
Abstract
Purpose
To estimate the highest power output at which predominant energy contribution is derived from the aerobic system (aerobic limit power: ALP) and to compare ALP with the upper boundary of the severe intensity exercise domain.
Methods
Fifteen male individuals participated in this study. The upper boundary was estimated using i) linear relationship between time to achieve \(\dot{\text{V}}\)O2max and time to task failure (PUPPERBOUND), ii) hyperbolic relationships between time to achieve \(\dot{\text{V}}\)O2max vs. power output, and time to task failure vs. power output (PUPPERBOUND´), and iii) precalculated \(\dot{\text{V}}\)O2max demand (IHIGH). ALP was estimated by aerobic, lactic, and phospholytic energy contributions using \(\dot{\text{V}}\)O2 response, blood [lactate] response, and fast component of recovery \(\dot{\text{V}}\)O2 kinetics, respectively.
Results
ALP was determined as the highest power output providing predominant aerobic contribution; however, anaerobic pathways became the predominant energy source when ALP was exceeded by 5% (ALP + 5%) (from 46 to 52%; p = 0.003; ES:0.69). The \(\dot{\text{V}}\)O2 during exercise at ALP was not statistically different from \(\dot{\text{V}}\)O2max (p > 0.05), but \(\dot{\text{V}}\)O2max could not be attained at ALP + 5% (p < 0.01; ES:0.63). ALP was similar to PUPPERBOUND and PUPPERBOUND´ (383 vs. 379 and 384 W; p > 0.05). There was a close agreement between ALP and PUPPERBOUND (r: 0.99; Bias: − 3 W; SEE: 6 W; TE: 8 W; LoA: − 17 to 10 W) and PUPPERBOUND´ (r: 0.98; Bias: 1 W; SEE: 8 W; TE: 8 W; LoA: − 15 to 17 W). ALP, PUPPERBOUND, and PUPPERBOUND´ were greater than IHIGH (339 ± 53 W; p < 0.001).
Conclusion
ALP may provide a new perspective to intensity domain framework.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Severe intensity exercise is characterised by the attainment of maximal oxygen value (\(\dot{\text{V}}\)O2max) (Burnley and Jones 2007; Poole and Jones 2012; Poole et al. 2016). Thus, severe intensity exercise domain encompasses exercise intensities from maximal metabolic steady state, i.e., critical power (CP), to the highest power output at which \(\dot{\text{V}}\)O2max can be achieved (Hill et al. 2002; Caputo and Denadai 2008; Raimundo et al. 2019). Within the severe intensity exercise domain, time to achieve \(\dot{\text{V}}\)O2max is inversely related to exercise intensity (Margaria et al. 1965; Hill et al. 2002). In the extreme intensity exercise domain (intensities above the ‘upper boundary’ of the severe domain) task failure intervenes before \(\dot{\text{V}}\)O2max can be attained (Hill et al. 2002). Intrinsically, this upper bound represents a critical threshold for eliciting a combination of central and peripheral training adaptations based on \(\dot{\text{V}}\)O2 kinetics, EMG responses, etc. (Turnes et al. 2016b, a, c; Lisbôa et al. 2019; Norouzi et al. 2022). Furthermore, high-intensity interval training performed at the upper bound is effective not only in enhancing endurance performance but also in improving sprint performance (Turnes et al. 2016c). This synergistic effect develops both aerobic power and anaerobic capacity, thereby contributing to comprehensive physiological adaptations (Norouzi et al. 2022).
There are three essential approaches for assessing the upper boundary of the severe intensity exercise domain. One of these methods was proposed by Hill et al. (2002). In this method, the upper boundary is estimated by the relationship between the time to task failure (in x-axis) and time to achieve \(\dot{\text{V}}\)O2max (in y-axis), obtained from 3–4 exhaustive exercise performed within the severe intensity exercise domain. In this relation, the intersection of the projected line with the line of identity (i.e., y = x) provides an estimation for the exercise duration of the highest power output at which \(\dot{\text{V}}\)O2max can be attained momentarily (Fig. 1, panel A). Then, the external power associated with the upper boundary of the severe intensity exercise domain (i.e., PUPPERBOUND) is calculated by the point of the intersection between the hyperbolic power-time to task failure relationship and estimated time to task failure (Fig. 1, panel B). In addition, Hill and Ferguson (1999), Hill et al. (2002) and Hill et al., (2024) also reported that the upper boundary can be estimated by the intersection of the hyperbolic relationships between the time to achieve \(\dot{\text{V}}\)O2max vs. power output, and time to task failure vs. power output (i.e., PUPPERBOUND´ in present study) (Fig. 2). The upper boundary is associated with an exercise duration of 100–160 s (Hill et al. 2002, 2024; Hill and Stevens 2005).

Estimation of the upper boundary of the severe exercise domain as proposed by Hill et al. (2002). In panel A, the point of the intersection between the time to achieve \(\dot{\text{V}}\)O2max plotted against time to task failure and the line of identity demonstrates an exercise duration associated with the PUPPERBOUND. In panel B, the point of the intersection between the time to task failure plotted against power and estimated exercise duration is used to estimate the PUPPERBOUND. Data from a representative participant

The upper boundary of the severe exercise domain as the intersection of the hyperbolic relationships between the time to achieve \(\dot{\text{V}}\)O2max vs. power output (solid line) and time to task failure vs. power output (dashed line) as suggested by Hill and Ferguson (1999), Hill et al., (2002) and Hill et al., (2024). The projection of the intersection point of these two hyperbolic relationships provides an estimation for the time to task failure in y-axis and a power output in x-axis belonging to the upper boundary of the severe exercise domain (i.e., PUPPERBOUND´). Data from the same representative participant shown in Fig. 1
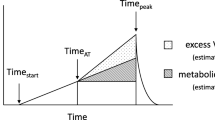
The third method for assessing the upper boundary of the severe intensity exercise domain was suggested by Caputo and Denadai (2008). In this method, the upper boundary is estimated by the data obtained from an exhaustive incremental exercise, and three further exhaustive exercise performed at constant work rates within the severe intensity exercise domain. Basically, the difference between the average of the highest \(\dot{\text{V}}\)O2 responses obtained from a total of four exercise and one typical error of measurement provides an attainable \(\dot{\text{V}}\)O2max value within the severe intensity exercise domain. Then, the highest exercise intensity that elicits the \(\dot{\text{V}}\)O2max is considered the upper boundary of the severe intensity exercise domain (i.e., IHIGH) (Fig. 3). Exercise duration at the IHIGH was reported as 2–3.5 min (Caputo and Denadai 2008).

