant to bendiocarb [11, 12]. This raises a concern for NMCPs because bendiocarb is currently being introduced in IRS formulation for malaria vector control in West African countries [51]. Indeed, bendiocarb resistance was observed in An. gambiae in Benin 3 years after the implementation of IRS [51] and such resistance may likely spread to An. funestus owing to its endophilic behavior.
A relative resistance to dieldrin was recorded in An. funestus mosquitoes from Pahou (mortality rate of 93%) while a susceptible status was observed in those from Kpome (mortality rate of 98.9%) [11, 12]. However, Both of them were resistant to DDT [11, 12, 20]. The recorded mortality after standard bioassay exposure ranged from 0 to 9.1% (Additional file 2). Meanwhile, a relative resistance was observed for DDT (mortality rate of 90 ± 3.18%) in Tanongou in northern Benin [20].
All An. funestus mosquitoes from southern Benin remain susceptible to malathion [11, 12]. Nevertheless, the eco-toxicity of this insecticide limits its adoption by national malaria vector control programmes. Overall, the multiple resistance in An. funestus species underline the complexity of malaria control in Benin.
Although, An. funestus became largely distributed in Benin and resistant to pyrethroid insecticides, it still remains less studied. Regular monitoring of insecticide susceptibility profiles of this malaria vector should be implemented to help NMCPs improve strategies made in place to control malaria.
Mechanisms underlying the insecticide resistance phenotypes
A ratio of mosquito individuals can tolerate lethal doses of insecticides in a normal population of the same species through different mechanisms, such as (i) metabolic resistance: insecticide can be broken down or metabolized by detoxification enzymes much faster in the resistant mosquitoes than in the susceptible ones, thereby quickly eliminated from the mosquito organism; (ii) target-site resistance: the insecticide target-site can be modified due to the presence of mutation that prevent the insecticide from binding thereby reducing lethal effect of such insecticide; (iii) penetration resistance: resistant mosquitoes may limit penetration of the insecticides than susceptible insects, or (iv) behavioural resistance (the less studied): mosquitoes avoid the insecticide contact [52]. Characterizing resistance mechanisms is an essential step in insecticide resistance management. This provides baseline data for designing control programmes and evidence-based choice of insecticides.
Basically, biochemical and molecular analysis as well as synergistic tests are usually used to determine insecticide resistance mechanisms in Anopheles vectors exposed to insecticides. These analysis and synergistic assays were used to characterize the mechanisms involved in insecticide resistance. Whole genome data has also recently become available for An. funestus studies [53] and advance molecular researches on the insecticide resistance mechanisms developed by this vector was uncovered.
Mechanisms underlying DDT, permethrin and deltamethrin resistance
A synergistic study conducted by Djouaka and collaborators [12], has suggested that P450 genes play very little role in the observed DDT resistance in An. funestus collected from Pahou (Southern Benin). Furthermore, authors showed that glutathione S-transferase (GSTe2) gene was overexpressed in these resistant An. funestus [12, 54]. Indeed, the GSTe2 gene is 44.8 time overexpressed in DDT-resistant mosquitoes when compared to the susceptible strain (FANG) [54]. The GSTe2 gene expression was higher than that of other GSTs genes in the same mosquito specimens indicating that GSTe2 is likely the main detoxification gene associated with DDT resistance in An. funestus mosquitoes collected from Pahou [12, 54]. As a result, further studies are need to explore the genomic pathway conferring this selective advantage for GSTe2 gene.
Further, high frequency (96%) of the L119 F-GSTe2 resistant allele in An. funestus collected from other localities of southern Benin was reported [11, 20]. However, a relatively lower frequency (35%) of this resistant allele was recorded in An. funestus collected from Tanongou (northern part of Benin) [20].
Interestingly a microarray-based genome-wide transcription and qRT-PCR analysis on An. funestus mosquitoes from Kpome (Southern Benin) showed that, overexpression of the GSTe2 gene is responsible for the observed DDT resistance [54, 55]. It was also reported the consistent difference for this gene between the population of southern Benin (Kpome, Pahou and Doukonta) and that of Tanongou (North Benin) [55] indicating that possible barriers to gene flow exist between these populations. This implies that barriers to gene flow likely to impact the design and implementation of resistance management strategies in this country.
A number of studies have revealed that L119F-GSTe2 resistant allele is near fixation in both DDT susceptible and resistant An. funestus mosquito populations from Kpome, Pahou and Doukonta (Southern Benin) [11, 55, 56]. On the other hand, there is a possibility that the L119F-GSTe2 allele could also be selected by pyrethroid-based interventions [11].
