
Abstract
Mechanical ventilation can improve hypoxemia, but can also cause the so-called ventilator-induced lung injury (VILI). Polyinosinic:polycytidylic acid (poly(I:C)), an analogue of natural double-strand RNA virus, can induce lung inflammation. The purpose of this study was to determine whether moderate tidal volume mechanical ventilation (MTV) augments poly(I:C)-induced lung injury, and if so, the mechanism responsible for it. Two µg/g poly(I:C) were instilled intratracheally in C57BL/6J wide type (WT) mice. They were then randomized to MTV (10 ml/kg tidal volume) or spontaneous breathing. Lung tissues and bronchoalveolar lavage fluid (BALF) were collected 4 h later for various measurements. Our results showed that MTV did not cause significant injury in normal lungs, but augmented poly(I:C)-induced lung injury. The expression level of WNT-induced secreted protein 1 (WISP1) was consistent with lung injury, and the amplification of lung injury by MTV could be alleviated by anti-WISP1 antibody treatment. MTV further increased poly(I:C)-induced integrin β3 expression in the lung. We performed coimmunoprecipitation, which showed there was an interaction between WISP1 and β3. WISP1 significantly increased poly(I:C)-induced TNF-α production in macrophages isolated from WT mice, but not in macrophages isolated from β3 knockout mice. Cotreatment with WISP1 and poly(I:C) markedly increased the phosphorylation of extracellular signal-related kinase (ERK) in macrophages. Pretreating macrophages with an ERK inhibitor, U0126, dose-dependently antagonized the synergistic effect of WISP1 on poly(I:C)-induced TNF-α release. In conclusion, MTV exaggerates poly(I:C)-induced lung injury in a WISP1- and integrin β3-dependent manner, involving, at least in part, the activation of the ERK pathway. The WISP1-integrin β3 pathway could be a novel therapeutic target.
Similar content being viewed by others
Introduction
Mechanical ventilation can protect injured lungs and improve hypoxemia, but can also cause ventilator-induced lung injury (VILI) because of different overlapping interactions, including: lung stress due to increased transpulmonary pressure; overdistension caused by high tidal volume; cyclic closing and opening of peripheral airways during tidal breath; and local and systemic release of lung-borne inflammatory mediators (1).
Double-stranded RNA (dsRNA) can be produced by many viruses during their replicative cycles (2). Polyinosinic-polycytidylic acid (poly(I:C)), a synthetic analog of dsRNA, initiates cascades of phosphorylation and transcriptional activation events associated with innate immunity. Poly(I:C) can induce loss of epithelial integrity, as well as the production of characteristic inflammatory cytokines and chemokines. It is a critical component in the modulation of infection-associated inflammatory diseases (3).
Moderate tidal volume mechanical ventilation (MTV) has been reported to function as a cofactor in the initiation of acute lung injury (ALI) by amplifying the inflammatory response induced by pathogens (4,5). A study by Chun CD et al. (6) found that mechanical ventilation at 10 ml/kg for 6 h in mice augmented poly(I:C)-induced cytokine release, polymorphonuclear (PMN) counts, 70-kD fluorescein isothiocyanate dextran concentration, and IgM level in bronchoalveolar lavage fluid (BALF). The amplification of the inflammatory response by MTV is related to the production of endogenous ligands, which are recognized by MyD88-dependent transmembrane receptors.
Our previous study has demonstrated that WNT-induced secreted protein 1 (WISP1), identified through a genome-wide approach, acts as an adjuvant adaptor molecule that contributes to VILI in an autocrine and paracrine fashion (7). WISP1 (Wnt1-inducible signaling pathway protein 1, also called CCN4 or Elm1), is a cysteine-rich, matricellular protein, allocated to the CCN protein family (8). Members of the CCN family are crucial for embryonic development and have important roles in inflammation, wound healing and injury repair in adulthood (9). Konigshoff et al. (10) also demonstrated that WISP1 is a potential therapeutic target for pulmonary fibrosis. Heise and colleagues (11) reported that WISP1 was induced by mechanical stretch in mouse alveolar type II cells, and the increased WISP1 expression level could be reduced in cells treated with a WISP1 antibody. These studies indicate that WISP1 contributes to the process of lung injury.
Integrins are transmembrane adhesion receptors that provide essential links between the extracellular environment and intracellular signaling pathways (12). The members of the integrin family play an important role in ALI through regulating lung inflammatory cytokines release and alveolar capillary permeability (13). Results from Wang B et al. (14) showed that inhibiting integrin ανβ3 reduced inflammation and attenuated VILI in rats. However, whether WISP1 and integrin β3 are involved in the amplification of MTV in poly(I:C)-induced lung injury remained unknown. We therefore hypothesize that MTV exacerbates poly(I:C)-induced lung injury via a WISP1-integrin β3-dependent pathway. The purpose of this study was to determine whether MTV augments poly(I:C)-induced lung injury, and if so, the role of the WISP1-integrin β3 signaling in this process.
