
Abstract
Background
Chromosomally encoded AmpC β-lactamases may be acquired by transmissible plasmids which consequently can disseminate into bacteria lacking or poorly expressing a chromosomal bla AmpC gene. Nowadays, these plasmid-mediated AmpC β-lactamases are found in different bacterial species, namely Enterobacteriaceae, which typically do not express these types of β-lactamase such as Klebsiella spp. or Escherichia coli. This study was performed to characterize two E. coli isolates collected in two different Portuguese hospitals, both carrying a novel CMY-2-type β-lactamase-encoding gene.
Findings
Both isolates, INSRA1169 and INSRA3413, and their respective transformants, were non-susceptible to amoxicillin, amoxicillin plus clavulanic acid, cephalothin, cefoxitin, ceftazidime and cefotaxime, but susceptible to cefepime and imipenem, and presented evidence of synergy between cloxacilin and cefoxitin and/or ceftazidime. The genetic characterization of both isolates revealed the presence of bla CMY-46 and bla CMY-50 genes, respectively, and the following three resistance-encoding regions: a Citrobacter freundii chromosome-type structure encompassing a blc-sugE-bla CMY-2-type -ampR platform; a sul1-type class 1 integron with two antibiotic resistance gene cassettes (dfrA1 and aadA1); and a truncated mercury resistance operon.
Conclusions
This study describes two new bla CMY-2-type genes in E. coli isolates, located within a C. freundii-derived fragment, which may suggest their mobilization through mobile genetic elements. The presence of the three different resistance regions in these isolates, with diverse genetic determinants of resistance and mobile elements, may further contribute to the emergence and spread of these genes, both at a chromosomal or/and plasmid level.
Similar content being viewed by others

Background
AmpC β-lactamases, along with Class A β-lactamases, are a major group of clinically important enzymes [1,2]. They belong to class C according to the Ambler classification and to group 1 following the functional classification of Bush-Jacoby [3,4], whose prevalence is increasing worldwide [1]; these β-lactamases are associated with infections caused by pathogenic Gram-negative bacteria, particularly Escherichia coli and Klebsiella pneumoniae. The identification of isolates containing plasmid-mediated AmpC-β-lactamase (PMAβ) is epidemiologically and clinically relevant due to the limitations of treatment options [5].
AmpC enzymes hydrolyse amino- and ureidopenicillins, and cephamycins (cefoxitin and cefotetan) and, at a low level, oxyiminocephalosporins (ceftazidime, cefotaxime, and ceftriaxone) and aztreonam: they are not inhibited by β-lactamase inhibitors such as clavulanic acid [1]. AmpC-producing isolates are susceptible to carbapenems and to zwitterionic cephalosporins (cefepime and cefpirome).
In this study, we performed the phenotypic and molecular characterization of two new CMY-2-types (designated CMY-46 and CMY-50), both encoded by probably chromosomal inducible ampC genes, produced by two clinical E. coli isolates. The genetic environment of bla CMY-46 and bla CMY-50 was also investigated.
Methods
Bacterial isolate collection
Two clinical E. coli strains (INSRA1169 and INSRA3413) were isolated, in 1999, from urine samples of two patients of 77 years and 7 months old, in two different hospitals in Portugal. E. coli DH5α (pBK-CMY-2) strain was used as control for antimicrobial susceptibility tests.
Antimicrobial susceptibility tests
Minimal inhibitory concentrations were determined by a microdilution method according to guidelines of the French Society of Microbiology (SFM 2013, http://www.sfm-microbiologie.org/) against seven β-lactams, alone or in combination with clavulanic acid, and against ciprofloxacin, gentamicin and trimethoprim. Isolates non-susceptible to one third-generation cephalosporin, cefoxitin and/or exhibiting synergy with boronic acid and/or cloxacillin, were considered as presumptive AmpC producers. Imipinem and clavulanic acid were used in order to identify induction effect of AmpC [1,6]. Disks of inducing agents (imipenem 10 μg and amoxicillin plus clavulanic acid 25 + 10 μg) and disks of cephalosporins (cefotaxime 30 μg and ceftazidime 30 μg) were placed on Mueller–Hinton agar plates, 20 mm apart. Positive induction was demonstrated by the antagonism effect surrounding the cephalosporin disks adjacent to the inducers.
