
Abstract
Understanding about regional versus local changes in vegetation is critical in answering archaeological questions, in particular at a time when humans are assumed to have caused higher disturbances at local scales rather than regional scales; this is the case during the Neolithic. The aim of this paper is to assess the impact of Neolithic land use on regional and local vegetation dynamics, plant composition and disturbance processes (e.g. fire) in eastern Fennoscandia. We apply the Landscape Reconstruction Algorithm (LRA) to high-resolution pollen records from three lacustrine sediment cores that cover the Neolithic period. We calculate changes in vegetation composition and the rate of plant compositional change. Fire dynamics are estimated as an indicator of land use, although fire can result from both natural and anthropogenic disturbances. Our results show that during the Early Neolithic, changes were mainly driven by natural and climate-induced factors and vegetation composition and fire activity were similar at both regional and local scales. From ca. 4000 bc onwards, trends in vegetation and fire dynamics start to differ between regional and local scales. This is due to local land uses that are overshadowed at the regional scale by climate-induced factors. The use of the LOVE model in pollen analyses is therefore very useful to highlight local land uses that are not visible by using REVEALS.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Holocene sediment archives from lakes, ponds and peat bogs in Europe provide evidence of human activities during the last ca. 12,000 years. Deforestation practices aimed at expanding areas for agriculture are commonly identified in pollen records from these archives via a decrease in tree pollen percentages and an increase in herbs and crop pollen types (e.g. Ojala and Alenius 2005; Gaillard et al. 2015; Dubois et al. 2017). Additionally, deforestation and land use cause erosion processes that can be observed in sediment archives (e.g. disturbances in accumulation rates). Anthropogenic impact on vegetation is related to the spread of human activities in a catchment area. It is related to local activities that can expand through time to a regional scale. Based on pollen-based modelling and anthropogenic land cover change scenarios, we have increase our understanding about regional and subcontinental-scale vegetation and land use patterning in Europe, although this information mostly refers to deforestation (e.g. Trondman et al. 2015; Kaplan et al. 2017; Marquer et al. 2017; Roberts et al. 2018; Zanon et al. 2018).
The development of pollen databases (e.g. Fyfe et al. 2009; Giesecke et al. 2014) allows the assessment of changes in Holocene vegetation composition and dynamics at regional and subcontinental scales (e.g. Marquer et al. 2014, 2017; Giesecke et al. 2017). These variations are basically due to climate, environmental (e.g. soil characteristics, characteristic traits of plant species, extreme disturbance events) and/or land use factors. Recent studies have shown that climate and land use are important drivers of vegetation change during the Holocene in northern Europe and Fennoscandia (Reitalu et al. 2013; Kuosmanen et al. 2016, 2018; Marquer et al. 2017). While climate is the dominant factor during the first half of the Holocene, land use becomes one of the primary drivers during the last 2,500 years.
Pollen-based reconstruction of past vegetation at regional and subcontinental scales represents a summary of the outcomes from all the interactions between vegetation dynamics, climate influences and impacts of land use within 104 km2 or larger (Prentice 1985). Land use impacts on vegetation composition and dynamics based on pollen are certainly recorded differently at local scales than at regional scales. The understanding of land use consequences at a local scale is particularly important to explore changes in the landscape-scale vegetation cover during the Neolithic; Neolithic populations, with subsistence based on hunting and gathering, are assumed to have had lower impact on vegetation at a regional scale than at a local scale in some areas. Furthermore, recent studies show that forest landscapes may have been shaped by human manipulation over millennia, although to a small extent. These human activities have caused local changes in biodiversity, species distributions and biomass (Dambrine et al. 2007; Innes et al. 2013; Pini et al. 2017). A better knowledge of early anthropogenic impact at a local scale can provide insights into human-nature relationships at a time when populations were highly dependent on climate and environmental variabilities (Boivin et al. 2016). However, this information can only be gained by studying known spatial resolutions and using adequate indicators of vegetation change, such as land cover, plant diversity and composition indices.
Traditional pollen data expressed as proportions provide information about a mix of the regional and local pollen assemblages, thus making it difficult to assess the local changes in vegetation. The Landscape Reconstruction Algorithm (LRA, Sugita 2007a, b) was proposed to overcome this issue and provides pollen-based reconstructions of regional and local vegetation in quantitative terms, i.e. plant cover at a defined spatial scale (a few metres to kilometres from the center of the lake/peat bog). In particular, the LRA corrects for biases caused by basin size, inter-taxonomic differences in pollen productivity and dispersal/deposition. The LRA has already been applied in various environmental contexts (e.g. Sugita et al. 2010; Cui et al. 2014; Poska et al. 2014; Hjelle et al. 2015; Mazier et al. 2015; Abraham et al. 2017; Marquer et al. 2020). A better understanding about regional versus local changes in vegetation is critical in answering archaeological questions, in particular at a time when humans are assumed to have caused higher disturbances at local scales than regional scales, this being the case during the Neolithic.
Recent studies suggest that the Neolithization of Eurasia was a more complex process and the Neolithic a more diverse phenomenon than traditionally assumed (Barker 2006; Fuller et al. 2011; Herva et al. 2017; Nordqvist 2018). As pointed out by Bogucki (1995) and Smith (2001), an extensive and systematic manipulation of resources and some degree of control of wild species was already taking place during the Mesolithic. This was to such a degree that the static categories of “Mesolithic” and “Neolithic” are not correct characterizations defining the Stone Age (see also Nordqvist 2018). There are examples of societies where combined foraging and low-level use of cultivars were long-term and stable strategies that maintained foraging options rather than replacing wild foods. Smith (2001) has proposed the term “low-level-food production” for the “in-between territories” that are neither hunting, fishing and gathering economies based exclusively on wild plant and animals, nor farming relying heavily on agricultural products. In western Norway, Hjelle et al. (2006) have argued that cereals and domesticated animals were introduced by some hunter-fisher groups during the early and middle Neolithic without any significant changes in the traditional economy and settlement pattern. Additionally, according to Barker (2006) there was not a steady “wave of advance” of a Neolithic package of new technologies and new forms of subsistence, but rather a piecemeal process of adoption of such package components by the indigenous foraging populations in different environmental and cultural contexts. New insights around the above archaeological issues might be obtained by a deeper understanding of the consequences of land use at local scales, i.e. by studying the environment the human populations lived in.
Within this context, the aim of the present paper is to assess the impact of Neolithic land use on the regional and local vegetation dynamics, plant composition and disturbance processes (fire). The study focuses on eastern Finland, where the Neolithic period begins with the appearance of pottery, ca. 5300–5200 bc (Pesonen and Leskinen 2009; Pesonen et al. 2012; Halinen 2015). The impact of Neolithic land use has already been studied in this region by using qualitative information, i.e. conventional pollen percentages and concentrations (grains/cm3) from sediment archives. These previous studies indicate that during the Early and Middle Neolithic hunter-gatherers increasingly started to impact the flora and vegetation structure and to incorporate cultivars into their economy (Tolonen 1978; Reynaud and Hjelmroos 1980; Königsson and Possnert 1997; Vuorela et al. 2001; Poska et al. 2004; Poska and Saarse 2006; Alenius et al. 2013, 2017; Augustsson et al. 2013; Alenius et al. in press). Here, we apply the LRA to high-resolution pollen records from three lacustrine sediment cores that cover the Neolithic period. We estimate changes in vegetation composition, the rate of plant compositional change and fire activity. Fire occurrence is here considered as an indicator of land use (e.g. Scherjon et al. 2015), although fire can result from both natural and anthropogenic disturbances (see “Discussion”).
Materials and methods
Environmental setting and archaeological context
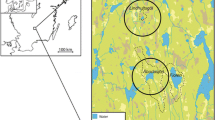
The study area (Fig. 1) is located in the southern boreal vegetation zone of southern Finland. It belongs to the Finnish Lake District, where lakes occupy about 25% of the region. The Lake District expands into most of central and eastern Finland and extends further east to the Karelian Isthmus and the Republic of Karelia in Russia. Forests in this area are dominated by Picea abies, Pinus sylvestris, Betula pendula, Betula pubescens, Populus tremula, Alnus incana and Alnus glutinosa (Alalammi 1988). The region is classified as lowland with a terrain relief between of 30 and 75 m a.s.l.