Materials and Methods
Experimental Animals
Male pathogen-free C57BL/6J wide type (WT) mice were purchased from the Laboratory Animal Research Center of Shanghai. Integrin β3 knockout mice were purchased from Jackson Laboratory and bred at the core facility at the University of Pittsburgh. Phenotype was confirmed by PCR before each experiment. A total of 160 male C57 BL/6J mice (8∼12 wks) were included in the experiment. WT mice (120 were randomized to various groups (n = 30 each group, half of the mice were used to determine alveolar-capillary permeability, half were used for other parameters): spontaneous breath (control); spontaneous breath with poly(I:C) treatment (tlrl-pic, Invivogen); moderate tidal volume mechanical ventilated group (MTV); mechanical ventilation with poly(I:C) pretreatment group (poly(I:C) + MTV). In the poly(I:C) + MTV group, another 20 mice were randomized to the anti-WISP1 antibody group or IgG group. For in vitro studies, fifteen C57 BL/6J mice and five integrin β3 knockout mice were used for isolation of peritoneal macrophages. The mice were housed in a temperature-controlled room on a 12 h light/dark cycle and fed on a standard Purina mouse chow diet. Animal experimentation was carried out in accordance with the Guide for the Care and Use of Laboratory Animals, (15). The Institutional Animal Care and Use Committee (IACUC) of Tongji University (TJMED-013-062) has approved all procedures.
Experimental Protocol
All mice were anesthetized by ketamine (45 mg/kg) and xylazine (10 mg/kg) intraperitoneally (i.p.). For intubation, mice were placed on a procedure bench equipped with a closed-loop system to control body temperature. The trachea was intubated with a 3-cm length of tube. Poly(I:C) was dissolved into 10 mg/mL with saline, then 2 µg/g per body weight of poly(I:C) were instilled through the catheter, mice were then randomized to MTV (10 mL/kg tidal volume, positive end-expiratory pressure of 0 cm H2O, and FiO2 0.21, 140 breaths/min), for 4 h with a volume-cycled mouse ventilator (Model Inspira, Harvard Apparatus) or spontaneous breath.
All animals were studied over 4 h of mechanical ventilation. The right lung was ligated and was collected for hematoxylin and eosin (H&E) stain, immunohistochemistry (IHC), RNA extraction, protein extraction and lung wet:dry weight ratios. The left lung was lavaged to determine cytokine levels, total protein levels and cell counts. A total of 5 ml isotonic sterile saline was used to wash the left lung for every mouse. Bronchoalveolar lavage fluid (BALF) was centrifuged immediately (4°C, 2000g for 15 min) for harvesting of the cells and the supernatant. Total protein levels in the supernatant were measured using bicinchoninic acid (BCA) and cytokine levels were assayed by enzyme-linked immunosorbent assay (ELISA). The deposits were collected for PMN counts by Wright-Giemsa-stained slides.
WISP1 Antibody Treatment
For the WISP1-blockade study, 20 mice were randomized to WISP1 antibody group or IgG group, and received 5 mg/kg body weight of anti-WISP1 neutralizing antibody (MAB1680, R&D Systems) or nonspecific isotype-specific antibody IgG (MAB002, R&D Systems) i.p., respectively. One hour later, animals were subjected to poly(I:C) + MTV as described previously. We use a pilot experiment to determine the dose of antibody.
Measurement of Alveolar-Capillary Permeability
The lung barrier function of the mice was assayed with Evans blue albumin (EBA). Phosphate-buffered saline (PBS; Sigma-Aldrich) without Ca2+/Mg2+ was used to dissolve Evans blue (0.5% EB, Sigma-Aldrich) and bovine serum albumin (Sigma-Aldrich). One hour before the mice were euthanized, EBA (2% in saline, 2 mL/kg) was intravenously administered. Blood was removed by cardiac puncture, plasma was immediately separated by centrifugation (30 min at 5000g). Lungs were perfused with 1 mL PBS (5 mmol EDTA), then removed and snap frozen in liquid nitrogen. Samples were then homogenized in 2 mL PBS, then incubated in 2 mL formamide at 60°C for 18 h. Optical density was determined at 620 and 750 nm. A standard curve normalized to plasma EB level was used to calculated the extravasated EB concentration. (16,17)
Lung Wet:Dry Weight Ratio
Lung edema was estimated by comparing wet:dry weight ratios. The wet weight of the middle lobe of the right lung was determined immediately after removal. The lobe was then placed in an oven at 65°C for 48 h, after which the dry weight was obtained.
Cytokines Assay in BALF
Concentrations of TNF-α and IL-6 in BALF were determined by mouse TNF-α and IL-6 ELISA kits (R&D Systems), respectively, according to the manufacturer’s protocol. In brief, assay diluent RD-14 was added to the center of each well. Sample, control and standards were inserted into the wells via pipette then mixed gently and incubated at room temperature for 2 h. Each well was washed five times with wash buffer, then TNF-α and IL-6 conjugate were added and the wells were incubated for 2 h at room temperature. The aspiration/wash steps were repeated, then a substrate solution was added to the wells, after which they were incubated for 30 min at room temperature and protected from light. Next, a stop solution was inserted into each well and a microplate reader set to 450 nm using a correction to determine the optical density of each well.
