
Abstract
Background
Fruits of Vitis vinifera L., commonly known as grapes, are largely consumed worldwide because of their high nutritional and medicinal benefits.
Context and purpose
The present study investigated effects of V. vinifera fruits in ovalbumin-induced animal model of bronchial asthma.
Methods
Male wistar rats (except group 1) were sensitized with allergen (ovalbumin, 40 mg/rat + aluminum hydroxide, 2 mg/rat). Groups of sensitized animals were treated orally with either vehicle (0.4 mL/kg), standard dexamethasone (2.5 mg/kg) or alcoholic extract of V. vinifera dried fruits (31 and 42.5 mg/kg) from day 1 to 28 (n = 6 for all groups). Inflammatory markers including cell counts, cytokines such as interleukin (IL)-4, IL-5, IL-1β, tumor necrosis factor, immunoglobulin E (IgE), leukotrienes and nitrite levels in both blood/serum and bronchoalveolar fluid were analysed. Breathing rate and tidal volume as lung function parameters were examined by spirometer. Lung tissues were studied for histamine content and histopathology.
Results
Treatment of sensitized animals with dexamethasone or two doses of V. vinifera fruits extract inhibited recruitment of inflammatory cytokines, IgE, nitrites and circulating cells particularly eosinophils in blood/serum and bronchoalveolar fluid (p < 0.001, p < 0.01 and p < 0.05). Dexamethasone and V. vinifera fruits extract treatment also normalized lung functions and histamine levels compared to ovalbumin-sensitized controls (p < 0.05 and p < 0.01). Moreover, both drugs exhibited protection against airway inflammation in lung histology.
Conclusion
Results of study demonstrate the effectiveness of V. vinifera fruits in allergic asthma possibly related to its ability to inhibit cellular response and subsequent production of inflammatory cytokines.
Similar content being viewed by others
Background
Vitis vinifera (Family: Vitaceae), commonly known as grapes, are widely utilized as natural dietary supplements due to their unique phytochemical composition and high nutritional value. Fruits are a good source of polyphenols [1], anthocyanins [2], flavanols [3], stilbenes (resveratrol) [4], phenolic acids, proteins, fats, vitamins (C and A) [5], minerals (calcium, boron phosphorus) [6], water, carbohydrates and fibers [7]. The medicinal value of the plant has long been recognized in folklore medicine. Documented evidences report anti-diabetic, cytotoxic [8], anti-aging [9], cardioprotective [10], hypolipdemic [11], anti-inflammatory [12] and antioxidant [13] properties of seeds and fruits of V. vinifera.
Regarding inflammatory process in chronic airway diseases such as asthma and chronic obstructive pulmonary disorder, disease pathology is directly associated with increased generation of reactive oxygen species in the lungs [14]. Several clinical studies suggest that supplementation of antioxidants benefit adults with mild to moderate asthma [15, 16]. Since, oxidative damage plays a significant role in the pathology of bronchial asthma therefore this process may represent a potential target of the management therapy in asthmatic patients [17].
Asthma is a chronic inflammatory disease of airways characterized by reversible constriction of the tracheobronchial tree and airway hyperresponsiveness to various stimuli such as environmental allergens, respiratory infection, cold air, exercise and some drugs [18, 19]. In human beings, allergic asthma is primarily initiated by a type I hypersensitivity reaction which represents increased susceptibility to produce immunoglobulin E (IgE) in response to external allergens, secretion and differentiation of which is dependent on CD4+ helper T cells (TH2-type) [20, 21]. Activated TH2 cells produce a number of cytokines such as interleukin (IL)-3, IL-4, IL-5, IL-13 which in turn play role in various process such as promoting production of IgE cells by B cells, growth of mast cells (IL-4) and survival of eosinophils by IL-5 [22]. IL-4 and IL-13 also stimulate epithelial cells to produce transforming growth factor alpha (TGF-α) which causes mucosal metaplasia and fibroblast proliferation [23]. Pro-inflammatory mediators including tumor necrosis factor (TNF) and granulocyte macrophage colony-stimulating factor (GM-CSF) stimulate the expression of vascular adhesion molecules on endothelial cells which result in an increased influx of inflammatory leukocytes into the bronchial tree [24].
With regard to presence of rich plethora of phenolic antioxidant constituents such as gallic acid and resveratrol in V. vinifera, the present study was conducted to examine therapeutic potential of dried fruits of V. vinifera in ovalbumin induced rat model of allergic asthma.
Methods
Drugs and chemicals
Ovalbumin (OVA), dexamethasone, gallic acid, heparin, methacholine, vecuronium bromide were purchased from Sigma Aldrich, St. Louis, MO, USA. Standard ELISA kits used for the determination of rat interleukin (IL)-4, tumor necrosis factor (TNF), IL-1β (Ray Biotech, Inc., IL, USA), IgE (Immunology Consultants Laboratory, Inc., Portland, OR), IL-5 and leukotrienes LTD4 (Cusabio Biotech, Hubei, China), Nitric oxide (NO) calorimetric kit (BioVision Research Products, USA) were purchased from commercial suppliers. All other chemicals were commercial products of analytical reagent grade.
Collection of plant and preparation of extract
Vitis vinifera L. dried fruits were collected from a local Indian supplier and botanically authenticated by Dr. H. B. Singh at National Institute of Science Communication and Information Resources, India. A sample voucher NISCAIR/RHMD/consult/-2011-12/1752/52) was submitted in herbarium of Jamia Hamdard, Hamdard University, New Delhi, India, for future reference. Dried fruits (1000 g) were homogenized and exhaustively extracted with ethanol for 3 days at 32 ± 2 °C. The extract was separated by filtration and concentrated in rotary vacuum evaporator (Buchi, USA) and then dried in lyophilizer (Uni-step, India, model: PPI –SX72) under reduced pressure. The yield obtained was 426.44 g of viscous dark brown residue (yield 42.64 % w/w). 250 mg extract was dissolved in purified water with the help of carboxymethyl cellulose (0.1 %). The prepared suspension of alcoholic extract of V. vinifera (VVHE) dried fruits was stored in refrigerator till administration to animals.