a Map of the study area in eastern Finland. The red rectangle in A shows the extent of map B. b Lake District showing the locations of sites used for REVEALS-based vegetation reconstructions. The red rectangle in B shows the extent of map C. c Local study area showing the locations of sites for LOVE-based vegetation reconstructions (lakes Huhdasjärvi A and C, and lake Katajajärvi). Regional fire dynamics are obtained from the lakes used for REVEALS, with the addition of charcoal data from lake Ahvenainen (marked with a gray circle in B). Local fire dynamics are obtained from the two sites used for LOVE (lakes Huhdasjärvi A and C)
Both lakes Katajajärvi and Huhdasjärvi belong to the drainage basin of the River Kymijoki, and were isolated from the Baltic Sea basin at around 9650 bc (Saarnisto 1970; Björck 1995; Lunkka et al. 2004). The bedrock includes primarily metavolcanic and metasedimentary rocks and several generations of granitoids (Lehtinen et al. 2005). The quaternary geology of the study area is dominated by till, sand, clay and organic deposits that mostly fill the depressions of crystalline bedrock. Valleys, eskers and drumlins are typical landforms in the area (Simonen 1980; Tyrväinen 1986; Alalammi 1988; Lunkka et al. 2004). Today, the length of the growing season is 160–175 days/year and the mean annual precipitation is 550 mm, of which about 200 mm is received as snow. The mean annual temperature varies between + 3.5 and + 4 °C, the warmest month is July (+ 17 °C), and the coldest are January and February (between − 9 and − 8 °C) (Alalammi 1988). The archaeological finds indicating Stone Age habitations around the lakes Huhdasjärvi and Katajajärvi are modest; most of the sites have not been dated more accurately than to the Stone Age or to the subsequent Early Metal Period, i.e. this corresponds to 1800 bc–ad 300 in the inland areas of Finland. More generally, eastern Finland is rich in rock paintings dated back to the Stone Age (Fig. 1), often located by the waterways and representing people, boats, elk, fish and human figures (Lahelma 2008; Gjerde 2010).
Pollen and charcoal data
Three sediment cores (Fig. 1) were sampled for pollen analyses. One of the cores was obtained from lake Katajajärvi (Alenius et al. 2009), and two (Huhdasjärvi A and C) from Pitkälahti bay about 13.4 km from lake Katajajärvi (Alenius et al. 2017). Pitkälahti bay is a small oblong area belonging to lake Huhdasjärvi. Coordinates, size of the lakes and references are given in Table 1. The two sediment cores obtained from Pitkälahti bay were situated 450 m apart. Empirical and theoretical studies in Scandinavia (e.g. Nielsen and Sugita 2005; Hellman et al. 2009; Hjelle and Sugita 2012; Cui et al. 2014; Hjelle et al. 2015; Mazier et al. 2015; Fredh et al. 2019) have demonstrated that the RSAPs (Relevant Source Areas of Pollen; sensu Sugita 1994) for small lakes correspond to a large range of values (ca. 900 to 3000 m) most likely resulting from different vegetation patch sizes, spatial structure of the local landscapes and the basin size. This means that the RSAPs of the two cores collected in Pitkälahti bay most likely overlap. An earlier analysis based on pollen proportions (ESM 1) has shown differences in the pollen data between these two cores (Alenius et al. 2017), which suggests that the two coring sites can be considered as distinct palaeoecological records. Furthermore, the squared chord distance analysis performed between the three sites shows that the differences in term of vegetation composition are higher between Huhdasjärvi A and C than between one or the other of the Huhdasjärvi sites and Katajajärvi (see Fig. 2 and “Discussion” below). This underlines that the pollen mixing during the deposition of sediment cannot be considered as homogeneous for the entire lake, but rather specific to locations defined by lake morphology. Such weak mixing of pollen within the entire lake would result in an overrepresentation of local pollen in the bays. This means that bays from a large lake might be assumed to be different small sites for LRA reconstructions. Note that a major assumption of the LRA is that lakes are of circular shape and the pollen grains, once deposited on the lake surface, are well mixed in the water column during the sedimentation processes. For the application of the LRA to our study, we therefore made the assumption that each site (Huhdasjärvi A and C) is an individual small lake of circular shape. However, this assumption has not been tested, i.e. comparison between LOVE reconstructions using modern surface pollen assemblages and actual plant cover around these sites; this might be a source of uncertainties in our LOVE estimates that should be taken into account in the interpretation of the results.

Differences (squared chord-distances) between: a the sites at the local scale (LOVE; LV); b the sites at the local and regional scales (REVEALS; RV); and c the sites that have been used for the REVEALS runs. HC: Huhdasjärvi C; HA; Huhdasjärvi A; K: Katajajärvi; KAAK: Kaakotinlampi; LAIH: Laihalampi;HUH: Huhdasjärvi 1; ORI: Orijärvi; KIRK:Kirkkolampi. For example, LVHA-HC means the differences between LOVE results from HA and HC for each time window
Charcoal data were collected from Huhdasjärvi A and C in order to reconstruct local fire dynamics at the two selected sites during the Holocene. The charcoal particles > 10 µm (measured along the longest axis) were counted on pollen slides and expressed as microcharcoal concentrations, i.e. number of fragments/cm3. Each charcoal record is standardized and normalized according to a protocol frequently used by the palaeofire community (Power et al. 2010). Charcoal influxes are then converted to Z-scores for an interpretation of individual values with respect to variation around the long-term mean (here the interval 1,000–6,000 bp, i.e. the period most representative of the entire database). Mean Z-scores are then computed at 200-year intervals. For the study of fire dynamics at the regional scale, we used the composite charcoal curve already published in Kuosmanen et al. (2018), constructed by grouping together all available records in southeast Finland. Differently from Kuosmanen et al. (2018), where the authors focused on the period 10,000–1,000 (i.e. the time where data on human population were available), in the present paper we extend the palaeofire reconstruction over the entire Holocene. Also in this case, charcoal data are expressed as transformed charcoal influx (hereafter referred as tCHAR) Z-scores.
Chronologies of Huhdasjärvi A and C are based on palaeomagnetic dating supported by three bulk AMS radiocarbon dates (Alenius et al. 2017). An age-depth model for Lake Katajajärvi uses four calibrated AMS radiocarbon ages and linear interpolation; the age-depth model agrees well with the known regional dates for the increase of Picea, Tilia and Alnus pollen percentages obtained from the literature (Alenius et al. 2009). Age-depth models, bathymetry and coring locations for Lake Katajajärvi, Huhdasjärvi A and C are presented in ESM 2.
The landscape reconstruction algorithm (LRA)
The LRA approach (Sugita 2007a, b) consists of two sub-models. The first sub-model is REVEALS for plant cover reconstruction at a regional scale of ca. 50–100 km radius around a large lake (radius > 50 ha) or a mix of several lakes and/or bogs of small sizes (radius < 50 ha; Marquer et al. 2014, 2017; Trondman et al. 2015, 2016). The second sub-model is LOVE for plant cover reconstruction at a local scale of about a few metres to kilometres from each of the small size sites from which pollen records are obtained. LOVE estimates correspond to vegetation composition within the RSAP at each site and are obtained by subtracting the regional background of pollen (calculated from the REVEALS estimates) from the overall pollen assemblages (Sugita 2007b).
For the LRA runs we use twenty-three pollen types and the relative pollen productivity estimates (RPPs) obtained from various regions in Europe (the LANDCLIM standard 2 dataset of Mazier et al. 2012) that have been previously applied for all sub-continental and regional pollen-based vegetation reconstructions (Marquer et al. 2014, 2017; Trondman et al. 2015; Kuosmanen et al. 2018). RPPs and their standard errors, fall speeds of pollen, and the different land cover categories used in this study are given in Table 2. Plant taxa are here grouped into four land cover categories: forest/woodland, meadow/grassland, cropland and wetland. The LRA results are expressed as percentage cover of plant types or land-cover categories with their standard errors. Standard errors of the REVEALS and LOVE estimates are obtained by a delta method (Stuart and Ord 1994).
Regional vegetation, REVEALS estimates
The REVEALS estimates used to reconstruct southeast Finland regional land-cover over the last 10,000 years are those already published in Kuosmanen et al. (2018). Note that in Kuosmanen et al. the data are presented only for the time period 1,000 to 12,000 years ago, but we here show the whole last 12,000 years. REVEALS estimates for the last 1,000 years were not shown in Kuosmanen et al. because of the focus of the study on a comparison between REVEALS and human population estimates that did not cover the last millennium. Time intervals of 200 years were used, except in the three uppermost time windows (ad 2000–1950, 1950–1900 and 1900–1850) where time intervals of 50 years were used. These estimates were based on five pollen records (lakes Orijärvi, Kirkkolampi, Laihalampi, Kaakotinlampi and Huhdasjärvi 1; Fig. 1, Table 1, ESM 3); for all details see Kuosmanen et al. 2018. The core Huhdasjärvi 1 (Alenius et al. 2013) was collected from the same lake as Huhdasjärvi A and C (used for the LOVE reconstructions). However, the Huhdasjärvi 1 sediment core was collected from a large open area and the pollen records are assumed to represent the regional vegetation. The Huhdasjärvi A and C cores were obtained from Pitkälahti bay and the pollen records from these cores are assumed to represent the local vegetation signals (see ‘Pollen and charcoal data’).
