
Abstract
This study utilises multi-isotope approaches to investigate early medieval diet and childhood origins of individuals interred in an unusual group burial from Lothian, Scotland. In 1976, the skeletal remains of nine adults and five infants were unearthed from the infill of a latrine of a bathhouse at the Roman fort at Cramond, Edinburgh. Originally thought to be later medieval (14th/15th century), but recently dated to the 6th century AD, these remains represent a rare opportunity to gain deeper insights into the dietary histories and lifetime mobility of the inhabitants of early medieval south-central Scotland. Bone and teeth from the adults (n=9) were sampled, along with local faunal bone from a range of species (n=12). Long-term dietary trends were explored using stable carbon (δ13C), nitrogen (δ15N) and sulphur (δ 34S) isotope analyses of bone collagen, focusing on inter-individual variability and the potential inclusion of marine protein in the diet. Sulphur (δ 34S) was also utilised as a possible indicator of later-life mobility. Strontium (87Sr/86Sr) and oxygen (δ18OCARB) ratios of tooth enamel were employed to identify potential locals and non-locals, as well as possible locations of childhood origin. No intra-group dietary variability was detected, and the results are similar to other contemporary populations from southern Britain. The sulphur isotope data indicate that all of the individuals likely lived locally in their last few decades of life. However, based on isotopic data from tooth enamel, at least one or perhaps two of the individuals likely spent their childhoods in other locations.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
The determination of isotopic ratios in preserved archaeological human and faunal tissues permits access to a valuable pool of information directly related to the individual, such as diet and mobility (DeNiro 1981; Lee-Thorp 2008; Müldner et al. 2014; Schoeninger et al. 1983). In combination with archaeological contextual information and other bioarchaeological data, isotope analyses can shed light on varied aspects of past lifeways, from social stratification, through sex-based dietary differentiation to the extent of intra-group mobility (Britton 2017; Lee-Thorp 2008). Gaining access to such a wide range of information is of particular importance when addressing historically elusive time periods, where little may be gleaned about lifeways of the contemporary populations from primary literary sources.
Information available about the early centuries of historic Scotland is notably reliant on archaeological studies due to the scarcity of contemporary sources available about the era (Breeze 1994; Fraser 2009). These centuries mark important cultural and social transformations in the region, including the development of more pronounced hierarchical structures, struggles between conflicting political powers and the spread of new ideologies (Fraser 2009). Considering the generally conservative nature of dietary customs, a deeper understanding of diet in early medieval Scotland could in turn improve our understanding of inter- or intra-group transformations during such tumultuous times (Eriksson and Lidén 2013). In recent years, stable isotope studies focusing on diet in early medieval England and Wales have provided valuable new perspectives (Haydock et al. 2013; Hemer et al. 2017; Hughes et al. 2014; Mays and Beavan 2012), while ongoing isotope studies have begun to address the issue in a Scottish context (Britton et al. 2016; Curtis-summers et al. 2014).
There have been various stable isotope studies conducted in England to explore dietary change in the later Middle Ages (see Müldner 2016). Variations in dietary trends, especially with reference to marine fish consumption, were identified in a number of publications focusing on past populations of England (Mays 1997; Müldner and Richards 2007). In comparison, however, relatively few studies address dietary change in Scotland (Milek and Jones 2012: 29-35), with some notable exceptions (Curtis-summers et al. 2014; Lamb et al. 2012; Müldner et al. 2009; Willows 2016). Furthermore, even fewer studies have been undertaken to date to address the question of mobility in early historic Scottish populations (Lamb et al. 2012; Müldner et al. 2009). It is therefore crucial to carry out new research to enable the comparative study across early historic Britain.
Here, we present dietary (δ13C, δ15N, δ34S) and mobility (87Sr/86Sr and δ18OCARB) data from nine adult individuals (five females, four males) unearthed from a Roman bathhouse’s latrine at Cramond, Edinburgh. Originally believed to have come from the 14th century AD (Hukelová 2010), the assemblage has recently been radiocarbon-dated to the 6th–7th century AD (see Supplementary Material). Furthermore, the recent osteological re-evaluation of the remains found evidence of interpersonal violence demonstrated by cranial trauma in the case of four individuals, raising further questions about their identity (Boyle 2021). The aims of the study are: (i) to assess the dietary habits of the individuals interred at Cramond, particularly with regard to the consumption of marine fish, and explore potential inter-individual variability within early medieval Scotland and England through comparison with previously published datasets and (ii) to identify potential places of childhood origin for the Cramond individuals, and to explore the extent of personal mobility in early medieval south-central Scotland.
Background
Early Medieval Scotland—a time of colliding political attitudes and religious beliefs
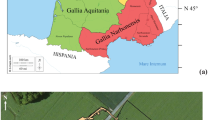
The first half of the 1st millennium AD saw a politically and ethnically divided Scotland. In the early Middle Ages, three main groups of people occupied the area (see Fig. 1): the Scotti in Dál Riata to the west, the Picts in most of northern Scotland and the Britons in the south (Foster 1996; Fraser 2009). From the northern territories, the Picts could sustain regular raids into the south, to the lands occupied by the various British tribes. It has previously been suggested that as a response, in the early 5th century the Britons invited Germanic tribes, referred to as Angles, Jutes and Saxons to fight against the northern threat (Foster 2014; Lowe 1999; Somerset-Fry 1982). Early discourse on this subject is often derived from documentary sources post-dating this event, such as references made by Bede to the Adventus Saxonum in his Historia ecclesiastica gentis anglorum (cited in Colgrave and Mynors 1969). While archaeological investigations have provided evidence supporting the arrival of people from the European continent in the 5th century AD, it should be noted that the impact and nature of this migration, as well as the process of the eventual development of a united identity, have been recognised as more complex than previously portrayed, and the model of the invasive replacement of the local population has been questioned (Hills 2017; Wood 2018). Thus, after the Roman withdrawal the region became characterised by power struggles and conflict, further exacerbated by the arrival and expansion of Anglian forces (Hayes 2005). Although the people of northern Britain did not represent a single political unit initially, by the sixth century the kingdoms of Goddodin, in present-day Lothian, and Strathclyde at Dumbarton Rock began to emerge (Foster 1996; Wormald 2005).

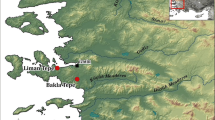
Map depicting the location of Cramond (above) and early medieval sites with published isotope data, including those discussed in this paper (left): Portmahomack, Westness and Bridge of Tilt in Scotland (Curtis-Summers et al. 2014; Czére et al. 2021; Barrett and Richards 2004), Auldhame (later early medieval) (Lamb et al. 2012) as well as sites in England (1—West Heslerton, 2—Lechlade Butler’s Field, 3—Berinsfield, 4—Tringhill Aston Clinton, 5—Dunstable Marina Drive, 6—Melbourn, 7—Barrington Edix Hill, 8—Lakenheath Eriswell, 9—Westgarth Gardens, 10—Coddenham, 11—Galey Hills, 12—Polhill, 13—Apple Down Compton, 14—Ford Laverstock, 15—Castledyke South, 16—Ipswich Buttermarket, 17—St Peter’s trip Broadstairs, 18—Finglesham, 19—Milhill Deal, 20—Dover Buckland) and Wales (21—Llandough, 22—Atlantic Trading Estate, 23—Porthclew, 24—Brownslade Barrow, 25—West Angle Bay, 26—St Patrick’s Chapel, 27—Ty Newydd, 28—Balladoole, 29—Cronk keeillane, 30—Peel Castle). Sites 1–14 are inland, 15, 16 are riverine and 17–20 are coastal (data published by Hannah et al. 2018; Leggett 2021; Mays and Beavan 2012; Privat et al. 2002); sites 21–22 are coastal sites located in southeast Wales, 23–26 in southwest Wales, 27–30 on the Isle of Man (data published by Hemer et al. 2017); and place names, major modern cultural centres in mid-6th century Scotland (after Foster 1996) and recorded kingdoms in England and Wales (after Carver 2019)
In addition to the politico-ethnic transformations of the time, Christianity began to influence the northern parts of Britain in the middle of the 1st millennium AD (Ritchie and Ritchie 1981; Wormald 2005). Latin inscriptions appearing on memorial stones, such as the Catstane near Edinburgh (c. 5 km away from the Roman Fort of Cramond) which was dated to the late 5th/early 6th century, evidence the spread of Christianity during this time (Ritchie and Ritchie 1981). These inscriptions are tied to the sub-Roman church of the time period. The earliest of the inscribed stones has been dated to c. 450 AD and is located in Whithorn, the site of the first recorded church in Scotland (Wormald 2005; Fraser 2009). Later historical sources suggest the southern areas of Scotland were introduced to the new belief system in the 6th century (Fraser 2009). The earliest upstanding churches of the country are much younger, however, than this period of conversion, largely dating to the 11th century or later. Therefore, in addition to the emergence of Latin inscriptions, changes in burial customs have also been suggested to relate to a changing belief system (Maldonado 2011).
The appearance and spread of East–West-oriented long-cist cemeteries are often argued to represent early Christian burials and therefore an early Christian influence (Lowe 1999; Ritchie and Ritchie 1981). Recent research into the burial practices in early medieval Scotland, however, suggests that the belief system and attitude towards death in the ‘Scottish’ early middle ages are more complex than previously thought (Maldonado 2011). Long-cist burials largely originate in the Iron Age, along with stone-lined graves and monumental burials, but only reached the peak of their popularity in the 5th–7th centuries AD (Lucy 2000; Maldonado 2013), with their use declining significantly afterwards. Their form and orientation were various, and so was the positioning of the body. East-facing supine burials became the norm gradually after the 5th century (Dunbar and Maldonado 2012; Maldonado 2013). Furnished early medieval burials are scarcely found in Scotland (Maldonado 2013). Furthermore, after the withdrawal of the Roman troops it was not uncommon to reuse Roman sites for burial purposes (Williams 1997). Roman masonry was often acquired to create stone-lined cists as evidenced in cemeteries at Thornybank (located in Midlothian), Hallow Hill (St Andrews) and Parkburn Quarry, Lasswade (Foster 2011; Henshall 1966-67; Maldonado 2013; Proudfoot 1996). The importance of the appropriate stonework and the geographical location of these burials is suggested to potentially signify familial ties or even territorial claims (Maldonado 2013).
Isotope approaches to diet and mobility in the early Middle Ages
The tumultuous nature of this time period, coupled with a general lack of domestic archaeological sites, makes the study of dietary customs and geographical origins of the people of early medieval Scotland all the more worthy of investigation. With regard to diet, zooarchaeological evidence suggests the importance of domesticates, in particular cattle (Bos taurus), during early medieval times in Scotland. In popularity, cattle were followed by pigs (Sus scrofa domesticus), sheep and goats (ovi-caprids) (Alcock 2003; Edwards and Ralston 2003). Agricultural production of barley and wheat became increasingly important as well (Gordon 1965; Härke 2002). There is little evidence, however, of the exploitation of marine food sources during this time period (Müldner 2016). In fact, the currently available published data suggest negligible fish consumption by early medieval communities throughout the British Isles, even at coastal locations (e.g. Barrett and Richards 2004; Curtis-summers et al. 2014; Hemer et al. 2017; Lamb et al. 2012; Leggett 2021; Mays and Beavan 2012). This is also evident, from faunal remains at least, throughout the late British pre-Roman Iron Age (Hambleton 2008; Sykes 2009). In fact, this has led some to even speculate that there may have been a contemporary cultural avoidance of marine fish (Dobney and Ervynck 2007), although the zooarchaeological record does indicate the presence of aquatic species at island Late Iron Age/ Early Pictish sites (Barrett et al. 1999).