Quantification of gallic acid by HPTLC
Preparation of sample and standards
10 mg of residue was dissolved in 1 mL of ethanol to obtain the concentration of 10 mg/mL. Stock solutions of gallic acid were prepared by dissolving 1 mg of gallic acid in 1 mL of ethanol and making dilutions to get the final concentration, 100 µg/mL of standard. Sample and standard solutions were filtered through a 0.22 µM membrane filter.
Chromatographic conditions
For quantifying gallic acid in ethanol extract of V. vinifera dried fruits, mobile phase with composition toluene: ethyl acetate: formic acid: methanol (3.5:3.5:0.8:0.5) and chamber saturation time 35 min was used. The standard and sample were spotted in the form of bands (width 8 mm with a CAMAG microliter syringe) on a pre-coated silica gel plate 60F-254 aluminum sheets (20 × 10 cm with 0.2 mm thickness; Merck KGaA, Darmstadt, Germany) using a CAMAG Linomat-V applicator (Muttenz, Switzerland) attached to CAMAG HPTLC system. The plates were pre-washed with methanol and activated at 105 °C for 5 min prior to chromatography. The standard and sample-loaded plates were kept in the TLC twin trough developing chamber (after saturating with solvent vapor for 15 min at 28 ± 2 °C) with respective mobile phase till plate run up to 80 mm. The developed plates were dried in hot air to evaporate solvents and scanned at 292 nm with CAMAG TLC densitometric scanner 3 operated by WinCATS software, using deuterium lamp. To quantify gallic acid 10 µl/spot of sample solution was applied on HPTLC plates.
Calibration curves for standards
From the stock solution of standard gallic acid (100 μg/mL) different volumes 1, 2, 4, 6, 8 and 10 μl, were spotted on a precoated TLC plate to obtain corresponding concentrations of 100, 200, 400, 600 and 800 and 1000 ng/spot of standard. Each application was done in triplicate. The regression equation for gallic acid was y = (1216 ± 2.863) + (6.484 ± 0.007)x; (Co-relation coefficient, r2 = 0.996 ± 0.0084).
Animals
The guidelines of Government of India regarding control and supervision of experiments on animals were followed. Approval was obtained from the Institutional Animal Ethics Committee (IAEC) before conducting the present study (Registration no. 173/CPCSEA/748). Male wistar rats (180–220 g; 8–10 weeks old) were procured from Central Animal House, Jamia Hamdard, New Delhi. Animals were housed in polypropylene cages under controlled conditions (room temperature 25 ± 2 °C; humidity 45 ± 5 %; and photoperiod of 12 h light: dark cycle). All the animals had free access to standard laboratory conditions of diet and water ad libitum throughout the study.
Experimental design
After acclimatization of male wistar rats to the standard housing conditions, animals were randomly divided into 5 groups (6 rats per group). Group 1 (SAL), the non-sensitized control, received vehicle (0.4 mL/kg); Group 2 (OVA), the OVA-sensitized or asthma control, was OVA-sensitized receiving vehicle only; Group 3 (OVA + dex), reference standard was OVA-sensitized received dexamethasone (2.5 mg/kg, b.w.); Group 4 and 5, the experimental groups (OVA + VVHE 1 and OVA + VVHE 2) were OVA-sensitized and received alcoholic extract of VVHE (31 and 42.5 mg/kg, b.w., respectively). Vitis vinifera fruits are an important constituent of various herbal formulations mentioned in Ayurvedic Pharmacopoeia of India (API). In view of this, dose of extract was calculated from human dose of drug mentioned in API [25]. Drugs or vehicle were administered orally from day 1 to 28 once daily in the morning hours.
Asthma was induced in rats using the method of Abdureyim et al., with some modifications (2011) [26]. All animals (except Group 1) were sensitized with intraperitoneal injection of allergen suspension (ovalbumin, 40 mg/rat + aluminum hydroxide, 2 mg/rat) on day 1. After 15 days of sensitization, animals were challenged by exposure to aerosol consisting of 1 % ovalbumin in normal saline for 20 min. Animals in the non-sensitized group were exposed to saline following the same protocol. Exposure to aerosolized solutions were done in a closed chamber (dimensions 40 × 32 × 32 cm) once daily for 8 consecutive days i.e., from day 15 to day 22, and thereafter, on day 25 and day 28.
Lung function and bronchoconstriction test
On day 28, 5 min after OVA exposure, rats were anaesthetized with an intraperitoneal injection of sodium pentobarbitone (105 mg/kg). Trachea was cannulated using a 12G cannula (2 mm internal diameter) as described previously [27]. Cannula was connected to a pneumotachograph (spirometer, Model no: FE141, ADInstruments, Pty, Australia) with attached flow head (Model: MLT 10L, 10 L/min, suitable for rats, ADInstruments, Pty, Australia) and differential pressure transducer to measure respiratory rate (f, breaths/min). Lungs tidal volume (VT, mL/s) were obtained by electronic integration of airflow signal. Data was stored in Power Lab System (ADInstruments, Pty, Australia) and acquired through LabChart Programme installed in the lab’s computer. Spirometer was calibrated before use to record data in terms of L/s and any drift or offsets in signal due to transducer was nullified to zero for accurate measurements. Femoral vein was cannulated with a 24G needle cannula filled with heparin for administration of methacholine (1.5 mg/kg). Changes in f and VT were recorded (1) before and after vehicle administration; (2) before and after methacholine treatment. 10–12 respiratory cycles were averaged to provide one data point. Vecuronium bromide (0.2 mg/kg) was injected intravenously to avoid incidence of spontaneous respiration. Excessive bronchial secretions were discharged using a small polyethylene tube without disturbing the trachea [28].
Bronchoalveolar lavage (BAL) fluid collection
Immediately after measurement of lung function parameters, lungs were lavaged for 3 times with 5 mL (5 mL × 3) aliquots of 0.9 % sterile saline solution via cannulated tracheal tube [29]. The BAL fluid recovered from each rat was pooled separately (approximately 11–12 mL/rat) and centrifuged (1500 rpm, 10 min at 4 °C). Supernatant was separated and stored at −80 °C until analysis for IgE and cytokines, whereas cell pellet was re-suspended in 1 mL of physiological saline for determination of total and differential leukocyte count as per method described by Jung et al. [30].