Estimation of the upper boundary of the severe exercise domain based on the method proposed by Caputo and Denadai (2008). The panel A indicates the highest exercise intensity at which precalculated \(\dot{\text{V}}\)O2max demand could be achieved (i.e., IHIGH as the upper boundary), and the panel B shows the lowest extreme-intense exercise at which precalculated \(\dot{\text{V}}\)O2max demand could not be achieved (i.e., IHIGH + 5% as a constant work rate exercise performed 5% above the IHIGH). Dashed line represents the precalculated \(\dot{\text{V}}\)O2max demand calculated by subtracting one typical error of measurement from the average of the highest 15-s mean \(\dot{\text{V}}\)O2 values in panel A and B. Data from a representative participant
It is well-known that exercise resulting in task failure in less than ~ 120–130 s is supported by a predominant anaerobic contribution (Åstrand and Rodahl 1986; Serresse et al. 1988; Medbø and Tabata 1989; Bangsbo et al. 1990; Withers et al. 1991, 1993). It has also been shown that PUPPERBOUND, the highest work rate associated with the attainment of \(\dot{\text{V}}\)O2max, is associated with a tolerable exercise duration of 100 to 160 s (Hill et al. 2002, 2024; Burnley and Jones 2007; Poole and Jones 2012; Ozkaya et al. 2023). Thus, we hypothesised that the upper boundary of the severe intensity exercise domain, i.e., the upper end of the aerobic exercise zone, may be considered the highest exercise intensity at which predominant, i.e., 50%+, energy contribution is derived from the aerobic energy system (aerobic limit power: ALP), and ALP may denote a transition from the severe to extreme intensity exercise domain. If so, it should be noticed that exercise performed just above the ALP may provide a more appropriate exercise stimulus to enhance anaerobic capacity rather than improving aerobic power due to 50%+ anaerobic energy contribution and insufficient time spent at \(\dot{\text{V}}\)O2max. However, to date, ALP has yet to be compared with previous methods that are typically used to assess the upper boundary of the severe intensity exercise domain. Consequently, the aim of this study was to estimate the ALP and compare it with the PUPPERBOUND, PUPPERBOUND´ and IHIGH to understand whether the ALP could be a boundary that partitions severe from the extreme intensity exercise domain.
Materials and methods
Ethical approval
This study was approved by university ethics committee (19-12 T/60). Experimental procedures were designed according to the rules and principles of the Helsinki Declaration. After explaining the study’s procedures, risks, and benefits to all individuals, written informed consent was received from each participant using the approved guidelines and documentations.
Participants
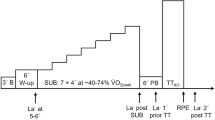
Fifteen physically active male individuals participated in this study (age: 31 ± 9 years; height: 1.77 ± 0.06 m; body mass: 70 ± 6 kg). They were required to visit the laboratory on 9–10 occasions over a 6- to 8-wk period with each visit separated by 24–72 h (Fig. 4). To ensure that circadian rhythm variance did not affect the results, the testing time was standardised (± 2 h) for each volunteer (Hill et al. 1992; Hill 2014). Over the course of the data collection, the participants did not alter their regular diet, sleep, or exercise habits. They were also instructed to refrain from any exhaustive exercise and to avoid drinking beverages containing alcohol or caffeine for 24 h before all trials. All participants had no history of systemic disease or injury and were not taking any medication during the study period.

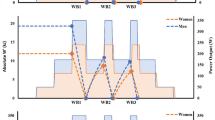
Schematic illustration of the experimental design. Over a period of 6–8-wk, each participant completed approximately nine excursions to the laboratory. These visits were separated by a 24–72-h rests. Following incremental exercise test session, each participant performed a series of constant work rate exercise tests until anaerobic energy metabolism became a major contributor. In this example, it seems that workload corresponding to 130% of PINC provides a 50%+ anaerobic contribution (i.e., it is performed within the extreme exercise intensity domain). PINC: power output corresponding the highest 15-s \(\dot{\text{V}}\)O2 response at the end of the incremental exercise, calculated using the equation proposed by Faina et al. (1997) (qq. Eq. 1)
Data collection
The tests were performed on an electromagnetically braked cycle ergometer (Lode Excalibur Sport, Groningen, the Netherlands). Pulmonary gas exchange data were measured by breath-by-breath (Cosmed Quark CPET, Rome, Italy). Before each exercise testing session, the gas analyser was calibrated using ambient air and gas mixture of known concentration (16% O2, 5% CO2, and balanced N2). The turbine flow meter was calibrated with a 3-L calibration syringe. The breath-by-breath data were filtered to remove occasional errant breaths caused by swallowing, coughing, and sighing. After removing data points positioned more than four standard deviations (SD) from local mean (i.e., 5-breath rolling mean) (Lamarra et al. 1987), the data were smoothed using a 5-breath moving average (Hill 2019). Capillary blood samples (20 μl) were collected from the finger prick and immediately analysed by an automated blood [lactate] analyser (Biosen C-line, EKF Diagnostics, GmbH, Barleben, Germany). Samples were taken at baseline and immediately after the termination of constant work rate exercise bouts, as well as at the first, third, fifth, and seventh min of recovery period. Resting \(\dot{\text{V}}\)O2 responses were evaluated for 10 min in a seated position before each constant work rate trial, while post-exercise \(\dot{\text{V}}\)O2 responses were taken throughout 30-min seated position just following the termination of exercise. All exercise tests were conducted in a well-ventilated laboratory, under standard conditions (~ 20 °C temperature, ~ 20.8% O2, ~ 0.05% CO2, and 50–60% relative humidity).
Procedures
Incremental tests
On their first visit to the laboratory, participants performed a multi-stage incremental exercise test. The tests commenced with 4 min of baseline cycling without resistance. Then, 0.5 W·kg−1 workload was applied to the system and increased every 3 min by 0.5 W·kg−1. The incremental test was terminated when participants allowed the cadence to fall below 70 rpm for more than 10 s, despite verbal encouragement. Power output corresponding the highest \(\dot{\text{V}}\)O2 response (PINC) was calculated using the equation proposed by Faina et al. (1997) (Eq. 1).
where P is the power output of the last step completed; ΔP is the increment of the power; and n is the number of seconds completed in the final stage.
Constant work rate exercise
Constant work rate exercise tests were initiated at 95% of PINC and the work rate was increased by 5% until anaerobic energy metabolism became a major contributor (50%+) on different days (qq. Fig. 4). Similar to the incremental exercise, the exercise tests commenced with 4 min of baseline cycling without resistance, then, workload was applied to the system. Each participant was encouraged to give maximum effort until the task failure. Exercise tests were terminated when participants allowed the cadence to fall below 70 rpm for more than 10 s. The highest 15-s \(\dot{\text{V}}\)O2 was recorded during tests.
Estimation of P UPPERBOUND
Data obtained from constant work rate exercise tests terminated between 2 and 10 min (i.e., approximately performed at 95%, 100%, 110%, and 125% of PINC) were used for the estimation of PUPPERBOUND. Time to achieve \(\dot{\text{V}}\)O2max was evaluated by nonlinear regression analysis (Sigma-Plot 14.0, Systat Software Inc., San Jose, CA, USA) using breath-by-breath \(\dot{\text{V}}\)O2 responses for each test (Eq. 2) (Hill et al. 2002; Hill and Stevens 2005).
where \(\dot{\text{V}}\)O2(t) is the \(\dot{\text{V}}\)O2 responses achievable at any time during exercise; \(\dot{\text{V}}\)O2baseline is the \(\dot{\text{V}}\)O2 obtained from the end of the baseline cycling period; Ap is the asymptotic amplitude of the primary phase which is the projected gain in \(\dot{\text{V}}\)O2; TDp is the time delay during primary phase; and τp represents the time constant which is related to 63% of the final amplitude in \(\dot{\text{V}}\)O2 responses.
