
Abstract
In this paper, we explore a stochastic non-autonomous one-predator–two-prey system with Beddington–DeAngelis functional response and impulsive perturbations. First, by using Itô’s formula, exponential martingale inequality, Chebyshev’s inequality and other mathematical skills, we establish some sufficient conditions for extinction, non-persistence in the mean, weak persistence, persistence in the mean and stochastic permanence of the solution of the stochastic system. Then the limit of the average in time of the sample path of the solution is estimated by two constants. Afterwards, the lower-growth rate and the upper-growth rate of the positive solution are estimated. In addition, sufficient conditions for global attractivity of the system are established. Finally, we carry out some simulations to verify our main results and explain the biological implications: the large stochastic interference is disadvantageous for the persistence of the population and the strong impulsive harvesting can lead to extinct of the population.
Similar content being viewed by others


1 Introduction
Predator–prey systems, competitive systems and cooperative systems, the three major systems in the ecosystem, play a vital role in promoting the stable operation of biological communities. Among them, predation and competition are the most common phenomena in nature, such as, tiger hunting rabbits, wolves catching deer, two trees in the same forest, eagle and snake feeding on the same mouse and so on. Many scholars have studied predation and competition systems (see [1,2,3,4,5,6,7,8,9,10,11,12,13]). Among them, the one-predator–two-prey system (see [14,15,16,17,18]) is the most common system in the ecosystem. Therefore, it is important and meaningful to consider dynamical behavior of the one-predator–two-prey system with interspecies competition. When modeling the one-predator–two-prey system, one of the most important factors should be involved is the functional response mechanism, which changes the prey density per unit time per predator as a function of prey or both prey and predator species. There are many kinds of famous functional response in the predator–prey system reported in the previous references, such as Holling types [19,20,21], Beddington–DeAngelis type [22,23,24,25], Michaelis–Menton type [26], Ivlev type [27], Hassell–Varley type [28], Crowley–Martin type functional response [29], which are suitable for different kinds of predator–prey systems, respectively. In 1975, Beddington [22] and DeAngelis [23] first introduced the Beddington–DeAngelis type predator–prey model taking the form
where x and y denote the population densities of prey and predator, respectively. The term \(\frac{c_{1}x}{a_{1}+a_{2} x+a_{3}y}\) represents the Beddington–DeAngelis functional response, which turns into the Holling-II functional response if \(a_{3} = 0\) and linear functional response if both \(a_{2} = 0\) and \(a_{3} = 0\). That is to say, the B-D functional response is affected by both predator and prey. Therefore, the effect of mutual interference on the dynamics of population is worth studying.
On the other hand, the population systems in the real world are always inevitably influenced by all kinds of environmental noises which are an important component in an ecosystem. Usually, there are two types of environmental noises: white noise and color noise. White noise arises from a nearly continuous series of small or moderate perturbations that have small effects on the intrinsic growth rates of the species. Therefore, it is essential to reveal how the environmental noise disturbs the population systems. In recent years, many scholars have proposed and investigated stochastic models with white noise perturbations, please refer to [30,31,32,33,34,35,36,37,38,39,40,41,42,43,44,45,46] and the references therein. For example, Ji et al. [30] considered a predator–prey model with modified Leslie–Gower and Holling type II schemes with stochastic perturbation and the condition for persistence and extinction of the system is established. Liu and Wang in [36] discussed a predator–prey system with Beddington–DeAngelis functional response with stochastic perturbation. They demonstrated that if the positive equilibrium of the deterministic system is globally stable, then the stochastic model will preserve this nice property provided the noise is sufficiently small.
However, periodic behavior often arises in implicit ways in various natural phenomena. For example, due to the seasonal variation, hunting, harvesting and so on, the birth rate, the mortality rate and other parameters in the population systems will not remain constant, but exhibit a more-or-less periodicity. Thus, it is natural to model the population by a periodic environment. Therefore, numerous authors have investigated the effect of seasonal variation and stochasticity (see [47,48,49]).
Furthermore, population growth in ecosystems is also affected by human activities, such as periodic harvesting or stocking for the species, which cannot be considered continuously. Stochastic systems that consider continuous phenomena are not suitable for these phenomena. Therefore, in this case, we should consider the effect of impulse in order to describe these phenomena more accurately. In recent decades, a variety of population dynamical systems with impulsive effects have been proposed and studied extensively (see [50,51,52,53,54,55,56]). For example, in [50] Liu and Wang concerned with an n-species stochastic nonautonomous Lotka-Volterra competitive system with impulsive effects. They obtained the sufficient conditions for stochastic permanence, extinction and global stability and investigated some dynamical properties. Zhang and Meng et al. [52] discussed a stochastic non-autonomous predator–prey system with impulsive effect. They concluded that the large stochastic disturbances can lead to the extinction of the population, and large impulse harvests can also result in the extinction of the population.
Taking all above influences into consideration, we focus on the stochastic non-autonomous one-predator–two-prey system with the Beddington–DeAngelis functional response and impulsive perturbations
where \(x_{i}(t)\) is the size of the ith population at time t, \(r_{i}(t)\) represents the intrinsic growth rate of the ith population, \(\alpha _{i}(t)\) stands for the density-dependent coefficients of the ith population, \(\beta _{1}(t)\) and \(\beta _{2}(t)\) are the competitive coefficient of \(x_{1}(t)\) and \(x_{2}(t)\), respectively, \(c_{j}(t)\) is the capturing rate of predator, \(e_{j}(t)\) represents the rate of conversion of nutrients into the reproduction of predator, \(B_{i}(t)\) (\(i=1,2,3\)) is for independent standard Brownian motions defined on a complete probability space and \(\sigma _{i}(t)\) is for the intensities of \(B_{i}(t)\). \(r_{i}(t)\), \(\alpha _{i}(t)\), \(\beta _{j}(t)\), \(a_{i}(t)\), \(b_{i}(t)\), \(c_{j}(t)\), \(e_{j}(t)\), \(\sigma _{i}(t)\) are positive, continuous and bounded functions defined on \(\mathbb{R}^{+}=(0, \infty )\), N denotes the set of positive integers, \(0< t_{1}< t_{2}<\cdots\) , \(\lim_{k\rightarrow +\infty }t_{k}=+\infty \), \(i=1, 2,3\), \(j=1, 2\), \(k \in N\).
We impose the following restriction on system (2) which is a reasonable way for giving biological meaning: \(h_{ik}+1>0\), \(i=1, 2,3\), \(k\in N\). When \(h_{ik} > 0\), the impulsive effects represent releasing the specie, but if \(h_{ik} < 0\), the impulsive effects denote harvesting for the ith population.
