
Abstract
Background
Rheumatoid arthritis (RA) is associated with a high concentration of extracellular DNA (ecDNA). This could be a consequence of the inflammation, but the ecDNA could also be involved in the unknown etiopathogenesis of RA. Clearance of ecDNA is hypothesized to prevent the development of RA. This study aimed to analyze the effects of exogenous deoxyribonuclease I (DNase I) administration in an animal model of RA.
Methods
The collagen antibody-induced arthritis (CAIA) model of RA was induced in adult female DBA/1J mice. CAIA mice were treated with saline or DNase I (10 mg/kg) every 12 h for the whole duration of the experiment. Arthritic scores were assessed. Paw volume and temperature were assessed using a plethysmometer and a thermal camera, respectively. Plasma ecDNA and its subcellular origin were analyzed using fluorometry and real-time PCR. DNase activity was quantified with single radial enzyme diffusion method.
Results
The CAIA model was successfully induced as proved by a higher volume, temperature and the overall arthritis score in comparison to controls. The administration of DNase I resulted in a nearly two-fold increase in serum DNase activity. Still, it did affect neither plasma ecDNA, nor the arthritis score or other measures of joint inflammation.
Conclusion
Our results suggest that exogenous DNase I does not prevent the development of CAIA in mice. Whether this is true for other animal models of arthritis or clinical RA requires further research. EcDNA does not seem to be involved in the pathogenesis of CAIA. Additional studies are also needed to elucidate the role of ecDNA in the development of RA, focusing especially on its origin and inhibition of ecDNA release.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Background
Rheumatoid arthritis (RA) is a chronic, autoimmune, inflammatory disease affecting around 1% of the population and its etiopathogenesis is still unknown [1]. The lack of knowledge about the real causes of RA leads to the absence of causal treatment [2]. The currently used biological disease-modifying anti-rheumatic drugs are the most effective treatment of RA in history. Still, side effects and a group of non-responders among patients require a further search for causal interventions [3, 4].
Similarly, to other autoimmune diseases, a higher concentration of circulating extracellular DNA (ecDNA) has been reported in patients with RA [5]. The release of DNA from cells can result from apoptosis, pyroptosis, autophagy, or the formation of neutrophil extracellular traps (NETs) [6]. NETs contribute to the increased ecDNA in RA, but other NETs such as citrullinated histones, myeloperoxidase, neutrophil elastase and antimicrobial peptides may play a role in the inflammatory responses typical for RA [7,8,9]. The fact that ecDNA release is driven by multiple mechanisms makes therapy targeting difficult. Potential protection of ecDNA from nucleases by extracellular vesicles further adds to the complexity. This could pose a significant challenge in developing treatments aiming to degrade ecDNA [10, 11]. Regarding subcellular sources, ecDNA can originate from the nucleus (nuclear DNA - ncDNA) or from the mitochondria (mitochondrial DNA - mtDNA) which has a higher potential to activate immune cells. EcDNA can be recognized as a damage-associated molecular pattern that induces an inflammatory response [12]. EcDNA is also present in healthy individuals, but in low concentration due to rapid removal by nucleases - deoxyribonucleases (DNases) [13]. The results of our previous study have shown that in RA patients, ecDNA concentrations positively correlate with the clinical parameters of disease severity, such as disease activity score (DAS28) and C-reactive protein [14, 15]. On the other hand, elevated ecDNA in RA might be more than just another biomarker of ongoing inflammation. It could be involved in the etiopathogenesis of RA, representing a potential causal therapeutic target [16].
As ecDNA is cleaved by nucleases, high ecDNA concentrations could be explained not only by increased release of DNA from cells but alternatively also by an insufficient clearance by DNases caused by their impairment or by the presence of their inhibitors as increased DNase I activity has been observed as a concomitant process in conditions where there is increased cell death [17,18,19,20]. Clinically, DNase is safely and efficiently used to treat cystic fibrosis. DNase reduces the viscosity of the mucus by digesting ecDNA in the alveoli [21, 22]. We and others have previously shown that exogenous DNase has beneficial effects in animal models of sepsis [23], hepatorenal injury [24], thrombosis [25] or ischemic-reperfusion injury [26]. Little is known about the effects of exogenous DNase in RA or animal models of RA. A recent study has shown promising results for the local intraarticular application of DNase [27]. However, RA is a systemic inflammatory disease and requires a systemic approach. Therefore, the present study aimed to investigate the effects of exogenous intraperitoneally administered DNase I in the mouse model of collagen antibody-induced arthritis (CAIA). In the current study DBA/IJ female mice were used to reflect the higher prevalence of RA in women compared to men [1]. We hypothesized that the clearance of ecDNA will prevent or reduce the severity of the induced arthritis.
Methods
Animals and housing conditions
In this study, female DBA/1J mice (12 months old) were used (Velaz, Prague, Czech Republic). Animals were group-housed (2–3 per cage) in polycarbonate cages (36.5 × 20.5 × 14 cm) and kept under standard laboratory conditions (temperature 22 ± 2 °C, humidity 55 ± 10%, 12:12 light-dark cycle). Animals had ad libitum access to food (standard diet for mice Eypy-KMK20, Bruzovice, Czech Republic) and water.
