
Abstract
Organs of future metastasis are not passive receivers of circulating tumor cells, but are instead selectively and actively modified by the primary tumor before metastatic spread has even occurred. Tumors orchestrate a pre-metastatic program by conditioning distant organs to create microenvironments that foster the survival and proliferation of tumor cells before their arrival, thereby establishing pre-metastatic niches. Primary tumor-derived exosomes modulate these pre-metastatic niches, generating a permissive environment that facilitates the homing and expansion of tumor cells. Moreover, microRNAs have emerged as a key component of exosomal cargo, serving not only to induce the formation of pre-metastatic niches but also to prime these sites for the arrival and colonization of specific secondary tumor populations. Against this backdrop, this review endeavors to elucidate the impact of tumor-derived exosomal microRNAs on the genesis of their individualized pre-metastatic niches, with a view towards identifying novel means of specifying cancer metastasis and exploiting this phenomenon for cancer immunotherapy.
Similar content being viewed by others

Introduction
Metastasis, a terminal stage of cancer progression characterized by the aberrant migration and colonization of tumor cells in distant organs, represents a profound and debilitating hallmark of cancer pathophysiology. This phenomenon frequently culminates in suboptimal therapeutic outcomes and exacerbates patient mortality. Despite a plethora of investigations into the underlying mechanisms, the metastatic cascade remains shrouded in complexity, with numerous aspects remaining poorly understood and in need of further elucidation [1,2,3]. The successful colonization of circulating tumor cells (CTCs) in secondary or distant organs represents a pivotal step in the metastatic process. However, this process is profoundly influenced by the local microenvironment of the target organ, which can either facilitate or impede tumor cell colonization [4].
To unravel the intricate molecular mechanisms governing cancer metastasis, Chaffer and Weinberg have posited a two-stage paradigm; the initial phase encompasses the physical dissemination of malignant cells from the primary tumor to the secondary tissue microenvironment, thereby initiating the metastatic cascade. This inaugural step is followed by a subsequent phase, which involves the colonization of these disseminated cells, requiring their successful adaptation to the novel tissue context and establishment of a self-sustaining population [3].
Primary tumors have been found to modulate the microenvironment of distant organs prior to CTC arrival, thereby establishing a pre-metastatic niche that primes the subsequent colonization of disseminated tumor cells [4]. A diverse array of molecular and cellular constituents has been pinpointed as critical mediators of pre-metastatic niche formation across various tumor models. These niche-inducing molecules are secreted by tumor cells, myeloid cells, and stromal cells, functioning in concert to initiate, polarize, and establish a pre-metastatic niche in distant organs, thereby generating a permissive environment for metastatic colonization [5, 6].
The extracellular vesicles derived from the tumor microenvironment assume a pivotal role in the formation of pre-metastatic niches within specific organs. Exosomes, small membrane-bound vesicles that are part of the extracellular vesicle population, play a vital role in pre-metastatic niche formation by functioning as a means of intercellular communication between tumor cells and their surrounding microenvironment. These extracellular vesicles are capable of transferring various biomolecules, including proteins, mRNAs, microRNAs, small RNAs, and DNA fragments, to recipient cells through a process known as horizontal gene transfer. This phenomenon enables the exchange of molecular information between tumor cells and their surroundings, thereby facilitating the establishment of a pre-metastatic niche [6]. Notably, microRNAs (miRNAs) assume a pivotal role in the multifaceted process of tumor growth and metastasis, encompassing all stages of carcinogenesis. Aberrant miRNA expression patterns are a hallmark of cancer cells, with numerous cancer types exhibiting either elevated or diminished miRNA levels. Cancer-secreted exosomes also display aberrant miRNA profiles, which can contribute to the creation of a permissive environment for tumorigenesis. By modulating the expression of oncogenes and/or tumor suppressor genes in recipient cells, these miRNAs can influence cellular processes and promote tumorigenesis. This underscores the potential significance of miRNA-containing exosomes as a key mechanism by which cancer cells exert their influence on the microenvironment, ultimately facilitating the development of cancer [7].
Selective metastasis
Cancer metastasis refers to the process through that cancer cells spread from the main tumor, settle, and develop at a location beyond the primary tumor site. For this purpose, cancer cells need to detach from their primary site and enter the bloodstream or lymphatic system. Once in circulation, they must survive the rigors of travel, including the immune response and blood flow, to reach their secondary sites [8,9,10,11]. At these new sites, they must adapt to their new environment by undergoing further changes that allow them to grow and thrive [12].
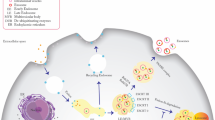
Captivatingly, most cancer types appear to have preferential locations to colonize, confirming that the spread of metastases is not random [12]. In fact, certain tissue microenvironments, known as ‘‘anti-metastatic niches,’’ are found to be inhospitable for formation of secondary tumor [4]. The metastatic localization of cancer cells is a complex process orchestrated by a multitude of pivotal mediators, including chemokines and secreted proteins. Another critical determinant of successful metastasis is the ability of metastatic cells to survive and adapt in their new environment. To enable successful engraftment, the secondary environment must undergo modifications to create a permissive microenvironment for cancer cell colonization (Fig. 1) [12]. Tumor-secreted factors, including tumor-derived extracellular vesicles and particles, circulate in the bloodstream and modulate distant organs, thereby influencing metastatic progression. Primary tumors can induce early changes in the microenvironment of secondary organs devoid of cancer cells, creating a permissive environment conducive to cancer cell settlement and metastasis initiation. This phenomenon gives rise to pre-metastatic niches (PMNs), which provide a fertile ground for the establishment of metastatic foci, preceding the arrival of cancer cells [4]. The seed and soil hypothesis, posits that cancer cells (the seeds) require a receptive microenvironment (the soil) for engraftment during metastasis [12]. It is established that the primary tumor can trigger the perpetuation of pre-metastatic niches (soil) through a process analogous to irrigation, where the tumor acts as the "watering can" that nourishes the soil, allowing it to become receptive to seed colonization.

Illustration of tumor microenvironment. Diverse range of cell types exist in tumor microenvironment, including immune cells, stem cells, and other cell subtypes that interact and influence tumor growth and progression
The PMN is shaped by intricate interactions between cancer-secreted factors and resident stromal cells at distant sites, as well as bone marrow-derived cells (BMDCs). The priming of a distant organ comprises a stepwise process that subverts tissue homeostasis, creating an environment receptive to circulating tumor cell (CTC) colonization. This multifaceted process involves the induction of vascular leakiness, lymphangiogenesis, extracellular matrix (ECM) remodeling, and the generation of an immunosuppressive microenvironment. These alterations collectively create a permissive environment that enables CTCs to establish a foothold and initiate metastatic growth [4]. It has been reported that the exosomes derived from tumor cells that are capable of migrating have the ability for interacting with blood vessels, stromal components, and immune cells to establish a pre-metastatic niche [13]. Therefore, the mechanisms by which tumor-derived exosomes sub-serve the tumor are still being investigated. Through the next section, we will delve into the most recent discoveries regarding the impact of cancer cell-derived exosomes and the miRNAs they transport, as one of the most important cargos, on selective and distant metastasis.
Role of cancer cells-derived exosomal miRNAs in selective metastasis
The metastatic potential of cancer cells is contingent upon a dynamic crosstalk between the tumor cells and their microenvironment (TME), where the tumor influences the microenvironment, which in turn shapes the behavior of cancer cells [14]. Moreover, it is conformed that the establishment of a pre-metastatic niche is orchestrated by the secretion of soluble factors from the primary tumor into the systemic circulation. Among these factors, tumor-derived exosomes (TDEs) are of particular significance, as they are small extracellular vesicles that encapsulate a diverse range of biomolecules, including RNA, DNA, proteins, metabolites, and microRNAs, derived from their parent cells [15, 16].
Liu and Cao have identified six key features of the pre-metastatic niche that facilitate tumor cell colonization and metastasis: inflammation, immunosuppression, organotropism, reprogramming, lymphangiogenesis, angiogenesis, and vascular permeability. These features collectively regulate the colonization and survival or dormancy of circulating tumor cells upon arrival in the target organ, influencing the ultimate fate of these disseminated cells [6]. Various studies have confirmed the contribution of exosomes in cancer metastasis, through their role in formation of the premetastatic niche, influencing tumor cells and TME, and determining specific organotropic metastasis [17, 18]. TDEs exhibit a predilection for homing to highly vascularized tissues, such as the lung and liver, owing to their distinctive surface integrin profiles [15]. Specifically, TDEs interact with inflammatory molecules to facilitate the formation of the PMN. Moreover, TDEs exert a multifaceted influence on the establishment of the PMN, encompassing immunosuppression and immune surveillance, promotion of angiogenesis and vascular permeability, activation of stromal cells, and remodeling of the extracellular matrix (ECM), as well as organotropic metastasis [19]. Given their ubiquity in bodily fluids and their potential as non-invasive biomarkers for cancer diagnosis, TDEs are being extensively explored as a novel therapeutic target in clinical settings [15].
There has been an increasing focus on the role of TDEs in mediating complex intratumoral communications within the tumor microenvironment [20]. TDEs have been shown to modulate tumor progression by secreting pro-inflammatory cytokines, promoting angiogenesis, triggering Toll-like receptor 3 (TLR-3)-dependent neutrophil infiltration, and recruiting myeloid-derived suppressor cells (MDSCs) [21]. The non-coding RNA cargo of TDEs, particularly microRNAs has been increasingly recognized as a critical component of the PMN. In fact, the intricate dialogue between tumor cells and the microenvironment is orchestrated by a complex interplay of signaling pathways, wherein miRNAs assume a pivotal role in shaping the pre-metastatic niche. TDEs have been shown to transfer miRNAs to surrounding cells or cells in distant metastatic niches by conditioning the pre-metastatic tumor microenvironment (TME) [14, 22]. Tumor-derived miRNAs selectively target key genes involved in immune cell suppression, angiogenesis, and EMT, thereby priming the pre-metastatic niche in distant organs (Fig. 2). This miRNA-mediated signaling cascade enables the primary tumor to establish a permissive environment for subsequent metastasis. Furthermore, organ-specific miRNA profiles play a crucial role in directing metastasis to specific tissues by regulating gene expression and modulating cellular processes [23, 24]. In fact, detection of specific miRNAs may serve as a potential biomarker for predicting or prognosticating metastasis, thereby providing a means to prevent or delay its onset [15, 19, 25, 26].

The effect of tumor microenvironment in preparation of pre-metastatic niche. Exosomal microRNA derived from tumor cells and tumor microenvironment can alter pre-metastatic niche in favor of tumor metastasis
With that context, through this section we will delve into the pivotal role of exosomal miRNAs in orchestrating organ-specific metastasis and PMN establishment across various cancer types, thereby highlighting the intricate molecular mechanisms underlying the heterogeneous patterns of tumor dissemination.
Colorectal cancer
Colorectal cancer (CRC) is the third most prevalent cancer globally. Despite advances in treatment, CRC has a significant mortality rate, with approximately 56% of patients succumbing to their disease. Notably, approximately 20% of patients present with metastatic disease at the time of diagnosis, a phenomenon that has remained relatively stable over the past two decades [27]. The most frequent sites of metastasis from CRC are the liver, lung, peritoneum, brain, and bone, with less common metastasis to the adrenal glands and spleen (Fig. 3) [28].

The effect of tumor-derived exosomes in selective metastasis. Organotropism in A colorectal cancer, B breast cancer, C lung cancer, D hepatocellular carcinoma
Exosomes derived from CRC cells can contain oncogenic miRNAs that suppress EMT inhibitors and triggers CRC metastasis [29]. For example, exosomal miR-335-5p originated from metastatic CRC cells are found to induce CRC metastasis through enhancing EMT by targeting RASA1 [30] and DLC-1 [31]. Besides, EMT-CRC-secreted exosomal miR-27b-3p triggers metastasis by inducing EMT through targeting p120 and vascular endothelial cadherin (VE-Cad) CRC cells [32]. In addition, exosomal components can be originated from or delivered to tumor cells or cells within the tumor microenvironment, elevating the metastatic activity through triggering the EMT process in tumor cells and effecting the microenvironment’s characteristic [29]. For instance, EMT-CRC-derived exosomal miR-29a enhances metastasis in endothelial cells in the TME by targeting KLF4 [33].