Local vegetation, LOVE estimates
The REVEALS estimates (see previous section) are used as inputs in LOVE for the regional vegetation abundance. The RSAP can be calculated by using an inverse modelling approach including in the LOVE model, however because of the low numbers of similarly-sized sites available for our analysis it would be difficult to obtain reliable RSAPs by using this approach. RSAPs have been estimated in southern Sweden by using a forward-modelling approach (Sugita 1994; Sugita et al. 1999) and hypothetical landscapes based on modern vegetation maps (Mazier et al. 2015). The results reveal predicted RSAPs of radii of 1,740 and 1,440 m from the centres of the lakes studied by Mazier et al. Because of similarly-sized sites and the use of pollen records from bays would overrepresent local pollen (see “Pollen and charcoal data” above), the lowest RSAPs of Mazier et al. would be relevant for our study. We have therefore considered a RSAP of radius 1,400 m as the spatial unit of the LOVE reconstructions around the sites. We further assume a constant RSAP through time. For the sizes of each lake the nearest distances from the shores have been measured to calculate the lake radii (Table 1). The time interval used for the LOVE runs is 200 years. Pollen counts falling within each time interval are pooled together. The longest time span (lasting from 7950 bc to ad 1580 with a total of 49 time windows) is obtained from Lake Katajajärvi, while the shortest span (lasting from 6550 bc to ad 450 with a total of 36 time windows) from Lake Huhdasjärvi A. We use the LOVE version 5.0 program (Sugita unpublished). Note that wind speed is set to 3 m s−1 and the maximum spatial extent of the regional vegetation from the centre of the study sites to 50 km. Neutral atmospheric conditions are considered for these runs. A Gaussian Plume Model (Sutton 1953; Tauber 1965; Prentice 1985; Sugita 1994) is used to describe pollen dispersal and deposition characteristics.
Indices of change in vegetation composition and CONISS
Rate of change in vegetation composition, vegetation turnover and evenness are calculated using the REVEALS- and LOVE-based results. Rate of change aims at identifying periods of stability and change in vegetation over time. Turnover represents the magnitude of change in vegetation over time. Evenness refers to how numerically equal the plant taxa are in their abundances (i.e. if all taxa are equal in their abundances the evenness is 1, maximum evenness). For methodological details see Marquer et al. (2014, 2017). In order to identify periods of similar vegetation composition, a stratigraphically constrained cluster analysis (CONISS; Grimm 1987) is used for both REVEALS and LOVE estimates. We further performed a squared chord distance analysis to assess the differences between the three “local sites” as well as between each of the local site and the REVEALS estimates. The squared chord distances have also been calculated between the five sites that have been used for REVEALS to test the assumption that in theory the sites used for REVEALS runs should be more similar to each other than to the LOVE sites.
Results
Changes in plant abundance and vegetation composition
Regional vegetation, REVEALS
Four major regional CONISS zones (CONISS R1 to R4) are observed based on the REVEALS estimates (Fig. 3). The main features of each CONISS zone are described in Table 3.

REVEALS-based estimates of the regional vegetation composition with their standard errors. Data are from Kuosmanen et al. (2018); the results are shown here for the whole of the last 12,000 years. The results of the CONISS analysis are given
The results for the rate of change, turnover and evenness are presented in Fig. 4. The rate of change remains low from 6650 to 5050 bc, slightly increases between 5050 and 3250 bc, then rises (higher variation than previously) from 3250 bc onwards with the highest value reached at the present time. A high turnover between 6850 and 4850 bc is followed by a decreasing trend until ad 1750. Then, an increase is recorded until the present time. Evenness is high between 6850 and 3450 bc, then a decreasing trend characterizes the period 3450 bc–ad 550. A new increase is visible from ad 550 until 1875.

Rate of change, turnover and evenness of vegetation at the regional scale (REVEALS; RV) and the local scales (LOVE; LV)
Local vegetation, LOVE
The four major CONISS zones identified in Huhdasjärvi C (HC1 to HC4), Huhdasjärvi A (HA1 to HA4) and Katajajärvi (K1 to K4) are shown in Fig. 5 and described in Table 3.

LOVE-based estimates of the local vegetation composition with their standard errors at lakes Huhdasjärvi A and C, and lake Katajajärvi
Considering the indices of change in vegetation composition, a low rate of change from 6650 bc until 2650 bc is followed by increasing and highly variable values between 2650 bc and ad 950. Turnover is high from 6850 to 3850 bc, and a decreasing trend characterizes the period between 3850 bc and ad 1350. Evenness is stable/slightly increasing from 6850 bc until 3850 bc, then it decreases during the period between 3850 bc and ad 1350.
Regional versus local vegetation dynamics
In general, similar trends in Holocene forest development (Fig. 6) are visible at both the regional and local scales. However, open land (meadow/grassland, cropland and wetland) and forest/woodland categories are respectively less and more abundant at the local scales than the regional scale. Some individual tree taxa (e.g. Picea) are, nevertheless, more common at the regional scale. In detail, one important difference is the spread of Picea. Between 5550 and 4750 bc in REVEALS Picea constitutes 1.8%, while LOVE estimates for this taxon are still very low (0.1–0.2%). Differences can also be seen in the relative abundances of tree taxa. Regionally, between 4750 and 1800 bc, Picea constitutes 26.8% (percentages are averaged REVEALS and LOVE values over the time interval) and Pinus 15% of forest/woodland vegetation, while at the local scales Picea abundance is lower than 6% and Pinus is more abundant (37.8–39%). With respect to broadleaved trees, during the same time period the regional abundance of Betula and Alnus together reaches 39%, while locally their vegetation cover is ca. 48%. Thermophilous trees (i.e. Corylus, Fraxinus, Tilia, Ulmus) are also more abundant regionally (9.7%) than locally (ca. 7%). Of meadow/grassland taxa, Juniperus shows an increase in Huhdasjärvi A around 4150 bc, Huhdasjärvi C around 4350 bc and in Katajajärvi around 3350 bc (more noticeably around 1750 bc), while such an increase is only visible at the regional scale at ad 550. Of wetland taxa, Salix is present in 0.3–0.8% in the local and regional reconstructions.

REVEALS and LOVE estimates for the land cover categories: forest/woodland, meadow/grassland, cropland and wetland. Thermophilous trees include noble deciduous taxa Corylus, Ulmus, Quercus, Tilia, Carpinus, Fraxinus and Fagus. Reconstruction of regional fire activity (tCHAR Z-scores) based on the “composite charcoal curve” from all lakes available in southeast Finland (see Kuosmanen et al. (2018) for details). Reconstruction of local fire activity (tCHAR Z-scores) based on “microscopic charcoal concentrations” from Huhdasjärvi A and C
Trends in evenness and turnover are similar between the local and regional scales. Differences are observed in terms of timing and amplitude of changes. The decrease in evenness between 3250 bc and ad 550 is more pronounced at the regional than at the local scales. While the local rate of change is in general higher than the regional one for the same time periods, values remain low at both the regional and local scales until 3250 and 2650 bc, respectively. Note that the rate of change at Lake Katajajärvi does not show an increasing trend.
The squared chord distance analysis (Fig. 2) shows few differences between the three local sites; the main differences are observed between Huhdasjärvi A and C. The dissimilarity between Huhdasjärvi A and C is essentially caused by differences in abundances of Betula, Picea, Pinus and Salix, in particular a high dissimilarity is observed at ca. 1400 bc due to Picea. Differences between the regional and local signals are increasing from ca. 4250 bc; they are higher than between the LOVE sites. The squared chord distances between the five sites that have been used for REVEALS (Fig. 2) support the assumption that in theory the sites used for REVEALS are similar to each other with a mean of squared chord distances lower than 0.2 and little variability between sites. There are more differences and variabilities between the REVEALS and LOVE estimates than between the sites that have been used for the REVEALS runs.
Fire dynamics at regional and local scales
Main trends of the regional and local fire-dynamics are presented in Fig. 6. At the regional scale, fire activity is above the long-term mean (i.e. positive tCHAR Z-scores) between 9750 and 3000 bc. A period with negative values occurs from 3000 bc until ad 600. Finally, fire activity is higher than the long-term mean during the last 1,400 years (between 600 ad and the present day).