Groups of animals and experimental design
Mice were divided into the following groups: control group – CTRL (n = 5) was injected with a saline (B. Braun Melsungen AG, Germany), CAIA group (n = 5) was injected with the mouse anti-type II collagen 5-clone monoclonal antibody cocktail (1.5 mg/mouse, Chondrex, Woodinville, WA, USA), recognizing the conserved epitopes of type II collagen. The last group of mice was the CAIA + DNase I group (n = 5). This group was injected with a monoclonal antibody cocktail as the CAIA group and DNase I (10 mg/kg, Sigma-Aldrich, Taufkirchen, Germany). Lyophilized DNase I was dissolved in 0.9% saline.
On day 0, CAIA females were injected i.p. with an anti-type II collagen 5-clone monoclonal antibody cocktail (1.5 mg/mouse, Chondrex, Woodinville, USA) followed by an i.p. lipopolysaccharide (25 µg/mouse, Chondrex, Woodinville, USA) injection on day 3 of the experiment. To assess the therapeutic potential of DNase I, the CAIA + DNase I group of females received an i.p. injection of bovine DNase I (10 mg/kg, Sigma-Aldrich, Taufkirchen, Germany) every 12 h from the beginning till the end of the experiment on day 8.
Clinical assessment of the severity of arthritis
Mice were monitored every second day during the whole experiment. The severity of arthritis was scaled from 0 to 4 points for every paw separately. Scoring was performed based on the mouse arthritis scoring system where three types of joints were observed - interphalangeal, metacarpophalangeal, carpal and tarsal joints. The scoring system was as follows: score 0 - normal/physiologic state, score 1 - one of the above-mentioned joints has redness and swelling, score 2 - two joints have redness and swelling, score 3 - all three joint types have redness and swelling, score 4 - achievement of maximal redness and swelling of the entire paw [28].
Plethysmometry
A plethysmometer (Ugo Basile, Gemonio, Italy) was used to evaluate paw swelling. The plethysmometer comprised two interconnected, clear acrylic tubes filled with a solution. Acrylic tubes were connected to a decoder that digitally presented the volume displaced after the paws of mice were placed into the prepared conductive solution. The paw volume was calculated as an average from three measurements for every paw [29].
Assessment of body temperature and temperature of paws
To assess the body temperature and temperature of all four paws a factory-calibrated handheld thermal imaging camera was used (Teledyne FLIR-E64501, Wilsonville, OR, USA). The body temperature correlates with the ocular surface temperature of mice, therefore, in the present study, the body temperature of mice was measured from the ocular surface [30]. During all temperature measurements, the room temperature was 24 ± 1 °C (humidity: 55 ± 10%), and the mice were handled by the same experimenter on a stable platform. The distance between the camera and the mouse was 20 cm. Three thermal images were recorded for each mouse at the same time of the day. The thermal images were analyzed using FLIR Tools software (FLIR Teledyne FLIR-E64501, Wilsonville, OR, USA). The ocular surface temperature was determined as an average of the maximum temperature of the left and right ocular surfaces. The final body temperature was identified as an average from 3 facial thermal images. To measure the temperature of paws, each paw was marked with an ellipse shape using marking tools within the software. The final temperature of the paws was calculated as an average of three thermal images of the paws [31].
Measurement of paw swelling using micro-computed tomography
After collection of front paws, paws were snap-frozen in liquid nitrogen and immediately stored in a -80 °C freezer. For more accurate verification of the extent of the swelling, micro-computed tomography (micro-CT) was used (IVIS® SpectrumCT, Caliper Life Sciences, Hopkinton, USA). During scanning, the paws were placed in the scanner with a long axis aligned with the axis of the scanner bed. Obtained scans were analyzed in RadiAnt DICOM Viewer (Medixant, Poznaň, Poland). The swelling of tissue was quantified as total paw thickness and also paw thickness inferior to metatarsal bones. The total paw thickness, in mm, was calculated as the distance across the whole paw including the metatarsal bones, the inferior thickness of the paw was measured from the metatarsal bones to the end of the soft tissue (Fig. 2G).

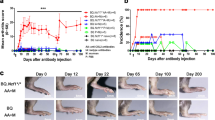
Development of arthritis in mice. A timeline of the present study. B dynamics of arthritis score representing arthritis incidence and severity. C representative images of the severity of arthritis. D dynamics of body temperature during the experiment. E, F incidence of the severity of arthritis in the form of the swelling of front and back paws
Bioluminescence
For noninvasive detection of inflammation, the bioluminescence of luminol is modified by the reaction with myeloperoxidase products. The luminol solution was prepared by dissolving powdered luminol sodium salt (Carbosynth, Berkshire, United Kingdom) in phosphate-buffered saline at a concentration of 50 mg/ml. To monitor the inflammation, the luminol solution (200 mg/kg body weight) was applied i.p. to mice [15]. Mice were anesthetized using isoflurane (Vetpharma Animal Health, S.L, Barcelona, Spain) inhalation (3% isoflurane, 97% oxygen) in the sleeping chamber during the measurements. Ten minutes after the application of the luminol solution, the mouse was placed into the chamber of the system IVIS® SpectrumCT (PerkinElmer, Waltham, MA, USA). The exposure time was set to 5 min. After the measurements, the bioluminescence signal from paws was quantified in the software LivingImage (PerkinElmer, Waltham, MA, USA) using average radiance (photons/s/cm2/sr) [32].
