
Abstract
Pancreatic cancer is one of the most lethal malignancies. Recent studies indicated that development of pancreatic cancer may be intimately connected with the microbiome. In this review, we discuss the mechanisms through which microbiomes affect the development of pancreatic cancer, including inflammation and immunomodulation. Potential therapeutic and diagnostic applications of microbiomes are also discussed. For example, microbiomes may serve as diagnostic markers for pancreatic cancer, and may also play an important role in determining the efficacies of treatments such as chemo- and immunotherapies. Future studies will provide additional insights into the various roles of microbiomes in pancreatic cancer.
Similar content being viewed by others
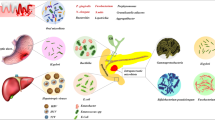

Background
Pancreatic cancer (PC), one of the most lethal malignancies, is the 10th most frequent cancer in men and the 9th most common in women in the United States. PC is responsible for the third-highest number of cancer-related deaths [1]. The incidence of PC has shown a rapid upward trend in recent years. PC onset is difficult to detect, and early symptoms are atypical. Many patients are diagnosed with local progression or distal metastasis and are not candidates for surgery, leading to a 5-year survival rate of less than 9% [1]. Therefore, it is very important to better understand the occurrence and development of PC to enable early diagnosis and treatment. In recent years, associations between microbiomes and the occurrence and development of PC have been identified, potentially representing an early screening and risk assessment factor. Furthermore, inflammation and immunosuppression caused by microbiome changes are recognized as mechanisms associated with cancer development [2,3,4]. In addition, the microbiome may also affect the metabolism of chemotherapy drugs, thereby modulating the effects of chemotherapy [4, 5]. This review will summarize these complex issues (Fig. 1).

Microbiomes play important roles in the development and treatment of pancreatic cancer. The blue arrow indicates that microbiome exposure activates inflammation to promote development of pancreatic cancer. The orange arrow shows that the microbiome leads to immune suppression. The green arrow shows that the microbiome influences the effects of cancer treatments. AHL-12, N-acetyl-dodecanoyl homoserine; T2R38, one of the family of bitter receptors; mTOR, mammalian target of rapamycin; LPS, lipopolysaccharide; TLR, Toll-like receptor; AP-1, Activator protein 1; STAT3, Signal transducers and activators of transcription 3; Th1/2, helper T cell 1/2; PD-1, programmed cell death-1; CDD, cytidine deaminase; 2′,2′-difluorodeoxycytidine, gemcitabine; 2′,2′-difluorodeoxyuridine, an inactive form of gemcitabine
Roles of microbiomes in development of PC
Infectious factors play a causative role in approximately 10–20% of all cancers worldwide [6]. However, in PC, no microbe has been identified as a causative agent. Many studies have suggested that changes in the diversity, proportions and dominant organisms of the microbiome (Porphyromonas, Actinomycetes, Neisseria, Streptococcus, Bacteroides Bifidobacteria and Fusobacterium species) may be associated with the occurrence and development of PC [7,8,9,10,11,12,13,14]. Particularly, Riquelme et al. used 16S rRNA gene sequencing to analyze the tumor microbiome composition of PDAC patients with short-term survival and long-term survival [15]. They supported Pseudoxanthomonas-Streptomyces-Saccharopolyspora-Bacillus clausii can highly predict the long-term survivorship, which showed the effect of microbiome on prognosis. Although it remains unclear whether these microbiome properties are directly associated with PC, the studies summarized in Table 1 have demonstrated some preliminary correlations.
The oral microbiome and PC
The oral cavity is a large reservoir of microbes including more than 700 types of bacteria, viruses and fungi [16, 32]. When conditions change, the commensal microbiomes can become pathogenic and lead diseases including PC [16]. Periodontal disease, an inflammation caused by oral microbes, has been regarded as a risk factor for PC. For example, Maisonneuve et al. conducted a meta-analysis of eight studies and suggested a significant link between periodontal disease and increased risk of PC [33]. The relative risks for PC in individuals with periodontitis and edentulism were 1.74 (95% confidence interval, CI 1.41–2.15) and 1.54 (95%CI 1.16–2.05), respectively [33]. Farrell et al. conducted an analysis using human oral microbiological microarrays to study variation in salivary microbiomes among 10 patients with resectable PC and 10 matched healthy controls. They identified 410 bacterial taxa [11], including 16 (3.9%) organisms such as Neisseria elongata and Streptococcus mitis whose frequencies differed significantly between the two groups. This study revealed significant changes in the microbial composition of PC patients and healthy controls [11]. In addition, Porphyromonas gingivalis, a commonly identified bacterium in patients with periodontal disease, is thought to increase risk of developing PC [17, 34]. To understand the connection between oral microbiomes and PC, Michaud et al. measured antibodies against oral bacteria in pre-diagnostic blood samples from 405 PC patients and 416 matched controls nested in the European Prospective Investigation into Cancer and Nutrition (EPIC) study [35]. The results showed that individuals with high levels of antibodies against P. gingivalis ATCC 53978 were at two-fold increased risk of PC compared with individuals with lower levels of these antibodies [35]. In addition, the authors found that individuals with consistently high levels of antibodies to common oral bacteria were at 45% lower risk for PC compared to those with lower antibody levels [35]. Similarly, other studies using 16S rRNA sequencing suggested that the presence of Haemophilus, Porphyromonas, Leptotrichia and Fusobacterium species in the oral cavity was also associated with increased risk of PC in humans [9, 18, 36].
As one of star oral microbiomes associated with PC, P. gingivalis has been extensively studied. Although several studies have shown that P. gingivalis is a new risk factor for PC, further researches are needed to explore the specific mechanisms leading to PC. Some researchers hypothesized that one potential mechanism might be that a peptidyl-arginine deiminase enzyme secreted by P. gingivalis leads to p53 and K-ras mutations following degradation of arginine [19], while others focus on the effect of oral bacteria on the systemic immune response, including IL-1β, IFNγ, and TNF [2, 37]. How, then, can the oral microbiome affect the pancreas? Gaiser et al. found higher loads of oral bacterial DNA in the cyst fluid of intraductal papillary mucinous neoplasms (a condition which can progress to PC), providing support for the relationship between oral and pancreatic microbes [20]. By feeding wild-type mice fluorescently-labeled Enterococcus faecalis or Escherichia coli, Pushalkar et al. found these bacteria accrued in the pancreas of mice, demonstrating that microbes can migrate to the pancreas and directly affect the pancreatic microenvironment [4]. However, no clear evidence has indicated which channels microbes use to reach the pancreas. In fact, although the pancreas belongs to distal organ of the digestive tract, it is still connected to the digestive tract via duodenum. Therefore, oral microbes are likely to enter the pancreas through the digestive tract. In addition, it is well known that oral microorganisms easily enter the blood, leading to bacteremia. Therefore, oral microorganisms may also enter the pancreas through blood circulation. In summary, the exact channel of oral microbes into the pancreas requires further researches to determine.
Helicobacter pylori and PC
Helicobacter pylori, a well-known bacterium that colonizes the human stomach, has been the subject of increasing attention over the last 30 years [38]. Many previous studies have attempted to correlate the presence of H. pylori with PC using serologic and culture-based methods. However, the vast majority of commensal microbes cannot be cultured, affecting the objective investigation of their role in pancreatic diseases [21, 39,40,41,42]. New techniques, such as next-generation sequencing and metagenomics, have provided a more representative assessment of the microbial community in health and disease and the dynamic interactions between microbiomes and their human hosts [22]. These techniques may help in understanding the association between H. pylori and PC. To determine whether H. pylori infection was associated with PC, Raderer et al. used enzyme-linked immunosorbent assays to analyze IgG antibodies against H. pylori in blood samples from 92 patients with PC, 30 patients with gastric cancer, 35 patients with colorectal cancer, and 27 healthy volunteers [39]. The results showed that 65% of patients with PC and 69% of patients with gastric cancer were seropositive, compared with only 45% of the other individuals [39]. Mei et al. found that H. pylori could be detected in the duodenum at higher frequencies in PC patients than in healthy controls [23]. Similarly, other studies used meta-analysis to confirm that H. pylori was associated with increased risk of PC in humans [24,25,26, 43, 44]. The probable mechanism of microbiome transmission involves changes in the intestinal microbial environment, which can alter the composition of the intestinal microbiome, increase intestinal permeability and permit microbial access to the bloodstream and distant organs [27, 45]. H. pylori may promote the development of PC by causing chronic mucosal inflammation as well as changes in cell proliferation and differentiation [28].