Although resistance genes confer the potential of surviving insecticide exposures, they are often associated with pleiotropic effects on various fitness-related traits in An. gambiae mosquitoes (e.g., trophic behaviour, fecundity, fertility, parasite transmission, longevity, and larval survivorship) [57, 58]. Influence of GST-metabolic resistance on vectorial competence in An. funestus should be more investigated.
Resistance to permethrin and deltamethrin in An. funestus across southern Benin was mainly attributed to cytochrome P450 monooxygenases [48, 56]. For instance, a significant over-expression of two duplicated P450s, CYP6P9a (Fold Change: 4.7) and CYP6P9b (Fold Change: 7) which can metabolize both permethrin and deltamethrin [59] was observed in An. funestus from Pahou (Southern Benin) [12]. Overall, there is a need to continue characterizing the insecticide resistance mechanisms in all localities where the presence of An. funestus have been reported in Benin to further capture the spread of this vector resistance and underlying mechanisms. This could help Benin’s NMCP to design and implement more suitable measures for resistance management in malaria vectors.
Mechanisms underlying bendiocarb and dieldrin resistance
Although, An. funestus populations from Pahou and Kpome were resistant to bendiocarb with a mortality rate ranging from 64 to 70%, no evidence of ace-1R resistant allele was recorded in this carbamate-resistant populations [11, 12]. A resistance profile of An. funestus to bendiocarb should be further investigated to provide updated information important in improving current vector control strategies in the country. In contrast, as resistance to dieldrin was recorded in Pahou, with susceptibility to the same insecticides in Kpome were recoded; pyrosequencing and PCR-RFLP analysis performed on mosquito specimens from Pahou, revealed a moderate level (16%) of 296 S-Rdlr mutation, a GABA receptor mutation conferring dieldrin resistance in An. funestus in Africa [60]. Genotyping of the A296S-Rdlr mutation in An. funestus from Kpome revealed 99% homozygous susceptible genotype with 1% heterozygous further confirming susceptibility profile to this insecticide [11]. To date, no other study has been conducted to update dieldrin susceptibility profile in An. funestus.
Perspectives on vector control strategies
A number of entomological, epidemiological and genomic studies on An. gambiae have significantly contributed to strengthen malaria control. The control of the major malaria vectors in endemics area is one of an important ways to interrupt the transmission of this disease [61]. Therefore, phase I (Cone and Tunnel tests) and Phase II (Experimental Hut Trials, EHTs) extend evaluation studies are also needed to appreciate the effectiveness of current and new designed vector control products targeting An. funestus mosquitoes.
In Benin, little is known about An. funestus responses to vector control tools. Bio-efficacy assays throughout standard cone and tunnel tests showed the loss of efficacy of currently used ITNs (PermaNet 2.0 and Olyset Net) against natural population of An. funestus from Kpome [48].
EHTs were also conducted in Kpome where more than 40% and 60% of An. funestus survived in presence of PermaNet 2.0 and Olyset Nets, indicating that these nets were able to provide only 46% and 17% personal protection, respectively [48]. Furthermore, it has been reported that current ITNs were still estimated to provide average ‘true’ personal protection of 80% against An. funestus bites in Lokohoue village in Ouidah in southern Benin [62]. All of these findings suggest that protective effect of currently used nets is compromised in Benin. Surprisingly, it was observed in Lokohoue that An. funestus likely continues to bite at dawn when people are no longer sleeping under mosquito nets [62]. This is a huge concern since current vector control strategies, rely only on the use of pyrethroid-treated nets to target nocturnal, endophilic malaria vectors.
However, the fact that An. funestus is still susceptible to bendiocarb in southern Benin and considering its high endophilic and endophagic behaviors, IRS in combination with current ITNs may be a promising control strategy, especially during the dry season where the density of An. funestus is high. Also, the major role played by metabolic resistance to pyrethroids in An. funestus in Benin suggests that combining the synergist PBO, such as PBO-Pyrethroid bednets, could help manage the pyrethroid resistance in this malaria vector. Interestingly, EHTs showed a benefit to use both PermaNet 3.0 and Olyset Plus (PBO-Based nets) to control resistant population of An. funestus in southern Benin [48]. A combination of the synergist PBO to pyrethroids makes treated-nets more efficient, as PBO is a potent cytochrome P450s enzyme inhibitor. Indeed, PermaNet 3.0 and Olyset Plus, were able to prevent blood feeding in 92% and 100% in resistant An. funestus, respectively [48]. High killing effect was also exhibited by these nets (100% for PermaNet 3.0 and 87% for Olyset Plus) [48]. The same trend have been observed for An. gambiae with 80% of personal protection in presence of PermaNet 3.0 [48].
