
Abstract
Osteosarcoma is the most common primary bone cancer in children and young adults. Limited progress has been made in improving the survival outcomes in patients with osteosarcoma over the past four decades. Especially in metastatic or recurrent osteosarcoma, the survival rate is extremely unsatisfactory. The treatment of osteosarcoma urgently needs breakthroughs. In recent years, immunotherapy has achieved good therapeutic effects in various solid tumors. Due to the low immunogenicity and immunosuppressive microenvironment of osteosarcoma, immunotherapy has not yet been approved in osteosarcoma patients. However, immune-based therapies, including immune checkpoint inhibitors, chimeric antigen receptor T cells, and bispecfic antibodies are in active clinical development. In addition, other immunotherapy strategies including modified-NK cells/macrophages, DC vaccines, and cytokines are still in the early stages of research, but they will be hot topics for future study. In this review, we showed the functions of cell components including tumor-promoting and tumor-suppressing cells in the tumor microenvironment of osteosarcoma, and summarized the preclinical and clinical research results of various immunotherapy strategies in osteosarcoma, hoping to provide new ideas for future research in this field.
Similar content being viewed by others


Introduction
Osteosarcoma is the most common primary malignant bone tumor that mostly occurs in children and young adolescents [1]. It originates from mesenchymal stem cells, characterized by osteoid product [2]. Osteosarcoma exhibits a predilection to occur in the metaphysis of long extremity bones, such as distal femur, proximal tibia, proximal femur, and proximal humerus [3]. It was reported that approximately 60% of patients are between 10 and 20 years old and osteosarcoma is considered as the second leading cause of death in this age group [1]. The incidence of osteosarcoma is 3–5 per million per year [4]. Despite their rare incidence, osteosarcoma has a high disability and mortality rate. The current standard treatment for osteosarcoma is neoadjuvant chemotherapy-surgery-consolidation chemotherapy treatment mode, which achieves great clinical success for patients with localized osteosarcoma [5]. The 5-year survival rate has reached a plateau in osteosarcoma patients with localized disease ranging from 60 to 70% since the introduction of systematic chemotherapy [6]. Unfortunately, osteosarcoma is a tumor with a high tendency for metastasis, and 15-20% of newly diagnosed osteosarcoma are detected with metastasis [7]. The most common metastatic site is lung. And the paitients with metastatic disease either at diagnosis or at the time of recurrence portends a dismal prognosis with 5-year survival of only 20-30% [8].
Limited progress has been made in improving the survival outcomes in patients with osteosarcoma over the past 40 years. Especially, the prognosis of metastatic or recurrent osteosarcoma is still unsatisfactory. Given these bottlenecks to conventional treatment regimens, there is an urgent need to exploit novel therapeutic strategies to improve the treatment situation of patients with osteosarcoma. In addition to traditional treatment methods including surgery, chemotherapy, and radiotherapy, new therapeutic approaches, such as targeted therapy and immunotherapy, are also under intense research for osteosarcoma. Currently, research on targeted therapy for osteosarcoma mainly revolves around anti-angiogenic small molecule inhibitors, with representative drugs being regorafenib and sorafenib. These anti-angiogenic drugs, do have certain effects on metastatic osteosarcoma, but they have short progression-free survival (PFS) time and insufficient objective response rate (ORR) [9].
In recent years, immunotherapies involving the application of immune checkpoint inhibitors (ICIs), genetically modified T cells, tumor vaccines, immune modulators and cytokines, have received considerable attention. Importantly, due to their impressive efficacy, ICIs are recommended for the treatment of several human malignancies, including melanoma [10], non-small-cell lung cancer [11], head and neck cancers [12] and so on. Chimeric antigen receptor T (CAR-T) cells as an adoptive cell therapy (ACT) have showed remarkable clinical outcome in the treatment of malignant hematological tumors, such as acute lymphoblastic leukemia [13], chronic lymphocytic leukemia [14], and non-hodgkin lymphoma [15]. Immunotherapy can more accurately target tumors and activate the host’s anti-tumor immune response. Compared to traditional chemotherapy, immunotherapy has fewer side effects and possesses long-lasting anti-tumor activity. The microenvironment of osteosarcoma has complex heterogeneity and immunosuppressive properties, which makes immunotherapy not yet approved for indications in osteosarcoma. However, it is undeniable that immunotherapy is becoming an increasingly attractive therapy strategy for osteosarcoma. In this review, we will present on the tumor microenvironment characteristics of osteosarcoma, including immune-suppressive related cells and tumor-suppressive related cells, as well as their roles in the osteosarcoma microenvironment. We will also summarize the results of preclinical and clinical research on immunotherapy, especially ICIs and CAR-modified immune cell therapies, as well as the immuno-combination strategies, with the aim of further understanding the immune characteristics of osteosarcoma from multiple perspectives and providing new directions for the application of immunotherapy for patients with osteosarcoma.
Immune microenvironment of osteosarcoma
The tumor microenvironment (TME) is a complex ecosystem with high heterogeneity, which includes tumor cells and a large number of non-tumor cells, embedded in altered extracellular matrix. In the same way, the TME of osteosarcoma also contains many components which are closely related to the escape of tumor cells from immune surveillance, uncontrolled proliferation, and metastasis. The immune cells in the TME not only play a crucial role in the occurrence and development of tumors, but also are important targets of anti-tumor immunotherapy [16]. There are various types of immune cells in osteosarcoma, mainly including tumor-associated macrophages (TAMs), tumor-associated neutrophils (TANs), myeloid-derived suppressor cells (MDSCs), T cells, B cells, natural killer cells (NK cells), and dendritic cells (DCs) and so on. In addition, molecular pathways, especially immune related molecules in the microenvironment of osteosarcoma, are closely related to the development of tumors and the prognosis of patients.
Characteristics of cell composition in the TME of osteosarcoma
The TME of osteosarcoma is distinctly suppressive, with immune cells exhibiting complex and diverse functions. Understanding the roles and mechanisms of different immune cells within it, and developing corresponding therapeutic strategies targeting specific elements, can help improve the prognosis for patients with osteosarcoma.
Tumor-promoting cells in TME
Tumor cells
Tumor cells, as the key components of TME, which can actively interact with the immune system and promote the formation of inhibitory immune networks [17]. In recent years, circulating tumor cells (CTCs) have been frequently mentioned. Under the impact of local and systemic treatment, some tumor cells can still survive in small amounts and escape from the primary lesion into the circulatory system, causing distant metastasis or recurrence of osteosarcoma [18]. Therefore, research on these tumor cells and the development of corresponding treatment methods will be benefit to reduce the metastasis and recurrence of osteosarcoma.
TAMs
TAMs are the most abundant immune cells in the immune microenvironment of osteosarcoma and account for approximately 50% of the immune cells [19]. The polarization state of TAMs has a significant impact on tumor cells. Generally, TAMs polarize into two types, M1 and M2, primarily due to the influence of different cytokines and signaling pathways in the tumor microenvironment [20]. In the TME of osteosarcoma, M2-type TAMs are the main proportion.and play a complex and crucial role. M2-type TAMs are typically associated with anti-inflammatory and tumor-promoting characteristics, and they participate in processes such as tumor angiogenesis, extracellular matrix remodeling, and immune defense and regulation [21]. The polarization state of M2-type TAMs can promote tumor progression by releasing specific cytokines and chemokines, such as producing anti-inflammatory cytokines like IL-10 and transforming growth factor-beta (TGF-β), which are involved in anti-inflammatory responses, and participate in tumor angiogenesis and extracellular matrix remodeling [22]. M2-type TAMs in the TME of osteosarcoma may also affect tumor immune evasion and drug resistance through interactions with other immune cells. For example, they may interact with cancer stem cells, supporting the function of cancer stem cells [23]. In addition, the presence and functional state of M2-type TAMs are closely related to the prognosis and therapeutic responsiveness of osteosarcoma. Studies have shown that the degree of infiltration of M2-type TAMs can serve as one of the biomarkers for judging the prognosis of osteosarcoma patients [21]. Various immune checkpoint proteins expressed on the surface of TAMs, such as PD-1 and CD47 receptor [19]. Therefore, therapeutic strategies targeting M2-type TAMs, such as promoting the polarization shift of M2-type TAMs to M1-type or targeting the immune checkpoints of TAMs, may provide new directions for targeted therapy of osteosarcoma [21]. In addition, the M3-TAMs, actually the FABP4+ TAMs previously identified as the alveolar macrophages in the lung, were dominant in the lung metastatic osteosarcoma lesion [24]. High expression of FABP4 provides fatty acids for tumor cells to promote the occurrence and development of cancer [25]. There has been some research on the mechanism of FABP4 action in tumors, but the specific mechanism is not yet fully understood. M3-TAMs contribute to the metastasis and colonization of osteosarcoma cells in the lungs [26], exacerbating the malignant progression of the tumor. Developing targeted therapeutic strategies against M3-TAMs may potentially suppress the pulmonary metastasis of osteosarcoma.
MDSCs
MDSCs are significant compotent in the TME of osteosarcoma, MDSCs attenuate anti-tumor immunity particularly the activity of T cells, by secreting factors such as arginase-1 (Arg-1), inducible nitric oxide synthase (iNOS), and reactive oxygen species (ROS), thereby facilitating tumor cells to evade immune surveillance [27]. MDSCs also contribute to tumor angiogenesis through the secretion of factors like vascular endothelial growth factor (VEGF) and matrix metalloproteinase 9 (MMP9) [28]. Moreover, MDSCs interact with other cells in the tumor microenvironment, such as TAMs and Tregs, to jointly promote immune evasion and tumor progression [29]. MDSCs establish pre-metastatic niches in distant organs, creating favorable conditions for the spread and colonization of tumor cells [30]. And they also secrete TGF-β and hepatocyte growth factor (HGF), which contribute to epithelial-mesenchymal transition (EMT), thereby promoting the invasion and metastasis of osteosarcoma [31]. Recent study has discovered that CD300ld, a surface molecule on MDSCs, plays a significant role in the immune-suppressive function of MDSCs and may potentially serve as a novel therapeutic target [32]. Given the critical role of MDSCs in immune suppression and tumor progression, they have become potential targets for immunotherapy. Treatment strategies targeting MDSCs may help to lift immune suppression and enhance the efficiency of the anti-tumor immune response. However, that need more exploration before clinical application [33].
Treg cells
In the osteosarcoma microenvironment, Treg cells exert their effects through various mechanisms. They secrete immunosuppressive cytokines such as IL-10, IL-35, and TGF-β to inhibit the activity of effector T cells, and inhibit the formation of osteoclasts through direct cell contact-dependent manners [34]. Interaction of CD4+ Tregs and osteoclasts significantly alters TME and is connected to poor prognosis of osteosarcoma [34]. And the interaction between T cell immunoglobulin and mucin-domain containing 3 (TIM-3)+ T cells and monocytes, naive CD4+ T cells, and Gal9-expressing CD4+ CD25+ Tregs could resulting in progressive suppression of Th1 responses [35]. Moreover, Zhou et al. Report that infiltration of T cell immunoreceptor with Ig and ITIM domains (TIGIT) expressing Treg cells in osteosarcoma tissue, providing a new target for the immunotherapy of osteosarcoma [4]. Treg cells not only help tumor cells evade immune surveillance but also participate in promoting tumor angiogenesis [36]. Furthermore, the heterogeneity of Treg cells in the osteosarcoma microenvironment and their proliferation and activation status are closely related to tumor development, potentially serving as targets for immunotherapy.
