
Abstract
A collection of approx. 150 isolated cheek teeth of the eomyid rodent Eomys antiquus which was obtained from a very thin layer in the Hamstead Member of the Bouldnor Formation at Bouldnor Cliff (Isle of Wight, England, UK) is described. The material is post-Grande Coupure, and earliest Oligocene in age. Comparison is made with and new data are given on teeth of Eomys from localities of a similar age situated elsewhere in Europe: Eomys antiquus from Hoogbutsel (Belgium), and Möhren 13, 19, and 20 (Germany), and Eomys aff. antiquus from Montalbán 1D (Spain), and Kocayarma (Turkish Thrace, Turkey). Teeth from Bouldnor Cliff and Hoogbutsel (Mammal Paleogene zone 21) are morphologically close to those from Möhren 13 (Mammal Paleogene zone 22), but tend to be larger. Size decrease in the course of time is also suggested by the smaller size of the teeth from Montalbán 1D (Mammal Paleogene zone 23). The morphological differences observed between the English, Belgian, and German material at the one side, and the Spanish and Turkish material at the other, confirm that the species from Montalbán 1D and Kocayarma is a different, although closely related form. The taxonomic history of Eomys antiquus is reviewed. The species is the commoner of the two oldest European Eomyidae known. General knowledge on the ecology of the eomyids suggests that at the time of deposition of the Eomys-containing layer in the Hampshire Basin forest was close by.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Eomyidae Winge, 1887 is a family of, generally small, extinct rodents, which lived in North America, Europe, and Asia. Most of them are known from their dentition only. Exceptional were the discovery of a complete skeleton with soft body outline identified as Eomys quercyi Comte and Vianey-Liaud, 1987 (Storch et al. 1996), and that of a partial skeleton with soft parts identified as Eomyodon volkeri Engesser, 1987 (Engler and Martin 2015), both at the late Oligocene locality Enspel in Germany. The family Eomyidae includes a large number of genera, some of which are represented by several species. Eomyids inhabited North America from the middle Eocene until the late Miocene (Flynn 2008). In Europe, eomyids have been found from the early Oligocene until the early Pleistocene (Engesser 1999). Because the oldest-known members of the family are from North America, it is generally assumed that the Eomyidae originated on the American continent, spreading henceforth, probably via the Bering route, to Eurasia. The recent identification of several eomyids of Oligocene and Miocene age in continental Asia and Japan is in accordance with this assumption (for a full review of genera identified in Asia, see Kimura et al. 2020).
In the present paper, we will describe and discuss cheek teeth of the oldest-known European eomyid, Eomys antiquus (Aymard, 1853), collected at the locality Bouldnor Cliff (Isle of Wight, England, UK). In addition, new or supplementary data will be provided respecting E. antiquus from the locality Hoogbutsel (Belgium), and E. aff. antiquus from the locality Montalbán 1D (Spain). Comparisons will be made with assemblages of E. antiquus teeth from the localities Möhren 13 and 20 (Germany), which were described by Maridet et al. (2010). Bouldnor Cliff and Hoogbutsel are localities with stratified sedimentary deposits, Möhren 13 and 20 are fissure fillings. In the system of European mammalian reference levels (MP: Mammal Paleogene; Schmidt-Kittler et al. 1987), the faunas from Bouldnor Cliff, Hoogbutsel, and Möhren 20 are placed in zone MP 21 (earliest Oligocene). The fauna from Möhren 13 is considered to be slightly younger (MP 22), while that from Montalbán 1D is referred to zone MP 23 (BiochroM’97 1997).
The genus Eomys comprises a large number of species. Differences in morphology between species are often subtle, and tooth size plays an important role in species identification. Fig. 1 illustrates the magnitude of the variation in size between type materials of the various recognised species.

Ranges and mean values of the length of M1/2 in the type assemblages of the formally named Eomys species. Data are based on the measurements given in the original descriptions. The numbers of teeth measured are indicated next to the horizontal lines. Species are placed in chronological order, following the age assignments by the authors. In case of E. major and E. zitteli, the age of the type locality (Gaimersheim, Germany) is as given in BiochroM’97 (1997). Correlation of the type locality of E. orientalis (Saint Jacques, Nei Mongol, China) to the European system of MP ages is uncertain. Because the type material of E. antiquus comprises only lower molars, data on M1/2 from Bouldnor Cliff (this paper) and Möhren 13 (Maridet et al. 2010) are used instead. The type material of E. minor likewise includes only lower molars. For this species, use is made of the length measurement of the single M1/2 from La Blache (France), provided by Maridet et al. (2010)
The Paleogene (middle/late Eocene-early Oligocene) continental strata of the Hampshire Basin of southern England have of old been an important source for the collection of fossil mammals. Rodents remained poorly known for a long time, because of their small size, and because they are mostly represented by bone fragments and isolated teeth only. Much progress was made by wet screening of large quantities of sediment, as was done by our group in the 1970s (Bosma 1974), and more recently by others (Hooker 1992; Hooker et al. 1995). The material to be described comes from the Hamstead Member of the Bouldnor Formation. This sequence is of particular interest because it correlates to Stehlin’s (1909) ‘Grande Coupure’, a major faunal turnover affecting mammals in Europe around the Eocene-Oligocene boundary (Hooker 2010). The material was collected with great care and perseverance by the late Richard L.E. Ford from Yarmouth (Isle of Wight).
Description of locality
Bouldnor Cliff is an approximately 55 m high cliff at the northwest coast of the Isle of Wight, between Bouldnor (Yarmouth) and Hamstead Ledge. The muds forming the cliff belong to the Bouldnor Formation (lithostratigraphical classification of Insole and Daley 1985). The material described in this paper comes from its middle unit, the Hamstead Member. The Hamstead Member of the Bouldnor Formation is equivalent to the Lower Hamstead Beds of Bristow et al. (1889). Because of continual and extensive sliding, sediments exposed at Bouldnor Cliff and its foreshore are usually not in situ. Table 1 lists the rodent species described from Bouldnor Cliff so far.
Richard Ford collected the Eomys teeth in the 1980s from a few centimeters thick fossiliferous layer, informally called the Eomys bed. The Eomys bed was exposed at the bottom of the cliff during low water in the summer (Ford, pers. comm., 1985; Fig. 2). It consists of bone-rich lenses in a succession of green to green-grey muds (Hooker, pers. comm.), and is situated in the upper part of the Hamstead Member. In Hooker’s (2021) description of the lithostratigraphy of the sections at Bouldnor Cliff, it is indicated as bed UHB10.

Lowermost portion of section at Bouldnor Cliff (Isle of Wight, England, UK) from which Eomys teeth were collected. The arrow indicates the position of the ‘Eomys bed’. Photograph placed at our disposal by R.L.E. Ford (1985)
Material and methods
Dental terminology and measurements
Upper deciduous premolars, permanent premolars, first, second, and third molars are indicated by D4, P4, M1, M2, and M3, respectively. Their lower counterparts are indicated by d4, p4, m1, m2, and m3. First and second molars are taken together as M1/2 or m1/2. The nomenclature used to describe details of the occlusal patterns of the teeth is that of Wang and Emry (1991). The length and width of the teeth were measured using a Leitz Orthoplan measuring microscope equipped with mechanical stage and measuring clocks. Width measurements were taken perpendicular to length measurements. All measurements are given in millimetres. In Figs. 3 and 4 teeth are depicted as left teeth. If a tooth is a right tooth, the accompanying letter is underlined. The orientation of the upper teeth is with the lingual side downward and the anterior side to the left, and the orientation of the lower teeth is with the buccal side downward and the anterior side to the left. Photographs were taken using a Jeol JCM-6000 scanning electron microscope.

