
Abstract
Vigorous CD8+ T cells play a crucial role in recognizing tumor cells and combating solid tumors. How T cells efficiently recognize and target tumor antigens, and how they maintain the activity in the “rejection” of solid tumor microenvironment, are major concerns. Recent advances in understanding of the immunological trajectory and lifespan of CD8+ T cells have provided guidance for the design of more optimal anti-tumor immunotherapy regimens. Here, we review the newly discovered methods to enhance the function of CD8+ T cells against solid tumors, focusing on optimizing T cell receptor (TCR) expression, improving antigen recognition by engineered T cells, enhancing signal transduction of the TCR-CD3 complex, inducing the homing of polyclonal functional T cells to tumors, reversing T cell exhaustion under chronic antigen stimulation, and reprogramming the energy and metabolic pathways of T cells. We also discuss how to participate in the epigenetic changes of CD8+ T cells to regulate two key indicators of anti-tumor responses, namely effectiveness and persistence.
Similar content being viewed by others
Introduction
The rapid development of cancer immunotherapy has proven to be effective in restoring T cell-mediated immune responses [1, 2]. The combined application of immune checkpoint blockade and adoptive cell therapy has not only achieved unprecedented clinical responses in patients with refractory solid tumors but also brought long-term clinical remission to patients who were considered incurable due to tumor metastasis [3,4,5]. The cornerstone of immunotherapy is Cluster of differentiation 8+ (CD8+) cytotoxic T lymphocytes, which can directly destroy cancer cells [6]. Therefore, promoting CD8+ T cell-specific immune responses is considered as the focus of current cancer immunotherapy, and effector CD8+ T cells (Teff) are the preferred immune cells against tumors.
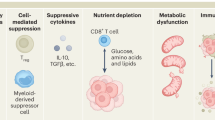
Naive CD8+ T cells, stimulated by tumor-associated antigens presented by the major histocompatibility complex (MHC) on the surface of tumor cells, are promoted to differentiate into Teff under the assistance of co-stimulatory molecules [7]. This process is accompanied by transcriptional, epigenetic, and metabolic reprogramming, as well as the acquisition of characteristic effector features, such as the ability to produce cytokines and cytotoxic molecules [8]. However, persistent tumor antigen stimulation can lead to the transition of CD8+ Teff to exhausted CD8+ T cells (Tex). T cell exhaustion was first characterized in lymphocytic choriomeningitis virus (LCMV)-specific T cell receptor (TCR) transgenic CD8+ T cells during chronic infection [9]. The characteristics of Tex in tumors primarily include the loss of effector function, persistent and elevated expression of inhibitory receptors (IRs), alterations in epigenetic and transcriptional profiles, and unique metabolic pathways [10, 11]. In most solid tumors, various reasons lead to a low burden of intracellular mutant antigens or down-regulation of MHC molecule expression in tumor cells, resulting in tumor cells being insufficient to be recognized by TCRs or unable to trigger the expansion and functional activation of CD8+ Teff after TCRs recognition [12,13,14]. The complex immune microenvironment of solid tumors hinders the high-throughput infiltration of CD8+ Teff, furthermore, the crosstalk between tumor cells, tumor matrix, and myeloid-derived suppressor cell populations (MDSCs) in the tumor microenvironment (TME) inhibits TCR signal transmission and limits Teff function [15]. Meanwhile, nutritional stress and metabolic waste accumulation caused by cancer cells [16] also induce energy and biosynthetic metabolic disorders, as well as alterations in transcriptome and epigenetic characteristics, thereby accelerating the exhaustion process of CD8+ Teff cells [17, 18].
With the rapid advancements in the fields of antigen identification, T cell biology, and gene therapy, a variety of anti-tumor immunotherapeutic strategies based on T cells have been proposed to enhance T cell function and redirect tumor-specific antigens [19]. The application of adoptive cell therapy, which involves the ex vivo expansion of tumor-infiltrating CD8+ T lymphocytes from patients or CD8+ T lymphocytes from donors stimulated by specific tumor antigens to sufficient numbers, and then reinfusion into patients who have undergone lymphocyte chemotherapy, has largely compensated for the low immunogenicity of “cold tumors” and the immune blockade of the suppressive tumor microenvironment [20,21,22]. The application of genetic engineering in adoptive cell therapy (ACT) has accelerated the progress of therapies involving chimeric antigen receptor (CAR) and engineered T cell receptor (TCR)-T cells. The antigen recognition of CAR-T relies on the single-chain variable fragment (ScFv) on the engineered receptor binding to the antigen on the membrane surface [23]. This presents challenges as it may not effectively recognize intracellular tumor antigens and cannot avoid the recognition of somatic common antigens, potentially leading to a low anti-tumor immune response and high off-target cytotoxicity [24, 25].TCR-T, which lyse tumor cells through exogenous affinity-matured TCRs, strictly recognizes antigen epitopes presented by MHC-I class molecules, making low mutation load neoantigens within tumor cells effective targets [26]. Importantly, a series of neoantigens have been identified, and multiple clinical trials have demonstrated the efficacy of TCR-T cell therapy against solid tumors [27,28,29].In addition to cell therapy, the application of engineered TCRs in T cell mobilization molecules, as well as the design of TCR mimic antibodies (TCRm Abs) based on the trajectory of TCR recognition of peptide-MHC (p-MHC) complexes, all contribute uniquely to the redirection of Teff in solid tumors [30,31,32]. In this review, we focus on the phenotypes of CD8+ T cells, discussing numerous feasible strategies to enhance CD8+ T cell therapy for solid tumors in the process of the initial expression and assembly of TCR to the activation and exhaustion of T cells, from the perspectives of T cell structure and genetic engineering modifications, and immune and metabolic program regulation.
Engineering of CD8+ T cells potentiate the anti-tumor efficacy
T cell receptor (TCR) engineered T cell therapy
The interaction between T cell receptors (TCRs) and their homologous tumor antigens is the core of anti-tumor immune responses. However, the inevitable heterogeneity of immune composition and phenotype spectrum within tumor-infiltrating lymphocytes [33,34,35] leads to low reactivity and variability in the intrinsic ability of TCRs to recognize autologous cancers [36, 37]. Neoantigen-specific TCRs (NeoTCR), due to their central tolerance, exhibit a higher affinity for mutated p-MHC with stronger interaction and stability [38, 39], and their ability to induce tumor regression has been characterized in various refractory solid tumors originating from different types of somatic cell mutations [27, 29, 31, 40]. Understanding the structural details of the interaction between TCR and its homologous p-MHCs provides a theoretical basis for exploring the structure-function relationship of neoantigen-reactive TCRs [41, 42]. NeoTCR-T cell therapy is currently a key area of exploration in immunotherapy. The development of individual cancer epitope identification technology has overcome the bias caused by the diversity of human leukocyte antigen (HLA) alleles and the huge spatial potential of peptide epitopes [43, 44]. The combination of isolating the original paired TCRαβ chains from polyclonal T cells and TCR response analysis based on next-generation sequencing (NGS) provides opportunities for high-throughput screening of endogenous reactive TCRs [45,46,47] but lacks control over the accuracy and frequency of individual TCRs in the library. The “sequence-and-synthesize” based ex vivo TCR gene editing [48, 49] and the “T-FINDER” [50], that combines neoantigen epitope identification and specific TCR screening, have made good supplements to this. Although the tumor neoantigen mutation burden (TMB) of individuals is directly related to the reactivity of NeoTCR-T cells [51], the heterogeneity of TMB emphasizes the importance of increasing the stability, affinity, and persistent functional activity of adoptively transferred NeoTCR-T cells in enhancing ACT [52].
Enhancing correct TCR membrane expression in engineered T cells
The primary success of tumor antigen-specific TCRs adoptive cell therapy is achieved through the transition to physiological T cells [53]. Previously, the transfer of exogenous TCR genes was primarily mediated by viruses [54], then semi-randomly expressed in engineered T cells, and assembled on the cell membrane with the CD3 signaling apparatus into an octameric TCR-CD3 complex with a 1:1:1:1 stoichiometry of TCRαβ:CD3γε:CD3δε:CD3ζζ [55]. A higher density of TCRs on the cell surface is believed to facilitate reaching the T cell activation threshold more easily [56]. However, due to the presence of endogenous TCRs, the expression of transgenic TCRs is often suppressed [57], or erroneous TCRαβ pairing occurs, causing engineered T cells to produce new antigenic properties, which may trigger auto-reactivity [58] (Fig. 1a). Therefore, ideally, successful TCR-T cells require efficient expression of the required αβ chains and are subject to physiological regulation, which is a major challenge in the field of TCR engineering.

Enhancement of Neoantigen-Specific Engineered TCR Expression Efficiency. (a) CD8+ T cells are transduced with neoantigen-specific engineered TCRs via lentivirus. Both endogenous and engineered TCRs are expressed on the cell surface, with a potential for mismatched TCR expression. (b) TCR domain engineering strategies are employed to reduce mismatched TCR expression. These strategies include the introduction of murine TCR constant domains, replacement of TCRαβ chain constant domains, or co-expression of TCRαβ chains with CD3ζ. Additionally, amino acid modifications such as the introduction of optimal amino acids, additional disulfide bonds, elimination of conserved N-glycosylation sites, increased hydrophobicity of the TCR transmembrane domain, and substitution of specific structural anchor amino acids in TCRαβ enhance engineered TCR expression. (c) CRISPR-Cas9 base editing is utilized to inhibit endogenous TCR expression, thereby reducing mismatched TCRs. CRISPR-Cas9 gene editing knocks out endogenous TCR genes and precisely integrates engineered TCR genes to promote their expression
For this reason, many biological modification methods applied to the TCRαβ chain structural domain have been attempted (Fig. 1b). Since the mouse TCR constant domain (C) α/β preferentially pairs, replacing its amino acid residues with the human TCR Cα/Cβ sequence can reduce the mismatch of human transgenic TCR [59], and minimally murinized TCR mutants also support the preferential pairing of transferred TCR chains [60]. Although murinized TCR chains may introduce potentially immunogenic exogenous sequences, no correlation has been found between immunogenicity and T cell functionality and clinical outcomes in patients receiving such engineered T cell therapy [61]. By swapping the therapeutic TCR Cα/Cβ structural domains, domain-swapped TCRs can assemble with CD3 and express on the T cell surface, reducing mismatches while excluding heterologous sequences in cell products [62]. A 3-domain single-chain TCRs framework [63] that depends on co-expression of the TCRα constant region can prevent unnecessary TCRαβ pairing and maintain a structure and functionality similar to the fully assembled double-chain TCR/CD3 complex [64]. In addition, co-transduction of TCR and CD3ζ increases the surface expression of the introduced TCR and reduces the mismatch with endogenous TCR chains [65], and its genetic tandem with auxiliary or co-stimulatory molecules can also enhance cell co-stimulation [66].
In addition to overall domain modifications, residue modifications between receptor chains are also expected to promote stable TCR expression. In this regard, various strategies have been designed and functionally validated, including optimal amino acid substitutions within the TCR framework region [67], the introduction of additional disulfide bonds at the TCR Cα/Cβ interface [68], selective removal of conservative N-glycosylation sites [69], and selective increase of TCRαβ Transmembrane domain (TMD) hydrophobicity [70]. Furthermore, swapping amino acid residues at the TCR Cα/Cβ interface that are linked to “knob-into-hole” structural interactions and electrostatic environments can also promote selective pairing of introduced TCRs [71]. Simultaneously, synonymous codon substitutions in the TCR coding sequence, as well as the omission of messenger ribonucleic acid (mRNA) instability motifs and implicit splice donor sites, allow for efficient expression of transgenic TCRs [72].
These efforts have indeed increased the cell membrane expression of transgenic TCRs, but mismatches cannot be eliminated. With the deep exploration of the field of genome editing, it has been found that actively knocking out endogenous TCR genes in engineered T cells fundamentally aids the efficient and stable expression of transgenic TCRs [73] (Fig. 1c). Initially, zinc-finger nucleases (ZFNs) [74] and transcription activator-like effector endonucleases (TALENs) [75] were used to disrupt endogenous TRAC and TRBC1/2 sites in engineered T cells, and subsequently emerged clustered regularly interspaced short palindromic repeats (CRISPR)– CRISPR-associated protein 9 (Cas9) [76] completed this work more efficiently [77]. However, these strategies induce double-strand DNA break behavior, which may lead to off-target genomic effects [78]. To change this situation, a CRISPR-based DNA base editor has been developed, which does not disrupt protein expression by inducing DNA double-strand breaks [79], and its application at TRAC, TRBC1/2 sites may provide a safer choice for genetically deleting endogenous TCR [80, 81]. In addition, CRISPR-Cas9 precise genome editing that does not rely on viral vectors can target the integration of NeoTCRs genes at the original TRAC site while knocking out endogenous TCR [29, 82, 83]. Gene-targeted integration is achieved through co-electroporation of human primary T cells, CRISPR-Cas9 ribonucleoprotein (RNP) complexes, and homologous double-stranded DNA repair templates. Compared with the semi-random insertion of NeoTCR genes by viral transduction, targeted integration allows NeoTCR genes to be physiologically regulated by endogenous promoters, providing engineered T cells with enhanced functionality and predictable safety, and reducing immune exhaustion [73]. Although the efficiency of targeted integration is lower than viral transduction, various improvement methods, such as single-stranded DNA (ssDNA) homology-directed repair (HDR) templates [84], nanoplasmid [85], stabilization of Cas9 RNP sand enhancement of template nuclear shuttling [86], have been designed to improve editing efficiency.
Increasing TCR effective recognition of antigen
The activation, proliferation, infiltration, and effector function of CD8+ T cells, as well as the duration of T cell responses, partly depend on the recognition strength of TCR to its homologous p-MHC complexes [87, 88]. This recognition strength is determined by the structural affinity based on the interaction of TCR with p-MHC complexes [89], which can be characterized by the dissociation kinetics of TCR with monomeric p-MHC complexes [90]. The non-covalent binding between different residues in TCR and p-MHC complexes reflects the contribution of this site to TCR antigen recognition. By analyzing the shared base sequence of TCR recognition-related core sequence, key conservative residues driving TCR recognition can be found, providing insights for analyzing the potential features of TCR epitope specificity spectrum [91]. The complementary determining regions (CDRs) in the variable structural domain of TCRαβ are crucial in the interaction with p-MHC complexes due to their antigen recognition and high variability. Usually, the CDR1 and CDR2 loops encoded by the same lineage gene have conservative interactions with MHC molecules, while the CDR3 loop is highly variable and mainly responsible for antigen peptide recognition [92]. The original structure of TCR can be artificially modified. The exploration of CDRs affinity maturation has been ongoing, and related work is dedicated to mutating CDRs residues to enhance TCR affinity and determine its basic characteristics and core base sequence [93, 94]. Mutation of a single CDR3 loop [95] or combined CDRs loops [96] to increase TCR affinity is a mature strategy, theoretically leading to up to 107 times higher affinity than the natural sequence [93], and its safety and effectiveness have been confirmed in subsequent clinical trials. In addition to CDRs directed evolution, the TCR affinity engineering platform constructed by ex vivo antigen-driven progenitor T cells has also been verified [97]. Changes in CDRs sequence may also change TCR antigen specificity, bringing antigen cross-reactivity while increasing affinity, which may cause fatal off-tumor or off-target toxicity [98, 99]. The combination of structure-guided positive and negative design, to fine-tune TCR in a way that reduces cross-reaction but keeps affinity close to the optimal level [100], seeks to redistribute attractive interactions on the TCR-p-MHC interface, thereby reducing the recognition of off-target peptides [101]. For TCR-p-MHC without readily available crystal structure, deep learning algorithms have been used for three-dimensional predictive analysis to train TCR’s binding ability to p-MHC complexes [102]. AlphaFold, a generative AI model using neural network architecture, can be applied to the development of a generalizable prediction model for TCR and p-MHC complexes specific binding [103]. With the iterative evolution of the model, the third-generation AlphaFold predicts the structure of TCR and p-MHC complexes interaction with unprecedented accuracy [104].
Mechanical transmission can enhance TCR’s recognition of p-MHC complexes. After TCR binds to the ligand, T cells change the TCR-p-MHC bond lifetime by applying endogenous forces - excitatory forces promote the formation of “catch bonds” to extend bond life, antagonistic forces promote the formation of “slip bonds” to shorten bond life [105]. More stable ligand recognition induces the formation of longer-lasting TCR-pMHC-CD8 three-molecule “catch bonds” [106, 107], which is a trans (lateral) heterodimer formed by the co-receptor CD8 interacting with the extracellular p-MHC complexes and the intracellular TCR-CD3 complexes and coupled by related kinases, which can transmit signals from the inside out [108]. It has been found that the FG loop conformation of the TCRβ chain constant structural domain controls the catch bond life of the variable structural domain module and controls peptide recognition through force-driven conformational changes [109]. Therefore, understanding the internal mechanism of TCR mechanosensing can explain why and how force enhances TCR antigen recognition and signal transduction without changing structural affinity [110]. Excitingly, the biophysics-based “catch bond” engineering that has emerged in response can increase TCR sensitivity while reducing potential adverse cross-reactivity. A feasible strategy is to introduce polar or charged amino acids into the CDRs residues that do not directly contact TCR and p-MHC complexes, to increase the molecular resistance of hydrogen bonds or salt bridges faced during the dissociation process of TCR-p-MHC trimer, thereby maintaining stable antigen epitope recognition and binding of p-MHC complexes [111] (Fig. 2).

Enhanced and Stabilized Neoantigen Recognition of TCR. Phage display platforms can be used to screen for mature, antigen-specific TCRs with high affinity, but there may be unknown reactivities. Structure-guided positive and negative selection of TCR design, which increases the connection between TCR and antigen epitopes while reducing the interaction between TCR and MHC, can significantly reduce cross-reactivity. Using a TCR affinity engineering platform built on progenitor T cells, it is possible to screen for neoantigen-reactive TCRs with increased affinity and low cross-reactivity. Using an iteratively evolved protein structure prediction model based on deep learning algorithms, it is possible to accurately predict the structure of non-readymade TCR-p-MHC complexes, guiding the analysis of neoantigen-specific TCRs. Based on the mechanical model of TCR-p-MHC interaction, stabilizing the FG loop structure of the TCRβ constant domain and introducing intermolecular forces at non-direct contact sites of the TCR and p-MHC complexes can induce the formation of catch bonds and extend their lifespan, enhancing TCR recognition of p-MHC complexes without significantly changing affinity
TCRmimic-engineered TCR-T cells
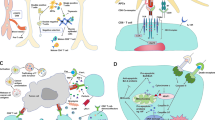
Synthetic TCR antigen receptor (STAR)/HLA-independent TCR (HIT) is a newly engineered receptor generated by replacing the variable domains of TCRα and β chains with the heavy and light chain variable domains of antibodies while retaining the complete TCR constant structural domain [112, 113]. Since only the variable domain of the receptor is changed, STAR/HIT can still form a complete TCR signal transmission module in conjunction with the CD3 complexes, and accept physiological regulation consistent with TCR. Compared to TCR-T and CAR-T, STAR/HIT-T cells, endowed with higher antigen affinity and superior tumor infiltration capabilities, demonstrate stronger antigen sensitivity and more stable cellular functions, enabling them to control, and even eradicate, tumor growth in various murine solid tumor models [112, 113], thereby providing a novel framework for the optimized design of engineered T cells. Co-stimulatory signals are a key factor in enhancing TCR signals. A switch fusion receptor called 80BB, composed of the extracellular structural domain of CD80 and the cytoplasmic structural domain of 41-BB, can transform signals from inhibitory receptors into co-stimulatory signals, enhance and maintain CD3-dependent T cell responses, and thereby improve the anti-tumor ability of STAR, HIT, or TCR-engineered T cells [114] (Fig. 3a). By concatenating modules that drive the nuclear factor (NF)-κB signal, namely MyD88 and CD40, to the C-terminus of the STAR β chain, a novel co-stimulation model is established, which gives rise to a variant of Co-STAR-T cells that not only exhibit enhanced sensitivity to antigens but also possess an improved ability to home to tumors over an extended period of time, thereby delivering superior anti-tumor efficacy, and paving the way for a new approach in T cell therapy specifically tailored for tumors characterized by an extremely low antigen density [115].

Affinity-enhanced TCR-mimic engineered T cells and bispecific protein molecules. a. The Synthetic TCR antigen receptor (STAR)/HLA-independent TCR (HIT), formed by replacing the physiological TCR variable domains with those of TCRm Abs or monoclonal antibodies (mAbs), recognize tumor-specific antigens (TSA) or tumor-associated antigens (TAA) and induce proximal phosphorylation signaling of the receptor-CD3 complex. The fusion protein 80BB, composed of the extracellular domain and transmembrane domain of CD80, and intracellular domain of 4-1BB, can convert the inhibitory signal from CTLA4 into a co-stimulatory signal, promoting proximal TCR signal transduction. The Myd88-cd40 co-stimulation module fused to the STARβ chain significantly enhances the antigen sensitivity of T cells. The bottom of the figure shows a schematic diagram of the gene arrangement of Co-STAR, STAR/HIT and 80BB, which are arranged in the order of proteins domains from the N-terminus to the C-terminus. Different chains are linked by the self-cleaving peptide sequence P2A. b. The immune-mobilizing monoclonal TCR against cancer (ImmTAC), composed of the soluble TCR targeting tumor-specific antigens and the single-chain fragment variable (ScFv) composed of the light and heavy chain variable structural domains of monoclonal antibodies targeting CD3ε, as well as the bispecific T cell engager (BiTE), composed of TCRm Abs or mAbs targeting tumor antigens and ScFv targeting CD3ε, induce polyclonal T cells to redirect against tumor cells and activate proximal TCR signals in a CD3-dependent manner. Bispecific protein molecules can be stably delivered to the tumor interior in the form of mRNA by lipid nanoparticle carriers (LNP). The bottom of the figure shows a schematic diagram of the gene arrangement of ImmTAC and BiTE, which are arranged in the order of proteins domains from the N-terminus to the C-terminus. Different chains are linked by the sequence of self-cleaving peptide P2A. Different domains within the same chain are connected by sequence of flexible linkers
Redirection of T cells potentiate the anti-tumor efficiency
Immune-mobilizing monoclonal TCR against cancer (ImmTAC)
In most solid tumors, low immunogenicity and a highly suppressive tumor microenvironment limit the infiltration and activation of CD8+ T cells. At the same time, small soluble proteins that target both tumor cells and T cells can overcome the biophysical characteristics of tumors, utilize natural antigen presentation pathways and specific antigen recognition structures to target tumor cells, trigger the relocation of polyclonal CD8+ T cells in the interstitium, promote the formation of immune synapses, and ultimately transform the immune rejection phenotype of the tumor into an immune inflammation phenotype (Fig. 3b).
