
Abstract
Background
Systemic lupus erythematosus (SLE) is an autoimmune disease characterized by a large production of autoantibodies and deficient clearance of cellular waste. The disease typically oscillates between episodes of elevated disease activity and quiescent disease. C-reactive protein (CRP) is a pentameric acute-phase protein usually reflecting inflammation and tissue damage. However, despite increased inflammation and elevated interleukin-6, the levels of CRP typically remain low or only slightly raised in SLE. Under certain conditions, pentameric CRP (pCRP) can dissociate into its monomeric isoform (mCRP), which mainly has been ascribed pro-inflammatory properties. The present study aims to investigate the potential relationship between pCRP and mCRP, respectively, with disease activity and clinical features of SLE.
Methods
The levels of pCRP and mCRP were measured, by turbidimetry (high-sensitive) and sandwich enzyme-linked immunosorbent assay (ELISA) respectively, in serum samples from 160 patients with SLE and 30 patients with antineutrophil cytoplasmic antibody-associated vasculitis (AAV). Twenty-two of the SLE cases were selected for analysis at two time-points; quiescent disease and active disease. The two CRP isoforms were evaluated in relation to disease activity and clinical features in the two diseases.
Results
Levels of pCRP and mCRP were significantly lower in SLE than AAV (p < 0.001) and the ratio of mCRP/pCRP was higher in SLE compared to AAV. The mCRP/pCRP ratio was higher for patients in remission and able to significantly separate between active/quiescent disease in paired, but not in non-paired, samples from patients with SLE. Significant correlations were observed with SLICC/ACR damage index for pCRP levels as well as inversely with the mCRP/pCRP ratio. Lower mCRP levels associated with malar rash.
Conclusion
As the interrelationship between the two isoforms appear to (a) discriminate between quiescent and active SLE and (b) differ between SLE and AAV, our data indicates that the two CRP isoforms could exert contrasting immunological effects and/or reflect different milieus. Given the biological effects of mCRP, it is possible that altered levels may indicate increased opsonization of immune complexes and apoptotic debris, and thereby prevent their deposition outside the reticuloendothelial system and manifestations such as lupus nephritis and lupus-related skin disease.
Similar content being viewed by others


Introduction
Systemic lupus erythematosus (SLE) is an autoimmune disease with highly heterogenous clinical presentation involving, e.g., skin, joints, and kidneys. Periods of raised disease activity may be followed by prolonged remission, and the disease severity ranges from mild skin and joint manifestations to life-threatening cytopenia, CNS disease, and/or renal involvement [1]. Autoantibody production, deposition of immune complexes (ICs), and complement activation are key components of the SLE pathogenesis. Clearance of dying cells and their constituents is normal and a strictly regulated part of the immune system’s homeostasis. The clearance is eventually carried out mainly by phagocytes, but opsonins, e.g., the acute-phase protein C-reactive protein (CRP), can facilitate this process. In SLE, a number of deficiencies in the waste disposal system has been found, including decreased phagocytic abilities of macrophages and polymorphonuclear cells [2]. Defective clearance is a source of autoantibody generation where the apoptotic cells and cellular debris fails to be eliminated properly, subsequently leading to chromatin and self-DNA being exposed for immune recognition [3]. These autoantibodies combined with soluble antigens form ICs that can accumulate in tissues, recruit complement, and ultimately lead to inflammation and tissue damage [4]. C1q is a complement protein which mitigates activation of the classical complement pathway, partly by CRP binding. Although extremely rare, homozygous deficiency of C1q is a strong risk factor of SLE, which implicates its importance in SLE pathogenesis and supports the waste disposal hypothesis. Furthermore, anti-C1q antibodies are a clinically valuable biomarker associated with SLE disease activity and lupus nephritis [5].
CRP is a highly conserved pattern recognition molecule, usually reflecting inflammation or tissue damage [6]. In contrast to many other inflammatory diseases, CRP appears to be an unreliable marker of raised disease activity in SLE. CRP is liver-derived, and its production is mainly regulated via interleukin-6 (IL-6), but genetic variants, e.g., the polymorphism rs1205, which is overrepresented in SLE, are also of importance for CRP levels [7]. Type I interferons can influence the levels of CRP in the circulation and may therefore be of interest when measuring CRP in serum [8]. However, type I interferons only partly explain the deviating levels of CRP in SLE. Structurally, CRP is a pentahomomeric protein with 5 Identical 206 amino acid subunits. Each subunit has a Ca2+-dependent phosphocholine binding site on one side and most likely binds C1q and Fcγ receptors on the other. CRP mediates a variety of pro- and anti-inflammatory biological effects, partly due to its ability to dissociate into a monomeric isoform (mCRP) [9]. CRP recognizes foreign pathogens and initiate clearance by binding to motifs such as phosphocholine, fibronectin, and chromatin, and activates the classical complement system via C1q [8, 10, 11]. The Fcγ receptor recognition aids in binding to Fcγ receptors and thereby activating phagocytotic cells and their clearance of foreign pathogens, apoptotic and damaged cells, and cellular waste [12]. Furthermore, CRP binds both apoptotic and necrotic cells, and low levels of CRP have been proposed to negatively affect the clearance of cellular debris [6, 13]. Since many of the observations do not pinpoint which of the CRP isoforms that are responsible for the certain effect, it is difficult to judge which isoform that associates with specific biological functions. In vitro, the pentameric CRP (pCRP) can dissociate into mCRP when exposed to denaturing conditions as occurs with exposure to membranes and lipoproteins, aqueous surface interfaces (as might occur with stirring and/or freeze/thaw cycles), or when exposed to heat or sufficient concentrations of urea [8, 14, 15]. In some of the above instances, calcium may regulate the dissociation of CRP into the mCRP isoform [16]. mCRP displays neoepitopes which are not exposed in the pentameric native form and asserts pro-inflammatory effector functions which are distinctly different from those of pCRP [17, 18], such as cytokine release, production of reactive oxygen species, and disrupting the alterative complement pathway through binding of lipid rafts and factor H [8, 19, 20]. We and others have shown that autoantibodies to mCRP are commonly found in SLE; especially associating with high disease activity and renal involvement [21, 22]. mCRP also plays an important role in waste disposal mechanisms and inflammatory processes by facilitating opsonization and activation of the classical complement pathway. Furthermore, mCRP has been shown to bind ICs at acidic pH [23], implicating its potential importance in IC-mediated diseases such as SLE. Other data indicate that mCRP in fluid phase (not membrane bound) may have a protective role against tissue damage inflicted by complement by binding of C1q, thereby hindering further activation of the classical complement pathway by C1q [20]. The same finding was obtained with high levels of pCRP (> 150 mg/L) in fluid phase [24]. However, mCRP bound to oxidized LDL has been shown to activate the classical complement pathway [25]. Furthermore, the levels of pCRP have been shown to associate with acquired organ damage in SLE, including the pulmonary and musculoskeletal systems [26]. In the general population, consistently elevated levels of CRP are associated with an increased risk of cardiovascular disease (CVD) such as myocardial infarction and stroke [27], and established CVD therapies has been shown to also lower the CRP concentration [28]. In addition, mCRP has been found in the arterial wall of normal human pulmonary tissue [29]. Although CVD continues to affect patients with SLE at young age, only few studies have been conducted to investigate the association of CVD and CRP specifically in SLE [8]. Recently, Wu et al. showed that the levels of mCRP were higher in patients with antineutrophil cytoplasmic antibody (ANCA)-associated vasculitis (AAV) than in healthy controls, and that these levels were related to CVD in the patients [30].
