
Abstract
Background
Dairy cows are at high risk of fatty liver disease in early lactation, but current preventative measures are not always effective. Cows with fatty liver have lower circulating branched-chain amino acid (BCAA) concentrations whereas cows with high circulating BCAA levels have low liver triglyceride (TG). Our objective was to determine the impact of BCAA and their corresponding ketoacids (branched-chain ketoacids, BCKA) on production performance and liver TG accumulation in Holstein cows in the first 3 weeks postpartum.
Methods
Thirty-six multiparous Holstein cows were used in a randomized block design experiment. Cows were abomasally infused for the first 21 d postpartum with solutions of 1) saline (CON, n = 12); 2) BCA (67 g valine, 50 g leucine, and 34 g isoleucine, n = 12); and 3) BCK (77 g 2-ketovaline calcium salt, 57 g 2-ketoleucine calcium salt, and 39 g 2-ketoisoleucine calcium salt, n = 12). All cows received the same diet. Treatment effects were determined using PROC GLIMMIX in SAS.
Results
No differences were detected for body weight, body condition score, or dry matter intake averaged over the first 21 d postpartum. Cows receiving BCK had significantly lower liver TG concentrations compared to CON (6.60% vs. 4.77%, standard error of the mean (SEM) 0.49) during the first 3 weeks of lactation. Infusion of BCA increased milk yield (39.5 vs. 35.3 kg/d, SEM 1.8), milk fat yield (2.10 vs. 1.69 kg/d, SEM 0.08), and lactose yield (2.11 vs. 1.67 kg/d, SEM 0.07) compared with CON. Compared to CON, cows receiving BCA had lower plasma glucose (55.0 vs. 59.2 mg/dL, SEM 0.86) but higher β-hydroxybutyrate (9.17 vs. 6.00 mg/dL, SEM 0.80).
Conclusions
Overall, BCAA supplementation in this study improved milk production, whereas BCKA supplementation reduced TG accumulation in the liver of fresh cows.
Graphical Abstract

Similar content being viewed by others


Introduction
The annual cost of fatty liver disease (FL) in dairy cows in the United States was estimated to be over $150 million 8 years ago when prevalence was assumed at 4.8% [1]. FL in dairy cows is marked by accumulation of triglyceride (TG) within hepatocytes. Higher liver TG concentration is usually associated with compromised liver function and lactation performance [1, 2]. Current FL treatments are ineffective, as affected cows respond poorly to therapy with high mortality rates [3]. To alleviate the high rates of FL, many nutritional preventative measures have been developed for fresh cows [4]. Unfortunately, these strategies are not always effective [5,6,7].
Cows with FL have lower circulating branched-chain amino acid (BCAA) concentrations whereas cows with high circulating BCAA levels have low liver TG [1]. This inverse relationship has been well-established [1], but BCAA supplementation has not been tested as a strategy to reduce liver TG in fresh cows. As major components of milk protein and precursors for nonessential AA [8], only a small portion of BCAA in lactating cows is directly removed by the liver [9, 10]. In sheep, BCAA also had the lowest proportional extractions by the liver compared to other AA [11]. Although BCAA may not substantially impact the liver (as little BCAA are taken up by cow liver), transamination products of BCAA—branched-chain ketoacids (BCKA)—are catabolized in the liver in most mammals [12]. To our knowledge, uptake of BCKA by liver has not been shown in cattle, but in sheep, as much as 75% of BCKA are catabolized in the liver [13]. Hence, BCKA shuttled to the liver may serve as mediators of the inverse relationship between circulating BCAA and liver TG concentration and BCKA supplementation might suppress accumulation of liver TG in early postpartum cows. Therefore, increasing BCKA availability to the liver, either by post-ruminal BCKA supplementation or by enhancing BCAA transamination by post-ruminal BCAA supplementation may reduce liver TG concentration in fresh cows.
Recent work in early-lactation dairy cows has also demonstrated that BCAA supplementation improved milk yield and milk components yield [14]. Increased post-ruminal BCAA supply likely contributes to these responses, as BCAA are estimated to make up 20.8% of total AA mass in milk [15]. Although little is known about the impact of BCKA on lactation performance, feeding keto-leucine increased milk fat yield in cows during mid-lactation [16]. Therefore, our working hypothesis was that post-ruminal BCKA supplementation would reduce the risk of FL whereas post-ruminal BCAA supplementation would improve lactation performance in fresh cows. To that end, the objectives of this study were to evaluate the effect of abomasal BCAA and BCKA infusion on liver TG concentrations and lactation performance in fresh cows.