TANs
TANs are a heterogeneous and functionally diverse subset of neutrophils that infiltrate the tumor microenvironment. TANs could also be polarized to the anti-tumor N1 phenotype and pro-tumor N2 phenotype. Infiltrating neutrophils combat tumor cells by orchestrating the recruitment of immune cells and also by facilitating antibody-dependent cellular cytotoxicity [37]. Yang et al. identified that neutrophils are more abundant at the primary tumor site than those at metastatic sites by detecting the specific marker CD11b [38]. It is reported that TANs significantly decresed in the highly hypoxic tumor microenvironment. Hypoxia may downregulate immune cells, thereby facilitating tumor immune evasion and metastasis [39]. Web-like chromatin structures known as neutrophil extracellular traps (NETs) plays important roles in immune protection, inflammatory and autoimmune diseases and cancer [40]. In osteosarcoma, a signature derived from NETs is associated with tumor recurrence and metastasis, and can predict patient prognosis [41, 42]. Currently, research on TANs in osteosarcoma is in its infancy, and there is a long road ahead in developing anti-tumor therapeutic strategies associated with TANs.
Cancer stem cells (CSCs)
CSCs are a subset of tumor cells that play a pivotal role in various stages of cancer development, including tumorigenesis, drug resistance, recurrence and metastasis [43, 44]. Extensive research suggests that osteosarcoma may arise from mesenchymal stem cells (MSCs), which contribute to modulate immune responses, facilitate cell fusion and differentiation [45]. MSCs are capable of secreting anti-inflammatory factors while suppressing pro-inflammatory agents, thereby aiding osteosarcoma cells in evading immune surveillance. This process is mediated by extracellular vesicles (EVs), particularly exosomes, which can be released through autocrine or paracrine mechanisms. MSC-EVs inhibit T cell proliferation and the immunological effects of B cells [46, 47]. TGF-β and interferon-γ (IFN-γ) associated with MSC-EVs can facilitate the transformation of mononuclear cells into Tregs [48]. Furthermore, MSCs can promote the polarization of TAMs towards the M2 phenotype by secreting IL-6 [49].
Tumor-associated fibroblasts (TAFs)
TAFs comprise a large proportion of cells in the microenvironment and produce components of the extracellular matrix that facilitate communication between cells, promote cell adhesion, and stimulate proliferation [50]. TAFs can be reprogrammed in lung under the influence of TGF-β1 in osteosarcoma-derived EVs, which promotes metastatic progression of osteosarcoma [51]. The interaction of osteosarcoma cells expressing CXCL14 and fibroblasts expressing integrin α11β1 promotes metastasis and demonstrate that targeting the CXCL14-integrin α11β1 axis is a potential strategy to inhibit osteosarcoma lung metastasis [52].
Tumor-suppressing cells in TME
T cells
T cells, particularly cytotoxic T cells, play a significant role in the immune microenvironment of osteosarcoma. T cell infiltration is the first step in its anti-tumor effectiveness. Analyses of TME of osteosarcoma demonstrate an immune cell infiltrate consisting of both macrophages and T cells [53, 54]. TIM-3(+)PD-1(+) T cells-specific immunosuppression that was amplified by M2-type tumor-associated macrophages. depletion of CD163(+) macrophages significantly improved T cell proliferation and proinflammatory cytokine production [55]. In osteosarcoma, the number of T cells in metastatic lesions is significantly higher than that in primary and recurrent lesions in situ [56]. An increased number of tumour-infiltrating T cells and PD-L1 expression in metastases compared with primary tumours, suggesting accessibility for T cells, could imply that osteosarcoma patients with metastatic disease may benefit from T-cell-based immunotherapy [56]. Tumor infiltrating lymphocytes (TILs) are mainly distributed in the region expressing human leukocyte antigen Class I, whereas CD4+ and CD8+ T cells concentrated at the pulmonary metastases interface [33]. The CD8+ T cells in TME of osteosarcoma are functionally inactive with increased expression of PD-1 and TIM-3. TIM-3 blockade restored the T cell alloreactive function of the CD8+ T cells in vitro and in vivo [57]. A study claimed that CD8+ T cells have a positive association with prognosis, whereas γδT cells are associated with poor prognosis [58]. As the main killer of tumor cells in the microenvironment, T cells have become the most important research object for developing immunotherapy.
DCs
DCs, as professional antigen-presenting cells (APCs), act crucial roles between innate and adaptive immunity [59]. DCs detect tumor antigens and present them to helper cells and cytotoxic T cells, thereby triggering the anti-tumor immune response. During this process, DCs also transform from an immature state to a mature state. During the initial phases, DCs engage in robust proliferation and maturation, which is essential for the activation of both helper and cytotoxic T cells. However, as the tumor progresses, osteosarcoma cells evolve to generate variants that are resistant to the actions of DCs and phagocytes. This resistance results in diminished DC activation, ultimately allowing the tumor to evade the immune surveillance [60]. A study found that in group with high immune scores, the quantity of resting DCs was significantly higher than that in the low immune score group, and the degree of DC activation was positively related with anti-tumor therapy response [61]. The role of DCs in the immune microenvironment is complex. A study reported that DCs drive the pathogenesis of osteosarcoma through oncogenes and the tumor suppressor glutamate metabotropic receptor 4 (GRM4) [62]. Compared with wild-type DCs, GRM4-knockout DCs secrete more IL-23 and IL-12, leading to rapid tumor growth and accelerated progression in vivo [63]. The proportion of CD1c+ DCs is larger in lung metastatic lesions than that in primary and recurrent lesions [4]. At present, the most popular therapeutic strategy for DCs is vaccination. In osteosarcoma, researchers tested the efficacy of DC vaccines in preclinical studies and demonstrated that DCs vaccines induced tumor suppression [64]. However, in clinical trials, DC vaccines showed limited effect for the treatment of osteosarcoma [65, 66].
NK cells
NK cells play key roles in the killing of tumor cells by releasing perforin, granzyme, and tumor necrosis factor-α (TNF-α) and expressing FasL [67]. But the anti-tumor effects of NK cells are also affected by the PD-1/PD-L1 interactions [68] In the osteosarcoma microenvironment, NK cells were suppressed, but TGF-β expression increased. TGF-β plays a key role in blunting the NKG2D-mediated tumor surveillance [69]. TIGIT also is expressed on NK cells. TIGIT inhibits human NK cytotoxicity against the tumor by interacting with the ligands, including CD155 and CD112 [70]. Therefore, targeting immune checkpoints and/or TGF-β pathway maybe can reinvigorate tumor elimination efficiency of NK cells. Additionally, stimulating the inherent tumor-killing function of NK cells is also a promising strategy. Adoptive NK cells showed initial success in the treatment of osteosarcoma [71].
B cells
B cells are not only the protagonists of humoral immunity by producing antibodies but are also a type of antigen-presenting cells involved in immunoregulation. Regulatory B cells are a type of B cells with immunosuppressive effects. Regulatory B cells inhibit CD4+ T cells, cytotoxic T lymphocytes (CTLs), macrophages, and DCs by secreting inhibitory cytokines such as IL-10, TGF-β, IL-35, and expressing membrane surface regulatory molecules such as FasL and CD1d, and promote the transformation of T cells into Tregs, thus weakening anti-tumor immune responses [72]. Osteosarcoma patients with high infiltration of B cells had a better prognosis and activated B cells were positively correlated with survival [73]. Research on B cells in osteosarcoma is still very limited, and new therapies based on B cells lack satisfactory results.
M1-type TAMs and C1Q + TAMs
TAMs are typed into M1 and M2. Generally, M1 macrophages participate in anti-tumor effects, while M2 macrophages promote carcinogenesis [20]. Although M2-TAMs are predominant in the osteosarcoma microenvironment, M1 macrophages also play their respective roles. M1 macrophages release pro-inflammatory mediators such as IL-1, IL-12, IL-18, and TNF-α, which have anti-tumor effects. In addition, a analysis of single-cell RNA datasets and bulk RNA datasets demonstrate that complete component 1q (C1Q)-positive TAMs have anti-tumor effect and predict a better prognosis for patients with osteosarcoma [74].
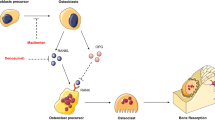
The cellular components infiltrating the tumor microenvironment are diverse (Fig. 1), and their functions are not singular but also varied. Immune cells in different states may primarily promote tumor progression, or they may exert anti-tumor effects when the surrounding environment changes. Therefore, it is necessary to view the cells in the microenvironment dialectically and develop appropriate anti-tumor treatment strategies based on their characteristics.

Schematic illustration of the cellular components infiltrated in the osteosarcoma microenvironment. The TME of osteosarcoma contains tumor cells and a large number of non-tumor cells, embedded in altered extracellular matrix. The cells in the TME can be divided into tumor-promoting cells and tumor-suppressing cells.They exert their effects through different mechanisms in the microenvironment, but their functions are dynamically changing. Compared to the primary lesion, the expression of immune checkpoint molecules in the lung metastatic foci is increased, and there is an increase in immunosuppressive cells
Molecular characteristics of osteosarcoma microenvironment
The bone microenvironment is closely related to the progression and malignancy of osteosarcoma. The analysis of TME landscapes in osteosarcoma suggested that patients with non-metastasis showed higher immune scores and better prognosis compared with those with metastatic osteosarcoma. In addition, high levels of tumor-infiltrating immune cells may suppress metastasis [38]. Wu et al. conducted a comprehensive genomic and immune characterization of post-treatment primary, local recurrence, and metastasis osteosarcoma specimens from 48 pediatric and adult patients. The results suggested that the high levels of genomic rearrangements and moderate point mutation burdens in osteosarcoma, which was not associated with immune infifiltrate levels and neoantigen expression. Besides, there are likely multiple immune-suppressive mechanisms in play in osteosarcoma [75]. The immunosuppressive mechanisms in osteosarcoma, including (1) Highly mutated and rearranged osteosarcoma genome may not generate sufficient neoantigens to elicit an immune response, (2) Aneuploidy and copy number changes alterations and (3) Deregulation of tumor-intrinsic immunosuppressive pathways such as IFNG, MAPK/PI3K/mTOR, and JAK/STAT signal pathways [76]. Yang et al. summarized the factors, including mesenchymal stem cells, hypoxia and acidic condition, chemokines and extracellular vesicles, that affect osteosarcoma metastasis in bone microenvironment through several signaling pathways including the PI3K/Akt, Wnt/β-Catenin, MAPK/ERK, Hedgehog, and Notch signaling pathways[PMID: 32977425]. TAM-derived cathepsins may mediate the activation of the nuclear factor-kappa B (NF-kB) signaling pathway and the signal transducer and activator of transcription 3 (STAT3) to facilitate therapeutic resistance [77]. Sundara et al. studied 85 formalin-fixed, paraffin-embedded blocks from 25 osteosarcoma patients and found that the high density of tumor-infiltrating T cells in metastatic osteosarcoma lesions compared to primary tumors and local relapses. Besides, positive PD-L1 expression was found in 13% of primary tumors, 25% of relapses and 48% of metastases and correlated with a high T-cell infiltration [56]. Wan et al. identified five immune subtypes in 87 osteosarcoma samples, and each of them was associated with distinct molecular characteristics and clinical outcomes [78]. The study of RNA sequencing of 100,987 individual cells from seven primary, two recurrent, and two lung metastatic osteosarcoma lesions provides a deeper insight into the cellular and molecular characteristics of osteosarcoma and its TME properties. The results showed that proinflammatory FABP4+ macrophages infiltration was found in lung metastatic osteosarcoma. And a large number of inactivated and exhausted T cells were observed, especially CD8+ T cells that highly express the inhibitory receptor TIGIT and lymphocyteactivation gene 3 (LAG-3). Compared to primary lesions, pulmonary metastatic lesions expressed more immune checkpoint and immunoregulatory molecules, including PD-1, PD-L1, LAG-3, TIM-3, indoleamine 2,3-dioxygenase (IDO1) and IFN-γ [33].