Furthermore, tumor-derived exosomes within serum can deliver miRNAs to distant sites and trigger metastasis. Altered regulation of exosomal miRNAs can promote metastatic cascade through upregulating EMT markers and phenotypic features of pro-metastatic cells. MiR‐221/222‐3p is an onco‐miRNA that is associated with poor prognosis in CRC individuals [34, 35]. Recently, Tian et al. [36] reported exosomal miR-221/222‐3p secreted from CRC cells enhances liver metastasis by positively increasing the expression level of hepatocyte growth factor through inhibiting SPINT1. Furthermore, exosomes originated from HCT116-TP53(R273H) cells are adsorbed by mouse embryonic fibroblasts (MEFs) and became activated. Furthermore, it has been shown that miR-21-3p and -769-3p are capable of activating fibroblasts and exerting a synergistic influence on the transforming growth factor-β (TGF-β)/Smad axis through their target genes. Overall, the mutant p53 CRC cells-derived exosomal miR-21-3p and -769-3p enhances pulmonary metastasis by activating stromal fibroblasts and premetastatic niche. In return, activated fibroblasts promotes tumor cell EMT by positively regulating TGF-β [37]. Exosomal miR-106b-3p derived from high invasive potential CRC cells contributes to lung metastasis in mice by targeting DLC-1 [31].
Immunosuppression is a critical factor for forming and developing the PMN and development of PMN, and the main contributor to tumors survival and development in vivo. plasma-derived exosome miR-203 is demonstrated to promote the differentiation of monocytes in distal organs into M2 TAMs of immunosuppressant phenotype [38]. Meanwhile, it has been established that exosomal miR-934 can induce the differentiation of normal phenotype M1 into M2 TAMs [39]. Downregulation of SOCS3 through miR-222-3p in TDEs is established to promote STAT3-mediated M2 and contribute to the immunosuppressive microenvironment [40]. Moreover, the upregulatory effect of exosomal miR-425-5p and -25-3p on M2 TAMs expression through the PI3K/AKT axis, have been demonstrated to trigger distant metastasis in CRC [41].
Noteworthy, inflammation is found to be implicated in suppression of the tumor progression as well as promotion of tumor occurrence and metastasis. For instance, inflammatory M1 TAMs are found to contribute to CRC development in colitis through their capability to have pro-inflammatory and immunostimulatory activity as well as producing anti-tumor factors, including IL-1β, IL-6 and TNF-α [42]. Hence, during tumor growth and metastasis, chronic inflammation that is the foundation of inflammatory microenvironment, can trigger the PMN formation in distant organs [43]. In fact, the aforementioned pro-inflammatory cytokines serve a crucial role in inflammatory microenvironment, which promotes tumor survival, proliferation and metastasis [43]. It has been established that low-density IL-1β can induce the local inflammatory response and result in protective immune responses, whereas high densities can trigger the inflammation-related cancer tissue damage [44]. On the other hand, during the immune response, IL-6 can activate T and B cells to perform an anti-inflammatory role [45]. It has been demonstrated that exosomal miRNAs derived from CRC tumor cells can upregulate IL-6 secretion, therefore elevating inflammatory responses [46]. In fact, high concentration of IL-6 has been identified in serum, live tumors or biopsies of cancer individuals, confirming that the inflammatory effects of this cytokine might be associated with the cancer occurrence [47]. Exosomal miR-21 can trigger the release of pro-inflammatory IL-6 and IL-21 and their presence within circulation, thereby promoting the formation of an inflammatory microenvironment [48]. More studies are shown in Tables 1 and 2.
In conclusion, the identification of specific exosomal microRNAs that promote EMT and tumor invasiveness, as well as those that contribute to immunosuppression and inflammation, may lead to the development of innovative therapeutic strategies aimed at disrupting these pathways and inhibiting CRC progression.
Breast cancer
Invasive breast cancer often exhibits a propensity for lymphatic and distant metastasis. At early-stage metastasis breast cancer, dissemination of tumor cells through both lymphatic and hematogenous systems occurs [14]. Multiple organs including lymph nodes, bone, lungs, brain, and liver are often found as receiver of the breast cancer metastasis (Fig. 3) [14]. The aberrant downregulation of miR-130a-3p has been identified in human breast cancer tissues and exosomes from circulating blood. The lower levels of exosomal miR-130a-3p are also found to be related to lymph node metastasis and advanced TNM stage [159]. In vitro study has established that exosomal miR-130a-3p can inhibit the cell proliferation, migration, and invasion of human breast cancer stem cells (BCSCs) through regulation of RAB5B/epidermal growth factor receptor signaling pathways [160]. Moreover, it has been established that exosomal miR-770 can be transmitted into tumor-associated macrophages, and subsequently increase the expression of miR-770 in macrophages [160]. In addition, upregulation of miR-770 is established to suppress the invasion and migration of Triple-negative breast cancer (TNBC) through targeting STMN1. On the other hand, Let-7a and c-Myc exhibit a negative correlation with BC. The exosomal Let-7a originated from MDA-MB-231 cell can suppress the proliferation, migration, and invasion both in vitro and in vivo through downregulating the c-Myc expression [161]. Moreover, it has been demonstrated that miR-188-5p can suppress breast cancer-cell proliferation and migration, through targeting IL-6 signal transducer (IL6ST) [162]. This data have confirmed the selective sorting of miR-188-5p into exosomes from malignant BC cells [14].
In the process of metastatic and diffusion, BC gains the capability to transmigrate through blood vessels via promoting alteration within the endothelial barrier [14]. Exosomal miR-939 in TNBC cells elevated tumor cell trans-endothelial migration and directly targeted vascular endothelial cadherin (VE-cadherin) in endothelial cells [163]. This suggests that BC-secreted exosomal miR-939 is implicated in the extracellular pro-tumorigenic characteristic and is correlated with a worse prognosis in TNBCs. MiR-105 was characteristically secreted by memory B cell (MBC) and was a potent migration regulator through targeting the tight junction protein zona occluden-1 (ZO-1) [164]. In endothelial monolayers, exosomal miR-105 secreted by BC cells disrupts the integrity of natural barriers and favors metastasis. Clinically, miR-105 have been detected in the circulation at the pre-metastatic stage, and its levels in the blood and tumor were associated with ZO-1 expression and metastasis in early-stage BC. Prior to neoadjuvant therapy, a comparative analysis of exosomal miRNA expression levels revealed significantly elevated levels of miR-21 and miR-105 in patients with metastatic breast cancer compared to those with non-metastatic disease and healthy donors [165].
MiR-155 is an oncogenic miRNA which its upregulation is commonly detected in BC and is implicated in the recurrence, metastasis, and resistance. MiR-155 is found to be abundant in cancer stem cells (CSCs) and resistant cells, and can be transmitted to BC cells through exosomes [166]. It has been demonstrated that exosomes can modify the migratory potential and enhance EMT in sensitive cells, partly through exosomal transfer of miR-155. A study by Gorczynski et al. found that both miR-155 and miR-205 play a crucial role in modulating the inflammatory response, thereby influencing the metastatic growth of BC cells in lung and liver metastasis models. Notably, elevated levels of BC exosomal miR-205 have been shown to suppress BC metastasis, whereas miR-155 has been found to have an opposite effect [167]. Through a bioinformatic analysis, Kia et al. identified that miR-9 and miR-155 were among the most highly expressed miRNAs in highly metastatic TNBC cells and their corresponding exosomes. This finding was subsequently validated through qRT-PCR experiments, providing further evidence for the potential role of these miRNAs in the biology of TNBC metastasis [83]. A luciferase assay confirmed that the miR-9 and miR-155, which were present in the exosomes of BC cells, specifically targeted the UTRs of PTEN and DUSP14 genes, respectively. In a subsequent study, when low-metastatic MCF-7 cells were treated with exosomes from highly metastatic MDA-MB-231 cells, they exhibited an enhanced metastatic phenotype, suggesting that the transfer of these exosomal miRNAs can modulate the migratory behavior of recipient cells [168]. The study revealed that the miR-155 shuttled by exosomes introduced a novel mechanism that promoted the development and metastasis of cancer. Furthermore, the researchers found that the levels of miR-7641 were elevated in the exosomes derived from BC cells and were also present in the plasma of breast cancer patients with distant metastases. Notably, this miR-7641 was found to stimulate tumor growth both in vitro and in vivo, suggesting a potential role for this miRNA in the progression of BC [169]. The study demonstrated that the miR-7641, secreted through exosomes, can promote the proliferation and invasion of BC cells. Additionally, miR-7641 can also induce epigenetic changes in recipient cells through exosome-mediated transfer. Similarly, another study found that the exosomal miR-1246, secreted from metastatic BC cells, plays a crucial role in promoting metastasis by inducing invasion in non-metastatic BC cells. Specifically, miR-1246 was shown to target the CCNG2 gene, leading to enhanced invasion capabilities in recipient cells. These findings suggest that exosomal miRNAs may play a significant role in modulating the behavior of BC cells and potentially contributing to disease progression [84].
In selective metastasis, tumor-derived exosomal miRNAs exhibit abnormal expression and play a key role in preparing the pre-metastatic niche by reprogramming the target organ, enhancing the likelihood of successful metastasis [14]. For instance, the expression of exosomal miR-19a and integrin-binding sialoprotein (IBSP) is significantly upregulated in the secretion of estrogen receptor-positive (ER +) bone-tropic breast cancer cell lines, as well as in ER + breast cancer patients with bone metastases [86]. Additionally, Wu and colleagues [86] has identified that exosomal miR-19a derived from ER + breast cancer cells enhances bone metastasis by promoting osteoclastogenesis through the targeting of PTEN. In the early metastatic niche, ER + breast cancer cells secrete integrin-binding sialoprotein (IBSP) as a chemoattractant, recruiting precursors of osteoclast (OC) cells and creating an OC precursor-enriched microenvironment. These OC precursors subsequently internalize exosomes from breast cancer cells, allowing miR-19a to be transported into the OC precursors, where it exerts its pro-tumorigenic effects [86]. Within the OC precursors, the internalized miR-19a suppresses the expression of phosphatase and tensin homolog (PTEN), activates the nuclear factor kappa B (NF-κB) and protein kinase B (AKT) signaling pathways, and promotes osteoclastogenesis. The resulting mature OC cells subsequently induce bone resorption, releasing growth factors from the bone matrix, which in turn facilitates the proliferation and survival of cancer cells [86]. Recently, Singh et al. [80] observed that miR-10b is significantly overexpressed in metastatic breast cancer cells compared to non-metastatic breast and non-malignant breast cells. Furthermore, upon internalization, miR-10b has been found to suppress the protein levels of its target genes, including homeobox D10 (HOXD10) and Krüppel-like factor 4 (KLF4), indicating its functional significance. Moreover, treatment with exosomes derived from metastatic breast cancer cells has been shown to induce metastasis by inducing invasive behavior in non-invasive breast epithelial HMLE cells through the targeting of HOXD10 [80]. More studies are shown in Tables 1 and 2.
Altogether, exosomal miRNAs have been identified as key mediators of selective metastasis, regulating gene expression, inducing EMT, and suppressing the immune response. These findings highlight the potential therapeutic significance of exosomal miRNAs as targets for the treatment of breast cancer.