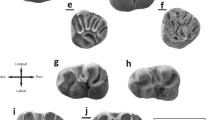
Eomys antiquus from Bouldnor Cliff. a left D4 (UU BC(FD) 76); b right D4 (UU BC(FD) 88); c right D4 (UU BC(FD) 90); d left P4 (UU BC(FD) 73); e left M1 or M2 (UU BC(FD) 91); f left M1 or M2 (UU BC(FD) 92); g right P4 (UU BC(FD) 83); h left P4 (UU BC(FD) 71); i left M1 or M2 (UU BC(FD) 96); j left M1 or M2 (UU BC(FD) 112); k left M1 or M2 (UU BC(FD) 111); l right M1 or M2 (UU BC(FD) 135); m left M3 (UU BC(FD) 141); n right M3 (UU BC(FD) 155); o left M3 (UU BC(FD) 145). Eomys aff. antiquus from Montalbán 1D. p right M3 (UU MO 748); q left M3 (UU MO 746); r left M3 (UU MO 747). Specimens indicated by an underlined letter are depicted in reverse

Eomys antiquus from Bouldnor Cliff. a right d4 (UU BC(FD) 161); b left p4 (UU BC(FD) 1); c left p4 (UU BC(FD) 19); d left p4 (UU BC(FD) 7); e right d4 (UU BC(FD) 163); f left m1 (UU BC(FD) 21); g right m1 (UU BC(FD) 45); h right m1 (UU BC(FD) 50); i left m2 (UU BC(FD) 26); j left m2 (UU BC(FD) 171); k right m2 (UU BC(FD) 38); l left m3 (UU BC(FD) 51); m left m3 (UU BC(FD) 53); n left m3 (UU BC(FD) 59). Specimens indicated by an underlined letter are depicted in reverse
Institutional abbreviations
BSPG: Bayerische Staatssammlung für Paläontologie und Geologie, München, Germany;
MNHN: Muséum national d‘Histoire naturelle, Paris, France;
MTA: Natural History Museum of the General Directorate of Mineral Research and Exploration (M.T.A.), Ankara, Turkey;
NAT: Naturalis Biodiversity Center, Leiden, The Netherlands;
NHMUK: Natural History Museum, London, UK;
UU: Department of Earth Sciences of Utrecht University, Utrecht, The Netherlands.
Other abbreviations
IOW: Isle of Wight
MP zone: Mammal Paleogene zone
Localities other than Bouldnor Cliff
Hoogbutsel (MP 21): The locality, situated in the Belgian Basin, was discovered and described by Glibert and De Heinzelin (1952). Data with respect to the rodent fauna are in Misonne (1957), Fahlbusch (1973), Freudenthal (1988), De Bruijn and Űnay (1989), Vianey-Liaud (1994), and Smith (2003).
Montalbán 1D (MP 23): This is the classical locality of Montalbán (Spain) (Freudenthal et al. 1990; Freudenthal 1996). Rodents from the Montalbán localities are dealt with in several publications, starting with that by Thaler (1969). Descriptions and discussions on Eomys from Montalbán are in Fahlbusch (1973), Comte and Vianey-Liaud (1989), and Maridet et al. (2010). The Eomys teeth included in this paper were collected by one of us (H.d.B.) during field work at Montalbán (Spain) in the 1960s. Fahlbusch (1973) figured several of these teeth.
Möhren 13 (MP 22), Möhren 20 (MP 21), and Möhren 19 (MP 21): The Möhren localities are karstic fissure fillings on the Franconian Alb (South Germany) (Heissig 1973, 1978). The Eomyidae from these fillings have been described in detail by Maridet et al. (2010). Data on other rodents are in Heissig (1973, 1978, 1987), Schmidt-Kittler and Vianey-Liaud (1979), Wang and Heissig (1984), Dienemann (1987), De Bruijn and Űnay (1989), and Cuenca-Bescós and Canudo (1992).
Comparative material
Eomys antiquus from Möhren 13
2 D4s (BSPG 1972 XI 3908, 4265);
47 P4s (BSPG 1972 XI 3901-3907, 3909-3947, 4267; measurements were taken from 20 randomly selected complete specimens);
92 M1/2s (BSPG 1972 XI 3948-3954, 3956-4036, 4126, 4146, 4154, 4203; measurements were taken from 30 randomly selected complete specimens);
21 M3s (BSPG 1972 XI 4037-4057; measurements were taken from 19 complete specimens);
3 d4s (BSPG 1972 XI 4261-4262, 4264);
48 p4s (BSPG 1972 XI 4058-4089, 4092-4107; measurements were taken from 20 randomly selected complete specimens);
95 m1/2s (BSPG 1972 XI 3955, 4108-4125, 4127-4135, 4137-4145, 4147-4153, 4155-4202, 4204, 4266, 4268; measurements were taken from 50 randomly selected complete specimens);
55 m3s (BSPG 1972 XI 4205-4229, 4231-4234, 4236-4260, 4269; measurements were taken from 20 randomly selected complete specimens).
Eomys antiquus from Möhren 20
7 P4s (BSPG 1975 XXII 752, 756-761; measurements were taken from 6 complete specimens);
13 M1/2s (BSPG 1975 XXII 762-768, 770-775; measurements were taken from 11 complete specimens);
3 M3s (BSPG 1975 XXII 753, 776-777);
2 d4s (BSPG 1975 XXII 751, 783);
4 p4s (BSPG 1975 XXII 778-781);
16 m1/2s (BSPG 1975 XXII 746-750, 784-785, 787-795; measurements were taken from 15 complete specimens);
10 m3s (BSPG 1975 XXII 754-755, 796-800, 832-834).
Material from Möhren 20 identified as Eomys sp. by Maridet et al. ( 2010 )
5 M1/2s (BSPG 1975 XXII 838, 847-850; measurements were taken from 4 complete specimens);
5 m1/2s (BSPG 1975 XXII 839-841, 843-844; measurements were taken from 3 complete specimens).
Eomys antiquus from Möhren 19
1 M1/2 (BSPG 1974 XXV 1995).
Eomys antiquus from Ronzon
Casts of type material: MNHN specimens nrs. LP. R6 and LP. R17.
Eomys fahlbuschi from Kocayarma (Turkish Thrace) (Űnay-Bayraktar 1989) (casts)
1 D4, 1 P4, 2 M1/2s, 1 d4, 3 m1/2s, figured by Űnay-Bayraktar (1989; Plate X, figs. 1-8). N.B. Kept at UU, original material stored at MTA.