In order to calculate the time to achieve \(\dot{\text{V}}\)O2max, data belonging to the first 20 s of exercise (i.e., cardio-dynamic phase) were excluded from the analyses (Krogh and Lindhard 1913; Weissman et al. 1982; Ozyener et al. 2001). The point at which the amplitude of the mono-exponential curve reached 99% was considered to represent the maximum value of the \(\dot{\text{V}}\)O2 response following the primary phase (Hill et al. 2002). Time to achieve \(\dot{\text{V}}\)O2max was evaluated as a value of 4.6 × τ. The point where the linear relationship (y = ax + b) between the time to achieve \(\dot{\text{V}}\)O2max and time to task failure intersects with the line of identity (y = x, i.e., time to achieve \(\dot{\text{V}}\)O2max = time to task failure) was accepted as an estimation for time to task failure (x = b × (1 − a)−1) of the upper boundary of the severe intensity exercise domain. For example, as shown in Fig. 1; x = 57.225 × (1 − 0.4336)−1 provided a time to task failure associated with PUPPERBOUND, i.e., 101 s for participant #9. Then, the point at which the hyperbolic relationship between the work rate and time to task failure intersects with the estimated exercise duration (i.e., 101 s) was considered the PUPPERBOUND, i.e., 458 W for participant #9 (Fig. 1) (Hill et al. 2002). On the other hand, as suggested by Hill and Ferguson (1999), Hill et al. (2002) and Hill et al. (2024), the upper boundary was also estimated as the point of the intersection of the hyperbolic relationships between the time to achieve \(\dot{\text{V}}\)O2max vs. power output (i.e., PUPPERBOUND = curvature constant × time to task failure–1 + CP) and time to task failure vs. power output (i.e., time to task failure = curvature constant × (PUPPERBOUND-CP)–1) (Fig. 2). Then, the projection of the intersection point of these two hyperbolic relationships provides an estimation for the time to task failure in y-axis and a power output (i.e., PUPPERBOUND´) in x-axis belonging to the upper boundary of the severe intensity exercise domain. For example, as shown in Fig. 2, PUPPERBOUND´ referred to 461 W and 104-s task failure for participant #9.
Estimation of I HIGH
Data obtained from incremental exercise and further constant work rate exercise tests performed at 95%, 100% and 110% of PINC were typically considered for the estimation of IHIGH (Caputo and Denadai 2008). \(\dot{\text{V}}\)O2max was calculated by subtracting one typical error of measurement from the average of the highest 15-s mean \(\dot{\text{V}}\)O2 values (Eq. 3). In order to calculate the typical error of measurement, the standard deviation of the highest 15-s mean \(\dot{\text{V}}\)O2 values obtained from incremental and constant work rate tests was divided by \(\sqrt{2}\) (Caputo and Denadai 2008).
where \(\dot{\text{V}}\)O2max is the difference between the average of the highest \(\dot{\text{V}}\)O2 values and one typical error of measurement; \(\dot{\text{V}}\)O2avg is an average of the highest 15-s \(\dot{\text{V}}\)O2 values of incremental exercise and constant work rate exercise tests performed at 95%, 100% and 110% of PINC; and \(\dot{\text{V}}\)O2sd is the standard deviation value of the mean.
Ultimately, participants implemented 2–4 further constant work rate trials until \(\dot{\text{V}}\)O2max could not be attained or maintained. The IHIGH was approved as the highest work rate and the shortest exercise duration providing the \(\dot{\text{V}}\)O2max.
Estimation of ALP
The ALP was estimated as the highest exercise intensity at which predominant energy contribution is derived from the aerobic energy system. The net energy contribution rates from aerobic, lactic, and phospholytic pathways were evaluated to estimate the ALP. The net energy contribution of the aerobic energy pathway (WAER) was evaluated by the difference between integrated exercise and resting \(\dot{\text{V}}\)O2 data over time (Eq. 4) (Åstrand 1981; Beneke et al. 2002; Bertuzzi et al. 2007; Ozkaya et al. 2014).
where t0 and t1 represent the time interval from the start to the end of the exercise; \(\dot{\text{V}}\)O2 is the total \(\dot{\text{V}}\)O2 responses during constant work rate exercise; \(\dot{\text{V}}\)O2resting is the \(\dot{\text{V}}\)O2 response during resting condition; and WAER is the net energy contribution of the aerobic energy pathway.
The net energy contribution of the phospholytic energy pathway (WPC) was assessed by the fast component of post-exercise \(\dot{\text{V}}\)O2 responses (Knuttgen 1970; Roberts and Morton 1978; Beneke et al. 2002; Bertuzzi et al. 2007; Ozkaya et al. 2014; Hill 2023). Post-exercise \(\dot{\text{V}}\)O2 response profile was evaluated using bi-exponential decay model (Eq. 5). The WPC was then calculated using the area under the curve of the fast component of post-exercise the \(\dot{\text{V}}\)O2 response (Eq. 6).
where \(\dot{\text{V}}\)O2(t) is the \(\dot{\text{V}}\)O2 responses achievable at any time during exercise; \(\dot{\text{V}}\)O2baseline is the \(\dot{\text{V}}\)O2 at baseline; Afc is the amplitude of the fast component of post-exercise \(\dot{\text{V}}\)O2 responses; τfc is the time constant of fast component of post-exercise \(\dot{\text{V}}\)O2 responses; TDfc is the time delay during the fast component; Asc is the amplitude of the slow component of post-exercise \(\dot{\text{V}}\)O2 responses; τsc is the time constant of slow component of post-exercise \(\dot{\text{V}}\)O2 responses; and TDsc is the time delay during the slow component.
where t0 and t1 represent the time interval from the peak \(\dot{\text{V}}\)O2 response of post-exercise recovery period to the end of the fast component; Afc is the amplitude of the fast component of post-exercise \(\dot{\text{V}}\)O2 responses; τfc is the time constant of the fast component of post-exercise \(\dot{\text{V}}\)O2 responses; TDfc is the time delay during the fast component; and WPC is the net energy contribution of the phospholytic energy pathway.
The net energy contribution of the lactic energy pathway (WLA) was calculated by multiplying body mass, peak delta (Δ) [lactate], O2-[lactate] equivalent, and the caloric equivalent of O2 (di Prampero 1981; Mader and Heck 1986; Beneke and Meyer 1997; Beneke et al. 2002; Bertuzzi et al. 2007; Ozkaya et al. 2014; Hill 2023). Peak Δ[lactate] was considered the difference between exercise and resting status. The accumulation of 1 mmol·L−1 of blood [lactate] was accepted as the equivalent to 3 mL·kg−1·mmol−1·L−1 (di Prampero 1981; di Prampero and Ferretti 1999). The contribution of each energy system was expressed in terms of mL·kg−1 and kJ, assuming the caloric quotient of 21.1 kJ·L−1. Total energy expenditure during constant work rate exercise (WTotal) was calculated as the sum of the energy derived from WAER, WPC, and WLA. Relative contributions of the energy systems were calculated as dividing the each of the WAER, WPC, and WLA value by the WTotal (WAER%, WPC%, and WLA%, respectively). Absolute and relative total anaerobic energy contributions (WANE and WANE%) were calculated as the sum of phospholytic and lactic contribution values. Note that the net energy contributions belonging to the ALP and IHIGH were directly calculated during constant work rate exercise tests; however, the net energy contributions of each energy system for the PUPPERBOUND and PUPPERBOUND´ were estimated by interpolated data obtained from the work rates just below and above the PUPPERBOUND or PUPPERBOUND´ (i.e., within the range of 5%), since the PUPPERBOUND and PUPPERBOUND´ was mathematically estimated.