Therefore, a combined pyrethroids-PBO nets showed a greater efficacy against resistant malaria vector populations and could be a promising strategy against pyrethroid-resistant Anopheles mosquitoes [63,64,65,66,67]. However, its efficacy can be impacted by other resistance mechanisms in mosquito vectors, such as GST-mediated metabolic resistance, which is not affected by PBO synergistic action [68]. Moreover, recent study revealed efficacy-loss of PBO–based nets (Olyset Plus) in highly pyrethroid-resistant An. funestus populations from Mozambique [69] and Cameroon [70]. This could also be occurring in Beninese mosquito populations. In fact, this reduced efficacy of PBO-based tools could be attributed to the overexpression of the cytochrome P450 genes that could allow mosquitoes to tolerate exposure to ITNs impregnated with pyrethroids and PBO in the nets. Efficacy against An. funestus of other new generation nets could also be explored as the case of An. gambiae where an interesting information have been reported in Benin and Ivory coast, highlights that these new generation nets have the potential to improve malaria vector control and provide better community protection against clinical malaria in pyrethroid resistant areas compared to standard pyrethroid-only ITNs [71,72,73].
The new generation nets combining pyrethroid and insect growth regulator (Pyriproxyfen) and a Royal Guard or Interceptor G2 impregnated with Chlorfenapyr (a pyrrole insecticide class), have shown a great efficacy against resistant An. gambiae in West African countries [71,72,73]. More recently, it was reported that Interceptor G2 provided a high lethal effect, blood-feeding inhibition, repellency and personal protection against An. funestus from Tanzania [74]. These new bednets could be a potential insecticide resistance management tool to prevent malaria transmission in areas compromised by the spread of pyrethroid resistance. Additionally, the insect growth factor combined with pyrethroid may also positively impact the reproductive success in An. gambiae [73]. Further investigations on how An. funestus exposure to these new nets can affect the fecundity, fertility and other life history traits, could provide additional key information on fitness effects.
Definitely, more robust entomological studies focusing on An. funestus should be implemented in Benin. Indeed, in addition to existing assays to measure the performance of ITNs which are rely on detection of rapid knockdown and 24-hour mortality, critical insights into the behavior of mosquitoes in response to ITNs can be gained by laboratory and semi-field studies that quantify important traits including net contact time, and blood-feeding behaviour, longevity and reproductive features. A suite of experimental procedures ranging from simple benchtop assays (e.g.Video Cone test, Thumb/Baited Box test) to large-scale video tracking [75, 76] to record the lifetime impact of exposure to an active ingredient in presence of a host could provide robust parameters to better appreciate the effectiveness of the current and new generation vector control tools.
On the other hand, alternative strategies using non-insecticide-based tools in combination with current ITNs against resistant pyrethroid resistant mosquitoes could be explored to improve progress towards malaria elimination. New tool in development uses combination of human-associated stimuli, including olfactory, visual and thermal cues, to lure and kill malaria vectors without insecticides. The strategy could be a promising integrated vector control strategy and have a real public health benefits.
In addition, considering the insecticide resistance phenomenon, novel innovative symbiotic control measure may be also explored [77, 78]. However, before implementing this control measure in natural An. funestus mosquito populations, it will be useful to better understand the bacterial diversity in this vector and their interactions with their hosts. Indeed, one of key factors determining vector competence is the gut microbiota of the mosquito [79]. The microbiota in mosquito midguts plays a crucial role in the development, reproduction, immunity, and vector competence as reviewed in An. gambiae populations [80, 81]. Recently, several research reports showed associations between the mosquito microbiota and resistance to the current insecticides used for vector control [82,83,84]. A good understanding of the role of An. funestus microbiota in insecticide resistance will allow improvement of techniques toward curbing the widespread of insecticide resistance in malaria vector [85].
Conclusion
This review provides synthesized information on the vectorial competence of An. funestus. Anopheles funestus became largely distributed in Benin with high density recorded during the driest period. The prevalence of Plasmodium infection in An. funestus was high with comparable entomological inoculation rate between this species and its counterpart An. gambiae. Furthermore, An. funestus was found to be resistant to permethrin, deltamethrin and bendiocarb, resistant to DDT but remain susceptible to malathion. GSte2 and P450 genes are mainly incriminated in the observed phenotypic resistance, highlighting the urgent need for further actions to strengthen malaria control strategies. Information provided on mechanisms underlying insecticide in An. funestus call for the development of more comprehensive resistance management and the implementation of alternative control interventions.
Availability of data and materials
All data generated or analysed during this study are included in this published article and additional files.