Serum preparation and cell count
Following collection of BAL fluid, blood was collected by cardiac puncture in two different portions. The first aliquot (2.5–3 mL) was collected in a non-heparinized tube; centrifuged (3000 rpm, 10 min) and serum stored at −80 °C for estimation of IgE and cytokines. The second aliquot (0.5 mL) was collected in a heparinized tube and stored at 4 °C until determination of total cell and differential count.
Within 30 min of collection of heparinized blood, total leukocyte count and differential cell count in blood was determined by autoanlyser (XP-100, Sysmex Corporation, Japan). Differential cell count was made from cytospin smears stained with Leishman’s stain (1.5 % in methanol for 6 min). A minimum of 500 cells were counted under an optical digital microscope (B1 series system, 400× magnification) and classified into eosinophils, neutrophils, lymphocytes, macrophages or monocytes using standard morphologic criterion. Absolute number of each cell type was calculated.
IgE, LTD4, cytokines, nitric oxide and nitrite levels in serum and BAL fluid
The levels of IgE, LTD4 and cytokines (TNF, IL-4, IL-5 and IL-1β) in serum (500 μl) and BAL fluid (5 mL) were measured using enzyme-linked immunosorbent assay (ELISA) kits. The samples were analyzed on an automated ELISA plate reader (Model no. ELX-80MS, Biotek, USA). Concentration of total NO and nitrite levels in serum and BAL fluid were determined by nitric oxide calorimetric kit.
Histamine analysis in lavaged lung tissue
After collection of BAL fluid lung tissue lobes from each animal were separately dissected out. One of the lobes was homogenized with 2.5 mL normal saline and processed for analysis of histamine as per method described by Shore et al., using spectrophotometer at 650 nm [30, 31].
Histological examination
Dissected lung tissues were washed with normal saline (NS) and fixed in 10 % neutral formaldehyde solution at 4 °C for 24 h. The specimens were embedded in paraffin wax and sectioned to 5 µm with a rotary microtome. Sections were stained with 1 % hematoxylin in distilled water and 1 % eosin dye in 90 % alcohol (H & E) for studying morphology. Tissues were mounted with glycerin jelly and cover slipped. The slides were examined under Labcon trinocular research microscope and photographs were taken with a Nikon Coolpix digital zoom camera (model no. S3300).
Ethics aspects
The research was approved by Institutional Animal Ethics Committee (IAEC) before conducting the present study under Registration no. 173/CPCSEA/748.
Statistical analysis
Results were reported as mean ± SEM. Statistical analyses were performed using one-way analysis of variance (ANOVA) followed by post hoc Tukey’s test. Differences were considered statistically significant at value of p < 0.05. All statistical analyses were performed using the Graph Pad software (San Diego, CA, USA).
Results
Quantification of gallic acid by HPTLC in V. vinifera dried fruits
The finger print profile of ethanol extract of V. vinifera dried fruits in developed solvent system is shown in Additional file 1: Figure S1. The ethanolic extract of V. vinifera showed 8 peaks where fifth peak at R f values 0.43 ± 0.027 with corresponding area of 2943.73 ± 4.542 coincides with standard gallic acid The mean content of gallic acid was calculated to be 2.66 ± 1.12 mg/gm of VVHE.
The developed analytical method was optimized by applying BBD-RSM (Box-Behnken Design-Response Surface Methodology). Analysis of variance for isolation and quantification of gallic acid confirmed that the analytical model was statistically significant with F-value 32.55 and depicted by the value of Prob > F less than 0.05. Quadratic equation generated from experimental parameters (Additional file 1: Table S1) showed a reasonable agreement of predicted R2 (0.8418) with the adjusted R2 (0.9404) (Additional file 1: Figures S2, S3) given by the following equations:
where, A, is amount of toluene (mL); B, is amount of ethyl acetate (mL); C, amount of formic acid (mL); and D, is time for saturation of developing chamber (min).
Effect of VVHE on lung function parameters
After methacholine injection, respiration rate was significantly (p < 0.001) increased in OVA-sensitized control group as compared to non-sensitized control group. Animals receiving VVHE 1 (31 mg/kg) or VVHE 2 (42.5 mg/kg), exhibited 13 or 21 % decrease (p < 0.05 or p < 0.01) in respiratory rate as compared to the OVA-sensitized control group. There was a considerable reduction in tidal volume (p < 0.001) in OVA-sensitized control animals as compared to non-sensitized control. Treating asthmatic rats with VVHE 2 (42.5 mg/kg, b.w.) markedly increased (p < 0.05) tidal volume by 11 % as compared to the OVA-sensitized control group ( Fig. 1a, b). Similar comparisons for dexamethasone treated animals exhibited improvement (p < 0.001) in breathing rate and tidal volume by 37 and 28 %, respectively.

Effect of VVHE on breathing rate (a) and tidal volume (b) in OVA-sensitized rats. Values shown are mean ± SEM. (n = 6); # p < 0.05, ### p < 0.001 and ns (non-significant) compared to the non-sensitized control (SAL); **p < 0.01 and ***p < 0.001, compared to the OVA-sensitized control (OVA). SAL non-sensitized and vehicle treated control group, OVA OVA-sensitized and vehicle treated control group, OVA + dex OVA-sensitized and dexamethasone, 2.5 mg/kg, b.w, treated group, OVA + VVHE 1 OVA-sensitized and VVHE, 31 mg/kg, treated group, OVA + VVHE 2 OVA-sensitized and VVHE, 42.5 mg/kg, b.w. treated group
Effect of VVHE on circulating cell count in blood
The total number of circulating leukocytes, eosinophils, and neutrophils in blood samples of OVA-control rats were markedly increased (p < 0.001) compared to non-sensitized control animals. In contrast, allergen sensitized and challenged rats showed lesser number of lymphocytes in blood compared to normal group animals. Elevated number of eosinophils and neutrophils significantly reduced after treatment with VVHE 2 (42.5 mg/kg, b.w., p < 0.05) and dexamethasone (p < 0.01) compared to asthma control group. Both drug treatments also normalized the lymphocytes count in blood of animals (Table 1).