Histological Examination
The right superior lobe lung of each animal was examined histologically. A light microscope was used for acquisition of images. To grade the degree of lung injury, a score system based on the following pathologic features was used: edema, neutrophil margination and tissue infiltration; hyperemia and congestion; intraalveolar hemorrhage and debris; and cellular hyperplasia. Each feature was graded as absent, mild, moderate, or severe, with a score of 0∼3 (18).
Western Blot Analysis
The right inferior lobe of the lung was removed rapidly. NP-40 buffer containing protease inhibitors (Roche) and phenylmethylsulfonyl fluoride (PMSF) were added to lyses tissues. Samples from each group were evenly divided between gels and transferred to nitrocellulose membrane. Membranes were blocked with 5% skim milk, incubated with mouse primary antibody against WISP1 (ab178547, Abcam), Integrin β3 (ab75872, Abcam), phosphorylation extracellular signal-related kinase (pERK) (ab 115617, Abcam) and β-actin (ab8226, Abcam) subsequently with a secondary antibody (Licor Biosciences).
Reverse Transcription and Polymerase Chain Reaction (PCR)
The lung was removed rapidly and placed in TRIzol (Invitrogen) total RNA isolation reagent according to the manufacturer’s instructions (Sigma-Aldrich). Total RNA was reverse-transcribed in a 20 µL reaction using the OmniScript RT kit (TaKaRa Bio Inc.). The Premix Taq Version 2.0 (TaKaRa Bio Inc.) was used to amplify the reverse transcription product. PCR products were confirmed on 2% agarose gel and visualized by SYBR green staining, Image Lab software (Bio-Rad) was used to quantitate the expression of mRNA. Following is the list of primers used: wisp1, 137 bp forward 5′-CAGCA CCACT AGAGG AAAGG A-3′, reverse 5′-CTGGG CACAT ATCTT ACAGC ATT-3′. Integrin β3, 107 bp, forward 5′-CCACA CGAGG CGTGA ACTC-3′, reverse 5′-CTTCA GGTTA CATCG GGGTG A-3′. β-actin, 154 bp, forward 5′-GGCTG TATTC CCCTC CATCG-3′, reverse 5′-CCAGT TGGTA ACAAT GCCAT GT-3′.
Coimmunoprecipitation (Co-IP)
To explore the interaction between WISP1 and integrin β3, Co-IP was performed. 500 µl of lysis buffer was added to lung homogenates and the samples were placed on ice for 30 min. Lysates were spun at 5000g at 4°C for 10 min. The supernatants were collected as input controls, or incubated with protein A/G beads overnight in the presence of anti-WISP1 (sc-8866, Santa Cruz), or anti-β3 (sc-6627, Santa Cruz) antibodies, respectively. After incubation, samples were washed with 500 µl of lysis buffer five times at 4°C. Supernatants were collected and 6 × sodium dodecyl sulfate (SDS) loading buffer was added into each sample. Before separating protein by gel electrophoresis using a 10% SDS-PAGE gels, the samples were boiled at 100°C for 5 min. The samples were then transferred to nitrocellulose membranes and blocked with 5% nonfat milk in tris buffered saline containing Tween-20 (TBST) at room temperature for 1 h. After blocking, membranes were incubated with either a rabbit monoclonal primary antibody against integrin β3 (ab75872, Abcam) or rabbit polyclonal primary antibody against WISP1 (ab178547, Abcam) overnight at 4°C. The membranes were washed three times with TBST and incubated with secondary antibody for 1 h at room temperature. The signals on the membrane were determined using an Odyssey image analysis system (Licor Biosciences) after extensive washing.
Peritoneal Macrophage Isolation
Macrophages were isolated from C57 BL/6J and integrin β3 knockout mice as described by Takeda et al. (19). Briefly, three days before euthanization, mice were injected i.p. with 2 mL 4% thioglycollate medium. Peritoneal macrophages were isolated by injection of 10 mL of RPMI 1640 medium (Sigma-Aldrich) into the peritoneal cavity and plated at a density of 5 × 105 cells/mL in RPMI 1640 medium containing 10% FBS, 50 U/mL penicillin G sodium and streptomycin sulfate. Overnight, the non-adherent cells were washed away with PBS and incubated in fresh medium. On the next day, the adherent macrophages were incubated in serum-free medium RPMI1640 at 37°C for 2 h and then treated with poly(I:C) in the presence or absence of recombinant WISP1 (1680-WS-050, R&D Systems).
Statistical Analysis
Statistical analyses were carried out using the GraphPad Prism 5 program. Data were presented as the means ± SEM and analyzed by either a one-way analysis of variance (ANOVA) and Student-Newman-Keuls test or independent samples group t test. P < 0.05 was considered statistically significant.
All supplementary materials are available online at www.molmed.org .