Histology scoring
Paws were fixed in neutral buffered 4% formalin for 6 h before the decalcification. Bones were decalcified for 1 week in 14% EDTA, pH = 7.2 at 4 °C. Sagittal sections were stained with hematoxylin and eosin and scored by a blinded observer [33]. The scoring system was as follows: inflammatory cells infiltrate 0 - no infiltrate detected, 1 - modest leukocyte infiltration in synovial tissue, no fluid leukocytes, 2 - moderate leukocyte infiltration in synovial tissue and fluid phase, with loss of synovial architecture, 3 - gross leukocyte infiltration in synovial membrane and fluid space, significant loss of synovial articular architecture. Synovial hyperplasia: 0 - no abnormalities detected, 1- synovial lining layer 2–4 cells thick, 2 - synovial lining layer 5 and more cells thick associated with a moderate expansion of the sub-lining layer, 3 - synovial lining layer 5 and more cells thick associated with significant expansion of the sub-lining layer zone and potentially with loss of synovial architecture. Erosion of cartilage and bone: 0 - no abnormalities, 1 - fibrillation of cartilage and/or mild erosive infiltration of periosteal and subchondral bone, nuclei intact within lacunae, 2 - moderate fibrillation and loss of cartilage and/or moderate erosive infiltration of periosteal and subchondral bone, 3 – a significant loss of cartilage and/or erosive infiltration of periosteal and subchondral bone, nuclei show apoptosis within lacunae across a wide area of cartilage/bone.
DNA isolation and quantification
To quantify the concentration of ecDNA blood samples were collected on the day of the sacrifice into tubes coated with lithium heparin (Microvette 500 LH, Sarstedt, Germany). To separate plasma, samples were centrifuged at 1600 × g for 10 min at 4 °C. Plasma was centrifuged again at 16000 × g for 10 min at 4 °C [34]. The supernatant was used for ecDNA isolation using QIAamp DNA Blood Mini Kit (Qiagen, Hilden, Germany). Quantification of isolated DNA was made by the Qubit 3.0 fluorometer and Qubit dsDNA high-sensitivity assay (Thermo Fisher Scientific, Waltham, MA, USA). The subcellular origin of ecDNA was determined using real-time PCR on the Mastercycler Realplex 4 (Eppendorf, Hamburg, Germany). The master mix used for the PCR reaction was SYBR Green Supermix (Bio-Rad Laboratories, Hercules, CA, USA). The PCR program was following: one cycle of 98 °C for 3 min, 40 cycles of 98 °C for 15 s for denaturation, 60 °C for 30 s for annealing, and 72 °C for 30 s for an extension. The sequence of primers to quantify the ncDNA (F:5′-TGTCAGATATGTCCTTCAGCAAGG-3′, R:5′-TGCTTAACTCTGCAGGCGTATG-3′), while for mtDNA we used primers with the sequence F:5′-CCCAGCTACTACCATCATTCAAGT-′, R:5′-GATGGTTTGGGAGATTGGTTGATGT-3′).
Measurement of serum DNase activity
For the determination of DNase activity in serum a single radial enzyme diffusion (SRED) method was used. DNA agarose gel (1%, 20 mM Tris–HCl, pH 7.5, 2 mM MgCl2, 2 mM CaCl2) with DNA isolated from chicken livers (0.035 mg/ml per gel) was prepared. For the visualization GoodView Nucleic Acid Stain (SBS Genetech, Beijing, China) and for the calibration values the DNase set with RDD buffer (Qiagen, Hilden, Germany) were used. 1 µl of the DNase stock and the samples were used. Prepared samples and standards (1 µl) were pipetted into the gel. Following an overnight incubation (at 37 °C in the dark) the gel was visualized using iBOX (Vision works LP Analysis Software, UVP, Upland, CA, USA). Radial enzyme diffusion was then calculated from the gel using the ImageJ software (NIH, Bethesda, MD, USA). The DNase activity was expressed in Kunitz units (K.u.) per ml of serum [35].
Statistical analysis
Statistical analysis was performed using GraphPad Prism 8.1 software (La Jolla, San Diego, CA, USA). Based on the Shapiro-Wilk test for normality of distribution, parametric One-way and Two-way Analysis of Variance (ANOVA) or Friedman’s test was used for respective parameters with a Bonferroni-corrected post-hoc t-test. P values lower than 0.05 were considered statistically significant. Data are presented as mean ± standard deviation (SD).
Results
RA score
Arthritis score based on inflammation signs constantly increased in the groups receiving monoclonal antibodies leading to a difference between the CAIA groups and the CTRL group of mice. A significant difference in the score was detected on day 6 of the experiment 8.33 ± 5.65 vs. 0 for CAIA and CTRL, respectively (t(5) = 3.616, p = 0.046 (Fig. 1B). Similar scores were observed on day 8 (t(5) = 5.918, p = 0.006). No significant differences in arthritis severity were found between the CAIA and CAIA + DNase I groups during the whole experiment.
