
Abstract
The fossil collections made by early explorers in South America have been fundamental to reveal the past diversity of extinct mammals and unravel their evolutionary history. One important early explorer in South America was the Swiss-Argentine palaeontologist Kaspar Jacob Roth, known as Santiago Roth (1850, Herisau, Switzerland-1924, Buenos Aires, Argentina), who made significant collections of fossil mammals that are housed in museums in Europe and Argentina. The important collections of Roth in Switzerland include iconic Pleistocene megafauna from the Pampean Region (Argentina). The palaeontological significance of the Pampean Region relies on its abundant record of fossil vertebrates that documents diversity dynamics and paleoenvironmental change in southern South America, serving as the basis for the South American biostratigraphical scale of the late Neogene and Quaternary. The South American native ungulates (SANUs) were hoofed placental mammals that radiated in South America. The clades Notoungulata and Litopterna include, among others, the last representatives of SANUs megafauna in the continent. We revise and describe for the first time the SANUs specimens from the Pampean Region of the Roth collections in Switzerland. The collections include two species of notoungulates (Toxodon cf. T. platensis and Mesotherium cristatum) and one litoptern species (Macrauchenia patachonica). The occurrences are restricted to the early and middle Pleistocene (pre-Lujanian Stages/Ages). Although the SANUs diversity in the Roth collections is low in comparison with other groups (e.g., xenarthrans), some of the specimens are very complete, including skulls and postcranial remains. The completeness of the Ma. patachonica material allows an update and reinterpretation of some of the details of the dentition and the postcranial skeleton of this iconic species. In addition to its historical importance, the SANU specimens from the Roth collections provide important information to study the paleobiology and evolution of South American megafauna and evaluate hypotheses about their extinction in the continent.
Resumen
Las colecciones de fósiles realizadas por los primeros exploradores en América del Sur han sido fundamentales para revelar la diversidad pasada de los mamíferos sudamericanos y desentrañar su historia evolutiva. Un importante explorador en América del Sur fue el paleontólogo suizo-argentino Kaspar Jacob Roth, conocido como Santiago Roth (1850, Herisau, Suiza-1924, Buenos Aires, Argentina), quien realizó importantes colecciones de mamíferos fósiles que se encuentran en museos de Europa y Argentina. Las colecciones de Roth en Suiza incluyen la icónica megafauna del Pleistoceno de la Región Pampeana (Argentina). La importancia paleontológica de la Región Pampeana se basa en su abundante registro de vertebrados fósiles que documenta la diversidad y el cambio paleoambiental en el sur de Sudamérica, sirviendo como base para la escala bioestratigráfica sudamericana del Neógeno tardío y Cuaternario. Los ungulados nativos sudamericanos (SANUs, por sus siglas en inglés) eran mamíferos placentarios con pezuñas que radiaron en Sudamérica. Los clados Notoungulata y Litopterna incluyen, entre otros, los últimos representantes de la megafauna de SANUs en el continente. En este trabajo, revisamos y describimos por primera vez los SANUs de la Región Pampeana de las colecciónes de Roth en Suiza. Las colecciónes incluyen dos especies de notungulados (Toxodon cf. T. platensis y Mesotherium cristatum) y una especie de litopterna (Macrauchenia patachonica). Las ocurrencias están restringidas al Pleistoceno temprano y medio (Pisos/Edades anteriores al Lujanense). Aunque la diversidad de SANU en las colecciónes Roth es baja en comparación con otros grupos (por ejemplo, xenarthros), algunos de los especímenes son muy completos, e incluyen cráneos y restos poscraneales. Lo completo del material de Ma. patachonica permite una actualización y reinterpretación de algunos de los detalles de la dentición y el esqueleto poscraneal de esta icónica especie. Además de su importancia histórica, los SANUs de la colección Roth brindan información importante para estudiar la paleobiología y evolución de la megafauna sudamericana y evaluar hipótesis sobre su extinción en el continente.
Similar content being viewed by others


Introduction
The “splendid isolation” and evolutionary history of South American mammals have fascinated palaeomammalogists for more than a century (Croft, 2016; Croft et al., 2020; Patterson & Pascual, 1968; Paula Couto, 1979; Simpson, 1980). The collections of fossil mammals made by early explorers in South America during the 19th and beginning of the 20th centuries have been fundamental to reveal the past diversity of South American mammals and unravel their evolutionary history (Defler, 2019; Fariña et al., 2013; Hatcher, 1985; Vizcaíno et al., 2017). These collections continue to be studied, some even recently highlighted for the first time (e.g., Carrillo-Briceño et al., 2016, 2021; Zurita-Altamirano et al., 2019), and have great historical and scientific value (Vizcaíno et al., 2017). One of the most important early fossil collectors in Argentina was the Swiss-Argentine palaeontologist Kaspar Jacob Roth, known as Santiago Roth (1850–1924), who made significant collections of fossil mammals that are housed in museums in Europe and Argentina (Bond, 1999a; Hansen, 2019; Sánchez-Villagra et al., 2023; Voglino et al., 2023).
Roth conducted numerous surveys in the Pampean Region of Argentina (Sánchez-Villagra et al., 2023; Voglino, 2020; Voglino et al., 2023) and published monographs and catalogues on the mammal fauna, geology and biostratigraphy of the region (e.g., Roth, 1888, 1889, 1904, 1921). Roth sold part of his collection to institutions in Denmark and Switzerland (Hansen, 2019), and today, the specimens are housed in the Zoological Museum of the University of Copenhagen (Hansen, 2019), the Natural History Museum of Geneva, the Cantonal Museum of Geology of the University of Lausanne, and the Palaeontological Institute and Museum of the University of Zurich (Voglino et al., 2023). The important collections of Santiago Roth in Switzerland include iconic Pleistocene megafauna from the Pampean Region, represented by a rich collection of giant glyptodonts and ground sloths, gomphotheres, and native ungulates (Carrillo-Briceño et al., 2023; Christen et al., 2023; Le Verger, 2023; Püschel & Martinelli, 2023; Roth, 1889).
The informal name of South American native ungulates (SANUs) includes several clades of hoofed placental mammals that radiated in South America during the Cenozoic (e.g., Croft et al., 2020 and references cited there). Notoungulata and Litopterna are the two SANU clades with the highest taxonomic diversity and longest temporal range, with a fossil record extending until the Late Pleistocene (Croft et al., 2020). Santiago Roth made a major contribution to the study of SANU evolution by defining the clade Notoungulata based on the anatomy of temporal bones (Roth, 1903).
Although the interrelationships between Notoungulata and Litopterna are not yet fully resolved, there have been important advances in the systematics of these clades (e.g., Billet, 2011; Carrillo & Asher, 2017; Carrillo et al., 2023; Cifelli, 1993; Deraco & García-López, 2016; McGrath et al., 2020b; Püschel et al., 2023; Shockey et al., 2022; Soria, 2001; Vera, 2015; Vera et al., 2019). Notoungulata includes two major sub-clades, Toxodontia (toxodonts and allies) and Typotheria, both of which have representatives that survived until the Late Pleistocene (Billet, 2011; Croft et al., 2020; Deraco & García-López, 2016). The two most diverse subclades within Litopterna are Proterotheriidae and Macraucheniidae, which also survived until the Late Pleistocene (Carrillo et al., 2023; Forasiepi et al., 2016; McGrath et al., 2020a; Püschel et al., 2023; Schmidt, 2015; Villafañe et al., 2006). Moreover, the Late Pleistocene Pampean record of Toxodon Owen, 1837 (Notoungulata) and Macrauchenia Owen, 1838 (Litopterna) provided key data that helped to clarify the phylogenetic relationships of these two extinct clades within placentals. Molecular data recovered from Toxodon and Macrauchenia supports a close relationship of notoungulates and litopterns with perissodactyls (Buckley, 2015; Welker et al., 2015; Westbury et al., 2017).
The palaeontological significance of the Pampean Region that attracted the attention of Roth and other explorers, relies on its abundant record of fossil vertebrates that documents diversity dynamics and paleoenvironmental change in southern South America and serves as the basis for the South American biochronological scale of the late Neogene and Quaternary (Cione & Tonni, 1995; Cione et al., 2015; Domingo et al., 2020; Gasparini et al., 2023; Pascual & Jaureguizar, 1990; Prado et al., 2021; Prevosti et al., 2021). The Pleistocene was a period of high mammal diversity in the Pampean Region, in particular megafauna, that was affected by paleoenvironmental changes (Le Verger, 2023; Prado et al., 2001; Prado et al., 2021). The SANUs megafauna survived in the Pampean Region until the Late Pleistocene/Holocene boundary (Prado et al., 2015), and its extinction in the region was likely the result of a synergy of paleoclimatic changes and anthropogenic impacts (Barnosky & Lindsey, 2010; Cione et al., 2003).
Here, we revise for the first time the Pleistocene SANUs specimens from the Pampean Region of the Roth collections in Geneva and Zurich. We provide a list of the referred material with identifications to the species level with anatomical descriptions, comments on the completeness of the specimens, and stratigraphic information when possible.
Materials and methods
We studied the SANUs specimens from the Roth collection housed in the Natural History Museum of Geneva (MHNG) and the specimens from the 5th catalogue of the fossils collected by Roth that were sold to the canton of Zurich (Roth, 1889) and currently housed at the Palaeontological Institute and Museum of the University of Zurich (PIMUZ). Descriptions were based on direct observations and current images from museum collections of relevant specimens. We revised their taxonomic identification according to the most recent studies for each taxon. We follow the criteria of Püschel et al. (2023) for dealing with the dental formula of litopterns and other SANUs. We follow Vera et al. (2019) on the dental terminology of Typotheria and Madden (1990) on Toxodontia. We follow Nessov et al. (1998) and Lobo et al. (2017) for the dental terminology of macraucheniids, although with some modifications and additions explained in the text and figures that update in a few instances the terminology previously used for this group. Measurements were taken in millimetres, either manually with a calliper, or digitally using Fiji (ImageJ v2.1.0) (Schindelin et al., 2012), to the nearest two decimal places. We followed the Nomina Anatomica Veterinaria (International Committee on Veterinary Gross Anatomical Nomenclature, 2017) for the anatomical terminology, and Bengston (1988) on the use of open nomenclature. Three-dimensional surface models of selected specimens described here are available at Sketchfab (https://sketchfab.com/PIMUZ).
All the specimens are from the Pampean Region in the Santa Fe and Buenos Aires Provinces, Argentina (Fig. 1). The precise stratigraphic provenance of the specimens is not known. Roth (1889) used the strata “Pampéen inférieur” (Lower Pampean), “Pampéen moyen” (Intermediate Pampean) and “Pampéen supérieur” (Upper Pampean). When possible, we provide the original strata provenance of each specimen as indicated by Roth in Table 1. Recent revisions on the chronostratigraphy and geochronology of the Pampean Region have considered that the age of the sediments, where Roth made his collections roughly correspond to the Ensenadan, Bonaerian, and Lujanian Ages/Stages (Table 1) (see Cione & Tonni, 1995; Cione et al., 2015; Voglino, 2020; Voglino et al., 2023; Le Verger, 2023; Fernández-Monescillo et al., 2023d and references therein).