Lung cancer
The majority of lung carcinomas are initially diagnosed at an advanced stage IV, often with widespread metastatic disease. Notably, lung carcinomas exhibit a propensity for metastasis via both lymphatic and hematogenous routes, reflecting their ability to spread through lymphatic vessels and the bloodstream [170]. The most common sites of metastasis for lung cancer are proposed to be the central nervous system, bone, liver, respiratory tract, and adrenal glands (Fig. 3) [171]. Recent studies have identified that certain exosomal miRNAs play a pivotal role in promoting lung cancer cell migration, invasion, tumor growth, and metastasis through multiple mechanisms [172, 173]. Notably, the dysregulation of these miRNAs can be targeted to counteract their oncogenic effects, such as through gene silencing strategies. Runx3 (Runt-related transcription factor 3) has been found to be downregulated in lung cancer tissues, and it is considered a tumor suppressor due to its ability to antagonize the activation of the Wnt signaling pathway [174]. Exosomes derived from non-small cell lung cancer (NSCLC) cells, which contain miR-210 (miR-210-Exo), have been shown to modulate the behavior of recipient lung cancer cells by inhibiting Runx3 expression and activating the PI3K/Akt signaling pathway. This results in enhanced proliferation, migration, and invasion of the recipient cells [175]. In vivo, exosomal miR-210 was found to target the Runx3/PI3K/Akt axis, thereby promoting the growth of transplanted tumors in nude mice. Consistently, silencing of miR-210 gene expression significantly attenuated the carcinogenic effect induced by exosomal miR-210, underscoring the potential therapeutic relevance of targeting this axis in cancer treatment [175].
Epitopic overexpression of miRNA-30a-5p in lung cancer cells was found to significantly inhibit cell proliferation, migration, and invasion by targeting the cell-cycle phase regulator cyclin E2. Notably, miR-30a-5p-containing exosomes produced by vascular endothelial cells from lung adenocarcinoma (LUAD) patients were also found to effectively suppress cell proliferation, migration, and invasion, highlighting the potential therapeutic utility of miR-30a-5p-Exo in LUAD treatment [176]. HEY-like protein (HEYL) is a member of the hairy and enhancer of the split-related (HESR) family, and functions as a downstream target molecule of the Notch signaling pathway. This pathway has been implicated in the regulation of cell growth and metastasis in lung cancer (LC), suggesting that HEYL may also play a role in these processes [177, 178].
A comparison of exosomes derived from benign pleural effusion (BPE) and malignant pleural effusion (MPE) in lung cancer (LC) patients revealed that the level of miR-665 is significantly higher in MPE-derived exosomes [179]. Both cultured cells and experiments in zebrafish have confirmed that exosomes enriched with lncRNA SCIRT and miR-665 can enhance the migratory and invasive abilities of lung cancer cells by specifically targeting and suppressing HEYL. Furthermore, these findings indicated that the plasma concentrations of miR-665 and lncRNA SCIRT are significantly elevated in patients with metastatic lung cancer compared to those with non-cancerous diseases or non-metastatic lung cancer, suggesting that these exosomal biomarkers may serve as valuable indicators for the diagnosis and monitoring of metastatic disease [179]. Therefore, exosomal miR-665 in serum or lung pleural effusion may serve as a specific marker for the early diagnosis of lung cancer metastasis. Additionally, FOXP4, a member of the human forkhead-box (FOX) family, has been found to play a key role in cell cycle regulation and tumorigenesis. Notably, it has been established that FOXP4 protein and mRNA levels are significantly elevated in NSCLCs compared to normal lung tissue, indicating that FOXP4 may be a potential biomarker for LC diagnosis [180]. Chen and colleagues demonstrated that exosomal miR-3180-3p derived from NSCLC cells can exert anti-tumor effects by suppressing cell proliferation, migration, and invasion in recipient NSCLC cells through the downregulation of FOXP4 expression. Furthermore, the in vivo administration of miR-3180-3p-exosomes was shown to impede the growth and metastatic potential of NSCLC xenografts in nude mice [153].
The androgen receptor (AR) is a member of the steroid hormone receptor family and acts as a nuclear transcription factor. When bound to a ligand, AR undergoes a conformational change, allowing it to translocate to the nucleus, where it regulates the transcription of genes responsive to AR signaling. Notably, male NSCLC patients who exhibit androgen pathway manipulation (APM) have been found to have a survival advantage, suggesting a potential therapeutic benefit from targeting the androgen axis in this patient population [181]. Female NSCLC patients who have higher levels of (AR) expression have a substantially better overall survival compared to those without AR expression [182]. This suggests that the relationship between steroid hormones and their receptors in lung cancer patient survival is complex and warrants further investigation. Recent research by Zhou and colleagues has made progress in this area, revealing that miR-224-5p is overexpressed in cancer tissues from NSCLC patients and cell lines. Additionally, the study found that miR-224-5p, specifically in the form of extracellular vesicle-derived miR-224-5p, is produced by lung cancer cells and exerts oncogenic effects, promoting metastasis and cell proliferation in both NSCLC cells and normal human lung cells. These findings suggest a potential role for miR-224-5p in the progression of lung cancer, highlighting the need for further research to elucidate the underlying mechanisms [183]. The study also demonstrated that the overexpression of miR-224-5p in NSCLC cells led to a suppression of AR expression. This repression of AR had a profound impact on the behavior of the cancer cells, as it promoted EMT, proliferation, migration, invasion, and resistance to apoptosis. Furthermore, this overexpression also drove the growth of lung cancer xenografts. Conversely, silencing the AR gene in NSCLC cells enhanced their migratory potential and increased their resistance to apoptosis. These findings suggest that miR-224-5p plays a key role in regulating the progression of lung cancer by targeting the AR pathway, which may have important implications for the development of novel therapeutic strategies [183]. This suggests that miR-224-5p-Exo can promote NSCLC progression by directly targeting AR. More studies are shown in Table 2.
Overall, these studies suggests that exosomal miRNAs play a pivotal role in the selective metastasis of lung cancer by facilitating the colonization of specific organs. Once internalized, these exosomal miRNAs reprogram recipient cells to promote their own proliferation, migration, and invasion, thereby creating a conducive microenvironment for metastatic lesion establishment. The targeting of specific organs by exosomal miRNAs is influenced by the expression profile of specific receptors or ligands on recipient cell surfaces, allowing lung cancer cells to selectively colonize particular organs and tissues. These findings underscore the significance of exosomal miRNAs in mediating selective metastasis in lung cancer and imply potential therapeutic strategies aimed at targeting these molecules to prevent or treat this disease.
Hepatocellular carcinoma
Hepatocellular carcinoma (HCC) is a prevalent cancer type, ranking sixth in terms of incidence and third in terms of mortality globally. Notably, lung metastasis is the most frequent extrhepatic manifestation of HCC, underscoring the importance of understanding the mechanisms underlying this phenomenon [184]. In addition, bone metastases (BM) are a common phenomenon in patients with metastatic HCC, affecting 2–25% of cases. The presence of BM is often associated with a poor prognosis in HCC, highlighting the need for more effective diagnostic and therapeutic strategies to address this complication [185]. It has been demonstrated that exosomal miRNAs derived from HCC cells can play a significant role in promoting lung and bone metastasis. For instance, liver cancer-derived exosomal miR-574-5p has been shown to exacerbate bone metastasis by enhancing osteoclastogenesis through direct targeting of the BMP2 gene [102]. Furthermore, exosomal miR-1247-3p derived from high-metastatic HCC cells has been found to contribute to the conversion of fibroblasts to cancer-associated fibroblasts (CAFs) by directly targeting the B4GALT3 gene. In turn, CAFs have been shown to enhance lung metastasis by promoting EMT and stemness, indicating a critical role for exosomal miRNAs in the crosstalk between cancer cells and stromal cells during metastatic progression [103].
Exosomes released by HCC cells have also been found to influence the metastatic behavior of tumor cells. Research suggests that exosomal miR-21 and exosomal miR-10b, which are induced by acidic microenvironments, promote the proliferation and metastasis of cancer cells. As such, these exosomal miRNAs may serve as potential prognostic molecular markers and therapeutic targets for HCC [101]. In addition, exosomal miR-125b has been found to exert anti-tumor effects by inhibiting the metastasis of HCC cells. Specifically, exosomal miR-125b has been shown to suppress EMT by targeting the SMAD2 gene, which is a key regulator of EMT [154]. Interestingly, exosomal miR-29a-3p released by high-metastatic HCC cells has been found to promote metastasis by enhancing EMT in low-metastatic HCC cells. This pro-metastatic effect is mediated by exosomal miR-29a-3p targeting the phosphatase and tensin homolog (PTEN) gene, a key regulator of EMT. This finding suggests that exosomal miR-29a-3p may act as a "metastatic bridge" between high-metastatic and low-metastatic HCC cells, promoting the dissemination of cancer cells through EMT activation [100].
Blood-borne metastasis, which refers to the spread of cancer cells through the bloodstream, is the primary cause of mortality in patients with cancer. The process of metastasis is facilitated by increased vascular permeability, which allows cancer cells to extravasate from the bloodstream and colonize distant organs. This highlights the critical role of the vasculature in promoting tumor dissemination and underscores the importance of understanding the molecular mechanisms underlying vascular permeability in cancer progression. Furthermore, targeting these mechanisms may provide a promising therapeutic strategy for inhibiting metastasis and improving patient outcomes [186]. A study by Fang et al. [92] demonstrated that exosomes secreted by HCC cells can transfer miR-103 to endothelial cells, thereby inhibiting the expression of key proteins involved in endothelial tight junction integrity, such as human vascular endothelial cadherin, zonula occludens 1, and p120-catenin. This led to a weakening of endothelial connectivity, increased vascular permeability, and accelerated metastasis. In a xenograft mouse model, high miR-103 expression was associated with an increased probability of intrahepatic and pulmonary metastasis. Notably, HCC patients with elevated serum miR-103 levels exhibited a higher metastatic potential compared to those with low miR-103 expression levels. These findings collectively suggest a positive correlation between exosomal miR-103 expression and the metastatic capacity of HCC [92, 187].
The mitogen-activated protein kinases (MAPKs) constitute a family of evolutionarily conserved serine/threonine protein kinases that play crucial roles in various cellular processes, including cell proliferation, differentiation, motility, and apoptosis [188]. The MAPK family, comprising p44/42 (ERK1/2), p46/p54 (JNK), and p38, serves as a critical component of protein kinase cascades, which are essential for regulating cell growth and differentiation, as well as modulating cellular responses to cytokines and stress signals [189]. The p44/42 MAPK (ERK1/2) signal transduction pathway is activated in response to a diverse range of extracellular stimuli, including mitogens, growth factors, and cytokines [190, 191], and it is an important target for cancer diagnosis and treatment [192]. Recent studies have demonstrated that exosomal miR-320a exhibits anti-tumor effects on HCC cells by suppressing cell proliferation, migration, and metastasis through the inhibition of the MAPK pathway, which ultimately leads to the induction of EMT and the upregulation of cyclin-dependent kinase 2 (CDK2) and matrix metallopeptidase 2 (MMP2) expression [135]. In the study conducted by Li Xiong et al., mast cells have been shown to inhibit the ERK1/2 pathway by delivering exosomal miR-490 to HCC cells, thereby suppressing the metastatic potential of HCC cells [158]. Activation of MAPK signaling pathway is a frequently observed event in the progression and metastasis of tumors, highlighting its significance as a potential therapeutic target for cancer treatment [193, 194]. These studies can provide new insights into the regulatory mechanism of HCC in the MAPK signaling pathway and identify potential ways of the therapeutic intervention for the disease.
Overall, these findings highlight the potential for exosomal miRNAs to serve as key mediators of metastatic spread in HCC, and underscores the importance of further investigating their mechanisms of actions.