Materials and methods
Experimental design and dietary treatments
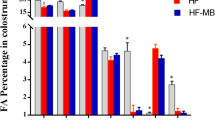
All procedures were approved by the Michigan State University Institutional Animal Care and Use Committee (protocol No. PROTO202000206). The experiment was conducted as a randomized complete block design with multiparous (lactation ≥ 2) Holstein dairy cows (n = 36) from the Michigan State University Dairy Cattle Teaching and Research Center (Lansing, MI, USA) between November 2021 and March 2022. A total of 36 cows were blocked according to expected calving date into 3 blocks. On average, cows enrolled had a 305-d mature equivalent milk yield of 13,421 ± 266 kg from previous lactation. Before enrollment, cows had an average body weight (BW) of 709 ± 8 kg and an average body condition score (BCS) of 3.3 ± 0.2. All cows underwent a rumenotomy procedure at 46.1 ± 2.2 d before expected calving date unless previously cannulated (n = 6). The number of animals included in this study were based on a power analysis to provide an 80% chance of detecting a 1.5% difference in liver TG concentration. Within each block, cows were randomly assigned to 1 of 3 treatments: control (CON, n = 12) receiving daily abomasal saline infusion without branched-chain amino acids or ketoacids; branched-chain amino acid (BCA, n = 12) receiving daily abomasal infusion of 67 g L-valine, 50 g L-leucine, and 34 g L-isoleucine (> 99.9% purity, ACP Chemicals, Montreal, QB, Canada); and branched-chain ketoacid (BCK, n = 12) receiving daily abomasal infusion of 77 g 2-ketovaline calcium salt, 57 g 2-ketoleucine calcium salt, and 39 g 2-ketoisoleucine calcium salt (> 99% purity, Stru Chem, Wujiang City, China). Using data from a previous study [17], plasma BCAA concentrations were on average 33% greater in cows with low hepatic TG (average 7.8 mg/g) whereas cows with high hepatic TG (average 50.8 mg/g) had lower plasma BCAA concentrations on d 4 postpartum. Therefore, dosage of BCA and BCK was based on a preliminary block of 6 cows (Fig. 1) that numerically increased plasma BCAA concentration by approximately 30%. The dose of BCA and corresponding BCK were matched on a 1:1 molar ratio across BCA and BCK treatments. Calcium chloride dihydrate (> 98% purity, Fisher Chemical) was added to CON and BCA infusate to match the calcium level in the BCK treatment. Treatments were delivered to cows daily via continuous abomasal infusion using a peristaltic pump [18, 19]. Seven liters of aqueous BCA (0.57 mol valine, 0.38 mol leucine, 0.26 mol isoleucine, pH = 2.5) or BCK (0.57 mol 2-ketovaline calcium salt, 0.38 mol 2-ketoleucine calcium salt, 0.26 mol 2-ketoisoleucine calcium salt, pH = 2.5) solution were administered to each cow daily. Calving day was considered d 0. Initiation of abomasal infusion for all animals occurred at 1200 h on d 1 after calving. Abomasal infusions were halted during milking for approximately 3–4 h/d. Abomasal infusion lines were flushed with 120 mL water after halting and before continuing infusion to ensure treatment delivery to abomasum.

Plasma valine, leucine, and isoleucine concentrations in a preliminary block of 6 cows (n = 2/treatment) on d 3 postpartum
Animal management
Animals were housed in individual sawdust-bedded tie stalls and were milked 3 times per day at approximately 0630, 1430, and 2230 h. Animals were fed ad libitum at 115% of expected intake. All cows received a common lactation diet (Table 1) once daily in the morning from calving to 21 d in milk (DIM). All cows enrolled in the study received the same close-up diet prior to calving. The lactation diet was formulated in AMTS Farm Cattle (Agricultural Modeling and Training Systems, LLC; Groton, NY, USA) to meet nutrient requirements for lactating cows at 14 DIM with an expected milk yield of 36.3 kg/d with 3.7% fat and 3.0% protein.
Body weight was measured weekly for each cow at the same time before the morning feeding. A body condition score (BCS; scale 1 = thin to 5 = obese, with quarter-point increments) was assigned to each cow weekly by 3 individuals and the average score was used for statistical analysis [20]. Dry matter intake was recorded daily. Milk yield was recorded daily during the first 21 DIM. Milk composition was analyzed, whereas energy-corrected milk (ECM) and fat-corrected milk (FCM) were calculated from calving to 21 DIM.
General cow housing and healthcare conform to approved standard operating procedures for the Michigan State University Dairy Cattle Teaching & Research Center. Attending veterinarians from the Michigan State University conducted diagnosis and, when needed, performed treatment for health problems.
Feed and milk sample collection and analyses
Dry matter (DM) of individual feed ingredients was determined weekly, and lactation diet was adjusted accordingly to maintain DM ratios of ingredients in the total mixed ration (TMR). Weekly samples of ingredients and TMR were frozen at –20 °C and composited monthly for analysis of DM, crude protein (CP), neutral detergent fiber (NDF), acid detergent fiber (ADF), amino acid (AA), lignin, ether extract, ash, Ca, P, Mg, K, Na by standard wet chemistry techniques [21] at a commercial laboratory (Cumberland Valley Analytical Services, Waynesboro, PA, USA) (Table 1). The values for net energy of lactation (NEL), rumen undegradable protein (RUP), rumen degradable protein (RDP), metabolizable protein (MP), Lys:Met ratio were also predicted using the National Academies of Sciences, Engineering, and Medicine (NASEM) 2021 model [15] with AA concentrations in the diet, dry matter intake (DMI), BW, BCS, milk production, and milk composition for each treatment as inputs (Table 2). Dry matter intake as a % of BW (DMI, % BW) was calculated daily using the weekly BW for each individual cow. Consecutive morning, midday, and evening milk samples were taken twice weekly. Milk samples were stored in a sealed tube with preservative (Bronopol tablet; D&F Control Systems, San Ramon, CA, USA) and stored at 4 °C until analyzed. Infrared spectroscopy was used to analyze and report milk fat (%), true protein (%), lactose (%), solids (%), and milk urea nitrogen (MUN) (mg/dL) and flow cytometry used to analyze somatic cell count (SCC) (1,000/mL) (Central Star Cooperative, Inc., Grand Ledge, MI, USA). Daily milk yield was summed over 3 milkings. Yields of 3.5% FCM and ECM were calculated as follows: FCM = 0.4324 × milk yield (kg) + 16.216 × fat yield (kg), ECM = 12.82 × fat yield (kg) + 7.13 × protein yield (kg) + 0.323 × milk yield (kg) [22, 23].