Given the suppressive immune microenvironment and high heterogeneity of osteosarcoma, achieving satisfactory therapeutic outcomes with immunotherapy may require approaches from the following aspects: (1) enhancing the immunogenicity of tumors to allow more immune cells to infiltrate the tumor tissue; (2) revitalizing the activity of anti-tumor immune cells within the microenvironment, turning “cold” tumors into “hot” tumors; (3) identifying biomarkers that can predict the responsiveness to immunotherapy, to select a population of osteosarcoma patients who are likely to benefit from such treatments; (4) considering the diverse immunosuppressive pathways in osteosarcoma, a single immunotherapy might have limited effects, while combined therapies could potentially break through this bottleneck.
The relationship between TME and prognosis in osteosarcoma
In order to detect the correlation between microenvironment and clinical parameters, Hong et al. calculated immune and stromal scores based on the ESTIMATE algorithm in 83 osteosarcoma samples and found that 137 genes were dysregulated, including 134 upregulated genes and three downregulated genes [79]. Hu et al. analyzed the data of 89 osteosarcoma samples and identified 769 TME-related genes by comparing the high-immune score and low-immune score osteosarcoma patients, which confirmed that patients with higher immune score had a favorable overall survival (OS) and disease free survival (DFS) [80]. Sheng et al. reviewed the potential mechanisms underlying osteosarcoma metastasis including microenvironment, osteoclast, angiogenesis, metabolism, immunity, and noncoding RNAs. For the immunity and metastasis, TAMs, TILs interactions between of PD-1 and PD-L1 putatively participate in immune response during osteosarcoma metastatic progression [81]. In addition, analysis of 85 osteosarcoma samples from TCGA dataset indicated that osteosarcoma cases with elevated immune cell infiltration in the microenvironment showed better prognosis [61].
In general, the TME of osteosarcoma is immunosuppressive, as increased expression of immunosuppressive molecules such as PD-1 and PD-L1 has been detected, especially in metastatic lesions. In theory, treatment with ICIs can be effective for osteosarcoma patients. Besides, osteosarcoma is a low immunogenic tumor that cannot attract a sufficient number of immune cells and tumor-specific T cells to infiltrate the tumor microenvironment. Therefore, combination strategies to improve the immunogenicity of osteosarcoma and then administering immunotherapy may have stronger anti-tumor effects. And discovering more potential tumor antigen that is specifically or at least relatively exclusively expressed in osteosarcoma will be beneficial for the development of immunotherapy strategies based on these targets.
Studies of ICIs for osteosarcoma
PD-1/PD-L1
Osteosarcoma has been shown to have variable PD-L1 expression and frequent deletions of MHC class I, which may promote immune evasion [56]. Metastatic osteosarcoma express higher PD-L1 than primary tumors. Blockade of PD-1/PD-L1 interactions significantly promotes the anti-tumor activity of tumor-infiltrating CTLs in vitro and in vivo [82]. Nivolumab (anti-PD-1 antibody) inhibits osteosarcoma metastasis in human PBMC-engrafted mouse models. The further study of mechanisms indicated that nivolumab promotes tumor lymphocyte infiltration (CD4+ and CD8+ lymphocytes) and enhances the cytolytic activity of CD8 lymphocytes in lung metastases [83]. Davis et al. initiated a phase I/II clinical study to test the safety, pharmacokinetics, and anti-tumor activity of nivolumab in children and young adults with recurrent or refractory non-CNS solid tumors or lymphoma [84]. The preliminary research results showed that nivolumab only has 10 -30% response rate in lymphoma but no response in other tumor types (including osteosarcoma), although the treatment related side effects were acceptable [84]. SARC028 trial (NCT02301039) is the first study to assess the activity and safety of anti-PD-1-antibodies in the treatment of soft-tissue sarcoma and bone sarcoma. This trail recruited 86 patients, and 80 of whom were treated with 200 mg intravenous pembrolizumab every three weeks. The results showed that two (5%) of 40 patients with bone sarcoma had an objective response, including one (5%) of 22 patients with osteosarcoma and one (20%) of five patients with chondrosarcoma. And the adverse events were acceptable [85]. Another single-arm, open-label, phase II trial (NCT03013127) was performed to evaluate the anti-tumor activity and safety of the PD-1 antibody pembrolizumab in patients with unresectable, relapsed osteosarcoma. Total 12 patients were enrolled in this study and accepted treatment of pembrolizumab 200 mg every 21 days. The results demonstrated that pembrolizumab was well tolerated but did not show clinical benefit [86]. Besides, a phase II study (NCT02406781) was conducted to test the safety and anti-tumor effects of pembrolizumab in combination with metronomic cyclophosphamide in patients with advanced osteosarcoma. It is a pity that only one patient experienced partial response (PR). Expression level of PD-L1 is not directly related to anti-tumor efficacy [87]. PD-1/PD-L1 antibodies not only block the interaction of PD-1/PD-L1, but also show anti-tumor effect through other ways. Liu et al. reported that atezolizumab (anti PD-L1 antibody) inhibits proliferation and induces immune-independent apoptosis of osteosarcoma cells through increasing the release of ROS and cytochrome-c. The excessive release of ROS could induce autophagy. They performed further studies in vitro and in vivo and demonstrated that blocking the protective autophagy induced by atezolizumab could significantly amplify its anti-tumor effect on osteosarcoma cells [88]. The safe dose of atezolizumab was confirmed in pediatric and young adult patients, supportive of weight-based dosing in pediatric patients [89]. However, the therapeutic effect of atezolizumab monotherapy was limited [90].
CTLA-4
Cytotoxic T lymphocyte-associated antigen-4 (CTLA-4) participates in the negative regulation of T cell activation and proliferation, which suppresses anti-tumor response [91]. The study suggested that CTLA-4 genetic polymorphisms also potentially associated with osteosarcoma risk [92, 93]. A phase I clinical trial was conducted by Merchant et al. to test the safety, pharmacokinetics, and immunogenicity of ipilimumab (anti-CTLA-4 antibody) in pediatric patients with advanced solid tumors, including osteosarcoma. The results suggested that ipilimumab showed incredible safety and increased numbers of activated and cycling T cells, but not regulatory T cells [94].
CD47
There is a large amount of macrophage infiltration in osteosarcoma, and the phagocytic effect of macrophages on tumor cells is inhibited by CD47 molecule. CD47 as a transmembrane protein inhibits macrophage phagocytosis when it binds to macrophage SIRPα [95]. Under physiological conditions, this is protection for normal tissues. But for tumors, CD47 is a checkpoint molecule that is overexpressed on tumor cells and inhibits macrophage anti-tumor activity. CD47 is also highly expressed in osteosarcoma and associated with the progression of tumor [96]. In vitro and in vivo osteosarcoma models demonstrated that anti-CD47 monoclonal antibody can block the CD47-SIRPα signaling pathway, thus enhancing the anti-tumor ability of macrophages [96].
TIM-3
TIM-3 was originally found to be expressed on the surface of f type 1 T helper (Th1) cells [97]. TIM-3, as a negative regulator, binds to its ligand galectin-9 (Gal-9) to induce the depletion of Th1 cells [98]. Subsequently, TIM-3 was found to be expressed on the surface of various immune cells, including CD8+T, CD4+T, Treg, macrophage, NK cells and DCs, playing different roles [98]. Blocking the TIM-3/Gal-9 signaling pathway can significantly increase the function of T cells. Therefore, the negative regulation of TIM-3 has attracted much attention for the anti-tumor therapy. TIM-3 and Gal-9 are expressed in osteosarcoma tissues, and the interaction of TIM-3 and Gal-9 promotes the apoptosis of CD4+ and CD8+ T cells in the TME of osteosarcoma, which is related with poor prognosis in osteosarcoma patients [99]. TIM-3 blockade impaired tumor growth in osteosarcoma models and decreased the number of tumor-infiltrating CD4+ T cells while increasing the numbers and functional activation of tumor-infiltrating CD8+ T cells [57]. To date, clinical research outcomes regarding TIM-3 inhibitors have not been published for osteosarcoma. But TIM-3 is a significant potential target for future advancements.
In addition to TIM-3, the expression of LAG-3 and IDO1 are significantly expressed in pulmonary metastatic foci of osteosarcoma and is closely related to immune suppression [33]. Another inhibitory molecule HHLA2, a newly defined B7 family member, widely expressed in osteosarcoma, which is associated with metastases and worse survival [100]. And the immunoreceptor inhibitory checkpoint marker TIGIT was highly expressed by the CD8+ T, CD4+ T, Treg and NKT cells in osteosarcoma lesions. Blocking TIGIT signaling can significantly enhance the cytotoxicity effects of CD3+ T cells with high expression of TIGIT against osteosarcoma cells, indicating potential therapeutic value of targeted TIGIT therapy for osteosarcoma [4]. Currently, there is more research on traditional immune checkpoints, such as PD-1, PD-L2 and CTLA-4, but emerging immune checkpoint molecules have tremendous potential and are worth further investigation in osteosarcoma. We summarized the clinical trials of ICIs in patients with osteosarcoma (Table 1). But the current research results show that immune checkpoint inhibitors have unsatisfactory anti-tumor effects in osteosarcoma. Perhaps exploring the underlying mechanisms of immunotherapy at a deeper level can provide new directions and assistance for the application of immunotherapy in osteosarcoma.
Studies of CAR-modified cellular therapies for osteosarcoma
CAR-T therapy
Adoptive T cell transfer, particularly CAR-engineered T cells, is one of the current research hotspots. At present, research on adoptive cell therapy represented by CAR-T is flourishing in various tumors, especially in hematological tumors, which have achieved good therapeutic effects. CAR-T also has shown great potential in the treatment of osteosarcoma. Targeting suitable molecules is the prerequisite for the anti-tumor effects of CAR-T. Human epidermal growth factor receptor-2 (HER2) CAR-T cells, disialoganglioside (GD2) CAR-T cells, and B7H3 CAR-T cells have been discussed the most in osteosarcoma [101].