Abbreviations
- DDT:
-
Dichlorodiphenyltrichloroethane
- GABA:
-
Gamma-aminobutyric acid
- GST:
-
Glutathione-S transferase
- IRS:
-
Indoor residual spraying
- ITNs:
-
Insecticide-Treated Nets
- PBO:
-
Piperonyl butoxide
- qPCR:
-
Quantitative polymerase chain reaction
- RFLP:
-
Restriction fragment length polymorphism
- EHT:
-
Experimental hut trials
References
Institut National de la Statistique et de l’Analyse Economique (INSAE). Et ICF. Cinquième Enquête Démographique et de Santé (EDSB-V) 2017–2018. Benin: Cotonou; 2018.
Akogbéto MC, Aïkpon RY, Azondékon R, Padonou GG, Ossè RA, Agossa FR, et al. Six years of experience in entomological surveillance of indoor residual spraying against malaria transmission in Benin: lessons learned, challenges and outlooks. Malar J. 2015;14:242.
Tokponnon FT, Aholoukpe B, Denon EY, Gnanguenon V, Bokossa A, N’Guessan R, et al. Evaluation of the coverage and effective use rate of long-lasting insecticidal nets after nation-wide scale up of their distribution in Benin. Parasit Vectors. 2013;6:265.
USAID/CDC PMI. Accelerating the reduction of malaria morbidity and mortality (ARM3) Benin behavior change communication (BCC). Report. 2018.
Bhatt S, Weiss DJ, Cameron E, Bisanzio D, Mappin B, Dalrymple U, et al. The effect of malaria control on Plasmodium falciparum in Africa between 2000 and 2015. Nature. 2015;526:207–11.
Djènontin A, Bio-Bangana S, Moiroux N, Henry M-C, Bousari O, Chabi J, et al. Culicidae diversity, malaria transmission and insecticide resistance alleles in malaria vectors in Ouidah-Kpomasse-Tori district from Benin (West Africa): a pre-intervention study. Parasit Vectors. 2010;3:83.
Gillies M, Meillon D. The Anophelinae of Africa south of the Sahara (Ethiopian zoogeographical region). Publ South African Inst Med Res. 1968.
Gillies MT, Coetzee M. A supplement to the Anophelinae of the South of the Sahara (Afrotropical Region). Publ S Afr Inst Med Res. 1987;55:1.
De Meillon B, van Eeden GJ, Coetzee L, Coetzee M, Meiswinkel R, du Toit CLN, et al. Observations on a species of the Anopheles funestus subgroup, a suspected exophilic vector of malaria parasites in northeastern Transvaal, South Africa. Mosq News. 1977;37:657–61.
Wilkes TJ, Matola YG, Charlwood JD. Anopheles rivulorum, a vector of human malaria in Africa. Med Vet Entomol. 1996;10:108–10.
Djouaka R, Riveron JM, Yessoufou A, Tchigossou G, Akoton R, Irving H, et al. Multiple insecticide resistance in an infected population of the malaria vector Anopheles funestus in Benin. Parasit Vectors. 2016;9:453.
Djouaka R, Irving H, Tukur Z, Wondji CS. Exploring mechanisms of multiple insecticide resistance in a population of the malaria vector Anopheles funestus in Benin. PLoS ONE. 2011;6:e27760.
Sougoufara S, Diédhiou SM, Doucouré S, Diagne N, Sembène PM, Harry M, et al. Biting by Anopheles funestus in broad daylight after use of long-lasting insecticidal nets: a new challenge to malaria elimination. Malar J. 2014;13:125.
Doucoure S, Thiaw O, Thiaw O, Wotodjo AN, Bouganali C, Diagne N, et al. Anopheles arabiensis and Anopheles funestus biting patterns in Dielmo, an area of low level exposure to malaria vectors. Malar J. 2020;19:230.
Nambunga IH, Ngowo HS, Mapua SA, Hape EE, Msugupakulya BJ, Msaky DS, et al. Aquatic habitats of the malaria vector Anopheles funestus in rural south-eastern Tanzania. Malar J. 2020;19:219.
Debrah I, Afrane YA, Amoah LE, Ochwedo KO, Mukabana WR, Zhong D, et al. Larval ecology and bionomics of Anopheles funestus in highland and lowland sites in western Kenya. PLoS ONE. 2021;16:e0255321.
Nkemngo FN, Mugenzi LMJ, Terence E, Niang A, Wondji MJ, Tchoupo M, et al. Elevated Plasmodium sporozoite infection and multiple insecticide resistance in the principal malaria vectors Anopheles funestus and Anopheles gambiae in a forested locality close to the Yaoundé airport, Cameroon. Wellcome Open Res. 2020;5:146.
Chanda J, Saili K, Phiri F, Stevenson JC, Mwenda M, Chishimba S, et al. Pyrethroid and carbamate resistance in Anopheles funestus Giles along Lake Kariba in southern Zambia. Am J Trop Med Hyg. 2020;103:90–7.