Results
MTV Exaggerated Poly(I:C)-induced Lung injury
To determine if mechanical ventilation would augment responses in the lung induced by exposure to dsRNA, mice received intratracheal poly(I:C) (2 µg/g) followed by MTV (10 ml/kg) for 4 h. The lung wet:dry weight ratio was used to assess lung edema. There were no statistically significant differences in lung EBA permeability (Figure 1A) or wet:dry weights ratios (Figure 1B) between the control group and the MTV alone group. Similarly, MTV alone had no significant effects on the total protein level (Figure 1C), TNF-α level (Figure 1D), IL-6 level (Figure 1E) and PMNs number (Figure 1F) in BALF. Although administration of poly(I:C) into the trachea alone significantly increased the lung wet:dry weight ratio, the total protein level, TNF-α level and IL-6 level in BALF, the addition of MTV further significantly increased these parameters. EBA permeability and PMN numbers in BALF in the poly(I:C) + MTV group increased significantly compared with those in poly(I:C) group (Figure 1).

Alterations in (A) EBA permeability, (B) wet:dry ratio of lungs, and (C) levels of total protein, (D) TNF-α and (E) IL-6, as well as (F) PMNs numbers in BALF at 4 h after poly(I:C) and MTV treatment. Data are presented as means ± SEM (n = 15) and compared by one-way ANOVA and Student-Newman-Keuls test. For comparing the poly(I:C) group with the untreated control group #P < 0.05, ##P < 0.01, ###P < 0.001. For comparing the poly(I:C) + MTV group with the poly(I:C) group **P < 0.01, ***P < 0.001.
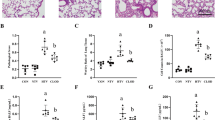
H&E staining of lung tissues revealed normal lung histology in control and MTV alone groups. Moderate inflammatory changes were observed in lung tissues from poly(I:C)-treated mice. In the poly(I:C) + MTV group, the lung injury score was markedly increased (Figure 2).

Morphologic alterations of the lungs as determined by photomicrography. Control: photomicrograph of a pulmonary section from a control mouse. MTV: photomicrograph of a lung section from a normal mouse treated with MTV. Poly(I:C): photomicrograph of a lung section from a poly(I:C) challenged mouse. Poly(I:C) + MTV: photomicrograph of a lung section from a poly(I:C)-challenged mouse treated with MTV. Original magnification: 400×. A total histopathologic score of lung injury was calculated for each animal. Data are presented as means ± SEM (n = 5) and compared by one-way ANOVA and Student-Newman-Keuls test. For comparing the poly(I:C) group with the untreated control group ***P < 0.001. For comparing the poly(I:C) + MTV group with the poly(I:C) group ***P < 0.01.
Taken together, these results indicated that mechanical ventilation with a tidal volume of 10 ml/kg for 4 h alone had no significant effects on normal lung, but amplified preexisting lung injury induced by intratracheal poly(I:C).
WISP1 Expression in the Lung
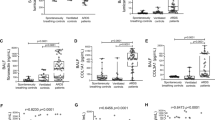
WISP1 is known to be induced by mechanical ventilation and contributes to the inflammatory response during VILI induced by high tidal volume. (7) As shown in Figures 3A–3B, MTV alone did not alter WISP1 expression in the lung. In the poly(I:C) alone group, wisp1 mRNA level in the lung was increased significantly; however, WISP1 protein levels were slightly elevated but not to a significant level compared to control mice. In contrast, the combination of intratracheal poly(I:C) followed by MTV resulted in a marked increase in lung WISP1 expression at mRNA and protein levels.

Alterations in pulmonary levels of wisp1 mRNA expressed as (A) the ratio to the housekeeping gene β-actin and protein and (B) the ratio to the housekeeping protein β-actin at 4 h after poly(I:C) and MTV treatment. Data are presented as means ± SEM (n = 15) and compared by one-way ANOVA and Student-Newman-Keuls test. For comparing the poly(I:C) group with the untreated control group **P < 0.01. For comparing the poly(I:C) + MTV group with the poly(I:C) group *P < 0.05, **P < 0.01.
Anti-WISP1 Antibody Attenuates Poly(I:C) + MTV-induced Lung injury
To determine the role of WISP1 in poly(I:C) + MTV-induced lung injury, an anti-WISP1 antibody or control IgG was injected i.p. 1 h before mechanical ventilation. As shown in Figures 4A–4F, administration of the WISP1 antibody prevented the increase in EBA permeability, the lung wet:dry weight ratio, as well as the increase in total protein and TNF-α, IL-6 levels and PMN counts in BALF induced by poly(I:C) + MTV.

Effects of anti-WISP1 treatment on (A) EBA permeability, (B) wet:dry ratio of lungs, and (C) levels of total protein, (D) TNF-α and (E) IL-6, as well as (F) PMNs numbers in BALF at 4 h after poly(I:C) and MTV treatment. Animals received the anti-WISP1 blocking antibody or control IgG i.p. 1 h before mechanical ventilation. Data are presented as mean ± SEM (n = 10) and compared by paired t test. For comparing the anti-WISP1 group with the IgG group *P < 0.05, **P < 0.01.