Geographic provenance of the SANUs specimens of the Roth collection from the Pampean Region, Argentina
Institutional Abbreviations. MACN-PV, Museo Argentino de Ciencias Naturales “Bernardino Rivadavia”, Colección Nacional de Paleontología Vertebrados, Buenos Aires, Argentina; MLP, Museo de La Plata, La Plata, Argentina; MHNG GEPI, Muséum d’histoire naturelle, Geneva, Switzerland. MNHN.F, Muséum national d’Histoire Naturelle, Palaeontological collection, Paris, France; PIMUZ A/V, Palaeontological Institute and Museum of the University of Zurich, Zurich, Switzerland; YPM VPPU, Yale Peabody Museum, Vertebrate Palaeontology Princeton University collections, New Haven, USA.
Other abbreviations. I/i, upper and lower incisor; ka, thousand years ago; M/m, upper and lower molar; P/p, upper and lower premolar; SANUs, South American native ungulates.
Systematic palaeontology
Mammalia Linnaeus, 1758
Eutheria Huxley, 1880
Panperissodactyla Welker et al., 2015
Notoungulata Roth, 1903
Typotheria Zittel, 1893
Mesotheriidae Alston, 1876
Mesotherium Serres, 1867
Mesotherium cristatum Serres, 1867
Referred material. PIMUZ A/V 467, partial skull and mandible (Catalogue No. 5, specimen 131; Roth, 1889; Table 1); PIMUZ A/V 4133, partial mandible (Catalogue No. 5, specimen 40; Roth, 1889; Table 1).
Provenance. PIMUZ A/V 467 and PIMUZ A/V 4133 come from San Nicolás, Buenos Aires Province, Argentina. PIMUZ A/V 467 comes from the Intermediate Pampean and PIMUZ A/V 4133 comes from the Lower Pampean (Roth, 1889; Table 1; Fig. 1).
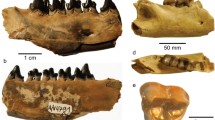
Remarks. Mesotherium cristatum is a common and iconic species in the South American fossil record. In the biostratigraphic scheme of the Pampean Region, it has been traditionally considered as a guide taxon of the Ensenadan Stage/Age (Cione & Tonni, 1995, 1999; Cione et al., 2015; Gasparini et al., 2023). Recently, the distribution of M. cristatum has been revised (Bond, 1999b; Fernández-Monescillo et al., 2023d), and the last record of the species has been documented at 220 ± 13 ka, which calls for a revision of the biostratigraphy of the Ensenadan Age (Fernández-Monescillo et al., 2023d). The taxonomic history and revisions of M. cristatum are complex due to the lack of recognition of ontogenetic variation (sometimes interpreted as inter-generic or intraspecific variation) and misidentification of specimens and taxa defined for Pliocene strata (Fernández-Monescillo et al., 2023b, 2023c; Simpson, 1940). A recent taxonomic revision concluded that variation among Middle and Early Pleistocene mesotheriinaes from the Pampean Region is consistent with ontogenetic and intraspecific variation of a single species (see Fernández-Monescillo et al., 2023b). We refer PIMUZ A/V 467 to M. cristatum based on the I1 mesiodistally elongated with a shallow lingual fold (Fig. 2A), the P4 lingually bilobed (Fig. 2B), the rostrally diverged lateral borders of the premaxilla (Fig. 2A), the inflated epitympanic theca (Fig. 2C), the lateral orientation of the parastyle in M3 (Fig. 2B), the outline of i1 nearly rectangular (Fig. 3D), the i1 having approximately five times the length of i2 (Table 2; Fig. 3D), and the mandibular symphysis with constrained lateral borders and concave ventral border (Fernández-Monescillo et al., 2023b). We refer PIMUZ A/V 4133 to M. cristatum based on the dental morphology, the constrained lateral border and straight ventral border of the mandibular symphysis (Fernández-Monescillo et al., 2023b). The dental and craniomandibular dimensions of PIMUZ A/V 467 are given in Tables 2 and 3.

Cranium of Mesotherium cristatum (PIMUZ A/V 467). A Ventral view. B Detail (left) and drawing (right) of the upper right dentition in occlusal view. C Lateral view. D Dorsal view. Notice the inflated epitympanic theca, the lateral borders of premaxilla diverging rostrally, the mesiodistally elongated I1 and the lateral orientation of the parastyle of M3 (see Fernández-Monescillo et al., 2023b)

Mandible of Mesotherium cristatum. PIMUZ A/V 467 (A–D). A Detail of the left lower dentition in occlusal view. Mandible in B dorsal view, C lateral, and D anterior views. E PIMUZ A/V 4133 in dorsal view. Notice the differences in size and occlusal outlines of i1 and i2, the constricted lateral borders and concave ventral border of the symphysis (see Fernández-Monescillo et al., 2023b)
Description. The upper dentition of PIMUZ A/V 467 is poorly preserved, but nevertheless some important diagnostic features can be observed (Fig. 2). The I1 is mesiodistally elongated, and it has a broad and shallow lingual fold, which is more clearly visible on the left I1 (Fig. 2A). The P3 is present, but the crown is damaged and details of the crown morphology cannot be seen (Fig. 2A, B). The P4 is lingually bilobed, with a mesiolabially oriented lingual sulcus (Fig. 2B). The ectoloph of the upper molars is oriented labially. In M1–2 the ectoloph is straight, whereas in M3, the ectoloph is concave with the parastyle strongly projected labially (Fig. 2B) (Fernández-Monescillo et al., 2023b; Patterson, 1952). There are two lingual sulci on M1–3.
The i1 has a nearly rectangular outline (similar longitudinal width) and is about five times longer than the i2 Table (Fig. 3; Table 2; see also Fernández-Monescillo et al., 2023a, 2023b). Only the crown of the left i2 is preserved, and it is single rooted and circular in cross section. The crown of the p4 is broken on the talonid, and the trigonid is narrow mesially (Fig. 3A). The m1–2 are similar to each other, with well-defined lingual and labial sulci separating the talonid and the trigonid, and the talonid being mesiodistally longer (Fig. 3A). The m3 is mesiodistally longer than m1–2 (Table 2), with an elongated talonid.
The skull is missing the zygomatic arches, the caudal portion of the palate and most of the basicranium (Fig. 2). The sagittal crest (crista sagittalis externa) is well-defined, and the nasals extend caudally until the level of the rostral border of the orbit on dorsal view (Fig. 2D). The skull shows two diagnostic features of M. cristatum: (a) the lateral borders of the premaxilla diverging rostrally in ventral view (Fig. 2A; Ameghino, 1918), and (b) the inflated epitympanic theca (Fig. 2C; Fernández-Monescillo, 2023b). The mandible is robust, reaching its maximum height at the level of m3 (Table 3; Fig. 3C). The coronoid process (processus coronoideus mandibulae) is high and the masseteric fossa (fossa masseterica) is wide. The mandibular symphysis extends caudally until the p4. The symphysis shows diagnostic features of M. cristatum, such as the constrained lateral borders, and the straight to concave ventral border (Fernández-Monescillo et al., 2023b).
Toxodontia Owen, 1853
Toxodontidae Gervais, 1847
Toxodon Owen, 1837
Toxodon cf. T. platensis Owen, 1837
Referred material. PIMUZ A/V 4163, partial mandible with left m1–3, fragments of left p3–4, alveoli of left p1–2, and right p1–4, and isolated right m1–2 (Catalogue No. 5, specimen 225; Roth, 1889; Table 1); PIMUZ A/V 4199, isolated right P2 and P3 (Catalogue No. 5, specimen 250; Roth, 1889; Table 1); PIMUZ A/V 4210, partial mandible with fragments of right and left m1–3, right p3–4, isolated right p1, fragments of right i1–3, and left i1, i3 (Catalogue No. 5, specimen 36; Roth, 1889; Table 1); PIMUZ A/V 4216 right femur and partial pelvis (Catalogue No 5, specimen 98; Roth, 1889; Table 1); PIMUZ A/V 4233, isolated right m2 (Catalogue No. 5, specimen 205; Roth, 1889; Table 1); PIMUZ A/V 4245, isolated left P4 (Catalogue No. 5, specimen 212; Roth, 1889; Table 1); PIMUZ A/V 4290, left ulna (Catalogue No. 5, specimen 62; Roth, 1889; Table 1); PIMUZ A/V 5697, partial skull (Catalogue No. 5, specimen 33; Roth, 1889; Table 1); MHNG GEPI V3665, partial skeleton, including atlas, axis, thoracic and lumbar vertebrae, ribs, sacrum, scapulae, partial left humerus, partial left ulna, and right femur (Table 1); MHNG GEPI V5791, two dental fragments, one lower premolar, possibly p2, and an upper molar fragment; MHNG GEPI V5792, right I2 (Table 1); MHNG GEPI V5793, fragmentary scapula and three thoracic vertebrae (Table 1), and MHNG GEPI V5794, partial left humerus (Table 1).
Provenance: PIMUZ A/V 4163 comes from Sánchez, Buenos Aires Province (Lower Pampean). PIMUZ A/V 4199 and PIMUZ A/V 4216 come from Arroyo Pavón, Santa Fe Province (Intermediate Pampean). PIMUZ A/V 4210, PIMUZ A/V 4233, and PIMUZ A/V 5697 come from Arroyo Pergamino, Buenos Aires Province (Intermediate Pampean). PIMUZ A/V 4245 comes from San Lorenzo, Santa Fe Province (Intermediate Pampean). PIMUZ A/V 4290 comes from Arroyo Merciel (Intermediate Pampean) (Roth, 1889; Table 1; Fig. 1).
Remarks. The first documented fossil remains of Toxodon were collected by Charles Darwin on the Voyage of the HMS Beagle in the 1830s, and later described by the Richard Owen (Fariña et al., 2013; Fernicola et al., 2009; Owen, 1837, 1838). Previous studies have described the skull and postcrania of Toxodon, and several Toxodon species have been described for the Pleistocene of the Pampean Region (Ameghino, 1887a; Bond et al., 1995; Liendo Lazarte, 1941; Miño-Boilini et al., 2006; Owen, 1837, 1838; Roth, 1895, 1898). Although there are not recent systematic revisions of the genus Toxodon, it has been proposed that only two species are likely valid for the Pleistocene of the Pampean Region, Toxodon platensis and the significantly smaller Toxodon gracilis Gervais & Ameghino, 1880 (Miño-Boilini et al., 2006). The other species would represent intraspecific and ontogenetic variation of a single species (T. platensis) (Miño-Boilini et al., 2006). We refer PIMUZ A/V 4163, 4199, 4233, 4245, 4290, and 5697 to Toxodon cf. T. platensis based on their size and overall morphology, as some of the described Toxodon species have not been formally designated as synonyms of T. platensis and following Bengston (1988) on the use of open nomenclature.
Description. PIMUZ A/V 5697 preserves only part of the cranium, including the occiput, occipitals, and part of the parietals (Fig. 4; Table 4). The sagittal (crista sagittalis externa) and the nuchal crest (crista nuchae) are well-developed, defining a wide temporal fossa (fossa temporalis) (Fig. 4A). The occipital condyles (condylus occipitalis) are large and oval, separated by a wide intercondylar notch (Fig. 4B). PIMUZ A/V 5697 shows no differences with the occipital part of the cranium described for other specimens of Toxodon (Roth, 1895, 1898).