HER2
HER2 expression has been reported in 60-70% of primary osteosarcoma [102] and is associated with adverse clinical outcomes [103]. Due to the low expression level of HER2, anti-HER2 monoclonal antibodies were ineffective for osteosarcoma patients with HER2-positive [104]. Therefore, Ahmed et al. developed HER2-specific CAR-T cells and tested the their ability to proliferate, produce immunostimulatory cytokines, and kill HER2-positive osteosarcoma cell lines in vitro. The HER2-positive CAR-T cells showed markedly regression of osteosarcoma both in locoregional and lung metastatic models [105]. In addition, HER2-specific CAR-T cells also decreased sarcosphere formation of drug resistant tumor-initialing cells [106]. In 2015, Ahmed et al. reported the results of a phase I/II clinical trail (NCT00902044) which to evaluate the safety and efficacy of HER2-CAR-T cells in patients with refractory or metastatic HER2-positive sarcoma. This trail included 19 patients with HER2-positive tumors (16 osteosarcomas, one Ewing sarcoma, one primitive neuroectodermal tumor, and one desmoplastic small round cell tumor). Patients received escalating doses of HER2-CAR-T cells (1 × 104 to 1 × 108 /m2) without the administration of IL-2 or lymphodepleting chemotherapy. The results demonstrated that infusion of up to 1 × 108 /m2 HER2-CAR-T cells was well tolerated, and 4 of 17 evaluable patients had stable disease for 12 weeks to 14 months. After removal of residual metastasis, three patients remain in remission at 6, 12, and 16 months, and one showing > 90% necrosis.The median OS time of all 19 patients is 10.3 months (range 5.1 to 29.1 months). Further analysis found that the CAR-T cells persisted for up to 18 weeks in peripheral blood, and they were detected in tumor sites [107].
GD2
Osteosarcoma cells express high level GD2 antigen that can be a potential target [108]. GD2-specific CAR-T cells exhibited therapeutic potency in Ewing sarcoma priclincal models [109]. Chulanetra et al. constructed CAR modified T cells targeting GD2 and demonstrated that the osteosarcoma cells expressing high levels of GD2 were be effectively targeted and killed by GD2-CAR-T cells in vitro. Besides, the combination of GD2-CAR-T cells with suboptimal chemotherapeutic treatment with docorubicin showed stronger anti-tumor efficacy. Unfortunately, further analysis suggested that the osteosarcoma cells and GD2-CAR-T cells were induced to up-regulate expression of PD-L1 and PD-1, respectively. And the interaction of PD-1 and PD-L1 promoted apoptosis of CAR-T cells [110]. This phenomenon suggests that the combination of PD-1/PD-l1 antibody and CAR-T cells may exhibit better anti-osteosarcoma effects.
ALCAM
Activated leukocyte cell adhesion molecule (ALCAM, CD166) is a 105 kDa trans-membrane glycoprotein that belongs to the immunoglobulin superfamily. The study demonstrated that ALCAM was associated with tumorigenesis in osteosarcoma [111]. ALCAM acts as a potential therapeutic target for osteosarcoma patients with ALCAM positivity. Wang et al. developed CAR-T cells targeting ALCAM and evaluated the anti-tumor efficacy. ALCAM-CAR-T cells exhibited specific and potent cytotoxicity against human osteosarcoma cells in vitro; the cytotoxicity was positively correlated with the level of ALCAM expression on the osteosarcoma cells. In murine models, ALCAM-CAR-T cells inhibited the tumor growth with no obvious toxicity [112].
B7-H3
B7-H3 (CD276) is a checkpoint molecule that contributes to tumor immune evasion, metastasis and poor prognosis [113, 114]. B7-H3 highly expressed on pediatric solid tumors, including osteosarcoma. Majzner et al. constructed a novel second-generation CAR-T cells directing B7-H3 and tested the anti-tumor efficacy in a MG63.3 xenograft model of osteosarcoma with strong metastatic potential. The results demonstrated that B7-H3-CAR-T cells mediated complete regression, eradication of osteosarcoma and lead to a significant survival advantage compared with control group. In addition, they found that CAR-T cell activity is dependent on B7-H3 molecule density, which reduced the off-target effects of CAR-T cells [115]. Zhang et al. conducted the third-generation CAR-T cells targeting B7-H3 and proved the killing effects for tumor cells in vitro and in vivo using the patientderived xenografts (PDXs) model of osteosarcoma [116,117,118,119,120,121,122,123,124,125].
EphA2
EphA2 is a tyrosine kinase receptor for Ephrin signalling during embryonic development. In osteosarcoma, overexpression of EphA2 has association with oncogenic signalling, the promotion of angiogenesis and tumor progression [117]. EphA2 is a promising target because of the high expression in tumors and low expression in normal tissue [118]. Hsu et al. generated EphA2-CAR-T cells that effectively killed EphA2-positive osteosarcoma cell lines in vitro. And in subcutaneous osteosarcoma mouse models, EphA2-CAR-T cells induced significant reduction or elimination of osteosarcoma and extended survival in a dose and delivery route-dependent manner. Besides, EphA2-CAR-T cells showed superior efficacy for liver and lung metastatic osteosarcoma cells when delivered systemically [119].
IL-11Rα
Interleukin-11 (IL-11), a member of a family of pleiotropic cytokines [120], specifically binds to IL-11 receptor α-chain (IL-11Rα) and active the signaling pathways involved in adipogenesis, osteoclastogenesis, neurogenesis, and megakaryocyte maturation and platelet production [121]. The overexpression of IL-11Rα was found in several cancer types, including osteosarcoma [122]. Based on that, Huang et al. developed IL-11Rα-CAR-T cells and tested the tumor suppression activity on osteosarcoma in vitro and in vivo. IL-11Rα-CAR-T cells not only killed the osteosarcoma cell lines expressing IL-11Rα, but also results in the regression of osteosarcoma lung metastases. Besides, the cytotoxicity of IL-11Rα-CAR-T cells correlated with level of IL-11Ra expression on osteosarcoma cells [123].
IGF1R and ROR1
Insulin-like growth factor 1 receptor (IGF1R) is a tetrameric transmembrane receptor tyrosine kinase and widely expressed by several solid tumors and hematologic malignancies. The binding of ligand and IGF1R contributes to the proliferation, survival, transformation, metastasis, and angiogenesis [124]. Overexpression of tyrosine kinase-like orphan receptor 1 (ROR1) involved in tumor cell migration and invasiveness [125]. And the limited expression of ROR1 in normal tissues is another advantage for it to be an attractive therapeutic target in sarcomas [126]. The preclinical study conducted by Huang et al. demonstrated that IGF1R and ROR1 CAR-T cells exhibited significantly tumor growth inhibition and survival extension on sarcoma [127].
NKG2D
CAR-T cells therapies are increasingly attractive for various malignant tumor. However, the severe cytokine release syndrome (CRS) caused by T-cell activation and expansion limited the clinical application. To solve the problem, Fernandez et al. used memory T cells to express an CAR. They chose natural killer cell group 2D (NKG2D) as the target of CAR-T cells [128]. NKG2D receptor is heterogeneously expressed in primary and metastatic osteosarcoma cells, but is rarely expressed by healthy tissues [129]. CD45RA+memory T cells expressing an NKG2D CAR effectively killed the osteosarcoma cells in vitro and inhibited the tumor growth in mouse models of metastatic osteosarcoma with no toxicity to healthy tissues [130]. Similarly, to diminish the risk of CRS, Lu et al. developed a CAR-T adaptor molecule (CAM)-based therapy using a bispecific low-molecular weight ligand EC17 (FITC- folic acid). EC17 penetrates solid tumors within minutes and strongly bind with folate receptor (FR), whereas unbound EC17 will be rapidly cleared [131]. Administration of intermitte dosing and/or dose-titration of EC17 CAM could drive CAR-T cell activation, proliferation, and persistence and control the amount of cytokine releasing, which is a safety mechanism for the application of CAR-T cells. For FR+ osteosarcoma cells, EC17 controlled CAR-T cells showed potent anti-tumor activity in mice models [132].
CAR-M therapy
Macrophages, as the most abundant immune cell component infiltrating the osteosarcoma microenvironment, hold great potential as a tool for adoptive immunotherapy. Compared with other immune cells, one major advantage of using macrophages for ACT is the propensity in migration and infiltration into tumors [133]. CAR macrophage (CAR-M) studies are mainly at the nascent stage with one phase I trial ongoing which uses autologous CAR-M targeting HER2 overexpressing solid tumors [134]. Pre-clinical study showed that anti-CD19 and anti-HER2 CAR-Ms phagocytose antigen-bearing tumor cells in an antigen-specific manner in vitro. In lung metastases mouse models, CAR-M treatment significantly inhibited tumor growth and prolonged the overall survival of the tumor-bearing mice [134]. In addition, CAR-Ms promoted an inflammatory state within the TME and were capable of antigen cross-presentation to tumor-specific CD8+ T cells [135]. In osteosarcoma, CAR-M is a promising tool, which specifically recognize and eliminate tumor cells [136, 137]. Moreover, engineered CAR-Ms have leaded to increase of anti-tumor cytokines, such as IL-6, and chemokines, such as CXCL18, in the TME of osteosarcoma [138, 139]. The production of these beneficial cytokines fosters help to convert cold tumors into hot tumors [140].
CAR-NK therapy
As previously mentioned, NK cells possess potent anti-osteosarcoma effects, but NK cells lack the appropriate machinery to recognize tumor antigens without binding Fc-γ receptors through CD16 to IgG-coated targets. When the NK cells are loaded a CAR targeting specific osteosarcoma antigen, this limitation can be overcome [141]. Currently, there are a number of clinical trials involving CAR-NK that have been planned or are ongoing. All these clinical trials are at the phase I/II trial stage [142]. At present, the early clinical trial results have not yet demonstrated consistent efficacy and safety. The trial in adults with chronic lymphocytic leukemia demonstrated that CAR-NK targeting CD19 are safe and effective [143]. However, the results from studies on targets other than CD19 in CAR-NK have been less than satisfactory, including both in vivo studies and clinical research. GD2-specific CAR-NK cells for Ewing sarcoma failed to eliminate GD2-positive sarcoma in xenografts models [71]. NK cells have a relatively short half-life, typically less than 10 days [144], which is a double-edge sword in CAR-NK therapy. This confers an advantage in case severe toxicity occurs, but also creates a challenge that repeated administrations may be needed to achieve durable response. Compared with CAR-T cells, CAR-NK therapy show several advantages, including (1) the ability to be derived from established cell lines or allogeneic NK cells without the major histocompatibility complex (MHC) restriction; (2) CAR-NK cells eliminate cancer cells through both CAR-dependent and CAR-independent manners; (3) reduced toxicity, particularly in terms of cytokine-release syndrome and neurotoxicity.
So far, CAR-NK and CAR-M therapies have not made promising progress in the research of osteosarcoma. It need to point out, these two therapies could theoretically compensate for the shortcomings of CAR-T treatment, but there is still a long way to go before their clinical application. At present, CAR-T has shown clear anti-tumor effects in preclinical studies for osteosarcoma, and a large number of clinical trials are still ongoing, with GD2, HER2, and B7-H3 as the main targets (Table 2). Compared to other cellular adoptive therapies, CAR-T research in osteosarcoma may achieve breakthrough progress first. We are looking forward to having inspiring results reported in the future.