TCRαβ chains are recombinantly expressed as soluble single-chain heterodimers, which are then fused with a single-chain variable fragment (ScFv) derived from an anti-CD3ε antibody through a flexible linker to generate a bispecific protein that can simultaneously target polyclonal T cells and tumor cells expressing specific p-MHC complexes, known as ImmTAC [116]. In vitro, ImmTAC fused with a TCR that has increased affinity after CDRs mutation by biological display technology can mediate CD8+ T cells to show specific cell lysis to tumor cells with low antigen density [31, 116, 117]. Tebentafusp, which specifically recognizes HLA-A*02:01 complexed with the melanoma-associated antigen gp100, is currently the only approved ImmTAC molecule for the treatment of unresectable or metastatic uveal melanoma and other solid tumors [118]. Propelled by the success of tebentafusp, the phase I/II clinical trials of ImmTAC molecules, which target the HLA complexed with the tumor antigens melanoma-associated antigen A4 (MAGE-A4)、New York esophageal squamous cell carcinoma 1 (NY-ESO-1) and L antigen family member-1 A (LAGE-1 A), have been completed (NCT03973333、NCT03515551), and concurrently, ImmTAC molecule that targets the HLA complexed with the tumor antigen preferentially expressed antigen in melanoma (PRAME) is preparing to enter clinical trials (NCT04262466).
Bispecific T cell engager (BiTE)
Another bispecific protein is the BiTE (Fig. 3b), which is composed of two ScFvs derived from antibodies - one recognizes the p-HLA epitope on the surface of tumor cells, and the other binds to the CD3 complexes on the surface of T cells, and is covalently connected by a peptide linker. It has achieved significant success in hematological malignancies [119, 120]. Compared to TCR, using ScFv as an antigen binding domain derived from TCR mimic Abs has the advantage that they are easy to improve in affinity and high-throughput synthesis by biological display technology [121]. With the advancement of high peptide specificity and MHC-restricted TCRm Abs screening and synthesis methods [30], a growing number of TCRm Abs targeting solid tumor neoantigens considered undrugneable and monoclonal Abs targeting solid tumor-associated antigens have been identified and used to synthesize BiTEs for anti-tumor immunity [122,123,124]. At present, a phase I clinical trial of a TCR-like antibody targeting MAGE-A4 in patients with recurrent or refractory (R/R) metastatic solid tumors is underway (NCT03132922) [125, 126]. Another delta-like ligand 3 (DLL3)/CD3 BiTE targeting DLL3, IMDELLTRA™ (tarlatamab-dlle), has completed phase I and II clinical trials (NCT03319940, NCT05060016) [127, 128], and has recently received accelerated approval from the U.S. Food and Drug Administration (FDA), becoming the first and only T cell engager for the treatment of extensive-stage Small-cell lung cancer (SCLC) [129]. Moreover, the tri-specific DLL3-targeting T-cell engager [130], which additionally targets the albumin binding domain to extend its half-life, is currently undergoing phase I / II clinical trials (NCT04471727) in patients with DLL3-associated cancers. Furthermore, bispecific T cell engagers targeting glypican-3 (GPC3), prostate-specific membrane antigen (PSMA) and claudin (CLDN) 18.2 are respectively undergoing phase I clinical trials in patients with hepatocellular carcinoma (HCC) (NCT05450562), metastatic castration-resistant prostate cancer (mCRPC) (NCT04740034) and pancreatic ductal adenocarcinoma (PDAC) and gastric and esophageal malignancies (NCT05164458, NCT05164458). Simultaneously, a bispecific T cell engager targeting the epithelial cell adhesion molecule (EpCAM) is in phase II clinical trials in patients with malignant ascites from epithelial cancers (NCT06266091). As well known, the tumor affinity of BiTE can be improved by changing the antibody structure to increase the number of binding elements targeting tumor cells. Cibisatamab, a bispecific T cell engager with two Fab structural domains recognizing carcinoembryonic antigen (CEA) and one Fab structural domain binding to CD3 [131], is currently undergoing a phase I clinical trial (NCT02324257) in patients with locally advanced or metastatic solid tumors expressing CEA.
Due to their relatively small molecular weight, ImmTAC and BiTE face a relatively high plasma clearance rate in therapeutic activities, leading to a short plasma half-life in patients, thereby preventing them from fully exerting their functions. In the past, a method of fusing small molecule proteins with Fc molecules modified by IgG to extend the half-life had been proposed [132]. With the development of nanocarrier technology, lipid nanoparticles (LNPs) with superior targeting and pharmacokinetics have been widely used for the in vivo delivery of mRNA vaccines [132]. Inspired by cancer mRNA vaccines, an mRNA (RiboMABs) containing 1-methyl pseudouridine for encoding His-tagged Bispecific T cell engager antibodies have been designed [133]. Recently, a clinical trial is underway to improve the efficacy of bispecific antibodies by applying BNT142, which is a lipid nanoparticle (LNP), formulated RiboMABs encoding claudin 6 (CLDN6) × CD3, in patients with late-stage solid tumors positive for the oncofetal antigen CLDN6 (NCT05262530) [134].
Effective TCR signaling is beneficial for T cell activity
Proximal signal transduction of TCR
The activation of CD8+ T cells largely depend on the signal transduction following TCR recognition of tumor antigens. The signal transduction process of antigen recognition is accompanied by conformational changes in the TCR-CD3 complex [135,136,137]. Therefore, the structural and functional integrity of the TCR-CD3 complex, as well as the coupling of LCK proto-oncogene, Src family tyrosine kinase (LCK) with signaling modules, is crucial for maintaining TCR signal transduction and activating T cells (Fig. 4a). The interaction of conserved polar residues within the TCRαβ TMD is key to the assembly of the TCR-CD3 modules [138]. The initial signal of antigen recognition is transmitted through the TCRαβ TMD to all modules of the TCR-CD3 complex, accompanied by structural changes in the signaling modules [139]. Understanding the structural changes in TM module interactions induced by ligand binding helps elucidate the mechanisms of TCR signal transduction and identify potential targets for enhancing TCR signaling. The membrane-embedded CD3ζζ contains a highly specific TM helical dimer interface, which is a key molecule coupling the TCR to intracellular signals [140]. In this process, the precise positioning of arginine within the TCRα TMD is critical [141]. Early experiments indicated that TCR binding triggers spatial changes between CD3ζζ subunits [142]. Subsequent studies defined a dynamic structural motion model of TMD related to the mechanobiology of the TCR-CD3 complex. This model revealed that force transmission induced by ligand recognition leads to a critical tilt of the TCRα TM helix, disrupting the interaction between Arg251 and CD3ζζ [143], causing the CD3ζζ tail to shift from lipid rafts to the cytoplasm, then providing tyrosine phosphorylation sites for LCK and promoting downstream signal transduction [144, 145]. Activated LCK recruits and phosphorylates ζ-chain-associated protein kinase 70 (ZAP-70) by phosphorylating immunoreceptor tyrosine-based activation motifs (ITAMs) [146]. The activated pZAP-70 further phosphorylates the key adaptor protein linker for activation of T cells (LAT) and promotes activation of TCR downstream signaling [147].
The strength of TCR signaling is a key determinant of CD8+ T cell anti-tumor responses. The availability of various proteins involved in TCR signal transduction can regulate T cell triggering, and their functional states under regulatory factors also influence the fate decisions of T cells post-triggering [148]. The protein tyrosine phosphatase Src homology region 2 domain-containing phosphatase (SHP) are key participants in the dephosphorylation reactions of the TCR signaling pathway of protein tyrosine phosphatases non-receptor (PTPN). They inhibit TCR signal transduction by directly or indirectly inactivating Src family kinases [149, 150]. The instability of SHP and PTPN is an important mechanism for enhancing TCR signal cascade reactivity. Studies have found that a serine/threonine kinase belonging to the STE-20 kinase family, Thousand-and-one-amino acid kinase 3 (TAOK3), acts as a rheostat for TCR signal transduction by regulating SHP-1 abundance, thereby determining the activation threshold of T lymphocytes [151]. Additionally, highly efficient and selective competitive inhibitors of PTPN active sites can achieve the loss of PTPN dephosphorylation function, thereby increasing T cell activation and recruitment [152]. Similarly, the suppressor of T cell signaling-1 (STS-1) encoded by UBASH3B can negatively transmit TCR signals by dephosphorylating pZAP-70 [153], and this biological function has been shown to be inhibited by the 2-(1 H)-quinolinone derivative Rebamipide [154]. Tense TCR signaling upregulates the expression of Cbl proto-oncogene, E3 ubiquitin protein ligase (CBL) in initial T cells [155], which then ubiquitinates and negatively regulates the LCK-ZAP-70-LAT axis to inhibit the transmission of proximal phosphorylation signals [156]. Understanding the negative feedback loop that exists between CBL and TCR signaling, which functions to temporally calibrate TCR signaling, helps selectively block CBL to enhance TCR signaling [157]. Furthermore, various LCK-dependent methods, such as CD61 and CD103 co-localization with TCR [158], CD146 homodimer-induced LCK autophosphorylation [159], and disulfiram increasing LCK kinase activity [160], have been shown to enhance proximal TCR signal transduction.
Distal signal transduction of TCR
Downstream signals transmitted by the TCR signal complexes, in conjunction with co-stimulatory signals including CD28 and environmental signals, synergistically regulate the functional activation of CD8+ T cells. Multiple adaptive immunity signaling pathways participate in TCR signal transduction (Fig. 4b). When antigen-MHC complexes bind to TCRs, protein tyrosine kinases (PTKs) are activated, leading to the activation of phospholipase C-γ1. Phospholipase C-γ1 hydrolyzes membrane phospholipid phosphatidylinositol 4,5-bisphosphate, generating Inositol triphosphate (IP3) and diacylglycerol (DAG) [161]. IP3 binds to receptors on the endoplasmic reticulum membrane, causing the release of Ca2+ in the endoplasmic reticulum and activating calcium channels to control intracellular Ca2+ concentration [162]. Ultimately, the influx of Ca2+ activates the nuclear factor of activated T cells (NFAT) pathway through Ca2+-related proteins, regulating T cell metabolism and inducing cytokine production [163]. When TCR/CD28 connect, Phosphoinositide-dependent kinase 1 (PDK1) effectively activates the protein kinase C (PKC) θ through the phosphatidylinositol 3-kinase pathway [164]. Activated PKC phosphorylates serine residues in the caspase recruitment domain-containing membrane-associated guanylate kinase protein 1 (CARMA1) [165]. Then, B cell lymphoma/leukemia 10 (BCL10), and mucosa-associated lymphoid tissue translocation protein 1 (MALT1) are recruited, forming an active CARMA1-BCL10-MALT1 signal complex [166]. This promotes the activation of the IĸB kinase (IKK) complex and the degradation of IĸB, thereby allowing the NF-κB to translocate to the nucleus, initiating T cell activation, promoting T cell proliferation and cytotoxicity [167]. After TCR upstream signals are transmitted to Ras, Ras is activated by guanosine triphosphate (GTP) exchange, then sequentially activates Raf kinase, Mek kinase/Ras-extracellular signal-related kinase (ERK) kinase 1/2 and ERK, promoting the transcription of c-Fos, basic leucine zipper activating transcription factor (ATF)-like transcription factor (BATF), leading to the formation of the activator protein (AP)-1 complex, promoting the T cell activation program [168]. At the same time, ERK1/2-mediated SMAD family member 4 (SMAD4) Ser367 site phosphorylation induces SMAD4 nuclear translocation, promoting the activation and effector function of CD8+ T cells [169]. The mammalian target of the rapamycin (mTOR) pathway can be activated by various signals, regulating T cell survival and metabolic differentiation [170]. The mTOR signal transduction includes two different complexes, the activation of mTOR complex 1 (mTORC1) leads to the phosphorylation of S6 kinase 1 and the translation of 4E-BP1, while the activation of mTORC2 leads to the phosphorylation of Akt kinase [171]. The tuberous sclerosis complex (TSC)1 and 2 are tumor suppressors, they form heterodimers to regulate downstream signals in T cells [172]. When TCR stimulation and PKC induce TSC2 Ser1365 phosphorylation, the GTPase activity of the TSC1/2 complex is inhibited, leading to the activation of mTORC1 [171]. Interestingly, the expression of a phosphorylation-silent mutant TSC2-S1365A (SA) can significantly increase mTORC1 activation and T cell effector function [173]. T cells also have the potential to trigger specific effector functions independent of antigen receptor signal transduction. Analysis of chromatin accessibility in T cell subsets found that transcription factors RUNX1, PU.1, and BCL11B cooperatively recruit the chromatin remodeling complex mammalians switch/sucrose non-fermentable (mSWI/SNF) to the Teff gene locus to establish a balanced chromatin landscape, supporting T cell activation [174].

Signal transduction of TCR recognition of tumor antigens in CD8+ T cells. (a) The recognition of ligands by TCRαβ dimers triggers the transmission of information through the noncovalently associated CD3δε, CD3γε, and CD3ζζ modules, which contain cytoplasmic ITAMs phosphorylated by the Src-family kinase LCK. This phosphorylation event recruits ZAP-70, which is subsequently phosphorylated and activated by LCK. Activated ZAP-70 then phosphorylates the adaptor protein LAT, leading to the recruitment of additional signaling effectors. (b) TCR signaling induces extensive cellular changes in T cells, including the activation of transcriptional regulators, alterations in protein synthesis, and metabolic shifts. These changes guide the proliferation and effector differentiation of naive T cells. The key signaling pathways, as shown from left to right in the figure, are: [1]. PLCγ1 Pathway: Phosphorylation of LAT recruits PLCγ1, which hydrolyzes PIP2 to generate IP3 and DAG. IP3 induces calcium release from the endoplasmic reticulum, while DAG activates PKC, leading to the activation of transcription factors such as NFAT [2]. CARMA1/BCL10/MALT1 Pathway: The CARMA1/BCL10/MALT1 complex is activated downstream of LAT, leading to the activation of the NF-kB pathway. This pathway is crucial for the transcription of genes involved in T cell survival and proliferation [3]. RasGRP1/Ras/Raf/MEK/ERK Pathway: LAT phosphorylation also recruits RasGRP1, which activates the Ras/Raf/MEK/ERK signaling cascade. This pathway culminates in the activation of the transcription factor AP-1, promoting gene expression changes necessary for T cell activation [4]. PI3K/AKT/mTOR Pathway: LAT recruits PI3K, which generates PIP3, leading to the activation of AKT. AKT activation promotes mTORC2 and subsequently mTORC1 activation, which regulates protein synthesis and cell metabolism, essential for T cell growth and differentiation
Reversing the exhausted state of CD8+ T cells
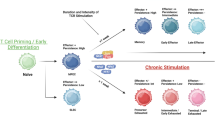
Effective CD8+ T cell responses are a key component of adaptive immune therapy for cancer. However, persistent TCR stimulation in solid tumors and the tumorigenic changes in the immune and metabolic landscape of the tumor microenvironment collectively drive naive CD8+ T cells to transition from Tex precursor population through Tex progenitor and Tex intermediate subset to ultimately differentiate into Tex terminally cells [175]. This primarily involves decreased cytokine secretion, sustained high expression of inhibitory receptors, metabolic changes, and unique transcriptional and epigenetic changes, ultimately leading to impaired effector function and reduced proliferation capacity of exhausted T cells [176,177,178]. The deepening understanding of the transcriptional, epigenetic, and metabolic characteristics that drive and maintain different exhaustion states of CD8+ T cells provide insights for designing immunotherapies that can reduce adverse events and improve tumor control.
The process of naive CD8+ T cells becoming progressively exhausted after receiving antigen stimulation signals is accompanied by changes in the Transcription factor (TF) motif, mainly characterized by a decrease in the availability of T-cell factor 1 (TCF1) (encoded by the TCF7 gene) and an increase in the availability of Thymocyte selection-associated high mobility group box protein (TOX) (encoded by the TOX gene) [179] (Fig. 5a). Compared with highly differentiated T cells, transferred T cell subsets originating from naive or stem cell-like cells exhibit stronger anti-tumor capabilities [180]. Among them, TCF1+ CD8+ T cells with stem cell-like characteristics are crucial for tumor control in cellular immunotherapy [4]. The HMG-box transcription factor TCF1, induced by the transcription factor Forkhead box O1 (FOXO1) binding to the TCF7 site [181], regulates the memory-like program of CD8+ T cells and is key to the development and maintenance of stem cell-like memory precursor T cells and Tex progenitor cells [182, 183]. The Inhibitor of DNA binding protein 2 (Id2) drives the stratified exhaustion of CD8+ T cells by disrupting the formation of the Tcf3-Tal1 transcription regulatory complex and recruiting the histone lysine demethylase 1 (LSD1), regulating the generation of Tex progenitor cells and their transition to Tex terminally cells, and promoting the maintenance of T cell stemness after adoptive cell therapy [184]. Another HMG-box transcription factor TOX is a central regulator of T cell exhaustion. When AP-1 is absent, the transcription factor NFAT imposes a negative feedback program on exhausted T cells, initiating TOX transcription and converting persistent stimulation signals into Tex-related transcriptional and epigenetic developmental programs [185, 186]. The interaction between BATF and interferon regulatory factor 4 (IRF4) can reduce the expression of TOX, promote the effector differentiation of the transcriptional and epigenetic landscape of exhausted T cells [187], thereby improving the anti-tumor response of engineered T cells [188].

The occurrence and rescue of CD8+ T cell exhaustion program within solid tumors. (a) In solid tumors, chronic antigen stimulation signals from the TCR are transduced and transformed into increased accessibility of TOX and decreased availability of TCF1, ultimately driving the occurrence of the exhaustion program characterized by increased expression of immunosuppressive receptors, which limits the anti-tumor ability of CD8+ T cells. (b) By blocking inhibitory receptor signals, regulating the availability of intracellular immune checkpoints, and knocking out or knocking in of gene loci with weakened or enhanced function, the exhausted state of CD8+ T cells can be effectively reversed, restoring their anti-tumor function. (c) The use of orthogonal cytokine receptor engineering and synthetic Notch (synNotch) receptor engineering to increase the availability and specificity of cytokines for CD8+ T cells enable engineered CD8+ T cells to achieve effective expansion and improved adaptability
Blocking immune checkpoint
Co-inhibitory receptors on the surface of T cells are a routine immune checkpoint in cancer immunotherapy and are key to enabling cancer cells to evade immune recognition and attack. Immune checkpoint blockade (ICB) therapy, by blocking the IRs signals on the surface of exhausted T cells, enables them to restore and maintain effector function to clear tumors, showing promise in the treatment of an increasing number of solid tumors [189] (Fig. 5b). Programmed death protein-1 (PD-1) and cytotoxic T lymphocyte antigen 4 (CTLA4) are the most well-known immune checkpoints, promoting immune tolerance by binding to their ligands PD-L1 and B7 family molecules respectively, and therapeutic antibodies targeting them have greatly changed cancer immunotherapy in the past [190]. Recent research has revealed a new structure-function relationship of PD-1, in which PD-1 and its ligand drive PD-1/PDL-1 cis-dimerization through non-covalent binding of the transmembrane structural domain, and enhance the functional inhibition of CD8+ Teff [191]. Therefore, manipulating the oligomeric state of the PD-1 axis is expected to provide an additional way to enhance cancer immunotherapy. Nevertheless, a portion of cancer patients still show low responsiveness to treatments targeting PD-1 and CTLA4, which has also promoted the exploration of the next generation of inhibitory receptors. Lymphocyte activation gene-3 (LAG3) is a co-inhibitory receptor highly expressed on exhausted T cells [192]. It colocalizes with the TCR during early T cell activation events [193], and increases the acidity of the immunological synapse through its cytoplasmic tail’s conserved EP motif, which is a tandem repeat of glutamic acid-proline sequences [194]. This leads to the uncoupling of LCK from CD8 and TCR-CD3 complexes, ultimately suppressing T cell activation [195]. T cell immunoglobulin-3 (Tim-3) is selectively expressed on CD8+ cytotoxic T lymphocytes (CTL) and is upregulated during CD8+ CTL dysfunction or exhaustion [196]. By interacting with its heterophilic ligand Carcinoembryonic antigen-related cell adhesion molecule-1 (CEACAM1), Tim-3 prevents the phosphorylation of TCR signaling modules within effector T cells, ultimately inhibiting T cell activation [197]. T cell immunoglobulin and immunoreceptor tyrosine-based inhibitory motif (ITIM) domain (TIGIT) is expressed on activated T cells, mainly connects to ligand CD155 in a CD226-independent manner, and forms nanoclusters located at the TCR immune synapse, inhibiting the acquisition of effector function and proliferation of effector T cells by downregulating the expression of TCR complex-related proteins and the central regulatory factors of the TCR signal cascade [198]. B and T lymphocyte attenuator (BTLA), a lymphocyte-specific cell surface co-inhibitory receptor, inhibits antigen-driven T cell activation and proliferation by binding to herpesvirus entry mediator (HVEM), and subsequently recruits SHP1/2 through the typical cytoplasmic domain of ITIM and immunoreceptor tyrosine-based activation motif (ITSM) [199, 200]. The clinical control of tumors has been enhanced to varying degrees in solid tumor therapy by blocking LAG3 [201, 202], Tim-3 [203, 204], TIGIT [205, 206], BTLA [207] to improve the adaptability of CD8+ tumor infiltrating lymphocytes (TILs). The CD200-CD200R axis is a potential target for antitumor immunomodulation. Typically, CD200+ CD8+ Teff exhibit improved antitumor functionality following immune checkpoint blockade therapy [208]. However, upon chronic antigen stimulation, the inhibitory receptor CD200R1 is upregulated on the surface of CD8+ Teff and suppresses T cell activity through its interaction with the ligand CD200 [209]. Selective blockade of CD200R1 can help restore the antitumor function of CD200R1+ CD8+ Teff [210]. V domain immunoglobulin suppressor of T cell activation (VISTA), a B7 family immune checkpoint protein expressed on T cells, drives T cell inhibition by binding to homologous ligands [211], it has been proven in tumor models that VISTA blockade can enhance the proliferation and cytotoxic function of CD8+ Teff [212]. Recently, a new immune checkpoint I-type single-pass membrane protein (ITPRIPL1) was characterized in tumors. The ITPRIPL1-CD3ε axis formed by the binding of ITPRIPL1 as a ligand on the surface of tumor cells with CD3ε can inhibit TCR-CD3 complexes signal transmission. Therapeutic antibodies targeting ITPRIPL1 can effectively enhance the cytotoxicity of tumor-specific T cells and achieve tumor growth control and are expected to be promising therapeutic targets for various types of tumors [213]. The development of bispecific antibody drugs that simultaneously target inhibitory receptors/ligands and another inhibitory receptor or co-stimulatory receptor, by selectively targeting immune checkpoints to rescue T cell exhaustion, provides additional options for enhancing the anti-tumor ability of CD8+ T cells [214]. More importantly, engineered synthetic ‘switch’ receptors-T cells that fuse inhibitory receptor structural domains to activation structural domains, including TIGIT-CD28 [215], CD200R-CD28 [216], TIM-3-CD28 [217], PD-1-CD28 [218], have shown superior anti-tumor capabilities in preclinical models or clinical trials, providing a realistic basis for remodeling solid tumor ACT.