Riveron JM, Tchouakui M, Mugenzi L, Menze BD, Chiang M-C, Wondji CS. Insecticide resistance in malaria vectors: an update at a global scale. In: Maguin S, Dev V, editors. Towards malaria elimination—a leap forward. London: InTech Open; 2018.
Djouaka R, Akoton R, Tchigossou GM, Atoyebi SM, Irving H, Kusimo MO, et al. Mapping the distribution of Anopheles funestus across Benin highlights a sharp contrast of susceptibility to insecticides and infection rate to Plasmodium between southern and northern populations. Wellcome Open Res. 2017;1:28.
Institut national de la statistique et de l’analyse économique (INSAE). Cahier des villages et quartiers de ville du département de l’Atacora (RGPH-4,2013). 2016;38.
INSAE. Cahier des villages et quartiers de ville département du BORGOU direction des Etudes Démographiques Cotonou. 2004.
Aïkpon R, Salako A, Ossè R, Aïkpon G, Sidick A, Sèwadé W, et al. The spread of malaria in Savannah area in Benin: the contribution of Anopheles gambiae and Anopheles funestus in the transmission. Int J Mosq Res. 2019;6:5–10.
Charlwood JD, Vij R, Billingsley PF. Dry season refugia of malaria-transmitting mosquitoes in a dry Savannah zone of East Africa. Am J Trop Med Hyg. 2000;62:726–32.
Moiroux N, Boussari O, Djènontin A, Damien G, Cottrell G, Henry MC, et al. Dry season determinants of malaria disease and net use in Benin, West Africa. PLoS ONE. 2012;7: e30558.
Aikpon R, Osse R, Govoetchan R, Sovi A, Oké-Agbo F, Akogbeto M. Entomological baseline data on malaria transmission and susceptibility of Anopheles gambiae to insecticides in preparation for indoor residual spraying (IRS) in Atacora, (Benin). J Parasitol Vector Biol. 2013;7:102–11.
Djegbe I, Abel MA, Djouaka R, Akogbeto M. Surveillance entomologique: dynamique de la population et de la résistance aux insecticides chez Anopheles gambiae s.l. en milieu de riziculture irriguée Au Sud Bénin. J Appl Biosci J Appl Biosci. 2017;111:10934–43.
Gnanguenon V, Govoetchan R, Agossa FR, Ossè R, Oke-Agbo F, Azondekon R, et al. Transmission patterns of Plasmodium falciparum by Anopheles gambiae in Benin. Malar J. 2014;13: 444.
Salako AS, Ossè R, Padonou GG, Dagnon F, Aïkpon R, Kpanou C, et al. Population dynamics of Anopheles gambiae s.l. and Culex quinquefasciatus in rural and urban settings before an indoor residual spraying campaign in Northern Benin. Vector-Borne Zoonotic Dis. 2019;19:674–84.
Tokponnon TF, Ossè R, Padonou GG, Affoukou CD, Sidick A, Sewade W, et al. Entomological characteristics of malaria transmission across Benin: an essential element for improved deployment of vector control interventions. Insects. 2023;14:52.
Yadouleton A, Aïkpon R, Houndeton G, Aboubacar S, Ursins F, Tchibçozo C, et al. Données entomologiques préliminaires pour la mise en place d’une pulvérisation intra-domiciliaire à Grande échelle Dans la commune de Corpargo Au Nord-Est Du Bénin. Int J Biol Chem Sci. 2018;12:1993–2003.
Yovogan B, Sovi A, Padonou GG, Adoha CJ, Akinro B, et al. Pre-intervention characteristics of the mosquito species in Benin in preparation for a randomized controlled trial assessing the efficacy of dual active-ingredient long-lasting insecticidal nets for controlling insecticide-resistant malaria vectors. PLoS ONE. 2021;16:e0251742.
WHO. World malaria report 2016. Geneva: World Health Organization; 2016.
Moiroux N, Gomez MB, Pennetier C, Elanga E, Djènontin A, Chandre F, et al. Changes in Anopheles funestus biting behavior following universal coverage of long-lasting insecticidal nets in Benin. J Infect Dis. 2012;206:1622–9.
Ossè RA, Tokponnon F, Padonou GG, Sidick A, Aïkpon R, Fassinou A, et al. Involvement of Anopheles nili in Plasmodium falciparum transmission in North Benin. Malar J. 2019;18:152.
Salako AS, Ahogni I, Kpanou C, Sovi A, Azondekon R, Sominahouin AA, et al. Baseline entomologic data on malaria transmission in prelude to an indoor residual spraying intervention in the regions of Alibori and Donga, northern Benin, West Africa. Malar J. 2018;17:392.