We hypothesized that inflammation per se may affect the ratios between the isoforms of CRP. Given the important biological function of CRP in relation to elimination of ICs, the present study focused on SLE. Thus, we aimed to evaluate the relationship between pCRP and mCRP and their reflection of disease activity and clinical features in cross-sectional and longitudinal samples from well-characterized patients with SLE. Samples from patients with AAV served as disease controls. To our knowledge, this is the first study to examine the relationship between mCRP and pCRP in SLE.
Patients and methods
Study population
The included patients with SLE (n = 160) were participants of a prospectively enrolling regional quality register entitled Clinical Lupus Register in North-Eastern Gothia (Swedish acronym “KLURING”) based at the University Hospital in Linköping [31]. All patients had a clinical diagnosis of SLE, where 138 (86%) fulfilled the 1982 American College of Rheumatology (ACR-82) classification criteria, and 160 (100%) met the 2012 Systemic Lupus International Collaboration Clinics (SLICC) criteria [32,33,34]. Disease activity was assessed using the SLE disease activity index 2000 (SLEDAI-2K) [35]. Irreversible organ damage was estimated by the SLICC/ACR damage index (SDI) [33]. Samples from approximately 50% of cases in the KLURING cohort were eligible for the present study to ascertain a broad range of disease activity. The characteristics are detailed in Table 1.
Twenty-two of the 160 patients with SLE were selected for paired analysis at two different time-points (visits); one sample with no clinical disease activity (SLEDAI-2K ≤ 4) and one sample with active disease (SLEDAI-2K ≥ 5) were analyzed. The median SLEDAI-2K (with IQR) values for each visit were 1 (0–2) and 12 (9–16), respectively.
AAV patients (n = 30), serving as disease controls, were included from the regional vasculitis register based at the University Hospital in Linköping [36]. The patients were recruited between years 2013 and 2020, had a clinical diagnosis of either microscopic polyangiitis (MPA) or granulomatosis with polyangiitis (GPA), and were classified according to the European Medicines Agency (EMA) algorithm [37]. Disease activity was assessed using the Birmingham Vasculitis Activity Score (BVAS) as shown in Table 1 [38].
For the mCRP assessment, 39 healthy controls (HC) with mean age 50 years (range 21–57), 33 females and 6 males, included in previous studies served as comparators [39,40,41,42]. For the anti-C1q antibody analyses, 100 anonymized blood donor sera from Uppsala University Hospital served as controls.
Detection of mCRP and pCRP
Serum concentrations of mCRP were measured using a sandwich enzyme-linked immunosorbent assay (ELISA). Immulon 2HB plates were coated overnight using a goat anti-human mCRP polyclonal antibody diluted in PBS (1:3000) and then blocked for 2 h at room temperature using a solution of 1% bovine serum albumin fraction V (BSA) (Sigma, St. Louis, MO, USA) in PBS-0.01% Tween-20 (blocking buffer). The wells were then washed with PBS-0.01% Tween-20 (washing buffer) and incubated with several concentrations of recombinant human mCRP (0.05–500 ng/mL; as described in Potempa et al. [43]) or serum samples diluted 1:5 in blocking buffer containing 1% normal goat serum for 2 h at room temperature. The wells were then washed three times with washing buffer and incubated with a mouse anti-human mCRP monoclonal antibody (8C10) diluted 1:100 in blocking buffer for 90 min at room temperature. Washing was repeated (3 ×), and the wells were then incubated with a 1:5000 dilution of a goat anti-mouse IgG antibody coupled to horseradish peroxidase (Abcam, Cambridge, UK) in blocking buffer. After a 1-h incubation at room temperature, the plates were washed (3 ×) and a substrate solution (3,3′,5,5′ tetramethylbenzidine [TMB], Sigma) was added to the wells and color allowed to develop. Reactions were stopped by addition of 1 M H2SO4 and optical density measured at 450 nm. Concentrations of mCRP in the samples were calculated based on the recombinant mCRP standard curve. Controls using purified pCRP at concentrations up to 50 µg/mL did not give any signal above background levels, showing specificity for mCRP.
pCRP in serum was measured using turbidimetry high-sensitive technique (detection limit 0.15 mg/L) at the routine laboratory, Clinical Chemistry laboratory of the University Hospital in Linköping. A cut-off level of 2.0 mg/L was applied according to the clinical routine for cardiovascular risk assessment [44].
