
Abstract
Background
Oxidative stress is known to contribute to the mechanisms underpinning the pathogenesis of neurodegenerative diseases. Previous studies have identified the presence of flavonoids as the major constituents of Passiflora edulis (PE) with antioxidant activity. This work aims at investigating the antioxidant, anti-neuroinflammatory, and neuroprotective effect of three PE fruit extracts, flavonoid fraction, and juice on neurodegenerative rat model.
Methods
Extracts were prepared using fruit pulp and peel and juice using pulp. Phytochemical contents (phenolic content and flavonoid) and in vitro antioxidant activity were evaluated through the DPPH radical scavenging capacity and the ability to reduce ferric ion. The neurocognitive dysfunction, activity of acetylcholinesterase (AChE), levels and activities of in vivo oxidant–antioxidant indices as well as neuroinflammatory markers were evaluated in the hippocampus and cortex of aluminum chloride (AlCl3) induced Alzheimer’s rats (AD).
Results
The highest total phenolic and flavonoids’ contents, the best DPPH scavenging activity and the ability to reduce ferric ion (Fe3+) were obtained with peel aqueous extract. The administration of the peel aqueous extract, juice, and flavonoid fraction resulted in a significant decrease (P < 0.05) in plasma and tissue levels of malondialdehyde compared to the positive control (PC). The levels of tumor necrosis factor-α (TNF-α), interleukin-6 (IL-6), cycyclooxygenase-2 (COX-2), and amyloid ß-42 (ß-42) were significantly reduced whereas the activities of catalase (CAT), superoxide dismutase (SOD), glutathione peroxidase (GPx), and glutathione level were significantly higher in the treated than that in the untreated Alzheimer’s rats (PC) groups (P < 0.05), respectively, in the hippocampus and in plasma, brain, and liver homogenates following the administration of juice, flavonoid fraction, and extracts (both doses). Treatment of AD-rats with PE ameliorated neurobehavioral changes, as evidenced by the improvement in brain function, as well as, modulation of AChE, and confirmed by the histological changes and Morris water maze test. The effect of aqueous extract was slightly greater than that of the flavonoids fraction, thus suggesting that flavonoids account for most of the Passiflora edulis antioxidant activity and neuroprotective effect.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Background
Oxidative stress is the primary cause of pathogenesis in inflammatory, partial ischemia, metabolic, and denatured cranial nerve disease [1]. Brain tissues are highly susceptible to oxidative damage, probably because of high oxygen consumption rate (20%), the presence of abundant polyunsaturated fatty acids in cell membranes, high iron (Fe) content, and low enzymatic antioxidants’ activities [2]. Aluminum is one of the well-known environmental heavy metal agents that affect the brain development. Although it is a relatively low redox mineral, it can induce oxidative damage through multiple mechanisms. It can bind to negatively charged brain phospholipids, which contain polyunsaturated fatty acids and are easily attacked by reactive oxygen species (ROS) [3]. Aluminum has the potential to be neurotoxic in humans and animals, and is present in many manufactured foods and medicines [4]. Alzheimer’s disease (AD) is an age-related progressive neurodegenerative disease characterized by the presence of intracellular amyloid aggregates and extracellular neurofibrillary tangles which results in neurocognitive decline and memory impairment.
Several studies have been conducted to prove the existence of a relationship between the prevention of neurodegenerative diseases and some foods [5]. In France, large study was conducted on 3777 men and women over 65 ages to show that an average consumption of flavonoids (antioxidants) of 14.4 mg/day (mainly from fruits and vegetables) leads to a significant reduction in the risk of dementia [6].
These observations prompted us to focus on the Passiflora edulis Sims (PE), a commercially grown plant whose fruits and leaves are used for medicinal and culinary purposes. Passiflora edulis fruit is commonly known as passion fruit, whose juice has shown antioxidant activity and improved the lipid profile in male rats [7]. Previous studies have identified the presence of flavonoids as the major constituents of P. edulis, mainly C-glycosylflavones. The flavonoids’ compounds that have been described in the fruit include isoorientin, quercetin and derivatives, luteolin, schaftoside, isoschaftoside, orientin, isovitexin, apigenin, kaempferol, rutin, saponarin, vitexin, and chrysin [8, 9]. Moreover, Da Silva et al. [9] have shown the in vitro and in vivo antioxidant activity of the leave aqueous extracts. In several studies, P. edulis extracts have exhibited potential effects for the treatment of inflammation, insomnia, pain, and as well as for attention-deficit hyperactivity disorder, hypertension, and cancer [10]. We therefore hypothesized that PE fruit could prevent the oxidative stress and AD induced by aluminum chloride. The present study aimed to evaluate antioxidant and neuroprotective effects of PE through phenolic content, the in vitro antioxidant activity of different extracts as well as the in vivo activity of AChE, oxidative–antioxidative indices of aqueous extracts, flavonoids fraction, and juice in aluminum chloride-induced AD rats.
Methods
Plant material and preparation of extracts and flavonoid-rich fraction
PE fruits were purchased from a local market in the west region of Cameroon and transported to the laboratory where they were washed before use. PE fruit peels and pulp aqueous, ethanolic, and hydroethanolic (20/80) extracts were obtained by soaking powders of corresponding parts in water, ethanol, and hydroethanolic mixture for 24 h with gentle stirring, after which the mixtures were filtered using a Whatman No. 4 filter paper. The resulting filtrates were dried at 45 °C using an air oven to obtain the respective extracts: peel aqueous extract (HP), pulp aqueous extract (HG), pulp hydroethanolic extract (HEG) and peel hydroethanolic extract (HEP), pulp ethanolic extract (EG), and peel ethanolic extract (EP).