Blood sample collection and analysis
Blood was sampled from the coccygeal vein on 0 (actual calving day before treatment), 3, 7, 14, and 21 d relative to parturition before the morning feeding. Samples were collected into evacuated tubes (BD Vacutainer®, Becton, Dickinson and Company, Franklin Lakes, NJ, USA) containing either clot activator or sodium heparin for serum and plasma, respectively. After blood collection, tubes with sodium heparin were placed on ice and tubes with clot activator were kept at 21 °C until centrifugation (~ 30 min). Serum and plasma were obtained by centrifugation at 2,000 × g for 30 min at 4 °C. Aliquots of serum and plasma were frozen (–20 °C) until further analysis.
Commercial kits were used to analyze concentrations of plasma glucose (Sigma Chemical Co., St. Louis, MO, USA; Catalog number: P7119), serum insulin (Mercodia AB, Uppsala, Sweden; Catalog number 10-1201-01), plasma non-esterified fatty acid (NEFA, NEFA HR(2) kit, Wako Chemicals USA, Richmond, VA, USA), and plasma β-hydroxybutyrate (BHBA, Stanbio Laboratory, Boerne, TX, USA; Catalog number 2440-058) as described previously [24, 25]. All samples were analyzed in triplicate. Insulin resistance indices were calculated according to the following formulas [26]: homeostasis model assessment of insulin resistance (HOMA) = glucose (mmol/L) × insulin (µU/mL); quantitative insulin sensitivity check index (QUICKI) = 1/{(log[glucose (mg/dL)] + log[insulin (µU/mL)]}; and revised QUICKI (RQUICKI) = 1/{log[glucose (mg/dL)] + log[insulin (µU/mL)] + log[NEFA (mmol/L)]}.
Liver sample collection and TG concentration quantification
Liver was sampled via puncture biopsy using a Hughes trocar over the 11th and 12th ribs [25] on d 1, 7, 14, and 21 relative to parturition. On biopsy days, animals were escorted from morning milking to animal handling chutes before having access to feed. Location of biopsy was determined by ultrasound. Biopsy incision site was at least 2 inches away from the previous incision site. An area of approximately 10 cm × 15 cm around the incision site was anesthetized with 10 mL of 2% lidocaine (Vet One, Boise, ID, USA). Biopsy samples were harvested and immediately frozen and stored in liquid nitrogen until further analysis for concentration of TG. Approximately 50 mg of liver was used for liver TG quantification in triplicates using the method previously described [25] except a five-point TG standard was created with a commercially available TG standard solution (Pointe Scientific, Canton, MI, USA; Catalog number: T7531STD).
Statistical analysis
All data were analyzed using PROC GLIMMIX of SAS v9.4 (SAS Institute, Cary, NC, USA) with the model: Y = µ + bi+ Tj+ Dk+ TDjk+ Al:ij+ еijklm where Y = the dependent, continuous variable, µ = overall mean, bi= random effect of the ith block, Ti = fixed effect of treatment, Dk = fixed effect of time, Al:ij = random effect of the lth cow nested within block × Treatment and еijklm= the residual error. Fixed effect of time is the day or week relative to parturition in which samples were collected for each variable. Parity (second vs. third lactation and greater) was kept in the model as covariate for all variables when significant (P ≤ 0.05). The blood and liver samples collected prior to administration of treatments (d 0 samples) were also included as covariate for NEFA, BHB, and liver TG concentration respectively if found to be significant. Blood metabolites and liver TG were analyzed at various time points that were not equally spaced with a heterogeneous variance over time. Therefore, the first order ante-dependence covariance structure ANTE(1), spatial power covariance structure SP(POW), or Toeplitz covariance structure (TOEP) was chosen based on Akaike’s information criterion values for repeated measures of liver TG and blood metabolites. For equally spaced data, autoregressive covariance structure AR(1) was used for repeated measures. Somatic cell count was log transformed for statistical analysis and presentation. Tukey’s post hoc test was used for multiple comparison correction of P-values for all pairwise comparisons of least squares means (LSM). Statistical differences were declared significant at P ≤ 0.05 and tendencies at 0.05 < P ≤ 0.10.
Results
Health
Health-related problems that occurred during the experiment are summarized in Table 3. Two cows (1 receiving BCA and 1 receiving BCK) developed peritonitis unrelated to experimental treatments (according to autopsy reports) and were removed from the study following the recommendation of attending veterinarians from the Michigan State University department of Large Animal Clinical Sciences. All data from these two cows were removed from the analysis. Five cows developed hypocalcemia; however, 4 of the 5 cases occurred before treatment (d 0 or d 1 relative to parturition). One cow from BCK had hypocalcemia on d 3 relative to parturition. One hypocalcemia cow from BCA also became ketotic on d 6 postpartum. The other 3 ketotic cows had moderate or high ketones during the second week of lactation.
Dry matter intake, body weight, and body condition score
Dry matter intake, DMI%BW, BW, BW change, and BCS are presented in Table 4. As expected, DMI of cows in all treatments gradually increased (P < 0.01) from calving to 21 d in milk. However, DMI did not differ between treatment groups (P = 0.22). When normalized to BW (DMI %BW), intake also did not differ between treatment groups (P = 0.18). Similarly, BW, BCS, and BCS change were not affected by either treatment (P > 0.58) during the experimental period.