Partial skull of Toxodon cf. T. platensis (PIMUZ A/V 5697). A Caudal portion of the cranium in dorsal view. B Occiput in caudal view
The two partial mandibles (PIMUZ A/V 4163 and 4210) are robust, as in T. platensis and unlike in T. gracilis (Ameghino, 1887a, 1887b; Miño-Boilini et al., 2006). The mandibular symphysis extends caudally until the level of m1 (Fig. 5A, C). In Toxodon, the symphysis extends until the p4 in juveniles and until the m1–2 in adults (Roth, 1898). The mandibular body (corpus mandibulae) in PIMUZ A/V 4163 is better preserved than in PIMUZ A/V 4210. The horizontal ramus of the mandible (ramus mandibulae) is deep and it has a mental foramen (formanen mentale) at the level of m1 (Fig. 5B).

Mandible and dentition of Toxodon cf. T. platensis. Mandible (PIMUZ A/V 4163) in A dorsal and B lateral views. C Mandible (PIMUZ A/V 4210) in dorsal view. D Right p1 (left) and m2 (right) in occlusal view (PIMUZ A/V 4163). E Right m2 in occlusal view (PIMUZ A/V 4233). F Left P4 in occlusal view (PIMUZ A/V 4245). G Right P3 in occlusal view (PIMUZ A/V 4199)
The I2 (MHNG GEPI V5792; Table 5) is developed as a tusk, it is poorly preserved, but two enamel bands are visible, on the mesial, and mesiolingual sides. The P2 (PIMUZ A/V 4199) is rhomboid in occlusal view, with labial and lingual enamel bands (Miño-Boilini et al., 2006). The P3 (PIMUZ A/V 4199) and P4 (PIMUZ A/V 4245) are similar in morphology, but the P4 is larger (Table 5). They have a lingual enamel fold oriented mesiolabially, lingual, and labial enamel bands (Fig. 5F, G). The lower incisors are only partially preserved in PIMUZ A/V 4210 and the base of the labial incisors (i3) are wider than the mesial ones (i1–2) (Fig. 5C), as in T. platensis (Ameghino, 1887a; Miño-Boilini et al., 2006; Roth, 1898). Only the crown of the right p1 is preserved, it is simple and oval in occlusal view (Fig. 5D). In the two partial mandibles (PIMUZ A/V 4163 and 4210) only parts of the crowns of p3 and p4 are preserved (Fig. 5A–C). They are elongated mesiodistally in occlusal view. The p4 has a labial groove (Fig. 5A). The lower molars have a buccal enamel fold on the labial side and anterior, meta-entoconid, and ento-hypoconulid folds on the lingual side (Fig. 5A, C–E). The size of the molars increases from m1 to m3 (Table 5). The m3 is elongated distomesially. The dental dimensions of the lower and upper teeth are witrange of known specimens of T. platensis (Table 5) (Miño-Boilini et al., 2006).
The limb bones of Toxodon have been described in previous works (e.g., Liendo Lazarte, 1941; Miño-Boilini et al., 2006; Roth, 1898), and the specimens of the Roth collections in Zurich and Geneva do not differ on the morphology or dimensions from elements previously described. The two right femora are similar in length (maximum length is 620 mm for MHNG GEPI V3665, and 600 mm for PIMUZ A/V 4216; Fig. 6). The humeri are robust, with a well-developed lateral epicondyle (epicondylus lateralis) (Fig. 6). Humeri dimensions are provided in Table 8. The left ulna (PIMUZ A/V 4290) is complete. The total length is 479 mm (Fig. 6), within the range of other Toxodon ulnae known (Roth, 1898). The shaft (corpus ulnae) is robust and is arched posteriorly (Fig. 6A). The olecranon is large and measures 163 mm. It is straight and longer than the trochlear notch (incisura trochlearis), which forms a crescent in lateral view (Fig. 6A). The anconeal process (processus anconeus) is large and projects anteriorly. The coronoid process (processus coronoideus ulnae) is larger than the anconeal process, and it extends medially (Fig. 6). PIMUZ A/V 4290 shows no differences with other described ulnae of Toxodon (Roth, 1898).

Limb bones of Toxodon cf. T. platensis from the Roth collections in Zurich and Geneva. Right femur (PIMUZ A/V 4216) in A anterior view, and B posterior view. Partial left humerus (MHNG GEPI V3665) in C anterior view, and D posterior view. Left ulna (PIMUZ A/V 4290) in E lateral view, and F medial view
Litopterna Ameghino, 1889
Macraucheniidae Gervais, 1855
Macraucheniinae Gervais, 1855
Macrauchenia Owen, 1838
Macrauchenia patachonica Owen, 1838
Referred material. PIMUZ A/V 4118, isolated right M2 with the roots missing (Catalogue No. 5, specimen 248; Roth, 1889; Table 1); PIMUZ A/V 4119, isolated left M2 with the tips of the roots broken (Catalogue No. 5, specimen 109; Roth, 1889; Table 1); PIMUZ A/V 5700, posterior portion of the cranium, complete series of seven cervical vertebrae (atlas, axis, C3–7), five thoracic vertebrae (T1–5), and right scapula and right forelimb, including the humerus, ulna-radius, carpals (scaphoid, lunate, cuneiform, pisiform, trapezium, trapezoid, magnum, and unciform), metacarpals (Mc II, Mc II, and Mc IV), and phalanges (proximal, intermediate, and distal; Catalogue No. 5, specimen 223; Roth, 1889; Table 1); MHNG GEPI V3659, anterior portion of right mandible with extremely worn i2, i3, and c, left fragment of maxilla with P4, M1–2, distal fragment of scapula, and rootless isolated right dP4 from a different individual; MHNG GEPI V3660, partial mandible with left m2–3; MHNG GEPI V3661, right partial mandible with p3–4. In the Zurich Roth collection, there is a right femur and pelvis fragment (PIMUZ A/V 4216) referred by Roth (1889) to Macrauchenia patachonica (Catalogue No. 5, specimen 98), which considering its size, proportions and anatomical features does not pertain to this taxon. We refer PIMUZ A/V 4216 to Toxodon cf. T. platensis.
Provenance. PIMUZ A/V 4118 comes from Villa Constitución, Santa Fe Province (Intermediate Pampean), PIMUZ A/V 4119 comes from Arroyo Ramallo, Buenos Aires Province (Intermediate Pampean), and PIMUZ A/V 5700 comes from Arroyo del Medio, Buenos Aires Province (Intermediate Pampean) (Roth, 1889; Table 1; Fig. 1). MHNG GEPI V3659 comes from an unknown locality of the Buenos Aires Province (Pampean), MHNG GEPI V3660 and MHNG GEPI V3661 come from Punta Santa Elena, Buenos Aires Province (Pampean) (Table 1; Fig. 1).
Remarks. Macrauchenia patachonica was first discovered by Darwin in 1834 in the vicinities of Puerto San Julián, Argentinean Patagonia, and later described by Richard Owen (Fernicola et al., 2009; Owen, 1838). Most of the skeleton of Ma. patachonica have been described in previous works (e.g., Burmeister, 1864, 1864–1869; Fernández de Álvarez, 1940; Gervais, 1855; Owen, 1838; Püschel & Martinelli, 2023; Sefve, 1925) or compared with earlier macraucheniids (e.g., Püschel et al., 2023; Scott, 1910), so here we focus only in relevant aspects previously unnoticed of some of the elements, expanding the anatomical knowledge of this iconic species. The craniodental and postcranial dimensions of PIMUZ A/V 5700, PIMUZ A/V 4118, PIMUZ A/V 4119, MHNG GEPI V3659, MHNG GEPI V3660, and MHNG GEPI V3661 are given in Tables 4, 5, 6, 7, 8, 9, and 10.
Description. PIMUZ A/V 5700 includes a fragmentary cranium that preserves the posterior portion, including the basicranium (Fig. 7). Cranial measurements of PIMUZ A/V 5700 are given in Table 4. The skull and dentition of Ma. patachonica have been already described in detail by Burmeister (1864, 1864–1869). More recently, some studies focusing on the cranial anatomy, including the petrosal and inner ear anatomy of other litopterns, have used specimens of Ma. patachonica in their comparisons revealing previously unknown aspects of macraucheniid’s cranial evolution (Billet et al., 2015; Dozo et al., 2023; Fernández-Monescillo, 2020; Forasiepi et al., 2016).

Cranium of Ma. patachonica (PIMUZ A/V 5700) in dorsal A and ventral B views. The cranium (PIMUZ A/V 5700) only preserves its posterior portion and basicranium, and was previously subjected to a poor restoration with plaster which covers some of the cranial structures
The specimens PIMUZ A/V 4119, PIMUZ A/V 4118, and MHNG GEPI V3659 represent different ontogenetic stages of the upper molars of Ma. patachonica (Fig. 8D–M). PIMUZ A/V 4119 and PIMUZ A/V 4118 are M2s with a different wear stage, the former being a relatively unworn left M2 (Fig. 8D–F) and the latter a worn right M2 (Fig. 8G–I). The M2 in MHNG GEPI V3659 (that also preserves the P4–M2 housed in a left maxilla fragment; Fig. 8M), presents higher degree of wear than PIMUZ A/V 4118 and PIMUZ A/V 4119, representing an older individual. However, there is an isolated and unworn deciduous P4 (dP4) stored in MHNG GEPI V collections under the same number (MHNG GEPI V3659; Fig. 8J–L, which it must be from a different and younger individual considering that the left maxilla fragment has a fully erupted P4 with considerable degree of wear (Fig. 8M). The dP4 is almost completely preserved, missing the lingual posteriormost portion of the crown and most of the roots. Overall, the dP4 is similar in morphology to an M1 as is seen in earlier macraucheniids (e.g., Theosodon lydekkeri [MACN A 9269-88]) and eutherian mammals in general (Jernvall, 1995). Key differences between the M1 and the dP4 in Ma. patachonica are a smaller size, and a considerably lower crown height in the dP4. The dP4 also presents a low ectocingulum or buccal cingulum which is possible to examine at a lower degree of wear (Fig. 8J, K). In contrast, in the permanent dentition the ectocingulum would be only possible to examine in the teeth with the highest degree of wear, considering the crown height and curvature of unworn molars, such as PIMUZ A/V 4119 (Fig. 8D–F). The preservation of PIMUZ A/V 4118 and PIMUZ A/V 4119 does not allow to evaluate the presence of an ectocingulum, as in both specimens the portion of the crown close to the roots and the roots are missing. In Ma. patachonica, the M2 is distinguished from M1 by being larger and having a consistently more buccally projected parastyle in comparison with the metastyle, whereas in M1, the parastyle and the metastyle tend to have a subequal buccal projection. The degree of projection of the parastyle in M2 varies with wear and it is easier to evaluate in upper molars in place in the maxilla. Dental measurements of PIMUZ A/V 4119, PIMUZ A/V 4118, MHNG GEPI V3659, and additional specimens that include lower dentition (i.e., MHNG GEPI V3659, MHNG GEPI V3660) are given in Table 5.