To obtain the flavonoid-rich ethylacetate fraction (EAFO), dried aqueous and ethanolic extracts (200 g each) of the peel were re-suspended in water then fractionated with n-hexane in a separating funnel, and n-hexane portion was discarded after separation. The dichloromethane was then added to the aqueous portion. After discarding the dichloromethane portion, the aqueous portion was collected and further extracted with ethyl acetate. The ethyl acetate fraction was collected and dried under rotary vacuum evaporator. The yield of the ethyl acetate portion was 35. 36% (w/w). This fraction was further subjected to qualitative chemical test and thin layer chromatography to confirm the presence of flavonoids.
Determination of in vitro antioxidant activity
Phenolic content
Total phenolic content of extracts were analyzed using the Folin Ciocalteu method as described by Dohou et al. [11]. Respectively 1.39 ml and 0.2 ml of distilled water and Folin reagent were added to 0.01 ml of extract 5 mg/ml. After 3 min of rest, 0.4 ml of sodium carbonate (Na2CO3, 20%) was added. The mixture was vortexed and incubated for 20 min at 40 °C using a water bath, thereafter the absorbance was read against a blank at 760 nm using a BioMate 6 UV–vis spectrophotometer (BIOMATE). The total phenolic content was determined using the standard curve (y = 0.022 x; r2 = 0.9945) obtained with Gallic acid. The contents were expressed as mg of Gallic Acid Equivalent/g of extract.
Determination of flavonoid content
Flavonoid content was determined according to a previously described method [12]. Briefly, 0.1 ml of extract was mixed with 1.4 ml of distilled water before the introduction of 0.03 ml of 5% sodium nitrite (NaNO2) solution. After 5 min resting, 0.2 ml of 10% (AlCl3) solution was added. After 5 min resting, 0.2 ml 10% (NaOH) solution and 0.24 ml of distilled water were added and the absorbance was measured at 510 nm using a BIOMATE. The flavonoid content was determined using the standard curve (y = 0.1972 x; r2 = 0.9972) obtained with catechin. The contents were expressed as mg CE/g of extract.
Evaluation of the antioxidant activity by the DPPH free radical scavenging assay
Antioxidant activity of different extracts was determined according to the method described by Mensor et al. [13]. 100 μl of ethanol and 100 μl of different extracts were mixed, afterwards, we proceeded to dilutions for the final concentrations of 200, 100, 50, 25, and 12.5 mg/ml. A volume of DPPH solution (0.1 mM) dissolved in ethanol was added. For each concentration, the test was done in triplicate. The mixture was then kept at room temperature and in the dark for 30 min. Absorbances were measured at 517 nm using a BIOMATE. The antioxidant activity, which expresses the ability to trap the radical DPPH° was estimated by the DPPH inhibition percentage. The antioxidant activity was calculated using the value of the EC50 (efficient concentration 50).
Assessment of the ferric reducing power
The ferric reducing ability of extracts was determined according to the method described by Oyaizu [14]. 100 μl of extracts solutions at the following concentrations: 2000, 1000, 500, 250 and 125 μg/ml were added to 500 μl of a phosphate buffer solution (0.2 M, pH 6.6) and 500 μl of aqueous potassium hexacyanoferrate solution [K 3 Fe (NC) 6] 1%. After 30 min of incubation at 50 °C in a water bath, 500 μl of trichloroacetic acid solution 10% were added. The mixture was centrifuged at 3000 g for 10 min, and 500 μl of supernatant was removed and mixed with 500 μl of distilled water followed by 100 μl of an ethanolic solution of FeCl3 0. 1%. The absorbance of the mixture was read at 700 nm against the blank using a BIOMATE. BHT was used as a standard.
In vivo antioxidant and neuroprotective effect of PE aqueous extract, EAFO, and crude juice on oxidative stress induced by aluminum chloride
Experimental animals, induction of Alzheimer’s disease (AD) in rats and diets
3-month Wistar rats weighing between 200 and 230 g were obtained from the Department Animal Centre and allowed to be accustomed to the new environment for 1 week. They were maintained in accordance with the guidelines of the OECD [15]. The animals were individually housed under controlled temperature (25 °C) and lighting (12-h light/12-h dark cycle) and had free access to water and diet. After acclimation, the induction of Alzheimer’s disease was performed by a daily intraperitoneal administration of 32.5 mg / kg bw of aluminum chloride (AlCl3) (except in the negative control group (NC)) during 60 consecutive days. At the end of the induction timeframe, the rats were randomly distributed into seven groups of 10 animals each (including two controls) and treated for another 28 days as follows:
-
Group (NC): normal, healthy rats served as negative controls and received water
-
Group (PC): AD-induced rats (positive control rats) received water
-
Group (A.E200): AD-induced rats received 200 mg/kg bw peel aqueous extract
-
Group (A.E400): AD-induced rats received 400 mg/kg bw peel aqueous extract
-
CJ: AD-induced rats received 1 ml crude juice per 200 g bw;
-
VE400: AD-induced rats received 400 mg/kg bw of Vitamin E
-
EAFO: AD-induced rats received 400 mg/kg bw of peel flavonoid fraction.
-
Animal were feed a basal diet composed as follows: corn flour (77.8%), fish flour (20%), bone (0.1%), palm olein (1%), vitamins (0.1%), and salt (1%). All experiments were carried out according to the regulations and ethical approval of the Experimental Animal Welfare and Ethics Committee of the Institution (No. 2017/056).
