
Abstract
Background
Because of its poor intrinsic repair capacity, articular cartilage seldom heals when damaged.
Main body
Regenerative treatment is expected for the treatment of articular cartilage damage, and allogeneic chondrocytes or cartilage have an advantage over autologous chondrocytes, which are limited in number. However, the presence or absence of an immune response has not been analyzed and remains controversial. Allogeneic-induced pluripotent stem cell (iPSC)–derived cartilage, a new resource for cartilage regeneration, reportedly survived and integrated with native cartilage after transplantation into chondral defects in knee joints without immune rejection in a recent primate model. Here, we review and discuss the immunogenicity of chondrocytes and the efficacy of allogeneic cartilage transplantation, including iPSC-derived cartilage.
Short conclusion
Allogeneic iPSC-derived cartilage transplantation, a new therapeutic option, could be a good indication for chondral defects, and the development of translational medical technology for articular cartilage damage is expected.
Similar content being viewed by others


Background
Articular cartilage covers the ends of bones and serves as a lubricant to ensure smooth joint movements. Articular cartilage consists of chondrocytes embedded in an abundant extracellular matrix (ECM), which is composed of type II, IX, and XI collagen molecules and proteoglycans. The ECM enables the mechanical functions of the cartilage. Cartilage has a limited regenerative capacity, and its damage tends to result in degenerative conditions, impairing joint function. Microfractures and autologous osteochondral transplantation have been used for relatively small defects (less than 2–3 cm2) as a treatment for articular cartilage damage [1, 2]. Healthy articular cartilage is called hyaline cartilage and is composed of hyaline cartilage rich in collagen II and proteoglycans. However, when cartilage ECM is lost due to injury or degeneration, hyaline cartilage degenerates into fragile fibrocartilage rich in collagen I, compromising its function as an articular cartilage. During cartilage repair after injury, fibrocartilage is formed due to aberrant collagen expression. Fibrocartilage is the result of cartilage fibrosis, and in many cases, repaired fibrocartilage, which lacks the original function, shows inferior mechanical properties, and even worsens osteoarthritis symptoms [3]. Microfracture induces progenitor cells from the bone marrow to repair the defect; however, the tissue repaired by microfracture consists of fibrocartilage which does not have the mechanical robustness of hyaline cartilage and is therefore vulnerable to mechanical forces of the joint. In one case series study, microfracture showed good short-term results in the treatment of small cartilage defects, but the deterioration of results began 18 months postoperatively and was most pronounced in the International Cartilage Repair Society (ICRS)-score [4]. Clinical outcomes of microfracture have tended to either plateau or deteriorate at longer follow-ups, raising concerns about long-term results [5]. Autologous osteochondral transplantation carries the risk of donor-site morbidity.
The transplantation of autologous cultured chondrocytes is the most commonly used cell-based therapy for treating human cartilage defects. However, chondrocytes lose their chondrocyte nature after expansion in culture, and most of the repaired tissue is fibrocartilage tissue [6,7,8]. As only a limited number of autologous chondrocytes are prepared, repair is thought to occur through the trophic effect of growth factors and other factors produced by transplanted cells that stimulate host cells. This treatment is a two-stage procedure and carries the risk of donor site morbidity. Chondrocytes have been shown to have limited major histocompatibility complex (MHC) expression and immunosuppressive potential in vitro [9], and allogeneic chondrocyte transplantation has also been studied. However, their in vivo immunogenicity remains controversial [10]. One of the other major cell sources for cartilage repair is mesenchymal stem cells (MSCs), which can be obtained from bone marrow, adipose tissue, and synovium. MSCs can differentiate into chondrocytes and can also achieve therapeutic effects through paracrine effects. Furthermore, MSCs are involved in the cartilage repair process by modulating the immune response when damaged cartilage is exposed to an inflammatory environment. Recent reviews have reported that functional heterogeneity of MSCs among donors, tissues, and MSC subpopulations leads to differences in cartilage repair capacity [11].