Taken together, these finding suggest that the rapid increase in WISP1 induced by poly(I:C) + MTV contributed to the lung injury and inflammation in this model.
Integrin β3 Expression in the Lung Is Increased by the Combination of poly(I:C) + MTV
Similar to WISP1 expression, MTV alone had no effects on integrin β3 expression in the lung (Figure 5). Although poly(I:C) alone upregulated integrin β3 mRNA expression in the lung, it did not significantly alter integrin β3 protein levels. Intratracheal poly(I:C) followed by MTV further upregulated integrin β3 mRNA and protein levels in the lung.

Alterations in pulmonary levels of integrin β3 mRNA expressed as (A) the ratio to the housekeeping gene β-actin and protein and (B) the ratio to the housekeeping protein β-actin at 4 h after poly(I:C) and MTV treatment. Data are presented as means ± SEM (n = 15) and compared by one-way ANOVA and Student-Newman-Keuls test. For comparing the poly(I:C) group and MTV group with the untreated control group: ###P < 0.001. And for comparing the poly(I:C) + MTV group with the poly(I:C) and MTV groups: ***P < 0.001.
Interaction Between WiSP1 and Integrin β3 in Poly(I:C) + MTV Mice
Co-IP was performed to determine if WISP1 interacts directly with integrin β3 following poly(I:C) + MTV treatment. Lung tissue lysates isolated at the end of MTV were immunoprecipitated with anti-WISP1 (Figure 6A) or anti-β3 (Figure 6B) antibodies and then analyzed for β3 or WISP1 by Western blot. Figure 6 shows that WISP1 interacts with integrin β3 at baseline and the interaction was enhanced in the poly(I:C) + MTV group.

Effects of poly(I:C) + MTV on WISP1/integrin β3 interaction. Lung tissues were harvested from C57BL/6J mice in the control and poly(I:C) + MTV groups. Total lysates were immunoprecipitated with an anti-WISP1 antibody or an anti-β3 antibody before immunoblot was performed. (A) Glycosylated form of integrin β3 (110 kD) was detected with an anti-integrin β3 antibody (IB: integrin β3) either in the whole lung tissue lysates (Total Lysates) or after immunoprecipitation with an anti-WISP1 antibody (IP: WISP1). (B) WISP1 was detected with an anti-WISP1 antibody (IB: WISP1) either in the whole lung tissue lysates (Total Lysates) or after immunoprecipitation with an anti-integrin β3 antibody (IP: integrin β3). Antibody species matched serum (Goat IgG) was used as a negative control for coprecipitation experiments. WISP1 and integrin β3 interact was enhanced in the poly(I:C) + MTV group as compared with the control group.
The Synergetic Effect of WiSP1 and Poly(I:C) on Macrophages
Peritoneal macrophages isolated from C57 BL/6J mice were treated with poly(I:C) in the presence or absence of recombinant WISP1 (1680-WS-050, R&D Systems) in serum-free medium for up to 24 h. TNF-α levels in the medium were measured by ELISA. We found that rWISP1 alone had no effects on TNF-α release, but significant enhanced poly(I:C)-induced TNF-α secretion. As shown in Figure 7A, the strongest synergistic effect was observed at 5 µg/mL poly(I:C), with no further increase in TNF-α levels at 10 µg/mL rWISP1. A time course study showed that WISPl-enhanced TNF-α production became significant by 16 h after macrophage exposure to rWISP1 + poly(I:C) (Figure 7B). These results confirmed that WISP1 can regulate poly(I:C)-induced inflammatory cytokine release in macrophages.

Effects of WISP1 on poly(I:C)-induced TNF-α production in macrophages. (A) Peritoneal macrophage isolated from C57BL/6J mice were treated with WISP1 (10 µg/mL) in the presence or absence of various concentrations (0–10 µg/mL) of poly(I:C) in serum-free medium for 16 h. TNF-α levels in the supernatant were measured by ELISA. (B) Peritoneal macrophage isolated from C57BL/6J mice were treated with WISP1(10 µg/mL) in the presence or absence of poly(I:C) (5 µg/mL) in serum-free medium for up to 24 h. TNF-α levels in the supernatant were measured by ELISA at 0 h, 4 h, 8 h, 12 h, 16 h, 20 h and 24 h after stimulation. Data are presented as mean ± SEM (n = 5) and compared by two-way ANOVA. For comparing the poly(I:C) + WISP1 group with the poly(I:C) alone group *P < 0.05, ***P < 0.001.