However, several other studies concluded that H. pylori was not associated with PC [25, 29, 46, 47]. One of the potential explanations for this discrepancy is that H. pylori has several variants, of which cytotoxin-associated gene A (Cag-A) positive strains have been the best studied. Cag-A has been linked to multiple diseases such as gastric inflammation and ulceration, gastric cancer and PC [48, 49]. However, it remains controversial whether Cag-A-positive or Cag-A-negative strains are associated with PC [50, 51]. An effect modification by ABO blood type was reported in a large case-control study: the association between PC and Cag-A-negative H. pylori was evident only in individuals with non-O blood types [41]. This finding may be explained by differences in the terminal antigens of gastrointestinal mucins in individuals with non-O blood types, which affects binding by H. pylori [2].
No single clear mechanism has been widely accepted to explain associations between H. pylori and PC. One hypothesis suggests that H. pylori infection leads to hyperchlorhydria and enhanced release of secretin, promoting pancreatic hyperplasia [52]. Conversely, another hypothesis suggested that H. pylori infection led to atrophic gastritis and hypochlorhydria, resulting in bacterial overgrowth and overproduction of N-nitrosamines [53]. In summary, the role of H. pylori in PC remains unclear. Further studies are required to consider other potential confounding risk factors and conclusively explore whether H. pylori is truly associated with the occurrence and development of PC.
The pancreatic microbiome
Traditionally, the pancreas has not been considered to have its own microbiome. Li et al. performed bacterial 16S rRNA gene-specific PCR to analyze the microbial constituents in the pancreatic cyst fluids, where Bacteroides, Escherichia/Shigella, and Acidaminococcus were predominant [30]. They reflected the local microbiota in the pancreas, and prove that pancreatic cyst fluid is a very important sample for microbial identification. Now, in addition to P. gingivalis and H. pylori, other microbes have been identified in PC tissues. Using 16S rRNA gene sequencing, Pushalkar et al. found high proportions of Proteobacteria (45%), Bacteroidetes (31%), and Firmicutes (22%) species in PC tissues [4]. Interestingly, they concluded that the microbiome proportions in PC tissue were quite different from those of normal pancreatic tissue. Some recent research also showed similar results [31, 54]. Thus, the pancreas is not sterile and has its own microbial environment which may affect the occurrence and development of PC. More complex mechanisms involving a large number of factors may alter the pancreatic microbiome. These alterations may occur via natural and non-natural channels.
Fungi and viruses in PC
Some studies have linked fungi and viruses to the development of PC. For example, a prospective cohort study conducted in Sweden showed that Candida infection in the oral cavity was associated with development of PC [55]. Similarly, another population-based cohort study in Taiwan showed that risk of PC was significantly higher among Candida-infected individuals [56]. However, the relationships between fungal infections and PC require further study. The roles of hepatitis viruses in the development of hepatocellular carcinoma are relatively clear. However, some evidence suggested that hepatitis viruses may also be associated with PC. Katakura et al. found increased serum levels of pancreatic enzymes in viral hepatitis patients [57], while Jin et al. identified a link between hepatitis B virus and chronic pancreatitis [58]. These studies link chronic hepatitis, chronic pancreatitis and PC and demonstrate that a potential role of viruses in PC cannot be ignored.
Mechanisms of microbiome involvement in development of PC
Microbes and pancreatic inflammation
Microbial infections often lead to inflammation [59]. Sometimes inflammation is a protective response to factors such as pro-inflammatory mediators, environmental toxins, or chronic infection [60,61,62]. However, inflammation is also a risk factor for development of many cancers. For example, Dejea et al. demonstrated that patients with familial adenomatous polyposis had higher frequencies of E. coli and Bacteroides fragilis cells in the colonic mucosa compared with healthy individuals. Moreover, tumor-prone mice had higher interleukin-17 (IL-17) and IL-23 levels and developed tumors faster upon stimulation by microbes [63, 64]. Inflammation of the pancreas also increases the incidence of PC. Patients with hereditary autoimmune pancreatitis are estimated to carry a lifetime risk of 40% of developing PC and patients with chronic pancreatitis, a recognized risk factor for PC, have a 13-fold higher risk of PC than other individuals [65, 66]. Multiple cohort studies have shown that acute pancreatitis is also associated with the development and progression of PC [67,68,69]. However, acute pancreatitis is not a direct factor, but the chronic prolongation of inflammation leads to the occurrence and development of PC [68]. Although there are no identified pathogens, this type of chronic inflammation can also be caused by microbial infections [70]. Microbial-induced inflammation leads to tumorigenesis through activation of tumor-related inflammatory signaling pathways.
Macropinocytosis and Wnt signaling
Microbes can trigger macropinocytosis, an endocytic process used by cells for antigen capture and presentation, to activate inflammation [71, 72]. For example, Bacillus Calmette-Guérin (BCG) can be internalized through macropinocytosis to treat bladder cancer [73, 74]. Moreover, Salmonella species can invade mammalian cells by inducing macropinocytosis through actin remodeling [72, 75]. Importantly, the process of macropinocytosis is closely related to the Wnt (Wingless/Integrated) signaling pathway, which is important for cell proliferation and differentiation during tumorigenesis, including PC [76]. For instance, Redelman-Sidi et al. demonstrated that Wnt-driven macropinocytosis occurred downstream of the β-catenin–dependent canonical Wnt pathway and was PAK1 dependent, contributing to cancer growth during the early phases of oncogenesis [77]. In PC cells, Wnt pathway activation is also relevant to macropinocytosis [71]. Although the details of this mechanism require further study, the influence of the Wnt signaling pathway on microbial uptake is clear.
Lipopolysaccharide stimulation of toll-like receptors may link microbiomes to inflammation
Dysbiotic microbial compositions could also interact with some receptors in cells to active inflammation and promote tumorigenesis. Lipopolysaccharide (LPS), a Gram-negative bacterial cell wall component, is specifically recognized by Toll-like receptor 4 (TLR4), a family member of pattern recognition receptors (PRR) [10, 78]. The interaction between LPS and TLR4 can activate the secretion of downstream pro-inflammatory cytokines [78], linking microbes to inflammation. Below, we discuss several molecules related to LPS-TLR signaling and their relevance to PC.
Physiological disorders resulting in destruction of the gut microbiome can lead to inflammatory conditions and some types of cancer. These pathologies are controlled by mammalian target of rapamycin (mTOR) [79], which is a downstream effector of TLRs [80]. Moreover, mTOR also plays a vital role in tumorigenesis, including in PC [81,82,83]. Phosphorylation of Extracellular regulated protein kinases 1/2 (ERK1/2) and mTOR was inhibited and pancreatic tumor size was reduced in mice if the engrafted pancreatic tumor cells were cultured in engineered-resistant starch [79, 84], which can shape the composition of the gut microbiomes. Therefore, the gut microbiome can influence the mTOR pathway and promote PC.
The nuclear factor kappa B/mitogen-activated protein kinase (NF-κB/MAPK) signaling pathway plays a major role in inflammation [17]. The NF-κB/MAPK signaling pathway, whose core complexes are c-fos/Jun and p50/p65, is associated with both inflammation and tumorigenesis (Fig. 2). The interaction between LPS and TLR4 can activate both NF-κB and Activator protein 1 (AP-1), leading to expression of pro-inflammatory cytokines and dysregulation of cellular processes [85, 86]. Similar to the role of LPS-TLR signaling in inflammation, Beller et al. found that Pseudomonas aeruginosa N-acetyl-dodecanoyl homoserine (AHL-12) could also activate downstream AP-1 by binding to the bitter receptor T2R38, which was initially identified in taste bud cells in the oral cavity [87,88,89]. Expression of T2R38 has also been described in pancreatic tumor cells [90]. Thus, T2R38 may represent one bridge connecting the microbiome with PC. Further research is needed to explore the relationship between T2R38 and PC.