Statistical analyses
Variables were evaluated using SPSS 19.0 (SPSS Inc., Chicago, USA). Shapiro–Wilk test was used to examine whether the data were normally distributed or not. A one-way repeated measures analysis of variance was used to assess the differences between \(\dot{\text{V}}\)O2 responses, work rates, time to task failures, absolute and relative energy contributions of 100% of PINC, PUPPERBOUND, PUPPERBOUND´, IHIGH, ALP, and work rate performed at 5% above the ALP (ALP + 5%). Pairwise comparisons were made with a Bonferroni adjustment. A Greenhouse-Geiser correction was used where sphericity was violated. Effect size (ES) values for repeated-measures ANOVA were calculated as eta squared (η2) and considered i) minimum effect size if 0.04 < η2 ≤ 0.25; ii) moderate effect size if 0.25 < η2 ≤ 0.64; and iii) strong effect size if η2 > 0.64 (Ferguson 2009). A Pearson product-moment correlation was computed to evaluate the relationship between PUPPERBOUND, PUPPERBOUND´ and IHIGH with the ALP. Additionally, standard error of estimation (SEE) and total error (TE) were used to examine the accuracy of estimations of the PUPPERBOUND, PUPPERBOUND´, IHIGH and ALP (Lohman 1981; Hopkins 2000). Bland–Altman plots were used to assess the limits of agreement (LoA) between the PUPPERBOUND, PUPPERBOUND´, IHIGH and ALP, and one-sample t-tests were used to determine whether the average difference between the values (i.e., Bias) was significantly different from zero (Bland and Altman 1986; Atkinson and Nevill 1998). Significance was set at p < 0.05. Results were reported as mean ± SD. According to the results of the post-hoc power analysis, based on a sample size of 15, alpha level of 0.05 and effect size of 0.84, the study had a sufficient statistical power (> 0.80) to detect meaningful differences (e.g., differences in work rates).
Results
The highest 15-s \(\dot{\text{V}}\)O2 average obtained from the 100% of PINC exercise (303 ± 48 W) was 54 ± 6 mL·min−1·kg−1, and there was no significant difference in \(\dot{\text{V}}\)O2 responses of 100% of PINC and ALP (p > 0.05). However, \(\dot{\text{V}}\)O2 response to ALP + 5% exercise was significantly decreased (p < 0.01; ES: 0.63). The mean values for work rates, \(\dot{\text{V}}\)O2 responses, and time to task failures obtained from the PUPPERBOUND, PUPPERBOUND´, IHIGH, ALP, and ALP + 5% are represented in Table 1.
The results and comparison of the markers showed that the PUPPERBOUND, PUPPERBOUND´ and ALP were similar in terms of both work rate and time to task failure values (p > 0.05), while the IHIGH was significantly underestimated PUPPERBOUND, PUPPERBOUND´ and ALP (p < 0.001; ES: 0.84). Note that the difference was not significant between PUPPERBOUND and PUPPERBOUND´, as expected (p > 0.05). Correlations, Bias, and LoA between the PUPPERBOUND, PUPPERBOUND´, IHIGH, and ALP are displayed in Fig. 5. According to the results, there were close agreements between ALP and PUPPERBOUND (r: 0.99; Bias: − 3 W; SEE: 6 W; TE: 8 W; LoA: − 17 W to 10 W), and ALP and PUPPERBOUND´ (r: 0.98; Bias: 1 W; SEE: 8 W; TE: 8 W; LoA: − 15 W to 17 W).

Correlation (A, C and E) and Bland–Altman analyses (B, D and F) between individual measures of the ALP and PUPPERBOUND, ALP and PUPPERBOUND´ and the ALP and IHIGH, respectively. In panels A, C and E, the solid lines represent the best-fit linear regression, and the dashed lines are the line of identity. In panels B, D and F, the solid lines represent the 95% limits of agreement, and the dashed lines are the mean difference between the two measures. PUPPERBOUND: the upper boundary of the severe exercise domain as the intersection of the relationship of the time to achieve \(\dot{\text{V}}\)O2max plotted against time to task failure and the line of identity as proposed by Hill et al. (2002); PUPPERBOUND´: the upper boundary of the severe exercise domain as the intersection of the hyperbolic relationships between the time to achieve \(\dot{\text{V}}\)O2max vs. power output and time to task failure vs. power output as suggested by Hill and Ferguson (1999), Hill et al., (2002) and Hill et al., (2024); IHIGH: the upper boundary of the severe exercise domain as the highest exercise intensity at which precalculated \(\dot{\text{V}}\)O2max demand can be achieved based on the method proposed by Caputo and Denadai (2008); ALP: the highest power output at which the predominant energy contribution is derived from the aerobic energy system; and SEE: standard error of estimate. Data based on group means
As expected, there was a greater absolute aerobic energy contribution at the IHIGH when compared to the PUPPERBOUND (p = 0.017), PUPPERBOUND´ (p = 0.009) or ALP (p = 0.006) (Table 2). However, there were similar absolute aerobic energy contributions for the ALP, PUPPERBOUND and PUPPERBOUND´ (p = 0.99). Otherwise, absolute contribution of aerobic energy system decreased significantly (Table 2), and therefore, relative anaerobic contribution became a predominant energy source (from 46 to 52%; p = 0.003; ES: 0.69) once the ALP was exceeded by 5%, i.e., during ALP + 5% exercise (Fig. 6).

Relative contribution of the aerobic and anaerobic energy system in overall energy expenditure belonging to the PUPPERBOUND, IHIGH, ALP, and ALP + 5% exercise. PUPPERBOUND: the upper boundary of the severe exercise domain based on the method proposed by Hill et al. (2002); IHIGH: the upper boundary of the severe exercise domain based on the method proposed by Caputo and Denadai (2008); ALP: the highest power output at which the predominant energy contribution is derived from the aerobic energy system; and ALP + 5%: constant work rate exercise performed 5% above the ALP. Note that calculated contribution rates of energy systems were quite similar between PUPPERBOUND and PUPPERBOUND´ (ES: 0.69 with < 0.9% differences between contribution rates) and not significantly different (p > 0.05). Data based on group means. a Significantly different from the IHIGH, p < 0.001. b Significantly different from the ALP + 5%, p < 0.01
Discussion
The aim of this study was to evaluate whether the highest exercise intensity at which predominant energy contribution is derived from the aerobic energy pathway (i.e., ALP) is a boundary that partitions the severe from extreme intensity exercise domain. Therefore, we compared the ALP to the PUPPERBOUND, PUPPERBOUND´ and IHIGH. To the best of our knowledge, the present study is the first to examine whether the ALP is associated with the upper boundary of the severe intensity exercise domain or not. Firstly, it should be noted that the ALP, as the upper end of the aerobic exercise zone, was sufficiently determined for each participant. While relative aerobic and anaerobic contribution rates of ALP intensity were 54% and 46% respectively, contribution rates were determined as 48% and 52%, once the ALP was exceeded by 5%, i.e., during ALP + 5% exercise. The principal novel findings demonstrated that the ALP was similar to the PUPPERBOUND and PUPPERBOUND´ in terms of both work rates and time to task failures. Indeed, there were close agreements between the ALP and PUPPERBOUND, and ALP and PUPPERBOUND´. Otherwise, ALP, PUPPERBOUND and PUPPERBOUND´ corresponded approximately 10–13% above the IHIGH, with a significant difference of 40–44 W between values (p < 0.001; ES: 0.84). In that case, it can be said that the ALP presents a novel approach to determine the upper boundary of the severe intensity exercise domain. Additionally, it offers a new perspective on the framework of exercise intensity domains, as it not only delineates the upper boundary of the severe intensity exercise domain but also indicates the upper end of the whole aerobic exercise zone including the moderate, heavy, and severe intensity exercise domains (Fig. 7).