Other immunotherapies for osteosarcoma
GD2 and HER2 as candidate target antigens due to their high expression level on osteosarcoma cells. However, clinical trials of trastuzumab (anti-HER2) or dinutuximab (anti-GD2) for metastatic or refractory osteosarcoma were not successful [145, 146]. Therefore, Park et al. constructed the bispecific antibodies (BsAbs) targeting CD3 and GD2 or HER2. GD2-BsAb and HER2-BsAb successfully directed T cells into tumor tissues and exerted potent anti-tumor activity against osteosarcoma. Besides, PD-1/PD-L1 blockade significantly enhanced anti-tumor activity of GD2-BsAb and HER2-BsAb [147]. Currently, only a small number of clinical studies of BsAbs have been conducted in osteosarcoma, summarized in Table 3. Besides, Mason et al. performed a phase I trial in canine osteosarcoma to evaluate the anti-tumor activity of recombinant Listeria vaccines expressing a chimeric human HER2/neu fusion protein (ADXS31-164). They found that ADXS31-164 induced HER2/neu-specifific immunity and reduced the incidence of metastatic disease and prolonged OS [148]. DCs are the most potent professional antigen-presenting cells and play a crucial role in the osteosarcoma microenvironment. Enhancing the anti-tumor capabilities of DCs holds promise as a novel therapeutic approach for osteosarcoma. Kawano et al. suggested that combined DCs and anti-TGF-β antibodies to treat osteosarcoma and detected enhanced systematic immune responses in mice models [149]. The synergistic action of DCs and agonistic antibodies against the glucocorticoid-induced tumor necrosis factor receptor (anti-GITR) can significantly amplify the systemic immune response. This strategy not only facilitates the elimination of regulatory T cells but also effectively inhibites osteosarcoma growth in vivo [150]. A variety of vaccines have shown encouraging results, including the CD1c+ DC vaccine, which harnesses the power of antigen-presenting DCs, and the innovative approach of vaccinating with polyinosinic: polycytidylic acid (poly I: C). This treatment activates and loads tumor antigens onto CD103+ myeloid/conventional DC1s, enhancing the immune system’s capacity to target cancer cells [151].
Research on immunotherapy for osteosarcoma is mostly in the early stages, and current results show that these treatment strategies have some therapeutic potential, but further validation and improvement are still needed. We present the current immunotherapy in osteosarcoma in Fig. 2.

The main immunotherapy strategies in osteosarcoma. In osteosarcoma, the relatively well-studied immunotherapies include: immune checkpoint inhibitors (with targets such as PD-1/PD-L1, CTLA-4, TIM-3, CD47 and TIGIT), adoptive cell therapies (such as CAR-T, CAR-M and CAR-NK), dendritic cell vaccines, and bispecific antibodies (targeting HER2 or GD2 and CD3). TAA: tumor-associated antigen
Combination therapy strategies
Combination of anti-PD-1/PD-L1 and anti-CTLA-4 treatment
Due to the presence of various immunosuppressive pathways in the microenvironment of osteosarcoma, treatment with a single-target ICI has not achieved satisfactory results. Like other solid tumors, trying a combination therapy of two ICIs in osteosarcoma may have better anti-tumor effects. At present, the combined application of ICIs is still mainly focused on classic anti-PD-1/PD-L1 and anti-CTLA-4 antibodies. Although combination therapy may bring breakthroughs in efficacy, its side effects also need to be paid more attention.
A study demonstrated that anti-PD-L1 treatment for metastatic osteosarcoma upregulate additional inhibitory receptors including CTLA-4, which contributed to the resistance of anti-PD-L1 therapy. Therefore, a combination therapy of anti-PD-L1 with anti-CTLA-4 antibody completely controlled metastatic osteosarcoma in an osteosarcoma mouse model [152]. A case report demonstrated that the combination of an anti-PD-1 antibody (nivolumab) and an anti-CTLA-4 antibody (ipilimumab) inhibited the progression of metastatic tumor for a young man with a metastatic osteosarcoma not responsive to several lines of standard chemotherapy. Before the application of dual checkpoint inhibition treatment, they detected the amplification of PD-L1 and PD-L2 in resected tumor tissue through next-generation sequencing. Thus it can be seen, it is important that identifying biomarkers to select patients with high response rates to immunotherapy [153]. The trial compared the efficacy of anti-PD-1 antibody only and the combination therapy of anti-PD-1 antibody plus anti-CTLA-4 antibody. Enrolled patients were allocated to receive either nivolumab monotherapy (43 patients) or nivolumab plus ipilimumab (42 patients). Nivolumab monotherapy showed limited efficacy for patients with advanced sarcoma. Nivolumab combined with ipilimumab demonstrated promising response rate with a manageable safety profile, which need further study [154]. 62 patients diagnosed as advanced or metastatic sarcoma were enrolled into the clinical trial (NCT02815995) to evaluate the safety and efficacy of the combination of durvalumab (anti-PD-L1 antibody) and tremelimumab anti-CTLA-4 antibody) [155]. Patients received 1500 mg intravenous durvalumab and 75 mg intravenous tremelimumab for four cycles, followed by durvalumab alone every 4 weeks for up to 12 months. The PFS at 12 weeks was 49%. Grade 3–4 treatment-related adverse events were reported in 21 patients [155]. It can be supposed that the combination therapy of different immune checkpoint inhibitors is a promising option for patients with osteosarcoma who have progressed after multi line therapy.
Combination of anti-PD-1/PD-L1 and anti-angiogenic treatment
Angiogenesis is significantly correlated with tumor progression and is an important part of the formation of the immunosuppressive microenvironment in osteosarcoma. Combining anti-angiogenic therapy with immune checkpoint inhibitor treatment is beneficial for lifting immunosuppression. At the same time, anti-angiogenic therapy promotes the normalization of tumor blood vessels, helping more anti-tumor drugs and immune cells to enter the tumor microenvironment and exert their effects.
Immunosuppression promoted by tumor angiogenesis, and more immune cells are related with angiogenesis [156]. Therefore, simultaneously targeting tumor blood vessels and antagonizing immune suppression may propose new therapeutic strategies for sarcoma. At 2020, Martin-Broto et al. reported the results of a phase Ib/II trial evaluating nivolumab (anti-PD-1 antibody) and sunitinib (inhibitor of anti-angiogenesis) combination in advanced soft tissue sarcomas (NCT03277924). 16 sarcoma patients (including osteosarcoma) entered into phase Ib study, and the results determined the recommended dose of sunitinib for phase II was 37.5 mg as induction and then 25 mg in combination with nivolumab. The 6-month PFS rate for soft-tissue sarcoma patients was 48% (95% CI: 41–55%). The most frequent treatment-related toxicities were fatigue (63.5%) and increased aspartate aminotransferase (48.0%). The most common grade 3 or 4 side effects were transaminitis (17.3%) and neutropenia (11.5%) [157]. We look forward to the publication of overall survival data of soft-tissue sarcoma patients and research results on osteosarcoma cohort. Yao et al. conducted a phase I trial (NCT04074564) to assess the safety and efficacy of multi-antigen stimulated cell therapy-I in combination with camrelizumab (anti-PD-1 antibody) and apatinib (a highly selective inhibitor targeting VEGFR2) in patients with unresectable recurrent or metastatic bone and soft-tissue sarcoma who had previously undergone at least one line of systemic therapy. Totally, 19 patients, including 6 osteosarcoma patients, were enrolled into the study. For the 6 patients with osteosarcoma, the ORR was 33.3%, the disease control response (DCR) was 50.0%, and median PFS was 5.7 months. The most common treatment-related adverse event was decreased neutrophil count. 11 (57.9%) experienced grade ≥ 3 treatmentrelated adverse events. No treatment-related deaths occurred [158]. Besides, Xie et al. reported the results of a single-arm phase II trial evaluating the safety and efficacy of apatinib (anti-VEGFR2 inhibitor) plus camrelizumab (anti-PD-1 antibody) in patients with chemotherapy-refractory osteosarcoma (NCT03359018). 43 patients enrolled this trial, and the ORR was 20.9% and two patients with durable disease control were observed. 13 of 43 patients were progression free at 6 months and the 6-month PFS rate was 50.9% [159]. In addition to ICIs, Park et al. conducted the preclinical study demonstrating that VEGF blockade enhanced the infiltration of tumor-antigens (GD2/HER2/GPC3) armed T cells into tumor microenvironment and increased CD8+ T cells survival and dispersion [160].
Combination of target-specific and non target-specific treatment
Target-specific therapies are notable for their strong targeting capabilities and significant effects, while non target-specific therapies enhance the overall immune response to fight cancer, potentially offering a broader range of applications. Non target-specific immune response, such as vaccine, DCs, IL-2 and TGF-β, combined with target-specific immunotherapy may exhibit stronger anti-tumor efficacy.
A study examined the anti-tumor efficacy of the combination of tumor lysate-pulsed DCs and CTLA-4 antibody in a C3H mouse osteosarcoma model. The combination therapy potently inhibited the accumulation of Tregs and promoted the infiltration of CD8+ T cells inside the metastatic settings. The synergistic effect of tumor lysate-pulsed DCs and CTLA-4 blockade is manifested in inhibiting the growth of metastatic tumors and prolonging survival time [161]. Guo et al. developed a new bifunctional fusion protein composed of a monoclonal antibody against PD-L1 fused with the extracellular domain of TGF-β receptor. However, the results of the phase Ib study showed no significantly improvement of the ORR in patients with recurrent osteosarcoma, while the side effects are acceptable [162]. Bempegaldesleukin, a CD122-preferential IL-2 pathway agonist, is associated with increased proliferation and activation of TILs among patients with solid tumors [163]. Bempegaldesleukin may be beneficial for optimizing the responsiveness of metastatic sarcoma to checkpoint blockade. Based on that, a study was conducted to evaulate the safety and efficacy of bempegaldesleukin combination with nivolumab for 84 patients with selected locally advanced or metastatic high-grade sarcoma, including 10 osteosarcoma patients. The combination strategy showed limited activity for most sarcoma. In other sarcoma subtypes, CD8+ T cell infiltration, PD-1 expression and upregulation of immunerelated pathways correlated with improved ORR. However, in osteosarcoma, although it has a higher tumor mutational burden, there is no significant response to the combination therapy [164].
As mentioned earlier, T cells are edited to express specific tumor antigens, allowing them to recognize and kill tumor cells. GD2-CAR-T therapy is being explored in extensive research in osteosarcoma, but the survival time of CAR-T cells in body is indeed limited. When T cells recognize viruses, they can exist in the body for many years. And when the same virus or virus vaccine enters the body again, these T cells can be quickly activated and cleared of the virus. Tanaka et al. generated the GD2-CAR-modified varicella zoster virus (VZV)-specific T cells and conducted a phase I trail (NCT01953900) to evaluate the activity of combined application of CAR-T cells and vaccine in advanced osteosarcoma and neuroblastoma [165]. At present, the results of this study have not been reported. It is hoped that through the multifaceted exploration of researchers, strategy can be found to control tumors for osteosarcoma patients, especially those with recurrent or metastatic disease.
Immunotherapy-combination stratagies have certain therapeutic potential, and discovering and verifying effective treatment combinations that achieve a “1 + 1 > 2” synergistic effect remains challenging. The exploration of advantageous combinations for immunotherapy-combination stratagy is a long and arduous task.
Conclusions and future directions
Osteosarcoma is the most common primary bone malignancy for children and young adults. The current standard care has improved the prognosis of patients with localized osteosarcoma, but the overall survival has not substantially improved over the past decades. For metastatic or relapsed osteosarcoma, even active treatment results in a 5-year survival rate of 20%. Therefore, it is urgent to find new and effective strategies to further improve the survival rate of osteosarcoma patients. The immunosuppressive TME of osteosarcoma, especially the lung metastatic foci, is infiltrated with a variety of cells that promote immunosuppression and express numerous immunosuppressive molecules, providing opportunities for the application of immunotherapy in osteosarcoma. However, it is precisely due to the unique inhibitory immune microenvironment of osteosarcoma, the responsiveness to immunotherapy is unsatisfactory. The immune checkpoint blockers (such as anti-PD-1/PD-L1 antibody and anti-CTLA-4 antibody) treatment only showed a limited therapeutic effect on osteosarcoma. CAR-T therapy in osteosarcoma is still in early stages of research. CAR-T therapy has not yet achieved the same success in clinical trials for osteosarcoma as it has in hematological malignancies, and other CAR-engineered adoptive cell therapies, such as CAR-NK and CAR-M, are still in the preclinical research phase. Based on current preclinical and clincial data on immunotherapy, there is still a long way to go for the application of immunotherapy in osteosarcoma. We need to explore more other biomarkers to predict the efficacy of immunotherapy in patients and choose the subpopulations that will be beneficial from immunotherapy. In addition, a single immunotherapy has low response rate in osteosarcoma. Combination therapy based on immunotherapy that converting a immune cold into an inflamed microenvironment may be a future direction.