Microbiomes are involved in the occurrence pancreatic cancer. Microbiomes can lead to development of inflammation, inhibit interactions between macrophages and T cells, and favor Th2 polarization of the T cell response. All of these factors can contribute to the occurrence of pancreatic cancer. GFR, growth factor receptor; TGF-β, transforming growth factor-β; TbR, transforming growth factor-β receptor; TLR, Toll-like receptor; TNF-α, tumor necrosis factor-α; TNFR, tumor necrosis factor receptor
Other molecules are involved in crosstalk between inflammation and tumorigenesis. For example, LPS-TLR signaling can also activate the STAT3 (Signal transducers and activators of transcription 3) pathway and trigger mutation of the Kirsten rat sarcoma viral oncogene (KRAS), which can promote PC progression [91,92,93] (Fig. 2). Therefore, these studies have found initial links between microbiomes, inflammation and PC, suggesting that microbial-associated inflammation could play an important role in development of PC.
Microbiomes and the immune system in PC
The occurrence and development of tumors are closely related to the immune system, and the impact of microbiomes on the immune system is a very hot topic. Recently, Riquelme et al. used human-into-mice fecal microbiota transplantation experiments from short-term survival, long-term survival, or control donors, and they found modulating the tumor microbiome can affect tumor growth as well as tumor immune infiltration [15]. Different studies have highlighted the various roles of microbiomes in the immune system, including effects on immune maturation and immune suppression.
The gut microbiomes and the immune system can affect one another in the gut lamina propria, and similar effects have recently been described at extraintestinal sites [94, 95]. Round et al. found that the immune systems of germ-free mice were deficient, with hypoplastic lymphoid organs and impaired immune cells. However, the immune systems of these mice matured after gut microbiome transplantation from specific-pathogen-free mice [96]. Mechanistically, microbiomes can act as antigens and activate the immune system. Damage to the intestinal mucosa allows microbes to enter the gut lamina propria and migrate to distant lymphoid organs, resulting in activation of the immune system. However, in the absence of intestinal microflora, the immune system cannot be activated [94, 97]. Responses to gut microbiomes mediated by IL22+ innate lymphoid cells, Th17 cells and regulatory T cells occurred in mice deficient in adaptive immunity, indicating that the gut microbiome can promote innate immunity [98]. Several studies showed that specific microbes, such B. fragilis and Bifidobacterium species, may be important factors for maturation of the immune system [99,100,101]. In lung cancer, bladder cancer, kidney cancer and melanoma, similar results have been reported [102,103,104].
However, other studies showed the opposite result, finding that immune systems became activated during antibiotic treatment in models of liver cancer, colon cancer and melanoma [105,106,107]. Immune cells are essential in the PC microenvironment, where they promote tumorigenesis along with related inflammatory factors, and thus immune cell infiltration has further effects on disease progression [95, 108]. Several studies have focused on immune cell infiltration in PC. Infiltration by different leukocyte subsets can have different effects on tumorigenesis and progression, either promoting tumor growth or inhibiting tumor progression [109]. Some studies have shown that Th1-polarized CD4+ and CD8+ T cells inhibit pancreatic tumor growth in a mouse model and are associated with prolonged survival in human PC [94, 106, 110]. In contrast, antigen-specific Th2-polarized CD4+ T cells can promote progression of PC in mice [106, 111], and are associated with shorter survival in human PC [112] (Fig. 2). Another study found that FOXP3+ regulatory T cells can promote immune escape in PC [113]. However, differentiation of T cells may be regulated by microbiome composition. For example, the number of pancreas-infiltrating CD45 immune cells was reduced in antibiotic-treated NOD/SCID (Non-obese diabetic/severe combined immunodeficient) mice [107]. In KC mice (K-rasLSL.G12D; Pdx1-Cre mice) and KPC mice (K-rasLSL.G12D; p53R172H/+; Pdx1-Cre mice), myeloid-derived suppressor cell infiltration was reduced during antibiotic treatment. Moreover, Th1 polarization of CD4+ T cells and cytotoxicity of CD8+ T cells were enhanced as shown by high T-BET, tumor necrosis factor (TNF)-α and interferon-γ expression [4] (Fig. 2).
Microbe-mediated immune suppression was associated with pattern recognition receptors, and inhibition of these receptors slowed tumor development [114]. As mentioned above, following interaction with LPS, TLR4 can activate the downstream NK-κB/MAPK pathway and lead to development of PC (Fig. 2). Several TLRs (TLR2, TLR4, TLR5 and TLR7) are associated with suppression of innate and adaptive immunity to promote development of PC [4, 111, 115]. Mechanistically, activation of TLRs results in inhibition of interactions between macrophages and lymphocytes, which are abrogated in the absence of TLRs signaling [4].
These opposite results may indicate that different microbiomes play different roles in immunity or in different tumor models. It is clear that further studies are needed to target the PC-associated microbiome to enhance immunotherapy. Moreover, the composition of the gut microbiome can divide patients into responders and nonresponders for immunotherapy, which demonstrates the significance of exploring specific microbial features as diagnostic markers in PC.
Microbiomes and metabolism in PC
Microbiomes play an important role in the body, especially in the metabolism of sugars, amino acids and fats as well as synthesis of vitamins or other nutrients [116, 117]. Microbiomes cause changes in human metabolism, contributing to a variety of metabolic diseases such as obesity and diabetes. Obesity and diabetes are also important factors in the development of PC. Papamichael et al. reported that colonization by H. pylori, which is a potential independent risk factor for PC, may be associated with obesity and diabetes [118]. Therefore, microbiomes also affect the development of PC via changes in metabolism.
Obesity is a risk factor for PC in both men and women [119,120,121,122]. Obesity affects the progression of pancreatic tumors by modifying the interactions between adipocytokines [123, 124], adiponectin [125,126,127], deoxycholic acid [128], and many other molecules. Furthermore, the gut microbiome is also believed to play an important role in connecting obesity and PC. Donohoe et al. found that the body mass indices of lean mice could be increased by transplantation of gut microbiomes from obese animals, which were able to digest more nutrients [129]. Therefore, we speculate that microbiomes may participate in the occurrence and development of cancer through some metabolic mechanisms. In addition to changes in microbial diversity [130], some microbial metabolites may be associated with the development of obesity. For example, short-chain fatty acids (SCFAs), which are enriched in obese individual, can activate the MAPK signaling pathway through G-protein-coupled receptors and lead to cancer cell proliferation [86]. In addition, obesity can promote release of LPS from the gut microbiome and therefore lead to endotoxemia [131]. Ren et al. demonstrated that PC patients had more LPS-producing bacteria than healthy controls, supporting a potential relationship between endotoxemia and PC [8]. In fact, obesity is also a type of inflammatory state. As mentioned above, microbiomes can influence the development and progression of PC through different mechanisms, and the NF-κB pathway is a common pathway in both inflammation and cancer. Pagliari et al. suggested that obesity is associated with the release of various pro-inflammatory cytokines, such as IL-6 and TNF, which activate the NF-κB pathway and regulate downstream cancer-associated signals [132].
Diabetes is also a risk factor for PC. In diabetic patients, the ratio of Firmicutes to Bacteroidetes species is relatively high, affecting metabolism of carbohydrates and production of SCFAs [133]. Perry et al. demonstrated that increased levels of acetate in the blood led to insulin resistance [134], while Devaraj et al. showed that decreased levels of butyrate in the intestine promoted low-level inflammation and caused insulin resistance [135]. Decreased levels of butyrate also impaired epithelial tight junctions in the intestinal mucosa and promoted entry of bacterial endotoxins into the blood [136]. This mechanism could link diabetes and PC through endotoxemia. This effect of butyrate level was similar in individuals of different races, and metformin was able to adjust the level of butyrate effectively [137]. However, more experimental evidence is needed to confirm connections between the microbiome, diabetes and PC.