Cartilage tissue is considered immune-privileged because it is avascular and chondrocytes are surrounded by ECM [9, 12]. The ECM inhibits immune cells from contacting chondrocytes, thus avoiding immunological reactions, even under allogeneic conditions. Allogeneic cartilage has been transplanted in clinical practice without human leukocyte antigen (HLA) matching or the use of immunosuppressive agents [13,14,15]. However, there are risks of donor shortage, heterogeneity in quality, and disease transmission. Recently, clinical trials using allogeneic human iPSCs have been conducted as a new regenerative medicine [16, 17]. iPSCs have unique pluripotency and self-renewal properties shared with embryonic stem cells (ESCs). iPSCs are created by introducing reprogramming factors into somatic cells, such as skin or blood cells, whereas ESCs are acquired from the internal cell mass of embryos. Human iPSCs do not bear the ethical issues associated with the sacrifice of embryos associated with human ESCs. iPSCs are a promising resource for the regenerative treatment of diseased or damaged organs and tissues, including articular cartilage damage, and the generation of iPSC-derived cartilage composed of chondrocytes and ECM has already been reported [18,19,20]. The self-renewal ability of iPS cells enables an unlimited supply of allogeneic iPSC-derived cartilage, solving the problems of allogeneic cartilage, such as the scarcity of donors and variations in cartilage quality among donors. Thus, allogeneic iPSC-derived cartilage transplantation may be a viable treatment option for articular cartilage damage. However, there have been reports of rejection after allogeneic chondrocyte transplantation [21,22,23,24], and it remains controversial whether transplanted allogeneic cartilage can cause an immune response. In this review, we summarize the immune response and efficacy of allogeneic cartilage transplantation for articular cartilage injury and discuss the recent advances in allogeneic iPSC-derived cartilage transplantation.
Immunogenicity of chondrocytes in vitro
Articular cartilage is generally considered immune-privileged because of its avascularity and because chondrocytes are embedded in the ECM (Fig. 1a). Previous in vitro studies have reported on the immunogenicity of chondrocytes prepared by the digestion of the ECM with collagenase or other agents. They reported that the co-culture of chondrocytes with allogeneic lymphocytes did not promote lymphocyte proliferation [9, 25,26,27]. Juvenile chondrocytes are less immunogenic than adult chondrocytes because of their lower HLA expression and have a stronger anabolic effect on ECM formation [9, 26, 28]. In addition, the immunosuppressive potential of chondrocytes has been reported, and the chondrocyte expression of B7 family members (B7-H1, B7-DC, B7-H2, B7-H3, and B7-H4), which act as inhibitory signals to T cells, chondromodulin-I, a T cell growth inhibitor, and indoleamine 2,3-dioxygenase, a mediator of immune evasion, has been suggested as a mechanism of immunosuppression [9]. These results suggest that chondrocytes are immune privileged, at least in vitro. However, juvenile chondrocytes stimulated with recombinant human interferon γ (IFNγ) show an increased expression of MHC Class I (HLA-ABC) (Fig. 1b); therefore, they can become immunogenic under inflammatory conditions, such as osteoarthritis [9].

Limited immunogenicity of articular cartilage. a Histological image of articular cartilage. ECM inhibits contact of immune cells with chondrocytes. b Expression of MHC-I molecules on chondrocytes. Chondrocytes stimulated with IFN-γ show increased expression of MHC-I, suggesting that in the absence of ECM, they can become immunogenic under inflammatory conditions. c Immune response in allogeneic iPSC-derived cartilage transplantation. In osteochondral defects, T cells are observed around the graft. On the other hand, the graft is engrafted without immune response in chondral defects
Immunogenicity of chondrocytes in vivo
There have been numerous reports of allogeneic chondrocyte transplants that are not rejected in vivo in animal models of knee cartilage defects [29,30,31,32,33,34,35,36]. However, some studies have observed certain immune responses, such as lymphocyte and macrophage aggregation [21,22,23,24, 37]; therefore, allogeneic chondrocyte transplantation in vivo is controversial. A combination of various factors, including differences in the animal species used, the method of preparation of chondrocytes for transplantation, and even the technique of defect preparation, may be responsible for these conflicting results [10]; however, the details of the immune response in allogeneic transplantation of chondrocytes have not been elucidated. Cartilage injuries are classified as osteochondral or chondral defects. One major difference between the two types of defects is that, in osteochondral defects, the graft is exposed to blood flow from the bone marrow. Blood flow can significantly affect immune responses. It has been reported that cartilage formed by the transplantation of allogeneic chondrocytes into articular cartilage defects in rats was infiltrated by immune cells migrating from the bone marrow, but not on the surface of transplants facing the joint cavity. This result suggests that the immune response occurs via the bone marrow and not the joint cavity [38].
Cellular infiltration involved in the rejection of cartilage formed by intramuscular allogeneic chondrocyte transplantation has been evaluated immunohistochemically in rats [39]. It has been suggested that activated monocytes, macrophages, and chondrocytes are involved in the lysis of the cartilage matrix during the rejection process and that the death of transplanted chondrocytes is mediated by infiltrating cytotoxic lymphocytes. There are no detailed reports on the immune responses of grafts to cartilage defects, and it is uncertain whether a similar process follows.