Length/width scatter plots for M1/2 and m1/2 of Eomys antiquus from Möhren 20 ( and
and  ) and Möhren 13 (○). For Möhren 13, measurements come from 30 randomly selected M1/2s and 50 randomly selected m1/2s. Specimens from Möhren 20 that were originally identified as Eomys sp. (Maridet et al., 2010) are indicated in grey. CV (coefficient of variation) values (in percentages) for the M1/2 from Möhren 20 are 7.72 (length) and 5.56 (width). CV values for the m1/2 from Möhren 20 are 7.91 (length) and 11.10 (width)
) and Möhren 13 (○). For Möhren 13, measurements come from 30 randomly selected M1/2s and 50 randomly selected m1/2s. Specimens from Möhren 20 that were originally identified as Eomys sp. (Maridet et al., 2010) are indicated in grey. CV (coefficient of variation) values (in percentages) for the M1/2 from Möhren 20 are 7.72 (length) and 5.56 (width). CV values for the m1/2 from Möhren 20 are 7.91 (length) and 11.10 (width)

Length/width scatter plots for M1/2 and m1/2 of Eomys antiquus from Bouldnor Cliff (△), Hoogbutsel (□), and Möhren 20 ( and
and  ). Specimens from Möhren 20 that were originally identified as Eomys sp. (Maridet et al., 2010) are indicated in grey
). Specimens from Möhren 20 that were originally identified as Eomys sp. (Maridet et al., 2010) are indicated in grey

Length/width scatter plots for upper (P4, M1/2, M3) and lower (p4, m1/2, m3) tooth elements of Eomys antiquus from Bouldnor Cliff (△), Hoogbutsel (□), Möhren 20 ( ), and Möhren 13 (○), and of Eomys aff. antiquus from Montalbán 1D (▽). For Möhren 13, randomly selected teeth were used in most instances: 20 P4s, 30 M1/2s, 20 p4s, 50 m1/2s, and 20 m3s. The scatter plot for M1/2 includes the single specimen from Möhren 19 (length 0.98 mm, width 1.06 mm) (◆). The scatter plot for m1/2 includes the two m1s and single m2 in the (type) material of Eomys antiquus from Ronzon (LP. R6 and LP. R17) (
), and Möhren 13 (○), and of Eomys aff. antiquus from Montalbán 1D (▽). For Möhren 13, randomly selected teeth were used in most instances: 20 P4s, 30 M1/2s, 20 p4s, 50 m1/2s, and 20 m3s. The scatter plot for M1/2 includes the single specimen from Möhren 19 (length 0.98 mm, width 1.06 mm) (◆). The scatter plot for m1/2 includes the two m1s and single m2 in the (type) material of Eomys antiquus from Ronzon (LP. R6 and LP. R17) ( )
)

Comparison of the mean length and the mean width of the successive permanent upper and lower dental elements of Eomys (aff.) antiquus from Bouldnor Cliff (BC), Hoogbutsel (HB), Möhren 20 (M20), Möhren 13 (M13), and Montalbán 1D (MO). Of the three M3s from Möhren 20, the exceedingly large one (see Fig. 7) has been excluded
Eomys sp. A from Kocayarma (Turkish Thrace) (Űnay-Bayraktar 1989) (casts)
1 D4, 2 M1/2s, 1 M3, 1 m1/2, 1 m3, figured by Űnay-Bayraktar (1989; Plate X, figs. 13-14, 16, 22-23, 26). N.B. Kept at UU, original material stored at MTA.
Eomys cf. sp. A from Kavakdere (Turkish Thrace) (Űnay-Bayraktar 1989) (casts)
3 M1/2s, 3 M3s, 2 m1/2s, figured by Űnay-Bayraktar (1989; Plate X, figs. 15, 17-21, 24-25). N.B. Kept at UU, original material stored at MTA.
Taxonomic history of Eomys antiquus
The first report of the species has been by Pictet (1853), who mentioned in the second edition of his Traité de paléontologie (tome I, p. 250) a genus indicated as ‘Les Decticus, Aymard’, with Decticus antiquus, Aymard as the only species. It was represented by a fragment of a lower jaw with incisor and cheek teeth, collected by Aymard from the ‘calcaire lacustre du Puy en Velay’ (Ronzon, France). Similar information, including Pictet’s (1853) short description of the specimen, was provided by Gervais (1859) in the second edition of his Zoologie et paléontologie françaises (p. 46). Filhol (1881), studying the mammal fauna of Ronzon, gave a more detailed description of the material, using the name Decticus antiquus (Aym.), and referring to Aymard in Pictet (1853). Filhol (1881) called the jaw ‘l’échantillon type’ (p. 18), and provided an (indistinct) drawing (fig. 4, pl. 6), mentioning a bad st preservation for the specimen.
Following a modern taxonomic approach, Lavocat (1951) assigned Decticus antiquus Aymard to Eomys, a genus created by Schlosser in 1884. At that time, Eomys comprised only one other species, the type species Eomys zitteli Schlosser, 1884. Lavocat formulated a diagnosis for Eomys antiquus, and gave measurements and detailed descriptions, not only of the material mentioned above (kept at MNHN, and registered as LP. R6), but also of a fragment of a lower jaw with m1 (kept at MNHN, and registered as LP. R17). For the latter specimen, no locality name was given. The cheek teeth of LP. R6, which proved to be an m1 and an m2, were figured by Lavocat (1951; pl. V, fig. 4), with the indication ‘Type’. Lavocat (1951)’s diagnosis was fully based on these cheek teeth, reading as follows (p. 40): ‘Espèce de très petite taille; M2 plus forte que M1. M1 de type Eomys normal. M2 sans bourrelet postérieur; pas de bras postérieur du protoconide, mais crête transversale antérieure.’
In their publication on Eomyidae from the Oligocene of western Europe, Comte and Vianey-Liaud (1989) presented an emended diagnosis for Eomys antiquus (Aymard). However, this diagnosis was based on material from localities in Spain and France that are younger (MP 23) than Ronzon (MP 21). One of these localities is Montalbán. Eomys from Montalbán has been considered a different form by Maridet et al. (2010), who described it as Eomys aff. antiquus. We agree with their conclusion (see later in this paper), and propose to maintain, for the time being, Lavocat (1951)’s diagnosis.
In several recent studies, eomyid material has been compared with, or been attributed to Eomys antiquus. Very useful have been the drawings of the cheek teeth in the type specimen LP. R6 as provided by some authors: Engesser (1987; figs. 2d and 5e), Comte and Vianey-Liaud (1989; fig. 37c), and Maridet et al. (2010; fig. 4A). Comte and Vianey-Liaud (1989; fig. 37b) presented in addition a drawing of the m1 in specimen LP. R17. Drawings of complete LP. R6 are in Engesser (1987; fig. 5e), and in Maridet et al. (2010; fig. 4A). We summarise the few published data on the measurements of the teeth in the type material of Eomys antiquus in Table 2.
Systematic palaeontology
Class Mammalia Linnaeus, 1758
Order Rodentia Bowdich, 1821
Family Eomyidae Winge, 1887
Genus Eomys Schlosser, 1884
Type species: Eomys zitteli Schlosser, 1884
Emended diagnosis: Fahlbusch (1970, p. 104).
Formally named species
Decticus antiquus Aymard, 1853
Eomys zitteli Schlosser, 1884
Eomys major Freudenberg, 1941
Eomys huerzeleri Engesser, 1982
Eomys quercyi Comte and Vianey-Liaud, 1987
Eomys minor Comte and Vianey-Liaud, 1987
Eomys gigas Comte and Vianey-Liaud, 1987
Eomys molassicus Engesser, 1987
Eomys ebnatensis Engesser, 1987
Eomys fahlbuschi Űnay-Bayraktar, 1989
Eomys orientalis Wang and Emry, 1991
Eomys alulghensis Cuenca and Canudo, 1994
Eomys reductus Ziegler, 1994
Eomys helveticus Engesser and Kälin, 2017
Eomys schluneggeri Engesser and Kälin, 2017
Remark: See Fig. 1 for proposed ages.