Effect of VVHE on inflammatory cellular counts in BAL fluid
The OVA-sensitized control group showed marked increase (p < 0.001) in total cells and differential cellular count in BAL fluid samples compared to non-sensitized control group. Number of eosinophils (p < 0.01), lymphocytes (p < 0.01), macrophages (p < 0.01) and neutrophils (p < 0.05) reduced significantly in VVHE 2 (42.5 mg/kg, b.w.) and dexamethasone (p < 0.001) treated animals compared to OVA-sensitized controls. VVHE at lower dose level (31 mg/kg) also reduced these inflammatory cells in lavaged fluid but the reduction was statistically significant only for lymphocytes and macrophages (p < 0.05) (Table 2).
Effect of VVHE on levels of LTD4 and cytokines in serum
Significant (p < 0.001) elevation in serum levels of LTD4 and all the cytokines, IL-4, IL-5, TNF and IL-1β was observed in OVA-sensitized control rats compared to the non-sensitized group controls. VVHE 1 (31 mg/kg, b.w.) and VVHE 2 (42.5 mg/kg, b.w.) treatment reduced serum levels of IL-4 by 17 and 24.2 % (p < 0.01), IL-5 by 17.1 and 28.2 % (p < 0.01), TNF by 17 and 30.2 % (p < 0.001), IL-1β by 5 and 15.3 % (p > 0.05 or p < 0.05) and LTD4 by 4.3 and 11 % (p > 0.05 or p < 0.05) compared to the OVA-sensitized control group (Fig. 2a). However, treating OVA-sensitized animals with dexamethasone showed significant decrease (p < 0.001) in serum levels of LTD4 by 44.7 % and all the cytokines tested in the study, IL-4 by 56.6 %, IL-5 by 47.6 %, TNF by 53.3 % and IL-1β by 57.11 %.

Effect of VVHE on cytokine levels in serum (a) and BAL fluid (b) of OVA-sensitized rats. Values shown are mean ± S.E.M. (n = 6); ### p < 0.001 compared to the non-sensitized control (SAL); *p < 0.05, **p < 0.01, ***p < 0.001, and ns (non-significant) compared to the OVA-sensitized control (OVA). SAL non-sensitized and vehicle treated control group, OVA OVA-sensitized and vehicle treated control group, OVA, OVA + dex OVA-sensitized and dexamethasone, 2.5 mg/kg, b.w, treated group, OVA + VVHE 1 OVA-sensitized and VVHE, 31 mg/kg, treated group, OVA + VVHE 2 OVA-sensitized and VVHE, 42.5 mg/kg, b.w. treated group
Effect of VVHE on levels of LTD4 and cytokines in BAL fluid
The levels of all the cytokines tested in BAL fluid of OVA-control group were significantly high (p < 0.001) compared to non-sensitized group animals. Substantial reduction in levels of cytokines by 30.5 % for IL-4 (p < 0.01); 23.2 % for IL-5 (p < 0.01); 10.6 % for IL-1β (p < 0.05); 27.3 % for TNF (p < 0.01) and 12.6 % for LTD4 (p < 0.05) was observed in VVHE 2 (42.5 mg/kg, b.w.) and dexamethasone (43.5-52.1 %, p < 0.001) treated groups. Treatment with VVHE 1, lower dose of the extract (31 mg/kg, b.w.) also attenuated all the cytokines and LTD4 levels in BAL fluid but inhibition was statistically significant only for IL-4, IL-5 and TNF (p < 0.05) (Fig. 2b). In contrast, animals treated with dexamethasone showed significant (p < 0.001) reduction in BAL fluid levels of LTD4 by 44.2 % and all the cytokines including IL-4 by 52.1 %, IL-5 by 43.5 %, TNF by 52.04 % and IL-1β by 50.5 % compared to OVA-sensitized control group.
Effect of VVHE on IgE levels of serum and BAL fluid
Allergen sensitization and challenge resulted in significant rise (p < 0.001) in IgE levels of both serum and BAL fluid samples of rats compared with non-sensitized control group. Compared to OVA-control group, animals receiving VVHE 1, 31 mg/kg, b.w., or VVHE 2, 42.5 mg/kg, b.w., exhibited substantial reduction by 17 % or 31 % in serum (p < 0.01) and 14 % or 26 % in BAL fluid (p < 0.01). Treatment with dexamethasone also caused significant (p < 0.001) reduction of IgE levels by 58 % in serum and 47.8 % in BAL fluid tested in the study compared to OVA-control group (Fig. 3a, b).

Effect of VVHE on IgE levels in serum (a) and BALF (b) of OVA-sensitized rats. Values shown are mean ± SEM. (n = 6); ### p < 0.001 compared to the non-sensitized control (SAL); *p < 0.05, **p < 0.01 and ***p < 0.001, compared to the OVA-sensitized control (OVA). SAL non-sensitized and vehicle treated control group, OVA OVA-sensitized and vehicle treated control group, OVA, OVA + dex OVA-sensitized and dexamethasone, 2.5 mg/kg, b.w, treated group, OVA + VVHE 1 OVA-sensitized and VVHE, 31 mg/kg, treated group, OVA + VVHE 2 OVA-sensitized and VVHE, 42.5 mg/kg, b.w. treated group
Effect of VVHE on total nitric oxide and nitrite concentration
A marked (p < 0.001) elevation of total NO and nitrite levels were observed in OVA-sensitized control animals as compared to non-sensitized control group. Compared with the asthmatic group animals receiving VVHE 2 (42.5 mg/kg, b.w.) showed 29.7 % and 31.4 % reduced levels of both nitric oxide and nitrites, respectively, in serum and 15 % and 27 % in BAL fluid samples (p < 0.01) (Fig. 4). Lower dose of extract (VVHE 1) also decreased (p < 0.05) levels of these analytes in both the body fluids. Treatment with reference drug, dexamethasone reduced (p < 0.001) elevated serum levels of total NO by 48.8 % and nitrites by 51.5 % and BAL fluids levels by 51 % and 55.6 %, respectively, compared to OVA-sensitized control animals.