At the local scale, trends in fire activity slightly differ between the two sites analyzed. Huhdasjärvi A is characterized by higher tCHAR Z-scores compared to the long-term mean between 6450 and 3800 bc, with a short period of lower fire activity between 5300 and 5000 bc. Then the values start to decrease, with negative tCHAR Z-scores during the period 3700–1200 bc. From 1200 bc to ad 400 the site is characterized by higher fire activity. Negative tCHAR Z-scores are finally recorded between ad 400 and 750. At Huhdasjärvi C, tCHAR Z-scores are characterized by higher values compared to the long-term mean between 6650 and 3400 bc. Despite some oscillations, fire activity between 3400 bc and ad 600 is similar to the long-term mean (tCHAR Z-scores around 0). Since ad 600, fire activity increases, with a peak at ad 950.
Discussion
Roles of climate and environmental factors in vegetation composition
Our results show similar trends in Holocene forest development at both the regional and local scales. Between 6850 and 4850 bc both the regional and local turnovers reach maximum values and the regional rate of change is the lowest. These changes likely relate to the spread of new tree taxa in the region, such as Tilia and Picea around 6000 bc. The highest abundances in thermophilous taxa (21–23%) are reached during the Early Neolithic, between 4750 and 4350 bc. This coincides with the Holocene Thermal Maximum (HTM), ca. 6000–3000 bc in southern Fennoscandia, when temperatures were ca. 1.5–2.5 °C higher than the present time (Heikkilä and Seppä 2003; Ojala et al. 2008; Seppä et al. 2009b).
The HTM was followed by an overall cooling and the southward retreat of noble deciduous tree taxa. A gradual decrease in thermophilous tree taxa is clear in both the regional and local vegetation reconstructions since around 4000 bc. This significant change in the Fennoscandian ecosystem is related to the spread of Picea after ca. 4000 bc (Giesecke and Bennett 2004; Seppä et al. 2009a). Oxygen isotope records in sediments show an increase in relative humidity after 3000 bc in the region (Hammarlund et al. 2003; Heikkilä and Seppä 2010; Kostrova et al. 2018), with an effect on the natural fire frequency. This is in agreement with our regional charcoal results, showing a long period of low fire activity from 3000 bc until ad 600. This period 4450 bc–ad 550 is further characterized by a gradual decline of turnover and evenness in vegetation. It can be argued that the reason for such changes is the progressive replacement of “former” species-rich mixed forests with more homogeneous “modern” boreal forests dominated by Picea, Pinus and Betula.
Compared to the regional abundances, local abundances are lower for Picea and thermophilous taxa (only 7.6% between 4850 and 4250 bc in Huhdasjärvi A), and higher for Pinus; this explains part of the increasing differences between the local and regional vegetation composition in Fig. 2. This difference is probably caused by the distribution of surface deposits. Till deposits dominate the area, while sorted glaciofluvial deposits, such as eskers, are less represented but favour Pinus that predominantly grows on these drier and nutrient-poor patches. Shade-tolerant Picea prefers moist and nutrient-rich soils (Sutinen et al. 2002) and forms the typical boreal forest zone on till-dominated soils of Finland. Thus it is not a surprise that Picea dominates the late Holocene regional vegetation based on REVEALS estimates from five basins across the area, while Pinus can be locally more common, in particular around Huhdasjärvi, located by a large area of glaciofluvial deposits. There are also differences between the two sites from Pitkälahti bay, as Picea has been (according to LOVE-based estimates of the local vegetation composition) slightly more common at the end of the bay than the middle part. We assume that the results reflect real differences in vegetation around the bay rather than major differences in pollen dispersal and/or deposition patterns, since the cores were collected from two individual small accumulation basins within the bay. This is supported by the uniform sediment accumulation rates between the cores through time.
The trend in the rate of vegetation change is increasing from 3250 bc at regional scale, and later (from 2650 bc) at local scales. Our results agree with turnover, evenness and rate of change observations studied at regional and subcontinental scales (Marquer et al. 2017) and are likely to be caused by Holocene climate changes.
Role of late Mesolithic and Neolithic land use in vegetation composition and fire activity
From late Mesolithic to Middle Neolithic (6450–3200 bc )
Fire activity
Between 6450 and 3800 bc both the regional and local fire activity are generally characterized by values above the long-term mean. Despite some oscillations, the local and regional fire activities are higher than the long-term mean until 3800 and 3000 bc respectively, suggesting the widespread use of fire. As there is plenty of evidence of human activity around the lake Huhdasjärvi during this period (detected from pollen and loss on ignition), human induced fires could be a reason for high values in fire activity. In the earlier study from Huhdasjärvi (analysed from Huhdasjärvi 1; Alenius et al. 2017) an abrupt erosion phase was dated to between 6340–5810 bc, and was found to coincide with phases of increased fire intensity. This was then followed by the increase in species richness from ca. bc 5260, simultaneous with a decrease in Pinus and Betula, indicating an increase in the openness of the landscape. Also in the cores obtained from Pitkälahti bay, the earliest human impact was earlier dated to between ca. 4400 and 3900 bc. It includes some use of fire and cultivation of hemp and barley. In addition, a sudden erosion phase was dated between 4165 and 4090 bc. In addition to the cultivated plants Hordeum and Cannabis, apophytes such as Plantago major/media, Rumex, Fabaceae, Campanula and Caryophyllaceae are recorded.
During the Neolithic burning practices are known to have played an important role in opening up landscapes to favour the growth of edible plants such as raspberry, juniper (Juniperus), hazel (Corylus) and wild strawberry (Fragaria vesca) (Vanhanen and Pesonen 2015). All these species have a competitive advantage in early successional communities but decline in abundance during later successional stages (Smith 2001). During this period fire could also have been used to clear forests for increasing grazing lands for large ruminants, such as moose that prefer young forests. Osteological material obtained from eastern Finland shows that people used moose and forest reindeer as mammal resources, in addition to aquatic resources, such as beaver and Saimaa ringed seal (Ukkonen 1996; Oinonen et al. 2014).
Early decrease of Tilia abundance at a regional scale
In the local reconstructions, Tilia abundances are decreasing in lakes Katajajärvi and Huhdasjärvi from ca. 4000–3200 bc. This decrease is also observed at the regional scale but a bit earlier than at the local scales, ca. 4600 bc. This decrease in Tilia abundance can be interpreted as a consequence of the natural gradual decrease in Tilia abundance due to the overall cooling and the southward retreat of noble deciduous tree taxa (Huntley and Birks 1983; Seppä et al. 2009a, b). These results suggest that the decreases in Tilia abundances have not been simultaneous in the studied region, which can be seen at the local scales. The difference in timing of local Tilia declines might be caused by land use, e.g. differences in the intensity and extent of land use between various locations. Tilia fluctuations at lake Huhdasjärvi, occurring synchronously with other settlement indicators (increase in Juniperus, Cannabis, Rumex), might suggest management of Tilia woodlands by small-scale human groups that can only be detected at the local scale. In the northern German Neolithic, the synchronous decline of Tilia values together with the simultaneous occurrence of settlement indicators has been interpreted as an anthropogenic influence on the woodland composition (Kirleis et al. 2012).
Manufacture of lime bast cordage has been an unbroken tradition from the Mesolithic to the present day in northern Europe (Myking et al. 2005). In eastern Baltic Sea regions, lime fibres are known in prehistory to be utilized for binding material for stationary wooden fishing structures (Bērzinš 2008) and lime bast is also widely used as a raw material for different textiles (Bender Jørgensen 1990; Schweingruber 1990; Rimkutė 2010; Mannering et al. 2012). Lime bast is usually obtained from young trees. Coppicing i.e. the periodic cutting-off at ground level is a traditional way to encourage the growth of multiple young shoots. Note that it has been demonstrated that the amount of pollen produced by Tilia cordata is heavily reduced as a result of periodic coppicing and is therefore likely to be ‘palynologically invisible’ (Waller et al. 2012). In addition to being used for bast, coppiced lime has also been used for leaf fodder for livestock in many parts of Europe (e.g. Rasmussen 1989). Pollen spectra from eastern and northern Denmark have revealed felling and burning of lime woodlands for livestock grazing and cereal cultivation (Andersen 2012). In Finland, osteological evidence of domesticated animals (excluding dogs) for this time period is missing. The earliest sheep/goat bone dates to end of the third millennium bc (Bläuer and Kantanen 2013). However, it should be noted that unburnt bone does not generally preserve in the acidic soil conditions of Finland and all the available osteological material is burnt.
Towards local semi-open landscapes from ca. 4000 bc
From ca. 4000 bc, differences in vegetation composition between the local and regional scales are increasing; see Fig. 2. The most notable difference is visible in Juniperus percentages, as the LOVE estimates for Huhdasjärvi and Katajajärvi show an increase in its abundance; from 4350 bc at Huhdasjärvi C, 4150 bc at Huhdasjärvi A and 3350 bc at Katajajärvi (with a more noticeable increase from 1750 bc onwards).