The early middle ages were a period of mass migration throughout Europe, and historical evidence supports the arrival of new groups of people to Scotland from Ireland and the advancement of Anglian tribes towards Strathclyde (Foster 2014; Härke 2002; Webster et al. 1997). Contemporary archaeological evidence attests to population movement into England from the continent in the 5th–6th century (Harland and M. 2019; Hills 2017), which is further supported by recent isotope studies (e.g. Hughes et al. 2018; Leggett 2021); in Scotland, however, such evidence is scarce, and little is known about individual lifetime movement patterns. Without direct bioarchaeological studies of skeletal material from the time period, little is known about personal movement in early medieval Scotland and beyond (Härke 2002; Ritchie and Ritchie 1981). Cramond is well situated concerning sea and riverine routes, providing an ideal opportunity for trade and contact (Cessford 2001). Considering the location of the site and the nature of the time period, the presence of non-local individuals would not be unusual.
Isotopic methods can be useful in providing additional evidence for individual- and population-level dietary habits and movement histories, based on the presumption that food and water consumed during life leave a specific signature in the body which can be traced back to their input source, evidencing diet, mobility patterns and even the contemporary environment (Britton 2017; Lee-Thorp 2008). The δ13C, δ15N and δ34S values of bodily proteins such as bone collagen reflect the δ13C, δ15N and δ34S values of ingested dietary protein (DeNiro and Epstein 1978; DeNiro 1981; Richards et al. 2003; Schoeninger and DeNiro 1984), albeit with minor contributions from other dietary macronutrients (Froehle et al. 2010; Warinner and Tuross 2009). Different tissues offer different temporal resolution: bone collagen can indicate long-term (~10 years or more) dietary isotopic averages (Ambrose and Norr 1993; Hedges et al. 2007). Carbon (δ13C) and nitrogen (δ15N) values of bodily proteins reflect the main sources of dietary protein consumed and can be used in combination to aid the differentiation between marine and terrestrial protein sources in the diet (Chisholm et al. 1982; Schoeninger et al. 1983). Carbon isotopes are brought into the biological system by plants and bacteria, using atmospheric or marine sources of carbon. Since atmospheric CO2 is depleted in carbon 13C in comparison with carbon originating from oceanic sources, marine organisms have elevated levels of 13C (DeNiro 1985; Schoeninger and Moore 1992). Nitrogen isotope (δ15N) values of various organisms, on the other hand, differ based on their trophic level, increasing with ~3–5 ‰ with each step up the food chain, whereas δ13C values are enriched by ~1 ‰ (Bocherens and Drucker 2003). δ15N values can also be used to indicate freshwater or marine dietary inputs, as aquatic ecosystems tend to have longer food chains (Schoeninger et al. 1983). While a great many studies have successfully utilised these approaches to investigate whether or not marine foods were consumed amongst past populations, a number of other factors have also been demonstrated to influence environmental and bodily stable light isotope ratios. These include certain animal husbandry practices, coastal proximity, soil pH and rainfall, and well as personal physiology and some health conditions (see Bocherens and Drucker 2003; Reitsema 2013; Szpak 2014).
Sulphur isotope ratios (δ34S) within human and animal tissues also relate to the isotopic composition of ingested food. Terrestrial organisms tend to have δ34S ratios between -5 and +10 ‰; however, close proximity to the sea and saline environments can elevate these values (Nehlich et al. 2010). As sulphur isotope ratios depend on the local lithology, in addition to dietary information, it is possible to predict the sulphur isotopic values of different regions, allowing to identify individual mobility (Richards et al. 2003; Nehlich 2015).
Strontium isotope approaches in archaeology are based on the relationship between the strontium isotope signatures of bodily tissues (normally tooth enamel), that of food consumed during formation of that tissue and ultimately the strontium isotope signature of the underlying lithology. Through comparison with geological or geochemical ‘isoscape’ maps, strontium isotope ratios of skeletal tissues can then potentially be related to specific geographical locations (see review in Bentley 2006). The 87Sr/86Sr isotopic ‘signature’ of soils or plants at any location (‘bioavailable’ strontium) is primarily influenced by the parent material/underlying lithology, although other local environmental processes are also known to influence local bioavailable 87Sr/86Sr (Bentley 2006; Britton et al. 2020; Greene et al. 2004; Montgomery et al. 2007; Vitousek et al. 1999). Furthermore, it should be noted that, while there is no anticipated trophic level ‘offset’ in 87Sr/86Sr due to fractionation, recent controlled feeding studies have highlighted the potential influence of feeding habits on strontium isotope values, largely due to variation in Sr content of diet and differential digestion (Lewis et al. 2017; Lengfelder et al. 2019; Weber et al. 2020). Despite these uncertainties, strontium isotope analysis is proving an increasingly popular means of the investigation of past animal and human movements, especially as part of multi-isotope studies or when teamed with other techniques (see recent reviews Britton et al. 2021; Sillen 2021). In archaeological and palaeoecological studies, estimations of bioavailable 87Sr/86Sr at a particular location often rely on the sampling of modern vegetation or soils, cross-referenced with geological maps and/or utilising modelling, and strontium ‘isoscape’ maps now exist for a number of countries (e.g. Bataille et al. 2018; Snoeck et al. 2020), including the UK (Evans et al. 2010, 2018), and have even been produced on a global scale (Bataille et al. 2020).
In bioarchaeological studies, oxygen isotope analysis is often utilised alongside strontium as a secondary indicator of geographical origin (see Britton 2020 for an overview, and Groves et al. 2013; Hughes et al. 2014; Lamb et al. 2012; Müldner et al. 2009 for specific examples). The approach is based on the premise that δ18O values present in the tissues of living organisms are regulated mainly by the isotopic ratios of water consumed and (to a smaller extent) by food ingested during tissue formation (Daux et al. 2005, 2008; Luz et al. 1984). The δ18O values of precipitation are determined by climatic and environmental factors, primarily local temperature, but also altitude, humidity, and proximity to the coast (e.g. Dansgaard 1964; Gat 1980, 1996). Where local water, derived from precipitation, is consumed, δ18O values of mineralised tissues can therefore be utilised to infer individual movement histories (Pederzani and Britton 2019; e.g. Müldner et al. 2009) and also the degree of broader mobility within past populations (e.g. Pellegrini et al. 2016). In archaeological studies, oxygen isotope measurements are most commonly determined in dental enamel, including both the phosphate (PO4) and carbonate (CO3) groups. Although the phosphate phase is generally thought to be less susceptible to diagenetic changes, it has been demonstrated that carbonate groups from tooth enamel are reliable sources of δ18O levels due to the protection of the hard crystalline structure of enamel (Bryant et al. 1995; Chenery et al. 2012; Sponheimer and Lee-Thorpe 1999). Nonetheless, the influence of anthropogenic factors on oxygen isotope values must be noted and considered, such as the consumption of domesticate milk and boiled, stewed products (see discussion in Pederzani and Britton 2019). For example, Brettell et al. (2012) identified a potential 2.3 ‰ enrichment in tooth enamel oxygen composition associated with the regular consumption of ale, teas and stews.
Previous isotopic work from early medieval Britain has focused largely on sites in England (Groves et al. 2013; Hannah et al. 2018; Hughes et al. 2014; Mays and Beavan 2012; Privat et al. 2002), although new studies have also included sites from Wales (Hemer et al. 2017) and Scotland (Curtis-Summers et al. 2014; Lamb et al. 2012). Stable carbon and nitrogen isotope data of bone collagen published to date have revealed a rather homogenous diet, with very little intra-group variability and little evidence of sex-based differences in diet (e.g. Curtis-Summers et al. 2014; Privat et al. 2002). Amongst English populations, while marine fish consumption was nearly universally low, some variability with location has been observed—notably at riverine and coastal sites, where freshwater and marine fish, respectively, may have formed a minor component of the diet (Mays and Beavan 2012). Overall depleted stable carbon and nitrogen isotope values were noted in early medieval Hampshire, further supporting a level of regional variability (Mallet 2016). Dietary information obtained from early medieval Wales also indicates a strong reliance on terrestrial food; however, there was enrichment observed in stable carbon and nitrogen isotope data from southwest coastal Wales and the Isle of Man, which was suggested to be the result of local foddering techniques (Hemer et al. 2017). A lack of measurable marine fish consumption was also evident in Scottish studies, including at 7th–10th Century Auldhame (Phase 1) (Lamb et al. 2014); amongst early Pictish (6–7th century) individuals at Portmahomack (Curtis-Summers et al. 2014) and Pictish burials at Westness (Barrett and Richards 2004) and Blair Atholl (Czére et al. 2021). Elevated nitrogen isotope ratios, in combination with ‘terrestrial’ carbon isotope values, were determined in individuals at Auldhame, Scotland, and accredited to either the consumption of freshwater fish or omnivorous protein, or indeed to soil improvement methods, which may have served to enrich local soils (Lamb et al. 2012). Strontium and oxygen isotope works at Auldhame were largely indicative of a local, static population, aside from two individuals which originated from later Medieval phases of the site. In England, a similar lack of intra-group variability during these early centuries in strontium and oxygen isotope ratios was determined at Berinsfield suggesting a local population, albeit with one outlier of possible continental European origin identified (Hughes et al. 2014). However, tentatively non-local individuals were identified at sites contemporary with the ‘migration period’ into England, at Ringlemere Farm in East Kent (Brettell et al. 2012), and Wasperton in Warwickshire (Montgomery et al. 2009), possibly associated with north-west European connections. In a more recent study, Leggett (2021) identified a large number of individuals at Finglesham, Kent, with oxygen isotope values not consistent with the local range, some possibly influenced by different dietary practices. In contrast, isotopic evidence for movement (at least within Scotland and possibility further afield) was determined in the analyses of only a limited number of Pictish burials (e.g. Czére et al. 2021; Parker Pearson et al. 2018; Walther et al. 2016). Revealing both shared inter-population trends across Britain and also some more local nuances, isotope studies are thus proving to yield valuable insights into early medieval lifeways. Further studies, particularly in Scotland—given the complex sociopolitical backdrop of the period—and at coastal locations, are, however, required.