Body temperature and paw inflammation
During the experiment, no significant differences were observed in body temperature between the studied groups (F = 0.1432, p = 0.8345) (Fig. 1D). On day 6 of the experiment, mice of the CAIA and CAIA + DNase I groups showed higher temperature of the front paws than the CTRL group (by 1.5 °C, CAIA: t(7.69) = 5.50, p = 0.002, CAIA + DNase I: t(8.26) = 5.26) (Fig. 2B). On day 8, only CAIA + DNase I females displayed significantly higher temperature of front paws by 1 °C compared to CTRL females (t(6.69) = 5.01, p = 0.005) (Fig. 2A and B). Regarding the back paws, CAIA mice vs. CTRL mice showed higher paw temperature by 1 °C, (t(9.24) = 3.07, p = 0.039) only on day 8. CAIA + DNase I females had warmer back paws compared to CTRL group on day 6 and day 8 of the experiment by 1 °C (day 6: t(9.93) = 3.32, p = 0.023, day 8: t(8.93) = 3.61 p = 0.017) (Fig. 2A and Fig. B).

Representation of arthritis signs at the end of the experiment. A representative image of the thermal camera pictures. B dynamics of the paw temperature. C representative images of the quantification of the inflammation-based bioluminescence. D dynamics of the paw bioluminescence. E representative images of the hematoxylin-eosin staining. F synovial hyperplasia and cartilage bone erosion. G representative pictures of the uCT, used for the assessment of tissue swelling. H representation of the swelling (total paw thickness and dorsal paw thickness)
Plethysmometer measurements
To verify peripheral inflammation and quantify the swelling of paws (paw volume), plethysmometer measurements were conducted (Fig. 1E). On day 6 of the experiment, front paws in CAIA + DNase I showed swelling by 18% higher than the CTRL group (t(6.84) = 3.28, p = 0.042) (Fig. 1E). On back paws we could observe the interactive effect of time (F(2.07, 29.5) = 6.18, p = 0.005) and treatment (F(2, 15) = 4.12, p = 0.037), nevertheless no significant differences were observed in the partial analysis of swelling between the groups (Fig. 1F).
Bioluminescence and micro-computed tomography
The extent of inflammation was measured by luminol-based bioluminescence imaging. Results showed more than 9 times higher signal of inflammation in CAIA mice in comparison to the CTRL group (t(12) = 3.08, p = 0.029). In addition, CAIA + DNase I females showed 1.5 times lower extent of the inflammation than CAIA mice however, these differences were not significant (t(12) = 0.82, p > 0.999). Regarding the back paws, the luminol-based bioluminescence imaging showed no significant differences in the extent of the inflammation between the CAIA and CTRL females (t(12) = 2.17, p = 0.152). The back paws of CAIA + DNase I showed a 5.25 higher bioluminescence signal than the CTRL group (t(12) = 3.05, p = 0.03) (Fig. 2C and D).
The development of edema in the CAIA model was also monitored using the micro-CT (Fig. 2G and H). Total paw thickness (paw tissue with bones) was significantly higher by 25% on the front paws in CAIA + DNase I in comparison to CTRL mice (t(14) = 3.75, p = 0.007,). In the inferior part of the front paws, both, CAIA and CAIA + DNase I showed more than 2 times increased tissue volume that the healthy paws in CTRL group (CTRL vs. CAIA: t(14) = 3.14, p = 0.022, CTRL vs. CAIA + DNase I: t(14) = 3.29, p = 0.016) (Fig. 2G and H).
ecDNA and DNase activity
Plasma concentrations of ecDNA were not affected by the application of monoclonal antibodies against collagen type II (F = 0.54, p = 0.59) (Fig. 3A). Similarly, no differences were observed in the subcellular origin of ecDNA (ncDNA: F = 3.08, p = 0.083, mtDNA: F = 0.46, p = 0.640) (Fig. 3C and D). Application of the exogenous DNase I led to higher DNase activity by 1.5 times between CAIA + DNase I and CAIA group, t(13) = 4.19, p = 0.003, (Fig. 3D). There were no significant differences in DNase activity between CAIA and CTRL mice (t(13) = 0.57, p > 0.999) (Fig. 3B).

Extracellular DNA (ecDNA) in plasma with its subcellular origin and deoxyribonuclease (DNase) activity. A concentration of total ecDNA in plasma. B DNase activity in serum after the application of the exogenous DNase. C plasma concentration of nuclear DNA (ncDNA), D plasma concentration of mitochondrial DNA (mtDNA)
Discussion
In this study we successfully induced the CAIA model in female mice. We tested the application of exogenous DNase as a possible intervention to prevent the development of arthritis. The administration of monoclonal antibodies led to inflammation - redness and swelling of the paws. Exogenous DNase, however, did not prevent the development of arthritis. No differences were seen in the arthritis score between the treated group and the mice with CAIA suggesting that ecDNA is not involved in the pathogenesis of this arthritis model.