In addition to cell surface inhibitory receptors, intracellular checkpoints are gradually being discovered as emerging targets (Table 1). Odd-skipped related 2 (Osr2) is a mechanoresponsive transcription factor, which is selectively induced in terminally exhausted tumor-specific CD8+ T cell subsets by TCR signal transduction and biomechanical stress mediated by the Piezo1/Ca2+/cyclic adenosine monophosphate(cAMP)-response element binding protein (CREB) axis. Subsequently, Osr2 recruits histone deacetylase 3 (HDAC3) to downregulate Histone H3 Lysine 27 acetylation (H3K27ac) signaling, inhibiting the expression of cytotoxic genes and exacerbating T cell exhaustion [219]. In solid tumor models, the absence of Osr2 can alleviate the exhausted state of transferred tumor-specific T cells and enhance their control of tumors, suggesting that Osr2, as a biomechanical checkpoint, holds promise as a new target for solid tumor immunotherapy. The co-stimulatory receptor CD226 is a key component of CD8+ T cell activation within tumors, and it can inhibit targets expressing CD112 and CD155 [220]. Overexpression of the transcription factor Eomesodermin leads to the loss of CD226 in CD8+ TILs, thereby blocking the effector program of T cells, and can serve as an intracellular immune checkpoint [221]. Regulating the expression and signal transduction of membrane receptors in CD8+ T cells through the control of post-translational processes of membrane proteins is an effective method to alter the anti-tumor effects of CD8+ T cells. The extracellular domain protease A disintegrin and metalloprotease 17 (ADAM17) inhibits the perception of interleukin-2 (IL-2) signals by CD8+ T cells by cleaving the membrane IL-2 Receptor β (CD122), thereby reducing the proliferation and effector differentiation of CD8+ T cells. In solid tumor models, the knockout or inhibition of ADAM17 can significantly upregulate the expression of CD122 on the membrane of CD8+ T cells and enhance their anti-tumor capabilities [222]. Additionally, CD8+ T cells have also generated their own regulatory signal transduction axis. In tumor-infiltrating exhausted CD8+ T cells, the immunosuppressive factor fibrinogen-like protein 2 (Fgl2) is expressed and positively regulated by FcγRIIB, which is the only inhibitory IgG-Fc receptor expressed in highly differentiated effector CD8+ T cells. Meanwhile, Fgl2 supports the suppression of the anti-tumor reactivity of CD8+ T cells by inducing caspase 3/7-mediated apoptosis through interaction with FcγRIIB [223]. It has been confirmed in mouse and human tumor models that inhibitory regulation of the Fgl2/FcγRIIB axis can effectively rescue disabled antigen-specific CD8+ T cells.
The operability of immune checkpoints is mainly described as a simple switching mechanism dependent on homologous ligand interactions, depending on intracellular factors that regulate the signal threshold of these receptors to inhibit T cell responses and is constrained by intracellular signal attenuators. T-lineage protein thymocyte-expressed molecule involved in selection (THEMIS) is an intracellular signal attenuator in T cells. It is recruited to the cytoplasmic structural domain of BTLA, and by interacting with growth factor receptor-bound protein 2 (GRB2) and SHP-1, it stabilizes the oxidation of SHP-1, thereby inhibiting BTLA signal transmission, ultimately promoting and maintaining the development of CD8+ T cells [224]. P-selectin glycoprotein-1 (PSGL-1) is an intrinsic immune checkpoint regulatory molecule of T cell exhaustion. It connects with TCR and inhibits ZAP-70 phosphorylation to block TCR proximal signal transmission by maintaining the expression of Ubiquitin-associated and SH3 domain-containing protein B, thereby limiting the activation of CD8+ T cell effector function and inducing the transition of T cells to exhaustion [225]. In addition, PSGL-1 can selectively bind to VISTA in the acidic tumor microenvironment, mediating the suppression of T cell activity [226]. Deletion or blockade of PSGL-1 allows CD8+ T cells to gain enhanced TCR sensitivity and metabolic characteristics, promoting their effector function and stem cell differentiation, and achieving successful tumor control in solid tumors resistant to PD-1 blockade [225].
Gene editing enhances the intrinsic adaptability of CD8+ T cells
Enhancing the adaptability of engineered therapeutic T cells under chronic antigen stimulation is also a promising strategy to improve anti-tumor efficacy. Due to the ability of the CRISPR/Cas9 gene editing system to precisely inactivate or knock-in target genes at the genomic DNA level, functional genomic screening based on CRISPR/Cas9 has been used to identify T cell therapy targets [181, 227] (Fig. 5b). Suppressor of cytokine signaling 1 (SOCS1), by negatively regulating the Janus kinase (JAK)/signal transducer and activator of transcription (STAT) signaling pathway, inhibits the accumulation of central memory T cells and Teff subsets in cancer and has been identified as an optimal T cell enhancement target. Targeting the SOCS1 SH2 structural domain with single guide RNA (sgRNA) via CRISPR/Cas9 to inactivate Socs1 in engineered T cell can enhance the reactivity of transferred T cells to cytokines and inhibit tumor growth in mice [228]. Intrinsic inflammatory regulatory factors Regnase-1, encoded by zinc finger CCCH-type containing 12A (ZC3H12A) and Roquin-1, encoded by ring finger and CCCH-type domains 1 (RC3H1) in T cells limit the expression of inflammatory genes by targeting and degrading the 3’-untranslated regions (UTRs) of inflammatory gene transcripts [229]. The absence of Regnase-1 reprograms CD8+ T cells to increase their inflammatory activity and persistence, thereby enhancing control of solid tumors [230], and co-knockout with Roquin-1 can significantly improve this anti-tumor function [231]. Protein tyrosine phosphatase PTPN2, encoded by PTPN2, regulates the generation of terminal exhaustion T cell subsets by weakening type I interferon signals. The absence of PTPN2 can restore the cytotoxicity and proliferative ability of exhausted T cells in a cytokine-independent manner [232], and enhance their control of solid tumors [150]. Moreover, small molecule inhibitors of the PTPN active site have achieved similar effects in preclinical models [233]. RAS P21 protein activator 2 (RASA2), encoded by RASA2, is a RAS GTPase-activating protein, which gradually increases in expression in CD8+ T cells under chronic antigen exposure. In the treatment of solid tumors with T cells in mice, the ablation of Rasa2 enhances the response of TIL to TCR stimulation through the mitogen-activated protein kinase (MAPK) signaling pathway, increases TIL antigen sensitivity, promotes its immune reprogramming, and ultimately achieves a more persistent effector function [234]. The family with sequence similarity 49 member B (FAM49B), encoded by CYFIP related Rac1 interactor B (CYRIB), is a regulator of proximal TCR signaling that inhibits the activation of CD8+ T cells by suppressing the active form of the small GTPase Rac and cytoskeletal actin dynamics [235]. The genetic elimination of CYRIB or the interaction between FAM49B and Rac can block the negative regulatory effect of FAM49B on T cells. The p38 kinase, encoded by MAPK14, has been identified as a central negative regulator of the effective anti-tumor phenotype characteristics of CD8+ T cells. Inhibiting Mapk14 is an effective method to promote the functional phenotype differentiation of T cells and enhance the anti-tumor effect of engineered T cells in mice [236]. The accessibility of the inositol requiring mutant 80 (INO80) and BRG1 or BRM-associated factor (BAF) chromatin remodeling complexes is related to the persistence of CD8+ T cells in tumors. The genetic deletion of chromatin remodeling factors, especially the BAF complex member Arid1a, can reduce the epigenetic characteristics related to CD8+ T cell exhaustion in mouses bearing melanoma, thereby increasing their function and persistence [237]. In addition to the majority of negative regulatory genes, some gain-of-function (GOF) gene targets within CD8+ T cells have also been screened. Proline dehydrogenase 2 (PRODH2), encoded by PRODH2, has been identified as a key regulatory enzyme for reprogramming proline metabolism in primary CD8+ T cells. Moreover, targeted knock-in of Prodh2 can reshape the metabolic group, transcriptome, and immune function of engineered T cells, thereby enhancing the anti-tumor efficacy in mice [238]. The caspase recruitment domain-containing protein 11 (CARD11)-phosphoinositide-3-kinase regulatory subunit 3 (PIK3R3) fusion gene, found in human skin T lymphocyte cancer, regulates T cell proliferation and differentiation by enhancing CARD11-BCL10-MALT1 complex signaling [239]. CD8+ engineered T cells carrying the Card11-Pik3r3 fusion gene have been shown to have increased functionality and stemness in animal model. Subsequently, in a screening of the whole genome of primary human T cells for functional key regulatory targets using sgRNA lentiviral infection with Cas9 protein electroporation (SLICE), casitas B-lineage lymphoma proto-oncogene B (CBLB) and CD5 were identified as negative regulators, and lymphocyte cytosolic protein (LCP) was identified as a positive regulator in T cell activation [240] (Table 2).
Recently, a programmable RNA targeting platform, multiplexed effector guide arrays (MEGA), was developed. This platform uses RNA-guided CRISPR-Cas13d with RNA targeting activity to perform highly multiplexed, quantitative, and reversible perturbation of the transcriptome of primary human T cells [241]. Combined with CRISPR combination screening, it can achieve synchronous regulation of paired regulatory factors affecting T cell function. Another modular platform for constructing DNA knock-in libraries, modular pooled knock-in screening (ModPoKI), evaluates hundreds to thousands of different T cell structures by merging hundreds of TFs and synthetic surface receptors (SRs) into different TCR-T cells, obtaining synthetic sequences targeting interested genomic binding sites [242]. In engineered T cell immunotherapy, this large-scale ModPoKI screening is a powerful method to accelerate cell state programming, helping to discover complex gene structures that program cell function, and successfully revealing the role of overexpressed TFAP4 and BATF in promoting the function and improving adaptability of engineered T cells in mouses bearing leukemic. In addition to genome editing, CRISPR-based base editing technology can be utilized for analyzing single-nucleotide variants (SNVs) in engineered T cells. Through this approach, gain-of-function mutations in genes encoding phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit delta (PIK3CD) and its related regulatory subunits have been identified to significantly enhance the anti-tumor capability of engineered CD8+ T cells ex vivo, which could potentially aid in guiding the engineering design of cell-based immunotherapy [243]. Engineered T cells expressing CD47 (Q31P) mediate an increase in macrophage-T cell crosstalk while evading macrophage phagocytosis, ultimately achieving more durable anti-tumor capabilities [244], it is anticipated to be rapidly constructed using CRISPR-based base editing technology.
Cytokine engineering reduces the occurrence of CD8+ T cell exhaustion
The binding of TCR/auxiliary receptors induces the activation of biochemical signaling pathways. These signaling pathways, in conjunction with signals from cytokine receptors, guide the outcome of the response. The lack of success of engineered TCR-T in solid tumor treatment may be related to the lack of available cytokine signals. Selectively enhancing the targeting of cytokines may provide new ideas for enhancing strategies for improved TCR-T cells [245] (Fig. 5c). However, some adoptive cell therapies supplemented with cytokines show off-target or systemic toxicity [246]. Therefore, strategies using cytokine engineering to actively regulate ILs/IL receptors (ILRs) expression are used to design the next generation of therapeutic T cells. An engineered IL-2 agonist, named H9T, can effectively bind to IL-2Rβ even in the absence of IL-2Rα [247]. It drives T cell proliferation while maintaining the expression of TCF-1, thereby promoting the maintenance of T cell stemness and preventing terminal differentiation of T cells [248]. Ultimately, it significantly prolongs the survival time of melanoma mouse models. Using orthogonal gene engineering, allowing T cells to express ortho IL-2Rβ, so that they can be selectively targeted by ortho IL-2 [249], or allowing T cells to secrete an IL-2 variant that can bind to IL-2Rβγ and the alarmin IL-33, can make adoptively transferred engineered T cells far from exhaustion and maintain superior effector function, ultimately improving the survival rate in mouses bearing advanced colon tumor [250]. Synthetic Notch (synNotch) receptor is a transmembrane receptor that converts signals from tumor-derived ligand-recognized extracellular structures into the release of designated transcription regulatory factors in intracellular structural domains, with orthogonality and flexibility [251]. Tumor-specific synNotch receptor engineered T cells based on the synNotch pathway can independently of T cell activation, rely on bypass signaling pathways to achieve IL-2 secretion, thereby enhancing T cell proliferation and effects while reducing potential cell toxicity risks [252]. It was demonstrated in immunosuppressive pancreatic cancer mouse models with significantly prolonged survival. In addition, the upregulation of membrane-tethered IL-15/IL-21 co-expression in engineered T cells [253], the selective expression of homodimeric IL-21R replaced by CD34 extracellular structural domain [254], and the fusion protein of IL-2 with IL-2Rα with selective affinity [255], can all effectively reduce the exhaustion of therapeutic T cells and enhance their anti-tumor function in different preclinical murine solid tumor models.
Metabolic programming regulation of CD8+ T cell functionality
Antigen recognition activates the TCR-CD3 complex signaling pathway, driving intricate metabolic programs within T cells. The balance of energy and material metabolism within T cells largely determines the activation and maintenance of their antitumor activities. Therefore, understanding the processes of energy production and biosynthesis in T cells, as well as the impact of these processes on T cell differentiation and functional maturation, will be beneficial in identifying new effective targets to enhance antitumor immunotherapy. Subsequently, by reprogramming T cell metabolism to regulate these proven “targets”, the transcriptome of T cells can be shifted towards characteristics that favor antitumor immunity. This represents an effective strategy for modifying T cells. (Fig. 6).

Metabolic pathways enhancing the anti-tumor function of CD8+ T cells. Appropriate metabolic program modulation of quiescent or dysfunctional CD8+ T cells can promote T cells to exit quiescence or rejuvenate exhausted T cells. The reprogramming pathways mainly include mitochondrial biogenesis, glucose metabolism, amino acid metabolism, lipid and sterol metabolism related to nutrient synthesis, and ion metabolism related to the regulation of cellular life activities
Metabolism of mitochondria
The tricarboxylic acid cycle (TCA) that occurs within mitochondria serves as the hub of cellular energy metabolism and is the final pathway for the metabolism of sugars, fats, and amino acids [256]. In CD8+ T cells, the TCA cycle, driven by glucose and glutamine, serves as the principal metabolic pathway for sustaining mitochondrial energy metabolism and intratumoral migration [257]. Variations in the expression of key metabolic enzymes and the availability of intermediate metabolites throughout this process can influence the homeostasis of energy metabolism [258, 259]. Reduced equivalents (FADH/NADH/NADPH) derived from glycolysis, the TCA cycle, and the pentose phosphate pathway participate in oxidative phosphorylation in mitochondria to directly supply energy for almost all life activities of T cells [260]. The survival and proliferation of CD8+ T cells depend on oxidative phosphorylation [261,262,263], and controlling the occurrence and progression of reduced equivalent generation reactions to regulate oxidative phosphorylation is an effective strategy. Among them, mTORC1 is an important regulatory factor, which induces the expression of transcription factors, such as IRF4/MYC Proto-Oncogene (MYC), and changes the metabolic state of T cells through transcription, translation, and post-translational modification [264,265,266]. Another key metabolic checkpoint is the mitochondrial pyruvate carrier (MPC), which transports pyruvate in the cytoplasm to mitochondria to enhance oxidative phosphorylation [267]. Inhibition of MPC will inhibit aerobic oxidation in T cells so that CD8+ T cells cannot maintain complex life activities [268]. At the same time, T cell energy metabolism is also closely related to TCR signals and CD28 co-stimulatory signals [269,270,271,272]. Mechanistically, TCR signals and CD28 co-stimulatory signals mediate the fusion of mitochondrial inner and outer membranes, driving more efficient electron transport chains to enhance oxidative phosphorylation [273, 274].
Mitochondria are also important sites for material metabolism. When T cells are activated, the mitochondrial one-carbon metabolism pathway is enhanced [275]. Serine is the main amino acid involved in one-carbon metabolism [276], and is crucial in promoting T cell proliferation and activation [275, 277]. The smooth entry of serine into mitochondria is the key to this metabolic process, and Sideroflexin1 (SFXN1) has been proven to be a key protein transporting serine into mitochondria [278]. The methionine cycle provides methyl for various biochemical reactions. S-adenosylmethionine (SAM) and its downstream metabolite 5-methylthioadenosine (MTA) can induce the occurrence of CD8+ T cell exhaustion program. And inhibition of MAT2A, a key enzyme for SAM synthesis, can effectively improve CD8+ T cell function in mouses bearing hepatocellular carcinoma and increase their overall survival [279]. Ketone bodies (KBs) are produced by the β-oxidation of fatty acids in mitochondria and are essential fuels for maintaining CD8+ T cell effector function. Endogenous KBs oxidation (ketolysis) drives CD8+ T cell effector-like metabolism. Mechanistically, KBs are preferentially utilized to supply substrates for the TCA cycle, thereby directly enhancing the TCA-dependent metabolic pathways of CD8+ Teff cells [280].
Mitochondrial biogenesis maintains the energy and metabolic homeostasis within CD8+ T cells, and proliferator-activated receptor (PPAR) gamma coactivator 1α (PGC-1α)—a transcriptional activator that promotes mitochondrial and nuclear interaction [281]—is a key regulatory factor [282]. The inhibitory tumor microenvironment often causes the expression of PGC-1α in TILs to be suppressed, leading to T cell disability [283]. The application of overexpressing PGC-1α in CD8+ T cells through genetic engineering can effectively enhance its anti-tumor effect in mouses bearing lung carcinoma [284, 285]. In addition, 4-1BB co-stimulation can upregulate PGC1α expression, and activate ATF2 through the p38-MAPK pathway, thereby enhancing mitochondrial fusion and biogenesis, and ultimately promoting the immunity and memory of CD8+ T cells in mouses bearing melanoma [286].
Metabolism of glucose and amino acid
Glucose and amino acid metabolism are crucial for T cell activation [261, 287]. Activated T cells regulate glucose uptake from the TME through Glucose transporter 1 (GLUT1) [288]. Then, glucose undergoes glycolysis to generate pyruvate, which can be transported to mitochondria to participate in the TCA [287]. Adjusting the availability of metabolic products and key metabolic enzymes of these biochemical reactions can guide T cell metabolic reprogramming. Phosphoenolpyruvate maintains T cell receptor-mediated Ca2+-NFAT signal transduction and effector function by inhibiting sarco/endoplasmic reticulum Ca2+-ATPase (SERCA) activity [289]. Phosphoenolpyruvate carboxykinase 1 (PCK1) not only enhances the impact of phosphoenolpyruvate on T cell function but also regulates T cell differentiation [290]. Pyruvate, the end product of glycolysis, is oxidized to lactate by lactate-dehydrogenase (LDH), reducing TCA substrate supply while increasing acid load. Moreover, lactate is a double-edged sword. On the one hand, physiological concentrations of lactate inhibit histone deacetylase activity, and increase H3K27 acetylation, thereby upregulating Tcf7 expression, ultimately increasing CD8+ T cell stemness and delaying colon tumor growth in mouses [291]. On the other hand, the Warburg effect of tumor cells that cells prefer to metabolize glucose to lactate even in the presence of oxygen, leads to a high lactate load in the TME [292]. TIL interaction with lactate in the TME is mediated by monocarboxylate transporter 1 (MCT1), a bidirectional proton-assisted transport protein encoded by SLC16A1, the directionality of which is determined by the concentration of substrate and H+ [293]. High concentrations of lactate promote the reduction of NAD+ to NADH, causing changes in NAD+ dependent enzyme-catalyzed reactions [294], thereby reducing the production of metabolites needed for T cell proliferation, ultimately limiting CD8+ T cell effector function [295, 296]. It has been substantiated that the stem cell-like memory differentiation of adoptively transferred CD8+ T cells can be effectively augmented by the inhibition of glycolysis [297, 298], an effect that is intimately associated with the preservation of FOXO1 activity within these T cells.