Data availability
No datasets were generated or analysed during the current study.
Abbreviations
- PFS:
-
Progression-free survival
- ORR:
-
Objective response rate
- ICIs:
-
Immune checkpoint inhibitors
- CAR-T:
-
Chimeric antigen receptor T
- ACT:
-
Adoptive cell therapy
- TME:
-
Tumor microenvironment
- TAMs:
-
Tumor-associated macrophages
- TANs:
-
Tumor-associated neutrophils
- MDSCs:
-
Myeloid-derived suppressor cells
- NK cells:
-
Natural killer cells
- DCs:
-
Dendritic cells
- CTCs:
-
Circulating tumor cells
- TGF-β:
-
Transforming growth factor-beta
- Arg-1:
-
Arginase-1
- iNOS:
-
Inducible nitric oxide synthase
- ROS:
-
Reactive oxygen species
- VEGF:
-
Vascular endothelial growth factor
- HGF:
-
Hepatocyte growth factor
- EMT:
-
Epithelial-mesenchymal transition
- TIM-3:
-
T-cell immunoglobulin and mucin domain-containing protein 3
- TIGIT:
-
T cell immunoreceptor with Ig and ITIM domains
- NETs:
-
Neutrophil extracellular traps
- CSCs:
-
Cancer stem cells
- MSCs:
-
Mesenchymal stem cells
- EVs:
-
Extracellular vesicles
- IFN-γ:
-
Interferon-γ
- TILs:
-
Tumor-infiltrating lymphocytes
- APCs:
-
Antigen-presenting cells
- TNF-α:
-
Tumor necrosis factor-α
- CTLs:
-
Cytotoxic T lymphocytes
- C1Q:
-
Complete component 1q
- NF-kB:
-
Nuclear factor-kappa B
- STAT3:
-
Signal transducer and activator of transcription 3
- LAG-3:
-
Lymphocyteactivation gene 3
- IDO1:
-
Indoleamine 2,3-dioxygenase
- OS:
-
Overall survival
- DFS:
-
Disease free survival
- PR:
-
Partial response
- CTLA-4:
-
Cytotoxic T lymphocyte-associated antigen-4
- Th1:
-
Type 1 T helper
- Gal-9:
-
Galectin-9
- HER2:
-
Human epidermal growth factor receptor-2
- GD2:
-
Disialogangliosides
- ALCAM:
-
Activated leukocyte cell adhesion molecule
- PDXs:
-
Patientderived xenografts
- IL-11:
-
Interleukin-11
- IL-11Rα:
-
IL-11 receptor α-chain
- IGF1R:
-
Insulin-like growth factor 1 receptor
- ROR1:
-
Tyrosine kinase-like orphan receptor 1
- CRS:
-
Cytokine release syndrome
- NKG2D:
-
Natural killer cell group 2D
- CAM:
-
CART adaptor molecule
- FR:
-
Folate receptor
- CAR-M:
-
CAR macrophage
- MHC:
-
Major histocompatibility complex
- BsAbs:
-
Bispecific antibodies
- GITR:
-
Glucocorticoid-induced tumor necrosis factor receptor
- DCR:
-
Disease control response
- VZV:
-
Varicella zoster virus
References
Beird HC, et al. Osteosarcoma Nat Rev Dis Primers. 2022;8(1):77.
Czarnecka AM et al. Molecular Biology of Osteosarcoma. Cancers (Basel), 2020. 12(8).
Isakoff MS, et al. Osteosarcoma: current treatment and a collaborative pathway to Success. J Clin Oncol. 2015;33(27):3029–35.
Zhou Y, et al. Single-cell RNA landscape of intratumoral heterogeneity and immunosuppressive microenvironment in advanced osteosarcoma. Nat Commun. 2020;11(1):6322.
Anderson ME. Update on Survival in Osteosarcoma. Orthop Clin North Am. 2016;47(1):283–92.
Arndt CA, et al. Common musculoskeletal tumors of childhood and adolescence. Mayo Clin Proc. 2012;87(5):475–87.
Bacci G, et al. High grade osteosarcoma of the extremities with lung metastases at presentation: treatment with neoadjuvant chemotherapy and simultaneous resection of primary and metastatic lesions. J Surg Oncol. 2008;98(6):415–20.
Sayles LC, et al. Genome-informed targeted therapy for Osteosarcoma. Cancer Discov. 2019;9(1):46–63.
Duffaud F, et al. Efficacy and safety of regorafenib in adult patients with metastatic osteosarcoma: a non-comparative, randomised, double-blind, placebo-controlled, phase 2 study. Lancet Oncol. 2019;20(1):120–33.
Long GV, et al. Overall survival and response with Nivolumab and Relatlimab in Advanced Melanoma. NEJM Evid. 2023;2(4):EVIDoa2200239.
Reck M, et al. Pembrolizumab versus Chemotherapy for PD-L1-Positive non-small-cell Lung Cancer. N Engl J Med. 2016;375(19):1823–33.
Ferris RL, et al. Nivolumab for Recurrent Squamous-Cell Carcinoma of the Head and Neck. N Engl J Med. 2016;375(19):1856–67.
Grupp SA, et al. Chimeric antigen receptor-modified T cells for acute lymphoid leukemia. N Engl J Med. 2013;368(16):1509–18.
Porter DL, et al. Chimeric antigen receptor T cells persist and induce sustained remissions in relapsed refractory chronic lymphocytic leukemia. Sci Transl Med. 2015;7(303):303ra139.
Kochenderfer JN, et al. Chemotherapy-refractory diffuse large B-cell lymphoma and indolent B-cell malignancies can be effectively treated with autologous T cells expressing an anti-CD19 chimeric antigen receptor. J Clin Oncol. 2015;33(6):540–9.
Liang H, et al. Advancements in osteosarcoma management: integrating immune microenvironment insights with immunotherapeutic strategies. Front Cell Dev Biol. 2024;12:1394339.
Zheng Y, et al. Mesenchymal stem cells in the osteosarcoma microenvironment: their biological properties, influence on tumor growth, and therapeutic implications. Stem Cell Res Ther. 2018;9(1):22.
Liu T, et al. Self-seeding circulating tumor cells promote the proliferation and metastasis of human osteosarcoma by upregulating interleukin-8. Cell Death Dis. 2019;10(8):575.
Huang Q, et al. The role of tumor-associated macrophages in osteosarcoma progression - therapeutic implications. Cell Oncol (Dordr). 2021;44(3):525–39.
Duluc D, et al. Interferon-gamma reverses the immunosuppressive and protumoral properties and prevents the generation of human tumor-associated macrophages. Int J Cancer. 2009;125(2):367–73.
Luo ZW, et al. Macrophages in Osteosarcoma Immune Microenvironment: implications for Immunotherapy. Front Oncol. 2020;10:586580.
He Z, Zhang S. Tumor-Associated macrophages and their Functional Transformation in the hypoxic Tumor Microenvironment. Front Immunol. 2021;12:741305.
Shao XJ, et al. Inhibition of M2-like macrophages by all-trans retinoic acid prevents cancer initiation and stemness in osteosarcoma cells. Acta Pharmacol Sin. 2019;40(10):1343–50.
Aran D, et al. Reference-based analysis of lung single-cell sequencing reveals a transitional profibrotic macrophage. Nat Immunol. 2019;20(2):163–72.
Herroon MK, et al. Bone marrow adipocytes promote tumor growth in bone via FABP4-dependent mechanisms. Oncotarget. 2013;4(11):2108–23.
Tang F, et al. Surgical Treatment of Osteosarcoma Induced Distant Pre-metastatic Niche in Lung to facilitate the colonization of circulating Tumor cells. Adv Sci (Weinh). 2023;10(28):e2207518.
Haist M et al. The functional crosstalk between Myeloid-Derived Suppressor Cells and Regulatory T Cells within the immunosuppressive Tumor Microenvironment. Cancers (Basel), 2021. 13(2).
Ran S, Wilber A. Novel role of immature myeloid cells in formation of new lymphatic vessels associated with inflammation and tumors. J Leukoc Biol. 2017;102(2):253–63.
Gabrilovich DI. Myeloid-derived suppressor cells. Cancer Immunol Res. 2017;5(1):3–8.
Umansky V et al. The role of myeloid-derived suppressor cells (MDSC) in Cancer Progression. Vaccines (Basel), 2016. 4(4).
Tsubakihara Y, Moustakas A. Epithelial-mesenchymal transition and metastasis under the control of transforming growth factor beta. Int J Mol Sci, 2018. 19(11).
Wang C, et al. CD300ld on neutrophils is required for tumour-driven immune suppression. Nature. 2023;621(7980):830–9.
Ligon JA et al. Pathways of immune exclusion in metastatic osteosarcoma are associated with inferior patient outcomes. J Immunother Cancer, 2021. 9(5).
Li F, et al. Interaction gene set between osteoclasts and regulatory CD4(+) T cells can accurately predict the prognosis of patients with osteosarcoma. Cancer Sci. 2023;114(7):3014–26.
Li X, et al. Tim3/Gal9 interactions between T cells and monocytes result in an immunosuppressive feedback loop that inhibits Th1 responses in osteosarcoma patients. Int Immunopharmacol. 2017;44:153–9.
Lucca LE, Dominguez-Villar M. Modulation of regulatory T cell function and stability by co-inhibitory receptors. Nat Rev Immunol. 2020;20(11):680–93.
Wu L et al. Tumor-Associated neutrophils in Cancer: going pro. Cancers (Basel), 2019. 11(4).
Yang B, et al. Identification of prognostic biomarkers associated with metastasis and immune infiltration in osteosarcoma. Oncol Lett. 2021;21(3):180.
Fu Y, et al. Development and validation of a Hypoxia-Associated Prognostic signature related to Osteosarcoma Metastasis and Immune Infiltration. Front Cell Dev Biol. 2021;9:633607.
Papayannopoulos V. Neutrophil extracellular traps in immunity and disease. Nat Rev Immunol. 2018;18(2):134–47.
Lin Y, et al. Development and validation of neutrophil extracellular traps-derived signature to predict the prognosis for osteosarcoma patients. Int Immunopharmacol. 2024;127:111364.
Tang H, et al. Osteosarcoma neutrophil extracellular trap network-associated gene recurrence and metastasis model. J Cancer Res Clin Oncol. 2024;150(2):48.
Brown HK, Tellez-Gabriel M, Heymann D. Cancer stem cells in osteosarcoma. Cancer Lett. 2017;386:189–95.
Chang AI, et al. Involvement of mesenchymal stem cells in cancer progression and metastases. Curr Cancer Drug Targets. 2015;15(2):88–98.