Head and neck cancer
Head and neck cancer (HNC) is a prevalent and debilitating neoplasm globally, with a significant burden on public health. Despite ongoing advances in therapeutic modalities, the 5-year overall survival rate for advanced HNC remains disappointingly low, hovering around 50%, underscoring the pressing need for innovative treatment strategies to improve patient outcomes [195,196,197,198]. HNC often originates from mucosal surfaces, specifically the oral cavity, which includes the tongue, lip, buccal mucosa, gingiva, and palate, as well as the oropharynx, larynx, and perioral skin, highlighting the importance of early detection and surveillance in these high-risk areas [199]. The majority of HNCs, exceeding 90%, are classified as head and neck squamous cell carcinomas (HNSCCs). Recent studies have uncovered a wealth of evidence indicating that tumor-derived exosomal miRNAs play a pivotal role in the oncogenic process, facilitating intercellular communication and signal transduction pathways that contribute to tumor development, progression, and treatment resistance [200, 201]. For instance, research has demonstrated that exosomes secreted by hypoxic oral squamous cell carcinoma (OSCC) cells transfer viral miR-21 to normoxic cells, inducing EMT and subsequently promoting cell migration and invasion [202]. Notably, research has shown that exosomal miR-21 derived from esophageal carcinoma cell line EC9706 promotes metastasis in esophageal cancer by enhancing migration and invasion through targeting the protein programmed cell death 4 (PDCD4) [115]. It has been demonstrated that miR-23b-3p derived from salivary adenoid cystic carcinoma (SACC) cells and packaged in exosomes can contribute to SACC metastasis by exacerbating angiogenesis through direct targeting of the tumor suppressor phosphatase and tensin homolog (PTEN) [111]. In addition, miR-34a-5p in CAF-derived exosomes in OSCC can stimulate the proliferation and metastasis of oral cancer cells by activating the AKT/glycogen synthase kinase-3 beta/β-catenin/Snail signaling cascade [136]. A recent investigation has shown that extreme metastatic oral squamous cell carcinoma cells secrete exosomes containing miR-1246 and -342-3p, which enhance the oncogenic growth, metastasis, and invasion of recipient cells [110]. The transformation of poor metastatic cells into aggressive metastatic cells is facilitated by the downregulation of the Multiple Acyl-CoA Dehydrogenase Deficiency (MADD)/DENN domain containing protein 2D (DENN2D) via the influence of exosomal miR-1246. Additionally, exosomes containing miR-21 exhibit increased expression of mesenchymal markers, such as vimentin and snail, and decreased expression of E-cadherin. These findings collectively suggest that OSCC malignant cell clusters undergo EMT and migrate to distant organs [202,203,204].
Lymphangiogenesis, the process of forming new lymphatic vessels, has been recognized as a novel prognostic indicator for predicting the risk of lymph node metastasis (LNM) [205]. During the progression of lymphatic metastasis, lymphatic vessels at the tumor periphery function as conduits for the dissemination of tumor cells to regional lymph nodes (LNs) [206]. Several cytokines, including vascular endothelial growth factor C (VEGF-C) and VEGF-D, engage with vascular endothelial growth factor receptor-3 (VEGFR-3) to stimulate the proliferation, tube formation, and migration of lymphatic endothelial cells (LECs) within the tumor microenvironment [207]. In various experimental tumor models, inhibition of the VEGF-C/VEGFR-3 signaling pathway was found to significantly reduce LNM by approximately 60–70% [208, 209], the effectiveness and side effects of these drugs still need to be carefully evaluatedNotably, some patients with esophageal squamous cell carcinoma (ESCC) with LNM display low levels of VEGF-C. Consequently, the identification of novel lymphangiogenesis regulators is crucial. Recent studies have revealed that miR-10527-5p, which is derived from ESCC-derived exosomes, exhibits potent inhibitory effects on the migration, invasion, and EMT of ESCC cells as well as the migration and tube formation of human lymphatic endothelial cells. This inhibitory activity prevents lymphatic metastasis and lymphangiogenesis of ESCC through the Wnt/β-catenin signaling pathway by directly targeting Rab10 [155]. In contrast, ESCC-derived exosomal miR-320b has been found to induce the metastasis of human lymphatic endothelial cells (HLECs) and lymphatic metastasis by promoting EMT through direct targeting of PDCD4 [114]. More studies about the role of exosomal miRNAs derived from other types of tumor cells are shown in Table 2.
Conclusion
The propensity of cancer cells to colonize specific organs and tissues, a phenomenon known as organ-specific metastasis, is a complex process governed by a delicate interplay of genetic and environmental factors. Tumor-derived exosomes exert a pivotal role in the orchestration of organ-specific metastasis by facilitating the transfer of a distinct repertoire of pro-metastatic cargo between cancer cells and the target organ microenvironment. Exosomes derived from primary tumors exhibit a selective enrichment for specific biomolecules, including transcriptional regulators, growth factors, and adhesion molecules, which are instrumental in inducing a pre-metastatic niche in recipient cells, thereby enabling them to undergo EMT and adapt to the foreign microenvironment. The selective loading of exosomes with pro-metastatic molecules enables cancer cells to "pre-condition" the target organ microenvironment, thereby increasing their likelihood of successful colonization and growth.
Exosomal miRNAs have been demonstrated to exert a profound impact on the selective metastatic process, facilitating the establishment of metastatic niches by modulating gene expression through selective targeting of recipient cells. Exosomal miRNAs orchestrate the preparation of the pre-metastatic niche, a complex microenvironment that facilitates the homing and colonization of cancer cells at distant sites. By regulating the expression of genes involved in inflammation, angiogenesis, and immunosuppression, exosomal miRNAs create a permissive microenvironment that fosters a pro-tumorigenic landscape conducive to metastasis. Conversely, these miRNAs also repress the expression of genes involved in immune surveillance and tissue repair, thereby generating an environment that enables cancer cells to establish a foothold and thrive. This dual function of exosomal miRNAs underscores their pivotal role in shaping the microenvironmental conditions that enable cancer cell metastasis, making them attractive targets for therapeutic exploitation [29, 210]. Furthermore, these exosomal miRNAs have been identified as potential biomarkers for cancer diagnosis and monitoring, owing to their unique profiles and dynamic expression patterns. In fact, the TDEs emerge as a prime candidate for the development of prognostic biomarkers for PMNs, boasting a trifecta of attributes that render them particularly well-suited for this purpose: their remarkable stability, widespread presence in bodily fluids, non-invasive accessibility, and tumor-specific expression patterns [211]. However, a striking disparity is observed in the research landscape regarding the role of exosomal microRNAs in selective metastasis across distinct cancer types. For instance, while their involvement in lung and breast cancer metastasis has been extensively explored, analogous investigations remain scarce for other tumor types, such as pancreatic or ovarian cancer. This observation underscores the need for comprehensive and organ-specific studies to elucidate the underlying mechanisms driving exosomal microRNA-mediated selective metastasis in each cancer type. By deciphering these mechanisms, researchers may uncover novel therapeutic targets and develop more effective strategies for preventing and treating cancer metastasis, ultimately improving patient outcomes.
Over the past three decades, groundbreaking discoveries in the elucidation of cancer metastasis have unveiled a plethora of novel targets for preventing this insidious process. Notably, significant strides have been made in modulating the biochemical pathways and signaling cascades governing cell adhesion, dissociation, migration, invasion, and the complex interactions between cancer cells and the tumor microenvironment (TME). These advances have significantly expanded our understanding of the intricate mechanisms underlying cancer dissemination, thereby providing a rich source of opportunities for therapeutic interventions aimed at thwarting the metastatic process [9]. The development of exosomal miRNA-based therapeutics holds great promise, with potential treatments focused on inhibiting the transfer of these molecules to prevent reprogramming of recipient cells. Elucidating the specific receptors and ligands involved in targeting will enable the design of targeted therapies to block this interaction. Moreover, exosomal miRNAs may serve as non-invasive biomarkers for early detection of metastasis, revolutionizing diagnostic capabilities. The exploration of combination therapies targeting multiple components will also be crucial in combating this complex disease. Ultimately, a deeper understanding of exosome biology will inform the development of novel therapeutic approaches, paving the way for improved patient outcomes and a more effective treatment landscape for cancer.
Availability of data and materials
Not applicable.
Data availability
No datasets were generated or analysed during the current study.
References
Fares J, Fares MY, Khachfe HH, Salhab HA, Fares Y. Molecular principles of metastasis: a hallmark of cancer revisited. Signal Transduct Target Ther. 2020;5(1):28.
Gerstberger S, Jiang Q, Ganesh K. Metastasis Cell. 2023;186(8):1564–79.
Chaffer CL, Weinberg RA. A perspective on cancer cell metastasis. Science. 2011;331(6024):1559–64.
Patras L, Shaashua L, Matei I, Lyden D. Immune determinants of the pre-metastatic niche. Cancer Cell. 2023;41(3):546–72.
Sceneay J, Smyth MJ, Möller A. The pre-metastatic niche: finding common ground. Cancer Metastasis Rev. 2013;32(3):449–64.
Liu Y, Cao X. Characteristics and significance of the pre-metastatic niche. Cancer Cell. 2016;30(5):668–81.
Chan S-H, Wang L-H. Regulation of cancer metastasis by microRNAs. J Biomed Sci. 2015;22(1):9.
Seyfried TN, Huysentruyt LC. On the origin of cancer metastasis. Crit Rev Oncog. 2013;18(1-2):43–73. https://doi.org/10.1615/critrevoncog.v18.i1-2.40.
Guan X. Cancer metastases: challenges and opportunities. Acta pharmaceutica sinica B. 2015;5(5):402–18.
Brabletz T, Kalluri R, Nieto MA, Weinberg RA. EMT in cancer. Nat Rev Cancer. 2018;18(2):128–34.
Weber GF. Why does cancer therapy lack effective anti-metastasis drugs? Cancer Lett. 2013;328(2):207–11.
Janssen LME, Ramsay EE, Logsdon CD, Overwijk WW. The immune system in cancer metastasis: friend or foe? J Immunother Cancer. 2017;5(1):79.
Whiteside TL. Tumor-Derived Exosomes and Their Role in Cancer Progression. Adv Clin Chem. 2016;74:103–41.
Tan Y, Luo X, Lv W, Hu W, Zhao C, Xiong M, et al. Tumor-derived exosomal components: the multifaceted roles and mechanisms in breast cancer metastasis. Cell Death Dis. 2021;12(6):547.
Morrissey SM, Zhang F, Ding C, Montoya-Durango DE, Hu X, Yang C, et al. Tumor-derived exosomes drive immunosuppressive macrophages in a pre-metastatic niche through glycolytic dominant metabolic reprogramming. Cell Metab. 2021;33(10):2040–58. e10.
Mirzaei H, Rahimian N, Mirzaei HR, Nahand JS, Hamblin MR. Exosomes. In: Mirzaei H, Rahimian N, Mirzaei HR, Nahand JS, Hamblin MR, editors. Exosomes and MicroRNAs in Biomedical Science. Cham: Springer International Publishing; 2022. p. 79–92.
Tian W, Liu S, Li B. Potential Role of Exosomes in Cancer Metastasis. Biomed Res Int. 2019;2019:4649705.
Jiang C, Zhang N, Hu X, Wang H. Tumor-associated exosomes promote lung cancer metastasis through multiple mechanisms. Mol Cancer. 2021;20(1):117.
Chen X, Feng J, Chen W, Shao S, Chen L, Wan H. Small extracellular vesicles: from promoting pre-metastatic niche formation to therapeutic strategies in breast cancer. Cell Communication and Signaling. 2022;20(1):141.
Li I, Nabet BY. Exosomes in the tumor microenvironment as mediators of cancer therapy resistance. Mol Cancer. 2019;18(1):32.
Bardi GT, Smith MA, Hood JL. Melanoma exosomes promote mixed M1 and M2 macrophage polarization. Cytokine. 2018;105:63–72.
Mirzaei H, Rahimian N, Mirzaei HR, Nahand JS, Hamblin MR. Exosomes and Cancer. In: Mirzaei H, Rahimian N, Mirzaei HR, Nahand JS, Hamblin MR, editors. Exosomes and MicroRNAs in Biomedical Science. Cham: Springer International Publishing; 2022. p. 93–113.
Mirzaei H, Rahimian N, Mirzaei HR, Nahand JS, Hamblin MR. MicroRNAs in Cancer. In: Mirzaei H, Rahimian N, Mirzaei HR, Nahand JS, Hamblin MR, editors. Exosomes and MicroRNAs in Biomedical Science. Cham: Springer International Publishing; 2022. p. 11–40.
Mirzaei H, Rahimian N, Mirzaei HR, Nahand JS, Hamblin MR. Exosomes and microRNAs in biomedical science. Morgan & Claypool Publishers; 2022
Chin AR, Wang SE. Cancer tills the premetastatic field: mechanistic basis and clinical implications. Clin Cancer Res. 2016;22(15):3725–33.