The burial site
Background
Cramond is situated to the south of the Firth of Forth estuary, in close proximity to the River Almond (Fig. 1). Archaeological evidence suggests that the area was occupied as early as the mid-9th Millennium BC (Lawson et al. 2020). The Roman Fort at Cramond to which the bathhouse was attached was established during the Antonine invasion of Scotland in 142AD as an outlying fort ‘defending’ the eastern flank of the Antonine Wall between it and Inveresk Roman Fort to the East. The fort continued to be occupied until the withdrawal of Roman forces in the 160s AD to Hadrian’s Wall. However, the strategic significance of the site was demonstrated when it was reoccupied acting as a major supply and military base during the early 3rd century (208-c.211AD) campaigns of Septimius Severus (Holmes et al. 2003). During both periods, the bathhouse was reoccupied and rebuilt.
Whilst the fort was again abandoned at the end of the Severan campaigns in Scotland around 211/212 AD, the site’s importance appears to have continued. Due to its geographical position, Cramond is well situated concerning sea routes which would have provided excellent opportunity for trade and contact. The discovery of a Celtic-style pin dated to the 3rd to 4th century AD from the bathhouse along with Byzantine coins of emperor Justinian (AD 527–565) and possible structural remains attest to the post-Severan use of the fort at Cramond (Cessford 2001; Holmes et al. 2003). The reuse of Roman structures has also been recorded throughout England, with a particular concentration of such sites in the South (Bell 2005). This custom has been suggested to utilise the abandoned structures as a reflection of status (Gerrard 2007). Although no definitive conclusions can be made about the shifting nature of the fort’s occupation, these stray finds of post-Roman coinage and the use of a former Roman structure point towards not only a continuous human presence in the vicinity of Din Eidyn (Edinburgh), but the significance of the site as well. Recent excavations provided further evidence of the continued occupation and importance of the fort between the 7th and 10th century AD, as well as human remains from an Anglian cemetery at the site (Cook et al. in prep).
The Roman latrine: excavation, archaeology and inhumations
Cramond’s Roman fort has been the subject of archaeological investigations since the 1950s most notably by Alan & Viola Rae (Rae et al. 1974) and by the City of Edinburgh Council’s Archaeology Service between 1975 & 1981 (Holmes et al. 2003). The latter programme of investigations was instigated by the chance discovery of the fort’s bathhouse in 1975 during an extension of a carpark located to the north of the fort adjacent to the shore. During the 1976 excavations of the bathhouse, the remains of nine adult individuals and five infants were found in the latrine attached to the northern side of its frigidarium (Fig. 2).

Position of the individual skeletons in the latrine of the Roman bathhouse (image: City of Edinburgh Council 2014)
Much of the original structure of the bathhouse had been destroyed by medieval and later occupation of the site and apparent stone robbing activities carried out in the 17th century. The upper deposits of the latrine including the human remains were recorded as secondary at the time containing both dismembered / disturbed human remains intermixed with medieval midden material similar to deposits overlying the bathhouse site as a whole. Consequently, due to the lack of separation amongst the remains and their close proximity to a layer of 14th-century pottery sherds, the burial was originally assigned to this era (Hukelová 2010). Recently, however, eight out of the nine human skeletons were radiocarbon dated using tooth samples, and it has been established that all these individuals belong to the 6th and 7th centuries AD, consistent with a rapid placement of these remains (see Supplementary Material, Table S1 and Fig. S1). Faunal remains were also found in the upper layers amongst large amounts of unstratified medieval and post-medieval pottery, likely displaced from the nearby village of Cramond (Barnetson 2003; Will 2003). The faunal remains are most likely not contemporary with the human remains but represent waste material from the later medieval period (Hukelová 2010). Zooarchaeological analysis of the faunal remains was undertaken by one of the authors (OCz). A large number of water voles (arvicola amphibious) in the assemblage and the presence of amphibian remains indicate that the area might have been affected by various flood episodes, which could have caused residual water to remain in this location. In terms of domestic species, ovi-caprids seem to have been dominant (by NISP), along with other domesticates such as cattle, pig and chicken (Fig. 3).

Faunal remains uncovered from the latrine detailing number of identified specimens (NISP)
The human remains
The skeletal remains were generally in good condition, albeit partly disarticulated. Due to the post-cranial elements having been mixed post-excavation, these have been treated as disarticulated deposits. The crania and mandibles, however, were associated with their original burial numbers and have been used for further studies (Boyle 2021). Five skulls have been identified as female or probable female and four as male or probable male. Age at death ranged from young adult (18–25 years) up to older adult (45+ years) (see summary in Table 1; Hukelová 2010; Museum of Edinburgh 2016). The uppermost remains displayed evidence of erosion and weathering, which suggests that for a period of time they were exposed, possibly as a consequence of the disturbance by stone robbers and the subsequent destruction of the latrine’s wall. This event also might have caused the disarticulation of some of the skeletal remains. The presence of well-preserved perinatal remains in the latrine has been suggested to indicate that this was not a case of a reburial, which would have impacted their preservation, but a hasty inhumation potentially following a dramatic event (Barnetson 2003). In the absence of radiocarbon dates associated with the perinates at this time, however, it cannot be excluded that these remains may belong to an earlier Roman phase of deposition (Boyle 2021). Due to the nature of the burial, it was not possible to associate the perinatal remains with particular adult individuals. Evidence of interpersonal violence was seen on the crania of at least four individuals. Cranium 1, an adult male aged 26–35 years, had a healed sharp-force injury on the right parietal and a possible depressed linear fracture on the posterior portion of the same bone. Peri-mortem sharp-force trauma was identified on the right parietal of cranium 9, also an adult male aged 26–35 years. Cranium 5, a young adult female aged 18–25 years, had suffered a severe peri-mortem blunt-force injury to the right side of her head. Cranium 6, an older adult female aged upwards of 46 years, had an ossified haematoma on the right parietal, possibly the results of a blow to the head and a possible healed fracture of the body of the right mandible.
Materials and Methods
Materials
Cranial bone was sampled from the nine adult individuals ahead of collagen extraction and the determination of stable carbon, nitrogen and sulphur isotope ratios. Although the bones sampled were not possible to identify to the specific cranial element in the case of three of the individuals (CR76-C-4, -5, and -6), a fragment from the left parietal was obtained from CR76-C-1, the sphenoid bone from CR76-C-2, the right temporal in the case of CR76-C-3, while the right frontal bone was sampled from individual CR76-C-7, and the left zygomatic and parietal from individuals CR76-C-8 and -9, respectively. In order to conduct strontium and oxygen isotope measurements, tooth enamel from second molars (M2) was sampled in the case of six individuals (CR76-C-1, -3, -4, -5, -7, -9) and from first molars (M1) from two further individuals (CR76-C-6, -8). Permanent first molars start to mineralise around the time of birth, and between the ages of 2 and 4 years in the case of the second premolars and the second molars. There is, however, some inter-individual variation involved (Scheuer and Black 2004; Hillson 1996). As M2 teeth begin mineralising at a later stage, these teeth were preferentially chosen for the purpose of this study, in order to avoid the ‘nursing effect’ which causes a trophic-level enrichment from mother to child, elevating the 18O levels of the latter (Britton et al. 2015). No teeth could be sampled from CR76-C-2 due to ante mortem tooth loss. In addition to enamel samples, the roots of two second molars (from CR76-C-1 and -7) were obtained for bulk strontium isotope analysis, as the roots of teeth—assumed to be affected by diagenetic alteration—can be a useful additional indication of strontium isotope ratios in local soils/the burial environment (Montgomery et al. 2007; Britton et al. 2011). Based on biosphere sampling and underlying lithologies, the expected 87Sr/86Sr values based in Scotland could range from 0.707 to 0.722. Cramond lies atop sedimentary bedrock originating in the Carboniferous Period and the local bioavailable Sr isotope values are expected to range between 0.709 and 0.711 (see Fig. 4 and Evans et al. 2018). Modern drinking water values in Scotland range from -4.5 to -8.5 ‰, with Cramond lying in the -7.0 to -8.0 ‰ region (see Fig. 4 and Darling et al. 2003).

Bone samples from the faunal skeletal material uncovered from the latrine were also sampled for carbon, nitrogen and sulphur isotope analysis, in order to provide a local ‘baseline’ for the interpretation of human diet. While the faunal assemblage from Cramond is likely associated with the later medieval pottery found at the site, and is thus not contemporary to the human remains (Barnetson 2003; see above), it can still be useful in providing an indication of local environmental isotope ratios. Bones of multiple species were chosen for the stable isotope analysis, including terrestrial herbivores—cattle (Bos Taurus), ovicaprids (Ovi-capridae), horse (Equidae), red deer (Cervus elaphus), water vole (Avricola amphibious); omnivores/carnivores – pig (Sus scrofa domesticus), chicken (Gallus gallus), fox? (Vulpes vulpes?), dog (Canis familiaris); and marine fish—cod (Gadidae). Where it was possible to identify two different individuals belonging to the same species, both were selected for analysis (Table 1).
Methods
Bone samples were prepared in the Archaeological Chemistry Laboratories of the Department of Archaeology, at the University of Aberdeen. Cranial bone from the nine adult human individuals was sampled for bulk bone collagen extraction and isotope analysis, along with bone of various skeletal elements from fauna from the site (n=12). Bone samples were cleaned using a tungsten carbide burr, and trabecular bone was removed during this process due to its susceptibility to diagenetic changes (Pye 2004).
Collagen was extracted using a modified Longin method (Collins et al. 2002; Longin 1971) with the addition of an ultrafiltration step by Brown et al. (1988), as described in Britton et al. (2012). After the demineralisation of the samples in 0.5 M HCl, they were gelatinised in 48 hours at 70 0C, in a ph3 HCl solution. Using Ezee® filters, the liquid section of the solution was isolated and subsequently ultrafiltered employing >30kDa filters to separate the fractions larger and smaller than 30kDa. Although both of these sections frozen and lyophilised (in order to calculate collagen yield), the sample fractions >30kDa were weighed out into tin capsules for analysis. The stable carbon and nitrogen analysis of the samples was performed at the Department of Archaeology, University of Reading, UK, using a Europa 20-20 IRMS with a SerCon elemental analyser using internal reference materials calibrated to international standards (IAEA-CH-6, -CH-7, -N1, -N2, USGS40). An analytical error of ± 0.2 ‰ (1σ) or better was established by repeat analysis of an internal collagen standard. Sulphur stable isotope analyses were analysed at Iso-Analytical Ltd, Cheshire, UK, using EA-IRMS. For quality control purposes test samples of IA-R061, IAEA-SO-5 (barium sulphate, δ 34SV-CDT = +0.50 ‰) and NBS-1577B (bovine liver, δ 34SV-CDT = +7.50 ‰) were measured as in-house quality control checks. Vanadium pentoxide (9-11 mg) was added to weighed collagen samples ahead of sulphur isotope analysis. The analytical error was ± 0.2 ‰ (1σ) or better.