Isoelectric point determination
β-Lactamases were characterized by isoelectric focusing of ultrasonicated bacterial extracts with the control strains expressing pI 5.2, 5.6, 7.6, 9.0, 9.2, as previously described [7].
Molecular characterization of ampC and ESBL-encoding genes
The presence of acquired ampC (bla CMY, bla MOX, bla FOX, bla LAT, bla ACT, bla MIR, bla DHA, bla MOR, bla ACC) and bla ESBL genes (bla TEM, bla SHV, bla OXA-1-type, bla CTX-M) was investigated by multiplex PCR assays with primers and conditions as described elsewhere [7-10], and those from Table 1. Controls were included in all assays.
Gene transfer experiments
Transferability of the bla CMY genes was attempted by both broth mating-out assays and electroporation. Conjugation experiments were performed at 37°C, using recipient strains E. coli C600 RifR, StrR and E. coli J53 NaN3 R, according to the antibiotic susceptibilities of the clinical isolates used as donor. Transconjugants were selected on MacConkey agar plates containing 250 μg/ml of rifampicin, 160 μg/ml of streptomycin or 160 μg/ml of sodium azide plus 10 μg/ml of cefoxitin. Plasmid DNA was extracted from clinical strains, using the Wizard Plus Midipreps DNA Purification kit (Promega), and used to transform electrocompetent E. coli DH5α ∆ampC by electroporation, as previously described [7]. Transformants were selected on Luria broth medium containing 10 μg/ml of cefoxitin.
Cloning experiments
The bla CMY-2, bla CMY-46 and bla CMY-50 genes were amplified with iProofTM High-Fidelity DNA Polymerase (Bio-Rad, Hercules, CA), using primers from Table 1. Amplicons (1169 bp) were ligated in the SmaI site of the phagemid pBK-CMV (Stratagene) downstream of its inducible lacZ promoter and transformed into electrocompetent E. coli DH5α ∆ampC cells. A gene Pulser II apparatus (Bio-Rad, Hercules, CA) was used for standard electroporation techniques, as previously described [7]. Recombinant bacteria were selected on LB agar plates containing 10 μg/ml of cefoxitin.
Genetic background characterization
The presence of class 1 integrons was determined in both isolates through PCR amplification of the integrase-specific intI1 gene with the same specific primers and conditions as reported previously [11] (Table 1). PCR-mapping and sequencing of the genetic environment of bla CMY-46 and bla CMY-50 was performed using primers targeting genes known for promoting antibiotic resistance and integrons (Table 1). Sequence alignments and generation of resistance cassette contigs were performed using Bionumerics (Applied Maths). Gene identity was confirmed at the NCBI website (http://www.ncbi.nlm.nih.gov/).
Findings
The two clinical E. coli isolates INSRA1169 and INSRA3413 were resistant to amoxicillin, amoxicillin plus clavulanic acid, cephalothin, cefoxitin, ceftazidime, cefotaxime, gentamicin and trimethoprim, but susceptible to cefepime and imipenem (Table 2). INSRA1169 was also nonsusceptible to ciprofloxacin. Synergy between cloxacillin and cefoxitin plus cefotaxime and boronic acid, along with the absence of synergy between extended-spectrum cephalosporins and clavulanic acid, suggest that the resistance to extended-spectrum cephalosporins was mediated by the overproduction of AmpC β-lactamases. The resistance phenotype was not transferable, neither in conjugation assays with E. coli C600 as a recipient, or in transformation assays by electroporation of plasmid-DNA preparations into E. coli DH5α. This might suggest a chromosomal location of AmpC-encoding genes.