The main goals of this paper are to investigate how impulsive perturbations and the white noises affect the permanence, persistence, extinction and global attractivity of system (2). The rest of the paper is organized as follows. In Sect. 2, we give some definition and prove the existence of a unique positive solution of the system. In Sect. 3, we will derive main theoretical results of this paper, such as sufficient conditions for the extinction, non-persistence in the mean, weak persistence, persistence in the mean and stochastic permanence of the system. Meanwhile the limit of the average in time of the sample path of the solution is estimated by two constants. In Sect. 4, the lower-growth rate and the upper-growth rate of the solutions are estimated. In Sect. 5, we investigate the global attractivity of the system. In Sect. 6, we give the conclusions and several examples and numerical simulations to illustrate our theoretical results.
2 Preliminary
Let \((\varOmega ,\mathcal{F}, \{\mathcal{F}\}_{t\geq 0}, \mathcal{P})\) be a complete probability space with a filtration \(\{\mathcal{F}_{t}\} _{t\geq 0}\) satisfying the common conditions (i.e. it is increasing and right continuous while \(\mathcal{F}_{0}\) contains all \(\mathcal{P}\)-null sets). Let \(B(t)=(B_{1}(t),B_{2}(t),B_{3}(t))^{T}\) be an n-dimensional Brownian motion defined on this probability space. Let \(\mathbb{R} _{+}^{3}=\{x\in \mathbb{R}^{3}:x_{i}>0 ,i\leq i\leq 3\}\). We define the norm as \(|x|=\sqrt{x_{1}^{2}+x_{2}^{2}+x_{3}^{2}}\).
If \(f(t)\) is a bounded continuous function on \([0,+\infty )\), define
For the constants \(m_{i}\), \(M_{i}\), \(f^{u}_{i}\), \(f^{l}_{i}\) (\(i=1,2,3\)), we denote
Definition 2.1
-
1.
\(x(t)\) is said to be extinctive if \(\lim_{t\rightarrow +\infty }x(t)=0\).
-
2.
\(x(t)\) is said to be non-persistent in the mean if \(\lim_{t\rightarrow +\infty }\frac{\int _{0}^{t}x(s)\,ds}{t}=0\).
-
3.
\(x(t)\) is said to be weakly persistent if \(\limsup_{t\rightarrow +\infty }x(t)>0\).
-
4.
\(x(t)\) is said to be persistent in the mean if \(\liminf_{t\rightarrow +\infty }\frac{\int _{0}^{t}x(s)\,ds}{t}>0\).
-
5.
\(x(t)\) is said to be stochastically permanent if for every \(\varepsilon \in (0,1)\) there are two constants \(\beta >0\), \(\delta >0\) such that
$$ \liminf_{t\rightarrow +\infty }\mathbb{P} \bigl\{ x(t)\geq \beta \bigr\} \geq 1- \varepsilon ,\qquad \liminf_{t\rightarrow +\infty }\mathbb{P}\bigl\{ x(t) \leq \delta \bigr\} \geq 1-\varepsilon . $$
Now we give an assumption which will be used in the following proof.
Assumption 2.1
There exist constants \(m_{i}\) and \(M_{i}\) (\(i = 1,2,3\)) such that
Remark 1
Assumption 2.1 is easy to satisfy. For example, if \(h_{ik}=e^{\frac{(-1)^{k+1}}{k^{2}}}-1\), \(i=1,2,3\), then \(1\leq \prod_{0< t_{k}< t}(1+h_{ik})\leq e\).
Theorem 2.1
For any given initial value \((x_{1}(0),x_{2}(0),x_{3}(0))^{T}\in \mathbb{R}_{+}^{3}\), system (2) exists a unique positive solution \(x(t)=(x_{1}(t),x_{2}(t),x_{3}(t))^{T}\) on \(\mathbb{R}^{+}\) and the positive solution will remain \(\mathbb{R}_{+}^{3}\) a.s.
Proof
Consider the following stochastic differential equations (SDEs) without impulses:
with initial value \((y_{1}(0),y_{2}(0),y_{3}(0))^{T}=(x_{1}(0),x_{2}(0),x _{3}(0))^{T}\). By the classic theory of SDEs without impulses (see [57]), system (3) has a unique global positive solution \(y(t)=(y_{1}(t),y_{2}(t),y_{3}(t))^{T}\).
Let \(x_{i}(t)=\prod_{0< t_{k}< t}(1+h_{ik})y_{i}(t)\), we show that \(x(t)=(x_{1}(t),x_{2}(t),x_{3}(t))^{T}\) is the solution of system (2) with initial value \((x_{1}(0),x_{2}(0),x_{3}(0))^{T}\).
In fact, since \(x_{1}(t)\) is continuous on each interval \((t_{k}, t _{k+1})\subset \mathbb{R}^{+}\) and for \(t\neq t_{k}\), \(k\in N\), we have
Similarly, we can obtain
And for each \(t_{k}\in \mathbb{R}^{+}\), it is not difficult to show that
Moreover,
Therefore\(,x(t)=(x_{1}(t),x_{2}(t),x_{3}(t))^{T}\) is the unique global positive solution of system (2). This completes the proof of Theorem 2.1. □
3 Extinction and persistence
In this section we will derive sufficient conditions for the extinction, non-persistence in the mean, weak persistence, persistence in the mean and stochastic permanence of the solutions of system (2).
Theorem 3.1
Suppose that \(x(t)=(x_{1}(t),x_{2}(t),x_{3}(t))^{T}\) is a solution of system (2), then
where
Particularly, if \(\delta _{i}^{*}<0\), then \(\lim_{t\rightarrow +\infty }x_{i}(t)=0\) a.s., namely, the ith species (\(i = 1, 2, 3\)) in system (2) is extinct.
Proof
Applying Itô’s formula to the first equation of system (3), we could find that
In the same way, combining with the last two equations of system (3) we have
which leads to
Integrating both sides of inequalities (4), (5) and (6) on the interval \([0,t]\), one can easily see that
where \(M_{i}(t)=\int _{0}^{t}\sigma _{i}(s)\,dB_{i}(s)\), \(i=1,2,3\). Note that \(M_{i}(t)\) are local martingales, whose quadratic variations are \(\langle M_{i}(t),M_{i}(t)\rangle =\int _{0}^{t}\sigma _{i}^{2}(s)\,ds \leq (\sigma _{i}^{u})^{2}t\). Making use of the strong law of large numbers for local martingales (see [58]) results in
On the other hand, it follows from (7) that
In other words, we can compute that
Taking superior limit on both sides of (8) and noting that \(\lim_{t\rightarrow +\infty }\frac{\ln y_{i}(0)}{t}=0\), we obtain
This completes the proof. □
Theorem 3.2
Suppose that \(x(t)=(x_{1}(t),x_{2}(t),x_{3}(t))^{T}\) is a solution of system (2), then
Particularly, if \(\delta _{i}^{*}= 0\), then \(\lim_{t\rightarrow + \infty }\frac{1}{t}\int _{0}^{t}x_{i}(s)\,ds=0\), that is, the ith species (\(i = 1, 2, 3\)) in system (2) is non-persistent in the mean.