Routine laboratory assessments
Laboratory analyses were carried out at the Clinical Chemistry unit, Linköping University Hospital or the Rudbeck Laboratory, Department of Immunology, Genetics and Pathology, Uppsala University, and included complement components (C3, C4, and C3d by nephelometry; classic hemolytic complement function), erythrocyte sedimentation rate (ESR), hemoglobin concentration, and blood cell counts (leukocytes, lymphocytes, neutrophils, basophils, monocytes, platelets). Urinalysis by urinary dipstick was assessed with regard to urinary erythrocytes (cells/µL) and urinary albumin (g/L). Renal function was measured by estimated glomerular filtration rate (eGFR) based on plasma creatinine [45]. Levels of myeloperoxidase (MPO)- and proteinase 3 (PR3)-ANCA were measured using sensitive fluorescence enzyme immunoassay (FEIA) [46]. Anti-C1q antibodies were analyzed using ELISA (Inova Diagnostics, San Diego, USA) [47].
The levels of C3, C4, and ESR were divided based on reference values from the accredited Clinical Chemistry unit, Linköping. C3 and C4 below 0.7 and 0.13 g/L, respectively, and ESR above 30 mm/h were considered subnormal/abnormal.
Ethical considerations
All included subjects had provided oral and written informed consent. The study protocol was approved by the Regional Ethics Review Board in Linköping regarding SLE (M75–08/2008) and AAV (2010/205–31).
Statistical analysis
The data was statistically analyzed using SPSS Statistics 26 (IBM Corp.; Armonk, NY, USA). The data was first tested for normality. Normally distributed data was thereafter examined for outliers. There were no data that fulfilled the requirements for parametrical testing. Testing between non-related groups was carried out using non-parametric Mann–Whitney U tests or Kruskal–Wallis test. Non-parametric correlation analyses were performed using Spearman’s rank correlation coefficient test. For comparisons between groups with paired data, Wilcoxon signed rank tests were used. For comparisons between binary data, exact χ2-test was carried out. Undetectable levels of mCRP were set to half the level of the detection limit (1.25 µg/L). A p-value of ≤ 0.05 was considered statistically significant.
Results
pCRP and mCRP
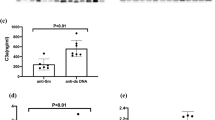
The median value (with IQR) for the levels of pCRP and mCRP in patients with SLE were 2.8 mg/L (1.3–8.7) and 0.0037 mg/L (0.0013–0.0074) respectively, and 26 mg/L (7.1–118) and 0.011 mg/L (0.0058–0.022) for AAV (Fig. 1). The levels of mCRP and pCRP did not correlate significantly with each other in either SLE (rho = − 0.002, p = 0.98) or AAV (rho = 0.30, p = 0.11).

The graphs show serum levels of pentameric C-reactive protein (pCRP; A) and monomeric CRP (mCRP; B), as well as in mCRP/pCRP ratios (C) in systemic lupus erythematosus (SLE; n = 160) and ANCA-associated vasculitis (AAV; n = 30). Panel D illustrates levels of autoantibodies against complement protein 1q (anti-C1q) in SLE as well as in healthy controls (HC; n = 100). In addition, panel B includes a group of healthy controls (HC; n = 39; B). The dotted line represents CRP cut-off level applied for cardiovascular risk assessment in clinical routine (2.0 mg/L; A) (* = p ≤ 0.05, *** = p < 0.001)
The subjects with SLE had lower levels of both CRP forms than the AAV patients (p < 0.001 for both comparisons). In addition, the ratio of mCRP/pCRP showed a significant difference between SLE and AAV (p < 0.01; Fig. 1), with mCRP/pCRP ratios showing median values (IQR) 3.7 × 10−3 (1.3 × 10−3–7.4 × 10−3) vs. 4.3 × 10−4 (1.2 × 10−4–1.2 × 10−3).
CRP in SLE
Among the 160 SLE cases, 65 (41%) had active disease (SLEDAI-2K ≥ 5) whereas 95 (59%) were in a quiescent phase of their disease. For paired samples, the ratios of mCRP/pCRP were lower in samples obtained from active compared to non-active disease (p ≤ 0.05). However, this comparison did not reach statistical significance in the cross-sectional cohort when samples were divided into active and non-active disease (p = 0.14). No significant differences were found regarding the levels of pCRP or mCRP between active disease and non-active disease neither for the paired nor the cross-sectional samples (Fig. 2).

Levels of pCRP (A, D) and mCRP (B, E) and ratios of mCRP/pCRP (C, F) between active and non-active systemic lupus erythematosus. Panels A, B, and C are based on 160 non-paired patient samples whereas D, E, and F represent paired samples from 22 patients (* = p ≤ 0.05)
Patients with normal and abnormal/subnormal levels of ESR, C3, and C4 based on reference intervals were separated into groups, and the levels of pCRP and mCRP as well as the ratios of mCRP/pCRP in each group were compared (Fig. 3). No significant differences were found for patients with normal or subnormal levels of C3 or C4. However, a highly significant difference (p < 0.001) was observed for both pCRP and the mCRP/pCRP ratios for patients with normal vs. abnormal ESR.

Comparisons of pCRP (A–D), mCRP (E–H), and mCRP/pCRP ratios (I–L) demonstrated between deviating levels of erythrocyte sedimentation rate (ESR), complement protein 3 (C3), C4, and negative/positive anti-C1q autoantibody test in the 160 patients with systemic lupus erythematosus. Abnormal ESR > 30 mm/h; subnormal C3 < 0.7 g/L; subnormal C4 < 0.13 g/L; positive anti-C1q > 20 units (*** = p < 0.001)
CRP in AAV
Significant differences in the levels of pCRP, mCRP, or the ratio of mCRP/pCRP between samples obtained from patients with MPA and GPA were not observed. Neither did any of the CRP measurements show significant correlations with disease activity assessed by BVAS. The levels of PR3-ANCA were inversely correlated with the mCRP/pCRP ratio (rho = − 0.42, p ≤ 0.05), but did not correlate significantly with pCRP or mCRP. MPO-ANCA levels correlated with the levels of mCRP (rho = 0.39, p ≤ 0.05), but not with pCRP or the mCRP/pCRP ratio. Furthermore, the mCRP/pCRP ratio correlated inversely with leukocyte count (rho = − 0.62, p < 0.001) and ESR (rho = − 0.76, p < 0.001, n = 19) (Fig. 4). Positive correlations were seen between pCRP levels and leukocyte count (rho = 0.46, p ≤ 0.05) as well as for ESR (rho = 0.84, p < 0.0001) (Fig. 4), but not with mCRP levels. In addition, mCRP levels correlated inversely with the age of the AAV patients (rho = − 0.38, p ≤ 0.05); this was not observed for pCRP levels, or the mCRP/pCRP ratios.