WIPS1 Enhances Poly(I:C)-induced TNF-α Release Dependent on integrin β3
To determine whether integrin β3 plays a role in WISP1 and poly(I:C)-induced TNF-α excretion in macrophages, we isolated peritoneal macrophage from C57BL/6J WT mice as well as β3 knockout mice. The cells were treated with poly(I:C) in the presence or absence of WISP1 for 16 h. WISP1 alone had no effects on TNF-α production in macrophages. As shown in Figure 8, WISP1 significantly increased poly(I:C)-induced TNF-α production in macrophages isolated from WT mice but not in macrophages isolated from β3 knockout mice, indicating that the synergistic effect of WISP1 and poly(I:C) on TNF-α production is dependent on integrin β3.

Effects of integrin β3 knockout on WISP1 plus poly(I:C)-induced TNF-α production in macrophages. Peritoneal macrophages isolated from C57BL/6J and integrin β3 knockout mice were treated with WISP1 (10 µg/mL) and poly(I:C) (5 µg/mL) for 16 h in serum free medium. TNF-α levels in the supernatant were measured by ELISA. Data are presented as mean ± SEM (n = 5) and compared by two-way ANOVA. ***P < 0.001 C57BL/6J versus integrin β3 knockout after WISP1 plus poly(I:C) treatment.
ERK Activation is Linked to the Synergetic Effect of WISP1 and Poly(I:C) on TNF-α Secretion
WISP1-induced signaling through integrins has been shown to involve the ERK (20). We therefore hypothesized that the ERK pathway may mediate the synergistic effect of WISP1 on poly(I:C)-induced TNF-α release. To test this idea, the effects of poly(I:C) and/or WISP1 on phosphorylation of ERK was examined in peritoneal macrophages isolated from C57 BL/6J WT mice. As shown in Figure 9A, cotreatment with WISP1 and poly(I:C) markedly increased the phosphorylation of ERK at 60 min. To establish the role of ERK in WISP1 plus poly(I:C)-induced TNF-α release, we pretreated macrophages with a specific ERK inhibitor, U0126, and measured TNF-α release in response to WISP1 and poly(I:C) treatment. As shown in Figure 9B, the ERK inhibitor dose-dependently suppressed WISP1 + poly(I:C)-induced TNF-α release. Thus, it appears that WISP1 and poly(I:C) act through the ERK-dependent signaling pathway to induce TNF-α secretion in macrophages.

Effects of WISP1 and poly(I:C) on ERK phosphorylation in macrophages (A) Peritoneal macrophages isolated from C57BL/6J mice were treated with WISP1 (10 µg/mL) or poly(I:C) in serum free medium for 15, 30 and 60 min. ERK phosphorylation was detected by Western blot. Effects of an ERK inhibitor, U0126, on WISP1 and poly(I:C)-induced TNF-α release in macrophages. (B) Peritoneal macrophage isolated from C57BL/6J mice were treated with WISP1 (10 µg/mL) and poly(I:C) (5 µg/mL) in the presence or absence of various concentrations (0–1.0 µmol/L) of U0126 in serum free medium for 16 h. TNF-α levels in the supernatant were measured by ELISA. Data are presented as means ± SEM (n = 5) and compared by two-way ANOVA. For comparing the poly(I:C) + WISP1 group pretreated with U0126 0.5 µmol/L with poly(I:C) + WISP1 untreated with U0126 group ***P < 0.001.
Discussion
WISP1 is downstream of Wnt/β-catenin signaling. Its expression is regulated by multiple signaling pathways which are activated by biochemical cues such as mechanical stretch. It remains unknown exactly how WISP1 modulates cellular function and through which receptors. Several studies have confirmed the functional interaction of WISP1 with integrins (21,22). Integrin β3 is expressed on endothelial cells, smooth muscle cells, fibroblasts, leukocytes and platelets. It binds to multiple ligands in the extracellular matrix such as WISP1, through interaction with the Arg-Gly-Asp motif (23). In this study, we show that integrin β3 is required for the amplification of inflammation and lung injury mediated through WISP1 when MTV is combined with intratracheal poly(I:C) treatment in mice.
The main findings of this study are: 1) MTV augmented poly(I:C)-induced ALI in mice; 2) this augmented response is associated with the broad modulation of WISP1 and β3 in the lung, partly through the ERK pathway; 3) WISP1 and poly(I:C)-induced inflammatory cytokine release in cultured macrophages depends on integrin β3.
Extracellular dsRNA, a potent stimulator of the innate immune response, induces inflammatory cytokine and chemokine production in many cell types. Our present study showed that MTV augmented poly(I:C)-induced lung injury by synergistically upregulating WISP1 and integrin β3 expression in mice. The result that an anti-WISP1 antibody attenuated lung injury further confirms that WISP1 is involved in the interaction between MTV and poly(I:C). Co-IP results showed that β3 and WISP1 interact in the lungs of mice exposed to both MTV and intratracheal poly(I:C). The synergistic effect of rWISP1 and poly(I:C) on TNF-α secretion is observed in macrophages isolated from WT mice, but not from integrin β3 knockout mice. These results confirm our hypothesis that mechanical ventilation with a tidal volume of 10 mL/kg enhances the expression of WISP1 induced by poly(I:C), as well as the receptor integrin β3, resulting in amplification of the inflammatory response from concurrently administered poly(I:C).