Mirzaei H, Rahimian N, Mirzaei HR, Nahand JS, Hamblin MR. MicroRNAs as Biomarkers. In: Mirzaei H, Rahimian N, Mirzaei HR, Nahand JS, Hamblin MR, editors. Exosomes and MicroRNAs in Biomedical Science. Cham: Springer International Publishing; 2022. p. 69–77.
Riihimäki M, Hemminki A, Sundquist J, Hemminki K. Patterns of metastasis in colon and rectal cancer. Sci Rep. 2016;6(1):29765.
Vatandoust S, Price TJ, Karapetis CS. Colorectal cancer: Metastases to a single organ. World J Gastroenterol. 2015;21(41):11767–76.
Danac JMC, Uy AGG, Garcia RL. Exosomal microRNAs in colorectal cancer: Overcoming barriers of the metastatic cascade (Review). Int J Mol Med. 2021;47(6):112. https://doi.org/10.3892/ijmm.2021.4945.
Sun X, Lin F, Sun W, Zhu W, Fang D, Luo L, et al. Exosome-transmitted miRNA-335-5p promotes colorectal cancer invasion and metastasis by facilitating EMT via targeting RASA1. Molecular Therapy-Nucleic Acids. 2021;24:164–74.
Liu H, Liu Y, Sun P, Leng K, Xu Y, Mei L, et al. Colorectal cancer-derived exosomal miR-106b-3p promotes metastasis by down-regulating DLC-1 expression. Clin Sci. 2020;134(4):419–34.
Dou R, Liu K, Yang C, Zheng J, Shi D, Lin X, et al. EMT-cancer cells-derived exosomal miR-27b-3p promotes circulating tumour cells-mediated metastasis by modulating vascular permeability in colorectal cancer. Clin Transl Med. 2021;11(12):e595.
Liu K, Dou R, Yang C, Di Z, Shi D, Zhang C, et al. Exosome-transmitted miR-29a induces colorectal cancer metastasis by destroying the vascular endothelial barrier. Carcinogenesis. 2023;44(4):356–67.
Yau TO, Wu CW, Dong Y, Tang CM, Ng SS, Chan FK, et al. microRNA-221 and microRNA-18a identification in stool as potential biomarkers for the non-invasive diagnosis of colorectal carcinoma. Br J Cancer. 2014;111(9):1765–71.
Liu S, Sun X, Wang M, Hou Y, Zhan Y, Jiang Y, et al. A microRNA 221- and 222-mediated feedback loop maintains constitutive activation of NFκB and STAT3 in colorectal cancer cells. Gastroenterology. 2014;147(4):847-59.e11.
Tian F, Wang P, Lin D, Dai J, Liu Q, Guan Y, et al. Exosome-delivered miR-221/222 exacerbates tumor liver metastasis by targeting SPINT1 in colorectal cancer. Cancer Sci. 2021;112(9):3744–55.
Ju Q, Zhao L, Gao J, Zhou L, Xu Y, Sun Y, et al. Mutant p53 increases exosome-mediated transfer of miR-21–3p and miR-769–3p to promote pulmonary metastasis. Chin J Cancer Res. 2019;31(3):533–46.
Takano Y, Masuda T, Iinuma H, Yamaguchi R, Sato K, Tobo T, et al. Circulating exosomal microRNA-203 is associated with metastasis possibly via inducing tumor-associated macrophages in colorectal cancer. Oncotarget. 2017;8(45):78598–613.
Zhao S, Mi Y, Guan B, Zheng B, Wei P, Gu Y, et al. Tumor-derived exosomal miR-934 induces macrophage M2 polarization to promote liver metastasis of colorectal cancer. J Hematol Oncol. 2020;13(1):156.
Ying X, Wu Q, Wu X, Zhu Q, Wang X, Jiang L, et al. Epithelial ovarian cancer-secreted exosomal miR-222-3p induces polarization of tumor-associated macrophages. Oncotarget. 2016;7(28):43076–87.
Wang D, Wang X, Si M, Yang J, Sun S, Wu H, et al. Exosome-encapsulated miRNAs contribute to CXCL12/CXCR4-induced liver metastasis of colorectal cancer by enhancing M2 polarization of macrophages. Cancer Lett. 2020;474:36–52.
Zhu W, Yu J, Nie Y, Shi X, Liu Y, Li F, et al. Disequilibrium of M1 and M2 macrophages correlates with the development of experimental inflammatory bowel diseases. Immunol Invest. 2014;43(7):638–52.
Guo Y, Ji X, Liu J, Fan D, Zhou Q, Chen C, et al. Effects of exosomes on pre-metastatic niche formation in tumors. Mol Cancer. 2019;18(1):39.
Lust JA, Lacy MQ, Zeldenrust SR, Dispenzieri A, Gertz MA, Witzig TE, et al. Induction of a chronic disease state in patients with smoldering or indolent multiple myeloma by targeting interleukin 1{beta}-induced interleukin 6 production and the myeloma proliferative component. Mayo Clin Proc. 2009;84(2):114–22.
Hong DS, Angelo LS, Kurzrock R. Interleukin-6 and its receptor in cancer: implications for translational therapeutics. Cancer. 2007;110(9):1911–28.
Pucci M, Raimondo S, Urzì O, Moschetti M, Di Bella MA, Conigliaro A, Caccamo N, La Manna MP, Fontana S, Alessandro R. Tumor-Derived Small Extracellular Vesicles Induce Pro-Inflammatory Cytokine Expression and PD-L1 Regulation in M0 Macrophages via IL-6/STAT3 and TLR4 Signaling Pathways. Int J Mol Sci. 2021;22(22):12118. https://doi.org/10.3390/ijms222212118.
Guillerey C, Nakamura K, Vuckovic S, Hill GR, Smyth MJ. Immune responses in multiple myeloma: role of the natural immune surveillance and potential of immunotherapies. Cellular and molecular life sciences : CMLS. 2016;73(8):1569–89.
Shi C, Yang Y, Xia Y, Okugawa Y, Yang J, Liang Y, et al. Novel evidence for an oncogenic role of microRNA-21 in colitis-associated colorectal cancer. Gut. 2016;65(9):1470–81.
Chen B, Sang Y, Song X, Zhang D, Wang L, Zhao W, et al. Exosomal miR-500a-5p derived from cancer-associated fibroblasts promotes breast cancer cell proliferation and metastasis through targeting USP28. Theranostics. 2021;11(8):3932–47.
Yan Z, Sheng Z, Zheng Y, Feng R, Xiao Q, Shi L, et al. Cancer-associated fibroblast-derived exosomal miR-18b promotes breast cancer invasion and metastasis by regulating TCEAL7. Cell Death Dis. 2021;12(12):1120.
Liu Y, Yang Y, Du J, Lin D, Li F. MiR-3613-3p from carcinoma-associated fibroblasts exosomes promoted breast cancer cell proliferation and metastasis by regulating SOCS2 expression. IUBMB Life. 2020;72(8):1705–14.
Wu H-J, Hao M, Yeo SK, Guan J-L. FAK signaling in cancer-associated fibroblasts promotes breast cancer cell migration and metastasis by exosomal miRNAs-mediated intercellular communication. Oncogene. 2020;39(12):2539–49.
Guo L, Li B, Yang J, Shen J, Ji J, Miao M. Fibroblast-derived exosomal microRNA-369 potentiates migration and invasion of lung squamous cell carcinoma cells via NF1-mediated MAPK signaling pathway. Int J Mol Med. 2020;46(2):595–608.
Wang S, Du P, Cao Y, Ma J, Yang X, Yu Z, et al. Cancer associated fibroblasts secreted exosomal miR-1290 contributes to prostate cancer cell growth and metastasis via targeting GSK3β. Cell Death Discovery. 2022;8(1):371.
Hu JL, Wang W, Lan XL, Zeng ZC, Liang YS, Yan YR, et al. CAFs secreted exosomes promote metastasis and chemotherapy resistance by enhancing cell stemness and epithelial-mesenchymal transition in colorectal cancer. Mol Cancer. 2019;18(1):91.
Zhang X, Sai B, Wang F, Wang L, Wang Y, Zheng L, et al. Hypoxic BMSC-derived exosomal miRNAs promote metastasis of lung cancer cells via STAT3-induced EMT. Mol Cancer. 2019;18(1):40.
Wang G, Ji X, Li P, Wang W. Human bone marrow mesenchymal stem cell-derived exosomes containing microRNA-425 promote migration, invasion and lung metastasis by down-regulating CPEB1. Regenerative Ther. 2022;20:107–16.
Liu W, Wang B, Duan A, Shen K, Zhang Q, Tang X, et al. Exosomal transfer of miR-769-5p promotes osteosarcoma proliferation and metastasis by targeting DUSP16. Cancer Cell Int. 2021;21(1):541.
Du Q, Ye X, Lu SR, Li H, Liu HY, Zhai Q, et al. Exosomal miR-30a and miR-222 derived from colon cancer mesenchymal stem cells promote the tumorigenicity of colon cancer through targeting MIA3. J Gastrointest Oncol. 2021;12(1):52–68.
Liu W, Long Q, Zhang W, Zeng D, Hu B, Liu S, et al. miRNA-221-3p derived from M2-polarized tumor-associated macrophage exosomes aggravates the growth and metastasis of osteosarcoma through SOCS3/JAK2/STAT3 axis. Aging. 2021;13(15):19760–75.
Yin Z, Ma T, Huang B, Lin L, Zhou Y, Yan J, et al. Macrophage-derived exosomal microRNA-501-3p promotes progression of pancreatic ductal adenocarcinoma through the TGFBR3-mediated TGF-β signaling pathway. J Exp Clin Cancer Res. 2019;38(1):310.
Wang Z, Zhang C, Guo J, Wang W, Si Q, Chen C, et al. Exosomal miRNA-223-3p derived from tumor associated macrophages promotes pulmonary metastasis of breast cancer 4T1 cells. Transl Oncol. 2023;35.
Chen Y, Lei Y, Li J, Wang X, Li G. Macrophage-derived exosomal microRNAs promote metastasis in pancreatic ductal adenocarcinoma. Int Immunopharmacol. 2024;129: 111590.
Li X, Chen Z, Ni Y, Bian C, Huang J, Chen L, et al. Tumor-associated macrophages secret exosomal miR-155 and miR-196a-5p to promote metastasis of non-small-cell lung cancer. Transl Lung Cancer Res. 2021;10(3):1338–54.
Zhang Z, Hu J, Ishihara M, Sharrow AC, Flora K, He Y, et al. The miRNA-21-5p Payload in Exosomes from M2 Macrophages Drives Tumor Cell Aggression via PTEN/Akt Signaling in Renal Cell Carcinoma. Int J Mol Sci. 2022;23(6):3005.
Wu L, Xue M, Lai S, Chen J, Lin Y, Ding N, et al. Hypoxia derived exosomes promote the proliferation and metastasis of colorectal cancer through the regulation of HIF-1α/miR-4299/ZBTB4. Life Sci. 2023;329: 121872.
Yu Y, Min Z, Zhou z, Linhong M, Tao R, Yan L, et al. Hypoxia-induced exosomes promote hepatocellular carcinoma proliferation and metastasis via miR-1273f transfer. Experiment Cell Res. 2019;385(1):111649.
Xia X, Wang S, Ni B, Xing S, Cao H, Zhang Z, et al. Correction to: Hypoxic gastric cancer-derived exosomes promote progression and metastasis via MiR-301a-3p/PHD3/HIF-1α positive feedback loop. Oncogene. 2021;40(41):6058.
Wang X, Luo G, Zhang K, Cao J, Huang C, Jiang T, et al. Hypoxic tumor-derived exosomal miR-301a mediates M2 macrophage polarization via PTEN/PI3Kγ to promote pancreatic cancer metastasis. Can Res. 2018;78(16):4586–98.
Xie L, Zhang K, You B, Yin H, Zhang P, Shan Y, et al. Hypoxic nasopharyngeal carcinoma-derived exosomal miR-455 increases vascular permeability by targeting ZO-1 to promote metastasis. Mol Carcinog. 2023;62(6):803–19.
Zhang Y, Qian K, Liu X, Zhao X, Zhao T, Lu G. Exosomal mir-625-3p derived from hypoxic lung cancer cells facilitates metastasis by targeting SCAI. Mol Biol Rep. 2022;49(10):9275–81.