Currently, the relationship between metabolic diseases and PC has been extensively studied. However, the relationships between microbiomes and metabolic diseases are not currently sufficient to draw firm conclusions. Promisingly, we still emphasize the importance of metabolic disorders associated with microbial diversity and microbial metabolites, which are worth further exploration.
Potential clinical application of microbiomes
Microbiomes as diagnostic markers
Despite many studies suggesting an association between oral microbial dysbiosis and PC, no convincing evidence has indicated whether oral microbial dysbiosis is causally related to or merely an effect of early PC [138]. However, further studies of bacterial markers of periodontal disease such as P. gingivalis and changes in microbial diversity may suggest non-invasive screening biomarkers for PC. Recent developments suggest that salivary RNA markers can be used to identify oral bacteria by high-throughput sequencing of bacterial small subunit ribosomal RNA (16S rRNA) genes [7, 139, 140]. Therefore, saliva testing, a non-invasive test of oral biomarkers, may become a convenient strategy to screen for PC in the future. However, the existing results must be confirmed in larger multicenter prospective studies [140].
In addition, other body fluids may contain diagnostic markers of the microbiome relevant to other cancers. For example, feces can be used as a biomarker for colorectal cancer while urine may contain biomarkers of bladder cancer [141, 142]. Therefore, other body fluids such as feces, blood and pancreatic juice may also provide diagnostic markers for PC. All of these biomarkers require more study to demonstrate their potential value.
Microbiomes as therapeutic targets
In Fusobacterium-associated colorectal cancer, metronidazole treatment could reduce not only the Fusobacterium load, but also cancer cell proliferation and patient-derived xenograft tumor growth [143]. Similarly, if there are a variety of microbes located in or associated with PC, these microbes could also become future therapeutic targets for PC. In this part, we will discuss the role of microbiomes in gemcitabine therapy, PD-1 targeted therapy, and antibiotics therapy, aiming to emphasize that some microbiomes can be seen as therapeutic targets in PC.
Microbiomes and gemcitabine chemoresistance
Chemotherapy is still the first-line treatment for PC of all stages, but the treatment effect differs widely in individual patients [144]. Recent studies revealed that the microbiome played an important role in determining the efficacy and side effects of chemotherapy [145, 146]. Chemotherapy could also affect the microbiome through multiple mechanisms.
Gemcitabine (2′,2′-difluorodeoxycytidine) is a representative chemotherapy drug that is widely used for treatment of various cancers including PC. However, bacteria can metabolize gemcitabine to 2′,2′-difluorodeoxyuridine, an inactive form [144], using cytidine deaminase (CDD) [147, 148]. Using deep sequencing of bacterial 16S rDNA, Geller et al. demonstrated that most of the microbes associated with pancreatic tumors were γ-proteobacteria, including Enterobacter and Pseudomonas species [144]. These microbes can produce CDD, leading to degradation of and resistance to gemcitabine [144]. In addition to CDD, the pyrimidine nucleoside phosphorylase (PyNP) produced by mycoplasmas also has a detrimental effect on the therapeutic efficacy of chemotherapeutic drugs by indirectly potentiating deamination of these drugs [149]. The natural pyrimidine nucleosides uridine, 2′-deoxyuridine and thymidine, which can inhibit deamination of gemcitabine, were removed by PyNP [149]. Moreover, in other cancers, certain microbes could also decrease the effect of gemcitabine. For instance, in laboratory culture, Mycoplasma hyorhinis contamination led to gemcitabine resistance [150, 151]. In addition, Panos et al. found that gemcitabine incubated with E. coli supernatants became less active [152]. Thus, the combination of antibiotics and gemcitabine may represent a new strategy to increase chemosensitivity in PC patients.
However, this does not mean the use of antibiotics is without challenges. In lymphoma, colon carcinoma and melanoma, Iida et al. showed that antibiotic-treated or germ-free mice engrafted with tumors failed to respond to CpG-oligonucleotide immunotherapy and platinum chemotherapy. Moreover, antibiotic-treated mice showed downregulation of genes related to antigen presentation and adaptive immune responses but upregulation of genes related to cancer [97]. Therefore, whether antibiotics can be used in cancer combination treatment regimens, and which antibiotics should be used, requires further study.
In addition to the ability of microbes to affect the activity of gemcitabine, the drug can also perturb the microbiome [153]. Chemotherapy is harmful to the gastrointestinal mucosa, where it may have direct cytotoxic effects on cells or produce changes in the microbiomes of the gut [154, 155]. Firmicutes and Bacteroidetes species, two dominant phyla of gut microbiomes in the normal intestine, were replaced by Proteobacteria and Verrucomicrobia in gemcitabine-treated mice, leading to gut inflammation and promoting the development of PC [5, 156,157,158,159]. Another study showed that treatment with gemcitabine can promoted infection by Clostridium difficile, which was undetectable in mice that were not treated with gemcitabine [5]. In addition to the microbiome itself, some studies also found that gemcitabine produces significant changes in the metabolomic profiles associated with specific microbes [160,161,162,163]. For example, Panebianco et al. found that inosine levels were significantly reduced in mice treated with gemcitabine; the mice also developed jaundice and had increased hypoxanthine levels [5]. Inosine is a natural metabolite of adenosine with anti-inflammatory and immunosuppressive functions, which has protective effects against LPS-induced inflammation [163, 164]. Therefore, destruction of the microbiome can occur during gemcitabine therapy, leading to a vicious cycle that accelerates tumor progression.
Although some progress has been made in this area, microbe-host-drug interactions are still not fully understood. Biological complexity remains a huge obstacle to precision treatment [165]. More research is needed to understand the role of the microbiome in chemotherapy resistance in PC, which has the potential to improve its poor prognosis.
The microbiome and PD-1-targeted therapies
Immunotherapy is effective against many malignant tumors. Immune checkpoint inhibitors can upregulate T cell responses by suppressing the T cell inhibitory receptors or their ligands on tumor cells [166]. Monoclonal antibodies targeting programmed death protein 1 (PD-1) are widely used and highly effective in melanoma, non–small cell lung cancer and renal cell carcinoma [102,103,104, 167, 168]. Interestingly, resistance to anti-PD-1 therapy has been observed and microbiomes may have a non-negligible effect in this process [167,168,169]. In non–small cell lung cancer and renal cell carcinoma, Routy et al. reported the antibiotic treatment significantly inhibited the efficacy of an anti-PD-1 monoclonal antibody [102]. They used quantitative metagenomics to explore the composition of the gut microbiomes and found that patients with Akkermansia muciniphila had better prognoses. Their results suggested that T helper cell 1 and cytotoxic T lymphocyte responses were positively associated with the presence of A. muciniphila. However, Pushalkar et al. reported opposite results regarding the effect of antibiotics in PC [4]. Their results revealed that antibiotics can enhance the anti-PD-1 effect of antibodies and enhance the activation of intratumoral CD4+ and CD8+ T cells via upregulation of PD-1 in T cells. They suggested that antibiotic therapy can be combined with checkpoint-directed immunotherapy, potentially representing a new strategy for treating patients with PC [4, 102].
In summary, these studies indicated that different microbiomes may play different roles in PC tumor microenvironments. Future studies should focus on specific categories of microbes to provide a theoretical basis for combining antibiotics with PD-1 therapy.
Microbiomes and antibiotics
As mentioned above, antibiotics may increase tumor sensitivity to drugs such as anti-PD-1 antibodies and gemcitabine. However, excessive exposure to antibiotics may cause dysbacteriosis and promote tumorigenesis. For instance, male patients who used tetracycline for 4 or more years had a significantly higher risk of prostate cancer. Moreover, increased risk was observed for all antibiotic classes in studies as well as in a subgroup analysis of patients who died from breast cancer [170, 171]. According to a population-based nested case-control study including 125,441 cases and 490,510 matched controls, use of penicillin was associated with elevated risk of PC [172]. The risk increased with the number of antibiotic courses but it then diminished over time [172]. By contrast, macrolides, cephalosporins, tetracyclines, antivirals, and antifungals were not associated with increased risk of PC [172]. Moreover, antibiotic-treated mice showed downregulation of genes related to antigen presentation and adaptive immune responses but upregulation of genes related to cancer [97].