Milk production and composition
Milk production and composition variables are presented in Table 4. Overall dietary treatment effect was observed for milk yield, milk fat yield, lactose yield, ECM, FCM, milk:DMI, milk protein percentage, and MUN (P < 0.05). Specifically, BCA significantly increased milk yield compared to CON (P = 0.05) and BCK (P = 0.04) (Fig. 2A). Although milk fat and lactose percentage were not impacted by BCA or BCK treatments (P > 0.28), BCA significantly increased milk fat yield and milk lactose yield (P ≤ 0.05, Fig. 3D) as well as led to a tendency towards higher milk protein yield (P = 0.10) due to higher milk yield in BCA cows. Similarly, although milk protein percentage was significantly lower in BCA than BCK cows (P = 0.01, Fig. 3A), BCA cows had similar milk protein yield as BCK cows. The significantly higher milk:DMI in BCA cows (Fig. 2B) was also mainly driven by greater milk yield from BCA cows. As shown in Fig. 2C, FCM for cows receiving BCAA infusion was greater compared to CON (P = 0.05) and tended to be higher compared to BCK (P = 0.08). A tendency towards higher ECM (Fig. 2D) was also observed in BCA cows when compared to CON (P = 0.06) and BCK (P = 0.10). BCA significantly increased MUN compared to BCK (P = 0.02) whereas it led to only a tendency towards lower MUN when compared to CON cows (P = 0.10). Milk fat, lactose, and solids percentage as well as SCC were not affected by treatments (P ≥ 0.28).

Effects of BCAA or BCKA infusion during early lactation on milk yield (A), milk:DMI (B), fat corrected milk (C) and energy corrected milk (D)

Effects of BCAA or BCKA infusion during early lactation on milk protein percentage (A), milk protein yield (B), milk fat yield (C), milk lactose yield (D) and milk MUN (E)
Blood biomarkers and liver TG concentration
Blood glucose, insulin, NEFA, BHBA, and liver TG concentration are presented in Table 5. Overall treatment effect was observed for plasma glucose, BHBA, NEFA, and liver triglyceride concentration (P < 0.05). Specifically, cows receiving BCAA had significantly lower (P < 0.01) plasma glucose compared to CON and BCK cows. In contrast, plasma BHBA level in BCA cows were significantly higher than CON (P = 0.02) and BCK cows (P = 0.01). Although neither BCA nor BCK altered plasma NEFA concentrations compared to CON, plasma NEFA concentrations in BCA cows were higher (P = 0.02) than in BCK cows (Fig. 4B). Insulin and insulin resistance indices were not altered by BCA or BCK (P > 0.20). Compared to CON, BCK infusion significantly decreased liver TG (P = 0.03). In contrast, liver TG concentration in BCA cows tended to be higher (P = 0.09) compared to BCK cows (Fig. 4D).

Effects of BCAA or BCKA infusion during early lactation on plasma concentrations of glucose (A), NEFA (B), BHBA (C), liver triglyceride (D), and insulin (E)
Discussion
Milk production and composition
Abomasal infusion of BCAA at the current dose increased milk yield by 4.2 kg/d compared to CON. Based on NASEM [15] predictions for our diets and assuming that 100% of infused AA were absorbed (Table 2), cows infused with BCAA were predicted to produce 2.5 kg more milk per day based on protein supply if no change was made in user-entered milk yield. It is well-established that milk yield is highly correlated with mammary plasma flow [27]. A recent jugular infusion study demonstrated that Leu (50 g/d) and Ile (22 g/d) supplementation increased mammary plasma flow by 24% and increased milk yield by 2.3 kg/d [14]. In the current study, we infused Leu at the same dose as Yoder et al. [14] and Ile at a higher dose (34 g/d) into the abomasum. Hence, the increased milk yield of BCA cows in this study might have been partly caused by a higher mammary plasma flow. Measurement of mammary plasma flow will be required to confirm this speculation.
Abomasal infusion of BCAA at the current dose also increased milk protein yield by 0.21 kg/d compared to CON. This outcome was expected considering fresh cows are in negative MP balance [28], and increasing the amount of AA available may alleviate MP shortage in dairy cows by providing more metabolizable AA available as precursors for milk protein. However, measurement of mammary AA uptake will be required to confirm this speculation. We suggest that supplementing BCAA may also have stimulated milk protein synthesis and milk yield via activation of intracellular signaling pathways, such as the mammalian target of rapamycin (mTOR) pathway, based on work in other species and in cultured mammary cells and tissue [29,30,31]. As a coordinator of cell growth and metabolism, mTOR signaling plays a central role in regulating many fundamental cell processes, especially protein synthesis [32]. Therefore, we expected BCAA supplementation to upregulate milk protein synthesis. However, the effect of BCAA supplementation on lactation performance in published literature has been inconsistent. Yoder et al. [14] found that milk yield increased in response to jugular infusion of Leu (50 g/d) and Ile (22 g/d) and speculated that this increase was likely due to changes in mammalian target of rapamycin complex 1 (mTORC1). In contrast, Leal Yepes et al. [33] observed no increase in milk yield with supplementation of rumen-protected BCAA (61 g/d Val, 101 g/d Leu, and 41 g/d Ile) during the first 35 d in milk. Such inconsistencies of BCAA effect on milk production could be due to differences in BCAA dosage and method of delivery between these experiments. In the present study, we infused BCAA via continuous abomasal infusion using the molar ratio (Val:Leu:Ile = 2.2:1.5:1) observed in plasma of fresh cows with low liver TG concentration from a previous transition cow study [17]. Because BCAA-infused cows produced 4.2 kg/d more milk than CON, with little change in milk protein and fat percentage in milk, they also produced more fat, and protein. In contrast to the effects of BCAA, infusion of BCKA at equal molar amounts and ratio did not alter milk production. The only previous report on BCKA infusion was a study demonstrating that dietary supplementation of keto-leucine at 0.75% of DM in a partially rumen-protected form increased milk fat yield in dairy cows [16]. It is possible that the results of that study were due to similarities between ketoleucine and isovalerate, one of the branched-chain VFAs required by cellulolytic bacteria in the rumen [34].
The milk:DMI ratio is an estimate of gross feed efficiency, although this ratio does not account for BW change in individual cows. With BW change similar between groups, milk:DMI provides a good estimate of feed efficiency in this study. Therefore, BCA cows were more efficient than CON cows, with a 4.2 kg/d increase in milk yield without increased DMI.