Toll-like receptors (TLRs) play an important role in pathogen recognition and innate immunity. Several TLRs including TLR2, TLR3, TLR6 and TLR10 have been reported to recognize poly(I:C) (24). Our previous study found that WISP1 acts as an endogenous signal through a TLR4-MyD88 pathway to increase alveolar capillary permeability in VILI (7). Studies from others show that cooperative signaling through both TLR4 and TLR3 mediated by MyD88 is involved in lung inflammation induced by mechanical ventilation (6). Consistent with the idea that TLR3 can induce inflammation not only through Trif but also MyD88, Alexopoulou et al. demonstrated that activation of TLR3 by poly(I:C) induces cytokine production through a signaling pathway dependent on MyD88 (2). To determine whether TLRS and signaling downstream were involved in poly(I:C)-induced lung injury, we measured TLR3, TRIF and MyD88 mRNA levels in poly(I:C)-administrated mice. The results showed that 6 h after endotracheal injection of poly(I:C), TLR3 and TRIF mRNA levels increased significantly, but MyD88 levels remained the same as the control (Supplementary Figure S1). This suggests that TLR3-Trif may be activated in our model, but selective deletion of TLR3-Trif would be required to confirm the involvement of this pathway.
Activation of the p38 MAPK pathway plays an important role in virus-induced acute lung injury. It upregulates the inflammatory responses and exaggerates lung injury (25). Stowell et al. showed that MAPK/ERK kinase kinase 1 were significantly upregulated in cells stimulated with dsRNA (26). Alexopoulou et al. also demonstrated that dsRNA can activate the MAPK pathway (2). Other research has shown that lung inflammation can negatively regulate the MAPK signaling (27–29). Since the MAPK signaling pathway is essential for lung injury and pathogen invasion, we measured the protein level of pERK in macrophages exposed to recombinant WISP1 and poly(I:C). As expected, when combined with WISP1, poly(I:C) increased the expression of pERK as compared with poly(I:C) alone. Furthermore, inhibition of ERK signaling inhibited TNF release by macrophages in response to WISP1 and poly(I:C). This result suggests that the synergistic effect of WISP1 and poly(I:C) on TNF-α production may be MAPK-dependent.
Conclusion
In summary, our study confirms that moderated volume mechanical ventilation enhances lung injury and inflammation in the setting of TLR3 activation. We extend work in this area to show that MTV does this through the up-regulation of the matricellular protein, WISP1. WISP1, in turn, signals through integrin β3, which itself is up-regulated in the model of MTV+poly(I:C). These observations suggest that the WISP1-integrin β3 axis should be further explored as both a bio-marker and a therapeutic target in acute lung injury in ventilated patients.
Disclosure
The authors declare that they have no competing interests as defined by Molecular Medicine, or other interests that might be perceived to influence the results and discussion reported in this paper.
References
Sutherasan Y, Vargas M, Pelosi P. (2014) Protective mechanical ventilation in the non-injured lung: review and meta-analysis. Crit Care. 18:211.
Alexopoulou L, Holt AC, Medzhitov R, Flavell RA. (2001) Recognition of double-stranded RNA and activation of NF-kappaB by Toll-like receptor 3. Nature. 413:732–8.
Takeda K, Akira S. (2004) Microbial recognition by Toll-like receptors. J. Dermatol. Sci. 34:73–82.
Altemeier WA, Matute-Bello G, Gharib SA, Glenny RW, Martin TR, Liles WC. (2005) Modulation of lipopolysaccharide-induced gene transcription and promotion of lung injury by mechanical ventilation. J. Immunol. 175:3369–76.
O’Mahony DS, et al. (2006) Mechanical ventilation interacts with endotoxemia to induce extrapulmonary organ dysfunction. Crit. Care. 10:R136.
Chun CD, Liles WC, Frevert CW, Glenny RW, Altemeier WA. (2010) Mechanical ventilation modulates Toll-like receptor-3-induced lung inflammation via a MyD88-dependent, TLR4-independent pathway: a controlled animal study. BMC. Pulm. Med. 10:57.
Li HH, et al. (2012) WNT1-inducible signaling pathway protein 1 contributes to ventilator-induced lung injury. Am. J. Respir. Cell Mol. Biol. 47:528–35.
Jun JI, Lau LF. (2011) Taking aim at the extracellular matrix: CCN proteins as emerging therapeutic targets. Nat. Rev. Drug. Discov. 10:945–63.
Hashimoto Y, et al. (1998) Expression of the Elm1 gene, a novel gene of the CCN (connective tissue growth factor, Cyr61/Cef10, and neuroblastoma overexpressed gene) family, suppresses In vivo tumor growth and metastasis of K-1735 murine melanoma cells. J. Exp. Med. 187:289–96.
Konigshoff M, et al. (2009) WNT1-inducible signaling protein-1 mediates pulmonary fibrosis in mice and is upregulated in humans with idiopathic pulmonary fibrosis. J. Clin. Invest. 119:772–87.