Eomys antiquus (Aymard, 1853)
Material from Bouldnor Cliff
8 D4s (UU BC(FD) 76-79, 87-90);
12 P4s (UU BC(FD) 71-75, 80-86); 1 P4 (NHMUK PV M 423 76);
38 M1/2s (UU BC(FD) 91-103, 105-107, 111-112, 121-140); 2 M1/2s (NHMUK PV M 423 73-74);
10 M3s (UU BC(FD) 141-145, 151-155);
3 d4s (UU BC(FD) 161-163);
20 p4s (UU BC(FD) 1-20);
39 m1/2s (UU BC(FD) 21-45, 47-48, 50, 171-181); 4 m1/2s (NHMUK PV M 346 15-16, M 423 75, M 423 77);
15 m3s (UU BC(FD) 51-57, 59, 61-67).
Material from Hoogbutsel (partly figured by Fahlbusch (1973) as E. cf. antiquus; pl. 6, figs. 3-6 pl. 8, figs. 1-6)
2 P4s (NAT St.149.072-St.149.073);
4 M1/2s (NAT St.149.074-St.149.077);
1 M3 (NAT St.149.078);
1 d4 (NAT St.149.068);
3 p4s (NAT St.149.065-St.149.067);
1 m1/2 (NAT St.149.069);
2 m3s (NAT St.149.070-St.149.071).
Measurements: Table 3.
Length/width scatter plots: Figs. 6 and 7.
Descriptions
D4
The occlusal pattern is similar to that of P4. A main addition is the well-developed anteroloph, which extends from the middle of the protoloph to the anterobuccal tooth corner.
The D4s from Bouldnor Cliff: In all teeth the mesoloph is short, reaching halfway or less than halfway to the buccal tooth border. The endoloph is interrupted or very low between the protocone and the place of origin of the mesoloph.
P4
The occlusal pattern is formed by the four heavily built main cusps (protocone, paracone, metacone, and hypocone), the three complete transverse crests (protoloph, metaloph, and posteroloph), the endoloph, and the mesoloph. The teeth show morphological variation with respect to: 1) the length and structure of the mesoloph, 2) the presence of a mesostyle, 3) the presence of an anteroloph, and 4) interruption of the endoloph.
The P4s from Bouldnor Cliff: In most teeth the mesoloph is short, extending less than halfway the buccal tooth border. In one tooth the mesoloph is double, with two very low and narrow branches, which reach to the centre of the central valley. In another tooth the mesoloph is long, interrupted in the middle, and ending in a small mesostyle. Most P4s are without an anteroloph, or possess an anteroloph which is represented by a slight elevation of the anterior tooth wall only. One specimen has a more developed, but still very low anteroloph which is composed of a very short lingual and a short buccal part (Fig. 3h). In another specimen the endoloph is interrupted between the place of origin of the mesoloph and the hypocone. Worn P4s possess a very large posterior wear facet in combination with loss of the posterior tooth segment.
The P4s from Hoogbutsel: In both specimens the mesoloph is short (Fahlbusch 1973; pl. 8, fig. 1). One tooth has a thickening at the anterior tooth wall which reminds of an anteroloph.
M1/2
The occlusal pattern is very regular. The four main cusps (protocone, paracone, metacone, and hypocone) are strongly built, the four main transverse crests (anteroloph, protoloph, metaloph, and posteroloph) are well developed and complete. The endoloph, although narrow in some specimens, is mostly uninterrupted. The morphological variation of the M1s and M2s relates to: 1) the length and height of the mesoloph, 2) the presence of a mesostyle, 3) the depth of the depression which is present at the anterolingual tooth wall between the lingual end of the anteroloph and the protocone, 4) interruption of the endoloph, and 5) the presence of a short additional crest originating from the endoloph in between the mesoloph and metaloph.
The M1/2s from Bouldnor Cliff: In about 50% of the teeth the mesoloph extends approximately halfway to the buccal tooth border. In most of the other specimens the mesoloph is shorter. In two teeth it is longer, without reaching the buccal tooth border. The mesoloph is often low or irregular at its buccal end. One M1/2 carries a mesostyle (Fig. 3f). The depression at the anterolingual tooth wall between the lingual end of the anteroloph and the protocone is shallow in most teeth, in four teeth it is relatively deep. In one tooth the endoloph is interrupted between the protocone and the place of origin of the mesoloph (Fig. 3k). In another five M1/2s the endoloph is interrupted or extremely low between the place of origin of the mesoloph and the hypocone.
The M1/2s from Hoogbutsel: In one tooth the mesoloph reaches about halfway to the buccal tooth border, in the other M1/2s it is shorter. The depression at the anterolingual tooth wall between the lingual end of the anteroloph and the protocone is relatively deep in all four specimens (Fahlbusch 1973; pl. 8, figs. 2-5). In one tooth the endoloph is interrupted between the protocone and the place of origin of the mesoloph. Another tooth possesses the short additional crest originating from the endoloph in between the mesoloph and metaloph.
M3
The occlusal pattern is dominated by the large protocone and paracone, and by the complete anteroloph, protoloph, and metaloph. All teeth possess a posteroloph. The posteroloph varies in length. The mesoloph originates from the middle of the metaloph. Variable features are: 1) the presence, length, and structure of the mesoloph, 2) the depth of the depression at the anterolingual tooth wall between the lingual end of the anteroloph and the protocone, and 3) interruption of the endoloph.
The M3s from Bouldnor Cliff: Most teeth possess a short and very low, irregularly built mesoloph. Three teeth are without a mesoloph. In one tooth the mesoloph is relatively long, reaching close to the buccal tooth border (Fig. 3m). The depression at the anterolingual tooth wall is shallow. In all but one specimen the endoloph is interrupted or very low between the protocone/protoloph and the hypocone/metaloph.
The M3 from Hoogbutsel: The single specimen (Fahlbusch 1973; pl. 8, fig. 6) has a mesoloph which extends halfway to the buccal tooth border, a relatively deep depression at the anterolingual tooth wall between the lingual end of the anteroloph and the protocone, and an interrupted endoloph.
d4
The main occlusal elements are the same as in p4. There is no ectostylid. The teeth are morphologically variable with regard to the following aspects: 1) the presence of a metalophid, 2) the presence of an anteroconid or anterolophid, and 3) the presence and length of the mesolophid.
The d4s from Bouldnor Cliff: In two of the three specimens the protoconid and the metaconid are connected by a metalophid. In the third tooth the protoconid and the metaconid are separated from each other by a deep valley. The latter tooth has neither an anteroconid nor an anterolophid. One d4 has an anterolophid which is present as a short crest running down from the protoconid in an anterolingual direction (Fig. 4a), and the other d4 has an anterolophid in the shape of a narrow vertical ridge which is situated against the anterior tooth wall (Fig. 4e). One tooth is without a mesolophid (Fig. 4e). In a second tooth the mesolophid is present as an indistinct spur (Fig. 4a). The third tooth has a mesolophid reaching to the lingual tooth border.