Glucose, insulin, and insulin resistance indices
It has been well established that a low-level of insulin could induce a metabolic condition favoring tissue and AA catabolism in cows, which might result in lactose yield increase because catabolism of gluconeogenic AA could increase the glucose available for mammary gland to produce lactose [35]. Although only a numerical decrease in insulin was observed in response to BCAA supplementation in this study, the observed increase in lactose yield may partially originate from increased AA catabolism. However, future evidence quantifying changes in gluconeogenesis in response to BCAA or BCKA supplementation may be required to support this hypothesis. Previous reports from both monogastric and ruminant studies have also illustrated the impact of BCAA supplementation on circulating glucose concentration [36, 37]. As the main precursor for lactose production in the mammary gland, mammary glucose uptake greatly impacts the circulating glucose concentration in dairy cows, especially during early lactation when cows enter a phase of negative energy balance [38, 39]. In this study, BCAA infusion decreased plasma glucose concentration, but significantly increased lactose yield compared with CON. A recent report in dairy cows revealed that Leu and Ile supplementation increased mammary plasma flow [14]. Although increase in mammary plasma flow could increase mammary glucose uptake, which, in turn, might have caused the lower plasma glucose concentration observed in this study, measurement of mammary arteriovenous glucose concentration difference will be required to confirm this speculation.
Recent advances in human research suggest that increases in BCAA plasma levels are associated with insulin resistance, and that elevated BCAA may cause insulin resistance by stimulating mTORC1 [30]. Contradictory to the suggested causal relationship, recent work in humans and rodents suggest that increased circulating BCAA concentrations are simply markers of insulin resistance, as physiological changes in BCAA concentrations are not sufficient to elicit insulin resistance [40, 41]. In dairy cows, homeorhetic adaptations to ensure sufficient glucose supply for the mammary gland induce insulin resistance in extra-mammary tissues during early lactation [42]. In this study, no change in insulin or various insulin resistance indices (HOMA, QUICKI, and RQUICKI) were observed in response to BCA or BCK, which could be because blood samples were collected after a short fasting period (~ 3 h). Hence, intravenous glucose and/or insulin tolerance test after overnight fasting may be required to determine the casual relationship between BCAA supplementation and insulin resistance in fresh cows.
Liver triglyceride concentration, BHBA, and NEFA
Fatty liver in dairy cattle is categorized into mild (1%–5% TG wet weight), moderate (5%–10% TG wet weight), and severe FL (> 10% TG wet weight) [1]. In the present study, BCK cows were in the mild FL category with an average 4.77% liver TG concentration whereas CON and BCA cows were in the moderate FL category with average liver TG concentration of 6.60% and 6.15%, respectively. Previous report from a murine model demonstrated that BCAA supplementation decreased liver TG [43]. Similarly, in dairy cattle, supplementation with BCAA and propylene glycol also reduced liver TG in early lactation [33]. With 40%–50% of dairy cows having liver TG concentration > 5% within the first month after calving, FL in dairy cows is usually associated with compromised lactation performance and increased susceptibility to health issues [1]. Based on these associations, strategies that effectively reduce liver TG concentration are expected to improve lactation performance in dairy cows by decreasing negative impact of FL disease. Despite the significantly lower liver TG concentration in BCK cows, lactation performance was not improved in the first 3 weeks of lactation. In contrast, with liver TG concentration in the moderate FL category, BCA increased milk yield and milk components yield compared to CON cows in the first 3 weeks of lactation. These results suggest that liver TG concentration may not be the best predictor for lactation performance, at least in fresh cows with liver TG concentration in the mild or moderate FL category. Instead, alterations in liver TG may reflect the degree of adipose tissue mobilization and consequent elevation in circulating NEFA concentrations in fresh cows. However, quantifiable measurements of adipose tissue mobilization will be required to confirm this speculation.
It is also noteworthy that abomasal infusion of BCAA at current dose did not decrease liver TG compared to CON, but infusing equal molar amount of BCKA (deaminated BCAA) successfully decreased liver TG concentration. This is likely because after absorption, BCAA were minimally removed by the liver in dairy cows whereas BCKA absorbed were metabolized primarily in the liver [44, 45]. Additionally, a proportion of BCAA was also likely utilized for milk and tissue protein synthesis during early lactation, which likely contributes to the discrepancy of BCA and BCK impact on liver TG concentration. Apart from the amount of BCKA reaching the liver, liver TG concentration is also greatly impacted by circulating NEFA concentrations during early lactation, as circulating NEFA concentration peaks soon after parturition in dairy cows [46]. Although circulating NEFA concentrations in BCA cows and BCK cows were similar to CON cows, BCA cows had significantly higher NEFA concentrations compared to BCK cows. Considering hepatic uptake of NEFAs is impacted by concentration of NEFAs in blood in cows [47], the significantly higher circulating NEFA level in BCA cows was likely associated with the observed tendency towards higher liver TG concentration in BCA cows compared to cows infused with BCK.
As a marker for adipose tissue lipolysis, very high levels of circulating NEFA concentrations in early lactation are associated with compromised health in dairy cows [48]. Nevertheless, NEFA are also important sources of energy and fatty acids for liver and mammary gland [28]. Therefore, a moderate increase in circulating NEFA concentration that provides appropriate amount of energy substrate without jeopardizing health outcome may be beneficial for fresh cows. Although circulating NEFA concentration was not altered in response to rumen-protected BCAA supplementation at a different dose in a recent fresh cow study [49], NEFA concentration in BCA cows was significantly higher than BCK cows and numerically higher than CON cows in this study. Meanwhile, milk fat yield in BCA cows was significantly higher than CON and BCK cows. Taken together, these results suggest that a higher circulating NEFA in BCA cows from this study could contribute to increased milk fat yield. However, data on mammary NEFA uptake will be required to confirm this speculation.