Allogeneic cartilage grafts in animal models
Studies of particulate juvenile allograft cartilage (PJAC) have been widely studied in allogeneic cartilage tissue transplantation. PJAC is composed of minced live cartilage allografts from juvenile donors containing chondrocytes within their native extracellular matrix. Juvenile cartilage is considered less immunogenic, and minced cartilage allows chondrocytes to diffuse out of the ECM to form new hyaline-like cartilage [9, 26, 28]. Juvenile chondrocytes show excellent ECM production capacity in vitro [26, 28], and increased fragmentation significantly enhances ECM production [40]. A study in a rabbit model showed that minced cartilage grafts embedded in atelocollagen gel repaired osteochondral defects to the same extent as autologous chondrocyte implantation (ACI) [41], whereas another study reported that PJAC repaired osteochondral defects with hyaline cartilage-like tissue and showed significantly higher modified O'Driscoll scores than controls [42]. Studies in pigs have shown that PJAC transplants repair full-thickness cartilage defects as well as autologous cartilage chips [43] and that the transplanted cells survive for at least 3 months [44]. In animal studies, including two osteochondral defect models and two full-thickness cartilage defect models, allogeneic cartilage transplantation resulted in good cartilage repair; however, the immune response was not investigated (Table 1).
Clinical studies in PJAC transplantation
Particulated juvenile allograft cartilage products (DeNovo Natural Tissue [NT], manufactured by Zimmer Biomet) have been available since 2007. By 2015, more than 8700 patients had been treated with DeNovo NT [47]. This product is a minced live cartilage graft from a juvenile donor that contains cartilage cells and surrounding ECM. Minced cartilage is expected to further promote ECM formation and repair by the graft itself. However, the data on mid- and long-term clinical outcomes are lacking. Several case series have reported short-term (2–3 years) improvements in clinical scores (KOOS, IKDC, VAS, etc.) and moderate to good filling of defects on post-transplantation MRI findings [13,14,15, 48,49,50]. Complications, such as graft hypertrophy, delamination, and displacement have been reported. However, no studies have evaluated the immune response or rejection. The evidence of PJAC transplantation for cartilage injuries remains insufficient; however, short-term studies have shown promising results (Table 2).
Immunogenicity in iPSC-derived cartilage
In a study on iPSC-derived chondrocytes, a new resource for cartilage regeneration, iPSC-derived chondrocytes showed limited HLA expression and did not induce lymphocyte proliferation in a mixed lymphocyte assay when co-cultured with allogeneic peripheral blood mononuclear cells [51]. Treatment with IFNγ induces the expression of major histocompatibility complex (MHC) class I, but not MHC class II, in iPSC-derived chondrocytes, being similar to juvenile chondrocytes and potentially immunogenic under inflammatory conditions [9, 51]. Thus, the immune response of iPSC-derived chondrocytes and juvenile chondrocytes in an in vivo inflammatory environment such as osteoarthritis should be further evaluated. These results collectively suggest that iPSC-derived chondrocytes have similar immunogenic properties to those of juvenile chondrocytes in vitro, so allogeneic iPSC-derived cartilage transplantation can be performed without the use of immunosuppressive agents as in PJAC transplantation.
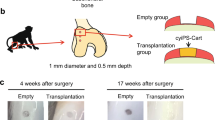
Allogeneic transplantation of iPSC-derived cartilage was performed in a primate model by mismatching the MHC, which is structurally similar to HLA, to verify the immune response in vivo. Cartilages generated from cynomolgus monkey iPS cells (cyiPSCs) were transplanted into chondral or osteochondral defects in the femoral trochlea of MHC-mismatched monkeys without the use of immunosuppressive drugs. Four weeks after allogeneic transplantation, although the graft remained intact, an accumulation of CD3-positive T cells was observed around the graft in osteochondral defects. In contrast, in chondral defects, the graft is engrafted without lymphocyte accumulation [45, 46]. It has been suggested that in chondral defects, the immune response is suppressed because there is no contact between the graft and bone marrow (Fig. 1c).
Allogeneic iPSC-derived cartilage transplantation for chondral defects
Allogeneic transplantation of cyiPSC-derived cartilage for chondral defects showed that the cyiPSC-derived cartilage was engrafted and contributed directly to hyaline cartilage-rich repair 4 months after transplantation [46].
Integration of the graft and host cartilage is essential for successful tissue replacement as it provides stable biological fixation and load distribution as well as adequate mechanotransduction necessary to maintain homeostasis [7]. However, cartilage-to-cartilage integration is exceedingly difficult to achieve because of the low metabolism of cartilage and the high density of the anti-adhesive ECM [52, 53]. Allogeneic cyiPSC-derived cartilage transplanted into chondral defects was well integrated with the host side cartilage, suggesting that stable biological fixation was achieved [46]. Human iPSC-derived cartilage has shown capacity for integration, and fibroblast growth factor (FGF) signals are involved in this integration [54]. RNA sequencing analysis showed a higher expression of FGF18 in the perichondrium-like membrane of human iPSC-derived cartilage. The addition of FGF18 promoted the integration of cartilages, whereas the addition of FGFR inhibitors inhibited it. These suggested that FGF18 secreted from the perichondrium-like membrane is involved in the integration of the human iPSC-derived cartilage [54].