Glutamine serves not only as the primary fuel for early CD8+ effector T cells but also as a crucial biosynthetic substrate. Glutamic-oxaloacetic transaminase 1 (Got1), which regulates de novo synthesis of aspartate from glutamine, is essential for sustaining the expansion and effector function of CD8+ effector T cells [299]. One of the essential amino acids, leucine, activates mTORC1 in a large amino acid transporter 1 (LAT1)/CD98-dependent manner to induce metabolic reprogramming, promoting CD8+ T cell activation [300]. Non-essential amino acids are usually synthesized from glucose or glutamine metabolic derivatives, participating in T cell metabolic activities [301]. The metabolic activity of amino acids is accompanied by the accumulation of metabolites. Kynurenine, a tryptophan metabolite, can be taken up by T cells through LAT1/CD98 regulation [302] and has been shown to inhibit T cell proliferation [303]. The enzyme co-factor tetrahydrobiopterin (BH4) can rescue the inhibition of kynurenine on T cells, upregulating the expression of GTP cyclohydrolase 1 (GCH1) and sepiapterin reductase-key enzymes in the BH4 synthesis process-significantly enhancing CD8+ T cell anti-tumor activity and controlling tumor growth in mouse bearing breast tumor [304]. L-arginine, a derivative of intracellular arginine metabolism, can enhance T cell adaptability, control tumor growth, and prolong overall survival in mouses bearing melanoma [305]. Mechanistically, L-arginine induce a shift in T cell respiration from glycolysis to oxidative phosphorylation. The key transcriptional regulators mediating this process are Bromodomain adjacent to zinc finger domain 1B (BAZ1B) and PC4 and SFRS1 interacting protein 1 (PSIP1). In CD8+ T cells, the absence of taurine, a cysteine metabolic derivative, increases endoplasmic reticulum stress and induces ATF4 transcription. This, in turn, activates immune checkpoint gene expression, leading to CD8+ T cell exhaustion [306].
Lipid and sterol metabolism
Glucose-derived pyruvate is synthesized into acetyl-CoA in mitochondria through the TCA cycle [307], and acetyl-CoA enters the mevalonate pathway to promote the synthesis of lipids and sterols [308]. When T cells encounter nutrient stress, AMP-activated protein kinase (AMPK), a serine/threonine kinase, is activated [262] then induces the biosynthesis of lipids and sterols in a mTORC1-dependent manner by upregulating sterol regulatory element-binding transcription factor (SREBP)1/2, maintaining the energy material supply for CD8+ T cells [309]. SREBP cleavage-activating protein (SCAP) is an activator of SREBP, which can promote the proliferation and functional activation of CD8+ T cells [310].
In solid tumors, rapidly proliferating tumor cells disproportionately compete with TILs for energy metabolites such as glucose and amino acids, leading to impaired T cell metabolic status and inability to maintain the energy synthesis required for survival and anti-tumor activity [17, 311]. At this time, fatty acid oxidation (FAO) becomes an effective way to produce energy, and it has been proven in mouses bearing melanoma that TILs rich in FAO are more effective in controlling cancer [312]. Lipids not only participate in energy synthesis but also are important regulators of T cell responses. Short-chain fatty acids (SCFAs) produced by gut microbiota metabolism, by inhibiting HDAC and regulating the mTOR-S6K pathway, induce T cell effector and memory differentiation [313], representing host-internal microbiota-induced metabolic reprogramming. Moreover, a diet-derived trans-vaccenic acid (TVA) inactivates the cell surface receptor G-protein-coupled receptor 43 (GPR43)-an immune regulatory G-protein-coupled receptor activated by its short-chain fatty acid ligand [314]-thereby activating the cAMP-PKA-CREB axis, enhancing CD8+ T cell cytotoxic effector function which suppress tumor growth in melanoma mouse models, and representing host-extrinsic reprogramming of CD8+ T cells [315]. When the glucose availability of CD8+ T cells is low, a short-chain saturated fatty acid called acetate participates in the synthesis of acetyl-CoA under the catalysis of acetyl-CoA synthetase, thereby enhancing metabolism to maintain the supply of acetyl-CoA, which is an important intermediate product of energy metabolism, ultimately promoting CD8+ T cell proliferation and effector function in preclinical breast cancer models [316]. Intracellular fatty acids can also regulate T cell activation and proliferation by binding to PPAR [317]. However, the abundance of long-chain fatty acids (LCFAs) in the TME will cause T cell dysfunction, and regulating the expression of VLC acyl-CoA dehydrogenase (ACADVL) can improve the metabolic adaptability of TILs [318]. It has been found in mouses bearing melanoma that leptin-a lipid factor in fat metabolism-binds to the upregulated leptin receptor on TILs, induces phospho-p38 MAPK activation, further activates transcription factor ATF2, ultimately increasing TIL oxidative activity and ability [319]. Another lipid mediator, prostaglandin E 2 (PGE2) is metabolized by the enzyme from the twenty-carbon unsaturated fatty acid arachidonic acid. And its signal transduction with E-type prostanoid receptor 2/4 (EP2/EP4) is the key to inhibiting CD8+ T cell response to IL-2 signals, leading to the inability to expand the number of stem cell-like CD8+ T cells, and even causing cell ferroptosis due to mitochondrial dysfunction. Blocking the PGE2-EP2/EP4 axis achieves immune control over various solid tumors in mice [320, 321].
Physiological levels of cholesterol are necessary to maintain normal T cell function. A naturally occurring sterol, 7-alpha-hydroxycholesterol (7α-HC), exerts a significant inhibitory effect on the TCR signaling pathway by enhancing the membrane binding of CD3ε, thereby hindering its aggregation with LCK and suppressing LCK-mediated TCR phosphorylation. A brief pre-treatment of engineered T cells with 7α-HC can induce a reduction in their in vivo signaling, leading to an increase in the number and proportion of memory cells, thereby enhancing their long-term anti-tumor capabilities in mouse models of melanoma [322]. The enrichment of oxysterols in the TME changes the expression ratio of SREBP2/liver X receptor (LXR) [323, 324], which is the key transcription factor for intracellular cholesterol metabolism, leading to cholesterol deficiency in T cells, thereby causing CD8+ T cell metabolic programming to be silenced and enhancing its autophagy, resulting in increased CD8+ T cell dysfunction and apoptosis [325]. High cholesterol levels upregulate the expression of immune checkpoints on T cells, such as PD-1, 2B4, in an ER-stress-X-Box Binding Protein 1-dependent manner, thereby limiting CD8+ Teff anti-tumor ability [326]. Cholesterol esters or cholesterol sulfate can comprehensively affect TCR signal transduction by changing TCR clustering or antigen affinity [327]. Therefore, regulating the types of fatty acids and cholesterol abundance in the TME is an effective way to regulate the anti-tumor ability of CD8+ T cells.
Inorganic ion metabolism
Minerals in the TME, often in the form of messenger molecules or co-factors of biological enzymes, participate in signal transduction and metabolic regulation within T cells, thereby participating in T cell fate. The proliferation, maturation, and activation of T cells strongly depend on the activity of calcium release-activated channels (CRAC) activated by stromal interaction molecule 1 (STIM1) and calcium release-activated calcium modulator 1 (ORAI1) [328]. When CTLs are activated by antigens, the endoplasmic reticulum Ca2+ store is rapidly depleted, and the secretion of vesicles containing toxic cytokines is calcium-dependent [329]. At this time, STIM1/ORAI1 dependent Ca2+ entry is crucial for maintaining the ability of CD8+ Teff to lyse tumor cells [330]. In addition, Store-operated Ca2+ entry guides CD8+ T cell metabolic reprogramming by activating NFAT and the PI3K-Akt kinase-mTOR nutrient-sensing pathway, supporting its anti-tumor response [331]. Mg+ can assist Ca2+ signal conduction. When T cells receive antigen stimulation, the expression of the magnesium transporter gene 1 (MAGT1) is upregulated, promoting Mg+ influx. The absence of MAGT1 causes a significant impairment of Ca2+ influx in T cells, leading to impaired response to antigen receptor binding [332]. Necrotic cells in tumors dissolve and release a large amount of K+, causing K+ accumulation in the TME. On the one hand, T cells perceive high extracellular K+ concentration and experience an obstacle to Akt-mTOR phosphorylation and effector programmes driven by TCR. Regulating the potassium channel Kv1.3 can improve the T cell dysfunction caused by this [333]. On the other hand, high extracellular K+ levels also inhibit T cell effector programs by limiting the uptake of nutrients, thereby inducing T cell autophagy, reducing the expression of exhaustion programs, and inducing CD8+ T cell stemness transformation and effector function inhibition [334]. Na+/K+ ATPase plays a key role in maintaining intracellular and extracellular K+ concentration. When the intracellular K+ level rises, it can prevent the destruction of reactive oxygen species (ROS), limit the exhausted state of T cells, and maintain their anti-tumor function, delay ultimately melanoma growth in mice [335]. Iron, as an important redox ion, participates in biochemical reactions within cells. When the activity of glutathione peroxidase 4 (GPX4) in cells decreases, lipid oxidation products cannot be metabolized by the glutathione reductase reaction catalyzed by GPX4, leading to a large accumulation of Fe2+ dependent lethal lipid ROS, eventually causing cell Ferroptosis [336]. It has been found that TILs are very sensitive to iron death, and GPX4 and ferroptosis suppressor protein 1 (FSP1) can enhance the iron death resistance of CD8+ TIL. In addition, a seizure risk-related protein DEP domain-containing protein5 (DEPDC5) protects CD8+ T cells from iron death and is necessary for CD8+ T cell-mediated anti-tumor immunity. Mechanistically, DEPDC5 defects cause an increase in mTORC1 activity in CD8+ T cells, thereby inducing ATF4 expression, and then increasing the expression of xanthine oxidase (XO) and lipid ROS, ultimately leading to spontaneous iron death [337]. Lithium salts, used clinically as mood stabilizers, have been found to have a role in regulating T cell metabolism [338]. Lithium carbonate has been confirmed to disrupt vacuolar ATPase, thereby halting lysosomal acidification and salvaging lysosomal diacylglycerol-PKCθ signaling, consequently promoting the localization of MCT1 to the mitochondrial inner membrane. This process further facilitates the entry of exogenous lactate into the mitochondria for oxidation, thereby improving the efficacy of T cell therapy for solid tumors in mouse [339].
Conclusion and perspectives
With the advancements in tumor neoantigen identification, reactive T cell receptor (TCR) screening technologies, and gene engineering, alongside the profound exploration of structural immunology regarding the structure-function relationship of engineered T cells, TCR-engineered T cells have propelled the immunotherapy of refractory solid tumors into a new phase with superior specificity and stable functional activity. Concurrently, the recruitment of tumor antigen-specific soluble bispecific molecules facilitates and promotes the redirection of pre-existing polyclonal CD8+ T cells towards tumor cells, offering a simpler alternative for T cell therapy against solid tumors. However, exhaustion remains an intrinsic aspect of the anti-tumor fate of CD8+ T cells, rendering the mitigation or reversal of CD8+ T cell exhaustion imperative for ensuring sustained efficacy of anti-tumor immune therapy. Manipulating immune checkpoints enhances the adaptability of CD8+ T cells within tumors, while diverse cytokine engineering provides novel avenues for restoring CD8+ T cell activity. Various innovative approaches have been explored in CAR-T cell therapy for cancer. For instance, overexpression of BCL-2 inhibits intrinsic apoptosis of T cells [340], overexpression of FOXO1 regulates memory-like differentiation of T cells [341], preconditioning with appropriate temperature improves intratumoral movement of T cells [257], and drug-delivery biodegradable scaffolds that mimic the physiological microenvironment of T cell activation sustain prolonged T cell stimulation [342]. The success of these cases will provide practical evidence for TCR-engineered T cell therapy in solid tumors.
In summary, cell therapy based on highly antigen-sensitive, microenvironment-adaptive, and functionally complete T cells may currently be the most promising approach for the immunotherapy of solid tumors. In the future, the development of T cell therapy should focus on optimizing the design of engineered T cells and improving the efficiency of their industrial-scale production. By combinatorially engineering individual T cells to stably express tumor-specific high-affinity TCRs, obtain efficient co-stimulatory signals, and tolerate immune negative regulation, these cells will participate in the treatment of solid tumors with high environmental adaptability and low cross-reactivity. To this end, the application of systems biology approaches in transcriptomics, epigenomics, chromatin accessibility, and metabolomics, especially at the single-cell level, will continue to provide new insights for T cell therapy of solid tumors.
Data availability
Not applicable.
Abbreviations
- ACT:
-
Adoptive cell transfer
- ACADVL:
-
VLC Acyl-CoA dehydrogenase
- ADAM17:
-
A disintegrin and metalloprotease 17
- AMPK:
-
AMP-activated protein kinase
- AP-1:
-
Activator protein 1
- ATF:
-
Activating transcription factor
- BAF:
-
BRG1 or BRM-associated factor
- BATF:
-
Basic leucine zipper ATF-like transcription factor
- BAZ1B:
-
Bromodomain adjacent to zinc finger domain, 1B
- BCL10:
-
B cell lymphoma/leukemia 10
- BH4:
-
Tetrahydrobiopterin
- BiTE:
-
Bispecific T cell engager
- BTLA:
-
B and T lymphocyte attenuator
- cAMP:
-
Cyclic adenosine monophosphate
- CAR:
-
Chimeric Antigen Receptor
- CARD11:
-
Caspase recruitment domain-containing protein 11
- CARMA1:
-
Caspase recruitment domain-containing membrane-associated guanylate kinase protein 1
- CBL:
-
Cbl proto-oncogene, E3 ubiquitin protein ligase
- CBLB:
-
Casitas B-lineage lymphoma proto-oncogene B
- CD:
-
Cluster of differentiation
- CDR:
-
Complementary determining region
- CEA:
-
Carcinoembryonic antigen
- CEACAM1:
-
Carcinoembryonic antigen-related cell adhesion molecule-1
- CLDN:
-
Claudin
- Co-STAR:
-
Co-stimulatory-STAR
- CRAC:
-
Calcium release-activated channels
- CREB:
-
cAMP- response element binding protein
- CRISPR- Cas9:
-
Clustered regularly interspaced short palindromic repeats- associated protein 9
- CTL:
-
Cytotoxic T lymphocytes
- CTLA4:
-
Cytotoxic T lymphocyte antigen 4
- CYRIB:
-
CYFIP related Rac1 interactor B
- DAG:
-
Diacylglycerol
- DEPDC5:
-
DEP domain-containing protein5
- DLL3:
-
Delta-like ligand 3
- DSF:
-
Disulfiram
- EpCAM:
-
Epithelial cell adhesion molecule
- ERK:
-
Extracellular signal-related kinase
- FAM49B:
-
The family with sequence similarity 49 member B
- FAO:
-
Fatty acid oxidation
- FDA:
-
Food and Drug Administration
- Fgl2:
-
Fibrinogen-like protein 2
- FOXO1:
-
Forkhead box O1
- FSP1:
-
Ferroptosis suppressor protein 1
- GTP:
-
Guanosine triphosphate
- GCH1:
-
GTP cyclohydrolase1
- GLUT1:
-
Glucose transporter 1
- GOF:
-
Gain-of-function
- Got1:
-
Glutamic-oxaloacetic transaminase 1
- GPC3:
-
Glypican-3
- GPR43:
-
G-protein-coupled receptor 43
- GPX4:
-
Glutathione peroxidase 4
- GRB2:
-
Growth factor receptor-bound protein 2
- H3K27ac:
-
Histone H3 Lysine 27 acetylation
- HCC:
-
Hepatocellular carcinoma
- HDAC3:
-
Histone deacetylase 3
- HDR:
-
Homology-directed repair
- HIT:
-
HLA-independent TCR
- HLA:
-
Human leukocyte antigen
- HVEM:
-
Herpesvirus entry mediator
- ICB:
-
Immune checkpoint blockade
- Id2:
-
Inhibitor of DNA binding protein 2
- IFN:
-
Interferon
- iICPs:
-
Inhibitory immune checkpoints
- IKK:
-
IĸB kinase
- IL:
-
Interleukin
- ILRs:
-
IL receptors
- ImmTAC:
-
Immune-mobilizing monoclonal TCR against cancer
- INO80:
-
Inositol requiring mutant 80
- IP3:
-
Inositol triphosphate
- IRF4:
-
Interferon regulatory factor 4
- IRs:
-
Inhibitory receptors
- ITAMs:
-
Immunoreceptor tyrosine-based activation motifs
- ITIM:
-
Immunoreceptor tyrosine-based inhibitory motif
- ITPRIPL1:
-
I-type single-pass membrane protein
- ITSM:
-
Immunoreceptor tyrosine-based activation motif
- JAK:
-
Janus kinase
- KBs:
-
Ketone bodies
- LAG3:
-
Lymphocyte activation gene-3
- LAGE-1A:
-
L antigen family member-1 A
- LAT:
-
linker for activation of T cells
- LAT1:
-
Large amino acid transporter 1
- LCFAs:
-
Long-chain fatty acids
- LCMV:
-
Lymphocytic choriomeningitis virus
- LCP:
-
Lymphocyte cytosolic protein
- LDH:
-
Lactate dehydrogenase
- LNPs:
-
Lipid nanoparticles
- LSD1:
-
Lysine demethylase 1
- LXR:
-
Liver X receptor
- LCK:
-
LCK proto-oncogene, Src family tyrosine kinase
- MAGE-A4:
-
Melanoma-associated antigen A4
- MAGT1:
-
Magnesium transporter gene 1
- MALT1:
-
Mucosa-associated lymphoid tissue translocation protein 1
- MAPK:
-
Mitogen-activated protein kinase
- mCRPC:
-
Metastatic castration-resistant prostate cancer
- MCT1:
-
Monocarboxylate transporter 1
- MDSCs:
-
Myeloid-derived suppressor cell populations
- MEGA:
-
Multiplexed effector guide arrays
- MHC:
-
Major histocompatibility complex
- ModPoKI:
-
Modular pooled knock-in screening
- MPC:
-
Mitochondrial pyruvate carrier
- mRNA:
-
Messenger ribonucleic acid
- mSWI/SNF:
-
Mammalians switch/sucrose non-fermentable
- MTA:
-
Methylthioadenosine
- mTOR:
-
Mammalian target of the rapamycin
- mTORC:
-
mTOR complex
- MYC:
-
MYC Proto-Oncogene
- NeoTCR:
-
Neoantigen-specific TCR
- NFAT:
-
Nuclear factor of activated T cells
- NF-κB:
-
Nuclear factor-ĸB
- NGS:
-
Next-generation sequencing
- NY-ESO-1:
-
New York esophageal squamous cell carcinoma 1
- ORAI1:
-
Calcium release-activated calcium modulator 1
- Osr2:
-
Odd-skipped related 2
- PCK1:
-
Phosphoenolpyruvate carboxykinase 1
- PD-1:
-
Programmed death protein-1
- PDAC:
-
Pancreatic ductal adenocarcinoma
- PDK1:
-
Phosphoinositide-dependent kinase 1
- PGC-1α:
-
PPAR gamma coactivator 1α
- PGE2:
-
Prostaglandin E 2
- PIK3CD:
-
Phosphatidylinositol-4,5-bisphosphate 3-kinase catalytic subunit delta
- PIK3R3:
-
Phosphoinositide-3-kinase regulatory subunit 3
- PKC:
-
Protein kinase C
- p-MHC:
-
Peptide- major histocompatibility complex
- PPAR:
-
Proliferator-activated receptor
- PRAME:
-
Preferentially expressed antigen in melanoma
- PRODH2:
-
Proline dehydrogenase 2
- PSGL-1:
-
P-selectin glycoprotein-1
- PSIP1:
-
PC4 and SFRS1 interacting protein 1
- PSMA:
-
Prostate-specific membrane antigen
- PTKs:
-
Protein tyrosine kinases
- PTPN:
-
Protein tyrosine phosphatases
- RASA2:
-
RAS P21 protein activator 2
- RNP:
-
Ribonucleoprotein
- ROS:
-
Reactive oxygen species
- RC3H1:
-
Ring finger and CCCH-type domains 1
- SAM:
-
S-adenosylmethionine
- SCAP:
-
SREBP cleavage-activating protein
- SCFAs:
-
Short-chain fatty acids
- ScFv:
-
Single-chain variable fragment
- SCLC:
-
Small-cell lung cancer
- SERCA:
-
Sarco/endoplasmic reticulum Ca2+-ATPase
- SFXN1:
-
Sideroflexin1
- sgRNA:
-
Single guide RNA
- SHP:
-
Src homology region 2 domain-containing phosphatase
- SLICE:
-
sgRNA lentiviral infection with Cas9 protein electroporation
- SMAD4:
-
SMAD family member 4
- SNVs:
-
Single-nucleotide variants
- SOCS1:
-
Suppressor of cytokine signaling 1
- SREBP:
-
Sterol regulatory element-binding transcription factor
- SRs:
-
Synthetic surface receptors
- ssDNA:
-
Single-stranded DNA
- STAR:
-
Synthetic TCR antigen receptor
- STAT:
-
Signal transducer and activator of transcription
- STE-20:
-
Serine/threonine-protein kinase 20
- STIM1:
-
Stromal Interaction Molecule 1
- STS-1:
-
Suppressor of T cell signaling-1
- SynNotch:
-
Synthetic Notch
- TAOK3:
-
Thousand-and- one–amino acid kinase 3
- TCA:
-
Tricarboxylic acid cycle
- TCF:
-
T-cell factor
- Cα/Cβ:
-
Constant domain α/β
- TCR:
-
T cell receptor
- TCRm:
-
TCRmimic
- Teff :
-
Effector T cell
- Tex :
-
Exhausted T cell
- TF:
-
Transcription factor
- THEMIS:
-
T-lineage protein thymocyte-expressed molecule involved in selection
- TALENs:
-
Transcription activator-like effector endonucleases
- TIGIT:
-
T cell immunoglobulin and ITIM domain
- TILs:
-
Tumor infiltrating lymphocytes
- TMB:
-
Tumor neoantigen mutation burden
- TMD:
-
Transmembrane domain
- TME:
-
Tumor microenvironment
- TOX:
-
Thymocyte selection-associated high mobility group box protein
- TRAC:
-
TCR constant domainα
- TRBC:
-
TCR constant domainβ
- TSC:
-
Tuberous sclerosis complex
- TVA:
-
Trans-vaccenic acid
- UTRs:
-
Untranslated regions
- VISTA:
-
V domain immunoglobulin suppressor of T cell activation
- XO:
-
Xanthine oxidase
- ZAP:
-
ζ-chain-associated protein
- ZFNs:
-
Zinc-finger nucleases
- ZC3H12A:
-
Zinc finger CCCH-type containing 12 A
- 7α-HC:
-
7-α-hydroxycholesterol
References
Sahin U, Derhovanessian E, Miller M, Kloke BP, Simon P, Löwer M, et al. Personalized RNA mutanome vaccines mobilize poly-specific therapeutic immunity against cancer. Nature. 2017;547(7662):222–6.