Proof
According to the definition of the limit, for arbitrary fixed \(\epsilon _{i} > 0\), there exists a constant \(T_{0} > 0\), for every \(t>T_{0}\), such that
Substituting above inequalities into (8) yields
for all \(t > T_{0}\), where \(\lambda _{i}=\delta _{i}^{*}+\epsilon _{i}\).
Denote \(g_{i}(t)=\int _{0}^{t}x_{i}(s)\,ds\), we get \(\frac{dg_{i}(t)}{dt}=x_{i}(t)\). Taking exponent on both sides of (9), we can show that
Integrating inequality (10) from \(T_{0}\) to t yields
Taking logarithm of both sides of inequality (11), we can derive that
Taking superior limit on (12) elicits that
Then it follows from L’Hospital’s rule that
This completes the proof of this theorem. □
Theorem 3.3
Suppose that \(x(t)=(x_{1}(t),x_{2}(t),x_{3}(t))^{T}\) is a solution of system (2). If \(\delta _{i}^{*}>0\), then the ith species (\(i = 1, 2, 3\)) in system (2) is weakly persistent a.s., i.e\(.\limsup_{t\rightarrow +\infty }x_{i}(t)>0\) a.s.
Proof
If this assertion is not true, then \(P(S)>0\), where S is the set \(S=\limsup_{t\rightarrow +\infty }x_{i}(t)=0\). It follows from (8) that
On the other hand, for \(\forall \omega \in S\), we have \(\lim_{t\rightarrow +\infty }x_{i}(t,\omega )=0\). Thus it follows from the boundedness of \(\alpha _{i}(t)\) that
Substituting these inequalities into (13) and making use of \(\lim_{t\rightarrow +\infty }\frac{M_{i}(t)}{t}=0\) a.s., one can obtain the contradiction \(0\geq \limsup_{t\rightarrow +\infty }\ln x_{i}(t, \omega )=\delta _{i}^{*}>0\). This completes the proof. □
Remark 2
Theorems 3.1–3.3 have an interesting biological interpretation. Observe that the extinction and persistence of species \(x_{i}(t)\) only depend on \(\delta _{i}^{*}\). If \(\delta _{i}^{*}> 0\), the population \(x_{i}(t)\) is weakly persistent. If \(\delta _{i}^{*}< 0\), the population \(x_{i}(t)\) goes to extinction.
Theorem 3.4
Suppose that \(x(t)=(x_{1}(t),x_{2}(t),x_{3}(t))^{T}\) is a solution of system (2), then
where
Particularly, if \({\theta _{i}}_{*}>0\), then the ith species (\(i = 1, 2, 3\)) in system (2) is persistent in the mean a.s.
Proof
Applying Itô’s formula to the first equation of system (3), we can observe that
Applying general calculations to (14), it is easy to verify that
It then follows from Theorem 3.2 that
Since \(\lim_{t\rightarrow +\infty }\frac{y_{i}(0)}{t}=0\), \(\lim_{t\rightarrow +\infty }\frac{M_{i}(t)}{t}=0\), \(i=1,2,3\), for \(\forall \epsilon _{1}>0\) there exists a \(T_{1} > 0\), such that
Substituting the above inequalities into (15), we get, for \(t>T_{1}\),
where \(\theta _{1}={\delta _{1}}_{*}- (\frac{\beta _{1}^{u}\delta _{2}^{*}}{\alpha _{2}^{l}}+\eta _{1} )-\epsilon _{1}\), and \(\eta _{1}=\min \{\frac{c_{1}^{u}}{a_{3}^{l}},\frac{c_{1}^{u} \delta _{3}^{*}}{a_{1}^{l}\alpha _{3}^{l}} \}\).
In the similar way, we can conclude that, for any \(\epsilon _{i}\), there exists some \(T_{i} > 0\) such that
where
and
Let \(T^{*}= \min_{1\leq i\leq 3}T_{i} > 0\), then from (16) and (17), we can easily see that
Denote \(g_{i}(t)=\int _{0}^{t}x_{i}(s)\,ds\), we get \(\frac{dg_{i}(t)}{dt}=x_{i}(t)\). Taking the exponent on both sides of (18), we can obtain
Integrating inequality (19) from \(T^{*}\) to t yields
Taking logarithm of both sides of inequality (20), it can be verified straightforwardly that
Taking superior limit on both sides of (21), we obtain
Then it follows from L’Hospital’s rule that
This completes the proof of this theorem. □
Theorem 3.5
If Assumption 2.1 holds and \((\check{\sigma })^{2}<2\hat{r}\), then system (2) is stochastically permanent.
Proof
First, we prove that, for arbitrary \(\varepsilon > 0\), there exists a constant \(\beta > 0\) such that
Define
where \(U(y)=\sum^{3}_{i=1}y_{i}(t)\), \(\varrho >0\), κ is a positive constant to be determined.
Applying Itô’s formula and system (3) once again, we can calculate that
Thus,
Substituting inequality \(y_{i}(t)y_{j}(t)\leq \frac{y_{i}^{2}(t)+y _{j}^{2}(t)}{2}\) (\(i,j=1,2,3\)) into the above inequality and making some estimations yield
where \(\phi = (\check{\alpha }+2\check{\beta }+\frac{{c_{1}^{u}}}{ {a_{1}^{l}}}+\frac{{c_{2}^{u}}}{{b_{1}^{l}}} )\).