Clearly, the use of antibiotics in patients with PC represents a major challenge. Whether the use of antibiotics can potentiate other treatments or promote tumor development may depend on the composition and proportion of microbiomes.
Conclusions
PC carries a poor prognosis. Our understanding of PC has gradually advanced, and there is now some evidence that occurrence, development and therapy of PC are all related to the microbiome in vivo. The study of microbes in the pancreatic tumor microenvironment may also have potential significance for treatment of PC. The use of probiotics/antibiotics may be combined with traditional treatments such as surgery, radiotherapy and chemotherapy, as well as emerging targeted therapies and immunotherapies, to yield novel treatment options. More study is required to understand the complex relationships between the microbiome and PC.
Availability of data and materials
Not applicable.
Abbreviations
- AHL-12:
-
N-acetyl-dodecanoyl homoserine
- AP-1:
-
Activator protein 1
- Cag-A:
-
Cytotoxin-associated gene-A
- CDD:
-
Cytidine deaminase
- CI:
-
Confidence interval
- EPIC:
-
European prospective investigation into cancer and nutrition
- ERK1/2:
-
Extracellular regulated protein kinases
- GFR:
-
growth factor receptors
- HOMIM:
-
Human oral microbiological identification microarrays
- IL:
-
interleukin
- KC mice:
-
K-rasLSL.G12D; Pdx1-Cre mice
- KPC mice:
-
K-rasLSL.G12D; p53R172H/+; Pdx1-Cre mice
- KRAS:
-
Kirsten rat sarcoma viral oncogene
- LPS:
-
Lipopolysaccharide
- MAPK:
-
Mitogen-activated protein kinase
- mTOR:
-
Mammalian target of rapamycin
- NF-κB:
-
Nuclear factor kappa B
- NOD/SCID:
-
Non-obese diabetic/severe combined immunodeficient
- PC:
-
Pancreatic cancer
- PCN:
-
pancreatic cystic neoplasm
- PD-1:
-
Programmed death-1
- PD-L1:
-
Programmed death-ligand 1
- PyNP:
-
Pyrimidine nucleoside phosphorylase
- SCFA:
-
short-chain fatty acids
- STAT3:
-
Signal transducers and activators of transcription 3
- TbR:
-
transforming growth factor-β receptors
- TGF-β:
-
transforming growth factor-β
- Th1/2:
-
Helper T cell 1/2
- TLRs:
-
Toll-like receptors
- TNFR:
-
tumor necrosis factor receptor
- TNF-α:
-
tumor necrosis factor-α
- Wnt pathway:
-
Wingless / Integrated pathway
References
Siegel RL, Miller KD, Jemal A. Cancer statistics, 2019. CA Cancer J Clin. 2019;69(1):7–34.
Michaud DS, Izard J. Microbiota, oral microbiome, and pancreatic cancer. Cancer J. 2014;20(3):203–6.
Zambirinis CP, et al. Pancreatic cancer, inflammation, and microbiome. Cancer J. 2014;20(3):195–202.
Pushalkar S, et al. The pancreatic Cancer microbiome promotes Oncogenesis by induction of innate and adaptive immune suppression. Cancer Discov. 2018;8(4):403–16.
Panebianco C, et al. Influence of gemcitabine chemotherapy on the microbiota of pancreatic cancer xenografted mice. Cancer Chemother Pharmacol. 2018;81(4):773–82.
Armstrong, H., et al., The Complex Interplay between Chronic Inflammation, the Microbiome, and Cancer: Understanding Disease Progression and What We Can Do to Prevent It. Cancers (Basel), 2018. 10(3).
Torres PJ, et al. Characterization of the salivary microbiome in patients with pancreatic cancer. PeerJ. 2015;3:e1373.
Ren Z, et al. Gut microbial profile analysis by MiSeq sequencing of pancreatic carcinoma patients in China. Oncotarget. 2017;8(56):95176–91.
Olson SH, et al. The oral microbiota in patients with pancreatic cancer, patients with IPMNs, and controls: a pilot study. Cancer Causes Control. 2017;28(9):959–69.
Leal-Lopes C, et al. Roles of commensal microbiota in pancreas homeostasis and pancreatic pathologies. J Diabetes Res. 2015;2015:284680.
Farrell JJ, et al. Variations of oral microbiota are associated with pancreatic diseases including pancreatic cancer. Gut. 2012;61(4):582–8.
Mitsuhashi K, et al. Association of Fusobacterium species in pancreatic cancer tissues with molecular features and prognosis. Oncotarget. 2015;6(9):7209–20.
Bultman SJ. Emerging roles of the microbiome in cancer. Carcinogenesis. 2014;35(2):249–55.
Memba R, et al. The potential role of gut microbiota in pancreatic disease: a systematic review. Pancreatology. 2017;17(6):867–74.
Riquelme E, et al. Tumor Microbiome Diversity and Composition Influence Pancreatic Cancer Outcomes. Cell. 2019;178(4):795–806.e12.
Jia G, et al. The oral microbiota - a mechanistic role for systemic diseases. Br Dent J. 2018;224(6):447–55.
Cani, P.D. and B.F. Jordan, Gut microbiota-mediated inflammation in obesity: a link with gastrointestinal cancer. Nat Rev Gastroenterol Hepatol, 2018.
Fan X, et al. Human oral microbiome and prospective risk for pancreatic cancer: a population-based nested case-control study. Gut. 2018;67(1):120–7.
Ogrendik M. Periodontal pathogens in the etiology of pancreatic Cancer. Gastrointest Tumors. 2017;3(3–4):125–7.
Gaiser RA, et al. Enrichment of oral microbiota in early cystic precursors to invasive pancreatic cancer. Gut. 2019.
Sharif R, et al. Impact of toll-like receptor 4 on the severity of acute pancreatitis and pancreatitis-associated lung injury in mice. Gut. 2009;58(6):813–9.
Fraher MH, O'Toole PW, Quigley EM. Techniques used to characterize the gut microbiota: a guide for the clinician. Nat Rev Gastroenterol Hepatol. 2012;9(6):312–22.
Mei QX, et al. Characterization of the duodenal bacterial microbiota in patients with pancreatic head cancer vs. healthy controls. Pancreatology. 2018;18(4):438–45.
Schulte A, et al. Association between helicobacter pylori and pancreatic cancer risk: a meta-analysis. Cancer Causes Control. 2015;26(7):1027–35.
Maisonneuve P, Lowenfels AB. Risk factors for pancreatic cancer: a summary review of meta-analytical studies. Int J Epidemiol. 2015;44(1):186–98.
Guo Y, Liu W, Wu J. Helicobacter pylori infection and pancreatic cancer risk: A meta-analysis. J Cancer Res Ther. 2016;12(Supplement):C229–c232.
Yan AW, Schnabl B. Bacterial translocation and changes in the intestinal microbiome associated with alcoholic liver disease. World J Hepatol. 2012;4(4):110–8.
Ding SZ, Goldberg JB, Hatakeyama M. Helicobacter pylori infection, oncogenic pathways and epigenetic mechanisms in gastric carcinogenesis. Future Oncol. 2010;6(5):851–62.
de Martel C, et al. Helicobacter pylori infection and development of pancreatic cancer. Cancer Epidemiol Biomark Prev. 2008;17(5):1188–94.
Li S, et al. Pancreatic cyst fluid harbors a unique microbiome. Microbiome. 2017;5(1):147.
Del Castillo E, et al. The microbiomes of pancreatic and duodenum tissue overlap and are highly subject specific but differ between pancreatic Cancer and noncancer subjects. Cancer Epidemiol Biomark Prev. 2019;28(2):370–83.
Garcia-Castillo V, et al. Microbiota dysbiosis: a new piece in the understanding of the carcinogenesis puzzle. J Med Microbiol. 2016;65(12):1347–62.