Tooth enamel was removed from teeth as a strip the length of the tooth crown, from the enamel-root junction to occlusal surface (buccal surface, where possible) using a diamond coated rotary saw at the University of Aberdeen. Dentine was mechanically separated from enamel using a tungsten carbide burr, and enamel samples (plus additional dentine samples from two individuals) were then ultrasonicated in ultrapure Milli-Q water and air-dried. The enamel and two additional dentine samples were then transferred to the NERC Isotope Geosciences Laboratory (NIGL) (now National Environmental Isotope Facility), Keyworth, UK, for further processing and isotope analysis, and samples were split for strontium and oxygen isotope analysis. In line with recommendations in Chenery et al. (2012) and Montgomery et al. (2007), no chemical pre-treatment was undertaken prior to oxygen and strontium isotope analyses, respectively, in line with many published studies (e.g. Evans et al. 2019; Leach et al. 2009). For strontium isotope analysis, subsamples of enamel and dentine samples were cleaned in ultrapure water and high-purity acetone, before being weighed into pre-cleaned Teflon beakers. Samples were then mixed with a 84Sr tracer solution and dissolved in high-purity nitric acid (8 M HNO3). Strontium was collected using Sr-spec resin columns (Eichrom AG50 X8) and then loaded onto single rhenium filaments following the method of Birck (1986). The isotope composition and concentrations were measured using determined by thermal ionisation mass spectrometry (TIMS). The international standard for 87Sr/86Sr, NBS987, gave a value of 0.710250 ± .000006 (n=8, 2σ) during the analysis of these samples. Blank values were in the region of 100pg (Evans and Stewart 2014).
The aliquot of enamel reserved for carbonate oxygen isotope analysis was ground to a fine powder using an agate mortar and pestle. Subsamples of powder were then loaded into glass vials, sealed and evacuated on a hot block set at 90 °C on the GV Multiprep system. CO2 was evolved from samples through reaction with anhydrous phosphoric acid (collection time 14 min) and transferred to a GV IsoPrime dual inlet mass spectrometer. The isotope values are treated as a carbonate (δ18O per mil) normalised to the PDB scale using a within-run calcite laboratory standard (KCM) calibrated against SRM19 and NIST reference material. These values were then converted to the VSMOW scale using the published conversion equation of Coplen (1988). Reproducibility, based on the analysis of in-house analytical reproducibility based on within run standards (KCM and an in-house enamel standard) for δ18OVSMOW was ±0.3 ‰ or better. Measured carbonate oxygen isotope data were then converted using equations established by Chenery et al. (2012) in order to predict the the δ18O signature of consumed drinking water.
Statistical analyses
Shapiro–Wilk test was performed on the stable carbon and nitrogen isotope data to assess the normality of dataset (Shapiro and Wilk 1965). Due to the sample size, a nonparametric Kruskal–Wallis test was performed to compare male and female dietary data. The significance level was set at 5%. These tests were performed using R version 4.0.0.
Results and Discussion
The results of all carbon, nitrogen and sulphur isotopic analyses conducted on the material from Cramond can be viewed in Table 1, and the strontium and oxygen isotope measurements are detailed in Table 2. The full isotopic dataset, including carbonate δ13C measurements, is available in the Supplementary material.
Carbon, nitrogen and sulphur isotope data: diet
As shown in Table 1, the %C, %N, % S and C:N, N:S, C:S of the majority of the samples that were included in this study, human and faunal, meet quality indicators (Ambrose 1990; van Klinken 1999; Nehlich and Richards 2009), confirming the integrity of the collagen extracted. While %C, %N and C:N ratios of the 9 human and 12 faunal samples are acceptable, the sulphur isotopic data from the chicken and cod sample analysed, however, are excluded from further discussion due to their C:S and N:S ratios, which did not meet quality indicators (Nehlich and Richards 2009). As shown in Table 1 and in Fig. 5, the δ13C values of the human bone collagen included in this study range from -21.4‰ to -20.7‰, with a mean at -21.1 ‰ ± 0.3 (1 s.d.) (n=9). The stable nitrogen isotope ratios are between 10.8 ‰ and 11.9 ‰, with a mean at 11.4 ‰ ± 0.4 (1 s.d.) (n=9). The values of δ 34S of the human remains range from 13.2 ‰ to 15.2 ‰ with a mean at ± 0.7 (1 s.d.) (n=9), and sulphur measured in faunal samples ranges from 12.8 ‰ to 15.6 ‰.

δ13C and δ15N ratios of bone collagen extracted from the human and faunal remains from Cramond
A wider range of variation in both carbon and nitrogen isotope values can be observed amongst the faunal material with the collagen extracted from herbivores (cattle, ovicaprids, horse, red deer) having δ13C values between -22.6 ‰ and -21.9 ‰ and δ 15N from 4.7 ‰ to 7.7 ‰ (n=6). The omnivorous/carnivorous species (chicken, pig, dog, fox) sampled range from -21.1‰ to -20.3 ‰, and 10.6 to 12.1 ‰ (n=4), for carbon and nitrogen isotope ratios, respectively, which is consistent with their trophic level and terrestrial context. The δ13C and δ 15N values of the cod sample are enriched in the heavier isotopes in comparison with those of the terrestrial animals, consistent with their marine origin and trophic position. The faunal data seem to be consistent with their expected trophic-level values noted in other publications based on contemporary sites (Barrett et al. 2011; Curtis-Summers et al. 2020; Leggett et al. 2021) (Fig. 6).

δ 13C and δ 34S ratios of bone collagen extracted from the human and faunal remains from Cramond
The δ13C and δ15N values obtained from the bone collagen of the individuals inferred at Cramond indicate that their dietary protein was largely terrestrial in origin, most likely consisting of domestic animal meat or secondary products, such as milk. This is suggested by the fact that the carbon isotopic values are all below -20 ‰. In addition, the nitrogen isotope values of 10.8 ‰ to 11.9 ‰ reflect a reliance on protein gained from local fauna, as the humans are enriched by ~4.8 ‰ relative to the herbivore average. Both stable carbon and nitrogen isotope data were proven to be normally distributed (δ13C (‰): W = 0.91563, p-value = 0.3573; δ15N (‰): W = 0.91563, p-value = 0.3573), and the results of the Kruskal–Wallis tests (δ13C (‰): chi-squared = 2.6, df = 1, p-value = 0.1069; δ15N (‰):chi-squared = 0.54454, df = 1, p-value = 0.4606) did not indicate a statistically significant pattern of dietary variation between the δ13C and δ15N ratios of males and females within this dataset.
Although contemporary data from sites from England are similar to the results obtained from Cramond and other sites from Scotland (i.e. not evidencing measurable marine fish consumption), the latter are somewhat enriched in 15N in comparison with the early medieval populations from England (e.g. Hannah et al. 2018; Leggett 2021; Privat et al. 2002; see Fig. 7). This may be the result of a slightly elevated ‘baseline’ soil and plant δ15N values in Scotland, relative to England, resulting in a small enrichment in 15N in the bone collagen of animals, and ultimately humans, regardless of trophic level. This can be observed in the enrichment of the fauna of Cramond in comparison with those from the inland 5th–7th century site of Berinsfield. Indeed, topsoils from arable lands and improved grasslands in Scotland have heavily enriched 15N values (Thornton et al. 2015). Improvement, and subsequent soil enrichment, was identified as a possible cause behind elevated δ15N values and lower ‘terrestrial’ δ13C in human bone collagen at early medieval Auldhame in Scotland (Lamb et al. 2012). Due to the coastal location of Cramond, sea spray could also affect δ15N values (Ambrose 1991). Soil enrichment has been noted as the potential cause of the elevated stable carbon and nitrogen isotope values detected in south west coastal Wales and the Isle of Man (Hemer et al. 2017). Intriguingly, these data most closely correspond to values measured in early medieval skeletal material in this and other Scottish studies. The data from the south-east coast of Wales, on the other hand, are more similar to the early medieval riverine assemblage from England. The exploitation of riverine resources, such as fish or wildfowl, may also explain these slightly elevated values (Nehlich and Richards 2009). This may be supported by the observed offset between human and herbivore bone collagen δ15N at Cramond (~4.8 ‰), which indicates the regular consumption of high trophic level food sources in this population. Indeed, slight elevation in δ15N values has also been determined in human bone collagen at riverine early medieval sites in England (Mays and Beavan 2012), which may suggest the importance of riverine resources at certain locales during this period, in particular in the case of lower status individuals (Privat et al. 2002). Sulphur stable isotope analysis can be a useful secondary indicator of the origin of dietary protein and has been used to identify the consumption of riverine fish in archaeological populations (Nehlich et al. 2010). Controlled feeding studies indicate that the sulphur isotopic ratios of terrestrial organisms tend to reflect sulphur values between -5 and 10 ‰; however, close proximity to the sea and saline environments can elevate these ratios (Nehlich 2015; Richards et al. 2003). This ‘sea-spray effect’ is the result of the redeposition of marine sulphur from sea salt sulphates and can reach as far as 30 km inland, while being further influenced by the local prevailing wind (Coulson et al. 2005; Nehlich 2015). Freshwater organisms are varied and can exhibit values ranging from −5 or lower to 10–14 ‰ (Nehlich and Richards 2009; Nriagu et al. 1991), and sulphur isotope values above 14 ‰ in mammalian bodily proteins are generally consistent with a significant marine influence within the diet (Nehlich 2015). All human values in this study range from 13 to 15 ‰ (see Fig. 6), which could indicate either some freshwater or marine component to the diet. However, terrestrial fauna measured in this study display similar values, including most of the herbivores studied. This likely indicates that these elevated values may instead be due to the ‘sea spray effect’ (Nehlich 2015; Richards et al. 2003), reflecting the proximity of the site to the sea.

Strontium and oxygen isotope data: mobility
The 87Sr/86Sr ratios of tooth enamel from the Cramond individuals are between 0.7077 and 0.7126. Cramond lies atop the carboniferous sedimentary rocks of the Strathclyde group, suggesting the local bioavailable strontium values range between 0.709 and 0.711 (see Fig. 4) (British Geological Survey 2006; Evans et al. 2010). The carboniferous coalfields located to the east of Edinburgh, however, reflect more radiogenic strontium isotope ratios of 0.711–0.712. The majority of enamel samples, with the exception of two individuals (CR76-C-1, CR76-C-8), align with the local signal. Strontium concentrations range from 46 to 157 ppm, which is within the expected range of human enamel samples from Britain (Evans et al. 2012). Measurements of dentine sampled from two individuals are within the local range (0.7098 and 0.7095) (Table 2, Fig. 8), with elevated Sr concentrations (306 and 405 ppm, respectively), suggesting their equilibration with the local burial environment.

Oxygen isotope data in Table 2 include both measured values (δ18OCARB) from enamel carbonate, and predicted δ18O drinking water values for each of the individuals, calculated using the methods described in Chenery et al. (2012: 309). The measured δ18OCARB values for the individuals range from 26.0 to 27.2 ‰ with a mean at 26.4 ‰ ± 0.4. The predicted oxygen isotopic values of drinking water (δ18Odw) of the human samples range from -7.4 to -5.5 ‰. Again, for the majority of individuals, measured δ18O values are consistent with the estimated local oxygen isotopic range based on the data produced by Darling et al. (2003), with the exception of CR76-C-8 (see Fig. 8).