The d4 from Hoogbutsel: In the single specimen (Fahlbusch 1973; pl. 6, fig. 3) the protoconid and the metaconid are connected by a metalophid. The tooth has an anterolophid as a distinct crest which runs down from the protoconid in anterolingual direction. The mesolophid extends halfway to the lingual tooth border.
p4
The most prominent elements of the occlusal structure are the protoconid, the metaconid, the hypoconid, the entoconid, the hypolophid, and the ectolophid. The protoconid and the metaconid are situated close together, and are connected by a complete, but narrow metalophid. The ectolophid is uninterrupted. All teeth have a posterolophid as described for m1/2. In most specimens the posterolophid is short and low. The teeth show morphological variation with respect to: 1) the presence of an anteroconid, 2) the length of the mesolophid, and 3) the presence of an ectostylid.
The p4s from Bouldnor Cliff: Six out of 19 specimens possess an anteroconid. The anteroconid is present as a small and low cuspule that is situated against the anterior tooth wall. The length of the mesolophid varies more in p4 than it does in m1/2. In eight out of 18 teeth the mesolophid extends approximately halfway to the lingual tooth border. In two teeth it is shorter. In four teeth the mesolophid reaches about ¾ of its maximum length. In another four teeth the mesolophid is more or less complete, ending in a low and/or irregular way at the lingual tooth border, or close to this border. Three specimens possess a small ectostylid or ectostylid-like structure that is situated low against the wall of the sinusid.
The p4s from Hoogbutsel: The only undamaged tooth (Fahlbusch 1973; pl. 6, fig. 4) possesses a small and low, but distinct anteroconid. The mesolophid reaches about halfway to the lingual tooth border. The tooth is without an ectostylid.
m1/2
The occlusal pattern is very regular. The four main cusps (protoconid, metaconid, hypoconid, and entoconid) are prominent, the metalophid and hypolophid are complete, strongly built crests. All teeth possess an anterolophid and a posterolophid. The posterolophid extends from the middle of the hypolophid in a lingual direction, ending low against the posterior wall of the entoconid. Its length is variable. The ectolophid is well developed and complete, but sometimes low between the place of origin of the mesolophid and the hypoconid. The morphological variation of the m1s and m2s relates for the main part to the following aspects: 1) the length of the mesolophid, 2) development of the buccal part of the anterolophid, 3) the position of the connection between the anterolophid and the protoconid/metalophid, and 4) the presence of an ectostylid.
The m1/2s from Bouldnor Cliff: In practically all specimens the mesolophid extends about halfway to the lingual tooth border. In two teeth it is longer, (nearly) reaching this border, but ending low and narrow. In approximately 50% of the m1/2s the buccal part of the anterolophid is short, in the remaining m1/2s it is longer. The anterolophid is connected to the protoconid in about 50% of the teeth, in most of the other teeth it is connected to the metalophid at a place adjacent to the protoconid. In three teeth the anterolophid occurs more or less isolated. Seven m1/2s have a small ectostylid at the bottom of the sinusid, close to the buccal tooth border.
The m1/2 from Hoogbutsel: In the single specimen (Fahlbusch 1973; pl. 6, fig. 5), probably an m2, the mesolophid reaches halfway to the lingual tooth border. The buccal part of the anterolophid is relatively long. The anterolophid is connected to the most buccal portion of the metalophid. The tooth does not possess an ectostylid.
m3
The occlusal pattern is dominated by the strongly built protoconid and hypoconid, the high and pointed metaconid, and the complete, well-developed metalophid and posterolophid. The entoconid is a low, often indistinct cusp. The mesolophid is directed towards the entoconid, or towards a place slightly anterior. The anterolophid has a relatively long buccal part. In most teeth the anterolophid is connected to the metalophid at a place adjacent to the protoconid. In some specimens the connection between the anterolophid and the metalophid is very low and narrow, which gives the anterolophid an isolated appearance. The ectolophid is complete. The outer synclinid is deep. None of the m3s possesses an ectostylid. The morphological variation relates mainly to the length of the mesolophid.
The m3s from Bouldnor Cliff: In eight out of 14 specimens the mesolophid reaches halfway to the lingual border of the occlusal surface, in two specimens it is slightly longer. In the remaining four teeth the mesolophid ends at or close to the lingual border or to the entoconid. One tooth (Fig. 4n) is exceptional: apart from being small, it has an anterolophid with a very short buccal part. Its mesolophid ends close to the entoconid.
The m3s from Hoogbutsel: In one of the two m3s (Fahlbusch 1973; pl. 6, fig. 6) the mesolophid extends halfway to the entoconid. In this tooth the anterolophid and the metalophid are strongly connected at some distance from the protoconid. This leads to a deep valley between the protoconid and the buccal part of the anterolophid. The other specimen, an incomplete tooth, has a mesolophid which reaches near to the entoconid.
Eomys aff. antiquus (Aymard, 1853)
(Fig. 3p-r)
Material from Montalbán 1D (partly figured by Fahlbusch (1973) as E. cf. antiquus; pl. 6, figs. 7-17; pl. 8, figs. 7-23)
3 D4s (UU MO 728-729, 740);
9 P4s (UU MO 701-707, 741-742);
14 M1/2s (UU MO 708-716, 743-745, 749A, 749B);
4 M3s (UU MO 746-749);
1 d4 (UU MO 730);
1 p4 (UU MO 717);
18 m1/2s (UU MO 718-726, 731-739);
1 m3 (UU MO 727).
Measurements: Table 3.
Length/width scatter plots: Fig. 7.
Descriptions
Remark: Only the variability of the morphology of the respective dental elements is discussed. For general morphological information see the corresponding sections on E. antiquus from Bouldnor Cliff and Hoogbutsel.
D4
In all three specimens the mesoloph is extremely short, or rather absent. One tooth possesses a small mesostyle which is situated close to the paracone. In the same tooth the anteroloph is very low. The endoloph is uninterrupted.
P4
The mesoloph is short, extending halfway or less than halfway to the buccal tooth border. Three teeth have a mesostyle, one tooth has two mesostyles. The tooth with the two mesostyles carries a very low anteroloph which is composed of a very short lingual and a short buccal part. The other P4s have no anteroloph or an anteroloph in the form of a thickening of the anterior tooth wall. The endoloph is complete. In one tooth it is low between the protocone and the place of origin of the mesoloph, in another tooth it is low between the place of origin of the mesoloph and the hypocone. One of the P4s (Fahlbusch 1973; pl. 8, fig. 14) has a divergent morphology: the anterior tooth wall runs round, while the mesoloph is directed towards the paracone.
M1/2
In most specimens the mesoloph is short, reaching less than halfway to the buccal tooth border. In two teeth it is longer, extending approximately halfway this border. One M1/2 carries an elongated cuspule in between the (short) mesoloph and the metaloph. Most specimens possess a mesostyle, which is sometimes present as an isolated cuspule, and sometimes as a low and narrow ridge connecting the paracone and metacone. The depression at the anterolingual tooth wall between the lingual end of the anteroloph and the protocone is shallow. It is somewhat deeper in one tooth. The endoloph is uninterrupted. In one M1/2 the endoloph is very low between the place of origin of the mesoloph and the hypocone.