Further, when \(y_{i}>0\), \(\sum^{3}_{i=1}y_{i}^{2}(t)<(\sum^{3}_{i=1}y _{i}(t))^{2}=U^{2}\), then from (22), we can derive that
On the other hand, it follows from Itô’s integration by parts formula and applying (23) that
We can choose positive constant κ small enough such that
Then
where
By the definition of κ, \(H(y)\) is upper bounded in \(\mathbb{R}^{+}\), we let \(H=\sup_{y\in \mathbb{R}^{+}}H(y)<+\infty \), we could find that
Integrating inequality (25) on the interval \([0, t]\), then multiplying \(e^{-\kappa t}\) and taking expectations on both sides, it is not difficult to show that
where \(y_{0}=\sum^{3}_{i=1}y_{i}(0)\). Thus,
On the other hand, since \(m\leq m_{i}\leq \prod_{0< t_{k}< t}(1+h_{ik})\leq M_{i}\leq M\) and by the previous transformation \(x_{i}(t)=\prod_{0< t_{k}< t}(1+h_{ik})y _{i}(t)\), we have
which yields
Consequently,
which leads to
Then, for any \(\varepsilon >0\), set \(\beta = (\frac{\kappa m^{2 \varrho }\varepsilon }{4^{\varrho }H} )^{\frac{1}{\varrho }}\), it follows from Chebyshev’s inequality (see [57]) that
In other words,
Next we show that, for arbitrary \(\varepsilon > 0\), there exists a constant \(\delta > 0\) such that
Let \(q>2\), applying Itô’s formula to the non-impulsive system (3),
Integrating (28) on the interval \([0, t]\) yields
Taking expectations on both sides of (29) we obtain
Denote
then we have
and
It is easy to see that \(g(y_{1})\) has a unique maximum \(y_{1}^{*}=\frac{1+q r_{1}^{u}+\frac{q(q-1)(\sigma _{1}^{u})^{2}}{2}}{(q+1) \alpha _{1}^{l}m_{1}}\) since
Therefore,
which yields
On the other hand, by applying Itô’s formula and the last two equations of system (3) then making some estimations, we can easily see that
Then, similar to the above discussions, we can also derive that
where
Combining (29) and (30), we can conclude that
Multiplying \(e^{-t}\) on both sides of (31) and taking the superior limit yield
This leads to
Then, for any \(\varepsilon >0\), let \(\delta =\sqrt{\frac{\varTheta }{ \varepsilon }}\), it follows from Chebyshev’s inequality that
where \(\varTheta =\sum_{i=1}^{3}\varTheta _{i}(q)(M_{i})^{2}\). As a consequence,
According to Definition 2.1, it follows from (27) and (33) that system (2) is stochastically permanent. □
Remark 3
From inequality (32), we can get
Therefore, system (2) has the property
4 Asymptotic properties
In this section we will discuss the asymptotic properties of the solution of system (2).
Theorem 4.1
If Assumption 2.1 holds and any solution \(x(t)=(x_{1}(t),x _{2}(t),x_{3}(t))^{T}\) of system (2) has the property that
and, moreover\(, 2\hat{r}-(\check{\sigma })^{2}>0\), then
Proof
It follows from Itô’s formula and combining with inequality (4), (5) and (6) that
Integrating above inequality (34) on the interval \([0, t]\) yields
where \(N_{i}(t)=\int _{0}^{t}e^{t}\sigma _{i}(s)\,dB_{i}(s)\) is the exponential martingale, whose quadratic variation is
Thus, it follows from the exponential martingale inequality (see [57]) that
By virtue of the Borel–Cantelli lemma, for almost all \(\omega \in \varOmega \), there exists \(k_{0}(\omega )\) such that, for every \(k\geq k_{0}(\omega )\),
for \(0\leq t\leq k\gamma \). Substituting inequality (36) into (35) and making some estimations yield
If we denote
Then \(f'(y_{i})=\frac{1}{y_{i}}-m_{i}\alpha _{i}^{l}\), \(f''(y_{i})=-\frac{1}{y _{i}^{2}}<0\), this means \(y_{i}^{*}=\frac{1}{m_{i}\alpha _{i}^{l}}\) is the unique maximum of the function \(f(y_{i})\), i.e\(.f(y_{i})\leq f(y _{i}^{*})\).
Thus,
Multiplying \(e^{-t}\) on both sides of (37) yields
For \((k-1)\gamma \leq t\leq k\gamma \) and \(k\geq k_{0}(\omega )\), if \(t\rightarrow \infty \), then \(k\rightarrow \infty \).
Therefore,
Let \(\rho \rightarrow 1\) and \(\gamma \rightarrow 0\), then \(\limsup_{t\rightarrow +\infty }\frac{\ln y_{i}(t)}{\ln t}\leq 1\). Since Assumption 2.1 holds,
Now, we prove the next part. By (26), there exists a constant \(C_{1} > 0\) such that
At the same time, it follows from (24) that
where \(C_{2}=\max \{|2\hat{r}-(2\varrho +1)(\check{\sigma })^{2}|,M \phi ,|3(\check{\sigma })^{2}-2\hat{r}|\}\). Let \(\mu > 0\) be sufficiently small for
Let \(k = 1, 2,\ldots \) , making use of (39) shows that
We compute that
On the other hand, by the famous Burkholder–Davis–Gundy inequality (see [57]), it is easy to derive that
Substituting (43) and (42) into (41) results in
Applying (38) and (40), we can show that
Let \(\epsilon > 0\) be arbitrary. Then, by the Chebyshev inequality, we obtain
By the Borel–Cantelli lemma [59], for almost all \(\omega \in \varOmega \), there exists an integer \(k_{0} = k_{0}(\omega )\) such that
for \(k\geq k-0\) and \((k-1)\mu \leq t\leq k\mu \). That is to say
Letting \(\epsilon \rightarrow 0\) gives
Consequently,
But this holds for any ϱ that satisfies \(2\hat{r}>(2\varrho +1)(\check{\sigma })^{2}\), we therefore have
It then follows that
This completes the proof of this theorem. □
Remark 4
Theorem 4.1 shows that, for any \(\epsilon > 0\), there exists a random variable \(T_{\epsilon }>0\) such that \(t^{-\frac{1}{2\hat{r}-(\check{\sigma })^{2}}+\epsilon }\leq |x(t)| \leq t^{1+\epsilon }\) for \(t\geq T_{\epsilon }\) almost surely. That is to say, the solution will not decay faster than \(t^{-\frac{1}{2 \hat{r}-(\check{\sigma })^{2}}+\epsilon }\) and will not grow faster than \(t^{1+\epsilon }\) with probability one. We are now in the position to estimate the limit of the average in time of the sample paths of solutions.
5 Global attractivity
In this section we give the definition of global attractivity and some useful lemmas to study the global attractivity of system (2).
Definition 5.1
Let \(x(t)=(x_{1}(t),x_{2}(t),x_{3}(t))^{T}\), \(z(t)=(z_{1}(t),z_{2}(t),z _{3}(t))^{T}\) be two arbitrary solutions of system (2) with initial values \(x(0),z(0)\in \mathbb{R}^{+}\), respectively. If \(\lim_{t\rightarrow +\infty }|x(t)-z(t)|=0\) a.s., then we say system (2) is globally attractive.
Lemma 5.1
(see [60])
Let \(X(t)\) be an n-dimensional stochastic process on \(t \geq 0\). Suppose that there exist positive constants α, β, c such that
Then there exists a continuous modification \(\overline{{X}}(t)\) of \(X(t)\) which has the property that for every \(\vartheta \in (0,\frac{ \beta }{\alpha } )\) there is a positive random variable \(h(\omega )\) such that
In other words, almost every sample path of \(\overline{{X}}(t)\) is locally but uniformly Hölder continuous with exponent ϑ.
Lemma 5.2
(see [60])
Let Assumption 2.1 hold. If \(y(t)=(y_{1}(t),y _{2}(t),y_{3}(t))^{T}\) is a solution of (3) with initial values \(y(0)\in \mathbb{R}^{+}\), then almost every sample path of \(y_{i}(t)\) (\(1 \leq i\leq 3\)) is uniformly continuous for \(t\geq 0\).