Transformants, obtained after cloning of DNA amplicons of INSRA1169 and INSRA3413 in pBK-CMV plasmid vector (Figure 1a), showed a resistance phenotype similar to that of clinical isolates. However, they were susceptible to ciprofloxacin, gentamicin and trimethoprim, like the control strain E. coli DH5α (pBK-CMY-2) (Table 2). Both the clinical strains and the transformants produced β-lactamases exhibiting an alkaline isoelectric point (pI 9.2) compatible with AmpC-type β-lactamases. Indeed, PCR revealed the absence of bla ESBL genes plus the presence of chromosomal E. coli ampC-type gene; the sequence of cloned DNA fragments identified two new CMY-2-like genes, which were not related to chromosome-mediated E. coli AmpC gene. The chromosomal location of such genes has only been observed in Salmonella spp. and Proteus mirabilis isolates [12-14].

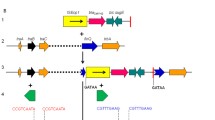
Schematic representation of the same three structures found within clinical isolates expressing bla CMY-46 and bla CMY-50 . The directions of transcription of the corresponding genes are depicted by arrows. A: sequence, including the genetic environment of bla CMY-type genes, compared with C. freundii chromosomal region (GenBank JH414884); B: class 1 integron, with attI1 site (grey circle) and the two attc regions (open circles); C: truncated mercury resistance operon.
The deduced amino acid sequences confirmed that the new genes encoding the β-lactamases CMY-46 (in INSRA1169) and CMY-50 (in INSRA3413), which were new variants of CMY-2, differed by 9 and 13 amino acid substitutions, respectively (Table 3). Two of these mutations (Q193K plus P208A for CMY-46 and N194S plus D198N for CMY-50) are in the Ω loop (between amino acids 178 and 226), which interacts by hydrogen bonding with helix H-2 close to the active Ser64. Substitutions in this region have been linked to the extension of the hydrolysis spectrum [15]. However, CMY-46 and CMY-50 β-lactamases did not confer resistance to cefepime and conferred low level of resistance to ceftazidime and cefotaxime, which suggests that, in contrast to extended-spectrum AmpCs, they have moderate or no extended-spectrum activity (Table 2) [15-18].
The study of sequences surrounding bla CMY-46 and bla CMY-50 revealed the presence of the blc gene (encoding an outer membrane lipoprotein) and the sugE gene (encoding a small MDR protein responsible for resistance to quaternary ammonium compounds) downstream of their open reading frames (Figure 1a). Upstream, an ampR gene encoding the usual transcriptional regulator of ampC genes was observed in an opposite direction of transcription (Figure 1a). The presence of an intact ampC-ampR segment in both new bla CMY genetic regions implied that the production of CMY-46 and CMY-50 is inducible, which was corroborated by the used phenotypic induction method. This ampC-ampR region was identical to the sequence flanking the bla AmpC gene in the C. freundii chromosome [6], except for AmpRCMY-46 (that had 4 amino acid substitutions), but none were located in the helix-turn-helix region or in other positions related to AmpR function [20,21]. The promoter regions of our bla CMY-2-type and ampR genes harbored no sequence element associated with increased strength of the promoter [20,21]. In addition, the frdD, frdC, and frdB genes that are usually adjacent to ampC-ampR in the C. freundii chromosome were not identified in the sequences flanking bla CMY-46 or bla CMY-50.
Class 1 integrons, also detected in INSRA1169 and INSRA3413 (Figure 1b), comprised the integrase-encoding gene intI1, two gene cassettes, aacA1 and dfrA1, and qacEΔ1 plus sul1, which were probably responsible for the observed resistances to trimethoprim and aminoglycosides. We also found a truncated mercury resistance operon (Figure 1c), which was previously reported as belonging to a “kan region” that included a kanamycin resistance gene [22]. This finding is of concern since mercury resistance may help to promote antibiotic resistance through indirect selection [23].