Conceptual advance of exercise intensity domain framework. The upper schematic illustration demonstrates exercise intensity domains and their boundaries. The lower schematic illustration presents conceptual advance of exercise intensity domains which are characterised by aerobic and anaerobic predominance. It should be considered that the ALP delineates predominantly aerobic intensity domain from anaerobic domain. Thus, it can be said that task failure intervenes in less than 120 s before aerobic energy contribution becomes main energy source within the extreme exercise domain. LT: lactate threshold, CP: critical power, PUPPERBOUND: the highest power output at which \(\dot{\text{V}}\)O2max can be achieved, ALP: the highest power output at which predominant energy contribution is derived from the aerobic system, i.e., aerobic limit power
It is known that \(\dot{\text{V}}\)O2 amplitude is constrained during an extreme intensity exercise, and anaerobic ATP resynthesis is relied upon to compensate for the energy deficit. In this case, anaerobic contribution preserves the energy contribution from oxidative metabolism. Indeed, the results of the present study indicated that once the ALP was exceeded by 5% (i.e., ALP + 5%), relative anaerobic energy measure became a major contributor (from 46 to 52%; p = 0.003). According to the literature, there could be a few reasons for the constriction of \(\dot{\text{V}}\)O2 response and increasing the anaerobic energy metabolism within the extreme intensity exercise domain. One plausible explanation for the decrease in final \(\dot{\text{V}}\)O2 amplitude observed during an extreme intensity exercise is the increase in relative contribution of type II fibres to muscular force production (Bottinelli and Reggiani 2000; He et al. 2000; Wilkerson et al. 2004). Type II fibres possess a lower oxidative capacity compared to type I fibres but have a higher glycolytic capability. Consequently, type II fibres may inherently exhibit a lower \(\dot{\text{V}}\)O2 amplitude and/or be compelled to fulfil a larger proportion of the energy requirements through the anaerobic energy pathways during exercise. Closely related with the increase in type II fibre recruitment, another plausible explanation is the fall in pH resulting from the accumulation of by-product in anaerobic metabolism (Poole et al. 1988; Vanhatalo et al. 2016; Bergstrom et al. 2017). This process inhibits oxidative phosphorylation and restricts the \(\dot{\text{V}}\)O2 rise during exercise (Conley et al. 2001; Jubrias et al. 2003). Therefore, anaerobic energy contribution in total energy expenditure increases during a typical extreme-intense exercise. The final explanation is the blood flow restriction (Krustrup et al. 2004). Depending on the increase in the work rate, the local motor unit recruitment increases at the constant work rate exercise performed within the extreme intensity exercise domain (Thomas et al. 2016; Zhang et al. 2021), thus the blood flow is restricted (Krustrup et al. 2004). That limits muscle oxygen delivery and obligates a greater anaerobic energy contribution (Wilkerson et al. 2004). These physiological factors may be the sign that there is a close relationship between the highest exercise intensity at which the predominant energy contribution is derived from the aerobic energy system and the highest exercise intensity at which \(\dot{\text{V}}\)O2max can be achieved.
It should be noted that projected time to achieve \(\dot{\text{V}}\)O2max was greater than realised time to task failure at which power output performed at ALP + 5% in present study (122 ± 18 vs. 104 ± 11 s; p > 0.001). This may also be a simplistic explanation to support that task failure occurred before \(\dot{\text{V}}\)O2max can be achieved during exercise performed within the extreme intensity exercise domain (Ozkaya et al. 2023), and the ALP is identical to the highest power output at which \(\dot{\text{V}}\)O2max can be attained, i.e., the upper boundary of the severe intensity exercise domain. This evidence also supports that the upper end of the aerobic exercise zone coincides with the upper boundary of the severe intensity exercise domain. Once the upper end is exceeded, neither a predominant (50%+) aerobic contribution nor a \(\dot{\text{V}}\)O2max response is able to be obtained until task failure.
Hill et al. (2002) claimed that the greater the exercise intensity within the severe intensity exercise domain, the shorter the time to achieve \(\dot{\text{V}}\)O2max. Therefore, a unique work rate, at which \(\dot{\text{V}}\)O2max would be attained, should be existed at the end of the domain. In this concept, the PUPPERBOUND (or PUPPERBOUND´) provides the highest exercise intensity where \(\dot{\text{V}}\)O2max can be achieved momentarily just before task failure. Indeed, based on our results, the PUPPERBOUND and PUPPERBOUND´ have emerged as much more valid and plausible approaches compared to the IHIGH. However, PUPPERBOUND and PUPPERBOUND´ include some mathematical approaches and presuppositions. For example, in these methods, the time to achieve \(\dot{\text{V}}\)O2max values are originally calculated based on the 4.6 × τ value. Nevertheless, in some of our measurements, projected time to achieve \(\dot{\text{V}}\)O2max values were higher than realised time to task failures belonging to the same exercise. For instance, for participant #4, projected time to achieve \(\dot{\text{V}}\)O2max for 125% of PINC was 136 s, while realised time to task failure associated with constant work rate exercise performed at 125% of PINC was 117 s. This situation demonstrated that some individuals may have a slower (or faster) primary phase in \(\dot{\text{V}}\)O2 kinetics than expected. In the present study, the slope of the regression that provides the relationship between the time to achieve \(\dot{\text{V}}\)O2max and time to task failure could not across the line of identity in some measurements. Therefore, we did exclude data obtained from two of our study applicants. In this respect, the inability to estimate the upper bound for the severe intensity exercise domain in some participants weakens the method.
Another limitation for the PUPPERBOUND may be considered as R2 values of the relationships i.e., to engender the lower-than-expected R2 values (R2 < 0.85) of the relationship used to calculate the estimated time to task failure associated with the PUPPERBOUND. Indeed, for participant #3, R2 value of the slope of the regression line was 0.33, while the R2 values were found as 0.47 and 0.79 for participant #6 and participant #12, respectively. Indeed, Caputo and Denadai (2008) previously speculated that the relationship between the time to achieve \(\dot{\text{V}}\)O2max value and time to task failure obtained from severe intensity exercise may be highly affected by aerobic training status of participants, and this may adversary affect R2 values of the relationships. Even though all these individual differences and mathematical presuppositions, the PUPPERBOUND was highly correlated with the ALP, and both corresponded approximately 40–44 W above the IHIGH. Furthermore, the PUPPERBOUND is a non-invasive method, while the ALP requires invasive approaches.