Significant correlations for both pCRP and the mCRP/pCRP ratio based on 160 patients with systemic lupus erythematosus (SLE; A) and 30 patients with ANCA-associated vasculitis (AAV; B). Black bars represent correlations with pCRP and white bars represent correlations with the mCRP/pCRP ratio. Left of midline represents inverse correlations whereas right of midline represents direct correlations. SDI, SLICC/ACR damage index; U-Ery, Urinary erythrocytes (SLE: n = 155); C3d, complement component 3d (n = 45); C4, complement protein 4 (n = 158); C3, complement protein 3 (n = 158); ESR, erythrocyte sedimentation rate (SLE: n = 155; AAV: n = 19); ns, not significant (* = p ≤ 0.05, ** = p < 0.01, *** = p < 0.001)
Associations between isoforms of CRP and damage accrual in SLE
The levels of pCRP and the mCRP/pCRP ratio correlated significantly with SDI score (rho = 0.33, p < 0.001; rho = − 0.23, p = < 0.01; Fig. 4). Significantly higher levels of pCRP were observed in patients with irreversible damage in the ocular (2.47 mg/L vs. 5.62, p ≤ 0.05), neuropsychiatric (2.25 mg/L vs. 6.34, p ≤ 0.001), peripheral vascular (2.47 mg/L vs. 7.01, p ≤ 0.05), and diabetes (2.67 mg/L vs. 8.41, p < 0.01) domains. The mCRP/pCRP ratio was significantly lower in patients with irreversible damage in the neuropsychiatric domain (1.49 × 10−3 vs. 4.16 × 10−4, p < 0.01). No significant change in mCRP levels were found for patients with any type of irreversible damage.
Associations between isoforms of CRP and clinical features
In patients with SLE, the levels of mCRP correlated inversely with disease duration (rho = − 0.31, p < 0.0001) but not with age. When the study population was separated into active/non-active disease, the inverse correlation remained in those with active disease (rho = − 0.49, p < 0.0001, n = 65) but not in the non-active. The levels of mCRP were significantly lower among patients meeting the ACR criterion for malar rash (p ≤ 0.05). Furthermore, pCRP were decreased in those meeting the criterion for photosensitivity (p ≤ 0.05); and higher in patients meeting the criterion for serositis (p ≤ 0.05; Fig. 5). Neither the mCRP/pCRP ratio, nor the levels of pCRP and mCRP, were associated with lupus nephritis (Supplementary Fig. 1). In addition, no significant associations between CRP isoforms and any ongoing medication were observed except for the prescribed corticosteroid dose, which significantly correlated with pCRP in SLE (rho = 0.26, p = 0.01), but not in AAV.

Comparisons of pCRP levels, mCRP levels, and mCRP/pCRP ratio between 160 patients with systemic lupus erythematosus meeting the ACR criteria for malar rash (A–C), photosensitivity (D–F) and serositis (G–I) compared to those that did not (* = p ≤ 0.05)
Associations between CRP isoforms and laboratory features
In SLE, levels of mCRP in the patient group in remission correlated inversely with classical complement function (rho = − 0.47, p ≤ 0.05, n = 22) and positively with basophil count (rho = 0.28, p < 0.01, n = 94). The pCRP levels were significantly higher in patients with anemia compared to those with hemoglobin concentrations within the reference limits (2.56 mg/L vs. 3.77, p ≤ 0.05), but no significant difference was detected for mCRP levels or the mCRP/pCRP ratio.
In AAV, there was no significant difference in pCRP levels or the mCRP/pCRP ratio between patients with anemia and those with hemoglobin concentration within the reference limit; however, significantly lower levels of mCRP were found in patients with anemia (0.024 mg/L vs. 0.0088, p ≤ 0.05). Differences in pCRP, mCRP, and mCRP/pCRP ratio were analyzed with regard to presence of proteinuria and hematuria, and with impaired renal function (eGFR < 60 mL/min/1.73 m2).
Significantly higher pCRP levels were found in SLE with hematuria compared to those without (p ≤ 0.05; Supplementary Fig. 1). The levels of mCRP were higher in AAV patients with albuminuria compared to those without (Supplementary Fig. 2). In addition, the levels of anti-C1q autoantibodies correlated weakly with the ratio of mCRP/pCRP (Fig. 4). No significant difference was found for anti-C1q autoantibody levels between SLE and healthy controls (Fig. 1D).
Discussion
Typically, pCRP levels remain low or only slightly elevated in SLE flares even if IL-6 is elevated, and these levels are often higher during infections than lupus flares [7, 8, 48]. Thus, pCRP is considered an unreliable marker of increased inflammation in SLE. To our knowledge, the present study is the first to evaluate the two isoforms of CRP in SLE.
In line with previous findings, we show that pCRP did not discriminate between active and non-active SLE. However, the mCRP/pCRP ratio met statistical significance in paired samples. Previously, it has been shown that mCRP possesses a number of distinct biological effects and can be of importance in relation to IC elimination via the reticuloendothelial system [23, 49]. The latter is of high relevance in SLE pathogenesis where extra-hepatic deposition of ICs is prevalent. Furthermore, it has been demonstrated that CRP together with C1q and dsDNA co-localize with IgG in electron-dense deposits of proliferative lupus nephritis [50], implicating that antibodies targeting dsDNA, CRP, and C1q are involved in the SLE pathogenesis. Presence of anti-CRP antibodies in SLE has been reported in varying frequencies by several groups [8]. These antibodies fluctuate over time but have consistently been found to associate with lupus nephritis and have the ability to bind apoptotic material and induce pro-inflammatory responses [21, 51, 52]. In addition, no significant correlation seem to exist between anti-CRP antibodies and CRP levels in serum [53]. Anti-mCRP antibodies have been shown to correlate with SLE disease activity [54]. Anti-CRP antibodies show a resemblance with anti-C1q antibodies, which bind to conformationally altered C1q and are strongly associated with lupus nephritis [5, 55]. The deposition of anti-C1q in glomeruli has previously been suggested to reveal neoepitopes not exposed elsewhere [5]. Prior observations of the antigen specificity have revealed that the anti-CRP antibodies found in SLE are directed towards mCRP, not pCRP, and that isolated ICs do not give rise to false positive anti-CRP [54, 56].