Proof
By (32), there exists \(T>0\), such that \(\mathbb{E}[y_{i}^{q}(t)] \leq \frac{3}{2}\varTheta _{1}(q)\) for all \(t\geq T\). Moreover, it follows from the continuity of \(\mathbb{E}[y_{i}^{q}(t)]\) that there is a \(\varTheta _{2}(q)>0\) such that \(\mathbb{E}[y_{i}^{q}(t)]\leq \varTheta _{2}(q)\) for \(t\geq T\). Let \(\varTheta (q)=\max \{\frac{3}{2}\varTheta _{1}(q), \varTheta _{2}(q) \}\), then, for all \(t\leq 0\),
Clearly, the first equation of system (3) is equivalent to the following equation:
Therefore,
By the famous moment inequality for stochastic integrals (see [58]), we obtain, for \(0\leq t_{1}\leq t_{2}\) and \(q>2\),
Then, for \(0< t_{1}< t_{2}<\infty \), \(t_{2}-t_{1}\leq 1\), \(\frac{1}{q}+ \frac{1}{p}=1\), we can derive that
where \(G_{2}(q)=\max \{G_{1}(q),[(\sigma _{1}^{2})^{u}]^{q} \varTheta (q) \}\). Then it follows from Lemma 5.1 that almost every sample path of \(y_{1}(t)\) is locally but uniformly Hölder continuous with exponent ϑ for every \(\vartheta \in (0,\frac{q-2}{2q} )\) and therefore almost every sample path of \(y_{1}(t)\) is uniformly continuous on \(t\geq 0\). Similarly, we can show that almost every sample path of \(y_{2}(t)\) and \(y_{3}(t)\) are uniformly continuous on \(t\geq 0\). □
Lemma 5.3
(see [61])
Let f be a non-negative function defined on \(t\geq 0\) such that f is integrable on \(t\geq 0\) and is uniformly continuous on \(t\geq 0\). Then \(\lim_{t\rightarrow +\infty }f(t)=0\).
Theorem 5.1
If Assumption 2.1 holds and
then system (2) is globally attractive.
Proof
Let \(x(t)=(x_{1}(t),x_{2}(t),x_{3}(t))^{T}\), \(z(t)=(z_{1}(t),z_{2}(t),z _{3}(t))^{T}\) be two arbitrary solutions of system (2) with initial values \(x(0),z(0)\in \mathbb{R}^{+}\), respectively. Let \(y(t)=(y_{1}(t),y_{2}(t),y_{3}(t))^{T}\), \(\overline{y}(t)=( \overline{y}_{1}(t),\overline{y}_{2}(t),\overline{y}_{3}(t))^{T}\) be two arbitrary solution of system (3) with initial values \(y(0),\overline{y}(0)\in \mathbb{R}^{+}\), respectively.
Then
Define
By Itô’s formula
Integrating both sides gives
Therefore
Making use of \(\overline{V}(t)\geq 0\) and (44) results in
Consequently, by Lemmas 5.2 and 5.3, one can observe that
Then
This completes the proof. □
6 Conclusion and numerical simulations
In this paper, a stochastic non-autonomous one-predator–two-prey system with Beddington–DeAngelis functional response and impulsive perturbations is proposed and investigated. First, we obtain some sufficient conditions for extinction, non-persistence in the mean, weak persistence, persistence in the mean and stochastic permanence of the solution, and we verify some asymptotic behaviors of the solutions of system (2), such as the limit of the average in time, the lower-growth rate, the upper-growth rate and global attractivity. Now we summarize the key results as follows:
(I):
-
(1)
If \(\delta _{i}^{*}=\limsup_{t\rightarrow +\infty } \frac{1}{t} [\sum_{0< t_{k}< t}\ln (1+h_{ik})+\int _{0}^{t}\delta _{i}(s)\,ds ]<0\), then the ith species (\(i = 1, 2, 3\)) in system (2) is extinct.
-
(2)
If \(\delta _{i}^{*}=0\), then the ith species (\(i = 1, 2, 3\)) in system (2) is non-persistent in the mean.
-
(3)
If \(\delta _{i}^{*}>0\), then the ith species (\(i = 1, 2, 3\)) in system (2) is weakly persistent.
-
(4)
If \({\theta _{i}}_{*}>0\), then the ith species (\(i = 1, 2, 3\)) in system (2) is persistent in the mean.
-
(5)
If \((\check{\sigma })^{2}<2\hat{r}\) and Assumption 2.1 holds, then system (2) is stochastically permanent.
(II): The solution \(x_{i}(t)\) (\(i=1,2,3\)) obeys
(III): Under Assumption 2.1, the solution of system (2) satisfies
In addition, if \(2\hat{r}-(\check{\sigma })^{2}>0\), then
(IV): If \(A,B,C>0\) and Assumption 2.1 holds, then system (2) is globally attractive. By our results, we can analyze that the smaller stochastic perturbations cannot affect the stochastic permanence and extinction of the population. However, if the stochastic perturbations are larger, the stochastic permanence of the populations will be extinct. Similarly, the small impulsive perturbations have a little influence on the stochastic permanence and extinction of the populations. However, if the impulsive perturbations are large, the stochastic permanence and extinction of the populations could be changed.
We will give some numerical experiments to verify our analytical results by using the Milstein method (see [62]) by supplementing impulsive perturbations into it. We choose the same initial value \((x_{1}(0), x_{2}(0), x_{3}(0))=(0.5,0.5,0.5)\) and the same parameters in the following numerical examples.
The parameters are as follows:
At first, we will discuss the effects of different stochastic perturbations to system (2) under the same impulse interference in following Examples 1–6.
Let \(h_{1k}=h_{2k}=h_{3k}=e^{-0.2}-1\), it is easy to verify that
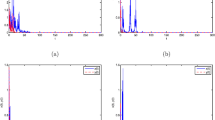
which means the Assumption 2.1 holds. In system (2) without stochastic perturbations, we can see that the prey and predator populations are all permanent (see Fig. 1).

(a) is the time sequence diagram and (b) the phase portrait of system (2) without stochastic perturbations and impulse\(.(x_{1}(0), x_{2}(0), x_{3}(0))=(0.5,0.5,0.5)\), \(\sigma ^{2}_{1}(t)= \sigma ^{2}_{2}(t)=\sigma ^{2}_{3}(t)=0\)
Example 1
Let \(\sigma _{1}^{2}(t)=\sigma _{2}^{2}(t)= \sigma _{3}^{2}(t)=0.1+0.04\sin t\). Then we get \((\check{\sigma })^{2}=0.14<2 \hat{r}=0.72\), and the Assumption 2.1 holds. According to Theorem 3.5, we can see that the prey population \(x_{1}(t)\), \(x_{2}(t)\) and the predator population \(x_{3}(t)\) are all stochastically permanent (see Fig. 2).