A similar investigation on behalf of the upper boundary of the severe intensity exercise domain was conducted by Caputo and Denadai (2008). The IHIGH method bases on an interrogating \(\dot{\text{V}}\)O2max demand. According to the authors, the \(\dot{\text{V}}\)O2max demand implies a lower limit that must be strictly reached during exercise performed within the severe intensity exercise domain. However, calculated lower limit for \(\dot{\text{V}}\)O2max has not always been attained during constant work rate exercise performed within the domain. For example, for participant #15, VO2 responses obtained from the four exercise tests (i.e., incremental exercise, 95%, 100% and 110% of PINC) were 4.04, 3.92, 3.99, and 3.89 L·min−1, respectively. Then, calculated i) VO2 average ii) standard deviation, and iii) typical error of measurement can be determined as 3.96 L·min−1, 0.07 L·min−1, 0.05 L·min−1, respectively. Finally, based on the IHIGH method (qq. Eq. 3), VO2max demand can be calculated as 3.91 L·min−1. However, calculated VO2max could not be attained at 110% of PINC exercise for participant #15. Moreover, it is also suspicious whether to attain precalculated \(\dot{\text{V}}\)O2max value by a constant work rate exercise performed at 5% above the CP (i.e., CP + 5%, which provides the lowest exercise intensity within the severe intensity exercise domain) (Turnes et al. 2016b). Indeed, even in original study, authors reported that there was no expected time spent at \(\dot{\text{V}}\)O2max during an interval exercise session performed at CP + 5% (Turnes et al. 2016b). Nevertheless, CP + 5% is a typical exercise intensity which provides the longest time spent at \(\dot{\text{V}}\)O2max. It can be said that the precalculated VO2 demand may be higher than the real VO2 response and could not represent to all severe intensity exercise tests, due to its relatively rigid methodology used in reliability evaluations.
Similar to the lactate threshold (i.e., LT) which delineates the transition from moderate to heavy exercise (Burnley and Jones 2007) or the maximal metabolic steady state (i.e., CP) which demarcates heavy from severe intensity domain (Jones et al. 2019), the identification of the upper boundary of the severe intensity exercise domain which denotes a transition from the severe to extreme, is of paramount importance for fitness assessment and training prescription (Hill et al. 2002; Hill and Stevens 2005). Some studies suggest that prescribing exercise intensity based on a certain fractional usage (%) of power output corresponding to \(\dot{\text{V}}\)O2max may lead to highly heterogeneous physiological responses among individuals within the same exercise program (McLellan and Skinner 1985; Coyle et al. 1985; Meyer et al. 1999; Baldwin et al. 2000; Scharhag-Rosenberger et al. 2010; Lansley et al. 2011; Iannetta et al. 2020). Indeed, Inglis et al., (2024) recently mentioned that implementing exercise prescription based on the intensity domain framework can mitigate the confounding variability in the exercise intensity prescription, thereby reducing inter-individual differences in physiological and perceptual responses to training programs. Consequently, determining the power output that partitions the severe from extreme intensity exercise domain is imperative for normalising exercise intensity, and potentially minimising inter-individual variability in physiological and perceptual responses to exercise program.
Conclusion
Firstly, PUPPERBOUND (or PUPPERBOUND´) is a gold standard for the estimation of the upper boundary of the severe intensity exercise domain. It theoretically identifies the highest power output and the shortest exercise duration in which \(\dot{\text{V}}\)O2max can be attained. Therefore, PUPPERBOUND serves as a practical and physiological marker in exercise physiology, aiding both in the assessment of aerobic fitness and in the design of effective training regimens. Secondarily, ALP may provide a novel approach to understanding the significance of the upper boundary of the severe intensity exercise domain. Furthermore, it provides a new perspective to intensity domain framework since the ALP also indicates the upper end of the whole aerobic zone, i.e., moderate, heavy, and severe intensity exercise domains, and it may denote a transition from predominantly aerobic intensity zone to anaerobic intensity zone. Finally, coaches, athletes, or individuals should consider that the total anaerobic energy contribution increases and becomes predominant energy source within the total energy turnover compared to the aerobic rate during exercise performed just above that upper bound. In this case, the extreme intensity exercise might not provide a sufficient exercise induce to enhance aerobic power, and it is more appropriate to develop anaerobic performance related indices.
Data availability
Data generated and/or analysed during this study are available from the corresponding author upon reasonable request.
Abbreviations
- Afc :
-
Amplitude of the fast component
- ALP:
-
Aerobic limit power
- Ap :
-
Amplitude of the primary phase
- Asc :
-
Amplitude of the slow component
- CP:
-
Critical power
- ES:
-
Effect size
- IHIGH :
-
The upper boundary of the severe intensity exercise domain based on the method proposed by Caputo and Denadai (2008)
- LoA:
-
Limits of agreement
- LT:
-
Lactate threshold
- P:
-
Power output
- PINC :
-
Power output corresponding the highest \(\dot{\text{V}}\)O2 response measured during incremental tests
- PUPPERBOUND :
-
External power associated with the upper boundary of the severe intensity exercise domain as proposed by Hill et al. (2002)
- PUPPERBOUND′ :
-
External power associated with the upper boundary of the severe intensity exercise domain as proposed by Hill and Ferguson (1999), Hill et al. (2002) and Hill et al., (2024)
- SD:
-
Standard deviation
- SEE:
-
Standard error of estimation
- TDfc :
-
Time delay during the fast component
- TDp :
-
Time delay during primary phase
- TDsc :
-
Time delay during slow component
- TE:
-
Total error
- TTF:
-
Time to task failure
- \(\dot{\text{V}}\)O2 :
-
Pulmonary O2 uptake
- \(\dot{\text{V}}\)O2(t) :
-
\(\dot{\text{V}}\)O2 responses achievable at any time during exercise
- \(\dot{\text{V}}\)O2avg :
-
Average of the highest 15-s \(\dot{\text{V}}\)O2 values of incremental exercise and constant work rate exercise tests performed at 95%, 100% and 110% of PINC
- \(\dot{\text{V}}\)O2baseline :
-
\(\dot{\text{V}}\)O2 obtained from the end of the baseline cycling period
- \(\dot{\text{V}}\)O2max :
-
Maximal oxygen uptake
- \(\dot{\text{V}}\)O2resting :
-
\(\dot{\text{V}}\)O2 response during resting condition
- \(\dot{\text{V}}\)O2sd :
-
Standard deviation of the highest 15-s \(\dot{\text{V}}{\text{O}_{2}}\)values of incremental exercise and constant work rate exercise tests performed at 95%, 100% and 110% of PINC
- WAER :
-
Energy contribution of the aerobic energy pathway
- WANE :
-
Energy contribution of the anaerobic energy pathway
- WLA :
-
Energy contribution of the lactic energy pathway
- WPC :
-
Energy contribution of phospholytic energy pathway
- WTotal :
-
Total energy expenditure
- η2 :
-
Eta squared
- τfc :
-
Time constant of fast component
- τp :
-