The early lactation period in dairy cows is typically marked with elevated plasma BHBA due to tissue mobilization [50]. Excessive circulating BHBA (subclinical or clinical ketosis) is typically associated with decreased milk production and potential health consequences, such as increased incidence of hyperketonemia and FL disease and decreasing β-oxidation, TCA cycle activity, and gluconeogenesis in hepatocytes [1, 51,52,53]. Although a previous report did not observe any change in circulating BHBA concentrations in response to BCAA supplementation [33], plasma BHBA concentrations were significantly higher in BCA cows compared with CON and BCK cows in this study. It is important to note that BHBA concentrations in the BCA cows of this study averaged 9.17 mg/dL (0.88 mmol/L). Therefore, these cows were not in a state of subclinical (1.2–2.9 mmol/L) or clinical (> 3.0 mmol/L) ketosis [54]. Considering BHBA is a main substrate for milk fat synthesis in the mammary gland [51], elevated circulating BHBA levels below the threshold for subclinical ketosis (1.2 mmol/L) may also contribute to the higher milk fat yield in BCA cows by providing more substrate for de novo FA synthesis in the mammary gland in this study. However, more direct evidence (e.g., mammary BHBA uptake or de novo milk FA output) will be required to support this speculation. As a main product of ketogenesis in the liver, higher circulating BHBA in BCA cows could also be an indicator of elevated incomplete FA oxidation in the liver. However, measurement of changes in abundance and activities of enzymes controlling ketogenesis are required to confirm such speculation.
Conclusions
With liver TG concentration in CON cows in the moderate FL category and TG concentration in BCK cows in the mild FL category, lower liver TG concentrations in BCK cows compared to CON cows suggests that post-ruminal BCKA supplementation at the current dose may be an effective measure for minimizing risk of moderate or severe FL in cows during early lactation. Although abomasal BCAA infusion did not decrease liver TG concentration, the greater milk yield and components yield compared to CON cows suggest that post-ruminal BCAA supplementation may improve lactation performance in fresh cows. The discrepancy in lactation performance and liver TG concentration between BCA and BCK cows in this study were likely due to differences in metabolic alterations in response to BCAA and BCKA, as BCA cows had significantly lower circulating glucose, higher NEFA, and higher BHBA levels compared to BCK cows. Considering these divergent responses were attained from same molar amounts of post-ruminal supplementation of BCAA and BCKA in this study, future research should focus on the interrelated metabolism of BCAA and BCKA to better understand the unique effects on lactation performance and underlying metabolic mechanisms.
Availability of data and materials
The datasets used and/or analyzed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- AA:
-
Amino acid
- ADF:
-
Acid detergent fiber
- BCAA:
-
Branched-chain amino acid
- BCA:
-
BCAA-infused group
- BCKA:
-
Branched-chain ketoacids
- BCK:
-
BCKA-infused group
- BCS:
-
Body condition score
- BHBA:
-
β-hydroxybutyrate
- BW:
-
Body weight
- CON:
-
Control group
- CP:
-
Crude protein
- DIM:
-
Days in milk
- DM:
-
Dry matter
- DMI:
-
Dry matter intake
- ECM:
-
Energy-corrected milk
- FCM:
-
Fat-corrected milk
- FL:
-
Fatty liver
- GLIMMIX:
-
Generalized linear mixed model
- HOMA:
-
Homeostasis model assessment
- MP:
-
Metabolizable protein
- MUN:
-
Milk urea nitrogen
- mTOR:
-
Mammalian target of rapamycin
- mTORC1:
-
Mammalian target of rapamycin complex 1
- NASEM:
-
National Academies of Sciences, Engineering, and Medicine
- NEFA:
-
Non-esterified fatty acid
- NDF:
-
Neutral detergent fiber
- NEL :
-
Net energy of lactation
- QUICKI:
-
Quantitative insulin sensitivity check index
- RQUICKI:
-
Revised QUICKI
- RUP:
-
Rumen undegradable protein
- RDP:
-
Rumen degradable protein
- SCC:
-
Somatic cell count
- SEM:
-
Standard error of the mean
- TG:
-
Triglyceride
- TMR:
-
Total mixed ration
References
Bobe G, Young JW, Beitz DC. Invited review: pathology, etiology, prevention, and treatment of fatty liver in dairy cows. J Dairy Sci. 2004;87(10):3105–24.
Petta S, Gastaldelli A, Rebelos E, Bugianesi E, Messa P, Miele L, et al. Pathophysiology of non alcoholic fatty liver disease. Int J Mol Sci. 2016;17(12):2082.
Adewuyi AA, Gruys E, van Eerdenburg FJ. Non esterified fatty acids (NEFA) in dairy cattle. A review. Vet Q. 2005;27(3):117–26.
Gruffat D, Durand D, Graulet B, Bauchart D. Regulation of VLDL synthesis and secretion in the liver. Reprod Nutr Dev. 1996;36(4):375–89.
Hartwell JR, Cecava MJ, Donkin SS. Impact of dietary rumen undegradable protein and rumen-protected choline on intake, peripartum liver triacylglyceride, plasma metabolites and milk production in transition dairy cows. J Dairy Sci. 2000;83(12):2907–17.
Piepenbrink MS, Overton TR. Liver metabolism and production of cows fed increasing amounts of rumen-protected choline during the periparturient period. J Dairy Sci. 2003;86(5):1722–33.