In clinical studies, RA patients have higher plasma ecDNA in comparison to healthy individuals [5, 16]. EcDNA can be removed by endogenous and exogenous DNase [5, 16, 36]. The known pro-inflammatory properties of ecDNA support the rationale for inducing clearance of ecDNA that could mitigate inflammation [16, 37]. In our previous study, DNase administration improved survival in an animal model of sepsis [37]. In the present study, we were not able to reproduce the beneficial effect of DNase seen in another inflammatory model. A possible explanation could be the different route of application. In the model of sepsis, DNase was applied into the tail vein, in the present study DNase was applied intraperitoneally. However, the assessment of serum DNase activity confirms that the treated animals had higher DNase activity. So, the difference might rather be related to the disease models and their pathogenesis.
There are only few published studies on DNase in RA [27, 38, 39]. In the study by Zervou et al. polymorphisms in the DNASEIL3 gene were studied in association with autoimmune disorders such as RA [38]. In the animal experiment by Kawane et al. mice with a deletion in Dnase2 gene suffered from chronic polyarthritis suggesting a similar cause in human patients with various types of arthritis [39]. Wang et al. in their study applied a hydrogel containing DNase to disrupt neutrophils extracellular traps to treat induced arthritis locally [27]. Only the latter one is an interventional experimental study. DNase fused with a hydrogel to increase its stability was applied locally to reduce arthritis in a mouse model [27]. This is an important difference to our study with systemic application of DNase that is not stabilized and so, has to be applied repeatedly. Local administration might have a better local outcome, but RA is a systemic disease affecting many joints and with severe extraarticular manifestations. So, we believe that a systemic approach to treatment is necessary also in the CAIA model, which despite all the differences in the pathogenesis is still used as an animal model of RA. Wang et al. also used collagen-induced arthritis - a slower, chronic model. In our study, we used CAIA with acute inflammation and fast progress. It is, thus, clear that the next step should be the testing of exogenous systemic DNase in the collagen-induced arthritis as well.
A reason for the lack of effect of DNase in CAIA could be the lack of effect of CAIA on ecDNA. In contrast to RA in human patients, CAIA induction in mice did not increase ecDNA suggesting a difference in the pathogenesis and pointing towards a major limitation of this model for the research of RA. Despite clear signs of inflammation in mice, CAIA did not affect ecDNA. The reasons could include the multiple, but local inflammation in CAIA vs. systemic inflammation in RA, but also the different duration of the inflammation in mice vs. human patients [1, 30]. In our study, the discrepancies regarding ecDNA concentrations might originate from the used experimental model. RA has a complex pathogenesis, making its modeling difficult. Consequently, various animal models of RA have been developed, each focusing on a specific aspect of the pathogenesis. No single animal model currently mimics the entire RA pathogenesis. Thus, ecDNA concentrations might vary between different models and the human disease [40]. We have studied the effect of DNase I treatment in collagen induced arthritis as this model was already established in our laboratory [41]. Unfortunately, we are not able to collect enough synovial fluid from mice to confirm higher local ecDNA and the potential effect of DNase on this ecDNA.
According to our knowledge, this is the first study describing the effect of intraperitoneal DNase administration in the CAIA model. Based on our results this new potential preventive treatment failed to affect the development of arthritis. Also, this is the first study to analyze ecDNA and its subcellular origin in the CAIA model. Again, our results suggest that ecDNA is not affected by CAIA and is not involved in its pathogenesis. However, due to differences between human disease and its animal models, this does not mean that DNase cannot be useful for the prevention or treatment of RA. To prove this hypothesis other models and potential clinical trials are needed. The source of ecDNA in inflammation are mostly neutrophil extracellular traps and these are elevated in patients with RA [36]. Neutrophils have an important role in the progression of RA, but maybe also in its initiation. NETs are web-like structures with a DNA scaffold coated with proteins which could act as auto-antigens [42]. RA is a chronic inflammatory disease primary affecting joints. Therefore, the research literature is focusing on the site of the inflammation and the neutrophils in the synovial fluid are investigated [43,44,45,46]. It has been reported that neutrophils in the synovial fluid show altered phenotype in RA resulting in higher NETs formation [43]. Furthermore, the presence of NETs in the synovial fluid was associated with osteaoclastogenesis leading to bone loss and articular damage [47, 48]. In addition, NETs drive T cell differentiation via activation of Toll-like receptors by NETs-associated histones and nucleosomes [49]. It is of importance that NETs contain citrullinated proteins stimulating the formation of anti-citrullinated antibodies [32]. Therefore, citrullinated proteins represents a strong association between the NETs and RA with a bridge to local inflammation primarily affecting the musculoskeletal apparatus. However, why does the phenotype of neutrophils in the synovial fluid change and what could be the reasons for the increased formation of NETs is still unclear [43].
Studies investigating the pathogenesis of RA suggest that NETs production could lead to higher concentration of ecDNA, which further triggers the inflammatory process [14, 15, 50]. The release of ecDNA inside vesicles or at least associated with vesicles such as exosomes represents a challenge for the removal of ecDNA [51]. Histones, cathelicidin and other antimicrobial peptides likely protect ecDNA derived from NETs from degradation [52]. Thus, components of NETs could cause the low efficacy of DNase I application in the present study. NETs as the major source of ecDNA in inflammation influence the accessibility of DNA to DNase I [53]. Efficacy of DNase I could be also be altered by its inhibitors, for example anti-DNase antibodies. In addition, the presence of anti-NETs antibodies could contribute to lower dissembling of NETs as it was reported in patients with systemic lupus erythematosus [54, 55]. The application of DNase I has already been shown to have beneficial effects in mouse models of sepsis and hepatorenal failure [24, 37]. However, differences in the structure of ecDNA in various diseases and disease models could cause the variability of DNase effects. Whether extracellular traps with ecDNA as their main compound are also higher in animal models of RA is not clear. Despite negative outcomes, our study might help to focus further research on RA linked to early biomarkers or more importantly to uncovering the unknown etiopathogenesis of RA. Despite the currently available biological therapy, there is still a large group of RA patients who would profit from a causal targeted treatment.