Example 2
Let \(\sigma _{2}^{2}(t)=\sigma _{3}^{2}(t)=0.1+0.04 \sin t\), \(\sigma _{1}^{2}(t)=2.56+0.04\sin t\). Then we get \(\delta _{1} ^{*}=-0.08<0\). By Theorem 3.1, we can see that the prey population \(x_{1}(t)\) will be extinct (see Fig. 3(a),(c)) and the population \(x_{2}(t)\), \(x_{3}(t)\) are all stochastically permanent (see Fig. 3(a),(d)).

Example 3
Let \(\sigma _{1}^{2}(t)=\sigma _{3}^{2}(t)=0.1+0.04 \sin t\), \(\sigma _{2}^{2}(t)=2.5+0.04\sin t\). Then we get \(\delta _{2} ^{*}=-0.13<0\). By Theorem 3.1, we can see that the prey population \(x_{2}(t)\) will be extinct (see Fig. 4(a),(c)) and the population \(x_{1}(t)\), \(x_{3}(t)\) are all stochastically permanent (see Fig. 4(a),(d)).

Example 4
Let \(\sigma _{1}^{2}(t)=\sigma _{2}^{2}(t)=0.1+0.04 \sin t\), \(\sigma _{3}^{2}(t)=2.2+0.04\sin t\). Then we get \(\delta _{3} ^{*}= -0.0376<0\). By Theorem 3.1, we can see that the predator population \(x_{3}(t)\) will be extinct (see Fig. 5(a),(c)) and the prey population \(x_{1}(t)\), \(x_{2}(t)\) are all stochastically permanent (see Fig. 5(a),(d)).

Example 5
Let \(\sigma _{1}^{2}(t)=\sigma _{2}^{2}(t)=0.1+0.04 \sin t\), \(\sigma _{3}^{2}(t)=2.1624+0.04\sin t\). Then we get \(\delta _{3}^{*}=0\). According to Theorem 3.2, we can see that the population \(x_{3}(t)\) is non-persistent in the mean (see Fig. 6(a),(c)).

Example 6
Let \(\sigma _{1}^{2}(t)=\sigma _{2}^{2}(t)= \sigma _{3}^{2}(t)=0.1+0.04\sin t\). By Theorem 3.1, 3.2 and 3.4, we can calculate that \(\delta _{1}^{*}=1.15\), \(\delta _{2}^{*}=1.07\), \(\delta _{3}^{*}=0.996\), \({\delta _{1}}_{*}=1.15\), \({\delta _{2}}_{*}=1.07\), \({\delta _{3}}_{*}=0.33\), \(\underline{x}_{1}^{*}=0.466\), \(\underline{x}_{2}^{*}=0.2784\), \(\underline{x}_{3}^{*}=0.7174\), \(\overline{x}_{1}^{*}=5\), \(\overline{x}_{2}^{*}=3.6897\), \(\overline{x}_{3}^{*}=2.2636\).
Denote \(x_{i}^{*}(t)=\frac{1}{t}\int _{0}^{t}x_{i}(s)\,ds\) (\(i=1,2,3\)). Since
then we have \(0.466\leq x_{1}^{*}(t)\leq 5\), \(0.2784\leq x_{2}^{*}(t) \leq 3.6897\), \(0.7174\leq x_{3}^{*}(t)\leq 2.2636\). In Fig. 7(a), we can see that the persistence in the mean of system (2). In Fig. 7(b), it is clear to see that the curve of \(x_{i}^{*}(t)\) gradually transcend the line \(\underline{x}_{i}^{*}(t)\) and stays between the \(\underline{x}_{i}^{*}(t)\) and \(\overline{x}_{i}^{*}(t)\) lines of the same color, which verify the conclusion.

Finally, we give Example 7 to discuss the effect of the impulsive perturbations on system (2), according to the choice of parameters in Example 1.
Example 7
Let \(h_{1k}=h_{2k}=e^{-1.2}-1\), \(h_{3k}=e^{-0.6}-1\). In Fig. 8, one can easily see that all of the species in system (2) become extinct gradually. This means suitable impulsive control strategy might be useful for the permanence of the system while arbitrary impulsive perturbations might lead to the extinction of system (2).

Therefore, through the numerical simulations given in Examples 1–6, we can see that the large stochastic disturbance is disadvantageous for the persistence of the population. However, the small stochastic perturbation is little effects on the permanence and extinction of the population. By Fig. 8, we can see that the small impulsive perturbations cannot affect the stochastic permanence and extinction of the prey and predator populations. But the large impulsive perturbations can lead to population extinction.
On the other hand, if \(a_{3}(t)=b_{3}(t)=0\), the Beddington–DeAngelis functional response converts to the Holling II functional response in system (2), then the system (2) becomes
Therefore, we can obtain the following results.
(I):
-
(1)
If \(\delta _{i}^{*}=\limsup_{t\rightarrow +\infty } \frac{1}{t} [\sum_{0< t_{k}< t}\ln (1+h_{ik})+\int _{0}^{t}\delta _{i}(s)\,ds ]<0\), then the ith species (\(i = 1, 2, 3\)) in system (45) is extinct.
-
(2)
If \(\delta _{i}^{*}=0\), then the ith species (\(i = 1, 2, 3\)) in system (45) is non-persistent in the mean.
-
(3)
If \(\delta _{i}^{*}>0\), then the ith species (\(i = 1, 2, 3\)) in system (45) is weakly persistent.
-
(4)
If \({\theta _{i}}_{*}>0\), then the ith species (\(i = 1, 2, 3\)) in system (45) is persistent in the mean, where
$$ \begin{aligned}[b] &{\theta _{1}}_{*}={\delta _{1}}_{*}- \biggl(\frac{\beta _{1}^{u}\delta _{2}^{*}}{\alpha _{2}^{l}}+\frac{c_{1}^{u}\delta _{3}^{*}}{a_{1}^{l} \alpha _{3}^{l}} \biggr), \qquad {{\theta _{2}}_{*}}={\delta _{2}}_{*}- \biggl(\frac{ \beta _{2}^{u}\delta _{1}^{*}}{\alpha _{1}^{l}}+\frac{c_{2}^{u}\delta _{3}^{*}}{b_{1}^{l}\alpha _{3}^{l}} \biggr), \qquad {{\theta _{3}}_{*}}={\delta _{3}} _{*}, \\ &{\delta _{i}}_{*}=\liminf_{t\rightarrow +\infty } \frac{1}{t} \biggl[\sum_{0< t_{k}< t}\ln (1+h_{ik})+ \int _{0}^{t} \biggl(r_{i}(s)- \frac{1}{2}\sigma ^{2}_{i}(s) \biggr)\,ds \biggr], \quad i=1,2,3. \end{aligned} $$ -
(5)
If \((\check{\sigma })^{2}<2\hat{r}\) and Assumption 2.1 holds, then system (45) is stochastically permanent.