Increase in Juniperus percentage cover indicates a change in vegetation structure towards semi-open landscapes, where light-demanding Juniperus grows better. This might be due to the local land use that is not visible at the regional scale. Changes in Juniperus abundance might be more influenced by climate and environmental factors at the regional scale than at the local scale. Juniperus increases at the local scale were probably caused by land use. This hypothesis of change in land use towards semi-open landscapes is supported by the simultaneous, yet sporadic evidence of cultivation of Cannabis and Hordeum, earlier dated in lake Huhdasjärvi pollen sequences to between ca. 4400 and 3200 bc, also indicating longer occupation at the site (Alenius et al. 2017). Earlier, increase in Juniperus percentage cover was likewise dated to between 4480 and 3250 bc in Lake Bol’shoye Zavetnoye (about 180 km to the east in the Karelian Isthmus, Russia) and was interpreted as indicating intensified human activity and prolonged human occupation at the site (Alenius et al. 2020).
In timing, this change in vegetation pattern towards semi-open landscapes agrees with the arrival of a new pottery tradition called Typical Comb Ware (TCW in Finland: 4000–3400 bc). During the early TCW period, there was an increasing tendency to locate housepit sites in unsheltered and aquatically-oriented locations, coincident with the appearance of village-like clusters of housepits. These changes have been interpreted as reflecting a growing degree of sedentism (Mökkönen 2011). The appearance of TCW is marked by new contact networks that are in some regions explained through small-scale migration and in other areas through cultural diffusion, the movement of new ideas and material goods (Nordqvist and Herva 2013; Herva et al. 2014; Nordqvist and Kriiska 2015).
If we accept the identification of the earliest Cerealia- and Cannabis-types of pollen as a proof of the growing of domesticated plant taxa, interesting questions regarding the nature and scale of these activities arise. The term cultivation usually refers to the growing of domesticated crops for food and other purposes, but it can also refer to non-agricultural contexts of wild-plant exploitation. The meaning has expanded to include a range of activities that promote plant growth such as tillage, land clearance, plant sowing, weeding, harvesting and deliberate burning of vegetation (Harris 2007; Harris and Fuller 2014; Fuller and Lucas 2017). If the sites were settled for a longer period than before, it is likely that shifting cultivation was not taking place but, instead, cultivation took place in longer-term, permanent plots. Sometimes the terms horticulture and garden cultivation are used in the literature for traditional agricultural systems. According to Harris (2007), gardens are smaller and are floristically more diverse than fields. In that sense, the growing of domesticated species around lakes Huhdasjärvi and Katajajärvi might relate to garden cultivation in permanent plots and possibly included other wild-plant management and activities. Nevertheless, it is clear that between ca. 4400 and 3200 bc cultivation did not form the basis of subsistence in southeastern Finland (a region situated in the boreal forest zone beyond 60° N latitude) and was not in the form of intensive field cultivation as in Central Europe (see also Núñez 1999; Mökkönen 2010; Nordqvist and Kriiska 2015). Hunting, fishing and gathering remained as subsistence sources even when the knowledge of cultivation had reached the area.
The highest HTM summer temperatures and the lowest relative humidity took place in Fennoscandia at ca. 4000 bc (Seppä et al. 2009b; Heikkilä et al. 2010; Salonen et al. 2014). The climatic conditions appear to correlate with: (1) the increase in Juniperus (from 4350–4150 bc onwards) and (2) the earliest sporadic signals of cultivation of domesticated species (in lake Huhdasjärvi between ca. 4400 and 3200 bc), both suggesting in situ human occupation, together with (3) the spread of TCW. In this respect, at that time there should not have been climatic limitations for the outset of small-scale, sporadic growing of cultivars, especially with the onset of a new, more sedentary, style of life. Coincident with the HTM and related high productivity of terrestrial, lacustrine and marine ecosystems, human population proxies indicate a substantial growth in hunter–gatherer populations supported by an increase in resource availability (Oinonen et al. 2010; Tallavaara et al. 2010; but see also Crombé and Robinson 2014).
From local to regional land use: from Late Neolithic (3200–1800 bc ) to Preindustrial period
A significant increase in the rate of change takes place from 2450 bc onwards in Huhdasjärvi A and 2050 bc onwards at Huhdasjärvi C. This increase corresponds to a first decrease in Betula and increases in the abundances of Picea and Pinus, based on LOVE reconstructions. A climate-induced change in vegetation composition might be suggested as a main explanation for this increase in the rate of change; an increase in boreal forest components, particularly Picea, at that time is commonly assumed to be climate-induced.
However, an increase in organic matter input into the bay, potentially caused by erosion due to human activities near to the shores of Huhdasjärvi, was recorded by earlier studies at both coring sites between ca. 2600 and 1840 bc, with the highest peak in organic content dated to ca. 2300 bc (Alenius et al. 2013, 2017). Furthermore, a Hordeum-type pollen grain was identified at ca. 2160 bc (Alenius et al. 2017), also indicating human occupation around the lake. The rates of change at the regional scale and at lake Katajajärvi are not as notable as in Huhdasjärvi A and C. All of this might indicate a local-scale land-use activity located near Huhdasjärvi.
Both the regional and local trends in fire activity decrease during this period. At the regional scale, the charcoal composite curve shows values lower than the long-term mean between 3000 bc and ad 600. Locally, Huhdasjärvi A is characterized by negative tCHAR Z-scores between 3800 and 1200 bc, while negative values are recorded at Huhdasjärvi C between 3700 bc and ad 600. This might suggest few human-induced fires at those times.
From 1750 bc onwards the local vegetation near lakes Huhdasjärvi A and Katajajärvi are both characterized by a decline in Betula, Ulmus and Corylus and an increase in Picea and Juniperus. The plausible reason might be the grazing of domesticated animals and those feeding in broadleaved forests (Behre 1981; Pykälä 2001). In timing, the LOVE results agree with the earliest osteological evidence. In Finland, the earliest secure appearance of livestock dates to the end of the Late Neolithic, ca. 2500–950 bc (Bläuer and Kantanen 2013; Cramp et al. 2014; Ukkonen and Mannermaa 2017; Ahola et al. 2018). The REVEALS results however do not show similar patterns, change in forest plant composition, suggesting that the land use has remained small scale and local. Only from ad 550 onwards does the regional vegetation show a significant reduction of forests along with the presence of domesticated cereals, meadows and grasslands and, from ad 750 onwards, the spread of slash-and-burn practices. The expansion of slash-and-burn practices becomes visible in the regional charcoal composite curve and, locally, at Huhdasjärvi A. The expansion of slash-and-burn practices during the Iron Age is also known from other sources (e.g. Taavitsainen 1987; Taavitsainen et al. 1998). All of this indicates a shift towards agricultural communities, where farming is the predominant activity and determines the main diet, although hunting, fishing and gathering may have continued. Both slash-and-burn and permanent field cultivation were in use from the 17th century onwards in eastern parts of Scandinavia, and slash-and-burn cultivation remained the major method of cereal crop cultivation in eastern Finland until the late 19th century (Soininen 1974; Taavitsainen 1987; Taavitsainen et al. 1998; Alenius and Laakso 2006; Alenius et. al. 2017). The timing of the shift towards agricultural communities suggested by the REVEALS results from eastern Finland agrees with the date range ad 400–1000 that is obtained genetically with ancient DNA at the time of the actual burst of animal husbandry in north-east Baltic Sea region (Niemi 2018). The rise and expansion of human activities from ca. ad 400 have further affected the regional plant composition, and in particular the evenness index. The regional climate-induced evenness change, i.e. high (6850–3450 bc) and decreasing evenness (3450 bc–ad 550), starts to increase from ad 500 until ad 1875. This increase in REVEALS evenness followed by a decline after ad 1875 is similar to the pattern reported by Fredh et al. (2017) in southern Sweden. Fredh et al. state that the highest evenness refers to the period of traditional cultural landscape while later evenness decrease refers to modern land use over the last century.
Conclusions
This article contributes to the discussion about Neolithic land use at local and regional scales in eastern Fennoscandia. The LRA allows us to compare the regional- vs. local-scale land use from fossil pollen records, and to underline main differences in the local land use between the study sites; the traditional use of pollen percentage/concentration data alone cannot achieve these objectives. Detailed, local scale investigations are therefore crucial in demonstrating the varieties of land use practices among hunting, fishing and gathering societies, and in providing a long-term background of human modifications and sustainable use of local resources.
In particular, we show that during the Early Neolithic, vegetation composition and fire activity were similar between the regional and local scales, mainly driven by natural and climate-induced factors. From ca. 4000 bc onwards, the trends in vegetation and fire dynamics start to differ between the regional and local scales, probably due to local land use being overshadowed at the regional scale by climate-induced factors. The LOVE model is therefore useful in pollen-based reconstruction of past vegetation to highlight the local land uses that are not visible through the REVEALS applications.