Chang X, et al. New perspective into mesenchymal stem cells: molecular mechanisms regulating osteosarcoma. J Bone Oncol. 2021;29:100372.
Lagerweij T, Perez-Lanzon M, Baglio SR. A preclinical mouse model of Osteosarcoma to define the Extracellular vesicle-mediated communication between Tumor and mesenchymal stem cells. J Vis Exp, 2018(135).
Khare D, et al. Mesenchymal stromal cell-derived exosomes affect mRNA expression and function of B-Lymphocytes. Front Immunol. 2018;9:3053.
Zhang Q, et al. Exosomes originating from MSCs stimulated with TGF-beta and IFN-gamma promote Treg differentiation. J Cell Physiol. 2018;233(9):6832–40.
Jia XH, et al. Activation of mesenchymal stem cells by macrophages promotes tumor progression through immune suppressive effects. Oncotarget. 2016;7(15):20934–44.
Wang JW, et al. Exosomal miR-1228 from Cancer-Associated fibroblasts promotes Cell Migration and Invasion of Osteosarcoma by directly targeting SCAI. Oncol Res. 2019;27(9):979–86.
Mazumdar A et al. Osteosarcoma-Derived Extracellular vesicles induce lung fibroblast reprogramming. Int J Mol Sci, 2020. 21(15).
Xu Y, et al. Osteosarcoma cells secrete CXCL14 that activates integrin alpha11beta1 on fibroblasts to form a lung metastatic niche. Cancer Res. 2024;84(7):994–1012.
Wolf-Dennen K, Gordon N, Kleinerman ES. Exosomal communication by metastatic osteosarcoma cells modulates alveolar macrophages to an M2 tumor-promoting phenotype and inhibits tumoricidal functions. Oncoimmunology. 2020;9(1):1747677.
Corre I et al. The Osteosarcoma Microenvironment: a Complex but Targetable Ecosystem. Cells, 2020. 9(4).
Han Q, Shi H, Liu F. CD163(+) M2-type tumor-associated macrophage support the suppression of tumor-infiltrating T cells in osteosarcoma. Int Immunopharmacol. 2016;34:101–6.
Sundara YT, et al. Increased PD-L1 and T-cell infiltration in the presence of HLA class I expression in metastatic high-grade osteosarcoma: a rationale for T-cell-based immunotherapy. Cancer Immunol Immunother. 2017;66(1):119–28.
Sun CY, et al. T cell exhaustion drives osteosarcoma pathogenesis. Ann Transl Med. 2021;9(18):1447.
Le T, Su S, Shahriyari L. Immune classification of osteosarcoma. Math Biosci Eng. 2021;18(2):1879–97.
Collin M, Bigley V. Human dendritic cell subsets: an update. Immunology. 2018;154(1):3–20.
Le T et al. Data-Driven Mathematical Model of Osteosarcoma. Cancers (Basel), 2021. 13(10).
Zhang C, et al. Profiles of immune cell infiltration and immune-related genes in the tumor microenvironment of osteosarcoma. Aging. 2020;12(4):3486–501.
Jones KB. Dendritic cells drive osteosarcomagenesis through newly identified Oncogene and Tumor suppressor. Cancer Discov. 2019;9(11):1484–6.
Kansara M, et al. Infiltrating myeloid cells drive Osteosarcoma Progression via GRM4 regulation of IL23. Cancer Discov. 2019;9(11):1511–9.
He YT, et al. In vitro generation of cytotoxic T lymphocyte response using dendritic cell immunotherapy in osteosarcoma. Oncol Lett. 2016;12(2):1101–6.
Himoudi N, et al. Lack of T-cell responses following autologous tumour lysate pulsed dendritic cell vaccination, in patients with relapsed osteosarcoma. Clin Transl Oncol. 2012;14(4):271–9.
Miwa S, et al. Phase 1/2 study of immunotherapy with dendritic cells pulsed with autologous tumor lysate in patients with refractory bone and soft tissue sarcoma. Cancer. 2017;123(9):1576–84.
Prager I, Watzl C. Mechanisms of natural killer cell-mediated cellular cytotoxicity. J Leukoc Biol. 2019;105(6):1319–29.
Zhang ML, et al. PD–L1/PD–1 axis serves an important role in natural killer cell–induced cytotoxicity in osteosarcoma. Oncol Rep. 2019;42(5):2049–56.
Lazarova M, Steinle A. Impairment of NKG2D-Mediated tumor immunity by TGF-beta. Front Immunol. 2019;10:2689.
Chauvin JM, Zarour HM. TIGIT in cancer immunotherapy. J Immunother Cancer, 2020. 8(2).
Kailayangiri S, et al. Targeting Ewing sarcoma with activated and GD2-specific chimeric antigen receptor-engineered human NK cells induces upregulation of immune-inhibitory HLA-G. Oncoimmunology. 2017;6(1):e1250050.
Sarvaria A, Madrigal JA, Saudemont A. B cell regulation in cancer and anti-tumor immunity. Cell Mol Immunol. 2017;14(8):662–74.
Li GQ, et al. Application of Immune Infiltration Signature and machine learning model in the Differential diagnosis and prognosis of Bone-Related Malignancies. Front Cell Dev Biol. 2021;9:630355.
Tu J, et al. Single-cell RNA datasets and bulk RNA datasets analysis demonstrated C1Q + tumor-associated macrophage as a major and antitumor immune cell population in osteosarcoma. Front Immunol. 2023;14:911368.
Wu CC, et al. Immuno-genomic landscape of osteosarcoma. Nat Commun. 2020;11(1):1008.
Wu CC, Livingston JA. Genomics and the Immune Landscape of Osteosarcoma. Adv Exp Med Biol. 2020;1258:21–36.
Larionova I, et al. Interaction of tumor-associated macrophages and cancer chemotherapy. Oncoimmunology. 2019;8(7):1596004.
Wan B, et al. Analysis of Immune Gene expression subtypes reveals Osteosarcoma Immune Heterogeneity. J Oncol. 2021;2021:p6649412.
Hong W, et al. Immune-related prognosis biomarkers associated with osteosarcoma microenvironment. Cancer Cell Int. 2020;20:83.
Hu C, et al. Comprehensive analysis of prognostic tumor microenvironment-related genes in osteosarcoma patients. BMC Cancer. 2020;20(1):814.
Sheng G, et al. Osteosarcoma Metastasis Front Oncol. 2021;11:780264.
Lussier DM, et al. Enhanced T-cell immunity to osteosarcoma through antibody blockade of PD-1/PD-L1 interactions. J Immunother. 2015;38(3):96–106.
Zheng B, et al. PD-1 axis expression in musculoskeletal tumors and antitumor effect of nivolumab in osteosarcoma model of humanized mouse. J Hematol Oncol. 2018;11(1):16.
Davis KL, et al. Nivolumab in children and young adults with relapsed or refractory solid tumours or lymphoma (ADVL1412): a multicentre, open-label, single-arm, phase 1–2 trial. Lancet Oncol. 2020;21(4):541–50.
Tawbi HA, et al. Pembrolizumab in advanced soft-tissue sarcoma and bone sarcoma (SARC028): a multicentre, two-cohort, single-arm, open-label, phase 2 trial. Lancet Oncol. 2017;18(11):1493–501.
Boye K, et al. Pembrolizumab in advanced osteosarcoma: results of a single-arm, open-label, phase 2 trial. Cancer Immunol Immunother. 2021;70(9):2617–24.
Le Cesne A, et al. Programmed cell death 1 (PD-1) targeting in patients with advanced osteosarcomas: results from the PEMBROSARC study. Eur J Cancer. 2019;119:151–7.
Liu Z, et al. Targeting autophagy enhances atezolizumab-induced mitochondria-related apoptosis in osteosarcoma. Cell Death Dis. 2021;12(2):164.
Shemesh CS, et al. Population pharmacokinetics, exposure-safety, and immunogenicity of atezolizumab in pediatric and young adult patients with cancer. J Immunother Cancer. 2019;7(1):314.
Geoerger B, et al. Atezolizumab for children and young adults with previously treated solid tumours, non-hodgkin lymphoma, and Hodgkin lymphoma (iMATRIX): a multicentre phase 1–2 study. Lancet Oncol. 2020;21(1):134–44.
Egen JG, Kuhns MS, Allison JP. CTLA-4: new insights into its biological function and use in tumor immunotherapy. Nat Immunol. 2002;3(7):611–8.
He J, et al. Association between CTLA-4 genetic polymorphisms and susceptibility to osteosarcoma in Chinese Han population. Endocrine. 2014;45(2):325–30.
Liu J, et al. Effect of cytotoxic T-lymphocyte antigen-4, TNF-alpha polymorphisms on osteosarcoma: evidences from a meta-analysis. Chin J Cancer Res. 2013;25(6):671–8.
Merchant MS, et al. Phase I clinical trial of Ipilimumab in Pediatric patients with Advanced Solid tumors. Clin Cancer Res. 2016;22(6):1364–70.
Tsai RK, Discher DE. Inhibition of self engulfment through deactivation of myosin-II at the phagocytic synapse between human cells. J Cell Biol. 2008;180(5):989–1003.
Xu JF, et al. CD47 blockade inhibits tumor progression human osteosarcoma in xenograft models. Oncotarget. 2015;6(27):23662–70.
Wen Y, et al. Immune checkpoints in osteosarcoma: recent advances and therapeutic potential. Cancer Lett. 2022;547:215887.
Zhao L, et al. TIM-3: an update on immunotherapy. Int Immunopharmacol. 2021;99:107933.
Ge W, et al. Tim-3 as a diagnostic and prognostic biomarker of osteosarcoma. Tumour Biol. 2017;39(7):1010428317715643.
Koirala P, et al. HHLA2, a member of the B7 family, is expressed in human osteosarcoma and is associated with metastases and worse survival. Sci Rep. 2016;6:31154.
DeRenzo C, Gottschalk S. Genetically modified T-Cell therapy for Osteosarcoma: into the Roaring 2020s. Adv Exp Med Biol. 2020;1257:109–31.
Tabak SA, Khalifa SE, Fathy Y. HER-2 immunohistochemical expression in bone sarcomas: a New Hope for Osteosarcoma patients. Open Access Maced J Med Sci. 2018;6(9):1555–60.
Wang SL, et al. Prognostic significance of the expression of HER family members in primary osteosarcoma. Oncol Lett. 2018;16(2):2185–94.
Colomer R, et al. Herceptin: from the bench to the clinic. Cancer Invest. 2001;19(1):49–56.
Ahmed N, et al. Immunotherapy for osteosarcoma: genetic modification of T cells overcomes low levels of tumor antigen expression. Mol Ther. 2009;17(10):1779–87.
Rainusso N, et al. Immunotherapy targeting HER2 with genetically modified T cells eliminates tumor-initiating cells in osteosarcoma. Cancer Gene Ther. 2012;19(3):212–7.
Ahmed N, et al. Human epidermal growth factor receptor 2 (HER2) -Specific chimeric Antigen receptor-modified T cells for the Immunotherapy of HER2-Positive sarcoma. J Clin Oncol. 2015;33(15):1688–96.
Picci P, et al. Relationship of chemotherapy-induced necrosis and surgical margins to local recurrence in osteosarcoma. J Clin Oncol. 1994;12(12):2699–705.
Charan M, et al. GD2-directed CAR-T cells in combination with HGF-targeted neutralizing antibody (AMG102) prevent primary tumor growth and metastasis in ewing sarcoma. Int J Cancer. 2020;146(11):3184–95.