Effect of VVHE on nitric oxide (NO) and nitrite level in serum and BALF of ovalbumin-sensitized rats. Values shown are mean ± S.E.M. (n = 6); ### p < 0.001 compared to the non-sensitized control (SAL); *p < 0.05, **p < 0.01 and ***p < 0.001, compared to the OVA-sensitized control (OVA). SAL non-sensitized and vehicle treated control group, OVA OVA-sensitized and vehicle treated control group, OVA, OVA + dex OVA-sensitized and dexamethasone, 2.5 mg/kg, b.w, treated group, OVA + VVHE 1 OVA-sensitized and VVHE, 31 mg/kg, treated group, OVA + VVHE 2 OVA-sensitized and VVHE, 42.5 mg/kg, b.w. treated group
Effect of VVHE on histamine levels in lung tissues
Analysis of histamine levels in lung tissues homogenates of OVA-control rats were significantly high (p < 0.001) compared to non-sensitized controls. VVHE at both dose levels, 31 and 42.5 mg/kg, reduced elevated histamine levels by 25 and 41.6 % (p < 0.01) compared with the OVA-control group (Fig. 5). Whereas treatment with standard dexamethasone caused reduction in histamine levels of lung tissue homogenates by 66.6 % compared to that of OVA-control group (p < 0.001).

Effect of treatments on lung tissue histamine levels in ovalbumin-sensitized rats. Values shown are mean ± SEM. (n = 6); ### p < 0.001 compared to the non-sensitized control (SAL); *p < 0.05, **p < 0.01 and ***p < 0.001, compared to the OVA-sensitized control (OVA). SAL non-sensitized and vehicle treated control group. OVA OVA-sensitized and vehicle treated control group, OVA, OVA + dex OVA-sensitized and dexamethasone, 2.5 mg/kg, b.w, treated group, OVA + VVHE 1 OVA-sensitized and VVHE, 31 mg/kg, treated group, OVA + VVHE 2 OVA-sensitized and VVHE, 42.5 mg/kg, b.w. treated group
Effect of treatments on histopathology of lung tissues
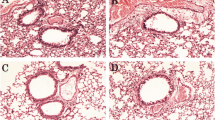
The histological examination of lung tissues from the OVA-control group showed reduced bronchiolar lumen, infiltration of inflammatory cells into the peribronchial tissues and epithelial desquamation. Treatment with VVHE and standard reference drug, dexamethasone elicited protection against all these pathological features as evidenced by the improvement in lumen size and reduced cellular infiltration (Fig. 6).

Effect of treatments on the histopathological changes in rat lung tissues. Representative hematoxylin and eosin-stained sections of the rat lungs (×10). SAL non-sensitized and vehicle treated control rat showing normal lung histology, OVA OVA-sensitized and vehicle treated control rat showing altered bronchiolar lumen and cellular infiltration, OVA, OVA + dex OVA-sensitized and dexamethasone, 2.5 mg/kg, b.w, treated rat, OVA + VVHE 1 OVA-sensitized and VVHE, 31 mg/kg, treated rat, OVA + VVHE 2 OVA-sensitized and VVHE, 42.5 mg/kg, b.w. treated rat
Discussion
Despite all the recent developments, use of herbal drugs is increasing as an adjunctive therapy to the conventional standard treatments in several chronic diseases such as asthma. These health supplements possess a number of medicinal properties which need to be investigated by adapting a translational approach using modern experimental tools. Ovalbumin-induced rat model of asthma is widely used for investigating anti-asthmatic potential of drugs in pre-clinical experimental trials [32, 33]. The main characteristic feature of this model is bronchial hyperresponsiveness to specific stimuli accompanied with airway inflammation and constriction.
In our study, ovalbumin sensitization of rats showed significant increase in breathing rate followed by reduction in tidal volume indicating features of broncho-constriction. Furthermore, all the animals in the asthma control group exhibited sneezing, hyperrhinorrhea and irritability. Protection against methacholine-induced bronchoconstriction in VVHE and dexamethasone treated animals demonstrated the bronchodilatory effect of the drugs. Amelioration of lung functions after treatment with VVHE may be due to reduced airway inflammation resulting in decrease in lung resistance to air flow. Airway hyperresponsiveness with clinical symptoms of high breathing rate and reduced tidal volume is considered as a primary target in the treatment of asthma [34].
Allergen-induced airway hyperresponsiveness is directly associated with TH2-driven eosniphilic airway inflammation [35]. Ovalbumin challenge of sensitized rats elicited an increase in number of total cells, eosinophils, and neutrophils in both the body fluids (BAL fluid and serum) compared to that of non-sensitized control group with exception of lymphocyte count in blood. The results were consistent with the study of Schster and group which explained decrease in lymphocytes number may be due to migration of these cells from blood to bronchial fluid after allergen provocation [36]. Oral treatment with VVHE (42.5 mg/kg b.w.) and dexamethasone for 28 days significantly reversed the OVA-induced infiltration of all the inflammatory cells particularly eosinophils, into rat airways, implying the possible role of extract in allergen mediated eosniphilic interventions.