Conclusion
Application of the DNase I did not cause major and clinically relevant changes between the treated and the untreated groups of CAIA mice. Results from this study suggest that ecDNA is not involved in the pathogenesis of rheumatoid arthritis, however, this could be influenced by the selected animal model, which represents only, very acute, and local inflammation. EcDNA thus can be increased in the site of inflammation but within the laboratory rodents, we are limited with the extraction of the fluids. Therefore, new analysis with different animal models of rheumatoid arthritis should be performed to prove if the ecDNA stands behind rheumatoid arthritis.
Data availability
The datasets used and/or analysed in the current study are available from the corresponding author upon reasonable request.
Abbreviations
- CAIA:
-
Collagen antibody-induced arthritis
- DAS28:
-
Disease activity score
- DNase I:
-
Deoxyribonuclease I
- DNases:
-
Deoxyribonucleases
- ecDNA:
-
Extracellular DNA
- micro-CT:
-
Micro-computed tomography
- mtDNA:
-
Mitochondrial DNA
- ncDNA:
-
Nuclear DNA
- RA:
-
Rheumatoid arthritis
References
Smolen JS, Aletaha D, Barton A, Burmester GR, Emery P, Firestein GS, et al. Rheumatoid arthritis. Nat Rev Dis Primers. 2018;4:18001.
Burmester GR, Pope JE. Novel treatment strategies in rheumatoid arthritis. Lancet. 2017;389(10086):2338–48.
Smolen JS, Emery P, Fleischmann R, van Vollenhoven RF, Pavelka K, Durez P, et al. Adjustment of therapy in rheumatoid arthritis on the basis of achievement of stable low disease activity with adalimumab plus methotrexate or methotrexate alone: the randomised controlled OPTIMA trial. Lancet. 2014;383(9914):321–32.
Kavanaugh A, van Vollenhoven RF, Fleischmann R, Emery P, Sainsbury I, Florentinus S, et al. Testing treat-to-target outcomes with initial methotrexate monotherapy compared with initial tumour necrosis factor inhibitor (adalimumab) plus methotrexate in early rheumatoid arthritis. Ann Rheum Dis. 2018;77(2):289–92.
Rykova E, Sizikov A, Roggenbuck D, Antonenko O, Bryzgalov L, Morozkin E, et al. Circulating DNA in rheumatoid arthritis: pathological changes and association with clinically used serological markers. Arthritis Res Ther. 2017;19(1):85.
Galluzzi L, Vitale I, Aaronson SA, Abrams JM, Adam D, Agostinis P, et al. Molecular mechanisms of cell death: recommendations of the nomenclature Committee on Cell Death 2018. Cell Death Differ. 2018;25(3):486–541.
Brinkmann V, Reichard U, Goosmann C, Fauler B, Uhlemann Y, Weiss DS, et al. Neutrophil extracellular traps kill bacteria. Science. 2004;303(5663):1532–5.
Lee KH, Kronbichler A, Park DD-Y, Park Y, Moon H, Kim H, et al. Neutrophil extracellular traps (NETs) in autoimmune diseases: a comprehensive review. Autoimmun rev. 2017;16(11):1160–73.
Sur Chowdhury C, Giaglis S, Walker UA, Buser A, Hahn S, Hasler P. Enhanced neutrophil extracellular trap generation in rheumatoid arthritis: analysis of underlying signal transduction pathways and potential diagnostic utility. Arthritis Res Therapy. 2014;16:1–14.
Lichá K, Pastorek M, Repiská G, Celec P, Konečná B. Investigation of the Presence of DNA in human blood plasma small extracellular vesicles. Int J Mol Sci. 2023;24(6).
Kustanovich A, Schwartz R, Peretz T, Grinshpun A. Life and death of circulating cell-free DNA. Cancer Biol Ther. 2019;20(8):1057–67.
Boyapati RK, Tamborska A, Dorward DA, Ho GT. Advances in the understanding of mitochondrial DNA as a pathogenic factor in inflammatory diseases. F1000Res. 2017;6:169.
Han DSC, Lo YMD. The Nexus of cfDNA and nuclease Biology. Trends Genet. 2021;37(8):758–70.
Macáková K, Illésová J, Mlynáriková V, Lesayová A, Konečná B, Vlková B, et al. The dynamics of extracellular DNA associates with treatment response in patients with rheumatoid arthritis. Sci Rep. 2022;12(1):21099.
Lauková L, Konečná B, Vlková B, Mlynáriková V, Celec P, Šteňová E. Anti-cytokine therapy and plasma DNA in patients with rheumatoid arthritis. Rheumatol Int. 2018;38:1449–54.