M3
The four M3s have a complex and irregular aspect in comparison with the M3s from Bouldnor Cliff. This is partly caused by the presence of very low and narrow longitudinal ridges connecting the main transverse crests. The mesoloph is long and regular, extending halfway to the buccal tooth border (two teeth) or reaching to this border (two teeth). In one tooth the mesoloph originates from a complete, strong endoloph instead of from the metaloph (Fig. 3r). The endoloph is very low and narrow between the protocone and metaloph in the remaining teeth. The little-worn specimens carry a mesostyle. The depression at the anterolingual tooth corner between the lingual end of the anteroloph and the protocone is either shallow (two teeth) or deeper (two teeth).
d4
In the only d4 (Fahlbusch 1973; pl. 6, fig. 7) the protoconid and the metaconid are connected by a metalophid. The tooth possesses a short and low anterolophid which runs down from the protoconid in a lingual direction, and a mesolophid which extends halfway to the lingual tooth border.
p4
The single p4 (Fahlbusch 1973; pl. 6, fig. 8) is rather worn. This means that the only observable occlusal elements are the four main cusps, the hypolophid, the posterolophid, and the ectolophid. The tooth appears to be without an anteroconid.
m1/2
In nearly all teeth the mesolophid extends approximately halfway to the lingual tooth border. In one tooth it is longer, without reaching this border. In 50% of the specimens the buccal part of the anterolophid is short, in the other 50% it is longer. The anterolophid is mostly connected to the metalophid at a place adjacent to the protoconid instead of directly to the protoconid. In five teeth the anterolophid occurs more or less isolated. None of the m1/2s possesses an ectostylid.
m3
In the single specimen (Fahlbusch 1973; pl. 6, fig. 17) the mesolophid reaches halfway to the entoconid. The anterolophid is connected to the metalophid next to the protoconid. There is a relatively deep depression between the protoconid and the buccal part of the anterolophid.
Results, discussion, and conclusions
Early occurrences of Eomyidae in Europe
Eomys antiquus from Bouldnor Cliff and Hoogbutsel are amongst the oldest Eomyidae presently known from the Eurasian continent. Other eomyids from the earliest Oligocene (MP 21 and 22) of Europe are E. antiquus from the type locality Ronzon (France), and E. antiquus from the fissure fillings Möhren 19, 20, 13, 21, 25, and Ronheim in southern Germany (Maridet et al. 2010). The single M1/2 from the MP 21 locality Dětaň (Czech Republic), figured by Fejfar (1987) as E. cf. zitteli, may belong to E. antiquus. From Spain, there are reports of Eomys sp. collected at the MP 21/22 localities Ciruelos 1A, Olalla 4A, and Santpedor-2 (Ferriz et al. 1993; Freudenthal 1996; Vianey-Liaud et al. 2019).
A distinct unnamed species of eomyid was described from Möhren 20 as Eomyidae gen. et sp. indet. by Maridet et al. (2010). The M3 from Möhren 19, identified as Eomys sp., is considered by us to represent this unnamed species (Maridet et al. 2010, p. 241; BSPG specimen nr. 1974 XXV 1994).
A species of Eomys, known from Europe, and probably also dating back to the early Oligocene, is E. fahlbuschi. The species was described by Űnay-Bayraktar (1989) from Kocayarma in Turkish Thrace. Unfortunately, the age of the Kocayarma fauna could not be determined precisely. Űnay-Bayraktar (1989) assessed correlation with part of the MP 23-27 interval. E. fahlbuschi differs from E. antiquus in its larger size (cf. Fig. 1), and in the more isolated anterolophid of the lower molars.
Comparison of Eomys antiquus assemblages
There is close resemblance in morphology as well as in size between the material from Bouldnor Cliff and that from Hoogbutsel (see Table 3, and Figs. 6 and 7). It may be concluded that the two assemblages represent populations which were very nearly related.
Among Eomys from Möhren 20 and 13, Maridet et al. (2010) distinguished two morphologically similar species: frequently occurring E. antiquus, and a rare, larger species, reported as Eomys sp. After re-examination of the material and plotting the measurements of M1/2 and m1/2 (Fig. 5), we propose that only one species, E. antiquus, is represented. The identification by Maridet et al. (2010) of a second species probably originated from the large variation in the size of the teeth from Möhren 20, especially when compared with the teeth from Möhren 13.
In Fig. 6, M1/2 and m1/2 from Möhren 20 are compared with M1/2 and m1/2 from Bouldnor Cliff and Hoogbutsel. The relatively large size range among the teeth from Möhren 20 may be related to their origin from a fissure filling, as opposed to the origin of the material from Bouldnor Cliff, which comes from a thin layer in a succession of stratified deposits.
The length/width scatter plots for the various permanent dental elements as presented in Fig. 7 illustrate that specimens from Bouldnor Cliff and Hoogbutsel are in general larger than specimens from Möhren 20 and 13 (see also Fig. 8). Notable is the relatively large size of the only Eomys tooth, an M1/2, available from Möhren 19. The age of Möhren 19 is considered to be corresponding to that of Bouldnor Cliff, Hoogbutsel, and Möhren 20 (MP 21).
Detailed analysis of the occlusal patterns of the teeth, as done by Maridet et al. (2010) for the Möhren material, and by us for the material from Bouldnor Cliff and Hoogbutsel, has revealed broad morphological variation, and has made known rare variants. Consistent differences between assemblages have, however, not been found.
Eomys aff. antiquus
In agreement with Maridet et al. (2010), we conclude from our material that Eomys from Montalbán 1D is a form differing from E. antiquus. To the data provided by other authors (Comte and Vianey-Liaud 1989; Maridet et al. 2010), we can add the following differential characters: 1) the main cusps of both the upper and lower teeth are less robust, 2) the P4 and m3 are larger in relation to the other elements in the same tooth row (see Fig. 8), and 3) the occlusal structure of the M3 is more complex (see for details the Systematic palaeontology section and Fig. 3p-r). Differences with E. antiquus are, however, difficult to substantiate, which seriously hampers the creation of a new, separate species.
Űnay-Bayraktar (1989) recognised in the fauna from Kocayarma a species of Eomys which she described as Eomys sp. A. Teeth identified as Eomys cf. sp. A were found in the slightly older locality Kavakdere (also Turkish Thrace). We agree with Űnay-Bayraktar (1989) that Eomys sp. A and Eomys from Montalbán are identical species.
Eomys aff. antiquus furthermore occurs at a number of MP 23/MP 24 localities in France: Itardies, Pech Crabit, Mounayne, Mège, Saint-Martin-de-Castillon C and F (Comte and Vianey-Liaud 1989; Maridet et al. 2010). Engesser and Mödden (1997) reported an E. aff. antiquus from the French MP 23 locality Lovagny (= Eomys nov. sp. 1 in Engesser 1990). It thus appears that E. aff. antiquus was widely distributed across southern Europe early in the Oligocene.
Palaeoecology
Eomyidae are generally thought to have been inhabitants of humid forests (Engesser 1999). This idea fully agrees with the observation that Eomys quercyi was a glider (Storch et al. 1996). Eomyodon volkeri, on the other hand, appears to have been a ground dweller (Engler and Martin 2015). The brachyodont and simple morphology of Eomys teeth is indicative of the consumption of non-abrasive food (Vianey-Liaud 1991; Maridet et al. 2011).
The depositional environment of the upper Eocene/lower Oligocene sediments of the Isle of Wight (Solent Group) was a coastal plain which fluctuated from lagoonal to lacustrine/palustrine to fluvial and was rarely open marine (Hooker et al. 2009). Mammalian assemblages in several horizons indicate habitats ranging from open woodland to closed forest (Hooker et al. 1995). It appears that at the time of deposition of the Eomys bed, forest was nearby.