Articular cartilage is a layered tissue that expresses proteoglycan 4 (PRG4), which functions as a lubricant in the superficial layer [55,56,57]. Post-transplantation, cyiPSC-derived cartilage showed high expression of PRG4 in the superficial layer. These results suggest that cyiPSC-derived cartilage acquires lubricating ability after transplantation and functions as articular cartilage [46]. One of the major differences between pre- and post-transplantation is that the graft is subjected to shear forces associated with knee joint motion in vivo after transplantation. Shear forces in vivo have been reported to stimulate PRG4 expression in chondrocytes via cyclic adenosine monophosphate (cAMP) signaling [58], and the expression of PRG4 in cyiPSC-derived cartilage after transplantation has been suggested to be associated with shear forces. Furthermore, salt-inducible kinase 3 (SIK3) has been suggested to be involved in PRG4 expression after transplantation [46]. SIKs inhibit nuclear translocation of cAMP response element binding protein (CREB)-regulated transcription coactivator (CRTC) by phosphorylating, thereby repressing CREB activation and gene transcription. Among the members of the SIK family, SIK3 functions primarily in chondrocytes. A recent study demonstrated that deletion of Sik3 further increased shear stress-induced Prg4 expression in mouse chondrocytes, suggesting that Sik3 negatively regulates Prg4 expression [46].
Suppression of immune response during allogeneic iPSC-derived cartilage transplantation into osteochondral defects
Allogeneic cyiPSC-derived cartilage transplantation for osteochondral defects showed that lymphocytes clustered around the graft 4 weeks postoperatively [45, 46]; however, the graft itself remained, suggesting a temporary immune response rather than complete immune rejection. Because chondrocytes express molecules that transduce inhibitory signals to T cells [9], these molecular mechanisms may contribute to the survival of allogeneic cyiPSC-derived cartilage in osteochondral defects.
The results of allogeneic iPSC-derived cartilage transplantation in a primate model with an immune system similar to that of humans suggested that chondral defects are a better indication for allogeneic iPSC-derived cartilage transplantation than osteochondral defects. Regarding osteochondral defects, an immune response has been observed in cases of MHC mismatches, which remains a long-term concern. One solution to prevent immune reactions is to minimize rejection by matching the HLA types of the donor and host cells. iPSC lines are established from donors whose major HLA types are homozygotes [59, 60]. These HLA-homo iPSC-derived products matched recipients with an identical set of HLA types in one allele. It is estimated that an iPSC line homozygous for the most frequent HLA types in the Japanese population would match 17% of the Japanese population. Therefore, preparing HLA-type homozygous iPSCs could minimize the influence of immune rejection [59, 60]. Another solution is to genome-edit iPS cells by using techniques, such as the CRISPR/Cas9 system [61]. It has been reported that B2M−/− cyiPSC-derived cartilage knocking out β2 microglobulin, which does not express MHC class I, was transplanted into osteochondral defects and showed immune response by NK cells [48]. This result is consistent with a previous finding that natural killer (NK) cells recognize and eliminate cells that fail to express MHC class I molecules [62]. Recent studies have established that human iPSCs lack HLA class I and II molecules and suppress NK cell attack by HLA-E transduction, CD47 overexpression, or PVR knockout [63,64,65,66,67]. Currently, the HLA genome-edited iPS stock is available for research use [67]. The use of cartilage tissue differentiated from HLA genome-edited iPS cells can suppress the immune response to allogeneic transplantation for osteochondral defects.
Safety and costs of allogeneic iPSC-derived cartilage transplantation
Transplantation of autologous iPSC-derived grafts is desirable in terms of avoiding immune reactions and minimizing the risk of spreading communicable viral infections; however, cost and manufacturing lead time is not practicable for commercialization. When the world’s first autologous transplant of iPSC-derived retinal pigment epithelial (RPE) cells was performed, the patient had to wait for more than 10 months from harvesting skin tissue to RPE sheet transplantation, which cost nearly 100 million yen [60, 68]. On the other hand, allogeneic transplantation approaches can reduce the cost and time of the iPSC manufacturing process. In the case of allogeneic iPSC-derived RPE sheet transplantation using cells from iPSC stock, the preoperative waiting period was approximately 1 month, and the total cost per patient was about one-fifth of autologous transplantation [60]. Similarly, the use of allogeneic iPSC stock reduces cost and time in iPSC-derived cartilage transplantation. One of the particular safety risks of iPSC-derived therapies, including iPSC-derived cartilage transplantation, is the concern of tumorigenicity because possible contamination of undifferentiated iPS cells can give rise to teratoma. The reprogramming process of the iPSC and the long culture time for cartilage differentiation could increase the potential for malignancy. The risk of tumorigenicity has been thoroughly evaluated in preclinical tests including in vitro expression analysis of iPS cell markers to detect contamination of iPS cells in the cartilage and in vivo tumorigenicity testing in which iPS cell-derived cartilage is transplanted into immunodeficient rats orthotopically for life-long observation. The evaluation of the tumorigenic risk of iPSC-derived cartilage using HeLa cells as the reference control suggested that the potential benefit of the therapy outweighs the risk of tumor formation and the clinical application of iPSC-derived cartilage in the knee joint was considered acceptable [69].