Assessment of animal behavior during treatment and Morris water maze test
Various physical symptoms of stress were observed daily 4 h after injection [15]. Sensitivity to pain was assessed by tail pinch. The motor activity was evaluated by observing the movement of the animal. Noise sensitivity was evaluated by causing a metallic noise impact using two iron rods next to the cage, the animal jumped sensitive. Aggressiveness was evaluated by stopping the animal.
Spatial learning and memory was assessed as previously described by Morris [16]. The experiment was based on the ability of the rats to learn how to escape from the pool by locating a transparent, submerged platform, climb, and stay on it in order to be returned to their cage. The Morris water maze test was carried out in a large circular pool (160 cm diameter) filled with water (30 cm depth) and included an acquisition trial and a probe trial. Four points around the edge of the tank were designated north (N), south (S), east (E), and west (W), thus providing four alternative start positions and dividing the pool into four quadrants (NW, NE, SE, and SW). The invisible platform was placed 2 cm below the water level of northeast quadrant. During the acquisition trial, the rats were trained to locate the platform (up to 90 s) thrice a day for 5 days. All the rats were first placed on the platform for 30 s, before being placed at a start point. If the rats reached the platform during the 90 s, they were allowed to remain for 30 s in the platform. When the rats failed to reach the platform during the 90 s, they were guided to the platform and then allowed to remain for 30 s. In the sixth day, the rats were individually subjected to a 90 s probe trial in which the platform was removed from the pool. The time spent swimming in target quadrant (within 90 s probe test time) was recorded. Rats that had no or less deteriorations in memory functions (negative control and treated groups) were expected to remember the platform location and to spend more time swimming within the target quadrant, looking for it, in comparison to the Alzheimer’s rats.
Tissue preparation for biochemical estimation
After 28 days of treatment, animals were sacrificed under anesthesia using steam chloroform. Blood was collected in EDTA tubes. Immediately after collection, the liver and brains were removed carefully. Different parts of the brain, i.e., hippocampus and cortex, were separated and homogenized in ice-cold phosphate buffer pH 7.4. Plasma was prepared from the collected blood and homogenates from the liver and brains. The brain homogenate were centrifuged at 800×g for 5 min at 4 °C to remove the nuclear debris. The supernatant was used for estimation of malondialdehyde (MDA) content and acetylcholine esterase activity. The remaining supernatant was further centrifuged at 10,000×g for 30 min at 4 °C to get the post-mitochondrial supernatant which was used for the estimation of reduced glutathione (GSH) and activities of SOD, catalase, and glutathione peroxidase (GPx).
Determination of thiobarbituric acid reactive substances (TBARS)
MDA was evaluated according to Yagi [17].Briefly, 100 μl of plasma or homogenate, 500 μl of 1% thiobarbituric acid (TBA reagent), and 500 μl of 1% phosphoric acid were introduced in the tubes. The mixture was heated in a water bath at 100 °C for 15 min and then cooled in a cold water bath for 30 min. They were centrifuged at 3000×g for 10 min, and the absorbance of the supernatant was read at 532 nm against the blank using a BIOMATE. The concentration of MDA was determined using the molar extinction coefficient (ε = 1.53 × 105 M− 1 cm− 1).
Determination of total protein
Total protein was done according to the protocol proposed by Gornall et al. [18]. In each tube were added 50 μl of plasma or homogenate, 2950 μl of 0.9% NaCl, and 3000 μl of Biuret reagent. All tubes were homogenized and incubated at room temperature for 15 min, and their optical density was read against the blank at 540 nm. Protein concentration of each sample (in milligram per milliliter or milligram per gram) was determined from the calibration curve.
Determination of glutathione
Glutathione was performed according to the method described by Ellman [19]. Homogenate (100 μl) was mixed to 900 μl of Ellman’s reagent prepared in tris-HCl buffer (0.1 M, pH 6.5). After homogenization, the mixture was incubated at room temperature for 30 min. Optical densities were read at 412 nm against the blank. The concentration of thiol group (SH) was determined using the molecular extinction coefficient DTNB (ε = 1.36 × 105 M− 1 cm− 1). The results were expressed in millimole per gram of proteins.
Assay of acetylcholinesterase activity
The enzyme activity was assessed according to the procedure of Ellman et al. [20]. Acetylthiocholine was hydrolyzed by acetylcholinesterase (AChE) to acetic acid and thiocholine. In brief, an aliquot of cortex or hippocampus homogenate (0.01 ml) was added to tubes containing 1.5 ml of phosphate buffer (100 mmol/l, pH 8.0), 0.01 ml of acetylthiocholine solution (75 nmol/l), and 0.05 ml of DTNB. The catalytic activity was measured by following the increase of yellow anion, 5-thio-2-nitrobenzoate, produced from thiocholine when it reacted with DTNB at 410 nm.
Assessment of catalase activity
Catalase was assayed according to the method of Sinha [21]. In brief, 50 μl of homogenate or plasma and 750 μl of phosphate buffer (0.01 M; pH 7.0) were added to 200 μl of hydrogen peroxide (200 mM). The reaction was stopped after 60 s by adding 2 ml of dichromate in acetic acid (1: 3v/v of 5% potassium dichromate with concentrated acetic acid). After heating at 100 °C for 10 min, tubes were cooled in an ice bath, and the optical densities were recorded at 570 nm against the blank (50 μl of 0.9% NaCl). The catalase activity was expressed in micromole of hydrogen peroxide consumed per minute per milligram of protein.