TH2 derived inflammatory and pro-inflammatory cytokines such as IL-4, IL-5, TNF and IL-1β are extracellular signaling proteins secreted by almost every cell under certain conditions. These mediators play a critical role in orchestrating all types of inflammatory responses in asthmatic airways [37]. Pharmacological therapies using specific receptors IL-4, IL-5, TNF and IL-1β blockers are likely to constitute a considerable development in asthma management [38, 39]. Present study showed presence of prominent TH2 type cytokines in OVA-control group indicating persistent airway inflammation. VVHE at both dose levels 31 and 42.5 mg/kg b.w. for 28 days inhibited elevated levels of IL-4, IL-5, TNF and IL-1β in both serum and BAL fluid compared to OVA-control group. However, changes induced by VVHE were less than that observed in corticosteroid dexamethasone (2.5 mg/kg oral, for 28 days) treated group for inflammation markers in present experiment. Our findings with VVHE suggest that the V. vinifera fruits could act in asthma via its neutralizing effects on TH2 derived pro-inflammatory (TNF and IL-1β) and inflammatory (IL-4 and IL-5) cytokines, key elements in the pathophysiology of bronchial asthma. Cysteinyl leukotrienes (cys LT’s), LTB4, LTC4, LTD4, and LTE4, represent a heterogeneous group of biologically active mediators produced from arachidonic acid. Cys LT’s - D4 and E4 are stable potent end products which account for clinical features of asthma [40]. VVHE (42.5 mg/kg b.w p.o.) elicited substantial inhibitory effects on cytokine LTD4 in both serum and BAL fluid compared to OVA-control animals which may also add to bronchodilation effects of the VVHE in the study.
Increased production of immunoglobulin (IgE) in allergic asthma (atopy) is the strongest detectable predisposing factor in the development of bronchial asthma [41]. The anti-inflammatory effects of VVHE in asthmatic airways were confirmed in our study while evaluating its effect on serum and BAL fluid IgE levels in OVA-sensitized and challenged rats. At both dose levels, VVHE (31 and 42.5 mg/kg, p.o.) was found to be effective in inhibiting the IgE levels in body fluids tested. Marked reduction in levels of both LTD4 and IgE was also observed in dexamethasone (2.5 mg/kg oral, for 28 days) treated animals.
NO is a gaseous free radical molecule that is formed by a wide range of cells, including airway and vascular smooth muscle cells, endothelial cells, and epithelial cells nerves, activated macrophages. Inflammatory cells from asthma patients produce more reactive radical species including nitrites than those obtained from normal subjects [42] which perpetuate the ongoing airway inflammation. Some studies report that pH in the asthmatic airways falls during acute condition which facilitates the conversion of nitrite to NO. Hence, increased NO concentrations in the exhaled air of asthmatic patients may reflect nitrite conversion rather than iNOS activity [43, 44]. In the present study while evaluating the effects of VVHE on nitric oxide metabolites in serum and BAL fluid of asthmatic rats, VVHE and dexamethasone treatments expressed significant suppression of both NO and nitrite levels in these body fluids after 28 days treatment of OVA-sensitized rats. The data suggests that reduced levels of TH2 derived cytokines particularly IL-4 and IL-5 would had inhibited respective secretion of IgE by B cells and infiltration of eosinophils and other inflammatory cells into rat airways leading to suppression of airway inflammation and radical production in lungs.
Polyphenolic compounds present in natural drugs have been reported to elicit potent anti-oxidant and anti-inflammatory properties mediated through inhibition of IL-1β, TNF, COX-2 and production of NO2 and PGE2 [45–47]. Among them, gallic acid (3, 4, 5-trihydroxy benzoic acid) is one of the most important plant secondary metabolite present in numerous plants including Vitis vinifera L. The phytoconstituent is reported to exhibit various biological activities such as antioxidant, anti-inflammatory, anti-diabetic, anti-cancer, anti-microbial, anti-aging, cardio-protective, liver fibrosis [48–52]. In experimental studies on different models of allergic diseases, gallic acid is demonstrated to suppress of allergen induced hypersensitivity reactions in mice and inhibit release of histamine and helper T cell subtypes, IL-4, IL-5 and IL-2 form human mast cells [49, 53, 54]. Accumulating all these data i.e., results of our study on V. vinifera fruits and previous findings with gallic acid suggest that amelioration of allergic consequences associated with inflammatory processes in asthmatic rats treated with VVHE, in the present study, may be attributed to the presence of significant amount bioactive constituents including gallic acid in V. vinifera fruits.
Conclusion
From the results of the study, we may suggest that VVHE might play an important role in the management of bronchial asthma by (1) offering protection against ongoing inflammatory process underlying asthma through inhibition of histamine release, cytokine production; (2) improving lung functioning by counteracting allergen induced bronchial hyperresponsiveness; and (3) blocking the release of inflammatory cellular infiltration (eosinophils, lymphocytes, neutrophils) into airways; Further studies may be advised to confirm its use as a valuable anti-asthmatic drug in human beings.
References
Hertog MGL, Hollman PCH, Van de Putte B. Content of potentially anticarcinogenic flavonoids of tea infusions, wines, and fruit juices. J Agric Food Chem. 1993;41:1242–6.
Wang L, Waltenberger B, Pferschy-Wenzig EM, Blunder M, Liu X, Malainer C, Blazevic T, Schwaiger S. Natural product agonists of peroxisome proliferator-activated receptor gamma (PPARγ): a review. Biochem Pharmacol. 2014;55(1):73–89.
El-Ashmawy IM, Saleh A, Salama OM. Effects of marjoram volatile oil and grape seed extract on ethanol toxicity in male rats. Basic Clin Pharmacol Toxicol. 2007;101:320–7.
Rockenbach II, Rodrigues E, Gonzaga LV, Caliari V, Genovese MI, Gonckaves AE, Fett R. Phenolic compounds content and antioxidant activity in pomace from selected red grapes (Vitis vinifera L. and Vitis labrusca L.) widely produced in Brazil. Food Chem. 2011;127(1):174–9.
Sousa EC, Thomaz AM, Carioca JOB, Morais SM, Lima AD, Martins CG, Cristiane DACD, Ferreira PAT, Rodrigues ALM, Rodrigues SP, Silva JN, Rodrigues LL. Chemical composition and bioactive compounds of grape pomace (Vitis vinifera L.), Benitaka variety, grown in the semiarid region of Northeast Brazil. Food Sci Technol Campinas. 2014;34(1):135–42.
Yadav M, Jain S, Bhardwaj A, Nagpal R, Puniya M, Tomar R, Singh V, Parkash O, Prasad G, Marotta F, Yadav H. Biological and medicinal properties of grapes and their bioactive constituents: an update. J Med Food. 2009;12(3):473–84.
Camire ME, Dougherty MP. Raisin dietary fiber composition and in vitro bile acid binding. J Agric Food Chem. 2003;51:834–7.