References
Abraham V, Novak J, Houfkova P, Petr L, Dudova L (2017) A landscape reconstruction algorithm and pedoanthracological data reveal late holocene woodland history in the lowlands of the NE Czech Republic. Rev Palaeobot Palynol 244:54–64
Ahola M, Kirkinen T, Vajanto K, Ruokolainen J (2018) On the scent of an animal skin: new evidence on Corded Ware mortuary practices in Northern Europe. Antiquity 92(361):118–131
Alalammi P (1988) Atlas of Finland, Folio 141: vegetation and flora. Geographical Society of Finland, Helsinki
Alenius T, Laakso V (2006) Palaeoecology and archaeology of the village of Uukuniemi, eastern Finland. Acta Borealia 23:145–165
Alenius T, Lavento M, Saarnisto M (2009) Pollen-analytical results from Lake Katajajärvi: aspects of the history of settlement in the Finnish Inland Regions. Acta Borealia 26:136–155
Alenius T, Mikkola E, Ojala AEK (2008) History of agriculture in Mikkeli Orijärvi, eastern Finland as reflected by palynological and archaeological data. Veget Hist Archaeobot 17:171–183
Alenius T, Mökkönen T, Lahelma A (2013) Early farming in the Northern Boreal zone: reassessing the history of land use in Southeastern Finland through high-resolution pollen analysis. Geoarchaeology 28:1–24
Alenius T, Mökkönen T, Holmqvist E, Ojala A (2017) Neolithic land-use in the Northern Boreal Zone: high-resolution multiproxy analyses from Lake Huhdasjärvi, south-eastern Finland. Veget Hist Archaeobot 26:469–486
Alenius T, Gerasimov D, Kuznetsov D et al (2020) Human-environment interaction during the Holocene along the Ancient Lake Ladoga: a case study based on palaeoecological and archaeological material from the Karelian Isthmus, Russia. Holocene, https://doi.org/10.1177/0959683620941071
Andersen ST (2012) The cultural landscape of megalithic tombs in Denmark, reconstructed by soil pollen analysis. Dan J Archaeol 1:39–44
Augustsson A, Gaillard M-J, Peltola P, Mazier F, Bergback B, Saarinen T (2013) Effects of land use and climate change on erosion intensity and sediment geochemistry at Lake Lehmilampi, Finland. Holocene 23:1,247–1,259
Barker G (2006) The agricultural revolution in prehistory: why foragers become farmers? Reprint 2009. Oxford University Press, Oxford
Behre K-E (1981) Interpretation of anthropogenic indicators in pollen diagrams. Pollen Spores 23:225–245
Bender Jørgensen L (1990) Stone age textiles in Northern Europe. In: Walton P, Wild JP (eds) Textiles in Northern Archaeology: NESAT III: textile symposium in York, 6–9 May 1987. Archetype Publications, London, pp 1–10
Bērzinš V (2008) Sārnate: living by a coastal lake during the East Baltic Neolithic. Acta Universitatis Ouluensis, Humaniora B86. Oulu University Press, Oulu
Björck S (1995) A review of the history of the Baltic Sea, 13.0-8.0 ka BP. Quat Int 27:19–40
Bläuer A, Kantanen J (2013) Transition from hunting to animal husbandry in Southern, Western and Eastern Finland: new dated osteological evidence. J Archaeol Sci 40:1,646–1,666
Bogucki P (1995) Prelude to agriculture in north-central Europe. In: Campana D (ed) Before Farming: Hunter-Gatherer Society and Subsistence. MASCA Research Papers in Science and Archaeology, Supplement to Vol 12. University of Pennsylvania Museum of Archaeology and Anthropology, Philadelphia, pp 105–116
Boivin N, Zedar M, Fuller DMD et al (2016) Ecological consequences of human niche construction: examining long-term anthropogenic shaping of global species distributions. PNAS 113(23):6,388–6,396
Cramp LJE, Evershed RP, Lavento M et al (2014) Neolithic dairy farming at the extreme of agriculture in northern Europe. Proc R Soc B 281:20,140,819
Crombé P, Robinson E (2014) 14C dates as demographic proxies in Neolithisation models of northwestern Europe: a critical assessment using Belgium and northeast France as a case-study. J Archaeol Sci 52:558–566
Cui Q-Y, Gaillard M-J, Lemdahl G, Stenberg L, Sugita S, Zernova G (2014) Historical land-use and landscape change in southern Sweden and implications for present and future biodiversity. Ecol Evol 4:3,555–3,570
Dambrine E, Dupouey JL, Laüt L, Humbert L, Thinon M, Beaufils T, Richard H (2007) Present forest biodiversity patterns in France related to former Roman agriculture. Ecology 88:1,430–1,439
Dubois N, Saulnier-Talbot Ė, Mills K et al (2017) First human impacts and responses of aquatic systems: a review of palaeolimnological records from around the world. Anthropocene Rev 5:28–68
Fredh D, Lagerås P, Mazier F, Björkman L, Lindbladh M, Broström A (2019) Farm establishment, abandonment and agrarian practices during the last 1,300 years: a case study from southern Sweden based on pollen records and the LOVE model. Veget Hist Archaeobot 28:529–544
Fredh D, Mazier F, Bragée P, Lagerås P, Rundgren M, Hammarlund D, Broström A (2017) The effect of local land-use changes on floristic diversity during the past 1000 years in southern Sweden. Holocene 27:694–711
Fuller D, Willcox G, Allaby R (2011) Cultivation and domestication had multiple origins: arguments against the core area hypothesis for the origins of agriculture in the Near East. World Archaeol 43:628–652
Fuller DO, Lucas L (2017) Adapting crops, landscapes, and food choices: Patterns in the dispersal of domesticated plants across Eurasia. In: Bolvin N, Crassard R, Petraglia M (eds) Human dispersal and species movement: from prehistory to present. Cambridge University Press, Cambridge, pp 304–331. https://doi.org/10.1017/9781316686942.013
Fyfe RM, de Beaulieu J-L, Binney H et al (2009) The European Pollen Database: past efforts and current activities. Veget Hist Archaeobot 18:417–424
Gaillard M-J, Kleinen T, Samuelsson P et al (2015) Causes of regional change: land cover. In: The BACC II Author Team (eds) Second assessment of climate change for the baltic sea basin. Regional Climate Studies. Springer, Cham, pp 453–477
Giesecke T, Bennett KD (2004) The Holocene spread of Picea abies (L.) Karst. in Fennoscandia and adjacent areas. J. Biogeogr. 31:1,523–1,548
Giesecke T, Basil Davis B, Brewer S et al (2014) Towards mapping the late Quaternary vegetation change of Europe. Veget Hist Archaeobot 23:75–86
Giesecke T, Brewer S, Finsinger W, Leydet M, Bradshaw RHW (2017) Patterns and dynamics of European vegetation change over the last 15,000 years. J Biogeogr 44:1,441–1,456
Gjerde JM (2010) Rock art and landscapes: studies of Stone Age rock art from northern Fennoscandia. PhD Dissertation, Department of Archaeology and Social Anthropology, University of Tromsø, p. 248
Grimm E (1987) CONISS: a FORTRAN 77 program for stratigraphically constrained cluster analysis by the method of incremental sum of squares. Comput Geosci 13:13–35
Halinen P (2015) Kivikausi. In: Haggren G, Halinen P, Lavento M, Raninen S, Wessman A (eds) Muinaisuutemme jaljet. Suomen esi-ja varhaishistoria kivikaudelta keskiajalle, Gaudeamus, Helsinki, pp 367–536
Hammarlund D, Björck S, Buchardt B, Israelson C, Thomsen CT (2003) Rapid hydrological changes during the Holocene revealed by stable isotope records of lacustrine carbonates from Lake Igelsjön, southern Sweden. Quat Sci Rev 22:353–370
Harris D (2007) Agriculture, cultivation and domestication: exploring the conceptual framework of early food production. In: Denham TP, Iriarte J, Vrydaghs L (eds) Rethinking agriculture: archaeological and ethnoarchaeological perspectives. Left Coast Press, Walnut Creek, pp 16–35
Harris DR, Fuller DQ (2014) Agriculture: definition and overview. In: Smith C (ed) Encyclopedia of global archaeology. Springer, New York, pp 104–113
Heikkilä M, Seppä H (2003) A 11,000 yr palaeotemperature reconstruction from the southern boreal zone in Finland. Quat Sci Rev 22:541–554
Heikkilä M, Seppä H (2010) Holocene climate dynamics in Latvia, eastern Baltic region: a pollen-based summer temperature reconstruction and regional comparison. Boreas 39:705–719
Heikkilä M, Edwards TWD, Seppä H, Sonninen E (2010) Sediment isotope tracers from Lake Saarikko, Finland, and implications for Holocene hydroclimatology. Quat Sci Rev 29:2,146–2,160