Determination of the activity of GPx
Glutathione peroxidase (GPx) activity in brain tissues was assessed by the method of Rotruck et al. [22]. Briefly the reaction mixture contained 0.2 ml of tris–HCl buffer (0.4 mol/l, pH 7.0), 0.2 ml of reduced GSH (1 mmol/l), 0.1 ml of sodium azide (10 mmol/l), 0.1 ml of H2O2 (1 mmol/l), and 0.2 ml of tissue sample. After incubation at 37 °C for 10 min, reaction was stopped by the addition of 0.4 ml of 10% trichloroacetic acid, and tubes were subjected to centrifugation at 2400 r/min for 10 min. The supernatant (0.2 ml) was then added with 0.1 ml Ellman’s reagent (0.019 8 g of DTNB prepared in 0.1% sodium citrate). Absorbance was read at 340 nm.
Assay of SOD
The activity of superoxide dismutase (SOD) was assayed according to the method of Oberley [23]. Each 1 ml reaction mixture consisted of 960 μl of 100 mM sodium carbonate buffer (pH 7.8) containing 0.1 mM xanthine, 0.025 mM nitroblue-tetrazolium (NBT), and 0.1 mM EDTA, 20 μl of xanthine oxidase and 20 μl of the brain supernatant. Changes in absorbance were monitored spectrophotometrically at 560 nm following the production of blue formazan. One unit of SOD was defined as the quantity required to inhibit the rate of NBT reduction by 50%. The enzyme activity was expressed as unit per minute per milligram protein.
Estimation of beta-amyloid (Aβ) peptide and proinflammatory markers by ELISA in the hippocampus
Tumor necrosis factor-α (TNF-α), interleukin-6 (IL-6), and cycyclooxygenase-2 (COX-2) levels were assayed by the method described in the commercial ELISA kit instructions purchased from Ray Biotech, Canada. Sandwich ELISA was used for the quantification of Aß40 and Aß42 in the hippocampus using ELISA kit according to the manufacturer’s protocol of Ray Biotech, Canada. The absorbances were measured immediately at 450 nm against blank using an ELISA reader.
Histological analysis of hippocampal CA1 region
After the experimental period, the brains were dissected out and post-fixed overnight in paraformaldehyde. Coronal brain sections of 8 μm thick were cut in a coronal plane using microtome. The sections were stained with cresyl violet and mounted. The stained sections of the rat hippocampal CA1 region were examined under the light microscope and photographed using a digital camera for studying the morphological changes of hippocampal CA1 pyramidal neurons.
Statistical analysis
Statistical analysis was performed using SPSS program version 21. In vitro experiments were performed in triplicate. Results were expressed as mean ± standard deviation (SD). One-way analysis of variance (ANOVA) with Bonferroni test was used for statistical analysis of the mean difference among groups. The post hoc Tukey helped highlighting the significant differences between the threshold averages. Differences were considered significant at P < 0.05 (at 95% confidence interval).
Results
Result of in vitro antioxidant activity
Total phenolic compounds and flavonoid contents
Table 1 shows the total phenolic and flavonoid contents of different extracts of PE.
HP presented the greatest value of phenolic compounds and flavonoids (54.6 ± 3.84 mg GAE/g of extract and 29.78 ± 3.1 mg CE/g of extract respectively), followed by HEG (38.25 ± 1.98 GAE mg/g of extract) and the EP (33.8 ± 0.82 mg GAE/g of extract). The lowest value was recorded with HG (22.85 ± 4. 36 mg GAE/g of extract). The lowest value was recorded with HEG (4.7 ± 1.32 mg CE/g of extract). There was no significant difference between HEP and HEG and between EG and H.G. Aqueous extract that presented the highest flavonoid content also showed the highest content in phenols.
DPPH scavenging (2,2-diphenyl-1-picrylhydrazyl)
Figure 1 shows the antiradical activity of PE extracts at various concentrations compared with that of vitamin C. It is clear from this figure that PE extracts exhibited antiradical activity. Similarly, all these extracts showed lower antiradical activity than vitamin C. Table 2 showed the values of EC 50 (efficient concentration 50) above that of vitamin C (4.40 ± 0.02 μg/ml). The antiradical activity and EC50 were highest with HP. The lower antiradical activity was recorded with HA.

Change in DPPH scavenging activity of different extracts of PE at different concentrations compared to that of vitamin C. HP aqueous extract of the peel; HG aqueous extract of the pulp; HEP hydro-ethanol extract of the peel; HEG hydro-ethanol extract of the pulp; EP ethanol extract of the peel; EG ethanol extract of the pulp; VITC vitamin C
Ferric reducing antioxidant power (FRAP)
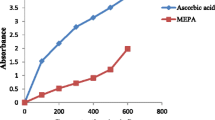
Figure 2 shows the trends in the reducing power of various extracts of PE at different concentrations compared to vitamin C. The reducing power of vitamin C was significantly (P < 0.05) higher than that of PE extracts. Similarly the reducing power of HP was greater and significantly different (P < 0.05) from HEP, EP, EG, and HG. Overall, the reducing power was correlated to the polyphenol contents and DPPH scavenging activity.