Kim DH, Park S, Kim H, Choi YJ, Kim SY, Sung KJ, et al. Tumor-derived exosomal miR-619–5p promotes tumor angiogenesis and metastasis through the inhibition of RCAN1.4. Cancer Letters. 2020;475:2–13.
Yu F, Liang M, Huang Y, Wu W, Zheng B, Chen C. Hypoxic tumor-derived exosomal miR-31-5p promotes lung adenocarcinoma metastasis by negatively regulating SATB2-reversed EMT and activating MEK/ERK signaling. J Exp Clin Cancer Res. 2021;40(1):179.
Yang C, Dou R, Wei C, Liu K, Shi D, Zhang C, et al. Tumor-derived exosomal microRNA-106b-5p activates EMT-cancer cell and M2-subtype TAM interaction to facilitate CRC metastasis. Mol Ther. 2021;29(6):2088–107.
Pei W, Wei K, Wu Y, Qiu Q, Zhu H, Mao L, et al. Colorectal cancer tumor cell-derived exosomal miR-203a-3p promotes CRC metastasis by targeting PTEN-induced macrophage polarization. Gene. 2023;885: 147692.
Wang D, Wang X, Si M, Yang J, Sun S, Wu H, et al. Corrigendum to “Exosome-encapsulated miRNAs contribute to CXCL12/CXCR4-induced liver metastasis of colorectal cancer by enhancing M2 polarization of macrophages” [Canc. Lett. 474 (2020) 36–52]. Cancer Lett. 2022;525:200–2.
Zeng Z, Li Y, Pan Y, Lan X, Song F, Sun J, et al. Cancer-derived exosomal miR-25-3p promotes pre-metastatic niche formation by inducing vascular permeability and angiogenesis. Nat Commun. 2018;9(1):5395.
Wang D, Wang X, Song Y, Si M, Sun Y, Liu X, et al. Exosomal miR-146a-5p and miR-155-5p promote CXCL12/CXCR7-induced metastasis of colorectal cancer by crosstalk with cancer-associated fibroblasts. Cell Death Dis. 2022;13(4):380.
Gu P, Sun M, Li L, Yang Y, Jiang Z, Ge Y, et al. Breast tumor-derived exosomal microRNA-200b-3p promotes specific organ metastasis through regulating CCL2 expression in lung epithelial cells. Front Cell Dev Biol. 2021;9.
Singh R, Pochampally R, Watabe K, Lu Z, Mo Y-Y. Exosome-mediated transfer of miR-10b promotes cell invasion in breast cancer. Mol Cancer. 2014;13(1):256.
Ding J, Xu Z, Zhang Y, Tan C, Hu W, Wang M, et al. Exosome-mediated miR-222 transferring: An insight into NF-κB-mediated breast cancer metastasis. Exp Cell Res. 2018;369(1):129–38.
Yang S-S, Ma S, Dou H, Liu F, Zhang S-Y, Jiang C, et al. Breast cancer-derived exosomes regulate cell invasion and metastasis in breast cancer via miR-146a to activate cancer associated fibroblasts in tumor microenvironment. Exp Cell Res. 2020;391(2): 111983.
Kia V, Paryan M, Mortazavi Y, Biglari A, Mohammadi-Yeganeh S. Evaluation of exosomal miR-9 and miR-155 targeting PTEN and DUSP14 in highly metastatic breast cancer and their effect on low metastatic cells. J Cell Biochem. 2019;120(4):5666–76.
Li XJ, Ren ZJ, Tang JH, Yu Q. Exosomal MicroRNA MiR-1246 promotes cell proliferation, invasion and drug resistance by targeting CCNG2 in breast cancer. Cell Physiol Biochem. 2018;44(5):1741–8.
Xing F, Liu Y, Wu S-Y, Wu K, Sharma S, Mo Y-Y, et al. Loss of XIST in breast cancer activates MSN-c-Met and reprograms microglia via exosomal miRNA to promote brain metastasis. Can Res. 2018;78(15):4316–30.
Wu K, Feng J, Lyu F, Xing F, Sharma S, Liu Y, et al. Exosomal miR-19a and IBSP cooperate to induce osteolytic bone metastasis of estrogen receptor-positive breast cancer. Nat Commun. 2021;12(1):5196.
Wang B, Mao J-h, Wang B-y, Wang L-x, Wen H-y, Xu L-j, et al. Exosomal miR-1910–3p promotes proliferation, metastasis, and autophagy of breast cancer cells by targeting MTMR3 and activating the NF-κB signaling pathway. Cancer Lett. 2020;489:87–99.
Wang J, Zhang Q, Wang D, Yang S, Zhou S, Xu H, et al. Microenvironment-induced TIMP2 loss by cancer-secreted exosomal miR-4443 promotes liver metastasis of breast cancer. J Cell Physiol. 2020;235(7–8):5722–35.
Sun L, He M, Xu N, Xu D-H, Ben-David Y, Yang Z-Y, et al. Regulation of RAB22A by mir-193b inhibits breast cancer growth and metastasis mediated by exosomes. Int J Oncol. 2018;53(6):2705–14.
Wang L, He J, Hu H, Tu L, Sun Z, Liu Y, et al. Lung CSC-derived exosomal miR-210-3p contributes to a pro-metastatic phenotype in lung cancer by targeting FGFRL1. J Cell Mol Med. 2020;24(11):6324–39.
Tiong T-Y, Chan M-l, Wang C-H, Yadav VK, Pikatan NW, Fong I-H, et al. Exosomal miR-21 determines lung-to-brain metastasis specificity through the DGKB/ERK axis within the tumor microenvironment. Life Sci 2023;329:121945.
Fang JH, Zhang ZJ, Shang LR, Luo YW, Lin YF, Yuan Y, et al. Hepatoma cell-secreted exosomal microRNA-103 increases vascular permeability and promotes metastasis by targeting junction proteins. Hepatology. 2018;68(4):1459–75.
He S, Li Z, Yu Y, Zeng Q, Cheng Y, Ji W, et al. Exosomal miR-499a-5p promotes cell proliferation, migration and EMT via mTOR signaling pathway in lung adenocarcinoma. Exp Cell Res. 2019;379(2):203–13.
Mao S, Zheng S, Lu Z, Wang X, Wang Y, Zhang G, et al. Exosomal miR-375-3p breaks vascular barrier and promotes small cell lung cancer metastasis by targeting claudin-1. Transl Lung Cancer Res. 2021;10(7):3155–72.
Kim DH, Park H, Choi YJ, Kang M-H, Kim T-K, Pack C-G, et al. Exosomal miR-1260b derived from non-small cell lung cancer promotes tumor metastasis through the inhibition of HIPK2. Cell Death Dis. 2021;12(8):747.
Ma Z, Wei K, Yang F, Guo Z, Pan C, He Y, et al. Tumor-derived exosomal miR-3157-3p promotes angiogenesis, vascular permeability and metastasis by targeting TIMP/KLF2 in non-small cell lung cancer. Cell Death Dis. 2021;12(9):840.
Chen J, Zhang K, Zhi Y, Wu Y, Chen B, Bai J, et al. Tumor-derived exosomal miR-19b-3p facilitates M2 macrophage polarization and exosomal LINC00273 secretion to promote lung adenocarcinoma metastasis via Hippo pathway. Clin Transl Med. 2021;11(9): e478.
Chang R-M, Fu Y, Zeng J, Zhu X-Y, Gao Y. Cancer-derived exosomal miR-197-3p confers angiogenesis via targeting TIMP2/3 in lung adenocarcinoma metastasis. Cell Death Dis. 2022;13(12):1032.
Yang X, Yu F, Huang G, Ni Y, Zhang T, Zou Z, et al. Exosomal miR-133a-3p promotes the growth and metastasis of lung cancer cells following incomplete microwave ablation. Int J Hyperth. 2023;40(1):2190065.
Yang B, Feng X, Liu H, Tong R, Wu J, Li C, et al. High-metastatic cancer cells derived exosomal miR92a-3p promotes epithelial-mesenchymal transition and metastasis of low-metastatic cancer cells by regulating PTEN/Akt pathway in hepatocellular carcinoma. Oncogene. 2020;39(42):6529–43.
Tian X-P, Wang C-Y, Jin X-H, Li M, Wang F-W, Huang W-J, et al. Acidic microenvironment up-regulates exosomal miR-21 and miR-10b in early-stage hepatocellular carcinoma to promote cancer cell proliferation and metastasis. Theranostics. 2019;9(7):1965.
Li T, Li M, Peng Y, Li X, Chen Y, Zhang J, et al. Liver cancer cell-secreted exosomes promote bone metastasis of liver cancer by facilitating osteoclast differentiation through the miR-574–5p/BMP2 axis. Pathol Res Pract. 2023;245:154485.
Fang T, Lv H, Lv G, Li T, Wang C, Han Q, et al. Tumor-derived exosomal miR-1247-3p induces cancer-associated fibroblast activation to foster lung metastasis of liver cancer. Nat Commun. 2018;9(1):191.
Lin X-M, Wang Z-J, Lin Y-X, Chen H. Decreased exosome-delivered miR-486-5p is responsible for the peritoneal metastasis of gastric cancer cells by promoting EMT progress. World J Surg Oncol. 2021;19(1):312.
Feng C, She J, Chen X, Zhang Q, Zhang X, Wang Y, et al. Exosomal miR-196a-1 promotes gastric cancer cell invasion and metastasis by targeting SFRP1. Nanomedicine. 2019;14(19):2579–93.
Yang H, Fu H, Wang B, Zhang X, Mao J, Li X, et al. Exosomal miR-423-5p targets SUFU to promote cancer growth and metastasis and serves as a novel marker for gastric cancer. Mol Carcinog. 2018;57(9):1223–36.
Li Q, Li B, Li Q, Wei S, He Z, Huang X, et al. Exosomal miR-21-5p derived from gastric cancer promotes peritoneal metastasis via mesothelial-to-mesenchymal transition. Cell Death Dis. 2018;9(9):854.
Qiu S, Xie L, Lu C, Gu C, Xia Y, Lv J, et al. Gastric cancer-derived exosomal miR-519a-3p promotes liver metastasis by inducing intrahepatic M2-like macrophage-mediated angiogenesis. J Exp Clin Cancer Res. 2022;41(1):296.
Zhu M, Zhang N, He S, Lu X. Exosomal miR-106a derived from gastric cancer promotes peritoneal metastasis via direct regulation of Smad7. Cell cycle (Georgetown, Tex). 2020;19(10):1200–21.
Sakha S, Muramatsu T, Ueda K, Inazawa J. Exosomal microRNA miR-1246 induces cell motility and invasion through the regulation of DENND2D in oral squamous cell carcinoma. Sci Rep. 2016;6(1):38750.
Yang W, Tan S, Yang L, Chen X, Yang R, Oyang L, et al. Exosomal miR-205-5p enhances angiogenesis and nasopharyngeal carcinoma metastasis by targeting desmocollin-2. Molecular Therapy-Oncolytics. 2022;24:612–23.
Zhong Q, Nie Q, Wu R, Huang Y. Exosomal miR-18a-5p promotes EMT and metastasis of NPC cells via targeting BTG3 and activating the Wnt/β-catenin signaling pathway. Cell cycle (Georgetown, Tex). 2023;22(13):1544–62.
Zhu Y, Li F, Wan Y, Liang H, Li S, Peng B, Shao L, Xu Y, Jiang D. Cancer-Secreted Exosomal MiR-620 Inhibits ESCC Aerobic Glycolysis via FOXM1/HER2 Pathway and Promotes Metastasis. Front Oncol. 2022;12:756109. https://doi.org/10.3389/fonc.2022.756109.
Liu T, Li P, Li J, Qi Q, Sun Z, Shi S, et al. Exosomal and intracellular miR-320b promotes lymphatic metastasis in esophageal squamous cell carcinoma. Molecular Therapy-Oncolytics. 2021;23:163–80.
Liao J, Liu R, Shi Y-J, Yin L-H, Pu Y-P. Exosome-shuttling microRNA-21 promotes cell migration and invasion-targeting PDCD4 in esophageal cancer. Int J Oncol. 2016;48(6):2567–79.