Distribution and dispersal patterns
Eomys antiquus from Bouldnor Cliff is not only one of the oldest, but also the northwesternmost representative of the genus in Europe. It belongs to the group of mammals which arrived in the area of the Hampshire Basin immediately after the ‘Grande Coupure’ (Hooker 2010). Interestingly, Eomys appears to have had a wide distribution in central and western Europe already during the earliest part of the Oligocene (see data given above).
The differences in tooth size between the assemblages of Eomys antiquus from England, Belgium, and Germany, as reported in this paper, most probably reflect differences between populations which were not only geographically, but also chronologically and/or ecologically separated. The assemblages from Bouldnor Cliff, Hoogbutsel, and Möhren 20 are oldest (MP 21). The assemblage from Möhren 13 is slightly younger (MP 22). It may be concluded that size decrease took place during MP 21/MP 22. The smaller size of the teeth of E. aff. antiquus from Montalbán 1D (MP 23) (Table 3, Figs. 7 and 8), a form most probably originating from E. antiquus, is in line with this assumption.
In Europe, including Turkish Thrace, Eomys underwent considerable radiation: the number of species increased largely (to 14), reaching its peak in the late Oligocene (MN 28 and MN 29) (Engesser 1999; see also Fig. 1). The first clear signs of diversification are the occurrence of E. minor, a species probably not originating from E. antiquus, in the MP 24/25 faunas from Belgarric and La Blache (France) (Comte and Vianey-Liaud 1987, 1989; Maridet et al. 2010), and the presence of two clearly different species, E. fahlbuschi and E. aff. antiquus, in the fauna from Kocayarma (Űnay-Bayraktar 1989). The latter fauna was correlated with uncertainty to MP 25 (Űnay-Bayraktar 1989; p. 93). Eomys disappeared in Europe before the end of the Oligocene.
The Asian record of Eomys merely consists of the type material of E. orientalis from Inner Mongolia (China) (Wang and Emry 1991), and the material from the Valley of Lakes (Central Mongolia) described as E. cf. orientalis, E. aff. orientalis, and Eomys sp. by Maridet et al. (2015). The age of the type material of E. orientalis was indicated to be ‘late middle Oligocene’. The age of the material from the Valley of Lakes is early Oligocene (local biozones A and B). Wang and Emry (1991) as well as Maridet et al. (2015) emphasised the similarity of their material to E. antiquus. It appears plausible, but needs further proof, that Eomys dispersed from Asia to Europe before or during the earliest Oligocene in the shape of an antiquus-like form.
The faunas from Möhren 19 and 20 comprise, next to Eomys antiquus, the eomyid indicated as Eomyidae gen. et sp. indet. (Maridet et al. 2010; this paper). The few teeth available display a distinctive morphology, which made Maridet et al. (2010) to suggest relationship with the Asian late Oligocene/early Miocene genus Asianeomys Wu, Meng, Ye, and Ni, 2006. It can only be speculated whether this little-known, probably new family member dispersed from Asia to Europe alongside E. antiquus.
Data Availability
All data and material are kept in public collections.
References
Alston, E. R. (1876). On the classification of the order Glires. Proceedings of the Zoological Society of London, 44, 61–98.
Aymard, A. (1853). Mentioned in F. J. Pictet, Traité de paléontologie (tome 1), seconde édition (pp. 1–584). Paris: J. B. Baillière.
Bahlo, E. (1975). Die Nagetierfauna von Heimersheim bei Alzey (Rheinhessen, Westdeutschland) aus dem Grenzbereich Mittel-/Oberoligozän und ihre stratigrafische Stellung. Abhandlungen des Hessischen Landesamtes für Bodenforschung, 71, 1–182.
Berger, G. (2008). Die fossilen Schlafmäuse (Gliridae, Rodentia, Mammalia) aus süddeutschen Spaltenfüllungen des Obereozäns und Unteroligozäns. Münchner Geowissenschaftliche Abhandlungen A, 41, 1–128.
BiochroM’97. (1997). Synthèses et tableaux de corrélations. In J. P. Aguilar, S. Legendre, & J. Michaux (Eds.), Actes du Congrès BiochroM’97 (pp. 769–805). Mémoires et Travaux de l’Institut de Montpellier de l’École Pratique des Hautes Études No. 21.
Bonis, L., de (1964). Étude de quelques mammifères du Ludien de La Débruge (Vaucluse). Annales de Paléontologie (Vertébrés), 50, 121–154.
Bosma, A. A. (1974). Rodent biostratigraphy of the Eocene-Oligocene transitional strata of the Isle of Wight. Utrecht Micropaleontological Bulletins, Special publication, 1, 1–128.
Bosma, A. A., & Bruijn, H., de (1979). Eocene and Oligocene Gliridae (Rodentia, Mammalia) from the Isle of Wight, England. Part I. The Gliravus priscus - Gliravus fordi lineage. Proceedings of the Koninklijke Nederlandse Akademie van Wetenschappen, Series B, 82, 367–384.
Bowdich, T. E. (1821). An Analysis of the Natural Classifications of Mammalia, for the Use of Students and Travellers (pp. 1–115). Paris: J. Smith.
Bristow, H.W., Reid, C., & Strahan, A. (1889). The Geology of the Isle of Wight (2nd ed.) (pp. 1–349). London: Memoirs of the Geological Survey. England and Wales. Her Majesty’s Stationery Office.
Bruijn, H., de & Űnay, E. (1989). Petauristinae (Mammalia, Rodentia) from the Oligocene of Spain, Belgium, and Turkish Thrace. In C.C. Black, & M.R. Dawson (Eds.), Papers on Fossil Rodents. In Honor of Albert Elmer Wood (pp. 139–145). Los Angeles, California: Science Series 33, Natural History Museum of Los Angeles County.
Comte, B., & Vianey-Liaud, M. (1987). Nouveaux Eomyidae du Paléogène d'Europe occidentale. Comptes rendus de l’Académie des Sciences, Paris, Série II, 304, 951–954.
Comte, B., & Vianey-Liaud, M. (1989). Eomyidae (Rodentia) de l’Oligocène d'Europe Occidentale. Palaeontographica, Abteilung A, 209, 33–91.
Cuenca, G., & Canudo, J. I. (1994). Les micromammifères de l’Oligocène supérieur des calcaires de Hinojosa de Jarque (Montalbán-Aliaga, Teruel, Espagne). Geobios, 27, 353–384.
Cuenca-Bescós, G., & Canudo, J. I. (1992). Los Sciuridae (Rodentia, Mammalia) del Oligoceno inferior de Montalbán y Olalla (Teruel, España). Discusión sobre el origen de los esciúridos. Boletín de la Real Sociedad Española de Historia Natural, Sección Geológica, 87, 155–169.
Dienemann, A. (1987). Die Gattungen Eucricetodon und Pseudocricetodon (Rodentia, Mammalia) aus dem Oligozän Süddeutschlands. Abhandlungen der Bayerischen Akademie der Wissenschaften, Mathematisch-Naturwissenschafliche Klasse. Neue Folge, 165, 1–158.
Engesser, B. (1982). Le plus grand représentant du genre Eomys (Rodentia, Mammalia) de l’Oligocène de l’Europe: Eomys huerzeleri nov. sp. Geobios, 15, 261–266.