Schumacher TN, Schreiber RD. Neoantigens in cancer immunotherapy. Sci (New York NY). 2015;348(6230):69–74.
Stadtmauer EA, Fraietta JA, Davis MM, Cohen AD, Weber KL, Lancaster E et al. CRISPR-engineered T cells in patients with refractory cancer. Sci (New York NY). 2020;367(6481).
Siddiqui I, Schaeuble K, Chennupati V, Fuertes Marraco SA, Calderon-Copete S, Pais Ferreira D, et al. Intratumoral Tcf1(+)PD-1(+)CD8(+) T cells with stem-like Properties promote Tumor Control in Response to Vaccination and Checkpoint Blockade Immunotherapy. Immunity. 2019;50(1):195–e21110.
Rosenberg SA, Restifo NP. Adoptive cell transfer as personalized immunotherapy for human cancer. Sci (New York NY). 2015;348(6230):62–8.
Raskov H, Orhan A, Christensen JP, Gögenur I. Cytotoxic CD8(+) T cells in cancer and cancer immunotherapy. Br J Cancer. 2021;124(2):359–67.
Opferman JT, Ober BT, Ashton-Rickardt PG. Linear differentiation of cytotoxic effectors into memory T lymphocytes. Sci (New York NY). 1999;283(5408):1745–8.
McLane LM, Abdel-Hakeem MS, Wherry EJ. CD8 T cell exhaustion during chronic viral infection and Cancer. Annu Rev Immunol. 2019;37:457–95.
Moskophidis D, Lechner F, Pircher H, Zinkernagel RM. Virus persistence in acutely infected immunocompetent mice by exhaustion of antiviral cytotoxic effector T cells. Nature. 1993;362(6422):758–61.
Doering TA, Crawford A, Angelosanto JM, Paley MA, Ziegler CG, Wherry EJ. Network analysis reveals centrally connected genes and pathways involved in CD8 + T cell exhaustion versus memory. Immunity. 2012;37(6):1130–44.
Schietinger A, Greenberg PD. Tolerance and exhaustion: defining mechanisms of T cell dysfunction. Trends Immunol. 2014;35(2):51–60.
McGranahan N, Rosenthal R, Hiley CT, Rowan AJ, Watkins TBK, Wilson GA, et al. Allele-specific HLA loss and Immune escape in Lung Cancer Evolution. Cell. 2017;171(6):1259–e7111.
Rooney MS, Shukla SA, Wu CJ, Getz G, Hacohen N. Molecular and genetic properties of tumors associated with local immune cytolytic activity. Cell. 2015;160(1–2):48–61.
Yamamoto K, Venida A, Yano J, Biancur DE, Kakiuchi M, Gupta S, et al. Autophagy promotes immune evasion of pancreatic cancer by degrading MHC-I. Nature. 2020;581(7806):100–5.
Binnewies M, Roberts EW, Kersten K, Chan V, Fearon DF, Merad M, et al. Understanding the tumor immune microenvironment (TIME) for effective therapy. Nat Med. 2018;24(5):541–50.
Hanahan D, Weinberg RA. Hallmarks of cancer: the next generation. Cell. 2011;144(5):646–74.
Chang CH, Qiu J, O’Sullivan D, Buck MD, Noguchi T, Curtis JD, et al. Metabolic competition in the Tumor Microenvironment is a driver of Cancer Progression. Cell. 2015;162(6):1229–41.
Siska PJ, Rathmell JC. T cell metabolic fitness in antitumor immunity. Trends Immunol. 2015;36(4):257–64.
Guedan S, Ruella M, June CH. Emerging Cellular therapies for Cancer. Annu Rev Immunol. 2019;37:145–71.
Paijens ST, Vledder A, de Bruyn M, Nijman HW. Tumor-infiltrating lymphocytes in the immunotherapy era. Cell Mol Immunol. 2021;18(4):842–59.
Olson DJ, Odunsi K. Adoptive cell therapy for nonhematologic solid tumors. J Clin Oncology: Official J Am Soc Clin Oncol. 2023;41(18):3397–407.
Morgan RA, Dudley ME, Wunderlich JR, Hughes MS, Yang JC, Sherry RM, et al. Cancer regression in patients after transfer of genetically engineered lymphocytes. Sci (New York NY). 2006;314(5796):126–9.
Hong M, Clubb JD, Chen YY. Engineering CAR-T cells for Next-Generation Cancer Therapy. Cancer Cell. 2020;38(4):473–88.
Flugel CL, Majzner RG, Krenciute G, Dotti G, Riddell SR, Wagner DL, et al. Overcoming on-target, off-tumour toxicity of CAR T cell therapy for solid tumours. Nat Reviews Clin Oncol. 2023;20(1):49–62.
Thistlethwaite FC, Gilham DE, Guest RD, Rothwell DG, Pillai M, Burt DJ, et al. The clinical efficacy of first-generation carcinoembryonic antigen (CEACAM5)-specific CAR T cells is limited by poor persistence and transient pre-conditioning-dependent respiratory toxicity. Cancer Immunol Immunotherapy: CII. 2017;66(11):1425–36.
Holler PD, Holman PO, Shusta EV, O’Herrin S, Wittrup KD, Kranz DM. In vitro evolution of a T cell receptor with high affinity for peptide/MHC. Proc Natl Acad Sci USA. 2000;97(10):5387–92.
Kim SP, Vale NR, Zacharakis N, Krishna S, Yu Z, Gasmi B, et al. Adoptive Cellular therapy with autologous tumor-infiltrating lymphocytes and T-cell receptor-Engineered T cells targeting common p53 neoantigens in human solid tumors. Cancer Immunol Res. 2022;10(8):932–46.
Leidner R, Sanjuan Silva N, Huang H, Sprott D, Zheng C, Shih YP, et al. Neoantigen T-Cell receptor gene therapy in pancreatic Cancer. N Engl J Med. 2022;386(22):2112–9.
Foy SP, Jacoby K, Bota DA, Hunter T, Pan Z, Stawiski E, et al. Non-viral precision T cell receptor replacement for personalized cell therapy. Nature. 2023;615(7953):687–96.
Yang X, Nishimiya D, Löchte S, Jude KM, Borowska M, Savvides CS, et al. Facile repurposing of peptide-MHC-restricted antibodies for cancer immunotherapy. Nat Biotechnol. 2023;41(7):932–43.
Poole A, Karuppiah V, Hartt A, Haidar JN, Moureau S, Dobrzycki T, et al. Therapeutic high affinity T cell receptor targeting a KRAS(G12D) cancer neoantigen. Nat Commun. 2022;13(1):5333.
Goebeler ME, Bargou RC. T cell-engaging therapies - BiTEs and beyond. Nat Reviews Clin Oncol. 2020;17(7):418–34.
Lavin Y, Kobayashi S, Leader A, Amir ED, Elefant N, Bigenwald C, et al. Innate Immune Landscape in Early Lung Adenocarcinoma by Paired single-cell analyses. Cell. 2017;169(4):750–e6517.
Chevrier S, Levine JH, Zanotelli VRT, Silina K, Schulz D, Bacac M, et al. Immune Atlas Clear Cell Ren Cell Carcinoma Cell. 2017;169(4):736–e4918.
Simoni Y, Becht E, Fehlings M, Loh CY, Koo SL, Teng KWW, et al. Bystander CD8(+) T cells are abundant and phenotypically distinct in human tumour infiltrates. Nature. 2018;557(7706):575–9.
Scheper W, Kelderman S, Fanchi LF, Linnemann C, Bendle G, de Rooij MAJ, et al. Low and variable tumor reactivity of the intratumoral TCR repertoire in human cancers. Nat Med. 2019;25(1):89–94.
Sharma P, Hu-Lieskovan S, Wargo JA, Ribas A. Primary, adaptive, and Acquired Resistance to Cancer Immunotherapy. Cell. 2017;168(4):707–23.
Oliveira G, Stromhaug K, Klaeger S, Kula T, Frederick DT, Le PM, et al. Phenotype, specificity and avidity of antitumour CD8(+) T cells in melanoma. Nature. 2021;596(7870):119–25.
George JT, Kessler DA, Levine H. Effects of thymic selection on T cell recognition of foreign and tumor antigenic peptides. Proc Natl Acad Sci USA. 2017;114(38):E7875–81.
Bobisse S, Genolet R, Roberti A, Tanyi JL, Racle J, Stevenson BJ, et al. Sensitive and frequent identification of high avidity neo-epitope specific CD8 (+) T cells in immunotherapy-naive ovarian cancer. Nat Commun. 2018;9(1):1092.
Finnigan JP, Newman JH, Patskovsky Y, Patskovska L, Ishizuka AS, Lynn GM et al. Structural basis for self-discrimination by Neoantigen-Specific TCRs. Res Square. 2023.
Wu D, Gallagher DT, Gowthaman R, Pierce BG, Mariuzza RA. Structural basis for oligoclonal T cell recognition of a shared p53 cancer neoantigen. Nat Commun. 2020;11(1):2908.
Lee MN, Meyerson M. Antigen identification for HLA class I- and HLA class II-restricted T cell receptors using cytokine-capturing antigen-presenting cells. Science immunology. 2021;6(55).
Fujiwara K, Shao Y, Niu N, Zhang T, Herbst B, Henderson M, et al. Direct identification of HLA class I and class II-restricted T cell epitopes in pancreatic cancer tissues by mass spectrometry. J Hematol Oncol. 2022;15(1):154.
Spindler MJ, Nelson AL, Wagner EK, Oppermans N, Bridgeman JS, Heather JM, et al. Massively parallel interrogation and mining of natively paired human TCRαβ repertoires. Nat Biotechnol. 2020;38(5):609–19.
Fahad AS, Chung CY, López Acevedo SN, Boyle N, Madan B, Gutiérrez-González MF, et al. Cell activation-based screening of natively paired human T cell receptor repertoires. Sci Rep. 2023;13(1):8011.
Vazquez-Lombardi R, Jung JS, Schlatter FS, Mei A, Mantuano NR, Bieberich F et al. High-throughput T cell receptor engineering by functional screening identifies candidates with enhanced potency and specificity. Immunity. 2022;55(10):1953-66.e10.
Moravec Z, Zhao Y, Voogd R, Cook DR, Kinrot S, Capra B et al. Discovery of tumor-reactive T cell receptors by massively parallel library synthesis and screening. Nat Biotechnol. 2024.
Genolet R, Bobisse S, Chiffelle J, Arnaud M, Petremand R, Queiroz L, et al. TCR sequencing and cloning methods for repertoire analysis and isolation of tumor-reactive TCRs. Cell Rep Methods. 2023;3(4):100459.
Cetin M, Pinamonti V, Schmid T, Boschert T, Mellado Fuentes A, Kromer K, et al. T-FINDER: a highly sensitive, pan-HLA platform for functional T cell receptor and ligand discovery. Sci Adv. 2024;10(5):eadk3060.
Samstein RM, Lee CH, Shoushtari AN, Hellmann MD, Shen R, Janjigian YY, et al. Tumor mutational load predicts survival after immunotherapy across multiple cancer types. Nat Genet. 2019;51(2):202–6.
Jardim DL, Goodman A, de Melo Gagliato D, Kurzrock R. The challenges of Tumor Mutational Burden as an Immunotherapy Biomarker. Cancer Cell. 2021;39(2):154–73.
Clay TM, Custer MC, Sachs J, Hwu P, Rosenberg SA, Nishimura MI. Efficient transfer of a tumor antigen-reactive TCR to human peripheral blood lymphocytes confers anti-tumor reactivity. Journal of immunology (Baltimore, Md: 1950). 1999;163(1):507 – 13.
Dembić Z, Haas W, Weiss S, McCubrey J, Kiefer H, von Boehmer H, et al. Transfer of specificity by murine alpha and beta T-cell receptor genes. Nature. 1986;320(6059):232–8.
Dong D, Zheng L, Lin J, Zhang B, Zhu Y, Li N, et al. Structural basis of assembly of the human T cell receptor-CD3 complex. Nature. 2019;573(7775):546–52.
Viola A, Lanzavecchia A. T cell activation determined by T cell receptor number and tunable thresholds. Sci (New York NY). 1996;273(5271):104–6.
Thomas S, Xue SA, Cesco-Gaspere M, San José E, Hart DP, Wong V et al. Targeting the Wilms tumor antigen 1 by TCR gene transfer: TCR variants improve tetramer binding but not the function of gene modified human T cells. Journal of immunology (Baltimore, Md: 1950). 2007;179(9):5803-10.
van Loenen MM, de Boer R, Amir AL, Hagedoorn RS, Volbeda GL, Willemze R, et al. Mixed T cell receptor dimers harbor potentially harmful neoreactivity. Proc Natl Acad Sci USA. 2010;107(24):10972–7.
Cohen CJ, Zhao Y, Zheng Z, Rosenberg SA, Morgan RA. Enhanced antitumor activity of murine-human hybrid T-cell receptor (TCR) in human lymphocytes is associated with improved pairing and TCR/CD3 stability. Cancer Res. 2006;66(17):8878–86.
Sommermeyer D, Uckert W. Minimal amino acid exchange in human TCR constant regions fosters improved function of TCR gene-modified T cells. J Immunol (Baltimore Md: 1950). 2010;184(11):6223–31.
Davis JL, Theoret MR, Zheng Z, Lamers CH, Rosenberg SA, Morgan RA. Development of human anti-murine T-cell receptor antibodies in both responding and nonresponding patients enrolled in TCR gene therapy trials. Clin cancer Research: Official J Am Association Cancer Res. 2010;16(23):5852–61.
Bethune MT, Gee MH, Bunse M, Lee MS, Gschweng EH, Pagadala MS et al. Domain-swapped T cell receptors improve the safety of TCR gene therapy. eLife. 2016;5.
Chung S, Wucherpfennig KW, Friedman SM, Hafler DA, Strominger JL. Functional three-domain single-chain T-cell receptors. Proc Natl Acad Sci USA. 1994;91(26):12654–8.
Voss RH, Thomas S, Pfirschke C, Hauptrock B, Klobuch S, Kuball J, et al. Coexpression of the T-cell receptor constant alpha domain triggers tumor reactivity of single-chain TCR-transduced human T cells. Blood. 2010;115(25):5154–63.
Degirmencay A, Thomas S, Holler A, Stauss HJ. Exploitation of CD3ζ to enhance TCR expression levels and antigen-specific T cell function. bioRxiv: the preprint server for biology. 2024.
Govers C, Sebestyén Z, Roszik J, van Brakel M, Berrevoets C, Szöőr Á, et al. TCRs genetically linked to CD28 and CD3ε do not mispair with endogenous TCR chains and mediate enhanced T cell persistence and anti-melanoma activity. J Immunol (Baltimore Md: 1950). 2014;193(10):5315–26.
Thomas S, Mohammed F, Reijmers RM, Woolston A, Stauss T, Kennedy A, et al. Framework engineering to produce dominant T cell receptors with enhanced antigen-specific function. Nat Commun. 2019;10(1):4451.
Cohen CJ, Li YF, El-Gamil M, Robbins PF, Rosenberg SA, Morgan RA. Enhanced antitumor activity of T cells engineered to express T-cell receptors with a second disulfide bond. Cancer Res. 2007;67(8):3898–903.
Kuball J, Hauptrock B, Malina V, Antunes E, Voss RH, Wolfl M, et al. Increasing functional avidity of TCR-redirected T cells by removing defined N-glycosylation sites in the TCR constant domain. J Exp Med. 2009;206(2):463–75.
Haga-Friedman A, Horovitz-Fried M, Cohen CJ. Incorporation of transmembrane hydrophobic mutations in the TCR enhance its surface expression and T cell functional avidity. J Immunol (Baltimore Md: 1950). 2012;188(11):5538–46.
Voss RH, Willemsen RA, Kuball J, Grabowski M, Engel R, Intan RS, et al. Molecular design of the calphabeta interface favors specific pairing of introduced TCRalphabeta in human T cells. J Immunol (Baltimore Md: 1950). 2008;180(1):391–401.
Scholten KB, Kramer D, Kueter EW, Graf M, Schoedl T, Meijer CJ, et al. Codon modification of T cell receptors allows enhanced functional expression in transgenic human T cells. Clin Immunol (Orlando Fla). 2006;119(2):135–45.
Schober K, Müller TR, Gökmen F, Grassmann S, Effenberger M, Poltorak M, et al. Orthotopic replacement of T-cell receptor α- and β-chains with preservation of near-physiological T-cell function. Nat Biomedical Eng. 2019;3(12):974–84.
Provasi E, Genovese P, Lombardo A, Magnani Z, Liu PQ, Reik A, et al. Editing T cell specificity towards leukemia by zinc finger nucleases and lentiviral gene transfer. Nat Med. 2012;18(5):807–15.
Berdien B, Mock U, Atanackovic D, Fehse B. TALEN-mediated editing of endogenous T-cell receptors facilitates efficient reprogramming of T lymphocytes by lentiviral gene transfer. Gene Ther. 2014;21(6):539–48.
Mali P, Esvelt KM, Church GM. Cas9 as a versatile tool for engineering biology. Nat Methods. 2013;10(10):957–63.
Legut M, Dolton G, Mian AA, Ottmann OG, Sewell AK. CRISPR-mediated TCR replacement generates superior anticancer transgenic T cells. Blood. 2018;131(3):311–22.
Nahmad AD, Reuveni E, Goldschmidt E, Tenne T, Liberman M, Horovitz-Fried M, et al. Frequent aneuploidy in primary human T cells after CRISPR-Cas9 cleavage. Nat Biotechnol. 2022;40(12):1807–13.
Komor AC, Kim YB, Packer MS, Zuris JA, Liu DR. Programmable editing of a target base in genomic DNA without double-stranded DNA cleavage. Nature. 2016;533(7603):420–4.
Webber BR, Lonetree CL, Kluesner MG, Johnson MJ, Pomeroy EJ, Diers MD, et al. Highly efficient multiplex human T cell engineering without double-strand breaks using Cas9 base editors. Nat Commun. 2019;10(1):5222.
Preece R, Pavesi A, Gkazi SA, Stegmann KA, Georgiadis C, Tan ZM, et al. CRISPR-Mediated Base Conversion allows discriminatory depletion of endogenous T cell receptors for enhanced synthetic immunity. Mol Therapy Methods Clin Dev. 2020;19:149–61.
Ruggiero E, Carnevale E, Prodeus A, Magnani ZI, Camisa B, Merelli I, et al. CRISPR-based gene disruption and integration of high-avidity, WT1-specific T cell receptors improve antitumor T cell function. Sci Transl Med. 2022;14(631):eabg8027.
Roth TL, Puig-Saus C, Yu R, Shifrut E, Carnevale J, Li PJ, et al. Reprogramming human T cell function and specificity with non-viral genome targeting. Nature. 2018;559(7714):405–9.
Shy BR, Vykunta VS, Ha A, Talbot A, Roth TL, Nguyen DN, et al. High-yield genome engineering in primary cells using a hybrid ssDNA repair template and small-molecule cocktails. Nat Biotechnol. 2023;41(4):521–31.
Oh SA, Senger K, Madireddi S, Akhmetzyanova I, Ishizuka IE, Tarighat S et al. High-efficiency nonviral CRISPR/Cas9-mediated gene editing of human T cells using plasmid donor DNA. J Exp Med. 2022;219(5).