In summary, this study describes two new bla CMY-2-type genes located within a C. freundii-derived fragment. Considering that CMY-type β-lactamases, detected in E. coli, are derived from the C. freundii chromosomal AmpC [1] and that chromosome-derived genes are usually mobilized by MGE [24,25], the presence of three resistance regions with diverse resistance determinants and MGE in this study, suggests the dynamics of bacteria in the transference of antibiotic resistance. In addition, they might also trigger the future emergence and spread of these resistant determinants both at a chromosomal or/and plasmid level.
Availability of supporting data
The data set supporting the results of this article is included within the article.
Abbreviations
- PMAβ:
-
Plasmid-mediated AmpC-β-lactamase
- SFM:
-
French Society of Microbiology
- MDR:
-
Multidrug-resistance
- MGE:
-
Mobile genetic elements
- ESAC:
-
Extended-spectrum AmpC cephalosporinases
- ESBL:
-
Extended-spectrum β-lactamase
References
Jacoby GA. AmpC β-lactamases. Clin Microbiol Rev. 2009;22:161–82.
Rice LB, Bonomo RA. β-Lactamases: which ones are clinically important? Drug Resist Updat. 2000;3:178–89.
Ambler RP, Coulson AF, Frere JM, Ghuysen JM, Joris B, Forsman M, et al. A standard numbering scheme for the class A β-lactamases. Biochem J. 1991;276:269–70.
Bush K, Jacoby GA. Updated functional classification of β-lactamases. Antimicrob Agents Chemother. 2010;54:969–76.
Pai H, Kang CI, Byeon JH, Lee KD, Park WB, Kim HB, et al. Epidemiology and clinical features of bloodstream infections caused by AmpC-type-β-lactamase-producing Klebsiella pneumoniae. Antimicrob Agents Chemother. 2004;48:3720–8.
Lindquist S, Lindberg F, Normark S. Binding of the Citrobacter freundii AmpR regulator to a single DNA site provides both autoregulation and activation of the inducible ampC β-lactamase gene. J Bacteriol. 1989;171:3746–53.
Mendonça N, Leitão J, Manageiro V, Ferreira E, Antimicrobial Resistance Surveillance Program in Portugal (ARSIP), Caniça M. Spread of extended-spectrum β-lactamase CTX-M-producing Escherichia coli clinical isolates in community and nosocomial environments in Portugal. Antimicrob Agents Chemother. 2007;54:1946–55.
Pérez-Pérez FJ, Hanson ND. Detection of plasmid-mediated AmpC β-lactamase genes in clinical isolates by using multiplex PCR. J Clin Microbiol. 2002;40:2153–62.
Jones-Dias D, Manageiro V, Francisco AP, Martins AP, Domingues G, Louro D, et al. Assessing the molecular basis of transferable quinolone resistance in Escherichia coli and Salmonella spp. from food-producing animals and food products. Vet Microbiol. 2013;167:523–31.
Manageiro V, Ferreira E, Caniça M, Manaia CM. GES-5 among the β-lactamases detected in ubiquitous bacteria isolated from aquatic environment samples. FEMS Microbiol Lett. 2014;351:64–9.
Leverstein-Van Hall MA, Paauw A, Box AT, Blok HE, Verhoef J, Fluit AC. Presence of integron-associated resistance in the community is widespread and contributes to multidrug resistance in the hospital. J Clin Microbiol. 2002;40:3038–40.
Shahada F, Sekizuka T, Kuroda M, Kusumoto M, Ohishi D, Matsumoto A, et al. Characterization of Salmonella enterica serovar Typhimurium isolates harboring a chromosomally encoded CMY-2 β-lactamase gene located on a multidrug resistance genomic island. Antimicrob Agents Chemother. 2011;55:4114–21.
Zioga A, Whichard JM, Joyce KJ, Tzelepi E, Tzouvelekis LS, Miriagou V. Evidence for chromosomal and plasmid location of CMY-2 cephalosporinase gene in Salmonella serotype Typhimurium. J Antimicrob Chemother. 2008;61:1389–90.
D’Andrea MM, Literacka E, Zioga A, Giani T, Baraniak A, Fiett J, et al. Evolution and spread of a multidrug-resistant Proteus mirabilis clone with chromosomal AmpC-type cephalosporinases in Europe. Antimicrob Agents Chemother. 2011;55:2735–42.