Hellman S, Gaillard M-J, Bunting JM, Mazier F (2009) Estimating the relevant source area of pollen in the past cultural landscapes of southern Sweden: a forward modelling approach. Rev Palaeobot Palynol 153:259–271
Herva V-P, Mökkönen T, Nordqvist K (2017) A northern NEOLITHIC? Clay work, cultivation and cultural transformations in the boreal zone of North-Eastern Europé, c. 5300–3000 BC. Oxf J Archaeol 36:25–41
Herva V-P, Nordqvist K, Lahelma A, Ikäheimo J (2014) Cultivation of perception and the emergence of the Neolithic world. Nor Archaeol Rev 47:141–160
Hjelle KL, Hufthammer AK, Bergsvik KA (2006) Hesitant hunters: a review of the introduction of agriculture in western Norway. Environ Archaeol 11:147–170
Hjelle KL, Mehl IK, Sugita S, Andersen GL (2015) From pollen percentage to vegetation cover: evaluation of the Landscape Reconstruction Algorithm in western Norway. J Quat Sci 30:312–324
Hjelle K, Sugita S (2012) Estimating pollen productivity and relevant source area of pollen using lake sediments in Norway: how does lake size variation affect the estimates? The Holocene 22(3):313–324
Huntley B, Birks HJB (1983) An atlas of past and present pollen maps for Europe: 0–13,000 years ago. Cambridge University Press, Cambridge
Innes JB, Blackford JJ, Rowley-Conwy PA (2013) Late Mesolithic and early Neolithic forest disturbance: a high resolution palaeoecological test of human impact hypotheses. Quat Sci Rev 77:80–100
Kaplan JO, Krumhardt KM, Gaillard M-J et al (2017) Constraining the deforestation history of europe: evaluation of historical land use scenarios with pollen-based land cover reconstructions. Land 6:91
Kirleis W, Klooß S, Kroll H, Müller J (2012) Crop growing and gathering in the norhern German Neolithic: a review supplemented by new results. Veget Hist Archaeobot 21:221–242
Kostrova SS, Meyer H, Bailey HL et al (2019) Holocene hydrological variability of Lake Ladoga, northwest Russia, as inferred from diatom oxygen isotopes. Boreas. https://doi.org/10.1111/bor.12385
Königsson L, Possnert G (1997) Pollen analysis covering the past 4000 radiocarbon years of cultural landscape of the Novgorod Area, Russia. In: Miller U, Clarke H (eds) Environment and vikings, scientific methods and techniques. Birka Studies 4. Riksantikvarieämbetet and Statens Historiska Museer, Stockholm, pp 73–89
Kuosmanen N, Seppä H, Reitalu T et al (2016) Long-term forest composition and its drivers in taiga forest in NW Russia. Veget Hist Archaeobot 25:221–236
Kuosmanen N, Marquer L, Tallavaara M et al (2018) The role of climate, forest fires and human population size in Holocene vegetation dynamics in Fennoscandia. J Veget Sci 29:382–392. https://doi.org/10.1111/jvs.12601
Lahelma A (2008) A touch of red: archaeological and ethnographical approaches to interpreting Finnish rock paintings. Iskos 15. Finnish Antiquarian Society, Helsinki
Lehtinen M, Nurmi PA, Rämö OT (2005) Precambrian geology of Finland. Key to the evolution of the Fennoscanidian Shield. Developments in Precambrian Geology 14. Elsevier Scientific Publication, Amsterdam
Lunkka JP, Johansson P, Saarnisto M, Sallasmaa O (2004) The Glaciation of Finland. In: Ehlers J, Gibbard PL (eds) Quaternary glaciations: extent and chronology developments in quaternary sciences 2, Part 1. Elsevier B.V., Amsterdam, pp 93–100
Mannering U, Gleba M, Bloch Hansen M (2012) Denmark. In: Mannering U, Gleba M (eds) Textiles and textile production in Europe: from prehistory to ad 400. Ancient Textiles Series 11. Oxbow Books, Oxford, pp 138–150
Marquer L, Gaillard M-J, Sugita S et al (2014) Holocene changes in vegetation composition in northern Europe: why quantitative pollen-based vegetation reconstructions matter. Quat Sci Rev 90:199–216
Marquer L, Gaillard-Lemdahl M-J, Sugita S et al (2017) Quantifying the effects of land use and climate on Holocene vegetation in Europe. Quat Sci Rev 171:20–37
Marquer L, Mazier F, Sugita S et al (2020) Pollen-based reconstruction of Holocene land-cover in mountain regions: evaluation of the landscape reconstruction algorithm in the Vicdessos valley, Northern Pyrenees, France. Quat Sci Rev 228:1,06,049
Mazier F, Broström A, Bragee P et al (2015) Two hundred years of land-use change in the south Swedish uplands: comparison of historical map-based estimates with a pollen-based reconstruction using the landscape reconstruction algorithm. Veget Hist Archaeobot 24:555–570
Mazier F, Gaillard M-J, Kuneš P, Sugita S, Trondman A-K, Broström A (2012) Testing the effect of site selection and parameter setting on REVEALS-model estimates of plant abundance using the Czech Quaternary Palynological Database. Rev Palaeobot Palynol 187:38–49
Mökkönen T (2010) Kivikautinen maanviljely Suomessa. Suomen Museo 116:5–38
Mökkönen T (2011) Studies on Stone Age housepits in Fennoscandia (4000–2000 cal BC): Changes in ground plan, site location, and degree of sedentism. Unigrafia, Helsinki
Myking T, Hertzberg A, Skrøppa T (2005) History, manufacture and properties of lime bast cordage in northern Europe. Forestry 78:65–71
Nielsen AB, Sugita S (2005) Estimating relevant source area of pollen for small Danish lakes around AD 1800. Holocene 15:1,006–1,020
Niemi M (2018) Genetic Reconstruction of Ancient Northern European Cattle and Sheep Populations. Faculty of Medicine, University of Helsinki, Unigrafia, Helsinki, Helsinki
Nordqvist K (2018) The Stone Age of North-Eastern Europe 5500–1800 calBC. Bridging the gap between the East and the West. Acta Universitatis Ouluensis B Humaniora 160. University of Oulu, Oulu
Nordqvist K, Herva V-P (2013) Copper use, cultural change and Neolithization in north-eastern Europe (c. 5500–1800 BC). Eur J Archaeol 16:401–432
Nordqvist K, Kriiska A (2015) Towards Neolithisation: The Mesolithic-Neolithic transition in the central area of the eastern part of the Baltic Sea. In: Kabaciński J, Hartz S, Raemaekers DCM, Terberger T (eds) The Dąbki Site in Pomerania and the Neolithisation of the North European Lowlands (c. 5000–3000 cal bc). Archaeology and History of the Baltic 8. Leidorf, Rahden/Westf, pp 537–556
Núñez M (1999) Role of food production in Stone Age Finland. Bidrag till kännedom av Finlands natur och folk 153. Helsinki
Oinonen M, Pesonen P, Tallavaara M (2010) Archaeological radiocarbon dates for studying the population history in eastern Fennoscandia. Radiocarbon 52:393–407
Oinonen M, Pesonen P, Alenius T et al (2014) Event reconstruction through Bayesian chronology: massive mid-Holocene lake burst triggered large-scale ecological and cultural change. Holocene 24:1,419–1,427
Ojala A, Alenius T (2005) 10 000 years of interannual sedimentation recorded in the Lake Nautajärvi (Finland) clastic-organic varves. Palaeogeogr Palaeoclimatol Palaeoecol 219:285–302
Ojala A, Alenius T, Seppä H, Giesecke T (2008) Integrated varve and pollen-based temperature reconstruction from Finland: evidence for Holocene seasonal temperature patterns at high latitudes. Holocene 18:529–538
Pesonen P, Leskinen S (2009) Pottery of The Stone Age hunter-gatherers in Finland. In: Jordan P, Zvelebil M (eds) Ceramics before farming. The dispercial of pottery among prehistoric Eurasian hunter-gatherers. Left Coast Press, Walnut Creek, pp 299–318
Pesonen P, Oinonen M, Carpelan C, Onkamo P (2012) Early Subneolithic ceramic sequences in eastern Fennoscandia: a Bayesian approach. Radiocarbon 54:661–676
Pini R, Ravazzi C, Raiteri L, Guerreschi A, Castellano L, Comolli R (2017) From pristine forests to high-altitude pastures: an ecological approach to prehistoric human impact on vegetation and landscapes in the western Italian Alps. J Ecol 105:1,580–1,597
Poska A, Saarse L (2006) New evidence of possible crop introduction to North-Eastern Europe during the Stone Age. Cerealia pollen finds in connection with the Akali Neolithic settlement. East Estonia Veget Hist Archaeobot 15:169–179