Upper dentition of Macraucheniidae. A Illustration of a Cramauchenia normalis upper molar (M1) indicating the cusps and cristae. B Illustration of a Theosodon lydekkeri upper molar (M1) indicating the cusps and cristae. C Illustration of a Macrauchenia patachonica upper molar (M1) indicating the cusps and cristae. D–F Relatively unworn left upper molar (M2) of Ma. patachonica (PIMUZ A/V 4119) in occlusal (D), lingual (E), and mesial (F) views. G–I. Worn right upper molar (M2; here mirrored) of Ma. patachonica (PIMUZ A/V 4118) in occlusal (G), lingual (H), and mesial (I) views. J–L. Unworn right deciduous premolar (dP4; here mirrored) of Ma. patachonica (MHNG GEPI V3659) in occlusal (J), buccal (K), and lingual (L) views. M Left maxilla fragment with P4, M1–2 of Ma. patachonica (MHNG GEPI V3659). The illustration of C. normalis’ M1 in A of was based on the specimens MLP 83-III-2–1 and MLP 85-V-VII-3-38a. The illustration of T. lydekkeri’s M1 in B was based on the specimens MACN-A 9269–88 and YPM VPPU 015717. The illustration of Ma. patachonica’s M1 in C was based on the specimen MACN-PV 11361, among other specimens. In A–C, the labels of controversial structures are in bold and relevant cusps are highlighted in different colours: blue for paraconule, red for protocone, green for metaconule, yellow for hypocone, and dark yellow for hypocone–metaconule. Notice that even though the dP4 (J–L) and the maxilla fragment with P4, M1–2 (M) share the same specimen number (MHNG GEPI V3659), these are from different individuals considering their different and incompatible ontogenetic stage (see the extreme wear in the permanent P4 in M). dif distolingual fossette, ecg ectocingulum, ets entostyle, hy hypocone, hy-mtl hypocone–metaconule, lmef lingual median fossete, maf mesiolabial fossette, me metacone, mef median fossette, mif mesiolingual fossette, ms mesostyle, mt metastyle, mtl metaconule, pa paracone, pcg precingulum, pplc preparaconule crista, pprc preprotocrista, pr protocone, prl paraconule, ps parastyle, pscg postcingulum, psmlc postmetaconule crista, psplc postparaconule crista, psprc postprotocrista, tb trigon basin
The changes of tooth morphology, enamel fossetes, and wear during the ontogeny of Ma. patachonica have not been established as it has been in its Pleistocene–Holocene relative Xenorhinotherium bahiense Cartelle & Lessa, 1988 (Lobo et al., 2017). However, from observing the upper dentition of several specimens of Ma. patachonica in different ontogenetic stages (e.g., MACN-PV 2, MACN-PV 11361, MNHN.F PAM 75, MHNG GEPI V3659), the wear degrees and enamel fossettes patterns of change are overall similar in both taxa. Consequently, in terms of the wear degrees defined by Lobo et al. (2017) for X. bahiense, PIMUZ A/V 4119 would be between degrees 1 and 2, and PIMUZ A/V 4118, between degrees 2 and 3, which is also reflected in the crown height of both specimens (Fig. 8D–F, G–I). In Ma. patachonica, to these wear degrees, it may be added a degree 6, in which a new lingual median fossette is generated in the position of the protocone (e.g., M2 in MHNG GEPI V3659; Fig. 8M), and also a degree 7, in which the original median fossette disappears (e.g., M1 in MHNG GEPI V3659; Fig. 8M). In addition, it has even been observed in the M1 of a Ma. patachonica specimen with extreme wear the disappearance of the lingual median and mesiolingual fossettes (e.g., Sefve, 1925: Fig. 4), which if confirmed in more specimens could be considered a degree 8 of wear.
It is interesting to note that the general morphology and cusps present in unworn M1–2 of X. bahiense (Lessa, 1992; Lobo et al., 2017) are extremely similar to those of Ma. patachonica. Indeed, Lobo et al. (2017) cusps interpretations for M1–2 of X. bahiense in unworn specimens, are mostly similar to our own interpretations of the cusps for M1–2 in Ma. patachonica (Fig. 8C). However, the cusp they interpret as a protocone, we interpret it as a paraconule, and in the position of what they described as a protocone–hypocone crest, there is a cusp that we interpret as the actual protocone. The protocone of the M1–2 of X. bahiense can be observed more clearly in Lessa (1992: pl 10). The protocone and paraconule are also clearly present in the dP4 of Ma. patachonica (MHNG GEPI V3659; Fig. 8J, L). The justification for our identification of the cusps and cristae in the M1–2 of Ma. patachonica (Fig. 8C) is given by the M1–2 morphology and cusp configuration of the earlier macraucheniids Cramauchenia normalis Ameghino 1902 (Late Oligocene–Early Miocene/Deseadan–Colhuehuapian Ages; Fig. 8A) and Theosodon lydekkeri Ameghino, 1887b (Early Miocene/Santacrucian Age; Fig. 8B). Cramauchenia normalis has a M1–2 configuration that is relatively easy to interpret as it shares most of the cusps seen in early South American “condylarths” with well-defined conules (Muizon & Cifelli, 2000), presenting, in addition, a well-developed postcingulum-derived “true” hypocone, such as the one present in didolodontid Didolodus Ameghino, 1887b (Muizon et al., 2019). It is interesting to note that in C. normalis the trigon basin is reduced and shifted mesially, and the metaconule is extremely reduced and very close to the hypocone, being both cusps connected by a crista (prehypocrista; Fig. 8A). In T. lydekkeri (and other species of the genus Theosodon) the hypocone shifts mesially, occupying the position of the reduced metaconule of C. normalis, which could represent a fusion of the metaconule with the hypocone (Soria, 1981; Fig. 8B). Therefore, the prehypocrista is lost in T. lydekkeri, and the cristae from the trigon that were connected to the metaconule in C. normalis (postprotocrista and postmetaconule crista) are now connected to this hypocone–metaconule cusp. Macrauchenia patachonica and also X. bahiense keep the same M1–2 cusp configuration displayed by T. lydekkeri with an increased hypsodonty, and the presence of enamel fossettes, from which the mesial and distal ones are enclosed by cristae that probably derive from a raised pre and postcingulum (Fig. 8C). Therefore, the lingual cusps that Ma. patachonica and X. bahiense exhibit in M1–2 (only seen in unworn or slightly worn specimens) going from mesial to distal are: paraconule, protocone, and hypocone–metaconule. The same evolutionary trend occurs in the M3 of macraucheniids.
In relation to the vertebrae of Ma. patachonica, Burmeister (1864–1869) mostly based on MACN-PV 2, inferred the presence of seven cervical vertebrae, 17 thoracic vertebrae and seven sacral vertebrae. Sánchez-Villagra et al. (2007) mentioned that Ma. patachonica has 20–21 thoracolumbar vertebrae based on an unknown MLP specimen, (likely MLP 12-1424, considering that is the most complete Ma. patachonica specimen so far described). From examining MLP 12-1424 and PIMUZ A/V 5700 it can be confirmed that Ma. patachonica has seven cervical vertebrae. Assuming that the thoracic and lumbar vertebral series is complete in MLP 12-1424, Ma. patachonica has 16 thoracic vertebrae and five lumbar vertebrae, making a total of 21 thoracolumbar which is consistent with previous reports (Sánchez‐Villagra et al., 2007). From these, PIMUZ A/V 5700 only preserves the anterior portion (C1–7 and T1–5; Fig. 9). The atlas was previously described wrongly by Burmeister (1864–1869), so it is described in detail and figured correctly for the first time in a different publication of this series which deals specifically with this controversy (Püschel & Martinelli, 2023). Vertebrae measurements of C2–T5 of PIMUZ A/V 5700 are given in Table 6.

Cervical (C2–7) and thoracic (T1–5) vertebrae of Macrauchenia patachonica (PIMUZ A/V 5700). A1–3 Second cervical vertebra (C2) in anterior (A1), left lateral (A2), and ventral (A3) views. B1–3 Third cervical vertebra (C3) in anterior (B1), left lateral (B2; mirrored), and ventral (B3) views. C1–3 Fourth cervical vertebra (C4) in anterior (C1), left lateral (C2), and ventral (C3) views. D1–3 Fifth cervical vertebra (C5) in anterior (D1), left lateral (D2), and ventral (D3) views. E1–3 Sixth cervical vertebra (C6) in anterior (E1), left lateral (E2; mirrored), and ventral (E3) views. F1–3 Seventh cervical vertebra (C7) in anterior (F1), left lateral (F2), and ventral (F3) views. G1–3 First thoracic vertebra (T1) in anterior (G1), left lateral (G2; mirrored), and ventral (G3) views. H1–3 Second thoracic vertebra (T2) in anterior (H1), left lateral (H2), and ventral (H3) views. I1–3. Third thoracic vertebra (T3) in anterior (I1), left lateral (I2), and ventral (I3) views. J1–3. Fourth thoracic vertebra (T4) in anterior (J1), left lateral (J2), and ventral (J3) views. K1–3 Fifth thoracic vertebra (T5) in anterior (K1), left lateral (K2; mirrored), and ventral (K3) views. ant fov anterior costal fovea, dor dorsal, fov tran costal fovea of transverse process, nu can nutrient canal, odon proc odontoid process, postzyg postzygapophysis, pos fov posterior costal fovea, prezyg prezygapophysis, spin proc spinous process, tr for transverse foramen, tr proc transverse process, tub small posterior transverse process tubercle, vent ventral
The cervical vertebrae are in general elongated (except C6 and C7) with anteroposteriorly long transverse and spinous processes and almost flat faces of their bodies (Fig. 9A1–F3). Previous authors (Burmeister, 1864; Burmeister, 1864–1869; Owen, 1838) described in detail C2–4, and C6. Although the C5 and C7 of Ma. patachonica have not been formally described, previous studies have compared their anatomy with those of other macraucheniids, such as Theosodon and Micrauchenia Püschel et al., 2023 (Püschel et al., 2023; Scott, 1910). The body of C5 is dorsoventrally compressed and ventrally presents two divergent ridges (Fig. 9D1–3). The spinous process of C5 is mediolaterally thin and anteroposteriorly elongated. The transverse process is also anteroposteriorly elongated, but it is divided into two separate sections: a ventral transverse process located in the anterior portion of the vertebra and projects anteriorly from the body, and a dorsal transverse process which extends anteroposteriorly around the dorsoventral midsection of the body. In contrast to C3 and C4, the dorsal transverse process of C5 does not reach the posterior margin of the body, a feature also seen in the C5 of Theosodon and Micrauchenia (Püschel et al., 2023). As in C2–4 and C6, the transverse foramen (foramen transversarium) is on the medial side of the wall of the neural arch. Dorsally the prezygapophyses (processus articularis cranialis) and postzygapophyses (processus articularis caudalis) form a U-shape and V-shape, respectively. In terms of size, C5 is shorter than C2–4, but longer than C6–7 (Table 6).