Conclusions
Chondrocyte and cartilage transplantation are treatment options for cartilage defects. Although chondrocytes have been shown to be hypoimmunogenic in vitro, allogeneic chondrocyte transplantation is controversial due to reports of immunoreactivity in vivo. In contrast, allogeneic cartilage transplantation, as performed in clinical practice, tends to show promising results in the short term; however, the evidence is insufficient. There are two types of cartilage defects, osteochondral and chondral defects, which may differ in their immune responses during allogeneic transplantation. The immune response to chondrocytes can be triggered by contact with the bone marrow. Although the effect of immune reactions on clinical outcomes in osteochondral defects has not yet been determined, immune reactions should be controlled to ensure good repair. Allogeneic iPSC-derived cartilage transplantation, a new therapeutic option, could be a good indication for chondral defects without an immune response. HLA type matching or iPSC lines in which HLA genes are edited can provide a solution to suppress the immune response in osteochondral defects (Fig. 2).

Solution to prevent immune response by iPSC-derived cartilage in osteochondral defects. HLA-matched or HLA genome-edited hypoimmunogenic iPSC lines can suppress the immune response in osteochondral defects
Availability of data and materials
All the original data are available upon request from the authors.
References
Makris EA, Gomoll AH, Malizos KN, Hu JC, Athanasiou KA. Repair and tissue engineering techniques for articular cartilage. Nat Rev Rheumatol. 2015;11(1):21–34.
Dekker TJ, Aman ZS, DePhillipo NN, Dickens JF, Anz AW, LaPrade RF. Chondral lesions of the knee: An evidence-based approach. J Bone Joint Surg Am. 2021;103(7):629–45.
Li J, Jiang H, Lv Z, Sun Z, Cheng C, Tan G, et al. Articular fibrocartilage-targeted therapy by microtubule stabilization. Sci Adv. 2022;8(46):eabn8420.
Kreuz PC, Steinwachs MR, Erggelet C, Krause SJ, Konrad G, Uhl M, et al. Results after microfracture of full-thickness chondral defects in different compartments in the knee. Osteoarthr Cartil. 2006;14(11):1119–25.
Harris JD, Siston RA, Pan X, Flanigan DC. Autologous chondrocyte implantation: a systematic review. J Bone Joint Surg Am. 2010;92(12):2220–33.
Brittberg M, Lindahl A, Nilsson A, Ohlsson C, Isaksson O, Peterson L. Treatment of deep cartilage defects in the knee with autologous chondrocyte transplantation. N Engl J Med. 1994;331(14):889–95.
Huey DJ, Hu JC, Athanasiou KA. Unlike bone, cartilage regeneration remains elusive. Science (New York, NY). 2012;338:917–21.
Roberts S, Menage J, Sandell LJ, Evans EH, Richardson JB. Immunohistochemical study of collagen types I and II and procollagen IIA in human cartilage repair tissue following autologous chondrocyte implantation. Knee. 2009;16(5):398–404.
Adkisson HD, Milliman C, Zhang X, Mauch K, Maziarz RT, Streeter PR. Immune evasion by neocartilage-derived chondrocytes: Implications for biologic repair of joint articular cartilage. Stem Cell Res. 2010;4(1):57–68.
Moskalewski S, Hyc A, Osiecka-Iwan A. Immune response by host after allogeneic chondrocyte transplant to the cartilage. Microsc Res Tech. 2002;58(1):3–13.
Zha K, Li X, Yang Z, Tian G, Sun Z, Sui X, et al. Heterogeneity of mesenchymal stem cells in cartilage regeneration: from characterization to application. NPJ Regen Med. 2021;6(1):14.
Elves MW, Zervas J. An investigation into the immunogenicity of various components of osteoarticular grafts. Br J Exp Pathol. 1974;55(4):344–51.
Farr J, Tabet SK, Margerrison E, Cole BJ. Clinical, radiographic, and histological outcomes after cartilage repair with particulated juvenile articular cartilage: a 2-year prospective study. Am J Sports Med. 2014;42(6):1417–25.