Given the recent publication by Wu et al. [30] on mCRP and pCRP in AAV, we decided to include a group of well-characterized patients with AAV as disease controls. Not surprisingly, the pCRP and mCRP levels in SLE were lower than in AAV. The ratio of mCRP/pCRP also differed, showing higher ratios in SLE compared to AAV. In contrast to Wu et al. [30], mCRP in our study did not correlate with BVAS in AAV patients (rho = 0.17, p = 0.37). However, in general, the patients with AAV included herein overall had less severe disease, which could indicate that the levels of mCRP is greater at even higher degree of disease activity than the patients in this study had. Comparisons between this study and the study by Wu et al. [30] shows that the level of mCRP in AAV differs greatly, where the patients herein had considerably lower levels. However, the mCRP levels in our study are more approximate to those reported by Chen et al. [57], and in line with other autoimmune disorders and with patients of high CRP [58,59,60]. The levels of mCRP were shown to correlate with age in AAV, while also inversely correlate with disease duration in SLE. Interestingly, this correlation was stronger in the SLE patients with an active disease profile, while the correlation in the patients without active disease was non-significant. The in-house ELISA used for detecting mCRP in serum displayed highly specific results using the 8C10 clone and has not been previously published (Supplementary Fig. 3).
Data on mCRP have been reported in AAV [30, 57], autoimmune dermatological disorders [60], bacterial infections [57, 59], CVD [58], healthy volunteers [30, 58, 60], and in a limited number of patients with SLE and rheumatoid arthritis [57]. Increased pCRP has been reported to associate with organ damage [26]. Similarly, in the present study we observed significant correlations between SDI and pCRP as well as with mCRP/pCRP ratio. Furthermore, SLE patients without malar rash had higher levels of mCRP than those meeting that ACR criterion. pCRP levels were higher in those with serositis, and both pCRP and mCRP were associated with skin manifestations (photosensitivity and malar rash). Patients with other autoimmune skin disorders have been reported to show elevated levels of mCRP compared to healthy controls [60]. Previously, mCRP has been shown to activate neutrophils, monocytes, and platelets [8, 61]. In the present study however, the levels of mCRP were not correlated with either neutrophil, monocyte, or platelet counts. Although the biological function of mCRP entails activation of the classical complement pathway [62], disruption of the alternative complement pathway [17], facilitation of opsonization [63], and activation of endothelial cells [17], in our hands, the levels of mCRP did not correlate significantly with the levels of complement proteins (C3, C4, or C3d).
When planning this study, we initially hypothesized that the mCRP/pCRP ratio could reflect disease activity or clinical features in SLE. Some associations were indeed observed, but the mCRP/pCRP ratio seems to predominantly be influenced by the levels of pCRP.
Conclusions
As the interrelationship between the two isoforms appear to (a) discriminate between quiescent and active SLE, and (b) differ between SLE and AAV, our data indicates that the two CRP isoforms could exert contrasting immunological effects and/or reflect different milieus. Given the biological effects of mCRP, it is possible that altered levels may indicate increased opsonization of ICs and apoptotic debris, and thereby preventing their deposition outside the reticuloendothelial system and manifestations such as lupus nephritis and lupus-related skin disease.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- SLE:
-
Systemic lupus erythematosus
- CRP:
-
C-reactive protein
- mCRP:
-
Monomeric C-reactive protein
- pCRP:
-
Pentameric C-reactive protein
- AAV:
-
Antineutrophil cytoplasmic antibody-associated vasculitis
- ICs:
-
Immune complexes
- IL-6:
-
Interleukin-6
- CVD:
-
Cardiovascular disease
- ANCA:
-
Antineutrophil cytoplasmic antibody
- KLURING:
-
Swedish acronym for Clinical Lupus Register in North-Eastern Gothia
- ACR-82:
-
1982 American College of Rheumatology classification criteria
- SLICC:
-
2012 Systemic Lupus International Collaboration Clinics criteria
- SLEDAI-2K:
-
SLE disease activity index 2000
- SDI:
-
SLICC/ACR damage index
- IQR:
-
Interquartile range
- MPA:
-
Microscopic polyangiitis
- GPA:
-
Granulomatosis with polyangiitis
- EMA:
-
European Medicines Agency
- BVAS:
-
Birmingham Vasculitis Activity Score
- HC:
-
Healthy controls
- ELISA:
-
Enzyme-linked immunosorbent assay
- BSA:
-
Bovine serum albumin
- TMB:
-
Tetramethylbenzidine
- C3:
-
Complement component 3
- C4:
-
Complement component 4
- C3d:
-
Complement component 3d
- C1q:
-
Complement component 1q
- ESR:
-
Erythrocyte sedimentation rate
- eGFR:
-
Estimated glomerular filtration rate
- MPO:
-
Myeloperoxidase
- PR3:
-
Proteinase 3
- FEIA:
-
Fluorescence enzyme immunoassay
- IgG:
-
Immunoglobulin G
References
Kaul A, Gordon C, Crow MK, Touma Z, Urowitz MB, van Vollenhoven R, Ruiz-Irastorza G, Hughes G. Systemic lupus erythematosus. Nat Rev Dis Primers. 2016;2:16039.
Munoz LE, Gaipl US, Franz S, Sheriff A, Voll RE, Kalden JR, Herrmann M. SLE–a disease of clearance deficiency? Rheumatology (Oxford). 2005;44(9):1101–7.