Dia I, Guelbeogo MW. Advances and perspectives in the study of the malaria mosquito Anopheles funestus. In: Manguin S, editor. Anopheles mosquitoes—new insights into malaria vectors. London: InTechOpen; 2013.
Djamouko-Djonkam L, Nkahe DL, Kopya E, Talipouo A, Ngadjeu CS, Doumbe-Belisse P, et al. Implication of Anopheles funestus in malaria transmission in the city of Yaounde, Cameroon. Parasite. 2020;27:10.
Kopya E, Ndo C, Djamouko-djonkam L, Nkahe L, Awono-ambene P, Njiokou F, et al. Anopheles leesoni Evans 1931, a member of the Anopheles funestus group, is a potential malaria vector in Cameroon. Adv Entomol. 2022;10:99–109.
Awolola TS, Oyewole IO, Koekemoer LL, Coetzee M. Identification of three members of the Anopheles funestus (Diptera: Culicidae) group and their role in malaria transmission in two ecological zones in Nigeria. Trans R Soc Trop Med Hyg. 2005;99:525–31.
Aïkpon R, Ossè R, Dagnon F, Ahogni I, Tokponon F, Lyikirenga L, et al. Residual malaria transmission assessment: quantification of the relative risk of malaria transmission at different night-time and location in Atacora region in Benin, West Africa. J Entomol Zool Stud. 2018;6:1026–32.
Shaukat AM, Breman JG, McKenzie FE. Using the entomological inoculation rate to assess the impact of vector control on malaria parasite transmission and elimination. Malar J. 2010;9:122.
Kelly-Hope LA, McKenzie FE. The multiplicity of malaria transmission: a review of entomological inoculation rate measurements and methods across sub-Saharan Africa. Malar J. 2009;8:19.
Temu EA, Minjas JN, Tuno N, Kawada H, Takagi M. Identification of four members of the Anopheles funestus (Diptera: Culicidae) group and their role in Plasmodium falciparum transmission in Bagamoyo coastal Tanzania. Acta Trop. 2007;102:119–25.
Kamau L, Koekemoer LL, Hunt RH, Coetzee M. Anopheles parensis: the main member of the Anopheles funestus species group found resting inside human dwellings in Mwea area of central Kenya toward the end of the rainy season. J Am Mosq Control Assoc. 2003;19:130–3.
Mulamba C, Riveron JM, Ibrahim SS, Irving H, Barnes KG, Mukwaya LG, et al. Widespread pyrethroid and DDT resistance in the major malaria vector Anopheles funestus in East Africa is driven by metabolic resistance mechanisms. PLoS ONE. 2014;9:e110058.
Montoya LF, Martí-Soler H, Máquina M, Comiche K, Cuamba I, Alafo C, et al. The mosquito vectors that sustained malaria transmission during the Magude project despite the combined deployment of indoor residual spraying, insecticide-treated nets and mass-drug administration. PLoS ONE. 2022;17:e0271427.
Akoton R, Tchigossou GM, Djègbè I, Yessoufou A, Atoyebi MS, Tossou E, et al. Experimental huts trial of the efficacy of pyrethroids/piperonyl butoxide (PBO) net treatments for controlling multi-resistant populations of Anopheles funestus s.s. in Kpomè, southern Benin. Wellcome Open Res. 2018;3:71.
Djouaka R, Zeukeng F, Farrelle M, Soglo E, Adeoti R, Merdie O, et al. Heavy metal contamination and faecal coliforms in peri-urban market gardening sites in Benin and Cameroon. Int J Agric Environ Res. 2016;2:1013–44.
Yadouleton A, Martin T, Padonou G, Chandre F, Asidi A, Djogbenou L, et al. Cotton pest management practices and the selection of pyrethroid resistance in Anopheles gambiae population in northern Benin. Parasit Vectors. 2011;4:60.
Akogbéto MC, Padonou GG, Gbénou D, Irish S, Yadouleton A. Bendiocarb, a potential alternative against pyrethroid resistant Anopheles gambiae in Benin, West Africa. Malar J. 2010;9:204.
Namias A, Jobe NB, Paaijmans KP, Huijben S. The need for practical insecticide-resistance guidelines to effectively inform mosquito-borne disease control programs. Elife. 2021;10: e65655.
Serazin AC, Dana AN, Hillenmeyer ME, Lobo NF, Coulibaly MB, Willard MB, et al. Comparative analysis of the global transcriptome of Anopheles funestus from Mali, West Africa. PLoS ONE. 2009;4:e7976.