variation of reducing power of PE extracts at different concentrations compared to that of vitamin C. HP aqueous extract of the peel; HG aqueous extract of the pulp; HEP hydro-ethanol extract of the peel; HEG hydro-ethanol extract of the pulp; EP ethanol extract of the peel; EG ethanol extract of the pulp; VITC vitamin C
Effect of peel aqueous extract, EAFO, and crude juice of PE on animal behavior and cognitive function
Table 3 shows the effect of the crude juice and different doses of PE extract on the animal behavior. After induction by aluminum chloride, all groups showed little reaction to noise, decreased locomotion, and a marked sleepiness during the first 3 h. During the administration of the extract at different concentrations and crude juice, a reduction of the aggressiveness in the treated group was observed from the second week. We observed the presence of aggressive animals in the untreated group compared to those induced and treated with different doses of aqueous extracts (doses of 200 and 400 mg/kg bw).
Spatial memory effectiveness was assessed over five consecutive days using an invisible platform. All groups displayed a gradual improvement in learning dysfunction over the 5 days of testing period (result not shown). During the probe trial on the sixth day, rats treated with AlCl3 took more time to reach the platform (Fig. 3a) thus spending less time in the target quadrant (Fig. 3b) in comparison to the control group. But administration of PE significantly reduced the time taken to reach the platform in both trials and significantly improved the time spent in the target quadrant during the probe trial. The 400 dose of PE was more effective than that of vitamin E and EAFO though not significant.

– Effect of PE on the spatial memory by measuring the time (s) required to reach the platform (a) and time spent in the target quadrant (b) during Morris water maze test. USOD = amount required to inhibit 50% nitroblue tetrazolium reduction, UAChE nmol of substrate hydrolyzed/min/mg protein. UGPx = nmol of nicotinamide adenine dinucleotide oxidized/min/mg protein. Data are expressed as mean ± SD. Values with different letters differ significantly at P < 0.05
Effect of PE on some biochemical parameters
Effect of crude juice and aqueous extract of PE peel on the plasma and tissue proteins of male rat
Table 4 shows the effect of the crude juice and aqueous extracts of PE peel in different doses on the plasma and tissue proteins levels. In general, we found that the stress induction resulted in a significant decrease (P < 0.05) in the plasma, liver, and brain proteins in animals. However, administration of the juice, the extract at different doses, and vitamin E resulted in a significant increase (P < 0.05) plasma protein levels and homogenates compared to positive control group with the exception of brain homogenates which did not show a significant increase at the dose 200 mg/kg.
Effect of crude aqueous extract and juice of PE peel on the plasma and tissue malondialdehyde (MDA)
Table 5 below shows the effect of stress induction and administration of the extract and crude juice on the level of the plasma, liver and brain malondialdehyde in rats. It was noted that the stress induction resulted in a significant increase in the plasma, liver, and brain malondialdehyde. Overall, administration of vitamin E, EAFO extracts, and crude juice resulted in a significant decrease (P < 0.05) of malondialdehyde in the liver and brain compared to the positive control group.
Effect of flavonoid fraction, crude juice, and peel aqueous extract of PE on the activity of the plasma, liver, and brain catalase
Aluminum-treated rats exhibited high AChE activity and a significant decrease (P < 0.05) in CAT levels, SOD, and GPx activities in hippocampus and cortex in comparison to controls. However, PE administration significantly and dose dependently inhibited AChE activity in comparison to AlCl3 untreated group (Fig. 4). The effect of flavonoid fraction was comparable to that of aqueous extract but greater than that of crude juice and vitamin E. This suggests that flavonoids accounts for most of the bioactive compounds responsible for the effect of the aqueous extract.

Antioxidant effect of PE on the activities of superoxide dismutase (SOD) (a), glutathione peroxidase (GPx) (b), and AChE (c) in the hippocampus and cortex of control and experimental rats
Table 6 shows the effect of oxidative stress and treatments on the activity of the plasma, liver, and brain catalase. It was found that the stress induction resulted in a significant decrease (P < 0.05) activity of plasma and brain catalase. The administration of flavonoid fraction, vitamin E, extracts, and crude juice led to a significant increase (P < 0.05) in the activity of catalase in plasma in all treated groups compared to the positive control group, and for the brain and liver in the group that received the extract and flavonoid fraction. Likewise, oral administration of flavonoid fraction and aqueous PE extract during AlCl3 exposure showed an improvement in SOD and GPx by significantly increasing their values. Similar effects were observed with vitamin E and flavonoid fraction, though to a lesser extent compared to the higher dose of PE (400 mg/kg).
Effect of flavonoid fraction, crude juice, and aqueous extracts of PE peel on the plasma and tissue glutathione levels
Table 7 shows the effect of the flavonoid fraction, crude juice, and aqueous extracts of PE on the plasma, liver, and brain reduced glutathione levels in rats. In general, the stress induction resulted in a significant decrease in the plasma, liver, and brain reduced glutathione levels. However, administration of flavonoid fraction, vitamin E, extracts, and juice significantly increased (P < 0.05) the levels of reduced glutathione in the plasma and organs compared to that of the positive control.
Effect of flavonoid fraction, crude juice, and aqueous extracts of PE peel on the proinflammatory and amyloidogenic markers
Chronic exposure to AlCl3 significantly (P < 0.05) increased the production of brain proinflammatory molecules such as TNF-α, IL1-β, and COX-2 accompanied with augmentation of Aβ42 and suppression of Aβ40 (Fig. 5). PE co-administration attenuated the neuro-inflammation. It reduced the cytokines level in the hippocampus tissues that are associated with the amyloid beta protein. It also induced the anti-amyloidgenic protein Aβ40.