Barthomeuf C, Lamy S, Blanchette M, Boivin D, Gingras D, Béliveau R. Inhibition of sphingosine-1-phosphate- and vascular endothelial growth factor-induced endothelial cell chemotaxis by red grape skin polyphenols correlates with a decrease in early platelet-activating factor synthesis. Free Rad Biol Med. 2006;40:581–90.
Khan GM, Ansari SH, Bhat ZA, Ahmad F. Study of aging and hepatoprotective activity of Vitis vinifera L. seeds in albino rats. Asian Pac J Trop Med. 2012; S1770–S1774.
Cui J, Cordis GA, Tosaki A, Maulik N, Das DK. Reduction of myocardial ischemia reperfusion injury with regular consumption of grapes. Ann NY Acad Sci. 2002;957:302–7.
Masani YA, Mathew N, Chakraborty M, Kamath JV. Effects of Vitis vinifera against Trition-X 100 induced hyperlipidaemia in rats. Inter Res J Pharm. 2012;3(12):101–3.
Heba H, Hanafi R, Islam E, Mohamed E, Ahmed K, Michael L, Lalia M, Nahla A. Antiinflamamtory and cytooxic activites of dietary polyphenolics isolated from Corchorous olitorius and Vitis vinefera. J Funct Food. 2013;2(3):1204–16.
Karthikeyan JR, Manikandan M. Evaluating the Antibacterial Potential of Vitis vinifera and Punica granatum fruit peels. BMR Complement Altern Med. 2014;1(1):1–6.
Shokry DM, El-Tarahony SE. Oxidant-antioxidant balance in childhood asthma. Egypt J Pediatr Allergy Immunol. 2013;11(1):35–40.
Ochs-Balcom HM, Grant BJB, Muti PP, Sempos CT, Freudenheim JL, Browne RW, McCa SE, Trevisan M, Cassano PA, Iacoviello L, Schunemann HJ. Antioxidants, oxidative stress, and pulmonary function in individuals diagnosed with asthma or COPD. Eur J Clin Nutr. 2006;60:991–9.
Toan NV, Hanh TT. Retraction: improved treatment of Asthma by using natural sources of antioxidants. Springer Plus. 2014;3:558.
Dittrich AM, Meyer HA, Krokowski M, Quarcoo D, Ahrens B, Kube SM, Witzenrath M, Esworthy RS, Chu FF, Hamelmann E. Glutathione peroxidase-2 protects from allergen-induced airway inflammation in mice. Eur Respir J. 2009;35(5):1148–54.
Roh SS, Kim SH, Lee YC, Seo YB. Effects of radix adenophorae and cyclosporine A on an OVA-induced murine model of asthma by suppressing to T cells activity, eosinophilia, and bronchial hyper-responsiveness. Mediat Inflamm. 2008;781425:1–11.
Global Initiative for Asthma (GINA). Global strategy for asthma management and prevention. 2015. www.ginasthma.org.
Abul K, Abbas MD. Disease of immunity. In: Kumar V, Abbas AK, Aster JC, editors. Robbins and Cotran pathologic basis of disease. 7th ed. Philadelphia: Elsevier Saunders; 2005. p. 194–268.
Platts-Mills TAE. The role of immunoglobulin E in allergy and asthma. Am J Respir Crit Care Med. 2001;164:S1–5.
Bjermer L, Diamant Z. Current and emerging nonsteroidal anti-inflammatory therapies targeting specific mechanisms in asthma and allergy. Treat Respir Med. 2004;3(4):235–46.
Nakae S, Ho LH, Yu M. Mast cell-derived TNF-α contributes to airway hyperreactivity, inflammation, and Th2 cytokine production in an asthma model in mice. J Allergy Clin Immunol. 2007;120:48.
Zeiger RS, Schatz M, Li Q. High blood eosinophil count is a risk factor for future asthma exacerbations in adult persistent asthma. J Allergy Clin Immunol Pract. 2014;2(6):741–50.
Anonymous: Ayurvedic Pharmacopoeia of India. Ministry of Health and Family Welfare, Government of India. The Controller of Publications, Civil Lines, Delhi, India, Part II, vol. II, (formulations) 1st English ed. 2008, p. 31–2.
Abdureyim S, Amat N, Umar A, Upur H, Berke B, Moore N. Anti-inflammatory, immunomodulatory, and hemeoxygenase-1 inhibitory activities of Ravan Napas, a formulation of Uighur traditional medicine, in a rat model of allergic asthma. Evid Based Complement Alternat Med. 2011;725926:1–14.
Parasuraman S, Raveendran R. Measurement of invasive blood pressure in rats. J Pharmacol Pharmacother. 2012;3(2):172–8.
Prado CM, Leick-Maldonado EA, Kasahara DI, Capelozzi VL. Effects of acute and chronic nitric oxide inhibition in an experimental model of chronic pulmonary allergic inflammation in guinea pigs. Am J Physiol Lung Cell Mol Physiol. 2005;289:L677–83.
Huang TJ, Adcock IM, Chung KF. A novel transcription factor inhibitor, SP [100030], inhibits cytokine gene expression, but not airway eosinophilia or hyperresponsiveness in sensitized and allergen exposed rat. Br J Pharmacol. 2001;134:1029–36.
Jung CH, Lee JY, Cho CH, Kim CJ. Anti-asthmatic action of Quercetin and Rutin in conscious guinea-pigs challenged with aerosolized ovalbumin. Arch Pharm Res. 2007;30(12):1599–607.
Shore PA, Burkhalter A, Cohn VH. A method for fluorometric assay of histamine in tissues. J Pharmcol Exp Ther. 1959;127:182–6.
Zosky GR, Sly PD. Animal models of asthma. Clin Exp Allergy. 2007;37:973–88.
Canning BJ, Chou Y. Using guinea pigs in studies relevant to asthma and COPD. Pulm Pharmacol Ther. 2008;21(5):702–20.
Mahajan SG, Mehta AA. Suppression of ovalbumin-induced Th2-driven airway inflammation by β-sitosterol in a guinea pig model of asthma. Eur J Pharmacol. 2011;650:458–64.