Nordmann P, Mammeri H. Extended-spectrum cephalosporinases: structure, detection and epidemiology. Future Microbiol. 2007;2:297–307.
Rodríguez-Martínez JM, Poirel L, Nordmann P. Extended-spectrum cephalosporinases in Pseudomonas aeruginosa. Antimicrob Agents Chemother. 2009;53:1766–71.
Rodríguez-Martínez JM, Nordmann P, Ronco E, Poirel L. Extended-spectrum cephalosporinase in Acinetobacter baumannii. Antimicrob Agents Chemother. 2010;54:3484–8.
Rodríguez-Martínez JM, Fernández-Echauri P, Fernández-Cuenca F, Diaz-de-Alba P, Briales A, Pascual A. Genetic characterization of an extended-spectrum AmpC cephalosporinase with hydrolysing activity against fourth-generation cephalosporins in a clinical isolate of Enterobacter aerogenes selected in vivo. J Antimicrob Chemother. 2012;67:64–8.
Bauernfeind A, Stemplinger I, Jungwirth R, Giamarellou H. Characterization of the plasmidic β-lactamase CMY-2, which is responsible for cephamycin resistance. Antimicrob Agents Chemother. 1996;40:221–4.
Bartowsky E, Normark S. Interactions of wild-type and mutant AmpR of Citrobacter freundii with target DNA. Mol Microbiol. 1993;10:555–65.
Hanson ND, Sanders CC. Regulation of inducible AmpC β-lactamase expression among Enterobacteriaceae. Curr Pharm Des. 1999;5:881–94.
Call DR, Singer RS, Meng D, Broschat SL, Orfe LH, Anderson JM, et al. bla CMY-2-positive IncA/C plasmids from Escherichia coli and Salmonella enterica are a distinct component of a larger lineage of plasmids. Antimicrob Agents Chemother. 2010;54:590–6.
Baker-Austin C, Wright MS, Stepanauskas R, McArthur JV. Co-selection of antibiotic and metal resistance. Trends Microbiol. 2006;14:176–82.
Frost LS, Leplae R, Summers AO, Toussaint A. Mobile genetic elements: the agents of open source evolution. Nat Rev Microbiol. 2005;3:722–32.
Norman A, Hansen LH, Sørensen SJ. Conjugative plasmids: vessels of the communal gene pool. Phil Trans R Soc B. 2009;364:2275–89.
Acknowledgements
This study was supported by a grant from the Department of Infectious Diseases, National Institute of Health Dr. Ricardo Jorge (in 2012). V. Manageiro was supported by grant SFRH/BPD/77486/2011 from the Fundação para a Ciência e a Tecnologia, Lisbon, Portugal. This work was funded by “Strategic Plan for Environmental and Natural Sciences, Project UI/211–2011–2012” referenced as “Pest-OE/AGR/UI0211/2011” from CECA-ICETA.
The GenBank accession numbers for the new AmpC-encoding genes are FN556186 for bla CMY-46 and FN645444 for bla CMY-50.
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
VM performed the experiments, data interpretation and drafted the manuscript; EF and MF took part in the experiments; MP and FF worked on microbiology and clinical data; RB participated in data interpretation and reviewing of the manuscript; and MC conceived the study, and contributed to data interpretation and reviewing of the manuscript. All authors read and approved the final manuscript.
Rights and permissions
This article is published under an open access license. Please check the 'Copyright Information' section either on this page or in the PDF for details of this license and what re-use is permitted. If your intended use exceeds what is permitted by the license or if you are unable to locate the licence and re-use information, please contact the Rights and Permissions team.
About this article

Cite this article
Manageiro, V., Ferreira, E., Pinto, M. et al. Two novel CMY-2-type β-lactamases encountered in clinical Escherichia coli isolates. Ann Clin Microbiol Antimicrob 14, 12 (2015). https://doi.org/10.1186/s12941-015-0070-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12941-015-0070-8