Wang T, Belkin NS, Burge AJ, Chang B, Pais M, Mahony G, et al. Patellofemoral cartilage lesions treated with particulated juvenile allograft cartilage: a prospective study with minimum 2-year clinical and magnetic resonance imaging outcomes. Arthroscopy. 2018;34(5):1498–505.
Dawkins BJ, Shubin Stein BE, Mintz DN, Fabricant PD, Gomoll AH, Strickland SM, et al. Patellofemoral joint cartilage restoration with particulated juvenile allograft in patients under 21 years old. Knee. 2022;36:120–9.
Yamanaka S. Pluripotent stem cell-based cell therapy-promise and challenges. Cell Stem Cell. 2020;27(4):523–31.
Takahashi K, Tanabe K, Ohnuki M, Narita M, Ichisaka T, Tomoda K, et al. Induction of pluripotent stem cells from adult human fibroblasts by defined factors. Cell. 2007;131(5):861–72.
Yamashita A, Tsumaki N. Recent progress of animal transplantation studies for treating articular cartilage damage using pluripotent stem cells. Dev Growth Differ. 2021;63(1):72–81.
Yamashita A, Morioka M, Yahara Y, Okada M, Kobayashi T, Kuriyama S, et al. Generation of scaffoldless hyaline cartilaginous tissue from human iPSCs. Stem Cell Rep. 2015;4(3):404–18.
Yamashita A, Morioka M, Kishi H, Kimura T, Yahara Y, Okada M, et al. Statin treatment rescues FGFR3 skeletal dysplasia phenotypes. Nature. 2014;513(7519):507–11.
Bentley G, Greer RB 3rd. Homotransplantation of isolated epiphyseal and articular cartilage chondrocytes into joint surfaces of rabbits. Nature. 1971;230(5293):385–8.
Kawabe N, Yoshinao M. The repair of full-thickness articular cartilage defects. Immune responses to reparative tissue formed by allogeneic growth plate chondrocyte implants. Clin Orthop Relat Res. 1991;268:279–93.
Hyc A, Malejczyk J, Osiecka A, Moskalewski S. Immunological response against allogeneic chondrocytes transplanted into joint surface defects in rats. Cell Transplant. 1997;6(2):119–24.
Osiecka-Iwan A, Hyc A, Moskalewski S. Immunosuppression and rejection of cartilage formed by allogeneic chondrocytes in rats. Cell Transplant. 1999;8(6):627–36.
Jobanputra P, Corrigall V, Kingsley G, Panayi G. Cellular responses to human chondrocytes: absence of allogeneic responses in the presence of HLA-DR and ICAM-1. Clin Exp Immunol. 1992;90(2):336–44.
Adkisson HD, Martin JA, Amendola RL, Milliman C, Mauch KA, Katwal AB, et al. The potential of human allogeneic juvenile chondrocytes for restoration of articular cartilage. Am J Sports Med. 2010;38(7):1324–33.
Huey DJ, Sanchez-Adams J, Willard VP, Athanasiou KA. Immunogenicity of bovine and leporine articular chondrocytes and meniscus cells. Tissue Eng Part A. 2012;18(5-6):568–75.
Bonasia DE, Martin JA, Marmotti A, Amendola RL, Buckwalter JA, Rossi R, et al. Cocultures of adult and juvenile chondrocytes compared with adult and juvenile chondral fragments: in vitro matrix production. Am J Sports Med. 2011;39(11):2355–61.
Chestan PJ, Smith AU. Homotransplantation of articular cartilage and isolated chondrocytes. An experimental study in rabbits. J Bone Joint Surg Br. 1968;50(1):184–97.
Aston JE, Bentley G. Repair of articular surfaces by allografts of articular and growth-plate cartilage. J Bone Joint Surg Br. 1986;68(1):29–35.
Wakitani S, Kimura T, Hirooka A, Ochi T, Yoneda M, Yasui N, et al. Repair of rabbit articular surfaces with allograft chondrocytes embedded in collagen gel. J Bone Joint Surg Br. 1989;71(1):74–80.
Freed LE, Grande DA, Lingbin Z, Emmanual J, Marquis JC, Langer R. Joint resurfacing using allograft chondrocytes and synthetic biodegradable polymer scaffolds. J Biomed Mater Res. 1994;28(8):891–9.
Frenkel SR, Toolan B, Menche D, Pitman MI, Pachence JM. Chondrocyte transplantation using a collagen bilayer matrix for cartilage repair. J Bone Joint Surg Br. 1997;79(5):831–6.
Rahfoth B, Weisser J, Sternkopf F, Aigner T, von der Mark K, Bräuer R. Transplantation of allograft chondrocytes embedded in agarose gel into cartilage defects of rabbits. Osteoarthr Cartil. 1998;6(1):50–65.