Effect of PE on brain tumor necrosis factor-α (TNF-α), interleukin-6 (IL-6), cycyclooxygenase-2 (COX-2), amyloid ß-40 (Aß-40), and amyloid ß-42 (ß-42) in male rats treated with aluminum chloride. Values are expressed as means ± SD for 10 rats in each group. Means with different letters (a, b, c, d) vary significantly at P < 0.05. Values with the same letters are not significantly different at P < 0.05
Effect of flavonoid fraction, crude juice, and aqueous extracts of PE peel on the AlCl3-induced neurodegeneration in the rat hippocampal CA1 area
In order to correlate the PE neuroprotective effects with a typical Alzheimer event, we finally investigated the impact of the PE extract and flavonoid fraction on AlCl3-induced neurodegeneration in the hippocampal CA1 region (Fig. 6). The microscopic examination of this brain region revealed degenerating pyramidal neurons with pyknotic nuclei, neuron swelling, vacuolation, and apoptotic cells upon long exposure to AlCl3 (Fig. 6b), while there was a normal cytoarchitecture of matured pyramidal cells in normal control (Fig. 6a). Co-administration of AlCl3 and PE extract and flavonoid fraction significantly attenuated the AlCl3-induced degenerative and morphological changes of the pyramidal cells in the hippocampal CA1 region.

PE reduces AlCl3-induced neurodegeneration in the rat hippocampal CA1 region: after the experimental period, the brain sections were prepared, stained with cresyl violet, and cells were observed under a light microscope. a Healthy rat with round nucleated healthy pyramidal cells. b AlCl3-treated rat with degenerative cells. c A.E200. d A.E400. e CJ. f VE 400. g EAFO
Discussion
To recap, this work was a comparative study of the effect of the most active (in vitro antioxidant activity) extract and flavonoid fraction in a rat model of Alzheimer’s disease. Thus, the evaluation of the in vitro antioxidant activity of different extracts allowed us to conclude that these fruits contain phenolic compounds endowed with very important biological activities. We then continued the assessment of the in vivo antioxidant and neuroprotective activities of the peel aqueous extract of PE because it presented the best in vitro activity, along with flavonoid fraction and crude juice.
The reducing power of the PE is due to the presence of hydroxyl groups in the phenolic compounds such as flavonoids that serve as electron donor reducing ferric ions to ferrous ions thereby increasing the amount of ferrous ion and resulting in an increase of the reducing power of the extract. (Siddhuraju and Becker) [24]. These results are similar to those of El-Missiry et al. [25] who showed that the antioxidant capacity of the plants is mainly due to their high phenolic compounds, hence to their hydroxyl groups. With regards to Souri et al. [26] antiradical activity classification, we can conclude that our extracts had moderate antioxidant activity with the exception of the pulp aqueous extract that had a low antioxidant activity. The antiradical activity was due to the presence of hydroxyl groups in our extracts which transfers their proton to stabilize the radical DPPH.
Phenolic compounds are used by plants to overcome stress conditions; the observed variations are due to extraction solvents and parts of fruit that were used [27] According to Anokwuru et al. [28], for the same solvent used, the phenolic content may vary according to the parts used. Similarly, studies by Rudnicki et al. [29] showed a higher total phenolic content with ethanolic extract of PE leaves. This would be due to the fact that flavonoids belong to the phenolic group. Birt et al. [30] showed that the flavonoids represented about two-thirds of total polyphenols.
It was necessary to investigate the status of the glutathione antioxidants that are the first line of defense against the free radicals’ damage. These results are similar to those reported by Al-Hashem et al. [31] who showed that metals such as cadmium and mercury result in oxidative stress by reduction in renal and brain intracellular glutathione. Indeed, polyphenols, namely flavonoids, modulate the expression of an important enzyme gamma glutamyl synthetase in the cellular antioxidant defense. This enzyme is responsible for the rate of synthesis of glutathione. These studies are consistent with those of Moskaug et al. [32] who showed that flavonoids increase the expression of gamma-glutamyl synthetase in vitro and in vivo in mice transgenic strain.
The decrease in the plasma, liver, and brain protein levels in the positive control group compared to that of the treated groups was due to a decrease in food intake and the increase in catabolism of plasma proteins. Similarly, it could also be due to reactive oxygen species formed such as hydroxyl radicals responsible for the oxidation of the side chains of some amino acids resulting in proteins hydrolysis. These changes could affect their functions, antigenicity, and proteolytic degradation in the proteasome. These results are in agreement with those of Yokel and McNamara [33] who showed a decrease in protein levels after intraperitoneal injection of aluminum in rats at a dose of 40 mg/kg bw. However, the administration of the flavonoid fraction, aqueous extract, and juice of PE increased plasma protein levels, hepatic, and brain. This could be due to the appetite increase and the presence of phenolic compounds that transfer their proton to limit oxidation of the amino acids.
Increase in MDA levels in the positive control suggests the involvement of cellular damage caused by free radicals. Indeed, the aluminum leads to the reduction of the activity of several antioxidant enzymes in different parts of the brain thus facilitating the spread of lipid peroxidation. Similar results were reported by Sadhana [34] which showed that aluminum nitrate induces an increase level of MDA concentrations in the rat brain and liver. The treatment with the aqueous extract at a dose of 200 and 400 mg significantly reduced plasma MDA levels while the hepatic and brain levels of MDA were also reduced upon administration of the crude juice and flavonoid fraction.