Time constant of primary phase
- τsc :
-
Time constant of slow component
References
Åstrand PO (1981) Aerobic and anaerobic energy sources in exercise. In: di Prampero PE, Poortmans J (eds) Physiological Chemistry of Exercise and Training. Karger, Basel, pp 22–37
Åstrand PO, Rodahl K (1986) Physical training. In: Åstrand PO, Rodahl K (eds) Textbook of Work Physiology: Physiological Bases of Exercise. Human Kinetics, Champain IL
Atkinson G, Nevill AM (1998) Statistical methods for assessing measurement error (reliability) in variables relevant to sports medicine. Sports Med 26:217–238. https://doi.org/10.2165/00007256-199826040-00002
Baldwin J, Snow RJ, Febbraio MA (2000) Effect of training status and relative exercise intensity on physiological responses in men. Med Sci Sports Exerc 32:1648–1654. https://doi.org/10.1097/00005768-200009000-00020
Bangsbo J, Gollnick PD, Graham TE et al (1990) Anaerobic energy production and O2 deficit-debt relationship during exhaustive exercise in humans. J Physiol 422:539–559. https://doi.org/10.1113/jphysiol.1990.sp018000
Beneke R, Meyer K (1997) Walking performance and economy in chronic heart failure patients pre and post exercise training. Eur J Appl Physiol Occup Physiol 75:246–251. https://doi.org/10.1007/s004210050155
Beneke R, Pollmann C, Bleif I et al (2002) How anaerobic is the wingate anaerobic test for humans? Eur J Appl Physiol 87:388–392. https://doi.org/10.1007/s00421-002-0622-4
Bergstrom HC, Housh TJ, Cochrane-Snyman KC et al (2017) A model for identifying intensity zones above critical velocity. J Strength Cond Res 31:3260–3265. https://doi.org/10.1519/JSC.0000000000001769
Bland JM, Altman DG (1986) Statistical methods for assessing agreement between two methods of clinical measurement. The Lancet 327:307–310. https://doi.org/10.1016/S0140-6736(86)90837-8
Bottinelli R, Reggiani C (2000) Human skeletal muscle fibres: Molecular and functional diversity. Prog Biophys Mol Biol 73:195–262. https://doi.org/10.1016/s0079-6107(00)00006-7
Burnley M, Jones AM (2007) Oxygen uptake kinetics as a determinant of sports performance. Eur J Sport Sci 7:63–79. https://doi.org/10.1080/17461390701456148
Caputo F, Denadai BS (2008) The highest intensity and the shortest duration permitting attainment of maximal oxygen uptake during cycling: effects of different methods and aerobic fitness level. Eur J Appl Physiol 103:47–57. https://doi.org/10.1007/s00421-008-0670-5
Conley KE, Kemper WF, Crowther GJ (2001) Limits to sustainable muscle performance: interaction between glycolysis and oxidative phosphorylation. J Exp Biol 204:3189–3194. https://doi.org/10.1242/jeb.204.18.3189
Coyle EF, Coggan AR, Hopper MK (1985) Walters TJ (1988) Determinants of endurance in well-trained cyclists. J Appl Physiol 64:2622–2630. https://doi.org/10.1152/jappl.1988.64.6.2622
de Bertuzzi RC, M, Franchini E, Kokubun E, Kiss MAPDM, (2007) Energy system contributions in indoor rock climbing. Eur J Appl Physiol 101:293–300. https://doi.org/10.1007/s00421-007-0501-0
di Prampero PE (1981) Energetics of muscular exercise. Rev Physiol Biochem Pharmacol 89:143–222. https://doi.org/10.1007/BFb0035266
di Prampero PE, Ferretti G (1999) The energetics of anaerobic muscle metabolism: a reappraisal of older and recent concepts. Respir Physiol 118:103–115. https://doi.org/10.1016/s0034-5687(99)00083-3
Faina M, Billat V, Squadrone R et al (1997) Anaerobic contribution to the time to exhaustion at the minimal exercise intensity at which maximal oxygen uptake occurs in elite cyclists, kayakists and swimmers. Eur J Appl Physiol Occup Physiol 76:13–20. https://doi.org/10.1007/s004210050207
Ferguson CJ (2009) An effect size primer: A guide for clinicians and researchers. Prof Psychol Res Pract 40:532–538. https://doi.org/10.1037/a0015808
He ZH, Bottinelli R, Pellegrino MA et al (2000) ATP consumption and efficiency of human single muscle fibers with different myosin isoform composition. Biophys J 79:945–961. https://doi.org/10.1016/S0006-3495(00)76349-1
Hill DW (2014) Morning-evening differences in response to exhaustive severe-intensity exercise. Appl Physiol Nutr Metab 39:248–254. https://doi.org/10.1139/apnm-2013-0140
Hill DW (2019) Strategies to ensure accurate calculation of parameters of the V̇O2 response profile during heavy intensity cycle ergometer exercise. Int J Sport Stud Health. https://doi.org/10.5812/intjssh.98161
Hill DW (2023) Calculation of anaerobic capacity in running and cycling using post-exercise measures. Sci Sports 38:780–789. https://doi.org/10.1016/j.scispo.2022.04.004
Hill DW, Ferguson CS (1999) A physiological description of critical velocity. Eur J Appl Physiol 79:290–293. https://doi.org/10.1007/s004210050509
Hill DW, Stevens EC (2005) V̇O2 response profiles in severe intensity exercise. J Sports Med Phys Fitness 45:239–247
Hill DW, Borden DO, Darnaby KM et al (1992) Effect of time of day on aerobic and anaerobic responses to high-intensity exercise. Can J Sport Sci 17:316–319
Hill DW, Poole DC, Smith JC (2002) The relationship between power and the time to achieve V̇O2max. Med Sci Sports Exerc 34:709–714. https://doi.org/10.1097/00005768-200204000-00023
Hill DW, Glass LW, Vingren JL (2024) Critical speed, thresholds for V̇O2max and boundaries of the severe exercise intensity domain. Rev Bras Med Esporte. https://doi.org/10.1590/1517-8692202430012021_0311i
Hopkins WG (2000) Measures of reliability in sports medicine and science. Sports Med 30:1–15. https://doi.org/10.2165/00007256-200030010-00001
Iannetta D, Inglis EC, Mattu AT et al (2020) A critical evaluation of current methods for exercise prescription in women and men. Med Sci Sports Exerc 52:466–473. https://doi.org/10.1249/MSS.0000000000002147
Inglis EC, Iannetta D, Rasica L et al (2024) Heavy-, severe-, and extreme-, but not moderate-intensity exercise increase V̇O2max and thresholds after 6 weeks of training. Med Sci Sports Exerc. https://doi.org/10.1249/MSS.0000000000003406
Jones AM, Burnley M, Black MI et al (2019) The maximal metabolic steady state: redefining the “gold standard.” Physiol Rep 7:e14098. https://doi.org/10.14814/phy2.14098
Jubrias SA, Crowther GJ, Shankland EG et al (2003) Acidosis inhibits oxidative phosphorylation in contracting human skeletal muscle in vivo. J Physiol 553:589–599. https://doi.org/10.1113/jphysiol.2003.045872
Knuttgen HG (1970) Oxygen debt after submaximal physical exercise. J Appl Physiol 29:651–657. https://doi.org/10.1152/jappl.1970.29.5.651
Krogh A, Lindhard J (1913) The regulation of respiration and circulation during the initial stages of muscular work. J Physiol 47:112–136. https://doi.org/10.1113/jphysiol.1913.sp001616
Krustrup P, Söderlund K, Mohr M, Bangsbo J (2004) The slow component of oxygen uptake during intense, sub-maximal exercise in man is associated with additional fibre recruitment. Pflugers Arch 447:855–866. https://doi.org/10.1007/s00424-003-1203-z
Lamarra N, Whipp BJ, Ward SA, Wasserman K (1987) Effect of interbreath fluctuations on characterizing exercise gas exchange kinetics. J Appl Physiol 62:2003–2012. https://doi.org/10.1152/jappl.1987.62.5.2003
Lansley KE, Dimenna FJ, Bailey SJ, Jones AM (2011) A “new” method to normalise exercise intensity. Int J Sports Med 32:535–541. https://doi.org/10.1055/s-0031-1273754