Pisetsky DS, Lipsky PE. New insights into the role of antinuclear antibodies in systemic lupus erythematosus. Nat Rev Rheumatol. 2020;16(10):565–79.
Hosszu KK, Valentino A, Peerschke EI, Ghebrehiwet B. SLE: novel postulates for therapeutic options. Front Immunol. 2020;11: 583853.
Trendelenburg M. Autoantibodies against complement component C1q in systemic lupus erythematosus. Clin Transl Immunology. 2021;10(4): e1279.
Vogt B, Fuhrnrohr B, Muller R, Sheriff A. CRP and the disposal of dying cells: consequences for systemic lupus erythematosus and rheumatoid arthritis. Autoimmunity. 2007;40(4):295–8.
Enocsson H, Gullstrand B, Eloranta ML, Wetterö J, Leonard D, Rönnblom L, Bengtsson AA, Sjöwall C. C-reactive protein levels in systemic lupus erythematosus are modulated by the interferon gene signature and CRP gene polymorphism rs1205. Front Immunol. 2020;11: 622326.
Enocsson H, Karlsson J, Li HY, Wu Y, Kushner I, Wetterö J, Sjöwall C. The complex role of C-reactive protein in systemic lupus erythematosus. J Clin Med. 2021;10(24):5837.
Sproston NR, Ashworth JJ. Role of C-reactive protein at sites of inflammation and infection. Front Immunol. 2018;9:754.
Du Clos TW, Zlock LT, Marnell L. Definition of a C-reactive protein binding determinant on histones. J Biol Chem. 1991;266(4):2167–71.
Suresh MV, Singh SK, Agrawal A. Interaction of calcium-bound C-reactive protein with fibronectin is controlled by pH: in vivo implications. J Biol Chem. 2004;279(50):52552–7.
Lu J, Mold C, Du Clos TW, Sun PD. Pentraxins and Fc receptor-mediated immune responses. Front Immunol. 2018;9:2607.
Sheriff A, Gaipl US, Voll RE, Kalden JR, Herrmann M. Apoptosis and systemic lupus erythematosus. Rheum Dis Clin North Am. 2004;30(3):505–27 viii-ix.
Yao Z, Zhang Y, Wu H. Regulation of C-reactive protein conformation in inflammation. Inflamm Res. 2019;68(10):815–23.
Ji SR, Wu Y, Potempa LA, Qiu Q, Zhao J. Interactions of C-reactive protein with low-density lipoproteins: implications for an active role of modified C-reactive protein in atherosclerosis. Int J Biochem Cell Biol. 2006;38(4):648–61.
Potempa LA, Maldonado BA, Laurent P, Zemel ES, Gewurz H. Antigenic, electrophoretic and binding alterations of human C-reactive protein modified selectively in the absence of calcium. Mol Immunol. 1983;20(11):1165–75.
Li HY, Wang J, Meng F, Jia ZK, Su Y, Bai QF, Lv LL, Ma FR, Potempa LA, Yan YB, et al. An intrinsically disordered motif mediates diverse actions of monomeric C-reactive protein. J Biol Chem. 2016;291(16):8795–804.
Ji SR, Wu Y, Zhu L, Potempa LA, Sheng FL, Lu W, Zhao J. Cell membranes and liposomes dissociate C-reactive protein (CRP) to form a new, biologically active structural intermediate: mCRP(m). FASEB J. 2007;21(1):284–94.
Li QY, Li HY, Fu G, Yu F, Wu Y, Zhao MH. Autoantibodies against C-reactive protein influence complement activation and clinical course in lupus nephritis. J Am Soc Nephrol. 2017;28(10):3044–54.
Wu Y, Potempa LA, El Kebir D, Filep JG. C-reactive protein and inflammation: conformational changes affect function. Biol Chem. 2015;396(11):1181–97.
Sjöwall C, Eriksson P, Almer S, Skogh T. Autoantibodies to C-reactive protein is a common finding in SLE, but not in primary Sjogren’s syndrome, rheumatoid arthritis or inflammatory bowel disease. J Autoimmun. 2002;19(3):155–60.
Sjöwall C, Zickert A, Skogh T, Wetterö J, Gunnarsson I. Serum levels of autoantibodies against C-reactive protein correlate with renal disease activity and response to therapy in lupus nephritis. Arthritis Res Ther. 2009;11(6):R188.
Motie M, Brockmeier S, Potempa LA. Binding of model soluble immune complexes to modified C-reactive protein. J Immunol. 1996;156(11):4435–41.
Sjöwall C, Wetterö J, Bengtsson T, Askendal A, Almroth G, Skogh T, Tengvall P. Solid-phase classical complement activation by C-reactive protein (CRP) is inhibited by fluid-phase CRP-C1q interaction. Biochem Biophys Res Commun. 2007;352(1):251–8.
Ji SR, Wu Y, Potempa LA, Liang YH, Zhao J. Effect of modified C-reactive protein on complement activation: a possible complement regulatory role of modified or monomeric C-reactive protein in atherosclerotic lesions. Arterioscler Thromb Vasc Biol. 2006;26(4):935–41.
Lee SS, Singh S, Link K, Petri M. High-sensitivity C-reactive protein as an associate of clinical subsets and organ damage in systemic lupus erythematosus. Semin Arthritis Rheum. 2008;38(1):41–54.
Labarrere CA, Zaloga GP. C-reactive protein: from innocent bystander to pivotal mediator of atherosclerosis. Am J Med. 2004;117(7):499–507.
de Ferranti SD, Rifai N. C-reactive protein: a nontraditional serum marker of cardiovascular risk. Cardiovasc Pathol. 2007;16(1):14–21.
Diehl EE, Haines GK 3rd, Radosevich JA, Potempa LA. Immunohistochemical localization of modified C-reactive protein antigen in normal vascular tissue. Am J Med Sci. 2000;319(2):79–83.
Wu KL, Liang QH, Huang BT, Ding N, Li BW, Hao J. The plasma level of mCRP is linked to cardiovascular disease in antineutrophil cytoplasmic antibody-associated vasculitis. Arthritis Res Ther. 2020;22(1):228.