Maisonneuve P, Amar S, Lowenfels AB. Periodontal disease, edentulism, and pancreatic cancer: a meta-analysis. Ann Oncol. 2017;28(5):985–95.
Meurman JH. Oral microbiota and cancer. J Oral Microbiol. 2010;2.
Michaud DS, et al. Plasma antibodies to oral bacteria and risk of pancreatic cancer in a large European prospective cohort study. Gut. 2013;62(12):1764–70.
Lu H, et al. Tongue coating microbiome data distinguish patients with pancreatic head cancer from healthy controls. J Oral Microbiol. 2019;11(1):1563409.
Li X, et al. Systemic diseases caused by oral infection. Clin Microbiol Rev. 2000;13(4):547–58.
Wang C, Li J. Pathogenic microorganisms and pancreatic Cancer. Gastrointest Tumors. 2015;2(1):41–7.
Raderer M, et al. Association between helicobacter pylori infection and pancreatic cancer. Oncology. 1998;55(1):16–9.
Stolzenberg-Solomon RZ, et al. Helicobacter pylori seropositivity as a risk factor for pancreatic cancer. J Natl Cancer Inst. 2001;93(12):937–41.
Risch HA, et al. ABO blood group, helicobacter pylori seropositivity, and risk of pancreatic cancer: a case-control study. J Natl Cancer Inst. 2010;102(7):502–5.
Yu G, et al. Seropositivity to helicobacter pylori and risk of pancreatic cancer. Cancer Epidemiol Biomark Prev. 2013;22(12):2416–9.
Trikudanathan G, et al. Association between helicobacter pylori infection and pancreatic cancer. A cumulative meta-analysis. Jop. 2011;12(1):26–31.
Xiao M, Wang Y, Gao Y. Association between helicobacter pylori infection and pancreatic cancer development: a meta-analysis. PLoS One. 2013;8(9):e75559.
Yan AW, et al. Enteric dysbiosis associated with a mouse model of alcoholic liver disease. Hepatology. 2011;53(1):96–105.
Lindkvist B, et al. A prospective study of helicobacter pylori in relation to the risk for pancreatic cancer. BMC Cancer. 2008;8:321.
Liu H, et al. Helicobacter pylori infection, atrophic gastritis, and pancreatic cancer risk: a meta-analysis of prospective epidemiologic studies. Medicine (Baltimore). 2017;96(33):e7811.
Kalaf EA, et al. Study of the cytoxin-associated gene a (CagA gene) in helicobacter pylori using gastric biopsies of Iraqi patients. Saudi J Gastroenterol. 2013;19(2):69–74.
Chen S, et al. Helicobacter pylori cytotoxin-associated gene a protein upregulates alpha-enolase expression via Src/MEK/ERK pathway: implication for progression of gastric cancer. Int J Oncol. 2014;45(2):764–70.
Ertz-Archambault N, Keim P, Von Hoff D. Microbiome and pancreatic cancer: a comprehensive topic review of literature. World J Gastroenterol. 2017;23(10):1899–908.
Risch HA, et al. Helicobacter pylori seropositivities and risk of pancreatic carcinoma. Cancer Epidemiol Biomark Prev. 2014;23(1):172–8.
Risch HA. Etiology of pancreatic cancer, with a hypothesis concerning the role of N-nitroso compounds and excess gastric acidity. J Natl Cancer Inst. 2003;95(13):948–60.
Blaser MJ. Hypotheses on the pathogenesis and natural history of helicobacter pylori-induced inflammation. Gastroenterology. 1992;102(2):720–7.
Swidsinski A, et al. Bacterial biofilm within diseased pancreatic and biliary tracts. Gut. 2005;54(3):388–95.
Huang J, et al. A prospective cohort study on poor oral hygiene and pancreatic cancer risk. Int J Cancer. 2016;138(2):340–7.
Chung LM, et al. Cancer risk in patients with candidiasis: a nationwide population-based cohort study. Oncotarget. 2017;8(38):63562–73.
Katakura Y, et al. Pancreatic involvement in chronic viral hepatitis. World J Gastroenterol. 2005;11(23):3508–13.
Jin Y, et al. Identification and impact of hepatitis B virus DNA and antigens in pancreatic cancer tissues and adjacent non-cancerous tissues. Cancer Lett. 2013;335(2):447–54.
Blander JM, et al. Regulation of inflammation by microbiota interactions with the host. Nat Immunol. 2017;18(8):851–60.
Shacter E, Weitzman SA. Chronic inflammation and cancer. Oncology (Williston Park). 2002;16(2):217–26 229; discussion 230-2.
Rubin DC, Shaker A, Levin MS. Chronic intestinal inflammation: inflammatory bowel disease and colitis-associated colon cancer. Front Immunol. 2012;3:107.
Coussens LM, Werb Z. Inflammation and cancer. Nature. 2002;420(6917):860–7.
Dejea CM, et al. Patients with familial adenomatous polyposis harbor colonic biofilms containing tumorigenic bacteria. Science. 2018;359(6375):592–7.
Grivennikov SI, et al. Adenoma-linked barrier defects and microbial products drive IL-23/IL-17-mediated tumour growth. Nature. 2012;491(7423):254–8.
Bartsch DK, Gress TM, Langer P. Familial pancreatic cancer--current knowledge. Nat Rev Gastroenterol Hepatol. 2012;9(8):445–53.
Yadav D, Lowenfels AB. The epidemiology of pancreatitis and pancreatic cancer. Gastroenterology. 2013;144(6):1252–61.
Sadr-Azodi O, et al. Pancreatic Cancer following acute pancreatitis: a population-based matched cohort study. Am J Gastroenterol. 2018;113(11):1711–9.
Kirkegard J, et al. Acute pancreatitis and pancreatic Cancer risk: a Nationwide matched-cohort study in Denmark. Gastroenterology. 2018;154(6):1729–36.
Chung SD, et al. More than 9-times increased risk for pancreatic cancer among patients with acute pancreatitis in Chinese population. Pancreas. 2012;41(1):142–6.
Kleeff J, et al. Chronic pancreatitis. Nat Rev Dis Primers. 2017;3:17060.
Redelman-Sidi G, et al. The canonical Wnt pathway drives macropinocytosis in Cancer. Cancer Res. 2018.
Francis CL, et al. Ruffles induced by Salmonella and other stimuli direct macropinocytosis of bacteria. Nature. 1993;364(6438):639–42.
Redelman-Sidi G, et al. Oncogenic activation of Pak1-dependent pathway of macropinocytosis determines BCG entry into bladder cancer cells. Cancer Res. 2013;73(3):1156–67.
Schmid SC, et al. Wntless promotes bladder cancer growth and acts synergistically as a molecular target in combination with cisplatin. Urol Oncol. 2017;35(9):544.e1–544.e10.
Terebiznik MR, et al. Elimination of host cell PtdIns(4,5)P(2) by bacterial SigD promotes membrane fission during invasion by Salmonella. Nat Cell Biol. 2002;4(10):766–73.
Clevers H, Nusse R. Wnt/beta-catenin signaling and disease. Cell. 2012;149(6):1192–205.
Redelman-Sidi G, et al. The canonical Wnt pathway drives macropinocytosis in Cancer. Cancer Res. 2018;78(16):4658–70.
Mai CW, Kang YB, Pichika MR. Should a toll-like receptor 4 (TLR-4) agonist or antagonist be designed to treat cancer? TLR-4: its expression and effects in the ten most common cancers. Onco Targets Ther. 2013;6:1573–87.
Noureldein MH, Eid AA. Gut microbiota and mTOR signaling: insight on a new pathophysiological interaction. Microb Pathog. 2018;118:98–104.
Tanti JF, et al. Implication of inflammatory signaling pathways in obesity-induced insulin resistance. Front Endocrinol (Lausanne). 2012;3:181.
Huang J, Manning BD. A complex interplay between Akt, TSC2 and the two mTOR complexes. Biochem Soc Trans. 2009;37(Pt 1):217–22.