Nguyen DN, Roth TL, Li PJ, Chen PA, Apathy R, Mamedov MR, et al. Polymer-stabilized Cas9 nanoparticles and modified repair templates increase genome editing efficiency. Nat Biotechnol. 2020;38(1):44–9.
Allard M, Couturaud B, Carretero-Iglesia L, Duong MN, Schmidt J, Monnot GC et al. TCR-ligand dissociation rate is a robust and stable biomarker of CD8 + T cell potency. JCI Insight. 2017;2(14).
Sim MJW, Lu J, Spencer M, Hopkins F, Tran E, Rosenberg SA, et al. High-affinity oligoclonal TCRs define effective adoptive T cell therapy targeting mutant KRAS-G12D. Proc Natl Acad Sci USA. 2020;117(23):12826–35.
Schmidt J, Chiffelle J, Perez MAS, Magnin M, Bobisse S, Arnaud M, et al. Neoantigen-specific CD8 T cells with high structural avidity preferentially reside in and eliminate tumors. Nat Commun. 2023;14(1):3188.
Hebeisen M, Schmidt J, Guillaume P, Baumgaertner P, Speiser DE, Luescher I, et al. Identification of Rare High-Avidity, Tumor-reactive CD8 + T cells by Monomeric TCR-Ligand off-Rates measurements on living cells. Cancer Res. 2015;75(10):1983–91.
Dash P, Fiore-Gartland AJ, Hertz T, Wang GC, Sharma S, Souquette A, et al. Quantifiable predictive features define epitope-specific T cell receptor repertoires. Nature. 2017;547(7661):89–93.
Adams JJ, Narayanan S, Birnbaum ME, Sidhu SS, Blevins SJ, Gee MH, et al. Structural interplay between germline interactions and adaptive recognition determines the bandwidth of TCR-peptide-MHC cross-reactivity. Nat Immunol. 2016;17(1):87–94.
Li Y, Moysey R, Molloy PE, Vuidepot AL, Mahon T, Baston E, et al. Directed evolution of human T-cell receptors with picomolar affinities by phage display. Nat Biotechnol. 2005;23(3):349–54.
Robbins PF, Li YF, El-Gamil M, Zhao Y, Wargo JA, Zheng Z et al. Single and dual amino acid substitutions in TCR CDRs can enhance antigen-specific T cell functions. Journal of immunology (Baltimore, Md: 1950). 2008;180(9):6116-31.
D’Angelo SP, Melchiori L, Merchant MS, Bernstein D, Glod J, Kaplan R, et al. Antitumor Activity Associated with prolonged persistence of Adoptively Transferred NY-ESO-1 (c259)T cells in synovial sarcoma. Cancer Discov. 2018;8(8):944–57.
Zhang M, Xu W, Luo L, Guan F, Wang X, Zhu P, et al. Identification and affinity enhancement of T-cell receptor targeting a KRAS(G12V) cancer neoantigen. Commun Biology. 2024;7(1):512.
Schmitt TM, Aggen DH, Ishida-Tsubota K, Ochsenreither S, Kranz DM, Greenberg PD. Generation of higher affinity T cell receptors by antigen-driven differentiation of progenitor T cells in vitro. Nat Biotechnol. 2017;35(12):1188–95.
Morgan RA, Chinnasamy N, Abate-Daga D, Gros A, Robbins PF, Zheng Z et al. Cancer regression and neurological toxicity following anti-MAGE-A3 TCR gene therapy. Journal of immunotherapy (Hagerstown, Md: 1997). 2013;36(2):133 – 51.
Cameron BJ, Gerry AB, Dukes J, Harper JV, Kannan V, Bianchi FC, et al. Identification of a titin-derived HLA-A1-presented peptide as a cross-reactive target for engineered MAGE A3-directed T cells. Sci Transl Med. 2013;5(197):197ra03.
Hellman LM, Foley KC, Singh NK, Alonso JA, Riley TP, Devlin JR, et al. Improving T cell receptor On-Target specificity via structure-guided design. Mol Therapy: J Am Soc Gene Therapy. 2019;27(2):300–13.
Rosenberg AM, Baker BM. Engineering the T cell receptor for fun and profit: uncovering complex biology, interrogating the immune system, and targeting disease. Curr Opin Struct Biol. 2022;74:102358.
Lu T, Zhang Z, Zhu J, Wang Y, Jiang P, Xiao X, et al. Deep learning-based prediction of the T cell receptor-antigen binding specificity. Nat Mach Intell. 2021;3(10):864–75.
Bradley P. Structure-based prediction of T cell receptor:peptide-MHC interactions. eLife. 2023;12.
Abramson J, Adler J, Dunger J, Evans R, Green T, Pritzel A et al. Accurate structure prediction of biomolecular interactions with AlphaFold 3. Nature. 2024.
Liu B, Chen W, Evavold BD, Zhu C. Accumulation of dynamic catch bonds between TCR and agonist peptide-MHC triggers T cell signaling. Cell. 2014;157(2):357–68.
Hong J, Ge C, Jothikumar P, Yuan Z, Liu B, Bai K, et al. A TCR mechanotransduction signaling loop induces negative selection in the thymus. Nat Immunol. 2018;19(12):1379–90.
Jiang N, Huang J, Edwards LJ, Liu B, Zhang Y, Beal CD, et al. Two-stage cooperative T cell receptor-peptide major histocompatibility complex-CD8 trimolecular interactions amplify antigen discrimination. Immunity. 2011;34(1):13–23.
Liu Y, Blanchfield L, Ma VP, Andargachew R, Galior K, Liu Z, et al. DNA-based nanoparticle tension sensors reveal that T-cell receptors transmit defined pN forces to their antigens for enhanced fidelity. Proc Natl Acad Sci USA. 2016;113(20):5610–5.
Das DK, Feng Y, Mallis RJ, Li X, Keskin DB, Hussey RE, et al. Force-dependent transition in the T-cell receptor β-subunit allosterically regulates peptide discrimination and pMHC bond lifetime. Proc Natl Acad Sci USA. 2015;112(5):1517–22.
Choi HK, Cong P, Ge C, Natarajan A, Liu B, Zhang Y, et al. Catch bond models may explain how force amplifies TCR signaling and antigen discrimination. Nat Commun. 2023;14(1):2616.
Zhao X, Kolawole EM, Chan W, Feng Y, Yang X, Gee MH, et al. Tuning T cell receptor sensitivity through catch bond engineering. Volume 376. New York, NY: Science; 2022. p. eabl5282. 6589.
Liu Y, Liu G, Wang J, Zheng ZY, Jia L, Rui W et al. Chimeric STAR receptors using TCR machinery mediate robust responses against solid tumors. Sci Transl Med. 2021;13(586).
Mansilla-Soto J, Eyquem J, Haubner S, Hamieh M, Feucht J, Paillon N, et al. HLA-independent T cell receptors for targeting tumors with low antigen density. Nat Med. 2022;28(2):345–52.
Dobrin A, Lindenbergh PL, Shi Y, Perica K, Xie H, Jain N et al. Synthetic dual co-stimulation increases the potency of HIT and TCR-targeted cell therapies. Nat cancer. 2024.
Mog BJ, Marcou N, DiNapoli SR, Pearlman AH, Nichakawade TD, Hwang MS, et al. Preclinical studies show that Co-STARs combine the advantages of chimeric antigen and T cell receptors for the treatment of tumors with low antigen densities. Sci Transl Med. 2024;16(755):eadg7123.
Liddy N, Bossi G, Adams KJ, Lissina A, Mahon TM, Hassan NJ, et al. Monoclonal TCR-redirected tumor cell killing. Nat Med. 2012;18(6):980–7.
Boudousquie C, Bossi G, Hurst JM, Rygiel KA, Jakobsen BK, Hassan NJ. Polyfunctional response by ImmTAC (IMCgp100) redirected CD8(+) and CD4(+) T cells. Immunology. 2017;152(3):425–38.
Nathan P, Hassel JC, Rutkowski P, Baurain JF, Butler MO, Schlaak M, et al. Overall Survival Benefit with Tebentafusp in Metastatic Uveal Melanoma. N Engl J Med. 2021;385(13):1196–206.
Bargou R, Leo E, Zugmaier G, Klinger M, Goebeler M, Knop S, et al. Tumor regression in cancer patients by very low doses of a T cell-engaging antibody. Sci (New York NY). 2008;321(5891):974–7.
Malik H, Buelow B, Rangaswamy U, Balasubramani A, Boudreau A, Dang K, et al. TNB-486, a Novel fully human bispecific CD19 x CD3 antibody that kills CD19-Positive tumor cells with minimal cytokine secretion. Blood. 2019;134:4070.
Hoogenboom HR, de Bruïne AP, Hufton SE, Hoet RM, Arends JW, Roovers RC. Antibody phage display technology and its applications. Immunotechnology: Int J Immunological Eng. 1998;4(1):1–20.
Hsiue EH, Wright KM, Douglass J, Hwang MS, Mog BJ, Pearlman AH, et al. Targeting a neoantigen derived from a common TP53 mutation. Volume 371. New York, NY: Science; 2021. 6533.
Dao T, Pankov D, Scott A, Korontsvit T, Zakhaleva V, Xu Y, et al. Therapeutic bispecific T-cell engager antibody targeting the intracellular oncoprotein WT1. Nat Biotechnol. 2015;33(10):1079–86.
Douglass J, Hsiue EH, Mog BJ, Hwang MS, DiNapoli SR, Pearlman AH et al. Bispecific antibodies targeting mutant RAS neoantigens. Science immunology. 2021;6(57).
Hong DS, Van Tine BA, Biswas S, McAlpine C, Johnson ML, Olszanski AJ, et al. Autologous T cell therapy for MAGE-A4(+) solid cancers in HLA-A*02(+) patients: a phase 1 trial. Nat Med. 2023;29(1):104–14.
Jungmichel S, Scheifele F, Sobieraj A, Wendelspiess S, Schleier T, Knobel P, et al. Enhanced anti-tumor responses with a novel dual pMHC T-cell engager bispecific antibody. Cancer Res. 2022;82(12Supplement):2891.
Paz-Ares L, Champiat S, Lai WV, Izumi H, Govindan R, Boyer M, et al. Tarlatamab, a first-in-class DLL3-Targeted bispecific T-Cell engager, in recurrent small-cell lung Cancer: an Open-Label, phase I study. J Clin Oncology: Official J Am Soc Clin Oncol. 2023;41(16):2893–903.
Ahn MJ, Cho BC, Felip E, Korantzis I, Ohashi K, Majem M, et al. Tarlatamab for patients with previously treated small-cell Lung Cancer. N Engl J Med. 2023;389(22):2063–75.
Mullard A. FDA approves first DLL3 × CD3 bispecific T-cell engager for lung cancer. Nat Rev Drug Discovery. 2024.
Molloy ME, Aaron WH, Barath M, Bush MC, Callihan EC, Carlin K et al. HPN328, a trispecific T cell-activating protein construct targeting DLL3-Expressing solid tumors. Mol Cancer Ther. 2024.
Bacac M, Fauti T, Sam J, Colombetti S, Weinzierl T, Ouaret D, et al. A Novel Carcinoembryonic Antigen T-Cell bispecific antibody (CEA TCB) for the treatment of solid tumors. Clin cancer Research: Official J Am Association Cancer Res. 2016;22(13):3286–97.
Zong Y, Lin Y, Wei T, Cheng Q. Lipid nanoparticle (LNP) enables mRNA delivery for Cancer Therapy. Adv Mater (Deerfield Beach Fla). 2023;35(51):e2303261.
Stadler CR, Bähr-Mahmud H, Celik L, Hebich B, Roth AS, Roth RP, et al. Elimination of large tumors in mice by mRNA-encoded bispecific antibodies. Nat Med. 2017;23(7):815–7.
Stadler CR, Ellinghaus U, Fischer L, Bähr-Mahmud H, Rao M, Lindemann C, et al. Preclinical efficacy and pharmacokinetics of an RNA-encoded T cell-engaging bispecific antibody targeting human claudin 6. Sci Transl Med. 2024;16(748):eadl2720.
Ma Z, Janmey PA, Finkel TH. The receptor deformation model of TCR triggering. FASEB journal: official publication of the Federation of American Societies for Experimental Biology. 2008;22(4):1002–8.
Xu C, Gagnon E, Call ME, Schnell JR, Schwieters CD, Carman CV, et al. Regulation of T cell receptor activation by dynamic membrane binding of the CD3epsilon cytoplasmic tyrosine-based motif. Cell. 2008;135(4):702–13.
DeFord-Watts LM, Dougall DS, Belkaya S, Johnson BA, Eitson JL, Roybal KT et al. The CD3 zeta subunit contains a phosphoinositide-binding motif that is required for the stable accumulation of TCR-CD3 complex at the immunological synapse. Journal of immunology (Baltimore, Md: 1950). 2011;186(12):6839-47.
Call ME, Pyrdol J, Wiedmann M, Wucherpfennig KW. The organizing principle in the formation of the T cell receptor-CD3 complex. Cell. 2002;111(7):967–79.
Krshnan L, Park S, Im W, Call MJ, Call ME. A conserved αβ transmembrane interface forms the core of a compact T-cell receptor-CD3 structure within the membrane. Proc Natl Acad Sci USA. 2016;113(43):E6649–58.
Irving BA, Weiss A. The cytoplasmic domain of the T cell receptor zeta chain is sufficient to couple to receptor-associated signal transduction pathways. Cell. 1991;64(5):891–901.
Call ME, Schnell JR, Xu C, Lutz RA, Chou JJ, Wucherpfennig KW. The structure of the zetazeta transmembrane dimer reveals features essential for its assembly with the T cell receptor. Cell. 2006;127(2):355–68.
Lee MS, Glassman CR, Deshpande NR, Badgandi HB, Parrish HL, Uttamapinant C, et al. A mechanical switch couples T cell receptor triggering to the cytoplasmic juxtamembrane regions of CD3ζζ. Immunity. 2015;43(2):227–39.
Brazin KN, Mallis RJ, Boeszoermenyi A, Feng Y, Yoshizawa A, Reche PA, et al. The T Cell Antigen receptor α transmembrane domain coordinates triggering through regulation of Bilayer Immersion and CD3 Subunit associations. Immunity. 2018;49(5):829–e416.
Aivazian D, Stern LJ. Phosphorylation of T cell receptor zeta is regulated by a lipid dependent folding transition. Nat Struct Biol. 2000;7(11):1023–6.
Guo X, Yan C, Li H, Huang W, Shi X, Huang M, et al. Lipid-dependent conformational dynamics underlie the functional versatility of T-cell receptor. Cell Res. 2017;27(4):505–25.
van Oers NS, Killeen N, Weiss A. ZAP-70 is constitutively associated with tyrosine-phosphorylated TCR zeta in murine thymocytes and lymph node T cells. Immunity. 1994;1(8):675–85.
Zhang W, Sloan-Lancaster J, Kitchen J, Trible RP, Samelson LE. LAT: the ZAP-70 tyrosine kinase substrate that links T cell receptor to cellular activation. Cell. 1998;92(1):83–92.
Laletin V, Bernard PL, Costa da Silva C, Guittard G, Nunes JA. Negative intracellular regulators of T-cell receptor (TCR) signaling as potential antitumor immunotherapy targets. J Immunother Cancer. 2023;11(5).
Fawcett VC, Lorenz U. Localization of Src homology 2 domain-containing phosphatase 1 (SHP-1) to lipid rafts in T lymphocytes: functional implications and a role for the SHP-1 carboxyl terminus. Journal of immunology (Baltimore, Md: 1950). 2005;174(5):2849-59.
LaFleur MW, Nguyen TH, Coxe MA, Miller BC, Yates KB, Gillis JE, et al. PTPN2 regulates the generation of exhausted CD8(+) T cell subpopulations and restrains tumor immunity. Nat Immunol. 2019;20(10):1335–47.
Poirier A, Ormonde JVS, Aubry I, Abidin BM, Feng CH, Martinez-Cordova Z, et al. The induction of SHP-1 degradation by TAOK3 ensures the responsiveness of T cells to TCR stimulation. Sci Signal. 2024;17(817):eadg4422.
Liang S, Tran E, Du X, Dong J, Sudholz H, Chen H, et al. A small molecule inhibitor of PTP1B and PTPN2 enhances T cell anti-tumor immunity. Nat Commun. 2023;14(1):4524.
Mikhailik A, Ford B, Keller J, Chen Y, Nassar N, Carpino N. A phosphatase activity of Sts-1 contributes to the suppression of TCR signaling. Mol Cell. 2007;27(3):486–97.
Aziz F, Reddy K, Fernandez Vega V, Dey R, Hicks KA, Rao S, et al. Rebamipide and derivatives are potent, selective inhibitors of histidine phosphatase activity of the suppressor of T cell receptor signaling proteins. J Med Chem. 2024;67(3):1949–60.
Eggert J, Zinzow-Kramer WM, Hu Y, Kolawole EM, Tsai YL, Weiss A, et al. Cbl-b mitigates the responsiveness of naive CD8(+) T cells that experience extensive tonic T cell receptor signaling. Sci Signal. 2024;17(822):eadh0439.
Wallner S, Gruber T, Baier G, Wolf D. Releasing the brake: targeting Cbl-b to enhance lymphocyte effector functions. Clin Dev Immunol. 2012;2012:692639.
Tsai YL, Arias-Badia M, Kadlecek TA, Lwin YM, Srinath A, Shah NH, et al. TCR signaling promotes formation of an STS1-Cbl-b complex with pH-sensitive phosphatase activity that suppresses T cell function in acidic environments. Immunity. 2023;56(12):2682–e989.
Hamid M, Cespedes PF, Jin C, Chen JL, Gileadi U, Antoun E, et al. Unconventional human CD61 pairing with CD103 promotes TCR signaling and antigen-specific T cell cytotoxicity. Nat Immunol. 2024;25(5):834–46.
Duan H, Jing L, Jiang X, Ma Y, Wang D, Xiang J et al. CD146 bound to LCK promotes T cell receptor signaling and antitumor immune responses in mice. J Clin Investig. 2021;131(21).
Wang Q, Zhu T, Miao N, Qu Y, Wang Z, Chao Y, et al. Disulfiram bolsters T-cell anti-tumor immunity through direct activation of LCK-mediated TCR signaling. EMBO J. 2022;41(16):e110636.
Rhee SG, Choi KD. Regulation of inositol phospholipid-specific phospholipase C isozymes. J Biol Chem. 1992;267(18):12393–6.
Berridge MJ. Inositol trisphosphate and calcium signalling mechanisms. Biochim Biophys Acta. 2009;1793(6):933–40.
Oh-hora M, Rao A. The calcium/NFAT pathway: role in development and function of regulatory T cells. Microbes Infect. 2009;11(5):612–9.
Lee KY, D’Acquisto F, Hayden MS, Shim JH, Ghosh S. PDK1 nucleates T cell receptor-induced signaling complex for NF-kappaB activation. Sci (New York NY). 2005;308(5718):114–8.
Matsumoto R, Wang D, Blonska M, Li H, Kobayashi M, Pappu B, et al. Phosphorylation of CARMA1 plays a critical role in T cell receptor-mediated NF-kappaB activation. Immunity. 2005;23(6):575–85.
Blonska M, Lin X. NF-κB signaling pathways regulated by CARMA family of scaffold proteins. Cell Res. 2011;21(1):55–70.
Sun Z, Arendt CW, Ellmeier W, Schaeffer EM, Sunshine MJ, Gandhi L, et al. PKC-theta is required for TCR-induced NF-kappaB activation in mature but not immature T lymphocytes. Nature. 2000;404(6776):402–7.
Samelson LE. Signal transduction mediated by the T cell antigen receptor: the role of adapter proteins. Annu Rev Immunol. 2002;20:371–94.
Liu X, Hao J, Wei P, Zhao X, Lan Q, Ni L, et al. SMAD4, activated by the TCR-triggered MEK/ERK signaling pathway, critically regulates CD8(+) T cell cytotoxic function. Sci Adv. 2022;8(30):eabo4577.
Peterson TR, Laplante M, Thoreen CC, Sancak Y, Kang SA, Kuehl WM, et al. DEPTOR is an mTOR inhibitor frequently overexpressed in multiple myeloma cells and required for their survival. Cell. 2009;137(5):873–86.
Pollizzi KN, Powell JD. Regulation of T cells by mTOR: the known knowns and the known unknowns. Trends Immunol. 2015;36(1):13–20.
van Slegtenhorst M, Nellist M, Nagelkerken B, Cheadle J, Snell R, van den Ouweland A, et al. Interaction between hamartin and tuberin, the TSC1 and TSC2 gene products. Hum Mol Genet. 1998;7(6):1053–7.
Patel CH, Dong Y, Koleini N, Wang X, Dunkerly-Eyring BL, Wen J et al. TSC2 S1365A mutation potently regulates CD8 + T cell function and differentiation and improves adoptive cellular cancer therapy. JCI Insight. 2023;8(21).
Gamble N, Bradu A, Caldwell JA, McKeever J, Bolonduro O, Ermis E, et al. PU.1 and BCL11B sequentially cooperate with RUNX1 to anchor mSWI/SNF to poise the T cell effector landscape. Nat Immunol. 2024;25(5):860–72.
Baessler A, Vignali DAA. T Cell Exhaustion. Annual review of immunology. 2024.
Baitsch L, Baumgaertner P, Devêvre E, Raghav SK, Legat A, Barba L, et al. Exhaustion of tumor-specific CD8⁺ T cells in metastases from melanoma patients. J Clin Investig. 2011;121(6):2350–60.
Guo X, Zhang Y, Zheng L, Zheng C, Song J, Zhang Q, et al. Global characterization of T cells in non-small-cell lung cancer by single-cell sequencing. Nat Med. 2018;24(7):978–85.