In C6 is interesting to note that in PIMUZ A/V 5700, the transverse foramen is located in the midpoint of the wall of the neural arch and the transverse process (Fig. 9E1), in contrast to the specimen illustrated by Burmeister (1864, 1864–1869; MACN-PV 2) in which it is located on the medial side of the wall of the neural arch as in C2–5. We interpret these differences as intraspecific variation. This is interesting, because in Theosodon, the transverse foramen perforates the transverse process (Scott, 1910), so PIMUZ A/V 5700 presents an intermediate condition between Theosodon and MACN-PV 2. C7 is an anteroposteriorly short cervical vertebra (Fig. 9F1–3). The main differences between C7 with other cervical vertebrae are (a) the absence of a transverse foramen, (b) a continuous and undivided transverse process located at the dorsoventral midpoint of the vertebral body, (c) the presence of a marked ventral keel in the vertebral body located at the mediolateral midpoint, and (d) the presence of posterior costal fovea. In terms of size, C7 is by far the shortest cervical vertebra (Table 6). Detailed comparisons of C7 across different macraucheniids including Ma. patachonica (and PIMUZ A/V 5700) have been previously conducted (Püschel et al., 2023).
The thoracic vertebrae of Ma. patachonica have a body that in general is notably shorter than the average cervical vertebrae and a more robust spinous process (Fig. 9G1–K3). Burmeister (1864, 1864–1869) described three thoracic vertebrae of Ma. patachonica from the Bravard’s collection that he considered to be T6–8, and also T13 and T16 from the MACN collection (later included under the number MACN-PV 2 ), so the rest of the thoracic vertebrae of this taxon remain undescribed. However, Püschel et al. (2023) gave a detailed description of T2 in Micrauchenia that includes comparisons with other macraucheniids, including Macrauchenia (PIMUZ A/V 5700). PIMUZ A/V 5700 preserves T1–5 with a variable degree of completeness. From T1 to T5 is possible to see a gradual change in morphology from a more cervical-like thoracic vertebra (T1) to a more typical thoracic vertebra (T5). T1 is extremely similar to C7 in general shape and proportions (Fig. 9G1–3; Table 6), the main differences being (a) the absence of a prominent ventral keel, (b) the presence of anterior costal fovea, (c) a more prominent spinous process and (d) the presence of costal fovea of the transverse process. T2–5 have a more prominent spinous process than T1 and the transverse process is less laterally projected.
As Burmeister (1864–1869) previously observed, there are five gradual morphological changes from the first to the last thoracic vertebrae in many mammals, which apply to Ma. patachonica: (a) the anterior vertebrae tend to be larger than the posterior, (b) in the anterior vertebrae, the anterior and posterior costal foveae tend to more ventrally located and larger, but as the vertebrae series progress towards the last thoracic vertebrae, the costal foveae tend to be more dorsally located and smaller, (c) in the anterior vertebrae the transverse process tends to have a more ventral position in relation to the vertebral arch, but as the vertebrae series progress towards the last thoracic vertebrae, the transverse process tends to be more dorsally located, (d) the spinous process tends to change from a more posteriorly bent position in the anterior vertebrae to a more perpendicular position in the posterior vertebrae, and (e) the prezygapophyses and postzygapophyses tend to be overall flat or horizontal in the anterior vertebrae, but as the vertebrae series progress towards the last thoracic vertebrae, the prezygapophyses and postzygapophyses tend to have an angle, being the prezygapophysis somewhat concave and the postzygapophyses somewhat convex.
Comparing the thoracic vertebrae that Burmeister (1864–1869) tentatively referred to T6–8 with the T1–5 in PIMUZ A/V 5700 and the probably complete vertebral series of MLP 12–1424, Burmeister (1864–1869) vertebrae correspond to T4–6 instead of T6–8 considering the length and direction of their spinous processes and the position and size of their anterior and posterior costal foveae.
The scapula of Ma. patachonica has been previously described, particularly its distal portion (Owen, 1838), so what follows is an update of this description considering that the PIMUZ A/V 5700 has a more complete scapula (only missing part of the anterodorsal portion and parts of the scapular spine [spina scapulae]; Fig. 10A–C. Measurements of the scapula of PIMUZ A/V 5700, and MHNG GEPI V3659, that includes a poorly preserved distal fragment of a scapula, are given in Table 7. The scapula of Ma. patachonica presents a large and concave glenoid cavity (cavitas glenoidalis) which narrows anteriorly ending in a prominent supraglenoid tubercle (tuberculum supraglenoidale; also known as coracoid). Dorsal to the supraglenoid tubercle there is a wide and shallow scapular notch (incisura scapulae). Laterally, a strong scapular spine separates the posterior fossa infraspinata from the anterior fossa supraspinata, both being strongly concave. Although lateral-most portions of the scapular spine are missing (as in the holotype NHMUK-PVM 43402; Owen, 1838), two posterior bendings, one close to its dorsoventral midpoint and also the other one in the acromion suggest the presence of a tuber in the middle of the scapular spine (tuber spinae scapulae) and a caudal process of the acromion or suprahamatus process (processus suprahamatus). The ventral process of the acromion or hamatus process (processus hamatus) is small. Medially there is a marked concavity that extends dorsoventrally and it is most profound just above the supraglenoid tubercle, which we interpret as a subscapular fossa (fossa subscapularis) for the origin of the subscapularis muscle. Overall, the scapula of Ma. patachonica is very similar to other macraucheniids, such as Theosodon and Xenorhinotherium (Lessa, 1992; Scott, 1910).

Right forelimb of Macrauchenia patachonica (PIMUZ A/V 5700). A–C Scapula in lateral (A), medial (B), and distal (C) views. D–H Humerus in proximal (D), anterior (E), posterior (F), medial (G), and lateral (H) views. I–J Ulna-radius in anterior (I) and posterior (J) views. K Manus (carpals, metacarpals and phalanges) in anterior view. In E the olecranon fossa was restored, so it in PIMUZ is not as deep as it should be. In K the carpals and metacarpals were glued together with the different elements in the right position, but in many cases with a wrong orientation (e.g., pisiform and Mc IV). In addition, the ungual phalanx of the third digit is fragmentary, preserving only its distomedial portion. ae aliform expansion of the radius, bg bicipital groove; ca, capitulum, ce capitular eminence, ch crest of the humerus, ctm crista tuberculi minoris, cu cuneiform, dpc deltopectoral crest, dt deltoid tuberosity, gl glenoid cavity; gt, greater tubercle, h head of the humerus, hp hamatus process, ht humeral trochlea, if fossa infraspinata, it intermediate tubercle, le lateral epicondyle, lt lesser tubercle; lu lunate, ma magnum, mcp medial coronoid process (processus coronoideus medialis), me medial epicondyle, of olecranon fossa, ol olecranon (tuber olecrani), pa anconeal process (processus anconeus), pi pisiform, ra radius, rf radial fossa; rt, radial or bicipital tuberosity (tuberositas radii), sc scaphoid, scn scapular notch, sf fossa supraspinata, shp suprahamatus process, sp styloid process (processus styloideus), sr supracondylar ridge, st supraglenoid tubercle, tr trapezium, trd trapezoid, trl tricipital line, tsp tuber of the scapular spine, ttm tuberosity of the teres major, ul ulna, un unciform
The humerus of Macrauchenia was previously described briefly by Sefve (1925), because, in a previous contribution, he described the humerus of Macrauchenia ullomensis Sefvre 1914, from Ulloma, Bolivia, which he found was very similar to the humerus of Ma. patachonica (Sefve, 1914). However, Sefve (1914) asserted that overall, there were enough differences in the postcranial elements of both taxa to merit a species separation, which has been accepted by more recent workers (Guérin & Faure, 2004; Hoffstetter, 1986). Considering the description of Sefve (1914) and the additions of Sefve (1925), our description focuses mostly on aspects previously unnoticed. Measurements of the humerus of PIMUZ A/V 5700 are given in Table 8. The humerus of Ma. patachonica is overall wide but with a marked constriction close to the distal epiphysis (Fig. 10D–H). Proximally, in the posterior aspect, the humerus presents a mediolaterally wide head (caput humeri). In the anterior aspect of the proximal epiphysis, the medial side is dominated by a small lesser tubercle (tuberculum minus), whereas the lateral side is dominated by a large and anteriorly projected greater tubercle (tuberculum majus; Fig. 10D). The head is separated by a strong concavity from the lesser tubercle. The lesser and the greater tubercles are separated by a bicipital groove (sulcus intertubercularis), which in its mediolateral midpoint presents a small convexity that could be interpreted as a rudimentary intermediate tuberculum (tuberculum intermedium).
Anteriorly, the humerus exhibits a marked deltopectoral crest (crista tuberculi majoris) starting from the greater tubercle and running distally up to a moderately sized deltoid tuberosity (tuberositas deltoidea) laterally (Fig. 10E). Running distally from the deltoid tuberosity there is a low and subtle crest of the humerus (crista humeri). Medially, a low crest runs from the lesser tubercle (crista tuberculi minoris) up to a subtle tuberosity of the teres major (tuberositas teres major; Fig. 10G). Posteriorly, the tricipital line (linea m. tricipitatis) starts close to the midpoint between the head of the humerus and the greater tubercle and runs distolaterally ending in the deltoid tuberosity (Fig. 10F). Distal from the tricipital line, in the diaphysis, a supracondylar ridge (crista supracondylaris lateralis) runs laterally, starting as a low ridge and strengthening as it approaches and connects with the large lateral epicondyle (epicondylus lateralis) in the distal end of the humerus. Medially, the medial epicondyle (epicondylus medialis) is moderately developed being noticeably smaller than the lateral epicondyle. Distally, before reaching the condyle of the humerus (condylus humeri) there are deep fossae, the olecranon fossa (fossa olecrani) anteriorly, and radial fossa (fossa radialis) posteriorly. These two fossae do not connect through a supratrochlear foramen (foramen supratrochleare). Among macraucheniids, the absence of a supratrochlear foramen in Ma. patachonica is shared with Theosodon and Xenorhinotherium (Lessa, 1992; Scott, 1910), in contrast with Cramauchenia and Llullataruca McGrath et al., 2018, which have a marked supratrochlear foramen (Dozo & Vera, 2010; McGrath et al., 2018). In the condyle of the humerus, the capitulum (capitulum humeri) is markedly more distally developed than the trochlea (trochlea humeri), although there is no indication of separation between facets. The trochlea is somewhat anteroposteriorly thicker than the capitulum.
The specimen PIMUZ A/V 5700 preserves the ulna-radius (Fig. 10I, J) and an almost complete manus (Fig. 10K; only misses the reduced Mc V). Measurements of these elements are given in Tables 9and10. The ulna-radius and manus of Ma. patachonica have been described in detail in previous publications (Gervais, 1855; Owen, 1838; Parodi, 1931; Sefve, 1914, 1925). Elements of the forelimb of Ma. patachonica have been also recently analysed and compared with different macraucheniids that include inferences in palaeobiology and locomotion of this taxon and other members of this litoptern family (Püschel et al., 2023).