Among the antioxidant enzymes, catalase is crucial and directly eliminates hydrogen peroxide. Catalase is present in mammals’ peroxisomes, and the highest activity is located in red blood cells and in the liver. This increase in catalase activity could be due to the presence of bioactive compounds present in our extracts and juices that would give their proton to stabilize radicals formed during stress. These results corroborate those of De Souza et al. [7] who showed that the juice of PE reduced oxidative stress in Wistar rats. Acetylcholine esterase is the enzymatic marker for cholinergic neurons and is responsible for the breakdown of acetylcholine, a neurotransmitter related to short-term memory. Our results show that the aluminum treatment for 60 days significantly elevated the activity of AChE in both cortex and hippocampus a result that is consistent with previous findings [35, 36]. The inhibition of the activities of AchE and the reduction of the neuroinflammatory markers by the peel aqueous extract and flavonoids fraction increased the acetylcholine with positive impact on the cognitive function. Our study clearly showed that PE possesses anticholinergic, anti-neuroinflammatory, anti-amyloidgenic, and antioxidant properties thus could reverse aluminum-induced cognitive dysfunction. In this study, we also investigated the histological and behavioral changes caused by AlCl3 exposure and the possible effect of PE treatment. We observed that chronic aluminum chloride exposure caused a significant morphological changes in hippocampal CA1 region such as alteration of the pyramidal cellular arrangement. Co-administration of PE together with AlCl3 could alleviate the aluminum-induced degenerative changes. The stress induction was done by aluminum chloride which is a neurotoxin that exerts its toxic effect by interfering with pathways involved in metabolism and normal iron homeostasis [37]. In Morris water maze test, aluminum administration was associated with decreased spatial memory as evidenced by the results. The cognitive function was restored upon treatment with PE. The greater overall effect of the aqueous extract compared to the flavonoids fraction (though there was no significant difference) indicated that besides flavonoids, other bioactive compounds could also be responsible to the neuroprotective effect of this fruit.
Conclusion
This study suggests that the aqueous extract and flavonoid fraction of the peel of PE as well as crude juice have antioxidant compounds that could be used to alleviate oxidative stress, neuroinflammation, and Azheimer’s disease, thereby could prevent chronic diseases that occur during aging. The flavonoids are mainly responsible for its activity though other bioactive compounds may also participate. Thus, this fruit could be used as a nutraceutical.
Abbreviations
- AChE:
-
Acetylcholinesterase
- CAT:
-
Catalase
- DPPH:
-
2,2-Diphenyl-1-picrylhydrazyl
- FRAP:
-
Ferric reducing antioxidant power
- GPx:
-
Glutathione peroxidase
- GSH:
-
Reduced glutathione
- MDA:
-
Total malondialdehyde
- SOD:
-
Superoxide dismutase
References
Love S, Jenner P. Oxidative stress in neurological disease. Brain Pathol. 1999;9:55–6.
Youdim M. Iron in the brain implications for Parkinson's and Alzheimer's diseases. Mt Sinai J Med. 1988;55(1):97–101.
Verstraeten SV, Nogueira LV, Schreier S, Oteiza PI. Effect of trivalent metal ions on phase separation and membrane lipid packing: role in lipid peroxidation. Arch Biochem Biophys. 1997;338(1):121–7.
Fatma M. El-Demerdash. Antioxidant effect of vitamin E and selenium on lipid peroxidation, enzyme activities and biochemical parameters in rats exposed to Aluminium. J Trace Elem Med Biol. 2004;18(1):113–21.
Esposito E, Rotilio D, Di Matteo V, Di Giulio C, Cacchio M, Algeri S. A review of specific dietary antioxidants and the effects on biochemical mechanisms related to neurodegenerative processes. Neurobiol Aging. 2012;23:719–35.
Commenges D, Scotet V, Renaud S, Jacqmin-Gadda H, Barberger-Gateau P, Dartigues J. Intake of flavonoids and risk of dementia. Eur J Epidemiol. 2000;16:357–63.
De Souza M, Barbalho S, Damasceno D, Rudge M, De Campos K, Madi K. Effects of Passiflora edulis (yellow passion) on serum lipids and oxidative stress status of Wistar rats. J Med Food. 2012;15(1):78–82.
Dhawan K, Dhawan S, Sharma A. Passiflora: a review update. J Ethnopharmacol. 2004;94(1):1–23.
Da Silva J, Cazarin C, Colomeu T, Batista A, Meletti L, Paschoal J, Júnior S, Furlan M, Reyes F, Augusto F, Júnior M, Zollner R. Antioxidant activity of aqueous extract of passion fruit (Passiflora edulis) leaves: in vitro and in vivo study. Food Res Int. 2013;53:882–90.
Spencer KC, Seigler DS. Cyanogenesis of Passiflora edulis. J Agric Food Chem. 1983;31(4):794–6.
Dohou N, Yamni K, Tahrouch S, Idrissi-Hassani L, Badoc A, Gmira N. Screening phytochimique d'une endémique ibéro-marocaine, Thymelaealythroides. Bull Soc pharmacolo Bord. 2003;142:61–78.
Zhishen J, Mengcheng T, Jianming W. The determination of flavonoid contents in mulberry and their scavenging effects on superoxide radicals. Food Chem. 1999;64:555–9.
Mensor L, Menezez F, Leitao G, Reis A, Dos Santos T, Coube C, Leitao S. Screning of Brazilian plant extracts for antioxidant activity by the use of DPPH free radical method. Phytoth Res. 2001;15:127–30.