Riveron JM, Yunta C, Ibrahim SS, Djouaka R, Irving H, Menze BD, et al. A single mutation in the GSTe2 gene allows tracking of metabolically based insecticide resistance in a major malaria vector. Genome Biol. 2014;15: R27.
Tchigossou GM, Atoyebi SM, Akoton R, Tossou E, Innocent D, Riveron J, et al. Investigation of DDT resistance mechanisms in Anopheles funestus populations from northern and southern Benin reveals a key role of the GSTe2 gene. Malar J. 2020;19:456.
Tchigossou G, Djouaka R, Akoton R, Riveron JM, Irving H, Atoyebi S, et al. Molecular basis of permethrin and DDT resistance in an Anopheles funestus population from Benin. Parasit Vectors. 2018;11:602.
Medjigbodo AA, Djogbénou LS, Djihinto OY, Akoton RB, Abbey E, Kakossou RM, et al. Putative pleiotropic effects of the knockdown resistance (L1014F) allele on the life—history traits of Anopheles gambiae. Malar J. 2021;20:480.
Rivero A, Vézilier J, Weill M, Read AF, Gandon S. Insecticide control of vector-borne diseases: when is insecticide resistance a problem? PLoS Pathog. 2010;6: e1001000.
Riveron JM, Ibrahim SS, Chanda E, Mzilahowa T, Cuamba N, Irving H, et al. The highly polymorphic CYP6M7 cytochrome P450 gene partners with the directionally selected CYP6P9a and CYP6P9b genes to expand the pyrethroid resistance front in the malaria vector Anopheles funestus in Africa. BMC Genom. 2014;15:817.
Wondji CS, Dabire RK, Tukur Z, Irving H, Djouaka R, Morgan JC. Identification and distribution of a GABA receptor mutation conferring dieldrin resistance in the malaria vector Anopheles funestus in Africa. Insect Biochem Mol Biol. 2011;41:484–91.
Gari T, Lindtjørn B. Reshaping the vector control strategy for malaria elimination in Ethiopia in the context of current evidence and new tools: opportunities and challenges. Malar J. 2018;17:454.
Moiroux N, Damien GB, Egrot M, Djenontin A, Chandre F, Corbel V, et al. Human exposure to early morning Anopheles funestus biting behavior and personal protection provided by long-lasting insecticidal nets. PLoS ONE. 2014;9:e104967.
Adeogun AO, Olojede JB, Oduola AO, Awolola TS. Village-scale evaluation of PermaNet 3.0: an enhanced efficacy combination long-lasting insecticidal net against resistant populations of Anopheles gambiae s.s. Malar Chemother Control Elimin. 2012;1:235543.
Allossogbe M, Gnanguenon V, Yovogan B, Akinro B, Anagonou R, Agossa F, et al. WHO cone bio-assays of classical and new-generation long-lasting insecticidal nets call for innovative insecticides targeting the knock-down resistance mechanism in Benin. Malar J. 2017;16:77.
N’Guessan R, Asidi A, Boko P, Odjo A, Akogbeto M, Pigeon O, et al. An experimental hut evaluation of PermaNet?? 3.0, a deltamethrin-piperonyl butoxide combination net, against pyrethroid-resistant Anopheles gambiae and Culex quinquefasciatus mosquitoes in southern Benin. Trans R Soc Trop Med Hyg. 2010;104:758–65.
Pennetier C, Bouraima A, Chandre F, Piameu M, Etang J, Rossignol M, et al. Efficacy of Olyset® Plus, a new long-lasting insecticidal net incorporating permethrin and piperonil-butoxide against multi-resistant Malaria vectors. PLoS ONE. 2013;8:e75134.
Riveron JM, Watsenga F, Irving H, Irish SR, Wondji CS. High Plasmodium infection rate and reduced bed net efficacy in multiple insecticide-resistant malaria vectors in Kinshasa, Democratic Republic of Congo. J Infect Dis. 2018;217:320–8.
Menze BD, Kouamo MF, Wondji MJ, Tchapga W, Tchoupo M, Kusimo MO, et al. An experimental hut evaluation of PBO-based and pyrethroid-only nets against the malaria vector Anopheles funestus reveals a loss of bed nets efficacy associated with GSTe2 metabolic resistance. Genes. 2020;11:143.
Riveron JM, Huijben S, Tchapga W, Tchouakui M, Wondji MJ, Tchoupo M, et al. Escalation of pyrethroid resistance in the malaria vector Anopheles funestus induces a loss of efficacy of piperonyl butoxide-based insecticide-treated nets in Mozambique. J Infect Dis. 2019;220:467–75.
Tchouakui M, Mugenzi LMJ, Menze BD, Khaukha JNT, Tchapga W, Tchoupo M, et al. Pyrethroid resistance aggravation in Ugandan malaria vectors is reducing bednet efficacy. Pathogens. 2021;10:415.