The results of 87Sr/86Sr and the predicted δ18O values seen in Fig. 8 reveal that six of the individuals display ratios that correlate well with the predicted local geological and hydrological values, suggesting a local origin. Two of the individuals—CR76-C-1(adult male) and CR76-C-8 (older adult female)—however, are prominent outliers. Although CR76-C-1 displays oxygen values consistent with the anticipated local range (~-7 ‰ to -8 ‰), the 87Sr/86Sr values are significantly more radiogenic than those of the other individuals at the site. While the enamel sample from CR76-C-8 was enriched in 18O relative to the other individuals, it must be noted that this datum has been obtained from a first permanent molar, which due to its early formation period could reflect a breastfeeding signal-induced enrichment (Pederzani and Britton 2019; Wright and Schwarcz 1998). While this cannot be excluded, no such enrichment (relative to the other individuals in the study) was observed in the case of individual CR76-C-6, where a first molar was also analysed. It is also possible that certain culinary practices, such as the regular consumption of brewed or boiled liquids (e.g. Brettell et al. 2012; Lengfelder et al. 2019, see discussion in Pederzani and Britton 2019), could result in such enrichment. However, given that, in the case of CR76-C-8, elevated oxygen values are also coupled with a significantly lower strontium isotope ratio than any other individuals in the study, it is perhaps most likely that these data are instead consistent with the individual, a female, having spent at least a portion of their childhood (i.e. the period of M1 tooth formation) in a different region of the UK. Such depleted strontium values are relatively rare in the UK and are only found in a small number of areas. The lowest 87Sr/86Sr values in Scotland (0.7078) have been recorded in the Inner Hebrides and are associated with Tertiary volcanic geology (Montgomery et al. 2003; Evans et al. 2012). Furthermore, the low bioavailable strontium values detected in northeast Northern Ireland along the Lower Basalt Formation of Co. Antrim (0.706896 ± 0.000372) (Snoeck et al. 2020) would also be consistent with such depleted 87Sr/86Sr data. Although smaller patches of younger igneous rocks can be found closer by (e.g. around Inverclyde and East Renfrewshire) (British Geological Survey 1993), given the elevated oxygen value determined in this individual, a westerly origin is supported. The most parsimonious interpretation of these data may therefore indicate that this individual spent their childhood on Scotland’s West Coast, with both strontium and oxygen isotope ratios being consistent with the Isles of Skye, Canna and Rum. Alternatively, the Northern Isles of Shetland (i.e. Hunsta, Fetlar, Yell) could also represent a potential area of origin in the case of this individual. It should be noted that small areas of west Wales, Cornwall and Somerset England could also match with such depleted strontium and elevated oxygen values (Evans et al. 2018).
CR76-C-1, a young male with a robust skull and a healed sharp forced trauma, can also be identified as an outlier. This male individual reflects elevated bioavailable strontium isotope ratios, which can be found around ~30 km south and south east of Cramond, as well as further south west in Peeblesshire and southern Lanarkshire, along the Silurian sedimentary bedrock of the Gala group dominating the Southern Uplands of Scotland. The oxygen isotope values of this individual are similar to the majority of the small group and are within the locally anticipated range. Similar strontium and oxygen isotopic values could also be expected further North West in the northern regions of Loch Lomond, which lie on the metamorphic geology of the Southern Highland Group. Although geographical areas from farther regions (including those outside the British Isles) cannot be discounted, the locations presented in this study can perhaps be considered the most probable interpretation of the data due to their close proximity to the site itself.
Discussion
The data presented here suggest that the individuals buried at Cramond lived on a largely terrestrial protein-based diet with no sex-based differentiation of food sources, or at least none that can be detected using these isotopic methods. Although the human δ15N levels could be considered somewhat elevated based on the baseline values, they are within 3–5 ‰ range of enrichment from the herbivores analysed (Bocherens and Drucker 2003), supporting this hypothesis and suggesting no great intake of marine protein (Schulting 1998). The dietary offset between these individuals and the local herbivores, however, does indicate a diet rich in higher trophic-level food sources. Although the human δ 34S from Cramond is relatively high compared with the typically ‘terrestrial’ values, given that similar values were measured in the fauna from the site (including herbivores), it seems likely that this is instead related to the coastal proximity of the site and the so-called sea spray effect rather than the consumption of marine foods (Nehlich et al. 2010; Richards et al. 2003).
The reliance on terrestrial resources and the lack of measurable consumption of marine fish throughout the early medieval period, at both inland and coastal locations, is a trend that has been seen at other sites across England, Wales and Scotland in the early medieval period (see Fig. 7). This is consistent with the idea that marine fish consumption only becomes increasingly visible in both the zooarchaeological and dietary isotope evidence from the 10th century onwards. The reasons for this are probably varied, not only the increasing influence of Christian dietary rules, following the Benedictine reforms in England, but also increasing urbanisation and the concurrent/subsequent growth in the North Sea and Baltic fishing industries and commodities trades (Barrett 2016; Barrett et al. 1999; Orton et al. 2016; Barrett and Richards 2004; Müldner and Richards 2007; Müldner 2016; Richards et al. 2006).
The strontium and oxygen isotopic data suggest that the majority of these individuals can be identified as broadly ‘local’ to the site, with the exception of a male and a female (CR76-C-1 and CR76-C-8, respectively). Based on the high similarity of their δ 34S levels to the other individuals buried at Cramond, and to local fauna, although these individuals may have spent their childhoods in different regions, it seems likely they have all spent the years prior to death in a coastal area of Scotland, likely in close proximity to the site itself. This study provides evidence of early medieval mobility in Scotland and, while the majority of individuals buried at Cramond were likely local, the contemporary populations were, to a degree, mobile. This suggests that the social and cultural borders that may have existed between the different peoples of Scotland in the early medieval period were not rigid and mobility was socially as well as physically possible. One of the females (CR76-C-8) was most likely born on one of the isles of the Inner Hebrides. These areas could have been occupied by the Irish settlers, the Scotti, at the time; however, Picts were also present on these Isles before they were forced to retreat eastwards by the Scotti. Close connections between Scotland and Ireland have been noted throughout the early medieval period (Stevens 2017), furthering the significance to this possibility. Alternatively, this individual could be associated with origins in Inverclyde and Renfrewshire, areas under the control of the Strathclyde Britons. Two other studies have also noted the potential movement of people from the western regions of Scotland in the early medieval period based on strontium and oxygen isotope data. The long-cist burial of a male individual at Bridge of Tilt (Czére et al. 2021), and one of four contemporary individuals (a female) from Phase 1 at Portmahomack (Walther et al. 2016) both displayed isotope values which would correspond to a westerly origin, or in the case of the latter, possibly of central Britain (Walther et al. 2016). Peeblesshire, Lanarkshire and the surrounding region of Loch Lomond were identified as possible places of origin of the male individual (CR76-C-1). It is worth considering that while 87Sr /86Sr and δ18O isotope data may correlate with only a small number of areas in some instances, there is potential for a wide range of sites to fall within the identified biosphere ranges. In such an instance, the proximity of these locations to the site of deposition should be considered and the areas located within the shortest range should be regarded as the most plausible interpretation.
Cramond lies in a region that was on the frontline between different groups of people. Moreover, it was easily accessible through naval routes and Cramond’s importance as a harbour is unlikely to have diminished after the Roman withdrawal (Cessford 2001). Scots, Angles and Picts were all known for their raiding activities in the centuries following the Roman era (Alcock 2003), with contact between the different groups of the region perhaps being inevitable. The introduction of Christianity to Britain occurred at the end of the Roman occupation. As mentioned earlier, however, there are no contemporary written sources that could attest to when it began spreading in the north of Britain (Wormald 2005). It is worth noting, however, that Iona, which is in close proximity to the potential geographical origins of CR76-C-8, is often linked to St Columba and is possibly amongst the earliest locations settled by Christian populations in Scotland (Carver 2004; Gondek 2006).
It is also noteworthy that while one man and one woman buried at this site originated from geographically different locations, they shared similar dietary customs with the individuals identified as local. Their interment in the same structure could possibly suggest familial ties between these people, or the assimilation and acceptance of these newcomers as part of a community. It must also be stressed, however, that the group burial at Cramond diverges considerably from the local norms of the time. Although the reuse of prehistoric monuments in late Iron Age and early medieval Scotland for burial purposes is not unique, this practice was more common in England at the time, while long-cist cemeteries with a preference for a supine extended body position were the dominant tradition within the burial landscape of the region south of the Firth of Forth (Bell 2005; Foster 2014; Maldonado 2011, 2013; Williams 1997). Furthermore, violent death in early medieval Scotland was likely not uncommon due to territorial expansions and competition over the control of various goods and naval routes. Skeletal remains of victims of violent trauma from the time period are relatively rare, however (Alcock 2003; Maldonado 2011), although a recent reanalysis of 306 skeletons from early medieval Scotland indicates a prevalence rate of ~9 % (Boyle 2021). As mentioned above, four of the individuals buried at Cramond exhibited signs of violent skeletal trauma, enhancing the peculiarity of this group, in particular when considered alongside the unusual method of burial. Based on the osteological evidence at least one, but likely two of these injuries were fatal. Although there is no further evidence of such injuries on the other remains, fatal soft-tissue damage would not be possible to detect osteologically (Hukelová 2010; Museum of Edinburgh 2016). In the politically uncertain time period, the importance of various strongholds in the British kingdoms cannot be understated. From the fort, easy access is provided to Cramond tidal island during low tide and its location at the border of regional ethnic, cultural and political groupings would make this a strategically important site to occupy, making local settlements potential targets of raiding activities (Cessford 2001; Lowe 1999). Although this raises further questions about the identity of these people, assigning them to any particular group in the region would be unwise, as the boundaries of these territories were very malleable and traversable at the time (as demonstrated by the movement histories of at least two individuals from the Cramond burial).
The multi-isotope techniques employed in this study have helped shed light on the life of a small group of people in early medieval Scotland, but they also raised further questions. In future studies, the lack of reliance on marine sources throughout Britain during the second half of the first millennium should be explored in more detail using methods allowing to identify potential short-term variations in one's diet, e.g. through the incremental sampling and isotopic analyses of dentinal collagen. Although the utility of sulphur stable isotope analysis to identify freshwater fish consumption has been impeded by the sea-spray effect in the case of this study, this method would be useful at sites located further inland. It is also crucial to generate more comparative sulphur data throughout Britain, in particular from animals. Furthermore, the utility of the analysis of compound-specific carbon- and nitrogen-compositions as indicators of the consumption of aquatic foodstuffs has been demonstrated by a number of studies (Naito et al. 2016; Webb et al. 2018) and may be helpful in cases such as this in identifying more minor contributions. The currently available data on childhood and later-life mobility during this time period in Scotland are also still limited. Further studies could help identify patterns of movement from and to Scotland and beyond during centuries of migrations and political and social transformations as well as would allow to detect any potential inter-sex variations of mobility behaviour.