Frodlund M, Dahlström O, Kastbom A, Skogh T, Sjöwall C. Associations between antinuclear antibody staining patterns and clinical features of systemic lupus erythematosus: analysis of a regional Swedish register. BMJ Open. 2013;3(10): e003608.
Tan EM, Cohen AS, Fries JF, Masi AT, McShane DJ, Rothfield NF, Schaller JG, Talal N, Winchester RJ. The 1982 revised criteria for the classification of systemic lupus erythematosus. Arthritis Rheum. 1982;25(11):1271–7.
Gladman D, Ginzler E, Goldsmith C, Fortin P, Liang M, Urowitz M, Bacon P, Bombardieri S, Hanly J, Hay E, et al. The development and initial validation of the Systemic Lupus International Collaborating Clinics/American College of Rheumatology damage index for systemic lupus erythematosus. Arthritis Rheum. 1996;39(3):363–9.
Ighe A, Dahlström O, Skogh T, Sjöwall C. Application of the 2012 Systemic Lupus International Collaborating Clinics classification criteria to patients in a regional Swedish systemic lupus erythematosus register. Arthritis Res Ther. 2015;17:3.
Gladman DD, Ibanez D, Urowitz MB. Systemic lupus erythematosus disease activity index 2000. J Rheumatol. 2002;29(2):288–91.
Eriksson P, Jacobsson L, Lindell A, Nilsson JA, Skogh T. Improved outcome in Wegener’s granulomatosis and microscopic polyangiitis? A retrospective analysis of 95 cases in two cohorts. J Intern Med. 2009;265(4):496–506.
Watts R, Lane S, Hanslik T, Hauser T, Hellmich B, Koldingsnes W, Mahr A, Segelmark M, Cohen-Tervaert JW, Scott D. Development and validation of a consensus methodology for the classification of the ANCA-associated vasculitides and polyarteritis nodosa for epidemiological studies. Ann Rheum Dis. 2007;66(2):222–7.
Mukhtyar C, Lee R, Brown D, Carruthers D, Dasgupta B, Dubey S, Flossmann O, Hall C, Hollywood J, Jayne D, et al. Modification and validation of the Birmingham Vasculitis Activity Score (version 3). Ann Rheum Dis. 2009;68(12):1827–32.
Fernandez-Botran R, Plankey MW, Ware D, Bordon J. Changes in liver steatosis in HIV-positive women are associated with the BMI, but not with biomarkers. Cytokine. 2021;144: 155573.
Fernandez-Botran R, Uriarte SM, Arnold FW, Rodriguez-Hernandez L, Rane MJ, Peyrani P, Wiemken T, Kelley R, Uppatla S, Cavallazzi R, et al. Contrasting inflammatory responses in severe and non-severe community-acquired pneumonia. Inflammation. 2014;37(4):1158–66.
Fernandez-Botran R, Vega AR, Garcia Y, Tirumala CC, Srisailam P, Raghuram A, Peyrani P, Furmanek S, Tella MA, Ritzhentaler JD, et al. The elevated systemic cytokine levels in HIV patients are not associated with an elevated pulmonary cytokine environment. Cytokine. 2020;126: 154874.
Newton TL, Fernandez-Botran R, Miller JJ, Lorenz DJ, Burns VE, Fleming KN. Markers of inflammation in midlife women with intimate partner violence histories. J Womens Health (Larchmt). 2011;20(12):1871–80.
Potempa LA, Yao ZY, Ji SR, Filep JG, Wu Y. Solubilization and purification of recombinant modified C-reactive protein from inclusion bodies using reversible anhydride modification. Biophys Rep. 2015;1:18–33.
Lindahl B, Toss H, Siegbahn A, Venge P, Wallentin L. Markers of myocardial damage and inflammation in relation to long-term mortality in unstable coronary artery disease. FRISC Study Group. Fragmin during Instability in coronary artery disease. N Engl J Med. 2000;343(16):1139–47.
Björk J, Grubb A, Sterner G, Nyman U. Revised equations for estimating glomerular filtration rate based on the Lund-Malmo study cohort. Scand J Clin Lab Invest. 2011;71(3):232–9.
Bossuyt X, Cohen Tervaert JW, Arimura Y, Blockmans D, Flores-Suarez LF, Guillevin L, Hellmich B, Jayne D, Jennette JC, Kallenberg CGM, et al. Position paper: Revised 2017 international consensus on testing of ANCAs in granulomatosis with polyangiitis and microscopic polyangiitis. Nat Rev Rheumatol. 2017;13(11):683–92.
Sjöwall C, Bentow C, Aure MA, Mahler M. Two-parametric immunological score development for assessing renal involvement and disease activity in systemic lupus erythematosus. J Immunol Res. 2018;2018:1294680.
Kruse K, Janko C, Urbonaviciute V, Mierke CT, Winkler TH, Voll RE, Schett G, Munoz LE, Herrmann M. Inefficient clearance of dying cells in patients with SLE: anti-dsDNA autoantibodies, MFG-E8, HMGB-1 and other players. Apoptosis. 2010;15(9):1098–113.
Skogh T, Stendahl O. Complement-mediated delay in immune complex clearance from the blood owing to reduced deposition outside the reticuloendothelial system. Immunology. 1983;49(1):53–9.
Sjöwall C, Olin AI, Skogh T, Wetterö J, Morgelin M, Nived O, Sturfelt G, Bengtsson AA. C-reactive protein, immunoglobulin G and complement co-localize in renal immune deposits of proliferative lupus nephritis. Autoimmunity. 2013;46(3):205–14.
Janko C, Franz S, Munoz LE, Siebig S, Winkler S, Schett G, Lauber K, Sheriff A, van der Vlag J, Herrmann M. CRP/anti-CRP antibodies assembly on the surfaces of cell remnants switches their phagocytic clearance toward inflammation. Front Immunol. 2011;2:70.