Kawamura S, Wakitani S, Kimura T, Maeda A, Caplan AI, Shino K, et al. Articular cartilage repair. Rabbit experiments with a collagen gel-biomatrix and chondrocytes cultured in it. Acta Orthop Scand. 1998;69(1):56–62.
Fragonas E, Valente M, Pozzi-Mucelli M, Toffanin R, Rizzo R, Silvestri F, et al. Articular cartilage repair in rabbits by using suspensions of allogenic chondrocytes in alginate. Biomaterials. 2000;21(8):795–801.
Perka C, Sittinger M, Schultz O, Spitzer RS, Schlenzka D, Burmester GR. Tissue engineered cartilage repair using cryopreserved and noncryopreserved chondrocytes. Clin Orthop Relat Res. 2000;378:245–54.
Moskalewski S, Osiecka-Iwan A, Hyc A, Jozwiak J. Mechanical barrier as a protection against rejection of allogeneic cartilage formed in joint surface defects in rats. Cell Transplant. 2000;9(3):349–57.
Romaniuk A, Malejczyk J, Kubicka U, Hyc A, Olszewski WL, Moskalewski S. Rejection of cartilage formed by transplanted allogeneic chondrocytes: evaluation with monoclonal antibodies. Transpl Immunol. 1995;3(3):251–7.
Bonasia DE, Marmotti A, Mattia S, Cosentino A, Spolaore S, Governale G, et al. The degree of chondral fragmentation affects extracellular matrix production in cartilage autograft implantation: An in vitro study. Arthroscopy. 2015;31(12):2335–41.
Matsushita R, Nakasa T, Ishikawa M, Tsuyuguchi Y, Matsubara N, Miyaki S, et al. Repair of an osteochondral defect with minced cartilage embedded in atelocollagen gel: A rabbit model. Am J Sports Med. 2019;47(9):2216–24.
Bonasia DE, Martin JA, Marmotti A, Kurriger GL, Lehman AD, Rossi R, et al. The use of autologous adult, allogenic juvenile, and combined juvenile-adult cartilage fragments for the repair of chondral defects. Knee Surg Sports Traumatol Arthrosc. 2016;24(12):3988–96.
Ao Y, Li Z, You Q, Zhang C, Yang L, Duan X. The use of particulated juvenile allograft cartilage for the repair of porcine articular cartilage defects. Am J Sports Med. 2019;47(10):2308–15.
Zhang C, Ao Y, Cao J, Yang L, Duan X. Donor cell fate in particulated juvenile allograft cartilage for the repair of articular cartilage defects. Am J Sports Med. 2020;48(13):3224–32.
Okutani Y, Abe K, Yamashita A, Morioka M, Matsuda S, Tsumaki N. Generation of monkey induced pluripotent stem cell-derived cartilage lacking major histocompatibility complex class I molecules on the cell surface. Tissue Eng Part A. 2022;28(1-2):94–106.
Abe K, Yamashita A, Morioka M, Horike N, Takei Y, Koyamatsu S, et al. Engraftment of allogeneic iPS cell-derived cartilage organoid in a primate model of articular cartilage defect. Nat Commun. 2023;14(1):804.
Yanke AB, Tilton AK, Wetters NG, Merkow DB, Cole BJ. DeNovo NT particulated juvenile cartilage implant. Sports Med Arthrosc Rev. 2015;23(3):125–9.
Tompkins M, Hamann JC, Diduch DR, Bonner KF, Hart JM, Gwathmey FW, et al. Preliminary results of a novel single-stage cartilage restoration technique: particulated juvenile articular cartilage allograft for chondral defects of the patella. Arthroscopy. 2013;29(10):1661–70.
Buckwalter JA, Bowman GN, Albright JP, Wolf BR, Bollier M. Clinical outcomes of patellar chondral lesions treated with juvenile particulated cartilage allografts. Iowa Orthop J. 2014;34:44–9.
Grawe B, Burge A, Nguyen J, Strickland S, Warren R, Rodeo S, et al. Cartilage regeneration in full-thickness patellar chondral defects treated with particulated juvenile articular allograft cartilage: An MRI analysis. Cartilage. 2017;8(4):374–83.
Kimura T, Yamashita A, Ozono K, Tsumaki N. Limited immunogenicity of human induced pluripotent stem cell-derived cartilages. Tissue Eng Part A. 2016;22(23-24):1367–75.
Khan IM, Gilbert SJ, Singhrao SK, Duance VC, Archer CW. Cartilage integration: evaluation of the reasons for failure of integration during cartilage repair. A review. Eur Cell Mater. 2008;16:26–39.
van de Breevaart BJ, In der Maur CD, Bos PK, Feenstra L, Verhaar JA, Weinans H, et al. Improved cartilage integration and interfacial strength after enzymatic treatment in a cartilage transplantation model. Arthritis Res Ther. 2004;6(5):R469–76.