Conclusions
In this study, we aimed to explore early medieval diet and mobility in Scotland utilising multi-isotopic methods to analyse nine 6th–7th century AD individuals unearthed from a Roman latrine at Cramond, Edinburgh, alongside multiple animal bones from the site. Very little inter-individual variability was observed in carbon, nitrogen and sulphur isotope ratios obtained from the analysis of bone collagen, indicating that no dietary customs could be observed that could potentially indicate social diversification within this group (at least based on these techniques). The majority of the individuals likely spent their childhoods in the region surrounding the site, based on the strontium and oxygen carbonate values presented in this study, with the exception of two individuals (identified as a male and a female skeletally) who originated from two different areas—including the northernmost isles of the Inner Hebrides or Shetland (CR76-C-8), as well as areas of inland Scotland, along the Western Highlands, or Lanarkshire (CR76-C-1). Based on the sulphur isotope signatures, however, each of these individuals seems to have spent a significant period of time in a coastal location in the latter years of their lives, possibly in the region of Cramond. This is further evidenced by the fact that there were no dietary differences between those who spent their childhoods in the Cramond area, and those who had spent their childhoods elsewhere, suggesting these individuals were not recent immigrants.
The results of this project are consistent with dietary patterns observed during the early centuries of the Middle Ages throughout Britain, characterised by a general lack of marine fish consumption, despite easy access to marine and riverine routes. The site’s close proximity to the sea and the consequent sea-spray effect unfortunately hindered the utility of sulphur stable isotope analysis to potentially identify the incorporation of freshwater fish into the diet in the case of this study. The results presented here suggest that movement throughout the 6th–7th century politically divided Scotland was a clear possibility, even from the far Western Isles, providing one of the first insights into early medieval population mobility in Scotland. In order to identify patterns of mobility on a wider scale, however, the analysis of a larger dataset would be required, incorporating a larger number of individuals from a greater number of sites. Such analyses, on a wider range of comparative material, could potentially help further unravel variations in personal inter-population lifetime mobility throughout early medieval Britain.
Availability of data and material
Data generated during this study are included in this manuscript.
Code availability
Not applicable.
References
Alcock L (2003) Kings and Warriors, Craftsmen and Priests in Northern Britain AD 550–850. Society of Antiquaries of Scotland Monograph Series, Edinburgh
Ambrose SH (1990) Preparation and characterization of bone and tooth collagen for isotopic analysis. J Archaeol Sci 17:431–451
Ambrose SH (1991) Effects of diet, climate and physiology on nitrogen isotope abundances in terrestrial foodwebs. J Archaeol Sci 18:293–317
Ambrose SH, Norr L (1993) Experimental evidence for the relationship of the carbon isotope ratios of whole diet and dietary protein to those of bone collagen and carbonate. Prehistoric Human Bone:1–37
Barnetson L (2003) The human remains from the bath-house of the latrine. In: Holmes N, Collard M, Lawson JA (eds) Excavations of Roman Sites at Cramond Edinburgh. Burns & Harris Printers, Dundee
Barrett JH, Richards MP (2004) Identity, gender, religion and economy: new isotope and radiocarbon evidence for marine resource intensification in early historic Orkney, Scotland, UK. Eur J Archaeol 7(3):249–271
Barrett JH, Nicholson RA, Cerón-Carrasco R (1999) Archaeo-ichthyological Evidence for Long-term Socioeconomic Trends in Northern Scotland: 3500 BC to AD 1500. J Archaeol Sci 26:353–358
Barrett JH, Orton D, Johnstone C, Harland J, Van Neer W, Ervynck A, Roberts C, Locker A, Amundsen C, Enghoff IB, Hamilton-Dyer S, Heinrich D, Hufthammer AK, Jones AKG, Jonsson L, Makowiecki D, Pope P, O’Connell TC, de Roo T, Richards M (2011) Interpreting the expansion of sea fishing in medieval Europe using stable isotope analysis of archaeological cod bones. J Archaeol Sci 38:1516–1524
Barrett JH (2016) Studying medieval sea fishing and fish trade: how and why, in: Barrett, J.H. et al. (ed) Cod and herring. The archaeology and history of medieval sea fishing, pp 1–10
Bataille CP, Crowley BE, Wooller MJ, Bowen GJ (2020) Advances in global bioavailable strontium isoscapes. Palaeogeogr Palaeoclimatol Palaeoecol 555:109849
Bataille CP, von Holstein ICC, Laffoon JE, Willmes M, Liu X-M, Davies GR, Cotton J (2018) A bioavailable strontium isoscape for Western Europe: A machine learning approach. PLOS ONE 13(5):e0197386. https://doi.org/10.1371/journal.pone.0197386
Bede (1969) Historia Ecclesiastica Gentis Anglorum,Book I. In: Colgrave B, Mynors RAB (eds) Bede’s Ecclesiastical History of the English People. Oxford
Bell T (2005) The religious reuse of Roman structures in early medieval England. BAR Br Ser 390
Bentley RA (2006) Strontium isotopes from the Earth to the archaeological skeleton: a review. J Archaeol Method Theory 13:135–187
Birck JL (1986) Precision K-Rb-Sr isotopic analysis: application to Rb-Sr chronology. Chem Geol 56(1–2):73–83
Bocherens H, Drucker D (2003) Trophic level isotopic enrichment of carbon and nitrogen in bone collagen: case studies from recent and ancient terrestrial ecosystems. Int J Osteoarchaeol 13:46–53
Boyle A (2021) Cowboys and Indians? A biocultural study of violence and conflict in south-east Scotland c AD 400 to c AD 800, PhD Thesis. University of Edinburgh
Breeze DJ (1994) The Imperial Legacy – Rome and her neighbours. In: Crawford BE (ed) 1994, Scotland in Dark Age Europe: The proceedings of a Day Conference held on 20 February 1993. St John's House Papers, St Andrews, pp 13–20
Brettell R, Evans J, Marzinzik S, Lamb A, Montgomery J (2012) ‘Impious Easterners’: Can Oxygen and Strontium Isotopes Serve as Indicators of Provenance in Early Medieval European Cemetery Populations? Eur J Archaeol 15(1):117–145
Brettell R, Montgomery J, Evans J (2012) Brewing and stewing: the effect of culturally mediated behaviour on the oxygen isotope composition of ingested fluids and the implications for human provenance studies. J Anal At Spectrom 27(5):778
British Geological Survey (1993) 30E Glasgow, Geological Survey of Scotland, 1:63,360/1:50,000, geological map series
British Geological Survey (2006) 32W Livingston, Geological Survey of Scotland, 1:63,360/1:50,000, on geological map series
Britton K (2017) A stable relationship: isotopes and bioarchaeology are in it for the long haul. Antiquity 91(358):853–864
Britton K (2019) Constitution Street: Isotope analysis of bone collagen and tooth enamel. In: Franklin J, Troy C, Britton K, Wilson D, Lawson J, A. (eds) Past lives of Leith: Archaeological work for Edinburgh trams. The City of Edinburgh Council, Edinburgh
Britton K (2020) Isotope Analysis for Mobility and Climate Studies. In: Britton K, Richards M (eds) Archaeological Science: An Introduction. Cambridge, Cambridge University Press, pp 99–124
Britton K, Crowley BE, Bataille CP, Miller JH, Wooller MJ (2021) Silver Linings at the Dawn of a “Golden Age”. Front Ecol Evol 9
Britton K, Fuller BT, Tütken T, Mays S, Richards MP (2015) Oxygen isotope analyisis of human bone phosphate evidences weaning age in archaeological populations. Am J Phys Anthropol 157:226–241
Britton K, Gaudzinski-Windheuser S, Roebroeks W, Kindler L, Richards MP (2012) Stable isotope analysis of well-preserved 120,000-year-old herbivore bone collagen from the Middle Palaeolithic site of Neumark-Nord 2, Germany reveals niche separation between bovids and equids. Palaeogeogr Palaeoclimatol Palaeoecol 333:168–177
Britton K, Grimes V, Niven L, Steele TE, McPherron S, Soressi M, Kelly TE, Jaubert J, Hublin J, Richards MP (2011) J Hum Evol 61(2):176–185
Britton K, Müldner G, Roderick K, Richards M, Talamo S, Noble G (2016) Fish ‘n’ Picts: Reconstructing diet in medieval Scotland using stable isotope analysis. European Association of Archaeologists, Vilnius
Britton K, Willmes M, Moffat I, Grün R, Mannino MA, Woodward S, Jaouen K (2020) Sampling Plants and Malacofauna in 87Sr/86Sr Bioavailability Studies: Implications for Isoscape Mapping and Reconstructing of Past Mobility Patterns. Front Ecol Evol 8:579473
Brown TA, Nelson DE, Vogel JS, Southon JR (1988) Improved collagen extraction by modified Longin method. Radiocarbon 30:171–177
Bryant D, J. and Froelich, P. N. (1995) A model of oxygen isotope fractionation in body water of large mammals. Geochim Cosmochim Acta 59(21):4523–4537
Carver M (2004) An Iona of the East: The Early-medieval Monastery at Portmahomack. Tarbat Ness, Medieval Archaeology 48(1):1–30
Carver M (2019) Formative Britain. Routledge, Oxon
Cessford, C., 2001. "Post-Severan Cramond: A Late Roman and Early Historic British and Anglo-Saxon Religious centre?", [Online], Accessed: 25/02/2014. Available from: http://www.heroicage.org/issues/4/Cessford.html
Chenery CA, Pashley V, Lamb AL, Sloan HJ, Evans JA (2012) The oxygen isotope relationship between the phosphate and structural carbonate fractions of human bioapatite. Rapid Commun Mass Spectrom 26:309–319
Chisholm BS, Nelson DE, Schwarcz HP (1982) Stable-carbon isotope ratios as a measure of marine versus terrestrial protein in ancient diets. Science (New York, N.Y.) 216(4550):1131–1132
Collins MJ, Nielsen-Marsh CM, Hiller J, Smith CI, Roberts JP, Prigodich RV, Wess TJ, Csapo J, Millard AR, Turner-Walker G (2002) The survival of organic matter in bone: a review. Archaeometry 44(3):383–394. https://doi.org/10.1111/1475-4754.t01-1-00071
Cook, M., Lawson, J., Clemments, V., and Bird, L. (in prep). New evidence for the use and occupation of Cramond Roman Fort. Unpublished Report: AOC Archaeology Ltd.