(II) The solution \(x_{i}(t)\) (\(i=1,2,3\)) obeys
(III) Under Assumption 2.1, the solution of system (45) satisfies
In addition, if \(2\hat{r}-(\check{\sigma })^{2}>0\), then
By comparison, we see that the results of the B-D functional response are more accurate than those of the Holling II functional response. Now we show some simulations to verify our main results.
Example 8
The parameter values are the same as those given in Example 6. By the results of the Holling II functional response in system (45), we can calculate that \(\delta _{1}^{*}=1.15\), \(\delta _{2}^{*}=1.07\), \(\delta _{3}^{*}=0.996\), \({\delta _{1}}_{*}=1.15\), \({\delta _{2}}_{*}=1.07\), \({\delta _{3}}_{*}=0.33\), \(\underline{x}_{1} ^{*}=-0.4713<0\), \(\underline{x}_{2}^{*}=-0.164<0\), \(\underline{x}_{3} ^{*}=0.7174\), \(\overline{x}_{1}^{*}=5\), \(\overline{x}_{2}^{*}=3.6897\), \(\overline{x}_{3}^{*}=2.2636\). We can see that the values of \(\underline{x}_{1}^{*}\) and \(\underline{x}_{2}^{*}\) of system (45) are smaller than those of system (2). Therefore, our results can be verified in Fig. 9.

References
Cheng, K.S.: Uniqueness of a limit cycle for a predator–prey system. SIAM J. Math. Anal. 12(4), 541–548 (1981)
Mitra, D., Mukherjee, D., Roy, A., Ray, S.: Permanent coexistence in a resource-based competition system. Ecol. Model. 60(1), 77–85 (1992)
Freedman, H., Mathsen, R.: Persistence in predator–prey systems with ratio-dependent predator influence. Bull. Math. Biol. 55(4), 817–827 (1993)
Hsu, S.-B., Huang, T.-W.: Global stability for a class of predator–prey systems. SIAM J. Appl. Math. 55(3), 763–783 (1995)
Cantrell, R.S., Cosner, C.: On the dynamics of predator–prey models with the Beddington–Deangelis functional response. J. Math. Anal. Appl. 257(1), 206–222 (2001)
Liu, M., Yu, J., Mandal, P.S.: Dynamics of a stochastic delay competitive model with harvesting and Markovian switching. Appl. Math. Comput. 337, 335–349 (2018)
Liu, L., Meng, X.: Optimal harvesting control and dynamics of two species stochastic model with delays. Adv. Differ. Equ. 2018(1), 181 (2018)
Zhuo, X., Zhang, F.: Stability for a new discrete ratio-dependent predator–prey system. Qual. Theory Dyn. Syst. 17(1), 189–202 (2018)
Liu, M., He, X., Yu, J.: Dynamics of a stochastic regime-switching predator–prey model with harvesting and distributed delays. Nonlinear Anal. Hybrid Syst. 28, 87–104 (2018)
Liu, M., Zhu, Y.: Stationary distribution and ergodicity of a stochastic hybrid competition model with Lévy jumps. Nonlinear Anal. Hybrid Syst. 30, 225–239 (2018)
Zhu, F., Meng, X., Zhang, T.: Optimal harvesting of a competitive n-species stochastic model with delayed diffusions. Math. Biosci. Eng. 16(3), 1554–1574 (2019)
Li, Y., Meng, X.: Dynamics of an impulsive stochastic nonautonomous chemostat model with two different growth rates in a polluted environment. Discrete Dyn. Nat. Soc. 2019, 15 (2019)
Liu, G., Chang, Z., Meng, X.: Asymptotic analysis of impulsive dispersal predator–prey systems with Markov switching on finite-state space. J. Funct. Spaces 2019, 18 (2019)
Fujii, K.: Complexity–stability relationship of two-prey–one-predator species system model: local and global stability. J. Theor. Biol. 69(4), 613–623 (1977)
Vance, R.R.: Predation and resource partitioning in one predator–two prey model communities. Am. Nat. 112(987), 797–813 (1978)
Lakoš, N.: Existence of steady-state solutions for a one-predator–two-prey system. SIAM J. Math. Anal. 21(3), 647–659 (1990)
Feng, W.: Coexistence, stability, and limiting behavior in a one-predator–two-prey model. J. Math. Anal. Appl. 179(2), 592–609 (1993)
Ma, T., Meng, X., Chang, Z.: Dynamics and optimal harvesting control for a stochastic one-predator–two-prey time delay system with jumps. Complexity 2019, Article ID 5342031, 19 pages (2019)
Holling, C.S.: Some characteristics of simple types of predation and parasitism. Can. Entomol. 91(7), 385–398 (1959)
Skalski, G.T., Gilliam, J.F.: Functional responses with predator interference: viable alternatives to the Holling type ii model. Ecology 82(11), 3083–3092 (2001)
Jiang, Z., Zhang, W., Zhang, J., Zhang, T.: Dynamical analysis of a phytoplankton–zooplankton system with harvesting term and Holling iii functional response. Int. J. Bifurc. Chaos 28(13), 1850162 (2018)
Beddington, J.R.: Mutual interference between parasites or predators and its effect on searching efficiency. J. Anim. Ecol. 44(1), 331–340 (1975)
DeAngelis, D.L., Goldstein, R., O’neill, R.: A model for tropic interaction. Ecology 56(4), 881–892 (1975)
Jiang, Z., Bai, X., Zhang, T., Pradeep, B.: Global Hopf bifurcation of a delayed phytoplankton-zooplankton system considering toxin producing effect and delay dependent coefficient. Math. Biosci. Eng. 16(5), 3807–3829 (2019)
Bian, F., Zhao, W., Yue, Y.S.R.: Dynamical analysis of a class of prey-predator model with Beddington–Deangelis functional response, stochastic perturbation, and impulsive toxicant input. Complexity 2017, Article ID 3742197 (2017)
Hsu, S.-B., Hwang, T.-W., Kuang, Y.: Global analysis of the Michaelis–Menten-type ratio-dependent predator–prey system. J. Math. Biol. 42(6), 489–506 (2001)
Ivlev, V.S.: Experimental Ecology of the Feeding of Fishes. University Microfilms, Moscow (1961)
Hassell, M., Varley, G.: New inductive population model for insect parasites and its bearing on biological control. Nature 223(5211), 1133–1137 (1969)
Crowley, P.H., Martin, E.K.: Functional responses and interference within and between year classes of a dragonfly population. J. North Am. Benthol. Soc. 8(3), 211–221 (1989)
Ji, C., Jiang, D., Shi, N.: Analysis of a predator–prey model with modified Leslie–Gower and Holling-type ii schemes with stochastic perturbation. J. Math. Anal. Appl. 359(2), 482–498 (2009)
Meng, X., Li, F., Gao, S.: Global analysis and numerical simulations of a novel stochastic eco-epidemiological model with time delay. Appl. Math. Comput. 339, 701–726 (2018)