Kisfalvi K, et al. Insulin potentiates Ca2+ signaling and phosphatidylinositol 4,5-bisphosphate hydrolysis induced by Gq protein-coupled receptor agonists through an mTOR-dependent pathway. Endocrinology. 2007;148(7):3246–57.
Liauchonak I, et al. Int J Mol Sci. 2018;19(2).
Panebianco C, et al. Engineered Resistant-Starch (ERS) Diet Shapes Colon Microbiota Profile in Parallel with the Retardation of Tumor Growth in In Vitro and In Vivo Pancreatic Cancer Models. Nutrients. 2017;9(4).
Takayama S, et al. Effects of helicobacter pylori infection on human pancreatic cancer cell line. Hepatogastroenterology. 2007;54(80):2387–91.
Pothuraju R, et al. Pancreatic cancer associated with obesity and diabetes: an alternative approach for its targeting. J Exp Clin Cancer Res. 2018;37(1):319.
Beller M, et al. Lipid droplets: a dynamic organelle moves into focus. FEBS Lett. 2010;584(11):2176–82.
Gaida MM, et al. Expression of the bitter receptor T2R38 in pancreatic cancer: localization in lipid droplets and activation by a bacteria-derived quorum-sensing molecule. Oncotarget. 2016;7(11):12623–32.
Kahle NA, et al. Bacterial quorum sensing molecule induces chemotaxis of human neutrophils via induction of p38 and leukocyte specific protein 1 (LSP1). Immunobiology. 2013;218(2):145–51.
Wu SV, et al. Expression of bitter taste receptors of the T2R family in the gastrointestinal tract and enteroendocrine STC-1 cells. Proc Natl Acad Sci U S A. 2002;99(4):2392–7.
Huang H, et al. Oncogenic K-Ras requires activation for enhanced activity. Oncogene. 2014;33(4):532–5.
Daniluk J, et al. An NF-kappaB pathway-mediated positive feedback loop amplifies Ras activity to pathological levels in mice. J Clin Invest. 2012;122(4):1519–28.
Yu H, Pardoll D, Jove R. STATs in cancer inflammation and immunity: a leading role for STAT3. Nat Rev Cancer. 2009;9(11):798–809.
Viaud S, et al. The intestinal microbiota modulates the anticancer immune effects of cyclophosphamide. Science. 2013;342(6161):971–6.
Sethi V, et al. The Role of the Microbiome in Immunologic Development and its Implication For Pancreatic Cancer Immunotherapy. Gastroenterol. 2019;156(7):2097–2115.e2.
Round JL, Mazmanian SK. The gut microbiota shapes intestinal immune responses during health and disease. Nat Rev Immunol. 2009;9(5):313–23.
Iida N, et al. Commensal bacteria control cancer response to therapy by modulating the tumor microenvironment. Science. 2013;342(6161):967–70.
Mao K, et al. Innate and adaptive lymphocytes sequentially shape the gut microbiota and lipid metabolism. Nature. 2018;554(7691):255–9.
Vetizou M, et al. Anticancer immunotherapy by CTLA-4 blockade relies on the gut microbiota. Science. 2015;350(6264):1079–84.
Sivan A, et al. Commensal Bifidobacterium promotes antitumor immunity and facilitates anti-PD-L1 efficacy. Science. 2015;350(6264):1084–9.
Mazmanian SK, et al. An immunomodulatory molecule of symbiotic bacteria directs maturation of the host immune system. Cell. 2005;122(1):107–18.
Routy B, et al. Gut microbiome influences efficacy of PD-1-based immunotherapy against epithelial tumors. Science. 2018;359(6371):91–7.
Gopalakrishnan V, et al. Gut microbiome modulates response to anti-PD-1 immunotherapy in melanoma patients. Science. 2018;359(6371):97–103.
Matson V, et al. The commensal microbiome is associated with anti-PD-1 efficacy in metastatic melanoma patients. Science. 2018;359(6371):104–8.
Ma C, et al. Gut microbiome-mediated bile acid metabolism regulates liver cancer via NKT cells. Science. 2018;360(6391).
Sethi V, et al. Gut microbiota promotes tumor growth in mice by modulating immune response. Gastroenterol. 2018.
Thomas RM, et al. Intestinal microbiota enhances pancreatic carcinogenesis in preclinical models. Carcinog. 2018.
Clark CE, et al. Dynamics of the immune reaction to pancreatic cancer from inception to invasion. Cancer Res. 2007;67(19):9518–27.
Zheng L, et al. Role of immune cells and immune-based therapies in pancreatitis and pancreatic ductal adenocarcinoma. Gastroenterol. 2013;144(6):1230–40.
Fukunaga A, et al. CD8+ tumor-infiltrating lymphocytes together with CD4+ tumor-infiltrating lymphocytes and dendritic cells improve the prognosis of patients with pancreatic adenocarcinoma. Pancreas. 2004;28(1):e26–31.
Ochi A, et al. MyD88 inhibition amplifies dendritic cell capacity to promote pancreatic carcinogenesis via Th2 cells. J Exp Med. 2012;209(9):1671–87.
De Monte L, et al. Intratumor T helper type 2 cell infiltrate correlates with cancer-associated fibroblast thymic stromal lymphopoietin production and reduced survival in pancreatic cancer. J Exp Med. 2011;208(3):469–78.
Hiraoka N, et al. Prevalence of FOXP3+ regulatory T cells increases during the progression of pancreatic ductal adenocarcinoma and its premalignant lesions. Clin Cancer Res. 2006;12(18):5423–34.
Zambirinis CP, et al. TLR9 ligation in pancreatic stellate cells promotes tumorigenesis. J Exp Med. 2015;212(12):2077–94.
Ochi A, et al. Toll-like receptor 7 regulates pancreatic carcinogenesis in mice and humans. J Clin Invest. 2012;122(11):4118–29.
Gill SR, et al. Metagenomic analysis of the human distal gut microbiome. Science. 2006;312(5778):1355–9.
Thomas S, et al. The host microbiome regulates and maintains human health: a primer and perspective for non-microbiologists. Cancer Res. 2017;77(8):1783–812.
Papamichael KX, et al. Helicobacter pylori infection and endocrine disorders: is there a link? World J Gastroenterol. 2009;15(22):2701–7.
Chakraborty S, et al. Current status of molecular markers for early detection of sporadic pancreatic cancer. Biochim Biophys Acta. 2011;1815(1):44–64.
Momi N, et al. Interplay between smoking-induced genotoxicity and altered signaling in pancreatic carcinogenesis. Carcinogenesis. 2012;33(9):1617–28.
Gumbs AA. Obesity, pancreatitis, and pancreatic cancer. Obes Surg. 2008;18(9):1183–7.
Michaud DS, et al. Physical activity, obesity, height, and the risk of pancreatic cancer. Jama. 2001;286(8):921–9.
Berger NA. Obesity and cancer pathogenesis. Ann N Y Acad Sci. 2014;1311:57–76.
Khandekar MJ, Cohen P, Spiegelman BM. Molecular mechanisms of cancer development in obesity. Nat Rev Cancer. 2011;11(12):886–95.
Huang B, et al. Adiponectin promotes pancreatic cancer progression by inhibiting apoptosis via the activation of AMPK/Sirt1/PGC-1alpha signaling. Oncotarget. 2014;5(13):4732–45.
Yang JP, et al. Association analysis of genetic variants of adiponectin gene and risk of pancreatic cancer. Int J Clin Exp Med. 2015;8(5):8094–100.
Bao Y, et al. A prospective study of plasma adiponectin and pancreatic cancer risk in five US cohorts. J Natl Cancer Inst. 2013;105(2):95–103.
Yoshimoto S, et al. Obesity-induced gut microbial metabolite promotes liver cancer through senescence secretome. Nature. 2013;499(7456):97–101.
Donohoe DR, et al. The microbiome and butyrate regulate energy metabolism and autophagy in the mammalian colon. Cell Metab. 2011;13(5):517–26.