Ye Y, Li SL, Ma YY, Diao YJ, Yang L, Su MQ, et al. Exosomal miR-141-3p regulates osteoblast activity to promote the osteoblastic metastasis of prostate cancer. Oncotarget. 2017;8(55):94834–49.
Liu Y, Yang C, Chen S, Liu W, Liang J, He S, et al. Cancer-derived exosomal miR-375 targets DIP2C and promotes osteoblastic metastasis and prostate cancer progression by regulating the Wnt signaling pathway. Cancer Gene Ther. 2023;30(3):437–49.
Dai Y, Gao X. Inhibition of cancer cell-derived exosomal microRNA-183 suppresses cell growth and metastasis in prostate cancer by upregulating TPM1. Cancer Cell Int. 2021;21:1–15.
Zhao Q, Chen H, Li X, Zeng B, Sun Z, Liu D, et al. Low-metastatic melanoma cells acquire enhanced metastatic capability via exosomal transfer of miR-199a-1-5p from highly metastatic melanoma cells. Cell Death Discov. 2022;8(1):188.
Chen H, Zeng B, Li X, Zhao Q, Liu D, Chen Y, Zhang Y, Wang J, Xing HR. High-Metastatic Melanoma Cells Promote the Metastatic Capability of Low-Metastatic Melanoma Cells via Exosomal Transfer of miR-411-5p. Front Oncol. 2022;12:895164. https://doi.org/10.3389/fonc.2022.895164.
Liu D, Li X, Zeng B, Zhao Q, Chen H, Zhang Y, et al. Exosomal microRNA-4535 of melanoma stem cells promotes metastasis by inhibiting autophagy pathway. Stem Cell Rev Rep. 2023;19(1):155–69.
Wu M, Tan X, Liu P, Yang Y, Huang Y, Liu X, et al. Role of exosomal microRNA-125b-5p in conferring the metastatic phenotype among pancreatic cancer cells with different potential of metastasis. Life Sci. 2020;255: 117857.
Wu J. Pancreatic Cancer-Derived Exosomes Promote the Proliferation, Invasion, and Metastasis of Pancreatic Cancer by the miR-3960/TFAP2A Axis. J Oncol. 2022;2022:3590326.
Deng Z, Rong Y, Teng Y, Zhuang X, Samykutty A, Mu J, et al. Exosomes miR-126a released from MDSC induced by DOX treatment promotes lung metastasis. Oncogene. 2017;36(5):639–51.
He L, Zhu W, Chen Q, Yuan Y, Wang Y, Wang J, et al. Ovarian cancer cell-secreted exosomal miR-205 promotes metastasis by inducing angiogenesis. Theranostics. 2019;9(26):8206–20.
Mo Y, Leung LL, Mak CSL, Wang X, Chan W-S, Hui LMN, et al. Tumor-secreted exosomal miR-141 activates tumor-stroma interactions and controls premetastatic niche formation in ovarian cancer metastasis. Mol Cancer. 2023;22(1):4.
You X, Wang Y, Meng J, Han S, Liu L, Sun Y, et al. Exosomal miR-663b exposed to TGF-β1 promotes cervical cancer metastasis and epithelial-mesenchymal transition by targeting MGAT3. Oncol Rep. 2021;45(4):12.
Zhou C-F, Ma J, Huang L, Yi H-Y, Zhang Y-M, Wu X-G, et al. Cervical squamous cell carcinoma-secreted exosomal miR-221-3p promotes lymphangiogenesis and lymphatic metastasis by targeting VASH1. Oncogene. 2019;38(8):1256–68.
Cai Q, Zhu A, Gong L. Exosomes of glioma cells deliver miR-148a to promote proliferation and metastasis of glioblastoma via targeting CADM1. Bull Cancer. 2018;105(7):643–51.
Gong L, Bao Q, Hu C, Wang J, Zhou Q, Wei L, et al. Exosomal miR-675 from metastatic osteosarcoma promotes cell migration and invasion by targeting CALN1. Biochem Biophys Res Commun. 2018;500(2):170–6.
Zhang Y, Tedja R, Millman M, Wong T, Fox A, Chehade H, et al. Adipose-derived exosomal miR-421 targets CBX7 and promotes metastatic potential in ovarian cancer cells. J Ovarian Res. 2023;16(1):233.
Cai J, Gong L, Li G, Guo J, Yi X, Wang Z. Exosomes in ovarian cancer ascites promote epithelial–mesenchymal transition of ovarian cancer cells by delivery of miR-6780b-5p. Cell Death Dis. 2021;12(2):210.
Tyagi A, Wu S-Y, Sharma S, Wu K, Zhao D, Deshpande R, et al. Exosomal miR-4466 from nicotine-activated neutrophils promotes tumor cell stemness and metabolism in lung cancer metastasis. Oncogene. 2022;41(22):3079–92.
Han Q, Tan S, Gong L, Li G, Wu Q, Chen L, et al. Omental cancer-associated fibroblast-derived exosomes with low microRNA-29c-3p promote ovarian cancer peritoneal metastasis. Cancer Sci. 2023;114(5):1929–42.
Zhang Z, Li X, Sun W, Yue S, Yang J, Li J, et al. Loss of exosomal miR-320a from cancer-associated fibroblasts contributes to HCC proliferation and metastasis. Cancer Lett. 2017;397:33–42.
Li Y-y, Tao Y-w, Gao S, Li P, Zheng J-m, Zhang S-e, et al. Cancer-associated fibroblasts contribute to oral cancer cells proliferation and metastasis via exosome-mediated paracrine miR-34a-5p. EBioMedicine. 2018;36:209–20.
Li BL, Lu W, Qu JJ, Ye L, Du GQ, Wan XP. Loss of exosomal miR-148b from cancer-associated fibroblasts promotes endometrial cancer cell invasion and cancer metastasis. J Cell Physiol. 2019;234(3):2943–53.
Zhang Y, Zhao J, Ding M, Su Y, Cui D, Jiang C, et al. Loss of exosomal miR-146a-5p from cancer-associated fibroblasts after androgen deprivation therapy contributes to prostate cancer metastasis. J Exp Clin Cancer Res. 2020;39(1):282.
Xu G, Zhang B, Ye J, Cao S, Shi J, Zhao Y, et al. Exosomal miRNA-139 in cancer-associated fibroblasts inhibits gastric cancer progression by repressing MMP11 expression. Int J Biol Sci. 2019;15(11):2320–9.
Li T, Wan Y, Su Z, Li J, Han M, Zhou C. Mesenchymal Stem Cell-Derived Exosomal microRNA-3940-5p Inhibits Colorectal Cancer Metastasis by Targeting Integrin α6. Dig Dis Sci. 2021;66(6):1916–27.
Ono M, Kosaka N, Tominaga N, Yoshioka Y, Takeshita F, Takahashi RU, et al. Exosomes from bone marrow mesenchymal stem cells contain a microRNA that promotes dormancy in metastatic breast cancer cells. Sci Signal. 2014;7(332):ra63.
Jahangiri B, Khalaj-Kondori M, Asadollahi E, Purrafee Dizaj L, Sadeghizadeh M. MSC-Derived exosomes suppress colorectal cancer cell proliferation and metastasis via miR-100/mTOR/miR-143 pathway. Int J Pharm. 2022;627: 122214.
Jing L, Hua X, Yuanna D, Rukun Z, Junjun M. Exosomal miR-499a-5p Inhibits Endometrial Cancer Growth and Metastasis via Targeting VAV3. Cancer Manage Res. 2020;12(null):13541–52.
Yu S, Zhou Y, Niu L, Qiao Y, Yan Y. Mesenchymal stem cell-derived exosome mir-342-3p inhibits metastasis and chemo-resistance of breast cancer through regulating ID4. Genes Genom. 2022;44(5):539–50.
Shang D, Liu Y, Chen Z. Exosome-Transmitted miR-128 Targets CCL18 to Inhibit the Proliferation and Metastasis of Urothelial Carcinoma. Front Mol Biosci. 2022;8:760748. https://doi.org/10.3389/fmolb.2021.760748.
Xie H, Wang J. MicroRNA-320a-containing exosomes from human umbilical cord mesenchymal stem cells curtail proliferation and metastasis in lung cancer by binding to SOX4. J Recept Sign Transduct. 2022;42(3):268–78.
Lian XY, Zhang H, Liu Q, Lu X, Zhou P, He SQ, et al. Ovarian cancer-excreted exosomal miR-199a-5p suppresses tumor metastasis by targeting hypoxia-inducible factor-2α in hypoxia microenvironment. Cancer Commun (London, England). 2020;40(8):380–5.
Wang X, Qian T, Bao S, Zhao H, Chen H, Xing Z, et al. Circulating exosomal miR-363-5p inhibits lymph node metastasis by downregulating PDGFB and serves as a potential noninvasive biomarker for breast cancer. Mol Oncol. 2021;15(9):2466–79.
Song B, Hou G, Xu M, Chen M. Exosomal miR-122-3p represses the growth and metastasis of MCF-7/ADR cells by targeting GRK4-mediated activation of the Wnt/β-catenin pathway. Cell Signal. 2024;117: 111101.
Wei L, Wang G, Yang C, Zhang Y, Chen Y, Zhong C, et al. MicroRNA-550a-3-5p controls the brain metastasis of lung cancer by directly targeting YAP1. Cancer Cell Int. 2021;21(1):491.
Xu S, Zheng L, Kang L, Xu H, Gao L. microRNA-let-7e in serum-derived exosomes inhibits the metastasis of non-small-cell lung cancer in a SUV39H2/LSD1/CDH1-dependent manner. Cancer Gene Ther. 2021;28(3):250–64.
Tian W, Yang X, Yang H, Lv M, Sun X, Zhou B. Exosomal miR-338-3p suppresses non-small-cell lung cancer cells metastasis by inhibiting CHL1 through the MAPK signaling pathway. Cell Death Dis. 2021;12(11):1030.
Chen T, Liu Y, Chen J, Zheng H, Chen Q, Zhao J. Exosomal miR-3180-3p inhibits proliferation and metastasis of non-small cell lung cancer by downregulating FOXP4. Thorac Cancer. 2021;12(3):372–81.
Kim HS, Kim JS, Park NR, Nam H, Sung PS, Bae SH, Choi JY, Yoon SK, Hur W, Jang JW. Exosomal miR-125b Exerts Anti-Metastatic Properties and Predicts Early Metastasis of Hepatocellular Carcinoma. Front Oncol. 2021;11:637247. https://doi.org/10.3389/fonc.2021.637247.
Xiao Z, Feng X, Zhou Y, Li P, Luo J, Zhang W, et al. Exosomal miR-10527–5p Inhibits Migration, Invasion, Lymphangiogenesis and Lymphatic Metastasis by Affecting Wnt/β-Catenin Signaling via Rab10 in Esophageal Squamous Cell Carcinoma. International Journal of Nanomedicine. 2023;18(null):95–114.
Li M, Zhou J, Zhang Z, Li J, Wang F, Ma L, et al. Exosomal miR-485-3p derived from pancreatic ductal epithelial cells inhibits pancreatic cancer metastasis through targeting PAK1. Chin Med J. 2022;135(19):2326–37.
Hu Y, Li D, Wu A, Qiu X, Di W, Huang L, et al. TWEAK-stimulated macrophages inhibit metastasis of epithelial ovarian cancer via exosomal shuttling of microRNA. Cancer Lett. 2017;393:60–7.
Xiong L, Zhen S, Yu Q, Gong Z. HCV-E2 inhibits hepatocellular carcinoma metastasis by stimulating mast cells to secrete exosomal shuttle microRNAs. Oncol Lett. 2017;14(2):2141–6.
Kong X, Zhang J, Li J, Shao J, Fang L. MiR-130a-3p inhibits migration and invasion by regulating RAB5B in human breast cancer stem cell-like cells. Biochem Biophys Res Commun. 2018;501(2):486–93.
Li Y, Liang Y, Sang Y, Song X, Zhang H, Liu Y, et al. MiR-770 suppresses the chemo-resistance and metastasis of triple negative breast cancer via direct targeting of STMN1. Cell Death Dis. 2018;9(1):14.