Tulic MK, Christodoulopoulos P, Hamid Q. Small airway inflammation in asthma. Respir Res. 2001;2(6):333–9.
Schster M, Tschering T, Krug N. Lymphocytes migrate from the blood into the bronchoalveolar lavage and lung parenchyma in the asthma model of the Brown Norway rat. Am J Respir Crit Care Med. 2000;161:558–66.
Cai1 Y, Cao Y, Lu S. Infliximab alleviates inflammation and ex vivo airway hyperreactivity in asthmatic E3 rats. Int Immunol. 2011; 1–9. doi:10.1093/intimm/dxr032.
Nag SS, Xu LJ, Hamid Q. The effects of IL-5 on airway physiology and inflammation in rats. J Allergy Clin Immunol. 2003;1(3):558–66.
Kouro T, Takatsu K. IL-5- and eosinophil-mediated inflammation: from discovery to therapy. Int Immunol. 2009;21(12):1303–9.
Cohen P, Noveral JP, Bhala A. Leukotriene D4 facilitates airway smooth muscle cell proliferation via modulation of the IGF axis. Am J Physiol. 1995;269:L151–7.
Hamelmann E. The rationale for treating allergic asthma with anti-IgE. Eur Resp Rev. 2007;16(104):61–6.
Cho YS, Moon H. The role of oxidative stress in the pathogenesis of asthma. Allergy Asthma Immunol Res. 2010;2(3):183–7.
Kharitonov SA, Barnes PJ. Clinical aspects of exhaled nitric oxide. Eur Respir J. 2000;16:781–92.
Ricciardolo FL, Sterk PJ, Gaston B. Nitric oxide in health and disease of the respiratory system. Physiol Rev. 2004;84:731–65.
Na HJ, Lee G, Oh HY, Jeon KS, Kwon HJ, Ha KS, Lee H, Kwon YG, Kim YM. 4-O-Methyl gallic acid suppresses inflammation-associated gene expression by inhibition of redox-based NF-kappa B activation. Int Immunopharmacol. 2006;6:1597–608.
Kang MS, Jang HS, Oh JS, Yang KH, Choi NK, Lim HS, Kim SM. Effects of methyl gallate and gallic acid on the production of inflammatory mediators interleukin-6, interleukin-8 by oral epithelial cells stimulated with Fusobaterium nucleatum. J Microbiol. 2009;47:760–7.
Kuppan G, Balasubramanyam J, Monickaraj F, Srinivasan G, Mohan V, Balasubramanyam M. Transcritptional regulation of cytokines and oxidative stress by gallic acid in human THP-1 monocytes. Cytokines. 2010;49:229–34.
Yeh RD, Chen JC, Lai TY, Yang JS, Yu CS, Chiang JH, Lu CC, Yang ST, Yu CC, Chang SJ, Lin HY, Chung JG. Gallic acid induces G0/G1 phase arrest and apoptosis in human leukemia HL-60 cells through inhibiting cyclin D and E, and activating mitochondria-dependent pathway. Anticancer Res. 2011;31:2821–32.
Vo TS, Ngo DH, Kim SK. Gallic acid-grafted chitooligosaccharides suppress antigen-induced allergic reactions in RBL-2H3 mast cells. Eur J Pharm Sci. 2012;47:527–33.
Nabavi SF, Habtemariam S, Sureda A, Moghaddam AH, Daglia M, Nabavi SM. In vivo protective effects of gallic acid isolated from Peltiphyllum Peltatum against sodium fluoride-induced oxidative stress in rat erythrocytes. Arh Hig Rada Toksikol. 2013;64:553–9.
Chang YJ, Hsu SL, Liu YT, Lin YH, Lin MH, Huang SJ, Ho JA, Wu LC. Gallic acid induces Necroptosis via TNF-α signalling pathway in activated hepatic stellate cells. PLoS ONE. 2015;10(3):e0120713. doi:10.1371/journal.pone.0120713.
Nayeem N, Asdaq SMB, Salem H. AHEI-Alfqv S. Gallic acid: a promising lead molecule for drug development. J App Pharm. 2016;8:2.
Kato K, Yamashita S, Kitanaka S, Toyoshima S. Effect of gallic acid derivatives on secretion of Th1 cytokines and Th2 cytokines from anti CD3-stimulated spleen cells. Yakugaku Zasshi. 2001;121:451–7.
Kim SH, Jun CD, Suk K, Choi BJ, Lim H, Park S, Lee SH, Shin HY, Kim DK, Shin TY. Gallic acid inhibits histamine release and pro-inflammatory cytokine production in mast cells. Toxicol Sci. 2006;91:123–31.
Authors’ contributions
PA (AB, ES); SHA (ES); AKN (FG); VA (FG), SA (FG). All authors read and approved the final manuscript.
Acknowledgements
Authors would like to express their gratitude for funding the current research, Project Number 45/48/2011/BMS/TRM, dated, 28-05-2013.
Competing interests
The authors declare that they have no competing interests.
Availability of data and materials
Data may be shared but not copied due to related ongoing research projects in our lab.
Funding
Notice for SRF, 45/48/2011/BMS/TRM, dated 28-05-2013. Indian Council of Medical Research (ICMR), India.
Author information
Authors and Affiliations
Corresponding author
Additional file
13223_2016_145_MOESM1_ESM.docx
Additional file 1: Table S1. Factors and levels of the Box-Behnken experimental plan. Figure S1. HPTLC chromatogram of gallic acid (A) standard and (B) ethanol extract of Vitis vinifera L. dried fruits. Figure S2. Diagnostic plot representing observed (actual) response values versus the predicted response values in terms of quantification of gallic acid in VVHE. Figure S3. Response surface plots representing effects of different independent variables on isolated concentration of gallic acid in VVHE.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Arora, P., Ansari, S.H., Najmi, A.K. et al. Investigation of anti-asthmatic potential of dried fruits of Vitis vinifera L. in animal model of bronchial asthma. Allergy Asthma Clin Immunol 12, 42 (2016). https://doi.org/10.1186/s13223-016-0145-x
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13223-016-0145-x