Pesickova SS, Rysava R, Lenicek M, Vitek L, Potlukova E, Hruskova Z, Jancova E, Honsova E, Zavada J, Trendelenburg M, et al. Prognostic value of anti-CRP antibodies in lupus nephritis in long-term follow-up. Arthritis Res Ther. 2015;17:371.
Rosenau BJ, Schur PH. Antibodies to C reactive protein. Ann Rheum Dis. 2006;65(5):674–6.
Sjöwall C, Bengtsson AA, Sturfelt G, Skogh T. Serum levels of autoantibodies against monomeric C-reactive protein are correlated with disease activity in systemic lupus erythematosus. Arthritis Res Ther. 2004;6(2):R87-94.
Tsacheva I, Radanova M, Todorova N, Argirova T, Kishore U. Detection of autoantibodies against the globular domain of human C1q in the sera of systemic lupus erythematosus patients. Mol Immunol. 2007;44(8):2147–51.
Mathsson L, Åhlin E, Sjöwall C, Skogh T, Rönnelid J. Cytokine induction by circulating immune complexes and signs of in-vivo complement activation in systemic lupus erythematosus are associated with the occurrence of anti-Sjogren’s syndrome A antibodies. Clin Exp Immunol. 2007;147(3):513–20.
Chen T, Xu PC, Gao S, Hu SY, Wei L, Yan TK. Monomeric C-reactive protein promotes platelets to release mitochondrial DNA in anti-neutrophil cytoplasmic antibody-associated vasculitis. Mol Immunol. 2021;137:228–37.
Wang J, Tang B, Liu X, Wu X, Wang H, Xu D, Guo Y. Increased monomeric CRP levels in acute myocardial infarction: a possible new and specific biomarker for diagnosis and severity assessment of disease. Atherosclerosis. 2015;239(2):343–9.
Williams RD, Moran JA, Fryer AA, Littlejohn JR, Williams HM, Greenhough TJ, Shrive AK. Monomeric C-reactive protein in serum with markedly elevated CRP levels shares common calcium-dependent ligand binding properties with an in vitro dissociated form of C-reactive protein. Front Immunol. 2020;11:115.
Zhang L, Li HY, Li W, Shen ZY, Wang YD, Ji SR, Wu Y. An ELISA assay for quantifying monomeric C-reactive protein in plasma. Front Immunol. 2018;9:511.
Potempa LA, Zeller JM, Fiedel BA, Kinoshita CM, Gewurz H. Stimulation of human neutrophils, monocytes, and platelets by modified C-reactive protein (CRP) expressing a neoantigenic specificity. Inflammation. 1988;12(4):391–405.
Khreiss T, Jozsef L, Potempa LA, Filep JG. Conformational rearrangement in C-reactive protein is required for proinflammatory actions on human endothelial cells. Circulation. 2004;109(16):2016–22.
Khreiss T, Jozsef L, Hossain S, Chan JS, Potempa LA, Filep JG. Loss of pentameric symmetry of C-reactive protein is associated with delayed apoptosis of human neutrophils. J Biol Chem. 2002;277(43):40775–81.
Acknowledgements
The authors wish to thank Marianne Petersson for biobank administration, Dr. Lawrence A. Potempa and Dr. Ibraheem Rajab for providing the monoclonal detection antibody 8C10 and for valuable comments on the manuscript, Dr. Helena Enocsson, for valuable comments on the manuscript, Lars Valter for statistical advice, and all the patients and clinicians at the Rheumatology Unit, University Hospital in Linköping, for their efforts.
Funding
Open access funding provided by Linköping University. This work was supported by grants from the Swedish Rheumatism Association, the Region Östergötland (ALF Grants), the Gustafsson Foundation, the King Gustaf V’s 80-year Anniversary foundation and the King Gustaf V and Queen Victoria’s Freemasons foundation.
Author information
Authors and Affiliations
Contributions
JK performed the statistical analysis, contributed to study design, interpretation of data, and manuscript writing. JW contributed to study design, interpretation of data, and writing of the manuscript. MW contributed to AAV data and manuscript writing. JR contributed to the anti-C1q analysis and manuscript writing. RFB performed the mCRP ELISA and contributed to manuscript writing. CS contributed to the original idea, study design, interpretation of data, and manuscript writing. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
This study was carried out in accordance with the Declaration of Helsinki. Oral and written informed consent was obtained from all participants. The study protocol was approved by the Regional Ethics Review Board in Linköping, Sweden for SLE (Decision number M75–08) and for AAV (2010/205–31).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1:
Supplementary Figure 1. Comparisons of pentameric C-reactive protein (pCRP) (A–D), monomeric (m)CRP (E–H), and mCRP/pCRP ratio (I–L) in 160 patients with systemic lupus erythematosus with lupus nephritis vs. no nephritis (A, E, I), normal vs. subnormal estimated glomerular filtration rate (eGFR) (B, F, J), presence vs. absence of hematuria (C, G, K) and presence vs. absence of albuminuria (D, H, L). Abnormal eGFR <60 mL/min/1.73m2. (* = p≤0.05).
Additional file 2:
Supplementary Figure 2. Comparisons of pentameric C-reactive protein (pCRP) (A–C), monomeric (m)CRP (D–F), and mCRP/pCRP ratio (G–I) demonstrated between normal vs. subnormal estimated glomerular filtration rate (eGFR) (A, D, G), presence vs. absence of hematuria (B, E, H) and presence vs. absence albuminuria (C, F, I) in patients with ANCA-associated vasculitis. Abnormal eGFR <60 mL/min/1.73m2. (* = p≤0.05).
Additional file 3:
Supplementary Figure 3. Standard curves and reactivity of the monoclonal detection antibody 8C10 towards monomeric C-reactive protein (mCRP) and pentameric (p)CRP: (A) standard curve for 8C10 antibody with mCRP; (B) reactivity of 8C10 antibody with pCRP; and (C) standard curve for samples in the present study.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Karlsson, J., Wetterö, J., Weiner, M. et al. Associations of C-reactive protein isoforms with systemic lupus erythematosus phenotypes and disease activity. Arthritis Res Ther 24, 139 (2022). https://doi.org/10.1186/s13075-022-02831-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13075-022-02831-9