Chen X, Yamashita A, Morioka M, Senba T, Kamatani T, Watanabe A, et al. Integration capacity of human induced pluripotent stem cell-derived cartilage. Tissue Eng Part A. 2019;25(5-6):437–45.
Swann DA, Silver FH, Slayter HS, Stafford W, Shore E. The molecular structure and lubricating activity of lubricin isolated from bovine and human synovial fluids. Biochem J. 1985;225(1):195–201.
Rhee DK, Marcelino J, Baker M, Gong Y, Smits P, Lefebvre V, et al. The secreted glycoprotein lubricin protects cartilage surfaces and inhibits synovial cell overgrowth. J Clin Invest. 2005;115(3):622–31.
Ruan MZ, Erez A, Guse K, Dawson B, Bertin T, Chen Y, et al. Proteoglycan 4 expression protects against the development of osteoarthritis. Sci Transl Med. 2013;5(176):176ra34.
Ogawa H, Kozhemyakina E, Hung HH, Grodzinsky AJ, Lassar AB. Mechanical motion promotes expression of Prg4 in articular cartilage via multiple CREB-dependent, fluid flow shear stress-induced signaling pathways. Genes Dev. 2014;28(2):127–39.
Okita K, Matsumura Y, Sato Y, Okada A, Morizane A, Okamoto S, et al. A more efficient method to generate integration-free human iPS cells. Nat Methods. 2011;8(5):409–12.
Umekage M, Sato Y, Takasu N. Overview: an iPS cell stock at CiRA. Inflamm Regen. 2019;39:17.
Ran FA, Hsu PD, Wright J, Agarwala V, Scott DA, Zhang F. Genome engineering using the CRISPR-Cas9 system. Nat Protoc. 2013;8(11):2281–308.
Ljunggren HG, Kärre K. In search of the 'missing self': MHC molecules and NK cell recognition. Immunol Today. 1990;11(7):237–44.
Han X, Wang M, Duan S, Franco PJ, Kenty JH, Hedrick P, et al. Generation of hypoimmunogenic human pluripotent stem cells. Proc Natl Acad Sci U S A. 2019;116(21):10441–6.
Deuse T, Hu X, Gravina A, Wang D, Tediashvili G, De C, et al. Hypoimmunogenic derivatives of induced pluripotent stem cells evade immune rejection in fully immunocompetent allogeneic recipients. Nat Biotechnol. 2019;37(3):252–8.
Ye Q, Sung TC, Yang JM, Ling QD, He Y, Higuchi A. Generation of universal and hypoimmunogenic human pluripotent stem cells. Cell Prolif. 2020;53(12):e12946.
Wang B, Iriguchi S, Waseda M, Ueda N, Ueda T, Xu H, et al. Generation of hypoimmunogenic T cells from genetically engineered allogeneic human induced pluripotent stem cells. Nat Biomed Eng. 2021;5(5):429–40.
Kitano Y, Nishimura S, Kato TM, Ueda A, Takigawa K, Umekage M, et al. Generation of hypoimmunogenic induced pluripotent stem cells by CRISPR-Cas9 system and detailed evaluation for clinical application. Mol Ther Methods Clin Dev. 2022;26:15–25.
Mandai M, Watanabe A, Kurimoto Y, Hirami Y, Morinaga C, Daimon T, et al. Autologous Induced Stem-Cell-Derived Retinal Cells for Macular Degeneration. N Engl J Med. 2017;376(11):1038–46.
Takei Y, Morioka M, Yamashita A, Kobayashi T, Shima N, Tsumaki N. Quality assessment tests for tumorigenicity of human iPS cell-derived cartilage. Sci Rep. 2020;10(1):12794.
Acknowledgements
The authors thank Yuki Okutani and Shuichi Matsuda for their significant contributions to this study, and Takahiro Negishi for reading the article.
Funding
This study was supported by KAKENHI from the Japan Society for the Promotion of Science (JSPS) under Grant Number 23H03029 and by AMED under grant numbers JP23ym0126127 and JP23bm1223014.
Author information
Authors and Affiliations
Contributions
All authors contributed to the writing of the manuscript. All authors have consented to the publication of this manuscript. All the authors have read and approved the final version of the manuscript.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
All experiments were approved by the Institutional Review Board, the Institutional Animal Committee (as appropriate), and the Institutional Biosafety Committee of Kyoto University.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Abe, K., Tsumaki, N. Regeneration of joint surface defects by transplantation of allogeneic cartilage: application of iPS cell-derived cartilage and immunogenicity. Inflamm Regener 43, 56 (2023). https://doi.org/10.1186/s41232-023-00307-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s41232-023-00307-0