Parekh PJ, Balart LA, Johnson DA. The influence of the gut microbiome on obesity. Metabolic Syndrome and Gastrointestinal Disease Clin Transl Gastroenterol. 2015;6:e91.
Thomas LV, Ockhuizen T, Suzuki K. Exploring the influence of the gut microbiota and probiotics on health: a symposium report. Br J Nutr. 2014;112(Suppl 1):S1–18.
Pagliari D, et al. Gut microbiota-immune system crosstalk and pancreatic disorders. Mediat Inflamm. 2018;2018:7946431.
Komaroff AL. The microbiome and risk for obesity and diabetes. Jama. 2017;317(4):355–6.
Perry RJ, et al. Acetate mediates a microbiome-brain-beta-cell axis to promote metabolic syndrome. Nature. 2016;534(7606):213–7.
Devaraj S, Hemarajata P, Versalovic J. The human gut microbiome and body metabolism: implications for obesity and diabetes. Clin Chem. 2013;59(4):617–28.
van Olden C, Groen AK, Nieuwdorp M. Role of intestinal microbiome in lipid and glucose metabolism in diabetes mellitus. Clin Ther. 2015;37(6):1172–7.
Forslund K, et al. Disentangling type 2 diabetes and metformin treatment signatures in the human gut microbiota. Nature. 2015;528(7581):262–6.
Chen J, Domingue JC, Sears CL. Microbiota dysbiosis in select human cancers: evidence of association and causality. Semin Immunol. 2017;32:25–34.
Fabryova H, Celec P. On the origin and diagnostic use of salivary RNA. Oral Dis. 2014;20(2):146–52.
Sturque, J., et al., Interest of studying the saliva metabolome, transcriptome and microbiome in screening for pancreatic cancer. J Stomatol Oral Maxillofac Surg, 2019.
Li X, et al. The expression and significance of feces cyclooxygensae-2 mRNA in colorectal cancer and colorectal adenomas. Saudi J Gastroenterol. 2017;23(1):28–33.
Gourd E. TERT mutations in urine could predict bladder cancer recurrence. Lancet Oncol. 2017;18(8):e443.
Bullman S, et al. Analysis of Fusobacterium persistence and antibiotic response in colorectal cancer. Science. 2017;358(6369):1443–8.
Geller LT, et al. Potential role of intratumor bacteria in mediating tumor resistance to the chemotherapeutic drug gemcitabine. Science. 2017;357(6356):1156–60.
Nicholson JK, Holmes E, Wilson ID. Gut microorganisms, mammalian metabolism and personalized health care. Nat Rev Microbiol. 2005;3(5):431–8.
Li H, He J, Jia W. The influence of gut microbiota on drug metabolism and toxicity. Expert Opin Drug Metab Toxicol. 2016;12(1):31–40.
Choy ATF, et al. The microbiome of pancreatic cancer: from molecular diagnostics to new therapeutic approaches to overcome chemoresistance caused by metabolic inactivation of gemcitabine. Expert Rev Mol Diagn. 2018:1–5.
Geller LT, Straussman R. Intratumoral bacteria may elicit chemoresistance by metabolizing anticancer agents. Mol Cell Oncol. 2018;5(1):e1405139.
Vande Voorde J, et al. Nucleoside-catabolizing enzymes in mycoplasma-infected tumor cell cultures compromise the cytostatic activity of the anticancer drug gemcitabine. J Biol Chem. 2014;289(19):13054–65.
Wei MY, et al. The microbiota and microbiome in pancreatic cancer: more influential than expected. Mol Cancer. 2019;18(1):97.
Olarerin-George AO, Hogenesch JB. Assessing the prevalence of mycoplasma contamination in cell culture via a survey of NCBI's RNA-seq archive. Nucleic Acids Res. 2015;43(5):2535–42.
Lehouritis P, et al. Local bacteria affect the efficacy of chemotherapeutic drugs. Sci Rep. 2015;5:14554.
Paci A, et al. Review of therapeutic drug monitoring of anticancer drugs part 1--cytotoxics. Eur J Cancer. 2014;50(12):2010–9.
Zwielehner J, et al. Changes in human fecal microbiota due to chemotherapy analyzed by TaqMan-PCR, 454 sequencing and PCR-DGGE fingerprinting. PLoS One. 2011;6(12):e28654.
Bien J, Palagani V, Bozko P. The intestinal microbiota dysbiosis and Clostridium difficile infection: is there a relationship with inflammatory bowel disease? Ther Adv Gastroenterol. 2013;6(1):53–68.
Eckburg PB, et al. Diversity of the human intestinal microbial flora. Science. 2005;308(5728):1635–8.
Turnbaugh PJ, et al. An obesity-associated gut microbiome with increased capacity for energy harvest. Nature. 2006;444(7122):1027–31.
Ley RE, et al. Obesity alters gut microbial ecology. Proc Natl Acad Sci U S A. 2005;102(31):11070–5.
Ganesh BP, et al. Commensal Akkermansia muciniphila exacerbates gut inflammation in Salmonella Typhimurium-infected gnotobiotic mice. PLoS One. 2013;8(9):e74963.
Montrose DC, et al. Celecoxib alters the intestinal microbiota and Metabolome in association with reducing polyp burden. Cancer Prev Res (Phila). 2016;9(9):721–31.
Forsgard RA, et al. Chemotherapy-induced gastrointestinal toxicity is associated with changes in serum and urine metabolome and fecal microbiota in male Sprague-Dawley rats. Cancer Chemother Pharmacol. 2017;80(2):317–32.
Daliri EB, et al. The human microbiome and metabolomics: current concepts and applications. Crit Rev Food Sci Nutr. 2017;57(16):3565–76.
da Rocha Lapa F, et al. Anti-inflammatory effects of purine nucleosides, adenosine and inosine, in a mouse model of pleurisy: evidence for the role of adenosine A2 receptors. Purinergic Signal. 2012;8(4):693–704.
Gomez G, Sitkovsky MV. Differential requirement for A2a and A3 adenosine receptors for the protective effect of inosine in vivo. Blood. 2003;102(13):4472–8.
Alexander JL, et al. Gut microbiota modulation of chemotherapy efficacy and toxicity. Nat Rev Gastroenterol Hepatol. 2017;14(6):356–65.
Pardoll D. Cancer and the immune system: basic concepts and targets for intervention. Semin Oncol. 2015;42(4):523–38.
Motzer RJ, et al. Nivolumab versus Everolimus in advanced renal-cell carcinoma. N Engl J Med. 2015;373(19):1803–13.
Borghaei H, et al. Nivolumab versus Docetaxel in advanced nonsquamous non-small-cell lung Cancer. N Engl J Med. 2015;373(17):1627–39.
Topalian SL, et al. Safety, activity, and immune correlates of anti-PD-1 antibody in cancer. N Engl J Med. 2012;366(26):2443–54.
Sutcliffe S, et al. Acne and risk of prostate cancer. Int J Cancer. 2007;121(12):2688–92.
Velicer CM, et al. Antibiotic use in relation to the risk of breast cancer. Jama. 2004;291(7):827–35.
Boursi B, et al. Recurrent antibiotic exposure may promote cancer formation--another step in understanding the role of the human microbiota? Eur J Cancer. 2015;51(17):2655–64.
Acknowledgements
Not applicable.
Funding
This study was supported by grants from the National Natural Science Foundation of China (No. 81772639; 81672443; 81672960), the Natural Science Foundation of Beijing (No. 7192157), the CAMS Innovation Fund for Medical Sciences (CIFMS) (No.2016-I2M-1-001), and the Non-profit Central Research Institute Fund of Chinese Academy of Medical Sciences (No. 2018PT32014, No. 2018PT32002).
Author information
Authors and Affiliations
Contributions
All authors contributed equally to this writing and editing of this manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
All authors agree to publish the article.
Competing interests
The authors declare no potential conflicts of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Wang, Y., Yang, G., You, L. et al. Role of the microbiome in occurrence, development and treatment of pancreatic cancer. Mol Cancer 18, 173 (2019). https://doi.org/10.1186/s12943-019-1103-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12943-019-1103-2