Coplen TB (1988) Normalization of oxygen and hydrogen isotope data. Chem Geol Isotope Geosci Sect 72:293
Coulson JP, Bottrell SH, Lee JA (2005) Recreating atmospheric sulfur deposition histories from peat stratigraphy: diagenetic conditions required for signal preservation and reconstruction of past sulfur deposition in the Derbyshire Peak District, UK. Chem Geol 218:223–248
Curtis-summers S, Montgomery J, Carver M (2014) Stable Isotope Evidence for Dietary Contrast Between Pictish and Medieval Populations at Portmahomack, Scotland. Mediev Archaeol 58:21–43
Curtis-Summers S, Pearson J A, Lamb AL (2020) ‘From Picts to Parish: Stable isotope evidence of dietary change at medieval Portmahomack, Scotland’. J Archaeol Sci Rep 31:102303
Czére O, Fawcett J, Evans J, Sayle K, Müldner G, Hall M, Will B, Mitchell J, Noble G, Britton K (2021) Multi-isotope analysis of the human skeletal remains from Blair Atholl, Perth & Kinross, Scotland: insights into the diet and lifetime mobility of an early medieval individual. Tayside Fife Archaeol J
Dansgaard W (1964) Stable isotopes in precipitation. Tellus 16:436–468
Darling WG, Bath AH, Talbot JC (2003) The O & H stable isotopic composition of fresh waters in the British Isles. 2. Surface waters and groundwater. Hydrol Earth Syst Sci 7(2):183–195
Daux V, Lécuyer, Adam F, Martineau F, Vimeux F (2005) Oxygen Isotope Composition Of Human Teeth And The Record Of Climate Changes In France (Lorraine) During The Last 1700 Years. Clim Chang 70(3):445–464
Daux V, Lécuyer C, Héran MA, Amiot R, Simon L, Fourel F, Martineau F, Lynnerup N, Reychler H, Escarguel G (2008) Oxygen isotope fractionation between human phosphate and water revisited. J Hum Evol 55(6):1138–1147
DeNiro MJ, Epstein (1981) Isotopic composition from aquatic organisms. Geochim Cosmochim Acta 45(10):1885–1894
DeNiro MJ (1985) Postmortem preservation and alteration of in vivo bone collagen isotope ratios in relation to palaeodietary reconstruction. Nature 317:806–809
Deniro MJ, Epstein S (1978) Influence of diet on the distribution of carbon isotopes in animals. Geochim Cosmochim Acta 42(5):459–506
Dobney K, Ervynck A (2007) In: Haselgrove C et al (eds) To fish or not to fish? Evidence for the possible avoidance of fish consumption during the Iron Age around the North Sea. The Later Iron Age in Britain and beyond, pp 403–418
Dunbar LJ, Maldonado A (2012) A long cist cemetery near Auchterforfar Farm, Forfar, Angus-Christian or pre-Christian? Tayside Fife Archaeol J
Edwards KJ, Ralston IBM (eds) (2003) Scotland After the Ice Age: Environment, Archaeology and History, 8000 BC–AD 1000. Edinburgh University Press, Edinburgh
Eriksson G, Lidén K (2013) Dietary life histories in Stone Age Northern Europe. J Anthropol Sci 32(3):288–302
Evans, J. A., Stewart, C., 2014. Sr isotope analysis of Anglo Saxon teeth from Edinburgh, NIGL report. Unpublished
Evans JA, Chenery CA, Mee K, Cartwright CE, Lee KA, Marchant AP, Hannaford L (2018) Biosphere Isotope Domains GB (V1): Interactive Website. Br Geol Surv (Interactive Resource). https://doi.org/10.5285/3b141dce-76fc-4c54-96fa-c232e98010ea
Evans JA, Montgomery GW, Boulton N (2010) Spatial variation in biosphere 87Sr/86Sr in Britain. J Geol Soc 167:1–4
Evans JA, Chenery CA, Montgomery J (2012) A summary of strontium and oxygen isotope variation in archaeological human tooth enamel excavated from Britain. J Anal Atom Spectrom 27(5):754
Evans J, Parker Pearson M, Madgwick R, Sloane H, Albarella U (2019) Strontium and oxygen isotope evidence for the origin and movement of cattle at Late Neolithic Durrington Walls, UK. Archaeol Anthropol Sci 11:5181–5197
Foster SM (1996) Picts, Gaels and Scots. Historic Scotland, London
Foster SM (2014) Picts, Gaels and Scots : early historic Scotland. Birlin Ltd, Edinburgh
Foster, S., 2011. Physical Evidence For The Early Church In Scotland. In: Buildings for Worship in Britain: Celtic and Anglo-Saxon, Department of Continuing Education, Oxford, January 2010, (Unpublished).
Fraser JE (2009) From Caledonia to Pictland : Scotland to 795. Edinburgh University Press
Froehle AW, Kellner CM, Schoeninger MJ (2010) FOCUS: effect of diet and protein source on carbon stable isotope ratios in collagen: follow up to Warinner and Tuross. J Archaeol Sci 37(10):2662–2670
Gat JR (1980) The isotopes of hydrogen and oxygen in precipitation. In Fritz P., Fontes J. Ch. (eds) Handbook of Environmental Isotope Geochemistry, Oxford: Elsevier 1:21–47
Gat JR (1996) Oxygen and hydrogen isotopes in the hydrologic cycle. Annu Rev Earth Planet Sci 4(1):225–262
Gerrard J (2007) Rethinking the small pig horizon at York Minster. Oxf J Archaeol 26(3):303–307
Gondek M (2006) Investigating in Sculpture: Power in Early-historic Scotland. Mediev Archaeol 50(1):105–142
Gordon CA (1965) The Pictish animals observed. Proc Soc Antiqu Scotl 18:215–224
Greene EF, Tauch VE, Webb D (2004) Application of diffuse reflectance infrared Fourier transform spectroscopy (DRIFTS) for the identification of potential diagenesis and crystallinity changes in teeth. Microchem J 76(1–2):141–149
Groves SE et al (2013) Mobility histories of 7th-9th century AD people buried at early medieval Bamburgh, Northumberland, England. Am J Phys Anthropol 151(3):462–476
Hambleton E. (2008) Review of Middle Bronze Age to Late Iron Age Faunal Assemblages from Southern Britain, English Heritage Research Report 71/2008
Hannah EL, McLaughlin TR, Keaveney EM, Hakenbeck SE (2018) Anglo-Saxon diet in the Conversion period: A comparative isotopic study using carbon and nitrogen. J Archaeol Sci Rep 19:24–34
Härke GH (2002) Archaelogy, ideology and society: the German experience. 2nd, rev. edn. P. Lang, Frankfurt am Main
Harland J, M. (2019) Memories of migration? The ‘Anglo-Saxon’ burial costume of the fifth century AD. Antiquity 93(370):1–16
Haydock H, Clarke L, Craig-Atkins E, Howcroft R, Buckberry J (2013) Weaning at Anglo-Saxon raunds: Implications for changing breastfeeding practice in Britain over two millennia. Am J Phys Anthropol 151(4):604–612
Hayes A (2005) Anglian Leadership in Northumbria, 547 A.D. through 1075 A.D. Louisiana State University, Thesis
Hedges REM, Clement JG, Thomas CD, O’Connell TC (2007) Collagen turnover in the adult femoral mid-shaft: Modeled from anthropogenic radiocarbon tracer measurements. Am J Phys Anthropol 133(2):808–816
Hemer KA, Lamb AL, Chenery CA, Evans JA (2017) A multi-isotope investigation of diet and subsistence amongst island and mainland populations from early medieval western Britain. Am J Phys Anthropol 162(3):423–440
Henshall, A. S., 1966-67. Second report of cist burials at Parkburn, Lasswade, Midlothian, Proc Soc Antiqu Scotl 98:204-214.
Hills C (2017) The Anglo-Saxon migration to Britain: an archaeological perspective. Migration and Integration from Prehistory to the Middle Ages, Landesmuseum of Prehistory, Halle, Germany, pp 239–254
Hillson S (1996) Dental Anthropology. Cambridge University Press, Cambridge
Holmes N, Collard M, Lawson JA (eds) (2003) Excavations of Roman Sites at Cramond Edinburgh. Burns & Harris Printers, Dundee
Hughes SS, Millard AR, Chenery CA, Nowell G, Graham Pearson D (2018) Isotopic Analysis of Burials from the Early Anglo-Saxon Cemetery at Eastbourne, Sussex, U.K. J Archaeol Sci Rep 19:513–525
Hughes SS, Millard AR, Lucy SJ, Chenery CA, Evans JA, Nowell G, Pearson DH (2014) Anglo-Saxon origins investigated by isotopic analysis of burials from Berinsfield, Oxfordshire, UK. J Archaeol Sci 42:81–92
Hukelová Z (2010) Possible motives for the burial of human skeletons found in Cramond, Edinburgh, based on osteological examination. The University of Edinburgh, MSc Human Osteoarchaeology edn
Lamb AL, Evans JE, Buckley R, Appleby J (2014) Multi-isotope analysis demonstrates significant lifestyle changes in King Richard III. J Archaeol Sci 50:559–565
Lamb AL, Melikian M, Ives R, Evans J (2012) Multi-isotope analysis of the population of the lost medieval village of Auldhame, East Lothian, Scotland. J Anal Atom Spectrom 27(5):765
Lawson, J., Saville, A. and Engl, R. 2020. [Forthcoming]. Narrow blade pioneers – The Mesolithic occupation at Cramond, Edinburgh and the introduction of narrowblade microlithic technology during the 9th millennium BC, Scottish Archaeological Internet Reports
Leach S, Lewis M, Chenery C, Müldner G, Eckardt H (2009) Migration and diversity in Roman Britain: a multidisciplinary approach to the identification of immigrants in Roman York, England. Am J Phys Anthropol 140(3):546–561
Lee-Thorp JA (2008) On isotopes and old bones. Archaeometry 50(6):925–950
Leggett S (2021) Migration and Cultural Integration in the Early Medieval Cemetery of Finglesham, Kent, through Stable Isotopes. Archaeol Anthropol Sci 13(10):171
Leggett S, Rose A, Praet E, Le Roux P (2021) Multi-Tissue and Multi-Isotope (Δ13C, Δ15N, Δ18O and 87/86Sr) Data for Early Medieval Human and Animal Palaeoecology. Ecology 102(6):e03349
Lengfelder F, Grupe G, Stallauer A, Huth R, Söllner F (2019) Modelling strontium isotopes in past biospheres – Assessment of bioavailable 87Sr/86Sr ratios in local archaeological vertebrates based on environmental signatures. Sci Total Environ 648:236–252
Lewis J, Pike AWG, Coath CD, Evershed RP (2017) Strontium Concentration, Radiogenic (87Sr/86Sr) and Stable (Δ88Sr) Strontium Isotope Systematics in a Controlled Feeding Study. STAR 3. 1:45–57
Longin R (1971) New method of collagen extraction for radiocarbon dating. Nature. 230:241–242
Lowe C (1999) Angles, Fools and Tyrants: Britons and Anglo-Saxon in Southern Scotland AD 450-750. Canongate Books, Edinburgh
Lucy S (2000) The Anglo-Saxon way of death. Sutton Publishing, Stroud
Luz B, Kolodny Y, Horowitz M (1984) Fractionation of oxygen isotopes between mammalian bone-phosphate and environmental drinking water. Geochimica et Cosmochimica Acta 48(8):1689–1693
Maldonado RAD (2011) Christianity and burial in late Iron Age Scotland, AD 400-650. University of Glasgow PhD thesis
Maldonado RAD (2013) Burial in Early Medieval Scotland: New Questions. Mediev Archaeol 57:1–33
Mallet S (2016) Diets in England: Change, Continuity and Regionality between the Iron Age and Domesday Book (700 B.C -1086 A.D.). University of Oxford
Mays S, Beavan N (2012) An investigation of diet in early Anglo-Saxon England using carbon and nitrogen stable isotope analysis of human bone collagen. J Archaeol Sci 39(4):867–874
Mays SA (1997) Carbon stable isotope ratios in medieval and later human skeletons from northern England. J Archaeol Sci 24(6):561–568
Milek K, Jones R (eds) (2012) Science in Scottish Archaeology. Society of Antiquaries of Scotland, Scottish Archaeological Research Framework
Montgomery J, Evans JA, Neighbour T (2003) Sr isotope evidence for population movement within the Hebridean Norse community of NW Scotland. J Geol Soc 160:649–653
Montgomery J, Evans J, Cooper RE (2007) Resolving archaeological populations with Sr-isotope mixing models. Appl Geochem 22:1502–1514
Montgomery J, Evans JA, Chenery CA, Müldner G (2009) Stable isotope analysis of bone. Wasperton: A Roman, British and Anglo-Saxon Community in, Central England
Müldner G (2016) Marine Fish Consumption in Medieval Britain: The Isotope Perspective from Human Skeletal Remains. In: Barrett JH, Orton D (eds) Cod and Herring: The Archaeology and History of Medieval Sea Fishing. Oxbow, Oxford
Müldner, G., Britton, K. And Ervynck, A., 2014. Inferring animal husbandry strategies in coastal zones through stable isotope analysis: new evidence from the Flemish coastal plain (Belgium, 1ste15th century AD), J Archaeol Sci 41: 322-332.