Conclusion
We reviewed, described, and illustrated for the first time the SANU specimens from the Pampean Region of the historical Roth collection in Zurich. The collection includes two notoungulates (Toxodon cf. T. platensis and Mesotherium cristatum) and one litoptern (Macrauchenia patachonica), restricted to the “Lower” and “Intermediate” Pampean (Ensenadan to Bonaerian Stages/Ages). Although the SANUs diversity in the Roth collection is low in comparison with other groups, such as xenarthrans (Le Verger, 2023), Neartic ungulates (Carrillo-Briceño et al., 2023), and rodents (Kerber, 2023), some of the specimens are very complete, including skulls and postcranial remains. The completeness and quality of the fossil material allow us to examine and discuss previously unnoticed anatomical aspects, including new interpretations on patterns of dental wear and on the identity of the cusps present on the upper molars in Ma. patachonica that allow homologous comparisons with other macraucheniids and potentially other SANUs. In addition to its historical and patrimonial importance, the SANUs specimens from the Roth collection provide important information that have the potential to shed light on the study of the paleobiology and evolution of South American megafauna and evaluate hypotheses about their extinction in the continent.
Availability of data and materials
The data and measurements generated in this study are included in the published article.
References
Alston, E. R. (1876). On the Classification of the Order Glires. Proceedings of the Zoological Society of London, 44(1), 61–98. https://doi.org/10.1111/j.1096-3642.1876.tb02543.x
Ameghino, C. (1918). Los yacimientos fosilíferos del Valle de Santa María en Catamarca y Tucumán. Anales de la Primera Reunión Nacional de la Sociedad Argentina de Ciencias Naturales, 1, 1–1148.
Ameghino, F. (1887a). Observaciones generales sobre el Orden de mamíferos extinguidos sudamericanos llamados toxodontes (Toxodontia) y sinopsis de los géneros y especies hasta ahora conocidos. Anales del Museo de la Plata, 1, 1–66.
Ameghino, F. (1887b). Enumeración sistemática de las especies de mamíferos fósiles coleccionados por Carlos Ameghino en los terrenos eocenos de la Patagonia austral y depositados en el Museo La Plata. Boletín del Museo de la Plata, 1, 1–26.
Ameghino, F. (1889). Contribución al conocimiento de los mamíferos fósiles de la República Argentina. Actas de la Academia Nacional de Ciencias de la República Argentina en Córdoba, 6, 1–1027.
Ameghino, F. (1902). Première contribution à la connaissance de la faune mammalogique descouches à Colpodon. Boletín de la Academia Nacional de Ciencias en Córdoba, 17, 71–138.
Barnosky, A. D., & Lindsey, E. L. (2010). Timing of Quaternary megafaunal extinction in South America in relation to human arrival and climate change. Quaternary International, 217(1–2), 10–29.
Bengston, P. (1988). Open nomenclature. Palaeontology, 31(1), 223–227. https://doi.org/10.4039/Ent8431-1
Billet, G. (2011). Phylogeny of the Notoungulata (Mammalia) based on cranial and dental characters. Journal of Systematic Palaeontology, 9(4), 481–497. https://doi.org/10.1080/14772019.2010.528456
Billet, G., de Muizon, C., Schellhorn, R., Ruf, I., Ladevèze, S., & Bergqvist, L. (2015). Petrosal and inner ear anatomy and allometry amongst specimens referred to Litopterna (Placentalia). Zoological Journal of the Linnean Society, 173(4), 956–987. https://doi.org/10.1111/zoj.12219
Bond, M. (1999a). Santiago Roth. Revista Museo, Fundación Museo de la Plata, 2(13), 33–37.
Bond, M. (1999b). Quaternary Native Ungulates of Southern South America. A Synthesis. In E. P. Tonni & A. L. Cione (Eds.), (Vol. 11, pp. 177–205). Rotterdam: CRC Press.
Bond, M., Cerdeño, E., & López, G. M. (1995). Los ungulados nativos de América de Sur. In M. T. Alberdi, G. Leone, & E. P. Tonni (Eds.), Evolución biológica y climática de la región Pampeana durante los últimos cinco millones de años. Un ensayo de correlación con el Mediterráneo occidental (Vol. 12, pp. 259–275). Madrid: Consejo Superior de Investigaciones Científícas.
Buckley, M. (2015). Ancient collagen reveals evolutionary history of the endemic South American ‘ungulates.’ Proceedings of Royal Society of Biology, 282(1806), 20142671. https://doi.org/10.1098/rspb.2014.2671
Burmeister, G. (1864). Beschreibung der Macrauchenia patachonica Owen (Opisthorhinus falkoneri Brav.) nach A. Bravard’s Zeichnungen und den im Museo zu Buenos Aires vorhandenen Resten entworfen. AAbhandlung der Naturforscher Gesellschaft zu Halle, 1, 75–112.
Burmeister, G. (1864–1869). Descripción de la Macrauchenia patachonica. In Anales del Museo Público de Buenos Aires para dar a conocer los objetos de historia natural nuevos o poco conocidos conservados en este establecimiento (Vol. Tomo I, pp. 32–66). Buenos Aires: Imprenta de “La Tribuna.”
Carrillo, J. D., & Asher, R. J. (2017). An exceptionally well-preserved skeleton of Thomashuxleya externa (Mammalia, Notoungulata), from the Eocene of Patagonia, Argentina. Paleontologia Electronica, 20.2.34.A, 1–33. https://doi.org/10.26879/759
Carrillo, J. D., Suarez, C., Benites-Palomino, A., Vanegas, A. F., Link, A., Rincon, A. F., et al. (2023). New remains of Neotropical bunodont litopterns and the systematics of Megadolodinae (Mammalia: Litopterna). Geodiversitas, 45(15), 409–447. https://doi.org/10.5252/geodiversitas2023v45a15
Carrillo-Briceño, J. D., Aguilera, O. A., Benites-Palomino, A., Hsiou, A. S., Birindelli, J. L. O., Adnet, S., et al. (2021). A historical vertebrate collection from the Middle Miocene of the Peruvian Amazon. Swiss Journal of Palaeontology, 140(1), 26. https://doi.org/10.1186/s13358-021-00239-7
Carrillo-Briceño, J. D., Amson, E., Zurita, A., & Sánchez-Villagra, M. R. (2016). Hermann Karsten (1817–1908): A German naturalist in the Neotropics and the significance of his paleovertebrate collection. Fossil Record, 20(1), 21–36. https://doi.org/10.5194/fr-20-21-2016
Carrillo-Briceño, J. D., Vezzosi, R. I., Ming, K. M., Christen, Z. M., Mothé, D., Ruiz-Ramoni, D., & Sánchez-Villagra, M. R. (2023). Nearctic Pleistocene ungulates from the Pampean region (Argentina) in the historical collections of Santiago Roth in Switzerland: An overview. Swiss Journal of Palaeontology, 142, 12. https://doi.org/10.1186/s13358-023-00273-7
Cartelle, C., & Lessa, G. (1988). Descrição de um novo gênero e espécie de Macraucheniidae (Mammalia, Litopterna) do Pleistoceno do Brasil. Paula-Coutiana, 3, 3–26.
Cerdeño, E., Vera, B., Schmidt, G. I., Pujos, F., & Quispe, B. M. (2012). An almost complete skeleton of a new Mesotheriidae (Notoungulata) from the Late Miocene of Casira, Bolivia. Journal of Systematic Palaeontology, 10(2), 341–360. https://doi.org/10.1080/14772019.2011.569576
Christen, Z. M., Sánchez-Villagra, M. R., & Le Verger, K. (2023). Cranial and endocranial comparative anatomy of the Pleistocene glyptodonts from the Santiago Roth Collection. Swiss Journal of Palaeontology, 142, 14. https://doi.org/10.1186/s13358-023-00280-8
Cifelli, R. L. (1993). The phylogeny of the native South American ungulates. In F. S. Szalay, M. J. Novacek, & M. C. McKenna (Eds.), Mammal phylogeny: Placentals (pp. 195–214). Springer-Verlag.
Cione, A. L., Gasparini, G. M., Soibelzon, E., Soibelzon, L. H., & Tonni, E. P. (2015). The great American biotic interchange: A South American perspective. Springer. https://doi.org/10.1007/978-94-017-9792-4
Cione, A. L., & Tonni, E. P. (1995). Chronostratigraphy and “Land-Mammal Ages” in the Cenozoic of southern South America: Principles, practices, and the “Uquian” problem. Journal of Paleontology, 69(1), 135–159. https://doi.org/10.2307/1306287
Cione, A. L., & Tonni, E. P. (1999). Biostratigrapjhy and chronological scale of upper-most Cenozoic in the Pamean Area, Argentina. In J. Rabassa & M. Salemme (Eds.), Quaternary of South America and Antartic Peninsula (pp. 23–51). Rotterdam: A.A. Balkema.
Cione, A. L., Tonni, E. P., & Soibelzon, E. (2003). The broken zig-zag: Late Cenozoic large mammal and tortoise extinction in South America. Revista del Museo Argentino de Ciencias Naturales, 51(1), 1–19.
Croft, D. A. (2016). Horned armadillos and rafting monkeys. The fascinating fossil mammals of South America. Indiana University Press.
Croft, D. A., Gelfo, J. N., & López, G. M. (2020). Splendid innovation: The extinct South American native ungulates. Annual Review of Earth and Planetary Sciences, 48(1), 259–290. https://doi.org/10.1146/annurev-earth-072619-060126
Defler, T. (2019). History of terrestrial mammals in South America: How South American Mammalian fauna changed from mesozoic to recent times (Vol. 42). Springer International Publishing. https://doi.org/10.1007/978-3-319-98449-0
Deraco, V., & García-López, D. A. (2016). A new Eocene Toxodontia (Mammalia, Notoungulata) from northwestern Argentina. Journal of Vertebrate Paleontology, 36(1), e1037884. https://doi.org/10.1080/02724634.2015.1037884
Domingo, L., Tomassini, R. L., Montalvo, C. I., Sanz-Pérez, D., & Alberdi, M. T. (2020). The Great American Biotic Interchange revisited: A new perspective from the stable isotope record of Argentine Pampas fossil mammals. Scientific Reports, 10(1), 1608. https://doi.org/10.1038/s41598-020-58575-6
Dozo, M. T., Martínez, G., & Gelfo, J. N. (2023). Paleoneurology of Litopterna: Digital and Natural Endocranial Casts of Macraucheniidae. In M. T. Dozo, A. Paulina-Carabajal, T. E. Macrini, & S. Walsh (Eds.), Paleoneurology of Amniotes : New Directions in the Study of Fossil Endocasts (pp. 809–836). Cham: Springer International Publishing. https://doi.org/10.1007/978-3-031-13983-3_21
Dozo, M. T., & Vera, B. (2010). First skull and associated postcranial bones of Macraucheniidae (Mammalia, Litopterna) from the Deseadan SALMA (late Oligocene) of Cabeza Blanca (Chubut, Argentina). Journal of Vertebrate Paleontology, 30(6), 1818–1826.
Fariña, R. A., Vizcaíno, S. F., & De Iuliis, G. (2013). Megafauna. Giant Beasts of Pleistocene South America. Indiana University Press.
Fernández de Álvarez, E.H.E. (1940). Descripción de la Macrauchenia patachonica Owen y comparación con otros géneros terciarios (Theosodon, Scalabrinitherium y Promacrauchenia). Publicaciones de la FCEFN, UBA, serie B (Científico-Técnica) 19:1–144 + lam. XXV.