Pinotti L, Baldi A, Politis I, Rebucci R, Sangalli L, Dell’Orto V. Rumen-protected choline administration to transition cows: effects on milk production and vitamin E status. J Vet Med A Physiol Pathol Clin Med. 2003;50(1):18–21.
Webb LA, Sadri H, Schuh K, Egert S, Stehle P, Meyer I, et al. Branched-chain amino acids: abundance of their transporters and metabolizing enzymes in adipose tissue, skeletal muscle, and liver of dairy cows at high or normal body condition. J Dairy Sci. 2020;103(3):2847–63.
Raggio G, Pacheco D, Berthiaume R, Lobley GE, Pellerin D, Allard G, et al. Effect of level of metabolizable protein on splanchnic flux of amino acids in lactating dairy cows. J Dairy Sci. 2004;87(10):3461–72.
Lapierre H, Blouin JP, Bernier JF, Reynolds CK, Dubreuil P, Lobley GE. Effect of supply of metabolizable protein on whole body and splanchnic leucine metabolism in lactating dairy cows. J Dairy Sci. 2002;85(10):2631–41.
Lobley GE, Connell A, Revell DK, Bequette BJ, Brown DS, Calder AG. Splanchnic-bed transfers of amino acids in sheep blood and plasma, as monitored through use of a multiple U-13 C-labelled amino acid mixture. Br J Nutr. 1996;75(2):217–35.
Wu G. Amino acids: biochemistry and nutrition. 2nd ed. Boca Raton: CRC Press; 2022.
Pell JM, Caldarone EM, Bergman EN. Leucine and alpha-ketoisocaproate metabolism and interconversions in fed and fasted sheep. Metabolism. 1986;35(11):1005–16.
Yoder PS, Huang X, Teixeira IA, Cant JP, Hanigan MD. Effects of jugular infused methionine, lysine, and histidine as a group or leucine and isoleucine as a group on production and metabolism in lactating dairy cows. J Dairy Sci. 2020;103(3):2387–404.
National Academies of Sciences Engineering, and Medicine. Nutrient requirements of dairy cattle: eighth revised edition. Washington, DC: The National Academies Press; 2021. p. 502.
Vandehaar MJ, Flakoll PJ, Beitz DC, Nissen S. Milk production and composition in cows and goats fed alpha-ketoisocaproate. J Dairy Sci. 1988;71(12):3352–61.
Zhou Z, Vailati-Riboni M, Luchini DN, Loor JJ. Methionine and choline supply during the periparturient period alter plasma amino acid and one-carbon metabolism profiles to various extents: potential role in hepatic metabolism and antioxidant status. Nutrients. 2016;9(1):10.
Spires HR, Clark JH, Derrig RG, Davis CL. Milk production and nitrogen utilization in response to postruminal infusion of sodium caseinate in lactating cows. J Nutr. 1975;105(9):1111–21.
Westreicher-Kristen E, Susenbeth A. Technical note: an improved tool to insert lines for abomasal infusion in rumen cannulated cattle. J Dairy Sci. 2017;100(3):1951–4.
Wildman EE, Jones GM, Wagner PE, Boman RL, Troutt HF, Lesch TN. A dairy cow body condition scoring system and its relationship to selected production characteristics. J Dairy Sci. 1982;65(3):495–501.
AOAC International. Official methods of analysis of AOAC International. 17th ed. Gaithersburg, MD: AOAC International; 2000. p. 251–306.
Hutjens MF. Benchmarking your feed efficiency, feed costs, and income over feed cost. Adv Dairy Technol. 2010;22:3–10.
Tyrrell HF, Reid JT. Prediction of the energy value of cow’s milk. J Dairy Sci. 1965;48(9):1215–23.
Maldini G, Kennedy KM, Allen MS. Temporal effects of ruminal infusion of propionic acid on hepatic metabolism in cows in the postpartum period. J Dairy Sci. 2019;102(11):9781–90.
Zhou Z, Vailati-Riboni M, Trevisi E, Drackley JK, Luchini DN, Loor JJ. Better postpartal performance in dairy cows supplemented with rumen-protected methionine compared with choline during the peripartal period. J Dairy Sci. 2016;99(11):8716–32.
Vieira-Neto A, Zimpel R, Lopes FR Jr, Scheffler TL, Block E, Thatcher WW, et al. Duration and degree of diet-induced metabolic acidosis prepartum alter tissue responses to insulin in dairy cows. J Dairy Sci. 2021;104(2):1660–79.
Prosser CG, Davis SR, Farr VC, Lacasse P. Regulation of blood flow in the mammary microvasculature. J Dairy Sci. 1996;79(7):1184–97.
Bell AW, Burhans WS, Overton TR. Protein nutrition in late pregnancy, maternal protein reserves and lactation performance in dairy cows. Proc Nutr Soc. 2000;59(1):119–26.
Lynch CJ, Adams SH. Branched-chain amino acids in metabolic signalling and insulin resistance. Nat Rev Endocrinol. 2014;10(12):723–36.
Yoon MS. The emerging role of branched-chain amino acids in insulin resistance and metabolism. Nutrients. 2016;8(7):405.
Appuhamy JA, Knoebel NA, Nayananjalie WA, Escobar J, Hanigan MD. Isoleucine and leucine independently regulate mTOR signaling and protein synthesis in MAC-T cells and bovine mammary tissue slices. J Nutr. 2012;142(3):484–91.
Saxton RA, Sabatini DM. mTOR signaling in growth, metabolism, and Disease. Cell. 2017;168(6):960–76.
Leal Yepes FA, Mann S, Overton TR, Ryan CM, Bristol LS, Granados GE, et al. Effect of rumen-protected branched-chain amino acid supplementation on production- and energy-related metabolites during the first 35 d in milk in Holstein dairy cows. J Dairy Sci. 2019;102(6):5657–72.