Cillo AR, Kürten CHL, Tabib T, Qi Z, Onkar S, Wang T, et al. Immune Landscape of viral- and carcinogen-driven Head and Neck Cancer. Immunity. 2020;52(1):183–e999.
Belk JA, Daniel B, Satpathy AT. Epigenetic regulation of T cell exhaustion. Nat Immunol. 2022;23(6):848–60.
Klebanoff CA, Gattinoni L, Palmer DC, Muranski P, Ji Y, Hinrichs CS, et al. Determinants of successful CD8 + T-cell adoptive immunotherapy for large established tumors in mice. Clin cancer Research: Official J Am Association Cancer Res. 2011;17(16):5343–52.
Shalem O, Sanjana NE, Hartenian E, Shi X, Scott DA, Mikkelson T, et al. Genome-scale CRISPR-Cas9 knockout screening in human cells. Sci (New York NY). 2014;343(6166):84–7.
Weber BN, Chi AW, Chavez A, Yashiro-Ohtani Y, Yang Q, Shestova O, et al. A critical role for TCF-1 in T-lineage specification and differentiation. Nature. 2011;476(7358):63–8.
Tejera MM, Kim EH, Sullivan JA, Plisch EH, Suresh M. FoxO1 controls effector-to-memory transition and maintenance of functional CD8 T cell memory. Journal of immunology (Baltimore, Md: 1950). 2013;191(1):187 – 99.
Li Y, Han M, Wei H, Huang W, Chen Z, Zhang T, et al. Id2 epigenetically controls CD8(+) T-cell exhaustion by disrupting the assembly of the Tcf3-LSD1 complex. Cell Mol Immunol. 2024;21(3):292–308.
Martinez GJ, Pereira RM, Äijö T, Kim EY, Marangoni F, Pipkin ME, et al. The transcription factor NFAT promotes exhaustion of activated CD8⁺ T cells. Immunity. 2015;42(2):265–78.
Khan O, Giles JR, McDonald S, Manne S, Ngiow SF, Patel KP, et al. TOX transcriptionally and epigenetically programs CD8(+) T cell exhaustion. Nature. 2019;571(7764):211–8.
Tsao HW, Kaminski J, Kurachi M, Barnitz RA, DiIorio MA, LaFleur MW, et al. Batf-mediated epigenetic control of effector CD8(+) T cell differentiation. Sci Immunol. 2022;7(68):eabi4919.
Seo H, González-Avalos E, Zhang W, Ramchandani P, Yang C, Lio CJ, et al. BATF and IRF4 cooperate to counter exhaustion in tumor-infiltrating CAR T cells. Nat Immunol. 2021;22(8):983–95.
Robert C. A decade of immune-checkpoint inhibitors in cancer therapy. Nat Commun. 2020;11(1):3801.
Pardoll DM. The blockade of immune checkpoints in cancer immunotherapy. Nat Rev Cancer. 2012;12(4):252–64.
Philips EA, Liu J, Kvalvaag A, Mørch AM, Tocheva AS, Ng C, et al. Transmembrane domain-driven PD-1 dimers mediate T cell inhibition. Sci Immunol. 2024;9(93):eade6256.
Woo SR, Turnis ME, Goldberg MV, Bankoti J, Selby M, Nirschl CJ, et al. Immune inhibitory molecules LAG-3 and PD-1 synergistically regulate T-cell function to promote tumoral immune escape. Cancer Res. 2012;72(4):917–27.
Silberstein JL, Du J, Chan KW, Frank JA, Mathews II, Kim YB, et al. Structural insights reveal interplay between LAG-3 homodimerization, ligand binding, and function. Proc Natl Acad Sci USA. 2024;121(12):e2310866121.
Maeda TK, Sugiura D, Okazaki IM, Maruhashi T, Okazaki T. Atypical motifs in the cytoplasmic region of the inhibitory immune co-receptor LAG-3 inhibit T cell activation. J Biol Chem. 2019;294(15):6017–26.
Guy C, Mitrea DM, Chou PC, Temirov J, Vignali KM, Liu X, et al. LAG3 associates with TCR-CD3 complexes and suppresses signaling by driving co-receptor-lck dissociation. Nat Immunol. 2022;23(5):757–67.
Fourcade J, Sun Z, Benallaoua M, Guillaume P, Luescher IF, Sander C, et al. Upregulation of Tim-3 and PD-1 expression is associated with tumor antigen-specific CD8 + T cell dysfunction in melanoma patients. J Exp Med. 2010;207(10):2175–86.
Huang YH, Zhu C, Kondo Y, Anderson AC, Gandhi A, Russell A, et al. CEACAM1 regulates TIM-3-mediated tolerance and exhaustion. Nature. 2015;517(7534):386–90.
Worboys JD, Vowell KN, Hare RK, Ambrose AR, Bertuzzi M, Conner MA, et al. TIGIT can inhibit T cell activation via ligation-induced nanoclusters, independent of CD226 co-stimulation. Nat Commun. 2023;14(1):5016.
Gavrieli M, Watanabe N, Loftin SK, Murphy TL, Murphy KM. Characterization of phosphotyrosine binding motifs in the cytoplasmic domain of B and T lymphocyte attenuator required for association with protein tyrosine phosphatases SHP-1 and SHP-2. Biochemical and biophysical research communications. 2003;312(4):1236–43.
Sedy JR, Gavrieli M, Potter KG, Hurchla MA, Lindsley RC, Hildner K, et al. B and T lymphocyte attenuator regulates T cell activation through interaction with herpesvirus entry mediator. Nat Immunol. 2005;6(1):90–8.
Tawbi HA, Schadendorf D, Lipson EJ, Ascierto PA, Matamala L, Castillo Gutiérrez E, et al. Relatlimab and Nivolumab versus Nivolumab in Untreated Advanced Melanoma. N Engl J Med. 2022;386(1):24–34.
Wang-Gillam A, Plambeck-Suess S, Goedegebuure P, Simon PO, Mitchem JB, Hornick JR, et al. A phase I study of IMP321 and gemcitabine as the front-line therapy in patients with advanced pancreatic adenocarcinoma. Investig New Drugs. 2013;31(3):707–13.
Sakuishi K, Apetoh L, Sullivan JM, Blazar BR, Kuchroo VK, Anderson AC. Targeting Tim-3 and PD-1 pathways to reverse T cell exhaustion and restore anti-tumor immunity. J Exp Med. 2010;207(10):2187–94.
Ngiow SF, von Scheidt B, Akiba H, Yagita H, Teng MW, Smyth MJ. Anti-TIM3 antibody promotes T cell IFN-γ-mediated antitumor immunity and suppresses established tumors. Cancer Res. 2011;71(10):3540–51.
Johnston RJ, Comps-Agrar L, Hackney J, Yu X, Huseni M, Yang Y, et al. The immunoreceptor TIGIT regulates antitumor and antiviral CD8(+) T cell effector function. Cancer Cell. 2014;26(6):923–37.
Chauvin JM, Pagliano O, Fourcade J, Sun Z, Wang H, Sander C, et al. TIGIT and PD-1 impair tumor antigen-specific CD8⁺ T cells in melanoma patients. J Clin Investig. 2015;125(5):2046–58.
Sordo-Bahamonde C, Lorenzo-Herrero S, Granda-Díaz R, Martínez-Pérez A, Aguilar-García C, Rodrigo JP, et al. Beyond the anti-PD-1/PD-L1 era: promising role of the BTLA/HVEM axis as a future target for cancer immunotherapy. Mol Cancer. 2023;22(1):142.
Wang X, Zha H, Wu W, Yuan T, Xie S, Jin Z, et al. CD200(+) cytotoxic T lymphocytes in the tumor microenvironment are crucial for efficacious anti-PD-1/PD-L1 therapy. Sci Transl Med. 2023;15(679):eabn5029.
Rijkers ES, de Ruiter T, Baridi A, Veninga H, Hoek RM, Meyaard L. The inhibitory CD200R is differentially expressed on human and mouse T and B lymphocytes. Mol Immunol. 2008;45(4):1126–35.
Fenaux J, Fang X, Huang YM, Melero C, Bonnans C, Lowe EL, et al. 23ME-00610, a genetically informed, first-in-class antibody targeting CD200R1 to enhance antitumor T cell function. Oncoimmunology. 2023;12(1):2217737.
Xie X, Chen C, Chen W, Jiang J, Wang L, Li T, et al. Structural basis of VSIG3: the Ligand for VISTA. Front Immunol. 2021;12:625808.
Liu J, Yuan Y, Chen W, Putra J, Suriawinata AA, Schenk AD, et al. Immune-checkpoint proteins VISTA and PD-1 nonredundantly regulate murine T-cell responses. Proc Natl Acad Sci USA. 2015;112(21):6682–7.
Deng S, Zhang Y, Wang H, Liang W, Xie L, Li N, et al. ITPRIPL1 binds CD3ε to impede T cell activation and enable tumor immune evasion. Cell. 2024;187(9):2305–e2333.
Zhang T, Lin Y, Gao Q. Bispecific antibodies targeting immunomodulatory checkpoints for cancer therapy. Cancer Biology Med. 2023;20(3):181–95.
Hoogi S, Eisenberg V, Mayer S, Shamul A, Barliya T, Cohen CJ. A TIGIT-based chimeric co-stimulatory switch receptor improves T-cell anti-tumor function. J Immunother Cancer. 2019;7(1):243.
Oda SK, Daman AW, Garcia NM, Wagener F, Schmitt TM, Tan X, et al. A CD200R-CD28 fusion protein appropriates an inhibitory signal to enhance T-cell function and therapy of murine leukemia. Blood. 2017;130(22):2410–9.
Zhao S, Wang C, Lu P, Lou Y, Liu H, Wang T et al. Switch receptor T3/28 improves long-term persistence and antitumor efficacy of CAR-T cells. J Immunother Cancer. 2021;9(12).
Liu X, Ranganathan R, Jiang S, Fang C, Sun J, Kim S, et al. A chimeric switch-receptor targeting PD1 augments the efficacy of second-generation CAR T cells in Advanced Solid tumors. Cancer Res. 2016;76(6):1578–90.
Zhang J, Li J, Hou Y, Lin Y, Zhao H, Shi Y et al. Osr2 functions as a biomechanical checkpoint to aggravate CD8(+) T cell exhaustion in tumor. Cell. 2024.
Shibuya K, Shirakawa J, Kameyama T, Honda S, Tahara-Hanaoka S, Miyamoto A, et al. CD226 (DNAM-1) is involved in lymphocyte function-associated antigen 1 costimulatory signal for naive T cell differentiation and proliferation. J Exp Med. 2003;198(12):1829–39.
Weulersse M, Asrir A, Pichler AC, Lemaitre L, Braun M, Carrié N, et al. Eomes-dependent loss of the co-activating receptor CD226 restrains CD8(+) T cell anti-tumor functions and limits the efficacy of Cancer Immunotherapy. Immunity. 2020;53(4):824–e3910.
Sun L, Jiao A, Liu H, Ding R, Yuan N, Yang B, et al. Targeting a disintegrin and metalloprotease (ADAM) 17-CD122 axis enhances CD8(+) T cell effector differentiation and anti-tumor immunity. Signal Transduct Target Therapy. 2024;9(1):152.
Bennion KB, Liu D, Dawood AS, Wyatt MM, Alexander KL, Abdel-Hakeem MS, et al. CD8(+) T cell-derived Fgl2 regulates immunity in a cell-autonomous manner via ligation of FcγRIIB. Nat Commun. 2024;15(1):5280.
Mélique S, Vadel A, Rouquié N, Yang C, Bories C, Cotineau C, et al. THEMIS promotes T cell development and maintenance by rising the signaling threshold of the inhibitory receptor BTLA. Proc Natl Acad Sci USA. 2024;121(20):e2318773121.
Hope JL, Otero DC, Bae EA, Stairiker CJ, Palete AB, Faso HA, et al. PSGL-1 attenuates early TCR signaling to suppress CD8(+) T cell progenitor differentiation and elicit terminal CD8(+) T cell exhaustion. Cell Rep. 2023;42(5):112436.
Johnston RJ, Su LJ, Pinckney J, Critton D, Boyer E, Krishnakumar A, et al. VISTA is an acidic pH-selective ligand for PSGL-1. Nature. 2019;574(7779):565–70.
Wang T, Wei JJ, Sabatini DM, Lander ES. Genetic screens in human cells using the CRISPR-Cas9 system. Science (New York, NY). 2014;343(6166):80 – 4.
Schlabach MR, Lin S, Collester ZR, Wrocklage C, Shenker S, Calnan C et al. Rational design of a SOCS1-edited tumor-infiltrating lymphocyte therapy using CRISPR/Cas9 screens. J Clin Investig. 2023;133(24).
Anderson P. Post-transcriptional regulons coordinate the initiation and resolution of inflammation. Nat Rev Immunol. 2010;10(1):24–35.
Wei J, Long L, Zheng W, Dhungana Y, Lim SA, Guy C, et al. Targeting REGNASE-1 programs long-lived effector T cells for cancer therapy. Nature. 2019;576(7787):471–6.
Mai D, Johnson O, Reff J, Fan TJ, Scholler J, Sheppard NC, et al. Combined disruption of T cell inflammatory regulators Regnase-1 and Roquin-1 enhances antitumor activity of engineered human T cells. Proc Natl Acad Sci USA. 2023;120(12):e2218632120.
Flosbach M, Oberle SG, Scherer S, Zecha J, von Hoesslin M, Wiede F, et al. PTPN2 Deficiency enhances programmed T cell expansion and survival capacity of activated T cells. Cell Rep. 2020;32(4):107957.
Baumgartner CK, Ebrahimi-Nik H, Iracheta-Vellve A, Hamel KM, Olander KE, Davis TGR, et al. The PTPN2/PTPN1 inhibitor ABBV-CLS-484 unleashes potent anti-tumour immunity. Nature. 2023;622(7984):850–62.
Carnevale J, Shifrut E, Kale N, Nyberg WA, Blaeschke F, Chen YY, et al. RASA2 ablation in T cells boosts antigen sensitivity and long-term function. Nature. 2022;609(7925):174–82.
Shang W, Jiang Y, Boettcher M, Ding K, Mollenauer M, Liu Z, et al. Genome-wide CRISPR screen identifies FAM49B as a key regulator of actin dynamics and T cell activation. Proc Natl Acad Sci USA. 2018;115(17):E4051–60.
Gurusamy D, Henning AN, Yamamoto TN, Yu Z, Zacharakis N, Krishna S, et al. Multi-phenotype CRISPR-Cas9 screen identifies p38 kinase as a target for adoptive immunotherapies. Cancer Cell. 2020;37(6):818–e339.
Belk JA, Yao W, Ly N, Freitas KA, Chen YT, Shi Q, et al. Genome-wide CRISPR screens of T cell exhaustion identify chromatin remodeling factors that limit T cell persistence. Cancer Cell. 2022;40(7):768–e867.
Ye L, Park JJ, Peng L, Yang Q, Chow RD, Dong MB, et al. A genome-scale gain-of-function CRISPR screen in CD8 T cells identifies proline metabolism as a means to enhance CAR-T therapy. Cell Metabol. 2022;34(4):595–e61414.
Garcia J, Daniels J, Lee Y, Zhu I, Cheng K, Liu Q, et al. Naturally occurring T cell mutations enhance engineered T cell therapies. Nature. 2024;626(7999):626–34.
Shifrut E, Carnevale J, Tobin V, Roth TL, Woo JM, Bui CT, et al. Genome-wide CRISPR screens in primary human T cells reveal key regulators of Immune function. Cell. 2018;175(7):1958–e7115.
Tieu V, Sotillo E, Bjelajac JR, Chen C, Malipatlolla M, Guerrero JA, et al. A versatile CRISPR-Cas13d platform for multiplexed transcriptomic regulation and metabolic engineering in primary human T cells. Cell. 2024;187(5):1278–e9520.
Blaeschke F, Chen YY, Apathy R, Daniel B, Chen AY, Chen PA, et al. Modular pooled discovery of synthetic knockin sequences to program durable cell therapies. Cell. 2023;186(19):4216–e3433.
Walsh ZH, Shah P, Kothapalli N, Shah SB, Nikolenyi G, Brodtman DZ et al. Mapping variant effects on anti-tumor hallmarks of primary human T cells with base-editing screens. Nat Biotechnol. 2024.
Yamada-Hunter SA, Theruvath J, McIntosh BJ, Freitas KA, Lin F, Radosevich MT, et al. Engineered CD47 protects T cells for enhanced antitumour immunity. Nature. 2024;630(8016):457–65.
Drakes DJ, Rafiq S, Purdon TJ, Lopez AV, Chandran SS, Klebanoff CA, et al. Optimization of T-cell receptor-modified T cells for Cancer Therapy. Cancer Immunol Res. 2020;8(6):743–55.
Liu Y, Zhou N, Zhou L, Wang J, Zhou Y, Zhang T, et al. IL-2 regulates tumor-reactive CD8(+) T cell exhaustion by activating the aryl hydrocarbon receptor. Nat Immunol. 2021;22(3):358–69.
Levin AM, Bates DL, Ring AM, Krieg C, Lin JT, Su L, et al. Exploiting a natural conformational switch to engineer an interleukin-2 ‘superkine’. Nature. 2012;484(7395):529–33.
Mo F, Yu Z, Li P, Oh J, Spolski R, Zhao L, et al. An engineered IL-2 partial agonist promotes CD8(+) T cell stemness. Nature. 2021;597(7877):544–8.
Sockolosky JT, Trotta E, Parisi G, Picton L, Su LL, Le AC, et al. Selective targeting of engineered T cells using orthogonal IL-2 cytokine-receptor complexes. Volume 359. New York, NY: Science; 2018. pp. 1037–42. 6379.
Corria-Osorio J, Carmona SJ, Stefanidis E, Andreatta M, Ortiz-Miranda Y, Muller T, et al. Orthogonal cytokine engineering enables novel synthetic effector states escaping canonical exhaustion in tumor-rejecting CD8(+) T cells. Nat Immunol. 2023;24(5):869–83.
Morsut L, Roybal KT, Xiong X, Gordley RM, Coyle SM, Thomson M, et al. Engineering customized cell sensing and response behaviors using synthetic notch receptors. Cell. 2016;164(4):780–91.
Allen GM, Frankel NW, Reddy NR, Bhargava HK, Yoshida MA, Stark SR, et al. Synthetic cytokine circuits that drive T cells into immune-excluded tumors. Volume 378. New York, NY: Science; 2022. p. eaba1624. 6625.
Nguyen R, Doubrovina E, Mousset CM, Jin BY, Okada R, Zhang X, et al. Cooperative Armoring of CAR and TCR T Cells by T cell-restricted IL15 and IL21 universally enhances solid Tumor Efficacy. Clin cancer Research: Official J Am Association Cancer Res. 2024;30(8):1555–66.
Zhu W, Zhang Z, Chen J, Chen X, Huang L, Zhang X, et al. A novel engineered IL-21 receptor arms T-cell receptor-engineered T cells (TCR-T cells) against hepatocellular carcinoma. Signal Transduct Target Therapy. 2024;9(1):101.
Joerger M, Calvo E, Laubli H, Lopez J, Alonso G, de la Corral E et al. Phase 1 first-in-human dose-escalation study of ANV419 in patients with relapsed/refractory advanced solid tumors. J Immunother Cancer. 2023;11(11).
Baldwin JE, Krebs H. The evolution of metabolic cycles. Nature. 1981;291(5814):381–2.
Simula L, Fumagalli M, Vimeux L, Rajnpreht I, Icard P, Birsen G, et al. Mitochondrial metabolism sustains CD8(+) T cell migration for an efficient infiltration into solid tumors. Nat Commun. 2024;15(1):2203.
Ryan DG, Yang M, Prag HA, Blanco GR, Nikitopoulou E, Segarra-Mondejar M et al. Disruption of the TCA cycle reveals an ATF4-dependent integration of redox and amino acid metabolism. eLife. 2021;10.
Gudgeon N, Munford H, Bishop EL, Hill J, Fulton-Ward T, Bending D, et al. Succinate uptake by T cells suppresses their effector function via inhibition of mitochondrial glucose oxidation. Cell Rep. 2022;40(7):111193.
Balaban RS. Regulation of oxidative phosphorylation in the mammalian cell. Am J Physiol. 1990;258(3 Pt 1):C377–89.
Nakaya M, Xiao Y, Zhou X, Chang JH, Chang M, Cheng X, et al. Inflammatory T cell responses rely on amino acid transporter ASCT2 facilitation of glutamine uptake and mTORC1 kinase activation. Immunity. 2014;40(5):692–705.
Blagih J, Coulombe F, Vincent EE, Dupuy F, Galicia-Vázquez G, Yurchenko E, et al. The energy sensor AMPK regulates T cell metabolic adaptation and effector responses in vivo. Immunity. 2015;42(1):41–54.
Sena LA, Li S, Jairaman A, Prakriya M, Ezponda T, Hildeman DA, et al. Mitochondria are required for antigen-specific T cell activation through reactive oxygen species signaling. Immunity. 2013;38(2):225–36.
Tan H, Yang K, Li Y, Shaw TI, Wang Y, Blanco DB, et al. Integrative Proteomics and Phosphoproteomics Profiling reveals Dynamic Signaling Networks and Bioenergetics pathways underlying T cell activation. Immunity. 2017;46(3):488–503.
Wang R, Dillon CP, Shi LZ, Milasta S, Carter R, Finkelstein D, et al. The transcription factor Myc controls metabolic reprogramming upon T lymphocyte activation. Immunity. 2011;35(6):871–82.