Poska A, Saarse L, Veski S (2004) Reflections of pre-and early-agrarian human impact in the pollen diagrams of Estonia. Palaeogeogr Palaeoclimatol Palaeoecol 209:37–50
Poska A, Saarse L, Koppel K, Nielsen AB, Avel E, Vassiljev J, Vali V (2014) The Verijarv area, South Estonia over the last millennium: a high resolution quantitative land-cover reconstruction based on pollen and historical data. Rev Paleobot Palynol 207:5–17
Power MJ, Marlon JR, Bartlein PJ, Harrison SP (2010) Fire history and the global charcoal database: a new tool for hypothesis testing and data exploration. Palaeogeogr Palaeoclimatol Palaeoecol 291:52–59
Prentice C (1985) Pollen representation, source area, and basin size: toward a unified theory of pollen analysis. Quatern Res 23:76–86
Pykälä J (2001) Maintaining biodiversity through traditional animal husbandry. In: The Finnish environment 495. Nature and Natural Resources Finnish Environment Institute, Vammala
Rasmussen P (1989) Leaf-foddering of Livestock in the Neolithic. Archaeobotanical evidence from Weier. Switzerland J Dan Archaeol 8:51–71
Reitalu T, Seppä H, Sugita S et al (2013) Long-term drivers of forest composition in a boreonemoral region: the relative importance of climate and human impact. J Biogeogr 40:1,524–1,534
Reynaud C, Hjelmroos M (1980) Pollen evidence and radiocarbon dating of human activity within the natural forest vegetation of the Pohjanmaa region (northern Finland). Candollea 35:257–304
Rimkutė V (2010) The Neolithic Mats of the Eastern Baltic Littoral. In: Andersson Strand E, Gleba M, Mannering U, Munkholt C, Ringgaard M (eds) North European Symposium for Archaeological Textiles 10. Ancient Textile Series 5. Oxbow Books, Oxford, pp 217–220
Roberts N, Fyfe RM, Woodbridge J et al (2018) Europe’s lost forests: a pollen based synthesis for the last 11,000 years. Sci Rep 8:716
Saarnisto M (1970) The Late Weichselian and Flandrian history of the Saimaa Lake complex. Soc Sci Fennica 37:1–107
Salonen JS, Luoto M, Alenius T, Heikkilä M, Seppä H, Telford R, Birks J (2014) Reconstructing palaeoclimatic variables from fossil pollen using boosted regression trees: comparison and synthesis with other quantitative reconstruction methods. Quat Sci Rev 88:69–81
Scherjon F, Bakels C, MacDonald K, Roebroeks W (2015) Burning the land. An ethnographic study of off-site fire use by current and historically documented foragers and implications for the interpretation of past fire practices in the landscape. Curr Anthropol 56:299–326
Schweingruber FH (1990) Microscopic wood anatomy; structural variability of stems and twigs in recent and subfossil woods from Central Europe, 3rd edn. Eidgenossische Forschungsanstalt WSL, Birmensdorf
Seppä H, Alenius T, Bradshaw RHW, Giesecke T, Heikkilä M, Muukkonen P (2009a) Invasion of Norway spruce (Picea abies) and the rise of the boreal ecosystem in Fennoscandia. J Ecol 97:629–640
Seppä H, Bjune AE, Telford RJ, Birks HJB, Veski S (2009b) Last nine-thousand years of temperature variability in Northern Europe. Clim Past 5:523–535
Simonen A (1980) Suomen kallioperä. 1:1 000 000. Geologian tutkimuskeskus, Espoo
Smith BD (2001) Low-level food production. J Archaeol Res 9:1–43
Soininen AM (1974) Vanha maataloutemme. Maatalous ja maatalousväestö Suomessa perinnäisen maatalouden loppukaudella 1720-luvulta 1870-luvulle. Historiallisia tutkimuksia 96. Suomen historiallinen seura, Helsinki
Stuart A, Ord K (1994) Kendall’s advanced theory of statistics, Vol 1: distribution theory, 6th edn. Edward Arnold, London
Sugita S (1994) Pollen representation of vegetation in quaternary sediments: theory and method in patchy vegetation. J Ecol 82:881–897
Sugita S (2007a) Theory of quantitative reconstruction of vegetation I: pollen from large sites REVEALS regional vegetation composition. Holocene 17:229–241
Sugita S (2007b) Theory of quantitative reconstruction of vegetation II: all you need is LOVE. Holocene 17:243–257
Sugita S, Gaillard M-J, Broström A (1999) Landscape openness and pollen records: a simulation approach. Holocene 9:409–421
Sugita S, Parshall T, Calcote R, Walker K (2010) Testing the landscape reconstruction algorithm for spatially explicit reconstruction of vegetation in northern Michigan and Wisconsin. Quat Res 74:289–300
Sutinen R, Teirilä A, Pänttäjä M, Sutinen M-L (2002) Distribution and diversity of trees species with respect to soil electrical characteristics in Finnish Lapland. Can J For Res 32:1,158–1,170
Sutton OG (1953) Micrometeorology. McGraw-Hill, New York
Taavitsainen J-P (1987) Wide-range hunting and swidden cultivation as prerequisites of Iron Age colonization in Finland. Suomen Antropologi 4:213–233
Taavitsainen J-P, Simola H, Grönlund E (1998) Cultivation history beyond the periphery: early agriculture in the North European Boreal Forest. J World Prehist 12:199–253
Tallavaara M, Pesonen P, Oinonen M (2010) Prehistoric population history in eastern Fennoscandia. J Archaeol Sci 37:251–260
Tauber H (1965) Differential pollen dispersion and the interpretation of pollen diagrams. Danmarks Geol Under II RÆKKE 89:1–69
Tolonen M (1978) Palaeoecology of annually laminated sediments in Lake Ahvenainen, S. Finland. I. Pollen and charcoal analyses and their relation to human impact. Ann Bot Fenn 15:177–208
Trondman A-K, Gaillard M-J, Mazier F et al (2015) Pollen-based quantitative reconstructions of Holocene regional vegetation cover (plant-functional types and land-cover types) in Europe suitable for climate modelling. Glob Chang Biol 21:676–697
Trondman A-K, Gaillard M-J, Sugita S et al (2016) Are pollen records from small sites appropriate for REVEALS model-based quantitative reconstructions of past regional vegetation? An empirical test in southern Sweden. Veget Hist Archaeobot 25:131–151
Tyrväinen A (1986) Pre-Quaternary rocks of the Vuohijärvi map-sheet area. Explanation to the maps of Pre-Queaternary rocks. Geological map of Finland 1:100 000. Geological Survey of Finland, Espoo
Ukkonen P (1996) Osteological analysis of the refuse fauna in the Lake Saimaa area. Helsinki Pap Archaeol 8:63–92
Ukkonen P, Mannermaa K (2017) Jääkauden jälkeläiset: Suomen lintujen ja nisäkkäiden varhainen historia. Museovirasto
Vanhanen S, Pesonen P (2015) Wild plant gathering in Stone Age Finland. Quat Int 30:1–13
Vuorela I (1981) The vegetational and settlement history in Sysmä, central south Finland, interpreted on the basis of two pollen diagrams. Bull Geol Soc Finland 53:47–61
Vuorela I, Saarnisto M, Lempiäinen T, Taavitsainen J-P (2001) Stone Age to recent land-use history at Pegrema, northern Lake Onega, Russian Karelia. Veget Hist Archaeobot 10:121–138
Waller M, Grant MJ, Bunting MJ (2012) Modern pollen studies from coppiced woodlands and their implications for the detection of woodland management in Holocene pollen records. Rev Palaeobot Palynol 197:11–28
Zanon M, Davis BAS, Marquer L, Brewer S, Kaplan JO (2018) European forest cover duroing the past 12,000 Years: a palynological reconstruction based on modern analogs and remote sensing. Front Plant Sci 9:253
Acknowledgements
The study was financed by the Academy of Finland through the Academy Research Fellow project “Land use, cultivation and animal husbandry during the Neolithic in North-Eastern Europe between c. 6000 and 1000 BC” granted to Teija Alenius (Project 274851). We are grateful to Marie-José Gaillard and two anonymous reviewers for their revisions and suggestions of an earlier version of the manuscript.
Funding
Open access funding provided by University of Turku (UTU) including Turku University Central Hospital.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by M.-J. Gaillard.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Alenius, T., Marquer, L., Molinari, C. et al. The environment they lived in: anthropogenic changes in local and regional vegetation composition in eastern Fennoscandia during the Neolithic. Veget Hist Archaeobot 30, 489–506 (2021). https://doi.org/10.1007/s00334-020-00796-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00334-020-00796-w