Hashimoto T, Yoshida K, Hashiramoto A, Matsui K. Cell-free DNA in rheumatoid arthritis. Int J Mol Sci. 2021;22(16).
Gaipl US, Beyer TD, Heyder P, Kuenkele S, Böttcher A, Voll RE, et al. Cooperation between C1q and DNase I in the clearance of necrotic cell-derived chromatin. Arthritis Rheum. 2004;50(2):640–9.
Mathapathi S, Chu C-Q. Contribution of impaired DNASE1L3 activity to anti-DNA autoantibody production in systemic lupus erythematosus. Rheumatol Immunol Res. 2022;3(1):17–22.
Sadeghi M, Dehnavi S, Jamialahmadi T, Johnston TP, Sahebkar A. Neutrophil extracellular trap: a key player in the pathogenesis of autoimmune diseases. Int Immunopharmacol. 2023;116:109843.
Korzeneva IB, Kostuyk SV, Ershova LS, Osipov AN, Zhuravleva VF, Pankratova GV, et al. Human circulating plasma DNA significantly decreases while lymphocyte DNA damage increases under chronic occupational exposure to low-dose gamma-neutron and tritium β-radiation. Mutat Research/Fundamental Mol Mech Mutagen. 2015;779:1–15.
Shak S, Capon DJ, Hellmiss R, Marsters SA, Baker CL. Recombinant human DNase I reduces the viscosity of cystic fibrosis sputum. Proc Natl Acad Sci. 1990;87(23):9188–92.
Lazarus RA, Wagener JS. Recombinant human deoxyribonuclease I. Pharmaceutical biotechnology: fundamentals and applications. 2019:471 – 88.
Kmeťová K, Čonka J, Janko J, Illés J, Uličná O, Celec P. Plasma DNA and deoxyribonuclease are associated with glucose metabolism in healthy mice. PLoS ONE. 2022;17(4):e0265099.
Vokálová L, Lauková L, Čonka J, Melišková V, Borbélyová V, Bábíčková J, et al. Deoxyribonuclease partially ameliorates thioacetamide-induced hepatorenal injury. Am J Physiol Gastrointest Liver Physiol. 2017;312(5):G457–63.
Kumar R, Sonkar VK, Swamy J, Ahmed A, Sharathkumar AA, Pierce GL, et al. DNase 1 protects from increased thrombin generation and venous thrombosis during Aging: cross-sectional study in mice and humans. J Am Heart Association. 2022;11(2):e021188.
Peer V, Abu Hamad R, Berman S, Efrati S. Renoprotective effects of DNAse-I Treatment in a rat model of Ischemia/Reperfusion-Induced Acute kidney Injury. Am J Nephrol. 2016;43(3):195–205.
Wang N, Ma J, Song W, Zhao C. An injectable hydrogel to disrupt neutrophil extracellular traps for treating rheumatoid arthritis. Drug Delivery. 2023;30(1):2173332.
Cantley MD, Haynes DR, Marino V, Bartold PM. Pre-existing periodontitis exacerbates experimental arthritis in a mouse model. J Clin Periodontol. 2011;38(6):532–41.
Vetal S, Bodhankar SL, Mohan V, Thakurdesai PA. Anti-inflammatory and anti-arthritic activity of type-A procyanidine polyphenols from bark of Cinnamomum zeylanicum in rats. Food Sci Hum Wellness. 2013;2(2):59–67.
Vogel B, Wagner H, Gmoser J, Wörner A, Löschberger A, Peters L, et al. Touch-free measurement of body temperature using close-up thermography of the ocular surface. MethodsX. 2016;3:407–16.
Nosrati Z, Bergamo M, Rodríguez-Rodríguez C, Saatchi K, Häfeli UO. Refinement and validation of infrared thermal imaging (IRT): a non-invasive technique to measure disease activity in a mouse model of rheumatoid arthritis. Arthritis Res Therapy. 2020;22:1–16.
Gutowski MB, Wilson L, Van Gelder RN, Pepple KL. In vivo bioluminescence imaging for longitudinal monitoring of inflammation in animal models of uveitis. Investig Ophthalmol Vis Sci. 2017;58(3):1521–8.
Hayer S, Vervoordeldonk MJ, Denis MC, Armaka M, Hoffmann M, Bäcklund J, et al. SMASH’recommendations for standardised microscopic arthritis scoring of histological sections from inflammatory arthritis animal models. Ann Rheum Dis. 2021;80(6):714–26.
Lo YD, Corbetta N, Chamberlain PF, Rai V, Sargent IL, Redman CW, et al. Presence of fetal DNA in maternal plasma and serum. Lancet. 1997;350(9076):485–7.
Janovičová Ľ, Čonka J, Lauková L, Celec P. Variability of endogenous deoxyribonuclease activity and its pathophysiological consequences. Mol Cell Probes. 2022:101844.
Duvvuri B, Lood C. Cell-free DNA as a Biomarker in Autoimmune Rheumatic diseases. Front Immunol. 2019;10:502.
Lauková L, Konečná B, Bábíčková J, Wagnerová A, Melišková V, Vlková B, et al. Exogenous deoxyribonuclease has a protective effect in a mouse model of sepsis. Biomed Pharmacother. 2017;93:8–16.