Chulanetra M, et al. GD2 chimeric antigen receptor modified T cells in synergy with sub-toxic level of doxorubicin targeting osteosarcomas. Am J Cancer Res. 2020;10(2):674–87.
Federman N, et al. Enhanced growth inhibition of osteosarcoma by cytotoxic polymerized liposomal nanoparticles targeting the alcam cell surface receptor. Sarcoma. 2012;2012:p126906.
Wang Y, et al. Anti-CD166/4-1BB chimeric antigen receptor T cell therapy for the treatment of osteosarcoma. J Exp Clin Cancer Res. 2019;38(1):168.
Picarda E, Ohaegbulam KC, Zang X. Molecular pathways: Targeting B7-H3 (CD276) for Human Cancer Immunotherapy. Clin Cancer Res. 2016;22(14):3425–31.
Tekle C, et al. B7-H3 contributes to the metastatic capacity of melanoma cells by modulation of known metastasis-associated genes. Int J Cancer. 2012;130(10):2282–90.
Majzner RG, Cells Targeting CART, et al. B7-H3, a Pan-cancer Antigen, Demonstrate Potent Preclinical Activity against Pediatric Solid Tumors and Brain tumors. Clin Cancer Res. 2019;25(8):2560–74.
Zhang Q, et al. B7-H3 targeted CAR-T cells show highly efficient anti-tumor function against osteosarcoma both in vitro and in vivo. BMC Cancer. 2022;22(1):1124.
Fritsche-Guenther R, et al. De novo expression of EphA2 in osteosarcoma modulates activation of the mitogenic signalling pathway. Histopathology. 2010;57(6):836–50.
Posthumadeboer J, et al. Surface proteomic analysis of osteosarcoma identifies EPHA2 as receptor for targeted drug delivery. Br J Cancer. 2013;109(8):2142–54.
Hsu K, et al. Chimeric Antigen receptor-modified T cells targeting EphA2 for the immunotherapy of paediatric bone tumours. Cancer Gene Ther. 2021;28(3–4):321–34.
Campbell CL, et al. Increased expression of the interleukin-11 receptor and evidence of STAT3 activation in prostate carcinoma. Am J Pathol. 2001;158(1):25–32.
Schwertschlag US, et al. Hematopoietic, immunomodulatory and epithelial effects of interleukin-11. Leukemia. 1999;13(9):1307–15.
Lewis VO, et al. The interleukin-11 receptor alpha as a candidate ligand-directed target in osteosarcoma: consistent data from cell lines, orthotopic models, and human tumor samples. Cancer Res. 2009;69(5):1995–9.
Huang G, et al. Genetically modified T cells targeting interleukin-11 receptor alpha-chain kill human osteosarcoma cells and induce the regression of established osteosarcoma lung metastases. Cancer Res. 2012;72(1):271–81.
Pollak M. The insulin and insulin-like growth factor receptor family in neoplasia: an update. Nat Rev Cancer. 2012;12(3):159–69.
Cui B, et al. Targeting ROR1 inhibits epithelial-mesenchymal transition and metastasis. Cancer Res. 2013;73(12):3649–60.
Hudecek M, et al. The B-cell tumor-associated antigen ROR1 can be targeted with T cells modified to express a ROR1-specific chimeric antigen receptor. Blood. 2010;116(22):4532–41.
Huang X, et al. IGF1R- and ROR1-Specific CAR T cells as a potential therapy for high risk sarcomas. PLoS ONE. 2015;10(7):e0133152.
Fernandez L, et al. Activated and expanded natural killer cells target osteosarcoma tumor initiating cells in an NKG2D-NKG2DL dependent manner. Cancer Lett. 2015;368(1):54–63.
Spear P, et al. NKG2D ligands as therapeutic targets. Cancer Immun. 2013;13:8.
Fernandez L, et al. Memory T cells expressing an NKG2D-CAR efficiently Target Osteosarcoma cells. Clin Cancer Res. 2017;23(19):5824–35.
Amato RJ, et al. A phase I study of folate immune therapy (EC90 vaccine administered with GPI-0100 adjuvant followed by EC17) in patients with renal cell carcinoma. J Immunother. 2013;36(4):268–75.
Lu YJ, et al. Preclinical evaluation of bispecific adaptor molecule controlled folate receptor CAR-T cell therapy with Special Focus on Pediatric malignancies. Front Oncol. 2019;9:151.
Sanchez-Paulete AR, et al. Targeting macrophages with CAR T cells delays solid tumor progression and enhances Antitumor Immunity. Cancer Immunol Res. 2022;10(11):1354–69.
Klichinsky M, et al. Human chimeric antigen receptor macrophages for cancer immunotherapy. Nat Biotechnol. 2020;38(8):947–53.
Gajewski TF, Schreiber H, Fu YX. Innate and adaptive immune cells in the tumor microenvironment. Nat Immunol. 2013;14(10):1014–22.
Koksal H, et al. Treating osteosarcoma with CAR T cells. Scand J Immunol. 2019;89(3):e12741.
Moreno C, Alternative CAR, Therapies et al. Recent approaches in Engineering chimeric Antigen receptor Immune cells to Combat Cancer. Biomedicines, 2022. 10(7).
Brady RV, Thamm DH. Tumor-associated macrophages: prognostic and therapeutic targets for cancer in humans and dogs. Front Immunol. 2023;14:1176807.
Chen S, et al. Harnessing and enhancing macrophage phagocytosis for Cancer Therapy. Front Immunol. 2021;12:635173.
de Visser KE, Joyce JA. The evolving tumor microenvironment: from cancer initiation to metastatic outgrowth. Cancer Cell. 2023;41(3):374–403.
Quamine AE et al. Approaches to Enhance Natural Killer Cell-based immunotherapy for Pediatric Solid tumors. Cancers (Basel), 2021. 13(11).
Wu Y, Tian Z, Wei H. Developmental and Functional Control of Natural Killer cells by cytokines. Front Immunol. 2017;8:930.
Liu E, et al. Use of CAR-Transduced Natural Killer cells in CD19-Positive lymphoid tumors. N Engl J Med. 2020;382(6):545–53.
Zhang Y, et al. In vivo kinetics of human natural killer cells: the effects of ageing and acute and chronic viral infection. Immunology. 2007;121(2):258–65.
Ebb D, et al. Phase II trial of trastuzumab in combination with cytotoxic chemotherapy for treatment of metastatic osteosarcoma with human epidermal growth factor receptor 2 overexpression: a report from the children’s oncology group. J Clin Oncol. 2012;30(20):2545–51.
Yu AL, et al. Phase I trial of a human-mouse chimeric anti-disialoganglioside monoclonal antibody ch14.18 in patients with refractory neuroblastoma and osteosarcoma. J Clin Oncol. 1998;16(6):2169–80.
Park JA, Cheung NV. GD2 or HER2 targeting T cell engaging bispecific antibodies to treat osteosarcoma. J Hematol Oncol. 2020;13(1):172.
Mason NJ, et al. Immunotherapy with a HER2-Targeting Listeria induces HER2-Specific immunity and demonstrates potential therapeutic effects in a phase I Trial in Canine Osteosarcoma. Clin Cancer Res. 2016;22(17):4380–90.
Kawano M, et al. Anti-TGF-beta antibody combined with dendritic cells produce antitumor effects in osteosarcoma. Clin Orthop Relat Res. 2012;470(8):2288–94.
Kawano M, et al. Dendritic cells combined with anti-GITR antibody produce antitumor effects in osteosarcoma. Oncol Rep. 2015;34(4):1995–2001.
Zhou Y et al. Vaccine efficacy against primary and metastatic cancer with in vitro-generated CD103(+) conventional dendritic cells. J Immunother Cancer, 2020. 8(1).
Lussier DM, et al. Combination immunotherapy with alpha-CTLA-4 and alpha-PD-L1 antibody blockade prevents immune escape and leads to complete control of metastatic osteosarcoma. J Immunother Cancer. 2015;3:21.
Nuytemans L, et al. NGS-analysis to the rescue: dual checkpoint inhibition in metastatic osteosarcoma - a case report and review of the literature. Acta Clin Belg. 2021;76(2):162–7.
D’Angelo SP, et al. Nivolumab with or without ipilimumab treatment for metastatic sarcoma (Alliance A091401): two open-label, non-comparative, randomised, phase 2 trials. Lancet Oncol. 2018;19(3):416–26.
Somaiah N, et al. Durvalumab plus Tremelimumab in advanced or metastatic soft tissue and bone sarcomas: a single-centre phase 2 trial. Lancet Oncol. 2022;23(9):1156–66.
Motz GT, Coukos G. The parallel lives of angiogenesis and immunosuppression: cancer and other tales. Nat Rev Immunol. 2011;11(10):702–11.
Martin-Broto J et al. Nivolumab and sunitinib combination in advanced soft tissue sarcomas: a multicenter, single-arm, phase Ib/II trial. J Immunother Cancer, 2020. 8(2).
Zhou Y, et al. A pilot study of multi-antigen stimulated cell therapy-I plus camrelizumab and apatinib in patients with advanced bone and soft-tissue sarcomas. BMC Med. 2023;21(1):470.
Xie L et al. Apatinib plus camrelizumab (anti-PD1 therapy, SHR-1210) for advanced osteosarcoma (APFAO) progressing after chemotherapy: a single-arm, open-label, phase 2 trial. J Immunother Cancer, 2020. 8(1).
Park JA et al. Targeting tumor vasculature to improve antitumor activity of T cells armed ex vivo with T cell engaging bispecific antibody. J Immunother Cancer, 2023. 11(3).
Kawano M, et al. Enhancement of antitumor immunity by combining anti-cytotoxic T lymphocyte antigen-4 antibodies and cryotreated tumor lysate-pulsed dendritic cells in murine osteosarcoma. Oncol Rep. 2013;29(3):1001–6.
Xie L, et al. Exploratory study of an anti-PD-L1/TGF-beta antibody, TQB2858, in patients with refractory or recurrent osteosarcoma and alveolar soft part sarcoma: a report from Chinese sarcoma study group (TQB2858-Ib-02). BMC Cancer. 2023;23(1):868.
Diab A, et al. Bempegaldesleukin (NKTR-214) plus Nivolumab in patients with Advanced Solid tumors: phase I dose-escalation study of Safety, Efficacy, and Immune activation (PIVOT-02). Cancer Discov. 2020;10(8):1158–73.
D’Angelo SP, et al. Pilot study of bempegaldesleukin in combination with nivolumab in patients with metastatic sarcoma. Nat Commun. 2022;13(1):3477.
Tanaka M, et al. Vaccination targeting native receptors to enhance the function and proliferation of Chimeric Antigen Receptor (CAR)-Modified T cells. Clin Cancer Res. 2017;23(14):3499–509.
Acknowledgements
Not applicable.
Funding
This review was supported by National Natural Science Foundation of China (Grant No. 82203007) and the China Postdoctoral Science Foundation (No. 2023MD744162).
Author information
Authors and Affiliations
Contributions
SY searched the literatures and wrote the manuscript. XY designed the study and participated in the discussion. All authors reviewed the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
All authors have read and approved the final manuscript for publication.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Yu, S., Yao, X. Advances on immunotherapy for osteosarcoma. Mol Cancer 23, 192 (2024). https://doi.org/10.1186/s12943-024-02105-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12943-024-02105-9