Yang K, Shrestha S, Zeng H, Karmaus PW, Neale G, Vogel P, et al. T cell exit from quiescence and differentiation into Th2 cells depend on Raptor-mTORC1-mediated metabolic reprogramming. Immunity. 2013;39(6):1043–56.
Bricker DK, Taylor EB, Schell JC, Orsak T, Boutron A, Chen YC, et al. A mitochondrial pyruvate carrier required for pyruvate uptake in yeast, Drosophila, and humans. Volume 337. New York, NY: Science; 2012. pp. 96–100. 6090.
Yang C, Ko B, Hensley CT, Jiang L, Wasti AT, Kim J, et al. Glutamine oxidation maintains the TCA cycle and cell survival during impaired mitochondrial pyruvate transport. Mol Cell. 2014;56(3):414–24.
Man K, Miasari M, Shi W, Xin A, Henstridge DC, Preston S, et al. The transcription factor IRF4 is essential for TCR affinity-mediated metabolic programming and clonal expansion of T cells. Nat Immunol. 2013;14(11):1155–65.
Rabinowitz JD, Beeson C, Wülfing C, Tate K, Allen PM, Davis MM, et al. Altered T cell receptor ligands trigger a subset of early T cell signals. Immunity. 1996;5(2):125–35.
Frauwirth KA, Riley JL, Harris MH, Parry RV, Rathmell JC, Plas DR, et al. The CD28 signaling pathway regulates glucose metabolism. Immunity. 2002;16(6):769–77.
Jacobs SR, Herman CE, Maciver NJ, Wofford JA, Wieman HL, Hammen JJ et al. Glucose uptake is limiting in T cell activation and requires CD28-mediated Akt-dependent and independent pathways. Journal of immunology (Baltimore, Md: 1950). 2008;180(7):4476-86.
Klein Geltink RI, O’Sullivan D, Corrado M, Bremser A, Buck MD, Buescher JM, et al. Mitochondrial Priming CD28 Cell. 2017;171(2):385–e9711.
Buck MD, O’Sullivan D, Klein Geltink RI, Curtis JD, Chang CH, Sanin DE, et al. Mitochondrial Dynamics Controls T Cell Fate through Metabolic Programming. Cell. 2016;166(1):63–76.
Ron-Harel N, Santos D, Ghergurovich JM, Sage PT, Reddy A, Lovitch SB, et al. Mitochondrial Biogenesis and Proteome Remodeling promote one-Carbon metabolism for T cell activation. Cell Metabol. 2016;24(1):104–17.
Ducker GS, Chen L, Morscher RJ, Ghergurovich JM, Esposito M, Teng X, et al. Reversal of Cytosolic one-Carbon Flux compensates for loss of the mitochondrial folate pathway. Cell Metabol. 2016;23(6):1140–53.
Ma EH, Bantug G, Griss T, Condotta S, Johnson RM, Samborska B, et al. Serine is an essential metabolite for Effector T cell expansion. Cell Metabol. 2017;25(2):345–57.
Kory N, Wyant GA, Prakash G, Uit de Bos J, Bottanelli F, Pacold ME, et al. SFXN1 is a mitochondrial serine transporter required for one-carbon metabolism. Volume 362. New York, NY: Science; 2018. 6416.
Hung MH, Lee JS, Ma C, Diggs LP, Heinrich S, Chang CW, et al. Tumor methionine metabolism drives T-cell exhaustion in hepatocellular carcinoma. Nat Commun. 2021;12(1):1455.
Luda KM, Longo J, Kitchen-Goosen SM, Duimstra LR, Ma EH, Watson MJ, et al. Ketolysis drives CD8(+) T cell effector function through effects on histone acetylation. Immunity. 2023;56(9):2021–e358.
Puigserver P, Wu Z, Park CW, Graves R, Wright M, Spiegelman BM. A cold-inducible coactivator of nuclear receptors linked to adaptive thermogenesis. Cell. 1998;92(6):829–39.
Wu Z, Puigserver P, Andersson U, Zhang C, Adelmant G, Mootha V, et al. Mechanisms controlling mitochondrial biogenesis and respiration through the thermogenic coactivator PGC-1. Cell. 1999;98(1):115–24.
Scharping NE, Menk AV, Moreci RS, Whetstone RD, Dadey RE, Watkins SC, et al. The Tumor Microenvironment represses T cell mitochondrial Biogenesis to Drive Intratumoral T Cell metabolic insufficiency and dysfunction. Immunity. 2016;45(2):374–88.
Dumauthioz N, Tschumi B, Wenes M, Marti B, Wang H, Franco F, et al. Enforced PGC-1α expression promotes CD8 T cell fitness, memory formation and antitumor immunity. Cell Mol Immunol. 2021;18(7):1761–71.
Lontos K, Wang Y, Joshi SK, Frisch AT, Watson MJ, Kumar A et al. Metabolic reprogramming via an engineered PGC-1α improves human chimeric antigen receptor T-cell therapy against solid tumors. J Immunother Cancer. 2023;11(3).
Menk AV, Scharping NE, Rivadeneira DB, Calderon MJ, Watson MJ, Dunstane D, et al. 4-1BB costimulation induces T cell mitochondrial function and biogenesis enabling cancer immunotherapeutic responses. J Exp Med. 2018;215(4):1091–100.
Chang CH, Curtis JD, Maggi LB Jr., Faubert B, Villarino AV, O’Sullivan D, et al. Posttranscriptional control of T cell effector function by aerobic glycolysis. Cell. 2013;153(6):1239–51.
Macintyre AN, Gerriets VA, Nichols AG, Michalek RD, Rudolph MC, Deoliveira D, et al. The glucose transporter Glut1 is selectively essential for CD4 T cell activation and effector function. Cell Metabol. 2014;20(1):61–72.
Ho PC, Bihuniak JD, Macintyre AN, Staron M, Liu X, Amezquita R, et al. Phosphoenolpyruvate is a metabolic checkpoint of anti-tumor T cell responses. Cell. 2015;162(6):1217–28.
Ma R, Ji T, Zhang H, Dong W, Chen X, Xu P, et al. A Pck1-directed glycogen metabolic program regulates formation and maintenance of memory CD8(+) T cells. Nat Cell Biol. 2018;20(1):21–7.
Feng Q, Liu Z, Yu X, Huang T, Chen J, Wang J, et al. Lactate increases stemness of CD8 + T cells to augment anti-tumor immunity. Nat Commun. 2022;13(1):4981.
Vander Heiden MG, Cantley LC, Thompson CB. Understanding the Warburg effect: the metabolic requirements of cell proliferation. Sci (New York NY). 2009;324(5930):1029–33.
Boidot R, Végran F, Meulle A, Le Breton A, Dessy C, Sonveaux P, et al. Regulation of monocarboxylate transporter MCT1 expression by p53 mediates inward and outward lactate fluxes in tumors. Cancer Res. 2012;72(4):939–48.
Quinn WJ 3rd, Jiao J, TeSlaa T, Stadanlick J, Wang Z, Wang L, et al. Lactate limits T cell proliferation via the NAD(H) Redox state. Cell Rep. 2020;33(11):108500.
Angelin A, Gil-de-Gómez L, Dahiya S, Jiao J, Guo L, Levine MH, et al. Foxp3 reprograms T cell metabolism to function in Low-Glucose, high-lactate environments. Cell Metabol. 2017;25(6):1282–e937.
Fischer K, Hoffmann P, Voelkl S, Meidenbauer N, Ammer J, Edinger M, et al. Inhibitory effect of tumor cell-derived lactic acid on human T cells. Blood. 2007;109(9):3812–9.
Hermans D, Gautam S, García-Cañaveras JC, Gromer D, Mitra S, Spolski R, et al. Lactate dehydrogenase inhibition synergizes with IL-21 to promote CD8(+) T cell stemness and antitumor immunity. Proc Natl Acad Sci USA. 2020;117(11):6047–55.
Sukumar M, Liu J, Ji Y, Subramanian M, Crompton JG, Yu Z, et al. Inhibiting glycolytic metabolism enhances CD8 + T cell memory and antitumor function. J Clin Investig. 2013;123(10):4479–88.
Ma EH, Dahabieh MS, DeCamp LM, Kaymak I, Kitchen-Goosen SM, Oswald BM, et al. (13)C metabolite tracing reveals glutamine and acetate as critical in vivo fuels for CD8 T cells. Sci Adv. 2024;10(22):eadj1431.
Sinclair LV, Rolf J, Emslie E, Shi YB, Taylor PM, Cantrell DA. Control of amino-acid transport by antigen receptors coordinates the metabolic reprogramming essential for T cell differentiation. Nat Immunol. 2013;14(5):500–8.
Hosios AM, Hecht VC, Danai LV, Johnson MO, Rathmell JC, Steinhauser ML, et al. Amino acids rather than glucose account for the Majority of Cell Mass in proliferating mammalian cells. Dev Cell. 2016;36(5):540–9.
Sinclair LV, Neyens D, Ramsay G, Taylor PM, Cantrell DA. Single cell analysis of kynurenine and system L amino acid transport in T cells. Nat Commun. 2018;9(1):1981.
Frumento G, Rotondo R, Tonetti M, Damonte G, Benatti U, Ferrara GB. Tryptophan-derived catabolites are responsible for inhibition of T and natural killer cell proliferation induced by indoleamine 2,3-dioxygenase. J Exp Med. 2002;196(4):459–68.
Cronin SJF, Seehus C, Weidinger A, Talbot S, Reissig S, Seifert M, et al. The metabolite BH4 controls T cell proliferation in autoimmunity and cancer. Nature. 2018;563(7732):564–8.
Geiger R, Rieckmann JC, Wolf T, Basso C, Feng Y, Fuhrer T, et al. L-Arginine modulates T cell metabolism and enhances survival and anti-tumor activity. Cell. 2016;167(3):829–e4213.
Cao T, Zhang W, Wang Q, Wang C, Ma W, Zhang C, et al. Cancer SLC6A6-mediated taurine uptake transactivates immune checkpoint genes and induces exhaustion in CD8(+) T cells. Cell. 2024;187(9):2288–e30427.
Liu X, Cooper DE, Cluntun AA, Warmoes MO, Zhao S, Reid MA, et al. Acetate production from glucose and coupling to mitochondrial metabolism in mammals. Cell. 2018;175(2):502–e1313.
Mullen PJ, Yu R, Longo J, Archer MC, Penn LZ. The interplay between cell signalling and the mevalonate pathway in cancer. Nat Rev Cancer. 2016;16(11):718–31.
Peterson TR, Sengupta SS, Harris TE, Carmack AE, Kang SA, Balderas E, et al. mTOR complex 1 regulates lipin 1 localization to control the SREBP pathway. Cell. 2011;146(3):408–20.
Kidani Y, Elsaesser H, Hock MB, Vergnes L, Williams KJ, Argus JP, et al. Sterol regulatory element-binding proteins are essential for the metabolic programming of effector T cells and adaptive immunity. Nat Immunol. 2013;14(5):489–99.
Reinfeld BI, Madden MZ, Wolf MM, Chytil A, Bader JE, Patterson AR, et al. Cell-programmed nutrient partitioning in the tumour microenvironment. Nature. 2021;593(7858):282–8.
Hunt EG, Hurst KE, Riesenberg BP, Kennedy AS, Gandy EJ, Andrews AM, et al. Acetyl-CoA carboxylase obstructs CD8(+) T cell lipid utilization in the tumor microenvironment. Cell Metabol. 2024;36(5):969–e8310.
Park J, Kim M, Kang SG, Jannasch AH, Cooper B, Patterson J, et al. Short-chain fatty acids induce both effector and regulatory T cells by suppression of histone deacetylases and regulation of the mTOR-S6K pathway. Mucosal Immunol. 2015;8(1):80–93.
Bachem A, Makhlouf C, Binger KJ, de Souza DP, Tull D, Hochheiser K, et al. Microbiota-derived short-chain fatty acids promote the memory potential of Antigen-activated CD8(+) T cells. Immunity. 2019;51(2):285–e975.
Fan H, Xia S, Xiang J, Li Y, Ross MO, Lim SA, et al. Trans-vaccenic acid reprograms CD8(+) T cells and anti-tumour immunity. Nature. 2023;623(7989):1034–43.
Miller KD, O’Connor S, Pniewski KA, Kannan T, Acosta R, Mirji G, et al. Acetate acts as a metabolic immunomodulator by bolstering T-cell effector function and potentiating antitumor immunity in breast cancer. Nat cancer. 2023;4(10):1491–507.
Angela M, Endo Y, Asou HK, Yamamoto T, Tumes DJ, Tokuyama H, et al. Fatty acid metabolic reprogramming via mTOR-mediated inductions of PPARγ directs early activation of T cells. Nat Commun. 2016;7:13683.
Manzo T, Prentice BM, Anderson KG, Raman A, Schalck A, Codreanu GS et al. Accumulation of long-chain fatty acids in the tumor microenvironment drives dysfunction in intrapancreatic CD8 + T cells. J Exp Med. 2020;217(8).
Rivadeneira DB, DePeaux K, Wang Y, Kulkarni A, Tabib T, Menk AV, et al. Oncolytic viruses Engineered to enforce leptin expression Reprogram Tumor-infiltrating T cell metabolism and promote Tumor Clearance. Immunity. 2019;51(3):548–e604.
Lacher SB, Dörr J, de Almeida GP, Hönninger J, Bayerl F, Hirschberger A, et al. PGE(2) limits effector expansion of tumour-infiltrating stem-like CD8(+) T cells. Nature. 2024;629(8011):417–25.
Morotti M, Grimm AJ, Hope HC, Arnaud M, Desbuisson M, Rayroux N, et al. PGE(2) inhibits TIL expansion by disrupting IL-2 signalling and mitochondrial function. Nature. 2024;629(8011):426–34.
Ren Z, Wang K, Zhang Y, Chen H, Zhu Y, Li H, et al. Transient hydroxycholesterol treatment restrains TCR signaling to promote long-term immunity. Cell Chem Biology. 2024;31(5):920–e316.
Brown MS, Goldstein JL. The SREBP pathway: regulation of cholesterol metabolism by proteolysis of a membrane-bound transcription factor. Cell. 1997;89(3):331–40.
Zelcer N, Hong C, Boyadjian R, Tontonoz P. LXR regulates cholesterol uptake through Idol-dependent ubiquitination of the LDL receptor. Sci (New York NY). 2009;325(5936):100–4.
Yan C, Zheng L, Jiang S, Yang H, Guo J, Jiang LY, et al. Exhaustion-associated cholesterol deficiency dampens the cytotoxic arm of antitumor immunity. Cancer Cell. 2023;41(7):1276–e9311.
Ma X, Bi E, Lu Y, Su P, Huang C, Liu L, et al. Cholesterol induces CD8(+) T cell exhaustion in the Tumor Microenvironment. Cell Metabol. 2019;30(1):143–e565.
Wang F, Beck-García K, Zorzin C, Schamel WW, Davis MM. Inhibition of T cell receptor signaling by cholesterol sulfate, a naturally occurring derivative of membrane cholesterol. Nat Immunol. 2016;17(7):844–50.
Feske S, Giltnane J, Dolmetsch R, Staudt LM, Rao A. Gene regulation mediated by calcium signals in T lymphocytes. Nat Immunol. 2001;2(4):316–24.
Lyubchenko TA, Wurth GA, Zweifach A. Role of calcium influx in cytotoxic T lymphocyte lytic granule exocytosis during target cell killing. Immunity. 2001;15(5):847–59.
Maul-Pavicic A, Chiang SC, Rensing-Ehl A, Jessen B, Fauriat C, Wood SM, et al. ORAI1-mediated calcium influx is required for human cytotoxic lymphocyte degranulation and target cell lysis. Proc Natl Acad Sci USA. 2011;108(8):3324–9.
Vaeth M, Maus M, Klein-Hessling S, Freinkman E, Yang J, Eckstein M, et al. Store-operated ca(2+) entry controls clonal expansion of T cells through metabolic reprogramming. Immunity. 2017;47(4):664–e796.
Li FY, Chaigne-Delalande B, Kanellopoulou C, Davis JC, Matthews HF, Douek DC, et al. Second messenger role for Mg2 + revealed by human T-cell immunodeficiency. Nature. 2011;475(7357):471–6.
Eil R, Vodnala SK, Clever D, Klebanoff CA, Sukumar M, Pan JH, et al. Ionic immune suppression within the tumour microenvironment limits T cell effector function. Nature. 2016;537(7621):539–43.
Vodnala SK, Eil R, Kishton RJ, Sukumar M, Yamamoto TN, Ha NH, et al. T cell stemness and dysfunction in tumors are triggered by a common mechanism. Volume 363. New York, NY: Science; 2019. 6434.
Collier C, Wucherer K, McWhorter M, Jenkins C, Bartlett A, Roychoudhuri R et al. Intracellular K (+) limits T cell exhaustion and preserves antitumor function. bioRxiv: the preprint server for biology. 2023.
Dixon SJ, Lemberg KM, Lamprecht MR, Skouta R, Zaitsev EM, Gleason CE, et al. Ferroptosis: an iron-dependent form of nonapoptotic cell death. Cell. 2012;149(5):1060–72.
Drijvers JM, Gillis JE, Muijlwijk T, Nguyen TH, Gaudiano EF, Harris IS, et al. Pharmacologic screening identifies metabolic vulnerabilities of CD8(+) T cells. Cancer Immunol Res. 2021;9(2):184–99.
Li S, Ouyang X, Sun H, Jin J, Chen Y, Li L, et al. DEPDC5 protects CD8(+) T cells from ferroptosis by limiting mTORC1-mediated purine catabolism. Cell Discovery. 2024;10(1):53.
Ma J, Tang L, Tan Y, Xiao J, Wei K, Zhang X, et al. Lithium carbonate revitalizes tumor-reactive CD8(+) T cells by shunting lactic acid into mitochondria. Nat Immunol. 2024;25(3):552–61.
Korell F, Olson ML, Salas-Benito D, Leick MB, Larson RC, Bouffard A, et al. Comparative analysis of Bcl-2 family protein overexpression in CAR T cells alone and in combination with BH3 mimetics. Sci Transl Med. 2024;16(750):eadk7640.
Chan JD, Scheffler CM, Munoz I, Sek K, Lee JN, Huang YK, et al. FOXO1 enhances CAR T cell stemness, metabolic fitness and efficacy. Nature. 2024;629(8010):201–10.
Zhang DKY, Brockman JM, Adu-Berchie K, Liu Y, Binenbaum Y, de Lázaro I et al. Subcutaneous biodegradable scaffolds for restimulating the antitumour activity of pre-administered CAR-T cells. Nat Biomedical Eng. 2024.
Alan J., Korman Sarah C., Garrett-Thomson Nils, Lonberg (2022) The foundations of immune checkpoint blockade and the ipilimumab approval decennial. Nat Rev Drug Discovery. 21(7):509–528. https://doi.org/10.1038/s41573-021-00345-8.
Julia, Paik (2022) Nivolumab plus relatlimab: first approval. Drugs 82(8):925–931. https://doi.org/10.1007/s40265-022-01723-1.
Kumar J, Kumar R, Singh AK, Tsakem EL, Kathania M, Riese MJ, et al. Deletion of Cbl-b inhibits CD8 + T-cell exhaustion and promotes CAR T-cell function. J ImmunoTher Cancer. 2021;9(1):e001688. https://doi.org/10.1136/jitc-2020-001688.
Patel RP, Ghilardi G, Zhang Y, Chiang YH, Xie W, Guruprasad P, et al. CD5 deletion enhances the antitumor activity of adoptive T cell therapies. Sci Immunol. 2024;9(97):eadn6509. https://doi.org/10.1126/sciimmunol.adn6509.
Ta HM, Roy D, Zhang K, Alban T, Juric I, Dong J, et al. LRIG1 engages ligand VISTA and impairs tumor-specific CD8 + T cell responses. Sci Immunol. 2024;9(95):eadi7418. https://doi.org/10.1126/sciimmunol.adi7418.
Acknowledgements
Figures in the review were created with BioRender.com.
Funding
This work was funded by National Natural Science Foundation of China Grants 32270995 (to D.W.), and 32100985 (to D.W.), by Natural Science Foundation of Hunan Province (2023JJ10034, to D.W.), and by the Science and Technology Innovation Program of Hunan Province Grant 2022RC1209 (to D.W.). High-level Talent Project of Hunan Health Committee (20240304133, to D.W.), Scientific research key project of Hunan Education Department (21A0258), Natural Science Foundation of Hunan Province (Joint project of Science and Health) (No.2021JJ70039), Central Government Guided Local Science and Technology Development Fund Project in Xinjiang Uygur Autonomous Region, Scientific research Project of Health Commission of Hunan Province (No.20201064, 20201919), and the APC was funded by Hunan Provincial Clinical Medical Technology Innovation Guide Project (No.2020SK51817) and University of South China Clinical Research 4310 Program (20224310NHYCG01, 20214310NHYPY05), and University of South China Research Start-up Fund (to D.W.), and The First Affiliated Hospital of the University of South China Dual Professorship Fund (to D.W.).
Author information
Authors and Affiliations
Contributions
ZY wrote the original manuscript, YZ, CL, HJ, and HW designed the figures and tables, YZ, CD, GC and DW contributed to final editing of the work. All authors contributed to the original preparation of respective sections and revisions of the whole manuscript. All authors read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no potential conflicts of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Daichao Wu is Lead contact.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Yao, Z., Zeng, Y., Liu, C. et al. Focusing on CD8+ T-cell phenotypes: improving solid tumor therapy. J Exp Clin Cancer Res 43, 266 (2024). https://doi.org/10.1186/s13046-024-03195-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13046-024-03195-5