Zervou MI, Andreou A, Matalliotakis M, Spandidos DA, Goulielmos GN, Eliopoulos EE. Association of the DNASE1L3 rs35677470 polymorphism with systemic lupus erythematosus, rheumatoid arthritis and systemic sclerosis: structural biological insights. Mol Med Rep. 2020;22(6):4492–8.
Kawane K, Ohtani M, Miwa K, Kizawa T, Kanbara Y, Yoshioka Y, et al. Chronic polyarthritis caused by mammalian DNA that escapes from degradation in macrophages. Nature. 2006;443(7114):998–1002.
Choudhary N, Bhatt LK, Prabhavalkar KS. Experimental animal models for rheumatoid arthritis. Immunopharmacol Immunotoxicol. 2018;40(3):193–200.
Schuh BM, Macáková K, Feješ A, Groß T, Belvončíková P, Janko J et al. Sex differences in long-term effects of collagen-induced arthritis in middle-aged mice. Front Physiol.14:1195604.
Knight JS, Carmona-Rivera C, Kaplan MJ. Proteins derived from neutrophil extracellular traps may serve as self-antigens and mediate organ damage in autoimmune diseases. Front Immunol. 2012;3:380.
Wright HL, Lyon M, Chapman EA, Moots RJ, Edwards SW. Rheumatoid arthritis synovial fluid neutrophils drive inflammation through production of chemokines, reactive oxygen species, and neutrophil extracellular traps. Front Immunol. 2021;11:3364.
Metzemaekers M, Malengier-Devlies B, Yu K, Vandendriessche S, Yserbyt J, Matthys P, et al. Synovial fluid neutrophils from patients with juvenile idiopathic arthritis display a hyperactivated phenotype. Arthritis Rheumatol. 2021;73(5):875–84.
Varady NH, Schwab P-E, Kheir MM, Dilley JE, Bedair H, Chen AF. Synovial fluid and serum neutrophil-to-lymphocyte ratio: novel biomarkers for the diagnosis and prognosis of native septic arthritis in adults. JBJS. 2022;104(17):1516–22.
Zhang L, Yuan Y, Xu Q, Jiang Z, Chu C-Q. Contribution of neutrophils in the pathogenesis of rheumatoid arthritis. J Biomedical Res. 2020;34(2):86.
Schneider AH, Taira TM, Públio GA, da Silva Prado D, Donate Yabuta PB, Dos Santos JC, et al. Neutrophil extracellular traps mediate bone erosion in rheumatoid arthritis by enhancing RANKL-induced osteoclastogenesis. Br J Pharmacol. 2024;181(3):429–46.
O’Neil LJ, Barrera-Vargas A, Sandoval-Heglund D, Merayo-Chalico J, Aguirre-Aguilar E, Aponte AM, et al. Neutrophil-mediated carbamylation promotes articular damage in rheumatoid arthritis. Sci Adv. 2020;6(44):eabd2688.
Wilson AS, Randall KL, Pettitt JA, Ellyard JI, Blumenthal A, Enders A, et al. Neutrophil extracellular traps and their histones promote Th17 cell differentiation directly via TLR2. Nat Commun. 2022;13(1):528.
Hashimoto T, Yoshida K, Hashimoto N, Nakai A, Kaneshiro K, Suzuki K, et al. Circulating cell free DNA: a marker to predict the therapeutic response for biological DMARDs in rheumatoid arthritis. Int J Rheum Dis. 2017;20(6):722–30.
Phillipson M, Kubes P. The neutrophil in vascular inflammation. Nat Med. 2011;17(11):1381–90.
Radic M, Muller S. LL-37, a multi-faceted amphipathic peptide involved in NETosis. Cells. 2022;11(15).
Van Holde K, Allen J, Tatchell K, Weischet W, Lohr D. DNA-histone interactions in nucleosomes. Biophys J. 1980;32(1):271–82.
Hakkim A, Fürnrohr BG, Amann K, Laube B, Abed UA, Brinkmann V et al. Impairment of neutrophil extracellular trap degradation is associated with lupus nephritis. Proceedings of the National Academy of Sciences. 2010;107(21):9813-8.
Puccetti A, Madaio MP, Bellese G, Migliorini P. Anti-DNA antibodies bind to DNase I. J Exp Med. 1995;181(5):1797–804.
Funding
This work was supported by the Slovak Research and Development Agency (APVV-18-0366).
Author information
Authors and Affiliations
Contributions
Authors contributionsP.C., B.V., E.Š contributed to study concept and design. K.M., V.B., M.T., J. J., and M.P. performed in vivo and in vitro experiments. R.H. and N. M., performed the histology analysis. K. M., V.B., and M.T. analyzed the results and made the figures. K.M., V.B., and P.C., wrote the final draft of the article. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if you modified the licensed material. You do not have permission under this licence to share adapted material derived from this article or parts of it. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by-nc-nd/4.0/.
About this article

Cite this article
Macáková, K., Borbélyová, V., Tekeľová, M. et al. Effects of exogenous deoxyribonuclease I in collagen antibody-induced arthritis. J Inflamm 21, 36 (2024). https://doi.org/10.1186/s12950-024-00403-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12950-024-00403-z