Fernández-Monescillo, M. (2020). Reconstrucción digital de la cavidad endocraneal de Macrauchenia patachonica. In I. Martínez Rivera & O. Rojas Mondaca (Eds.), Macrauchenia Un enigmático animal extinto de la Cuenca de Calama (pp. 39–47). Santiago de Chile: Museo de Historia Natural y Cultural del Desierto de Atacama. https://ri.conicet.gov.ar/handle/11336/146428.
Fernández-Monescillo, M., Antoine, P.-O., Croft, D. A., & Pujos, F. (2023a). Intraspecific craniomandibular and dental analysis of Pseudotypotherium exiguum (Mesotheriidae, Notoungulata) from Monte Hermoso, late Neogene, Buenos Aires Province, Argentina. Journal of Vertebrate Paleontology. https://doi.org/10.1080/02724634.2023.2184269
Fernández-Monescillo, M., Croft, D. A., Pujos, F., & Antoine, P.-O. (2023b). Taxonomic history and intraspecific analysis of Mesotherium cristatum (Mammalia, Notoungulata, Mesotheriidae) from the Early-Middle Pleistocene of Buenos Aires Province, Argentina. Historical Biology, 35(6), 1028–1040. https://doi.org/10.1080/08912963.2022.2074844
Fernández-Monescillo, M., Croft, D. A., Pujos, F., & Antoine, P.-O. (2023c). Resolution of the long-standing controversy over the type species of the genus Pseudotypotherium (Notoungulata, Typotheria, Mesotheriidae). Palaeoworld, 32(3), 547–554. https://doi.org/10.1016/j.palwor.2022.09.009
Fernández-Monescillo, M., Martínez, G., García López, D., Frechen, M., Romero-Lebrón, E., Krapovickas, J. M., et al. (2023d). The last record of the last typotherid (Notoungulata, Mesotheriidae, Mesotherium cristatum) for the middle Pleistocene of the western Pampean region, Córdoba Province, Argentina, and its biostratigraphic implications. Quaternary Science Reviews, 301, 107925. https://doi.org/10.1016/j.quascirev.2022.107925
Fernicola, J. C., Vizcaíno, S. F., & De Iuliis, G. (2009). The fossil mammals collected by Charles Darwin in South America during his travels on board the HMS Beagle. Revista de la Asociación Geológica Argentina, 64(1), 147–159.
Forasiepi, A. M., MacPhee, R. D. E., Hernández del Pino, S., Schmidt, G. I., Amson, E., & Grohé, C. (2016). Exceptional skull of Huayqueriana (Mammalia, Litopterna, Macraucheniidae) from the late Miocene of Argentina: Anatomy, systematics, and paleobiological implications. Bulletin of the American Museum of Natural History, 404, 1–76. https://doi.org/10.5531/sd.sp.23
Gasparini, G. M., Soibelzon, E., Soibelzon, L. H., & Cruz, L. E. (2023). Bioestratigrafía basada en mamíferos: Cambios faunísticos e inferencias paleoclimáticas durante el Cuaternario en la región pampeana (Argentina). Publicación Electrónica de la Asociación Paleontológica Argentina, 23(1), 296–316. https://doi.org/10.5710/PEAPA.29.11.2022.408
Gervais, P. (1847). Observations sur les mammiféres fossiles du midi de la France. Annales de Sciences Naturelles, Zoologie, 3(8), 203–224.
Gervais, P. (1855). Recherches sur les mammifères fossiles de l’Amérique Méridionale. In F. de Castelnau (Ed.), Animaux nouveaux ou rares recueillis pendant l’expedition dans les parties centrales de l’Amérique du Sudde Rio de Janeiro á Lima, et de Lima au Para (pp. 1–63). Paris: P. Bertrand, Librarie-Editeur. Retrieved June 6, 2023 from http://archive.org/details/bub_gb_rAw1AQAAIAAJ
Gervais, H., & Ameghino, F. (1880). Los mamíferos fósiles de la América del Sur. Paris y Buenos Aires: Sabih e Igon.
Guérin, C., & Faure, M. (2004). Macrauchenia patachonica Owen (Mammalia, Litopterna) de la région de São Raimundo Nonato (Piauí, Nordeste brésilien) et la diversité des Macraucheniidae pléistocènes. Geobios, 37(4), 516–535. https://doi.org/10.1016/j.geobios.2003.04.006
Hansen, K. L. (2019). From the shadows out of time. A comparison and assessment of the fossil collections of P.W. Lund and V. Lausen based on historical archives and bone analysis. Additionally, thoughts and guidelines about future avenues for their usage are proposed. Natural History Museum of Denmark, Faculty of Science, University of Copenhagen., Copenhagen.
Hatcher, J. B. (1985). Bone hunters in Patagonia. Ox Bow Press.
Hoffstetter, R. (1986). High Andean Mammalian faunas during the Plio-Pleistocene. In F. Vuilleumier & N. Monasterio (Eds.), High altitude tropical biogeography (pp. 218–245). Oxford University Press.
Huxley, T. (1880). On the application of the laws of evolution to the arrangement of the Vertebrata, and more particularly of the Mammalia. Proceedings of the Zoological Society of London, 43, 649–662.
International Committee on Veterinary Gross Anatomical Nomenclature. (2017). Nomina Anatomica Veterinaria (6th ed.). Editorial Committee Hanover (Germany), Ghent (Belgium), Columbia, MO (U.S.A.), Rio de Janeiro (Brazil).
Jernvall, J. (1995). Mammalian molar cusp patterns: Developmental mechanisms of diversity. Acta Zoologica Fennica, 198, 1–61.
Kerber, L. (2023). Caviomorph rodents from the Pampean region (Argentina) in the historical Santiago Roth Collection in Switzerland. Swiss Journal of Palaeontology, 142, 8. https://doi.org/10.1186/s13358-023-00272-8
Le Verger, K. (2023). Xenarthrans of the collection of Santiago Roth from the Pampean Region of Argentina (Pleistocene), in Zurich. Switzerland. Swiss Journal of Palaeontology, 142, 3. https://doi.org/10.1186/s13358-023-00265-7
Lessa, G. M. (1992). Estudo descritivo de Xenorhinotherius bahiense Cartelle & Lessa e comparação com outras espécies de Macraucheniidae (Litopterna, Mammalia). Universidade Federal do Rio de Janeiro.
Liendo Lazarte, M. (1941). Toxodon: estudio analítico comparativo, osteografía, osteometría. Investigaciones generales en nuevos restos fósiles, conclusiones taxonómicas, filogenéticas, paleobiológicas. Facultad de Ciencias Exactas y Naturales. Universidad de Buenos Aires, Buenos Aires. Retrieved from http://digital.bl.fcen.uba.ar/Download/Tesis/Tesis_0262_LiendoLazarte.pdf
Linnaeus, C. (1758). Systema Naturae per Regna Tria Naturae, Secundum Classes, Ordines, Genera, Species cum Caracteribus, Differentiis, Synonymis, Locis (Tenth Edition., Vol. I). Stockholm: Laurentii Salvii.
Lobo, L. S., Lessa, G., Cartelle, C., & Romano, P. S. R. (2017). Dental eruption sequence and hypsodonty index of a Pleistocene macraucheniid from the Brazilian Intertropical Region. Journal of Paleontology, 91(5), 1083–1090. https://doi.org/10.1017/jpa.2017.54
Madden, R. H. (1990). Miocene Toxodontidae (Notoungulata, Mammalia) from Colombia, Ecuador and Chile (PhD diss.). Duke University.
McGrath, A. J., Anaya, F., & Croft, D. A. (2018). Two new macraucheniids (Mammalia: Litopterna) from the late middle Miocene (Laventan South American Land Mammal Age) of Quebrada Honda, Bolivia. Journal of Vertebrate Paleontology. https://doi.org/10.1080/02724634.2018.1461632
McGrath, A. J., Anaya, F., & Croft, D. (2020a). New Proterotheriids from the middle Miocene of Quebrada Honda, Bolivia, and body size and diversity trends in Proterotheriid and Macraucheniid Litopterns (Mammalia). Ameghiniana, 57(2), 159–188. https://doi.org/10.5710/amgh.03.03.2020.3268
McGrath, A. J., Flynn, J. J., & Wyss, A. R. (2020b). Proterotheriids and macraucheniids (Litopterna: Mammalia) from the Pampa Castillo Fauna, Chile (early Miocene, Santacrucian SALMA) and a new phylogeny of Proterotheriidae. Journal of Systematic Palaeontology, 18(9), 717–738. https://doi.org/10.1080/14772019.2019.1662500
Miño-Boilini, Á. R., Cerdeño, E., & Bond, M. (2006). Revisión del género Toxodon Owen, 1837 (Notoungulata: Toxodontidae) en el Pleistoceno de las provincias de Corrientes, Chaco y Santa Fe, Argentina. Revista Española de Paleontología, 21(2), 93. https://doi.org/10.7203/sjp.21.2.20483
Muizon, C. D., Billet, G., & Ladevèze, S. (2019). New remains of kollpaniine “condylarths” (Panameriungulata) from the early Palaeocene of Bolivia shed light on hypocone origins and molar proportions among ungulate-like placentals. Geodiversitas, 41(25), 841–874. https://doi.org/10.5252/geodiversitas2019v41a25
Muizon, C. D., & Cifelli, R. L. (2000). The “condylarths” (archaic Ungulata, Mammalia) from the early Palaeocene of Tiupampa (Bolivia): Implications on the origin of the South American ungulates. Geodiversitas, 22(1), 47–150.
Nessov, L. A., Archibald, J. D., & Kielan-Jaworoska, Z. (1998). Ungulate-like mammals from the Late Cretaceous of Uzbekistan and a phylogenetic analysis of Ungulatomorpha. Bulletin of Carnegie Museum of Natural, 34, 40–88.
Owen, R. (1837). A description of the cranium of the Toxodon platensis, a gigantic extinct mammiferous species, referrible by its dentition to the Rodentia, but with affinities to the Pachydermata and the herbivorous Cetacea. Proceeding of the Geological Society, 2, 541–542.
Owen, R. (1838). Part 1: Fossil Mammalia. In C. Darwin (Ed.), The Zoology of the Voyage of the HMS Beagle under the command of Captain Fitzroy, during the years 1832 to 1836 (Vol. 1, pp. 1–111). Smith, Elder and Co.
Owen, R. (1853). Description of some species of the extinct genus Nesodon, with remarks on the primary group (Toxodontia) of hoofed quadrupeds, to which that genus is referable. Philosophical Transactions of the Royal Society of London, 143, 291–310. https://doi.org/10.1098/rstl.1853.0011
Prado, J. L., Alberdi, M. T., Azanza, B., & Sánchez, B. (2001). Climate and changes in mammal diversity during the late Pleistocene-Holocene in the Pampean Region (Argentina). Acta Palaeontologica Polonica, 46(2), 261–276.
Parodi, L. J. (1931). Huesos de los miembros de los macroquénidos neoterciarios. Physis, 10, 294–304.
Pascual, R., & Jaureguizar, E. O. (1990). Evolving climates and mammal faunas in cenozoic South America. Journal of Human Evolution, 19(1), 23–60. https://doi.org/10.1016/0047-2484(90)90011-Y
Patterson, B. (1952). Notas acerca del cráneo de un ejemplar juvenil de Mesotherium cristatum. Revista del Museo Municipal de Ciencias Naturales y Tradicional de Mar del Plata, 1, 71–78.