Allison MJ, Bryant MP, Katz I, Keeney M. Metabolic function of branched-chain volatile fatty acids, growth factors for ruminococci. II. Biosynthesis of higher branched-chain fatty acids and aldehydes. J Bacteriol. 1962;83(5):1084–93.
Bertoni G. Feeding and bovine milk quality: endocrine and metabolic factors. Zoot Nutr Anim. 1996;22:205–14.
Rezaei R, Gabriel AS, Wu G. Dietary supplementation with branched-chain amino acids enhances milk production by lactating sows and the growth of suckling piglets. J Anim Sci Biotechnol. 2022;13:65.
Leal Yepes FA, Mann S, Overton TR, Behling-Kelly E, Nydam DV, Wakshlag JJ. Hepatic effects of rumen-protected branched-chain amino acids with or without propylene glycol supplementation in dairy cows during early lactation. J Dairy Sci. 2021;104(9):10324–37.
Annison EF, Linzell JL. Oxidation and utilization of glucose and acetate by mammary gland of goat in relation to their over-all metabolism and to milk formation. J Physiol-London. 1964;175(3):372–85.
Lin Y, Sun X, Hou X, Qu B, Gao X, Li Q. Effects of glucose on lactose synthesis in mammary epithelial cells from dairy cow. BMC Vet Res. 2016;12:81.
Piccolo BD, Comerford KB, Karakas SE, Knotts TA, Fiehn O, Adams SH. Whey protein supplementation does not alter plasma branched-chained amino acid profiles but results in unique metabolomics patterns in obese women enrolled in an 8-week weight loss trial. J Nutr. 2015;145(4):691–700.
Piccolo BD, Graham JL, Stanhope KL, Fiehn O, Havel PJ, Adams SH. Plasma amino acid and metabolite signatures tracking diabetes progression in the UCD-T2DM rat model. Am J Physiol Endocrinol Metab. 2016;310(11):E958–69.
De Koster JD, Opsomer G. Insulin resistance in dairy cows. Vet Clin North Am Food Anim Pract. 2013;29(2):299–322.
Arakawa M, Masaki T, Nishimura J, Seike M, Yoshimatsu H. The effects of branched-chain amino acid granules on the accumulation of tissue triglycerides and uncoupling proteins in diet-induced obese mice. Endocr J. 2011;58(3):161–70.
Lobley GE. Control of the metabolic fate of amino acids in ruminants: a review. J Anim Sci. 1992;70(10):3264–75.
Arriola Apelo SI, Knapp JR, Hanigan MD. Invited review: current representation and future trends of predicting amino acid utilization in the lactating dairy cow. J Dairy Sci. 2014;97(7):4000–17.
Bell AW. Lipid metabolism in liver and selected tissues and in the whole body of ruminant animals. Prog Lipid Res. 1979;18(3):117–64.
Grummer RR. Nutritional and management strategies for the prevention of fatty liver in dairy cattle. Vet J. 2008;176(1):10–20.
Contreras GA, Sordillo LM. Lipid mobilization and inflammatory responses during the transition period of dairy cows. Comp Immunol Microbiol Infect Dis. 2011;34(3):281–9.
Xu W, Kenez A, Mann S, Overton TR, Wakshlag JJ, Nydam DV, et al. Effects of dietary branched-chain amino acid supplementation on serum and milk metabolome profiles in dairy cows during early lactation. J Dairy Sci. 2022;105(10):8497–508.
Drackley JK, ADSA Foundation Scholar Award. Biology of dairy cows during the transition period: the final frontier? J Dairy Sci. 1999;82(11):2259–73.
McArt JA, Nydam DV, Oetzel GR, Overton TR, Ospina PA. Elevated non-esterified fatty acids and β-hydroxybutyrate and their association with transition dairy cow performance. Vet J. 2013;198(3):560–70.
Grum DE, Drackley JK, Younker RS, LaCount DW, Veenhuizen JJ. Nutrition during the dry period and hepatic lipid metabolism of periparturient dairy cows. J Dairy Sci. 1996;79(10):1850–64.
Rukkwamsuk T, Wensing T, Geelen MJ. Effect of fatty liver on hepatic gluconeogenesis in periparturient dairy cows. J Dairy Sci. 1999;82(3):500–5.
Duffield TF, Lissemore KD, McBride BW, Leslie KE. Impact of hyperketonemia in early lactation dairy cows on health and production. J Dairy Sci. 2009;92(2):571–80.
Acknowledgements
We thank Evonik Nutrition and Care GmbH (Hanau-Wolfgang, Germany) for donating branched-chain amino acids for this experiment. We also appreciate the assistance from the staff at Michigan State University Dairy Cattle Teaching and Research Center (East Lansing) during this experiment.
Funding
This work is supported by the Agriculture and Food Research Initiative competitive grant No. 2021-67015-33383 from the USDA National Institute of Food and Agriculture (Washington, DC) and USDA, AgBioResearch, Michigan State University.
Author information
Authors and Affiliations
Contributions
ZZ and MV designed the study. KG, IB, SN, and VM conducted the experiment and collected the samples. KG, CC, DM, GA, JD, and ZZ analyzed the samples. KG, JD, MV, and ZZ wrote and prepared the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All procedures were approved by the Michigan State University Institutional Animal Care and Use Committee (protocol No. PROTO202000206).
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Gallagher, K., Bernstein, I., Collings, C. et al. Abomasal infusion of branched-chain amino acids or branched-chain keto-acids alter lactation performance and liver triglycerides in fresh cows. J Animal Sci Biotechnol 15, 13 (2024). https://doi.org/10.1186/s40104-023-00973-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40104-023-00973-7