Lisbôa FD, Raimundo JAG, Salvador AF et al (2019) Acute cardiopulmonary, metabolic, and neuromuscular responses to severe-intensity intermittent exercises. J Strength Cond Res 33:408–416. https://doi.org/10.1519/JSC.0000000000002130
Lohman TG (1981) Skinfolds and body density and their relation to body fatness: a review. Hum Biol 53:181–225
Mader A, Heck H (1986) A theory of the metabolic origin of “anaerobic threshold.” Int J Sports Med 7(Suppl 1):45–65. https://doi.org/10.1055/s-2008-1025802
Margaria R, Manglli F, Cuttica F, Cerretelli P (1965) The kinetics of the oxygen consumption at the onset of muscular exercise in man. Ergonomics 8:49–54. https://doi.org/10.1080/00140136508930773
McLellan TM, Skinner JS (1985) Submaximal endurance performance related to the ventilation thresholds. Can J Appl Sport Sci 10:81–87
Medbø JI, Tabata I (1989) Relative importance of aerobic and anaerobic energy release during short-lasting exhausting bicycle exercise. J Appl Physiol 67:1881–1886. https://doi.org/10.1152/jappl.1989.67.5.1881
Meyer T, Gabriel HH, Kindermann W (1999) Is determination of exercise intensities as percentages of V̇O2max or HRmax adequate? Med Sci Sports Exerc 31:1342–1345. https://doi.org/10.1097/00005768-199909000-00017
Norouzi M, Cabuk R, Balci GA et al (2022) (2022) Assessing acute responses to exercises performed within and at the upper boundary of severe exercise domain. Res Q Exerc Sport. https://doi.org/10.1080/02701367.2022.2117268
Ozkaya O, Colakoglu M, Kuzucu EO, Delextrat A (2014) An elliptical trainer may render the wingate all-out test more anaerobic. J Strength Cond Res 28:643–650. https://doi.org/10.1519/JSC.0b013e3182a20f77
Ozkaya O, Jones AM, Burnley M et al (2023) Different categories of V̇O2 kinetics in the “extreme” exercise intensity domain. J Sports Sci 41:2144–2152. https://doi.org/10.1080/02640414.2024.2316504
Ozyener F, Rossiter HB, Ward SA, Whipp BJ (2001) Influence of exercise intensity on the on- and off-transient kinetics of pulmonary oxygen uptake in humans. J Physiol 533:891–902. https://doi.org/10.1111/j.1469-7793.2001.t01-1-00891.x
Poole DC, Jones AM (2012) Oxygen Uptake Kinetics Compr Physiol 2:933–996. https://doi.org/10.1002/cphy.c100072
Poole DC, Ward SA, Gardner GW, Whipp BJ (1988) Metabolic and respiratory profile of the upper limit for prolonged exercise in man. Ergonomics 31:1265–1279. https://doi.org/10.1080/00140138808966766
Poole DC, Burnley M, Vanhatalo A et al (2016) Critical power: An important fatigue threshold in exercise physiology. Med Sci Sports Exerc 48:2320–2334. https://doi.org/10.1249/MSS.0000000000000939
Raimundo JAG, Turnes T, de Aguiar RA et al (2019) The severe exercise domain amplitude: a comparison between endurance runners and cyclists. Res Q Exerc Sport 90:3–13. https://doi.org/10.1080/02701367.2018.1549356
Roberts AD, Morton AR (1978) Total and alactic oxygen debts after supramaximal work. Eur J Appl Physiol Occup Physiol 38:281–289. https://doi.org/10.1007/BF00423111
Scharhag-Rosenberger F, Meyer T, Gässler N et al (2010) Exercise at given percentages of V̇O2max: heterogeneous metabolic responses between individuals. J Sci Med Sport 13:74–79. https://doi.org/10.1016/j.jsams.2008.12.626
Serresse O, Lortie G, Bouchard C, Boulay MR (1988) Estimation of the contribution of the various energy systems during maximal work of short duration. Int J Sports Med 9:456–460. https://doi.org/10.1055/s-2007-1025051
Thomas K, Elmeua M, Howatson G, Goodall S (2016) Intensity-dependent contribution of neuromuscular fatigue after constant-load cycling. Med Sci Sports Exerc 48:1751–1760. https://doi.org/10.1249/MSS.0000000000000950
Turnes T, de Aguiar RA, de Cruz RS, O, et al (2016a) Short-term interval training at both lower and higher intensities in the severe exercise domain result in improvements in on-kinetics. Eur J Appl Physiol 116:1975–1984. https://doi.org/10.1007/s00421-016-3449-0
Turnes T, de Aguiar RA, de Cruz RS, O, Caputo F, (2016b) Interval training in the boundaries of severe domain: effects on aerobic parameters. Eur J Appl Physiol 116:161–169. https://doi.org/10.1007/s00421-015-3263-0
Turnes T, de Aguiar RA, de Oliveira Cruz RS et al (2016c) High-intensity interval training in the boundaries of the severe domain: Effects on sprint and endurance performance. Int J Sports Med 37:944–951. https://doi.org/10.1055/s-0042-109068
Vanhatalo A, Black MI, DiMenna FJ et al (2016) The mechanistic bases of the power-time relationship: Muscle metabolic responses and relationships to muscle fibre type. J Physiol 594:4407–4423. https://doi.org/10.1113/JP271879
Weissman ML, Jones PW, Oren A et al (1982) Cardiac output increase and gas exchange at start of exercise. J Appl Physiol 52:236–244. https://doi.org/10.1152/jappl.1982.52.1.236
Wilkerson DP, Koppo K, Barstow TJ, Jones AM (2004) Effect of work rate on the functional “gain” of Phase II pulmonary O2 uptake response to exercise. Respir Physiol Neurobiol 142:211–223. https://doi.org/10.1016/j.resp.2004.06.001
Withers RT, Sherman WM, Clark DG et al (1991) Muscle metabolism during 30, 60 and 90 s of maximal cycling on an air-braked ergometer. Eur J Appl Physiol Occup Physiol 63:354–362. https://doi.org/10.1007/BF00364462
Withers RT, Van der Ploeg G, Finn JP (1993) Oxygen deficits incurred during 45, 60, 75 and 90-s maximal cycling on an air-braked ergometer. Eur J Appl Physiol Occup Physiol 67:185–191. https://doi.org/10.1007/BF00376665
Zhang J, Iannetta D, Alzeeby M et al (2021) Exercising muscle mass influences neuromuscular, cardiorespiratory, and perceptual responses during and following ramp-incremental cycling to task failure. Am J Physiol Regul Integr Comp Physiol 321:238–249. https://doi.org/10.1152/ajpregu.00286.2020
Acknowledgements
This study was the master thesis of Arda Peker completed under the supervision of Prof. Dr. Ozgur Ozkaya. The experiment complies with the current laws of the country in which they were performed. Arda Peker was honoured with the “Best Young Researcher” award at the 20th International Sports Sciences Congress held in Antalya in 2022 with this work. The authors extend their gratitude to the coaches, athletes, and all those who provided support and participated in this study.
Funding
Open access funding provided by the Scientific and Technological Research Council of Türkiye (TÜBİTAK). This study was financially supported by the Scientific and Technological Research Council of Turkiye (TUBITAK-1002 program; project code: 222S455).
Author information
Authors and Affiliations
Contributions
OO and AP contributed to the study conception and design. Material preparation, data collection and analysis were performed by AP, HA, EK, and GAP. The first draft of the manuscript was written by OO and AP, and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that there is no conflict of interest.
Additional information
Communicated by Jean-René Lacour.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Peker, A., As, H., Kaya, E. et al. The highest work rate associated with a predominantly aerobic contribution coincides with the highest work rate at which VO2max can be attained. Eur J Appl Physiol (2024). https://doi.org/10.1007/s00421-024-05533-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00421-024-05533-z