Oyaizu M. Studies on products of browning reaction prepared from glucose amine products derived from bees. J Pharm Biomed Anal. 1986;41:1220–34.
OECD. Guidelines for chemicals trials: repeated dose oral toxicity studies for 28 days in rodents: OECD; 2008. p. 1–29. https://ntp.niehs.nih.gov/iccvam/suppdocs/feddocs/oecd/oecdtg407-2008.pdf
Morris R. Developments of a water-maze procedure for studying spatial learning in the rat. J Neurosci Methods. 1984;11:47–60.
Yagi K. Simple fluorometric assay for lipoperoxyde in blood plasma. Biol Med. 1976;15:212–6.
Gornall A, Gomez-Caravaca A, Gomez-Romero M, Arraez-Roman D, Segura-Carretero A, Barwill G, David M. Determination of serum protein by means of the buiret reaction. J Biol Chem. 1949;177:751–66.
Ellman GL. Quantitative determination of peptide by sulfhydryl (-SH) groups. Arch Biochemi and Biophys. 1959;82:70–7.
Ellman GL, Courtney KD, Andres V Jr, Featherstone RM. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochem Pharmacol. 1961;7:88–95.
Sinha K. Colorimetric assay of catalase. Biochem Anal. 1972;47:389–94.
Rotruck JT, Pope AL, Ganther HE, Swanson AB, Hafeman DG, Hoekstra WG. Selenium: biochemical role as a component of glutathione peroxidase. Science. 1973;179:588–90.
Oberley LW. Inhibition of tumor cell growth by overexpression of manganese-containing superoxide dismutase. Age. 1998;21:95–7.
Siddhuraju P, Becker K. The antioxidant and free radical scavenging activities of processed cowpea (Vigna unguiculata (L) Walp) seed extracts. Food Chem. 2007;101(1):10–9.
El-Missiry M, El Gindy A. Amelioration of Alloxan induced diabetes mellitus and oxidative stress in rats by oil of Eruca sativa seeds. Ann Nutr Metabol. 2000;44:97–100.
Souri E, Amin G, Farsam H, Barazandeh TM. Screening of antioxidant activity and phenolic content of 24 medicinal plant extracts. Daru. 2008;16(2):83–7.
Shan B, Cai Y, Sun M, Corke H. Antioxidant capacity of 26 spices extracts and characterization of their phenolic constituents. J Agric Food Chem. 2005;53(20):7749–59.
Anokwuru C, Ajibaye O, Adesuyi A. Comparative antioxidant activity of water extract of Azadiractha indica stem bark and Telfairia occidentalis. Leaf Med Res. 2010;41:197–200.
Rudnicki M, De Oliveira M, Pereira T, Reginatto F, Dal-Pizzol F, Moreira J. Antioxidant and antiglycation properties of Passiflora alata and Passiflora edulis extracts. Food Chem. 2007;100(2):719–24.
Birt D, Hendrich S, Wang W. Dietary agents in cancer prevention: flavonoids and isoflavonoids. Pharmacology. 2001;90:157–77.
Al-Hashem F, Dallak N, Bashir M, Abbas R. Camel’s milk protects against cadmium chloride induced toxicity in white albino rats. Am J Pharmacol Toxicol. 2009;4:107–17.
Moskaug J, Carlsen H, Myhrstad MC, Blomhoff R. Polyphenols and glutathione synthesis regulation. Am J Clin Nutr. 2005;81(1):277–83.
Yokel R, McNamara PJ. Elevated aluminum persists in serum and tissue of rabbits after a six-hour infusion. Toxicol Appl Pharmacol. 1989;99:133–8.
Sadhana S. S-allyl-cysteines reduce amelioration of aluminum induced toxicity in rats. Am Jo Biochemy Biotech. 2011;7(2):74–83.
Hussien HM, Abd-Elmegied A, Ghareeb DA, Hafez HS, Ahmed HEA, El-moneam NA. Neuroprotective effect of berberine against environmental heavy metals induced neurotoxicity and Alzheimer's-like disease in rats. Food Chem Toxicol. 2018;111:432–44.
Yuliani S, Mustofa, Partadiredja G. The neuroprotective effects of an ethanolic turmeric (Curcuma longa L.) extract against trimethyltin-induced oxidative stress in rats. Nutr Neurosci. 2018. https://doi.org/10.1080/1028415X.2018.1447267.
Banksa W, Niehoffa M, Dragob D, Zatta P. Aluminum complexing enhances amyloid β protein penetration of blood-brain barrier. Brain Res. 2006;1116:215–21.
Acknowledgements
The authors are thankful to the University of Dschang that provided lab facilities, and to all the friends, lab colleagues, and the fellows who directly or indirectly helped us during this experiment. No financial support was received from any sources for this work.
Availability of data and materials
All the generated and analyzed data are included in this article.
Author information
Authors and Affiliations
Contributions
DK and APNK are involved in designing the experiment. HTS, DK, and APNK performed the experimental work and statistical analysis. HTS and DK wrote the first and final draft. All the authors read and approved the final draft.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable as no study on human was performed.
Consent for publication
Not applicable
Competing interests
The authors declare that they have no competing interests.
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Doungue, H.T., Kengne, A.P.N. & Kuate, D. Neuroprotective effect and antioxidant activity of Passiflora edulis fruit flavonoid fraction, aqueous extract, and juice in aluminum chloride-induced Alzheimer’s disease rats. Nutrire 43, 23 (2018). https://doi.org/10.1186/s41110-018-0082-1
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41110-018-0082-1