Müldner G, Montgomery J, Cook G, Ellam R, Gledhill A, Lowe C (2009) Isotopes and individuals: diet and mobility among the medieval Bishops of Whithorn. Antiquity 83(322):1119–1133
Müldner G, Richards MP (2007) Stable isotope evidence for 1500 years of human diet at the city of York, UK. Am J Phys Anthropol 133(1):681–697
Museum of Edinburgh (2016) Dark goings on in Cramond, [Exhibition]. Museum of Edinburgh, Edinburgh
Naito YI, Bocherens H, Chikaraishi Y, Drucker DG, Wißing C, Yoneda M, Ohkouchi N (2016) An overview of methods used for the detection of aquatic resource consumption by humans: Compound-specific delta N-15 analysis of amino acids in archaeological materials. J Archaeol Sci Rep 6:720–732
Nehlich O (2015) The application of sulphur isotope analysies in archaeological research: A review. Earth Sci Rev 142:1–17
Nehlich O, Richards MP (2009) Establishing collagen quality criteria for sulphur isotope analysis of archaeological bone collagen. Archaeol Anthropol Sci 1(1):59–75
Nehlich O, Boric D, Stefanovic S, Richards MP (2010) Sulphur isotope evidence for freshwater fish consumption: A case study from the Danube Gorges, SE Europe. J Archaeol Sci 37(5):1131–1139
Nriagu CE, Rees VL, Mekhtieva AY, Lein P, Fritz RJ, Drimmie RG, Pankina RW, Robinson HR (1991) KrouseSulfur Isotopes in the Hydrosphere. In: Krouse HR, Grinenko VA (eds) Stable Isotopes: Natural and Anthropogenic Sulphur in the Environment. John Wiley & Sons, Chichester, pp 177–265
Orton DC, Locker A, Morris J, Barrett JH (2016) Fish for London. In: Barrett, J.H. et al. Cod and herring. The archaeology and history of medieval sea fishing, pp 205–214
Parker Pearson M, Williams J, Marshall P, Montgomery J, Evans J, Chenery C (2018) The Pictish burial cairn, cal ad 640–780. In: Parker Pearson M, Brennand M, Mulville J, Smith H (eds) Cille Pheadair: a Norse farmstead and Pictish burial cairn in South Uist. Oxbow Books, Oxford and Philadelphia, pp 21–41
Pederzani S, Britton K (2019) Oxygen isotopes in bioarchaeology: Principles and applications, challenges and opportunities. Earth Sci Rev 188:77–107
Pellegrini et al (2016) Tooth enamel oxygen “isoscapes” show a high degree of human mobility in prehistoric Britain. Sci Rep 6(1):34986
Privat KL, O’Connell TC, Richards MP (2002) Stable isotope analysis of human and faunal remains from the Anglo-Saxon cemetery at Berinsfield, Oxfordshire: dietary and social implications. J Archaeol Sci 29:779–790
Proudfoot E (1996) Excavations at the long cist cemetery on the Hallow Hill, St Andrews, Fife, 1975-7. Proc Soc Antiqu Scotl 126:387–454
Pye, K., 2004. Isotopic and trace element characterization of human teeth and bones for identification and provenance purposes, NITECRIME Workshop.
Rae and Rae, A and R. (1974) The Roman fort at Cramond, Edinburgh: excavations 1954-1966. Britannia 5:163–224
Reitsema LJ (2013) Beyond diet reconstruction: stable isotope applications to human physiology, health, and nutrition. Am J Hum Biol 25(4):445–456
Richards MP, Fuller BT, Sponheimer M, Robinson T, Ayliffe L (2003) Sulphur isotopes in palaeodietary studies: a review and results from a controlled feeding experiment. Int J Osteoarchaeol 13:37–45
Richards MP, Fuller BT, Molleson TI (2006) Stable isotope palaeodietary study of humans and fauna from the multi-period (Iron Age, Viking and Late Medieval) site of Newark Bay, Orkney. J Archaeol Sci 33:122–131
Ritchie JNG, Ritchie A (1981) Scotland, archaeology and early history. Edinbugh University Press, Edinburgh
Scheuer L, Black S (2004) The Juvenile Skeleton. Elsevier Academic Press, London
Schoeninger and DeNiro (1984) Nitrogen and carbon isotopic composition of bone collagen from marine and terrestrial animals. Geochim Cosmochim Acta 48:625–639
Schoeninger MJ, Moore KM (1992) Bone stable isotope studies in archaeology. J World Prehist 6:247–296
Schoeninger MJ, DeNiro MJ, Tauber H (1983) Stable nitrogen isotope ratios of bone collagen reflect marine and terrestrial components of prehistoric human diet. Science 220:1381–1383
Schulting RJ (1998) Slighting the sea: the Mesolithic-Neolithic transition in northwest Europe. Documenta Praehistorica 25:203–218
Shapiro SS, Wilk MB (1965) An analysis of variance test for normality (complete samples). Biometrika 52(3/4):591–611
Sillen A (2021) 87Sr/86Sr in Archeological and Paleobiological Research: A Perspective. Frontiers in Ecology and Evolution, p 9
Snoeck C, Ryan S, Pouncett J, Pellegrini M, Claeys P, Wainwright AN, Mattielli N, Lee-Thorp JA, Schulting RJ (2020) Towards a biologically available strontium isotope baseline for Ireland. Sci Total Environ 712:136248
Somerset-Fry P (1982) The history of Scotland. Routledge & Barnes & Noble, London
Sponheimer M, Lee-Thorpe JA (1999) Oxygen Isotopes in Enamel Carbonate and their Ecological Significance. J Archaeol Sci 26:723–728
Stevens P (2017) Early medieval jet-like jewellery in Ireland: production, distribution and consumption. Mediev Archaeol 61(2):239–276
Sykes N (2009) Worldviews in Transition: The Impact of Exotic Plants and Animals on Iron Age/Romano-British landscapes. Landscapes 10(2):19–36
Szpak P (2014) Complexities of nitrogen isotope biogeochemistry in plant-soil systems: implications for the study of ancient agricultural and animal management practices. Front Plant Sci 5:288
Thornton B, Martin G, Procee M, Miller DR, Coull M, Yao H, Chapman SJ, Hudson G, Midwood AJ (2015) Distributions of carbon and nitrogen isotopes in Scotland’s topsoil: a national-scale study. Eur J Soil Sci 66(6):1002–1011
Van Klinken GJ (1999) Bone Collagen Quality Indicators for Palaeodietary and Radiocarbon Measurements. J Archaeol Sci 26:687–695
Vitousek PM et al (1999) Weathering versus atmospheric sources of strontium in ecosystems on young volcanic soils. Oecologia 1212:255–259
Walther L, Montgomery J, Evans J (2016) Digest 4.4 Combined strontium and oxygen isotope data at Portmahomack. In: Carver M, Garner-Lahire J, Spall C (eds) Portmahomack on Tarbat Ness: Changing Ideologies in North-East Scotland, Sixth to Sixteenth Century AD. Society of Antiquaries of Scotland, Edinburgh
Warinner C, Tuross N (2009) Alkaline cooking and stable isotope tissue-diet spacing in swine: archaeological implications. J Archaeol Sci 36(8):1690–1697
Webb EC et al (2018) Compound-specific amino acid isotopic proxies for distinguishing between terrestrial and aquatic resource consumption. Archaeol Anthropol Sci 10(1):1–18
Weber M, Tacail T, Lugli F, Clauss M, Weber K, Leichliter J, Winkler DE, Mertz-Kraus R, Tütken T (2020) Strontium uptake and intra-population 87Sr/86Sr variability of bones and teeth—controlled feeding experiments with rodents (Rattus norvegicus. Frontiers in Ecology and Evolution, Cavia porcellus), p 434
Webster L, Brown M, Michelle P (eds) (1997) The transformation of the Roman world AD 400-900. University of California Press
Will (2003) Medieval and post-medieval pottery. In: Holmes N, Collard M, Lawson JA (eds) Excavations of Roman Sites at Cramond Edinburgh. Burns & Harris Printers, Dundee
Williams H (1997) Ancient Landscapes and the Dead: The Reuse of Prehistoric and Roman Monuments as Early Anglo-Saxon Burial Sites. Medieval Archaeol 41(1):1–32
Willows M (2016) Health status in Lowland Medieval Scotland: a regional analysis of four skeletal populations. The University of Edinburgh
Wood I (2018) Britannia and the Barbarians. IN TEMPORE 99
Wormald J (2005) Scotland: A history. Oxford University Press, Oxford
Wright LE, Schwarcz HP (1998) Stable carbon and oxygen isotopes in human tooth enamel: Identifying breastfeeding and weaning in prehistory. Am J Phys Anthropol 106(1):1–18
Acknowledgements
We thank the City of Edinburgh Council the University of Aberdeen and the University of Reading for funding the isotope analyses associated this research. KB and OC also thank the Leverhume Trust (PLP-2019-284 to KB) and the AHRC (award ref: 1775362 to OC) for their financial support during the preparation of this manuscript.
Funding
Funding for the isotopic research was provided to KB and OC by the City of Edinburgh Council the Development Trust Student Fund, University of Aberdeen. The Leverhume Trust (PLP-2019-284 to KB) and the AHRC (award ref: 1775362 to OC) provided financial support during the preparation of this manuscript.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflicts of interest/Competing interests
Not applicable.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
ESM 1
(XLSX 19 kb)
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Czére, O., Lawson, J.A., Müldner, G. et al. The Bodies in the ‘Bog’: A Multi-Isotope Investigation of Individual Life-Histories at an Unusual 6th/7th AD Century Group Burial from a Roman Latrine at Cramond, Scotland. Archaeol Anthropol Sci 14, 67 (2022). https://doi.org/10.1007/s12520-022-01509-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s12520-022-01509-2