Patterson, B., & Pascual, R. (1968). The fossil mammal fauna of South America. The Quarterly Review of Biology, 43(4), 409–451.
Paula Couto, C. (1979). Tratado de Paleomastozoologia. Academia Brasileira de Ciências.
Prado, J. L., Alberdi, M. T., & Bellinzoni, J. (2021). Pleistocene mammals from Pampean Region (Argentina). Biostratigraphic, biogeographic, and environmental implications. Quaternary, 4(2), 15. https://doi.org/10.3390/quat4020015
Prado, J. L., Martinez-Maza, C., & Alberdi, M. T. (2015). Megafauna extinction in South America: A new chronology for the Argentine Pampas. Palaeogeography, Palaeoclimatology, Palaeoecology, 425, 41–49. https://doi.org/10.1016/j.palaeo.2015.02.026
Prevosti, F. J., Romano, C. O., Forasiepi, A. M., Hemming, S., Bonini, R., Candela, A. M., et al. (2021). New radiometric 40Ar–39Ar dates and faunistic analyses refine evolutionary dynamics of Neogene vertebrate assemblages in southern South America. Scientific Reports, 11(1), 9830. https://doi.org/10.1038/s41598-021-89135-1
Püschel, H. P., Alarcón-Muñoz, J., Soto-Acuña, S., Ugalde, R., Shelley, S. L., & Brusatte, S. L. (2023). Anatomy and phylogeny of a new small macraucheniid (Mammalia: Litopterna) from the Bahía Inglesa Formation (late Miocene), Atacama Region, Northern Chile. Journal of Mammalian Evolution. 30, 415–460. https://doi.org/10.1007/s10914-022-09646-0
Püschel, H. P., & Martinelli, A. G. (2023). More than 100 years of a mistake: On the anatomy of the atlas of the enigmatic Macrauchenia patachonica. Swiss Journal of Palaeontology, 142, 16. https://doi.org/10.1186/s13358-023-00279-1
Roth, S. (1888). Beobachtungen über Entstehung und Alter der Pampasformation in Argentinien. Mit Tafel XXII u. XXIII (pp. 1–114). Berlin Sonderabdruck aus der Zeitschrift der deutschen geologischen Gesellschaft. https://www.biodiversitylibrary.org/item/53403
Roth, S. (1889). Fossiles de la Pampa, Amerique du Sud, collectionné par Santiago Roth. Catalogue No. 5. Zurich: Jean Meyer.
Roth, S. (1895). Rectificaciones sobre la dentición del toxodón. Revista del Museo de La Plata, 6. http://sedici.unlp.edu.ar/handle/10915/97123.
Roth, S. (1898). Catálogo de los mamíferos fósiles conservados en el Museo de La Plata: grupo Ungulata, órden Toxodontia. Revista del Museo de La Plata, 8, parte 1. http://sedici.unlp.edu.ar/handle/10915/99250.
Roth, S. (1903). Los ungulados sudamericanos. Anales del Museo de la Plata, 5, 1–36.
Roth, S. (1904). Noticias preliminares sobre nuevos mamíferos fósiles del Cretácico Superior y Terciario Inferior de la Patagonia. Revista del Museo de la Plata, 11, 135–158.
Roth, S. (1921). Investigaciones geológicas en la llanura argentina. Revista del Museo de la Plata, 25, 135–342.
Sánchez-Villagra, M. R., Bond, M., Reguero, M., & Bartoletti, T. J. (2023). From fossil trader to paleontologist: On Swiss-born naturalist Santiago Roth and his scientific contributions. Swiss Journal of Palaeontology, 142, 19. https://doi.org/10.1186/s13358-023-00282-6
Sánchez-Villagra, M. R., Narita, Y., & Kuratani, S. (2007). Thoracolumbar vertebral number: The first skeletal synapomorphy for afrotherian mammals. Systematics and Biodiversity, 5(1), 1–7. https://doi.org/10.1017/S1477200006002258
Schindelin, J., Arganda-Carreras, I., Frise, E., Kaynig, V., Longair, M., Pietzsch, T., et al. (2012). Fiji: An open-source platform for biological-image analysis. Nature Methods, 9(7), 676–682. https://doi.org/10.1038/nmeth.2019
Schmidt, G. I. (2015). Actualización sistemática y filogenia de los Proterotheriidae (Mammalia, Litopterna) del “Mesopotamiense” (Mioceno tardío) de Entre Ríos, Argentina. Revista Brasileira de Paleontologia, 18(3), 521–546. https://doi.org/10.4072/rbp.2015.3.14
Scott, W. B. (1910). Mammalia of the Santa Cruz Beds. Part I. Litopterna. In William B Scott (Ed.), Reports of the Princeton University Expeditions to Patagonia, 1896–1899 (Vol. 7, pp. 1–156). Princeton and Stuttgart: Schweizerbartsche Verlaghandlung (E. Nagele).
Sefve, I. (1914). Über eine neue Art der Gattung Macrauchenia. Bulletin of the Geological Institutions of the University of Uppsala, 12, 205–256.
Sefve, I. (1925). Macrauchenia patagonica. Bulletin of the Geological Institution of the University of Upsala, 19, 1–21.
Serres, E. (1867). De l’ostéographie de Mesotherium et de ses afffinités zoologiques. Comptes Rendus Hebdomadaires des Séances de l’académie des Sciences, 65, 140–148.
Shockey, B. J., White, E., Anaya, F., & McGrath, A. (2022). A new proterotheriid (Mammalia, Litopterna) from the Salla Beds of Bolivia (upper Oligocene): phylogeny and litoptern patellar pit knee locks. Journal of Vertebrate Paleontology, 42(2), e2162409. https://doi.org/10.1080/02724634.2022.2162409
Simpson, G. G. (1940). The names Mesotherium and Typotherium. American Journal of Science, 238(7), 518–521. https://doi.org/10.2475/ajs.238.7.518
Simpson, G. G. (1980). Splendid Isolation. The Curious History of South American Mammals. Yale University Press.
Soria, M. F. (1981). Los Litopterna del Colhuehuapense (Oligoceno tardío) de la Argentina. Revista del Museo Argentino de Ciencias Naturales “Bernardino Rivadavia,” 3, 1–54.
Soria, M. F. (2001). Los Proterotheriidae (Litopterna, Mammalia), sistemática, origen y filogenia. Monografías del Museo Argentino de Ciencias Naturales, 1, 1–167.
Vera, B. (2015). Phylogenetic revision of the South American notopithecines (Mammalia: Notoungulata). Journal of Systematic Palaeontology, (August), 1–20. https://doi.org/10.1080/14772019.2015.1066454
Vera, B., González Ruiz, L., Novo, N., Martin, G., Reato, A., & Tejedor, M. F. (2019). The Interatheriinae (Mammalia, Notoungulata) of the Friasian sensu stricto and Mayoan (middle to late Miocene), and the fossils from Cerro Zeballos, Patagonia, Argentina. Journal of Systematic Palaeontology, 17(13), 923–943. https://doi.org/10.1080/14772019.2018.1511387
Villafañe, A. L., Ortiz-Jaureguizar, E., & Bond, M. (2006). Cambios en la riqueza taxonómica y en las tasas de primera y última aparición de los Proterotheriidae (Mammalia, Litopterna) durante el Cenozoico. Estudios Geológicos, 62(1), 155–166.
Vizcaíno, S. F., Iuliis, G. D., Brinkman, P. D., Kay, R. F., & Brinkman, D. L. (2017). On an album of photographs recording fossils in the “Old collections” of the Museo de la Plata and Ameghino’s private collection at the beginning of the XXTH century. Publicación Electrónica de la Asociación Paleontológica Argentina, 17(1). https://doi.org/10.5710/PEAPA.21.06.2017.244
Voglino, D. (2020). Sitios geológicos y paleontológicos estudiados por Santiago Roth en el centro-este de la Argentina. Museo de Ciencias Naturales A. Scasso, 1–23.
Voglino, D., Carrillo-Briceño, J. D., Furrer, H., Balcarcel, A., Rangel de Lazaro, G., Aguirre Fernández, G., & Forasiepi, A. (2023). Pampean megamammals in Europe: The fossil collections from Santiago Roth. Swiss Journal of Palaeontology. https://doi.org/10.1186/s13358-023-00283-5
Welker, F., Collins, M. J., Thomas, J. A., Wadsley, M., Brace, S., Cappellini, E., et al. (2015). Ancient proteins resolve the evolutionary history of Darwin’s South American ungulates. Nature, 522, 81–84. https://doi.org/10.1038/nature14249
Westbury, M., Baleka, S., Barlow, A., Hartmann, S., Paijmans, J. L. A., Kramarz, A., et al. (2017). A mitogenomic timetree for Darwin’s enigmatic South American mammal Macrauchenia patachonica. Nature Communications, 8, 15951. https://doi.org/10.1038/ncomms15951
Zittel, K. A. von. (1893). Handbuch der palaeontologie. Paleozoologie. IV. Band, Vertebrata (Mammalia) (Vol. 4, pp. 1–836). München: R. Oldenbourg. https://www.biodiversitylibrary.org/item/124990
Zurita-Altamirano, D., Buffetaut, E., Forasiepi, A. M., Kramarz, A., Carrillo, J. D., Aguirre-Fernández, G., et al. (2019). The Allemann collection from the Santa Cruz Formation (late early Miocene), Argentina, in Zurich, Switzerland. Swiss Journal of Palaeontology, 138, 259–275. https://doi.org/10.1007/s13358-019-00185-5
Acknowledgements
We thank the curators of the following institutions for their kind assistance during museum visits of H.P.P.: Lionel Cavin (MHNG), Laura Chornogubsky (MACN), Agustín Martinelli (MACN), Marcelo Reguero (MLP), Susana Bargo (MLP), Guillaume Billet (MNHN), Gabriel Aguirre (PIMUZ), Marcelo Sánchez-Villagra (PIMUZ), Daniel Brinkman (YPM VPPU), and Vanessa Rhue (YPM VPPU). We thank Jorge Carrillo-Briceño for sharing pictures of specimens of the MHNG GEPI V collection that were useful in figuring and describing the specimens. We thank Gabriel Aguirre Fernández and Analía Forasiepi, editors of the special issue, and two anonymous reviewers for their valuable comments.
Funding
J.D.C is supported by the Swiss National Science Foundation Grant TMPFP2_209818. This work was also funded by the SwissCollNet grant SCN133-ZH to Gabriel Aguirre Fernández.
Author information
Authors and Affiliations
Contributions
JDC and HPP designed the study, identified the specimens, wrote the manuscript and prepared the figures. The two authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
We declare no competing interests.
Additional information
Editorial handling: Analía M. Forasiepi
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Carrillo, J.D., Püschel, H.P. Pleistocene South American native ungulates (Notoungulata and Litopterna) of the historical Roth collections in Switzerland, from the Pampean Region of Argentina. Swiss J Palaeontol 142, 28 (2023). https://doi.org/10.1186/s13358-023-00291-5
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13358-023-00291-5
Keywords
- Pleistocene
- Argentina
- Santiago Roth
- Palaeontological collections
- Switzerland
- Notoungulata
- Litopterna
- Mesotherium
- Toxodon
- Macrauchenia