Camara S, Ahoua Alou LP, Koffi AA, Clegban YCM, Kabran JP, Koffi FM, et al. Efficacy of Interceptor® G2, a new long-lasting insecticidal net against wild pyrethroid-resistant Anopheles gambiae s.s. from Côte d’Ivoire: a semi-field trial. Parasite. 2018;25:42.
N’Guessan R, Ngufor C, Kudom AA, Boko P, Odjo A, Malone D, et al. Mosquito nets treated with a mixture of chlorfenapyr and alphacypermethrin control pyrethroid resistant Anopheles gambiae and Culex quinquefasciatus mosquitoes in West Africa. PLoS ONE. 2014;9:e87710.
Ngufor C, Agbevo A, Fagbohoun J, Fongnikin A, Rowland M. Efficacy of royal guard, a new alpha-cypermethrin and pyriproxyfen treated mosquito net, against pyrethroid-resistant malaria vectors. Sci Rep. 2020;10:12227.
Tungu PK, Michael E, Sudi W, Kisinza WW, Rowland M. Efficacy of Interceptor® G2, a long-lasting insecticide mixture net treated with chlorfenapyr and alpha-cypermethrin against Anopheles funestus: experimental hut trials in north-eastern Tanzania. Malar J. 2021;20:180.
Parker JEA, Angarita-Jaimes N, Abe M, Towers CE, Towers D, McCall PJ. Infrared video tracking of Anopheles gambiae at insecticide-treated bed nets reveals rapid decisive impact after brief localised net contact. Sci Rep. 2015;5: 13392.
Angarita-Jaimes NC, Parker JEA, Abe M, Mashauri F, Martine J, Towers CE, et al. A novel video-tracking system to quantify the behaviour of nocturnal mosquitoes attacking human hosts in the field. J R Soc Interface. 2016;13:20150974.
Gabrieli P, Caccia S, Varotto-Boccazzi I, Arnoldi I, Barbieri G, Comandatore F, et al. Mosquito trilogy: microbiota, immunity and pathogens, and their implications for the control of disease transmission. Front Microbiol. 2021;12:630438.
Ricci I, Valzano M, Ulissi U, Epis S, Cappelli A, Favia G. Symbiotic control of mosquito borne disease. Pathog Glob Health. 2012;106:380–5.
Singh A, Allam M, Kwenda S, Khumalo ZTH, Ismail A, Oliver SV. The dynamic gut microbiota of zoophilic members of the Anopheles gambiae complex (Diptera: Culicidae). Sci Rep. 2022;12:1495.
Djihinto OY, Medjigbodo AA, Gangbadja ARA, Saizonou HM, Lagnika HO, Nanmede D, et al. Malaria-transmitting vectors microbiota: overview and interactions with Anopheles mosquito biology. Front Microbiol. 2022;13: 891573.
Jayakrishnan L, Sudhikumar AV, Aneesh EM. Role of gut inhabitants on vectorial capacity of mosquitoes. J Vector Borne Dis. 2018;55:69–78.
Dada N, Lol JC, Benedict AC, López F, Sheth M, Dzuris N, et al. Pyrethroid exposure alters internal and cuticle surface bacterial communities in Anopheles albimanus. ISME J. 2019;13:2447–64.
Dada N, Sheth M, Liebman K, Pinto J, Lenhart A. Whole metagenome sequencing reveals links between mosquito microbiota and insecticide resistance in malaria vectors. Sci Rep. 2018;8:2084.
Omoke D, Kipsum M, Otieno S, Esalimba E, Sheth M, Lenhart A, et al. Western Kenyan Anopheles gambiae showing intense permethrin resistance harbour distinct microbiota. Malar J. 2021;20:77.
Cansado-Utrilla C, Zhao SY, McCall PJ, Coon KL, Hughes GL. The microbiome and mosquito vectorial capacity: rich potential for discovery and translation. Microbiome. 2021;9:111.
Acknowledgements
We are grateful to Perugine Akoton for making the study map. We would like to thank the Global health for the grant (Grant No. INV024969) awarded to LSD.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
RA wrote the main manuscript text. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1.
Overall Plasmodium infection rates in Anopheles mosquito populations from Benin.
Additional file 2.
Evolution of insecticide resistance in An. funestus populations from Benin.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Akoton, R., Sovegnon, P.M., Djihinto, O.Y. et al. Vectorial competence, insecticide resistance in Anopheles funestus and operational implications for malaria vector control strategies in Benin Republic. Malar J 22, 385 (2023). https://doi.org/10.1186/s12936-023-04815-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12936-023-04815-9