Du J, Fan JJ, Dong C, Li HT, Ma BL. Inhibition effect of exosomes-mediated Let-7a on the development and metastasis of triple negative breast cancer by down-regulating the expression of c-Myc. Eur Rev Med Pharmacol Sci. 2019;23(12):5301-5314. https://doi.org/10.26355/eurrev_201906_18197.
Wang M, Zhang H, Yang F, Qiu R, Zhao X, Gong Z, et al. miR-188-5p suppresses cellular proliferation and migration via IL6ST: a potential noninvasive diagnostic biomarker for breast cancer. J Cell Physiol. 2020;235(5):4890–901.
Di Modica M, Regondi V, Sandri M, Iorio MV, Zanetti A, Tagliabue E, et al. Breast cancer-secreted miR-939 downregulates VE-cadherin and destroys the barrier function of endothelial monolayers. Cancer Lett. 2017;384:94–100.
Zhou W, Fong MY, Min Y, Somlo G, Liu L, Palomares MR, et al. Cancer-secreted miR-105 destroys vascular endothelial barriers to promote metastasis. Cancer Cell. 2014;25(4):501–15.
Rodríguez-Martínez A, de Miguel-Pérez D, Ortega FG, García-Puche JL, Robles-Fernández I, Exposito J, et al. Exosomal miRNA profile as complementary tool in the diagnostic and prediction of treatment response in localized breast cancer under neoadjuvant chemotherapy. Breast Cancer Res. 2019;21:1–9.
Santos JC, Lima NdS, Sarian LO, Matheu A, Ribeiro ML, Derchain SFM. Exosome-mediated breast cancer chemoresistance via miR-155 transfer. Scientific reports. 2018;8(1):829.
Gorczynski RM, Zhu F, Chen Z, Kos O, Khatri I. A comparison of serum miRNAs influencing metastatic growth of EMT6 vs 4THM tumor cells in wild-type and CD200R1KO mice. Breast Cancer Res Treat. 2017;162:255–66.
Mortazavi Y, Paryan M, Biglari A, Mohammadi-Yeganeh S. Exosomal miRNAs from highly metastatic cells can induce metastasis in non-metastatic cells. Life Sci. 2019;220:162–8.
Shen S, Song Y, Zhao B, Xu Y, Ren X, Zhou Y, et al. Cancer-derived exosomal miR-7641 promotes breast cancer progression and metastasis. Cell communication and signaling. 2021;19:1–13.
Popper HH. Progression and metastasis of lung cancer. Cancer Metastasis Rev. 2016;35(1):75–91.
Riihimäki M, Hemminki A, Fallah M, Thomsen H, Sundquist K, Sundquist J, et al. Metastatic sites and survival in lung cancer. Lung Cancer. 2014;86(1):78–84.
Hussen BM, Ahmadi G, Marzban H, Azar MEF, Sorayyayi S, Karampour R, et al. The role of HPV gene expression and selected cellular MiRNAs in lung cancer development. Microb Pathog. 2021;150:104692.
Rezaei M, Mostafaei S, Aghaei A, Hosseini N, Darabi H, Nouri M, et al. The association between HPV gene expression, inflammatory agents and cellular genes involved in EMT in lung cancer tissue. BMC Cancer. 2020;20:1–11.
Lee KS, Lee YS, Lee JM, Ito K, Cinghu S, Kim JH, et al. Runx3 is required for the differentiation of lung epithelial cells and suppression of lung cancer. Oncogene. 2010;29(23):3349–61.
Li Z, Chen X, Yi X. Tumor promoting effects of exosomal microRNA-210 derived from lung cancer cells on lung cancer through the RUNX3/PI3K/AKT signaling pathway axis. J Biol Regul Homeost Agents. 2021;35(2):473–84.
Tao K, Liu J, Liang J, Xu X, Xu L, Mao W. Vascular endothelial cell-derived exosomal miR-30a-5p inhibits lung adenocarcinoma malignant progression by targeting CCNE2. Carcinogenesis. 2021;42(8):1056–67.
Yi F, Amarasinghe B, Dang TP. Manic fringe inhibits tumor growth by suppressing Notch3 degradation in lung cancer. Am J Cancer Res. 2013;3(5):490–9.
Essandoh K, Yang L, Wang X, Huang W, Qin D, Hao J, et al. Blockade of exosome generation with GW4869 dampens the sepsis-induced inflammation and cardiac dysfunction. Biochem Biophys Acta. 2015;1852(11):2362–71.
Wang Z, Lin M, He L, Qi H, Shen J, Ying K. Exosomal lncRNA SCIRT/miR-665 Transferring Promotes Lung Cancer Cell Metastasis through the Inhibition of HEYL. J Oncol. 2021;2021:9813773.
Yang T, Li H, Thakur A, Chen T, Xue J, Li D, et al. FOXP4 modulates tumor growth and independently associates with miR-138 in non-small cell lung cancer cells. Tumour biology : the journal of the International Society for Oncodevelopmental Biology and Medicine. 2015;36(10):8185–91.
Harlos C, Musto G, Lambert P, Ahmed R, Pitz MW. Androgen pathway manipulation and survival in patients with lung cancer. Hormones and Cancer. 2015;6:120–7.
Berardi R, Morgese F, Santinelli A, Onofri A, Biscotti T, Brunelli A, et al. Hormonal receptors in lung adenocarcinoma: expression and difference in outcome by sex. Oncotarget. 2016;7(50):82648.
Li M, Zhou S, Wu Y, Li Y, Yan W, Guo Q, et al. Prenatal exposure to propylparaben at human-relevant doses accelerates ovarian aging in adult mice. Environ Pollut. 2021;285: 117254.
Comacchio GM, Melan L, Zambello G, Mammana M, Rea F. Lung metastases from hepatocellular carcinoma: multidisciplinary approach—narrative review. AME Surgical Journal, 2. 2022
Guo X, Xu Y, Wang X, Lin F, Wu H, Duan J, et al. Advanced Hepatocellular Carcinoma with Bone Metastases: Prevalence, Associated Factors, and Survival Estimation. Med Sci Mon. 2019;25:1105–12.
Hoshino A, Costa-Silva B, Shen T-L, Rodrigues G, Hashimoto A, Tesic Mark M, et al. Tumour exosome integrins determine organotropic metastasis. Nature. 2015;527(7578):329–35.
Sun W, Fu S, Wu S, Tu R. Growing Evidence of Exosomal MicroRNA-Related Metastasis of Hepatocellular Carcinoma. Biomed Res Int. 2020;2020:4501454.
Turjanski A, Vaque J, Gutkind J. MAP kinases and the control of nuclear events. Oncogene. 2007;26(22):3240–53.
Kim EK, Choi E-J. Compromised MAPK signaling in human diseases: an update. Arch Toxicol. 2015;89:867–82.
Meloche S, Pouysségur J. The ERK1/2 mitogen-activated protein kinase pathway as a master regulator of the G1-to S-phase transition. Oncogene. 2007;26(22):3227–39.
Roux PP, Blenis J. ERK and p38 MAPK-activated protein kinases: a family of protein kinases with diverse biological functions. Microbiol Mol Biol Rev. 2004;68(2):320–44.
Roberts PJ, Der CJ. Targeting the Raf-MEK-ERK mitogen-activated protein kinase cascade for the treatment of cancer. Oncogene. 2007;26(22):3291–310.
Liu C, Wu H, Mao Y, Chen W, Chen S. Exosomal microRNAs in hepatocellular carcinoma. Cancer Cell Int. 2021;21(1):254.
Kumar B, Sinclair J, Khandrika L, Koul S, Wilson S, Koul HK. Differential effects of MAPKs signaling on the growth of invasive bladder cancer cells. Int J Oncol. 2009;34(6):1557–64.
Ang KK, Harris J, Wheeler R, Weber R, Rosenthal DI, Nguyen-Tân PF, et al. Human papillomavirus and survival of patients with oropharyngeal cancer. N Engl J Med. 2010;363(1):24–35.
Bray F, Ferlay J, Soerjomataram I, Siegel RL, Torre LA, Jemal A. Global cancer statistics 2018: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. 2018;68(6):394–424.
Eslami M, Khazeni S, Khanaghah XM, Asadi MH, Ansari MA, Garjan JH, et al. MiRNA-related metastasis in oral cancer: moving and shaking. Cancer Cell Int. 2023;23(1):182.
Nahand JS, Rabiei N, Fathazam R, Taghizadieh M, Ebrahimi MS, Mahjoubin-Tehran M, et al. Oncogenic viruses and chemoresistance: What do we know? Pharmacol Res. 2021;170: 105730.
Lydiatt WM, Patel SG, O'Sullivan B, Brandwein MS, Ridge JA, Migliacci JC, et al. Head and neck cancers—major changes in the American Joint Committee on cancer eighth edition cancer staging manual. CA Cancer J Clin. 2017;67(2):122–37.
Wang Z, Chen J-Q, Liu J-L, Tian L. Exosomes in tumor microenvironment: novel transporters and biomarkers. J Transl Med. 2016;14:1–9.
Zhang H-G, Grizzle WE. Exosomes: a novel pathway of local and distant intercellular communication that facilitates the growth and metastasis of neoplastic lesions. Am J Pathol. 2014;184(1):28–41.
Li L, Li C, Wang S, Wang Z, Jiang J, Wang W, et al. Exosomes derived from hypoxic oral squamous cell carcinoma cells deliver miR-21 to normoxic cells to elicit a prometastatic phenotype. Can Res. 2016;76(7):1770–80.
Shan Y, You B, Shi S, Shi W, Zhang Z, Zhang Q, et al. Hypoxia-induced matrix metalloproteinase-13 expression in exosomes from nasopharyngeal carcinoma enhances metastases. Cell Death Dis. 2018;9(3):382.
Wang M, Yu F, Ding H, Wang Y, Li P, Wang K. Emerging function and clinical values of exosomal microRNAs in cancer. Molecular therapy-Nucleic acids. 2019;16:791–804.
Stacker SA, Williams SP, Karnezis T, Shayan R, Fox SB, Achen MG. Lymphangiogenesis and lymphatic vessel remodelling in cancer. Nat Rev Cancer. 2014;14(3):159–72.
Sundar SS, Ganesan TS. Role of lymphangiogenesis in cancer. J Clin Oncol. 2007;25(27):4298–307.
Hirakawa S, Brown LF, Kodama S, Paavonen K, Alitalo K, Detmar M. VEGF-C–induced lymphangiogenesis in sentinel lymph nodes promotes tumor metastasis to distant sites. Blood. 2007;109(3):1010–7.
Hwang SD, Song JH, Kim Y, Lim JH, Kim MY, Kim EN, et al. Inhibition of lymphatic proliferation by the selective VEGFR-3 inhibitor SAR131675 ameliorates diabetic nephropathy in db/db mice. Cell Death Dis. 2019;10(3):219.
Burton JB, Priceman SJ, Sung JL, Brakenhielm E, An DS, Pytowski B, et al. Suppression of prostate cancer nodal and systemic metastasis by blockade of the lymphangiogenic axis. Can Res. 2008;68(19):7828–37.
Cha DJ, Franklin JL, Dou Y, Liu Q, Higginbotham JN, Demory Beckler M, et al. KRAS-dependent sorting of miRNA to exosomes. eLife. 2015;4:e07197.
Tellez-Gabriel M, Knutsen E, Perander M. Current status of circulating tumor cells, circulating tumor DNA, and exosomes in breast cancer liquid biopsies. Int J Mol Sci. 2020;21(24):9457.
Acknowledgements
None declared.
Funding
No funding was received.
Author information
Authors and Affiliations
Contributions
M.B. was responsible for the literature search, writing the initial and edited draft of the manuscript, and visualization. J.S.N. proposed the topic and main idea, commented on and revised the paper, and finalized the manuscript. Both authors have read and approved the article.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Bayat, M., Sadri Nahand, J. Exosomal miRNAs: the tumor's trojan horse in selective metastasis. Mol Cancer 23, 167 (2024). https://doi.org/10.1186/s12943-024-02081-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12943-024-02081-0