Rudnicki, R., Pichór, K.: Influence of stochastic perturbation on prey–predator systems. Math. Biosci. 206(1), 108–119 (2007)
Feng, T., Qiu, Z., Meng, X.: Dynamics of a stochastic hepatitis c virus system with host immunity. Discrete Contin. Dyn. Syst., Ser. B (2019). https://doi.org/10.3934/dcdsb.2019143
Chang, Z., Meng, X., Zhang, T.: A new way of investigating the asymptotic behaviour of a stochastic sis system with multiplicative noise. Appl. Math. Lett. 87, 80–86 (2019)
Meng, X., Zhao, S., Feng, T., Zhang, T.: Dynamics of a novel nonlinear stochastic sis epidemic model with double epidemic hypothesis. J. Math. Anal. Appl. 433(1), 227–242 (2016)
Liu, M., Deng, M.: Permanence and extinction of a stochastic hybrid model for tumor growth. Appl. Math. Lett. 94, 66–72 (2019)
Feng, T., Qiu, Z.: Global dynamics of deterministic and stochastic epidemic systems with nonmonotone incidence rate. Int. J. Biomath. 11(7), 1850101 (2018)
Feng, T., Qiu, Z., Meng, X., Rong, L.: Analysis of a stochastic hiv-1 infection model with degenerate diffusion. Appl. Math. Comput. 348, 437–455 (2019)
Feng, T., Qiu, Z.: Global analysis of a stochastic tb model with vaccination and treatment. Discrete Contin. Dyn. Syst., Ser. B 24(6), 2923–2939 (2019)
Song, Y., Miao, A., Zhang, T., Wang, X., Liu, J.: Extinction and persistence of a stochastic sirs epidemic model with saturated incidence rate and transfer from infectious to susceptible. Adv. Differ. Equ. 2018(1), 293 (2018)
Gao, N., Song, Y., Wang, X., Liu, J.: Dynamics of a stochastic SIS epidemic model with nonlinear incidence rates. Adv. Differ. Equ. 2019, 41 (2019)
Liu, X., Li, Y., Zhang, W.: Stochastic linear quadratic optimal control with constraint for discrete-time systems. Appl. Math. Comput. 228, 264–270 (2014)
Ma, H., Jia, Y.: Stability analysis for stochastic differential equations with infinite Markovian switchings. J. Math. Anal. Appl. 435(1), 593–605 (2016)
Li, X., Lin, X., Lin, Y.: Lyapunov-type conditions and stochastic differential equations driven by g-Brownian motion. J. Math. Anal. Appl. 439(1), 235–255 (2016)
Feng, T., Qiu, Z., Meng, X.: Analysis of a stochastic recovery-relapse epidemic model with periodic parameters and media coverage. J. Appl. Anal. Comput. 9(3), 1–15 (2019)
Cai, Y., Jiao, J., Gui, Z., Liu, Y., Wang, W.: Environmental variability in a stochastic epidemic model. Appl. Math. Comput. 329, 210–226 (2018)
Jiang, D., Shi, N., Li, X.: Global stability and stochastic permanence of a non-autonomous logistic equation with random perturbation. J. Math. Anal. Appl. 340(1), 588–597 (2008)
Qi, H., Liu, L., Meng, X.: Dynamics of a nonautonomous stochastic sis epidemic model with double epidemic hypothesis. Complexity 2017, Article ID 4861391 (2017)
Wang, W., Cai, Y., Li, J., Gui, Z.: Periodic behavior in a fiv model with seasonality as well as environment fluctuations. J. Franklin Inst. 354(16), 7410–7428 (2017)
Liu, M., Wang, K.: Asymptotic behavior of a stochastic nonautonomous Lotka–Volterra competitive system with impulsive perturbations. Math. Comput. Model. 57(3), 909–925 (2013)
Liu, G., Wang, X., Meng, X.: Extinction and persistence in mean of a novel delay impulsive stochastic infected predator–prey system with jumps. Complexity 2017, Article ID 1950970 (2017)
Zhang, S., Meng, X., Feng, T., Zhang, T.: Dynamics analysis and numerical simulations of a stochastic non-autonomous predator–prey system with impulsive effects. Nonlinear Anal. Hybrid Syst. 26, 19–37 (2017)
Meng, X., Zhang, L.: Evolutionary dynamics in a Lotka–Volterra competition model with impulsive periodic disturbance. Math. Methods Appl. Sci. 39(2), 177–188 (2016)
Meng, X., Wang, L., Zhang, T.: Global dynamics analysis of a nonlinear impulsive stochastic chemostat system in a polluted environment. J. Appl. Anal. Comput. 6(3), 865–875 (2016)
Lv, X., Wang, L., Meng, X.: Global analysis of a new nonlinear stochastic differential competition system with impulsive effect. Adv. Differ. Equ. 2017(1), 296 (2017)
Zhang, T., Meng, X., Song, Y., Zhang, T.: A stage-structured predator–prey SI model with disease in the prey and impulsive effects. Math. Model. Anal. 18(4), 505–528 (2013)
Mao, X.: Stochastic Differential Equations and Applications. Horwood, Chichester (1997)
Liptser, R.: A strong law of large numbers for local martingales. Stochastics 3(1–4), 217–228 (1980)
Mao, X., Yuan, C.: Stochastic Differential Equations with Markovian Switching. Imperial College Press, London (2006)
Karatzas, I., Shreve, S.E.: Brownian Motion and Stochastic Calculus. Springer, Berlin (1991)
Barbalat, I.: Systems d’equations differentielles d’oscillations nonlineaires. Rev. Roum. Math. Pures Appl. 4, 267–270 (1959)
Higham, D.: An algorithmic introduction to numerical simulation of stochastic differential equations. SIAM Rev. 43(3), 525–546 (2001)
Funding
This work was supported by the Research Fund for the Taishan Scholar Project of Shandong Province of China, Shandong Provincial Natural Science Foundation of China (ZR2019MA003), the SDUST Research Fund (2014TDJH102), the SDUST Innovation Fund for Graduate Students (SDKDYC180226), the Postgraduate Research & Practice Innovation Program of Jiangsu Province (KYCX18_0370), the Scholarship Foundation of China Scholarship Council (201806840120), the National Natural Science Foundation of China (11671206), and the Fundamental Research Funds for the Central Universities (30918011339).
Author information
Authors and Affiliations
Contributions
All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Qi, H., Meng, X. & Feng, T. Dynamics analysis of a stochastic non-autonomous one-predator–two-prey system with Beddington–DeAngelis functional response and impulsive perturbations. Adv Differ Equ 2019, 235 (2019). https://doi.org/10.1186/s13662-019-2170-9
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13662-019-2170-9