Heise RL, Stober V, Cheluvaraju C, Hollingsworth JW, Garantziotis S. (2011) Mechanical stretch induces epithelial-mesenchymal transition in alveolar epithelia via hyaluronan activation of innate immunity. J. Biol. Chem. 286:17435–44.
Butcher EC, Picker LJ. (1996) Lymphocyte homing and homeostasis. Science. 272:60–6.
Ding X, et al. (2015) RGD peptides protects against acute lung injury in septic mice through Wisp1-integrin beta6 pathway inhibition. Shock. 43:352–60.
Wang B, Wan JY, Zhang L, Min S. (2012) Synthetic RGDS peptide attenuates mechanical ventilation-induced lung injury in rats. Exp. Lung Res. 38:204–10.
Institute of Laboratory Animal Resources; Commission on Life Sciences; National Research Council. (1996) Guide for the Care and Use of Laboratory Animals. Washington (DC): National Academy Press.
Moitra J, Sammani S, Garcia JG. (2007) Re-evaluation of Evans Blue dye as a marker of albumin clearance in murine models of acute lung injury. Transl. Res. 150:253–65.
Lu H, et al. (2015) Conditioned media from adipose stromal cells limit lipopolysaccharide-induced lung injury, endothelial hyperpermeability and apoptosis. J. Transl. Med. 13:67.
Wu R, et al. (2007) Ghrelin attenuates sepsis-induced acute lung injury and mortality in rats. Am. J. Respir. Crit. Care Med. 176:805–13.
Takeda Y, Bui VN, Iwasaki K, Kobayashi T, Ogawa H, Imai K. (2014) Influence of olive-derived hydroxytyrosol on the toll-like receptor 4-dependent inflammatory response of mouse peritoneal macrophages. Biochem. Biophys. Res. Commun. 446:1225–30.
Hou CH, Chiang YC, Fong YC, Tang CH. (2011) WISP-1 increases MMP-2 expression and cell motility in human chondrosarcoma cells. Biochem. Pharmacol. 81:1286–95.
Ono M, Inkson CA, Kilts TM, Young MF. (2011) Young WISP-1/CCN4 regulates osteogenesis by enhancing BMP-2 activity. J. Bone Miner Res. 26:193–208.
Desnoyers L, Arnott D, Pennica D. (2001) Pennica WISP-1 binds to decorin and biglycan. J. Biol. Chem. 276:47599–607.
Wang H, Lim D, Rudd CE. (2010) Immunopathologies linked to integrin signalling. Semin. Immunopathol. 32:173–82.
Sha Q, Truong-Tran AQ, Plitt JR, Beck LA, Schleimer RP. (2004) Activation of airway epithelial cells by toll-like receptor agonists. Am. J. Respir. Cell Mol. Biol. 31:358–64.
Wei D, et al. (2014) Roles of p38 MAPK in the regulation of the inflammatory response to swine influenza virus-induced acute lung injury in mice. Acta Virol. 58:374–9.
Stowell NC, et al. (2009) Long-term activation of TLR3 by poly(I:C) induces inflammation and impairs lung function in mice. Respir. Res. 10:43.
Finkin-Groner E, Moradov D, Shifrin H, Bejar C, Nudelman A, Weinstock M. (2015) Indoline-3-propionate and 3-aminopropyl carbamates reduce lung injury and pro-inflammatory cytokines induced in mice by LPS. Br. J. Pharmacol. 172:1101–13.
Xu Y, et al. (2014) Spred-2 deficiency exacerbates lipopolysaccharide-induced acute lung inflammation in mice. PLoS One. 9:e108914.
Yeh CH, Yang JJ, Yang ML, Li YC, Kuan YH. (2014) Rutin decreases lipopolysaccharide-induced acute lung injury via inhibition of oxidative stress and the MAPK-NF-kappaB pathway. Free Radic. Biol. Med. 69:249–57.
Acknowledgments
This study was supported by National Natural Science Foundation of China grant 81270135 (QL) and SN81300004 (ZXC). This work was performed at Transformation Center of Shanghai East Hospital, TongJi University; Central Laboratory of Shanghai Tenth People’s Hospital, TongJi University. Rongqian Wu helped to improve the language of this paper.
Author information
Authors and Affiliations
Corresponding author
Electronic Supplementary Material
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution-NonCommercial-NoDerivatives 4.0 International License, which permits any non-commercial use, sharing, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, and provide a link to the Creative Commons license. You do not have permission under this license to share adapted material derived from this article or parts of it.
The images or other third party material in this article are included in the article’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
To view a copy of this license, visit (http://creativecommons.org/licenses/by-nc-nd/4.0/)
About this article

Cite this article
Jin, S., Chen, Z., Ding, X. et al. Mechanical Ventilation Augments Poly(I:C)-Induced Lung Injury via a WISP1-Integrin β3-Dependent Pathway in Mice. Mol Med 22, 54–63 (2016). https://doi.org/10.2119/molmed.2015.00233
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.2119/molmed.2015.00233