Engesser, B. (1987). New Eomyidae, Dipodidae, and Cricetidae (Rodentia, Mammalia) of the lower freshwater molasse of Switzerland and Savoy. Eclogae Geologicae Helvetiae, 80, 943–994.
Engesser, B. (1990). Die Eomyidae (Rodentia, Mammalia) der Molasse der Schweiz und Savoyens. Schweizerische Paläontologische Abhandlungen, 112, 1–144.
Engesser, B. (1999). Family Eomyidae. In G. E. Rössner & K. Heissig (Eds.), The Miocene Land Mammals of Europe (pp. 319–335). München: Verlag Dr. Friedrich Pfeil.
Engesser, B., & Kälin, D. (2017). Eomys helveticus n.sp. and Eomys schluneggeri n.sp., two new small eomyids of the Chattian (MP 25/MP 26) subalpine lower freshwater molasse of Switzerland. Fossil Imprint, 73, 213–224.
Engesser, B., & Mödden, C. (1997). A new version of the biozation of the lower freshwater molasse (Oligocene and Agenian) of Switzerland and Savoy on the basis of fossil mammals. In J. P. Aguilar, S. Legendre, & J. Michaux (Eds.), Actes du Congrès BiochroM’97 (pp. 475–499). Mémoires et Travaux de l’Institut de Montpellier de l’École Pratique des Hautes Études No. 21.
Engler, T., & Martin, T. (2015). A partial skeleton of the eomyid Eomyodon volkeri Engesser, 1987 (Mammalia: Rodentia) from the late Oligocene Fossil-Lagerstätte of Enspel, Germany. In M. Wuttke, T. Schindler, & K. T. Smith (Eds.), The Fossil-Lagerstätte Enspel – reconstructing the palaeoenvironment with new data on fossils and geology. Palaeobiodiversity and Palaeoenvironments, 95(1), 133–147.
Fahlbusch, V. (1970). Populationsverschiebungen bei tertiären Nagetieren, eine Studie an oligozänen und miozänen Eomyidae Europas. Abhandlungen der Bayerischen Akademie der Wissenschaften, Mathematisch-Naturwissenschafliche Klasse. Neue Folge, 145, 1–136.
Fahlbusch, V. (1973). Die stammesgeschichtlichen Beziehungen zwischen den Eomyiden (Mammalia, Rodentia) Nordamerikas und Europas. Mitteilungen der Bayerischen Staatssammlung für Paläontologie und historische Geologie, 13, 141–175.
Fejfar, O. (1987). A lower Oligocene mammalian fauna from Dětaň and Dvérce NW Bohemia, Czechoslovakia. Münchner Geowissenschaftliche Abhandlungen A, 10, 253–264.
Ferriz, F. J., Freudenthal, M., & Martin Suarez, E. (1993). The oldest mammals from the Betic Cordillera: the Oligocene of the internal Prebetic (Nerpio, Albacete, SE Spain). Geobios, 26, 625–631.
Filhol, H. (1881). Étude des mammifères fossiles de Ronzon (Haute-Loire). Annales des Sciences Géologiques, Paris, 12, 1–270.
Flynn, L. J. (2008). Eomyidae. In C. M. Janis, G. F. Gunnell, & M. D. Uhen (Eds.), Evolution of Tertiary Mammals of North America, Volume 2: Small Mammals, Xenarthrans, and Marine Mammals (pp. 415–427). Cambridge University Press.
Freudenberg, H. (1941). Die oberoligocänen Nager von Gaimersheim bei Ingolstadt und ihre Verwandten. Palaeontographica, Abteilung A, 92, 99–164.
Freudenthal, M. (1988). Redescription of Eucricetodon atavus (Misonne, 1957) (Rodentia) from Hoogbutsel (Belgium). Bulletin de l’Institut royal des Sciences naturelles de Belgique, Sciences de la Terre, 58, 199–207.
Freudenthal, M. (1996). The Early Oligocene rodent fauna of Olalla 4A (Teruel, Spain). Scripta Geologica, 112, 1–67.
Freudenthal, M., Lacomba, J. I., Martínez-Salanova, J., & Sacristán, A. (1990). Nueva sucesión de niveles con micromamíferos en el Oligoceno de Montalbán (Prov. Teruel, España). Acta Salmanticensia, 68, 133–140.
Gervais, P. (1859). Zoologie et paléontologie françaises, deuxième édition (pp. 1–544). Paris: A. Bertrand.
Glibert, M., & De Heinzelin, J. (1952). Le gîte des vertébrés tongriens de Hoogbutsel. Bulletin de l’Institut royal des Sciences naturelles de Belgique, 28, 1–22.
Heissig, K. (1973). Oligozäne Vertebraten aus der Spaltenfüllung “Möhren 13” bei Treuchtlingen, Fränkischer Jura. Mitteilungen der Bayerischen Staatssammlung für Paläontologie und historische Geologie, 13, 177–182.
Heissig, K. (1978). Fossilführende Spaltenfüllungen Süddeutschlands und die Ökologie ihrer oligozänen Huftiere. Mitteilungen der Bayerischen Staatssammlung für Paläontologie und historische Geologie, 18, 237–288.
Heissig, K. (1987). Changes in the rodent and ungulate fauna in the Oligocene fissure fillings of Germany. Münchner Geowissenschaftliche Abhandlungen A, 10, 101–108.
Hemprich, W. F. (1820). Grundriss der Naturgeschichte für höhere Lehranstalten (pp. 1–432). Berlin: Rücker.
Hooker, J. J. (1992). British mammalian paleocommunities across the Eocene-Oligocene transition and their environmental implications. In D. R. Prothero & W. A. Berggren (Eds.), Eocene-Oligocene Climatic and Biotic Evolution (pp. 494–515). Princeton University Press.
Hooker, J. J. (2010). The ‘Grande Coupure’ in the Hampshire Basin, UK: taxonomy and stratigraphy of the mammals on either side of this major Palaeogene faunal turnover. In J. E. Whittaker & M. B. Hart (Eds.), Micropalaeontology, Sedimentary Environments and Stratigraphy: A Tribute to Dennis Curry (1912–2001) (pp. 147–215). London: The Micropalaeontological Society.
Hooker, J.J. (2021). The mammals of the late Eocene-early Oligocene Solent Group, part 1 (pp. 1-147). Monographs of the Palaeontographical Society, Publication No. 659, issued as part of Vol. 175 for 2021.
Hooker, J. J., Collinson, M. E., Van Bergen, P. F., Singer, R. L., De Leeuw, J. W., & Jones, T. P. (1995). Reconstruction of land and freshwater palaeoenvironments near the Eocene-Oligocene boundary, southern England. Journal of the Geological Society, London, 152, 449–468.
Hooker, J.J., Grimes, S.T., Mattey, D.P., Collinson, M.E., & Sheldon, N.D. (2009). Refined correlation of the UK Late Eocene-Early Oligocene Solent Group and timing of its climate history. In C. Koeberl, & A. Montanari (Eds.), The Late Eocene Earth - Hothouse, Icehouse, and Impacts. The Geological Society of America, Special Paper 452, 179–195.
Hugueney, M. (1967). Les Gliridés (Mammalia, Rodentia) de l'Oligocène supérieur de Coderet-Branssat (Allier). Compte rendu sommaire des séances de la Société géologique de France, 1967(3), 91–92.
Illiger, C. (1811). Prodromus systematis mammalium et avium additis terminis zoographicis utriusque classis eorumque versione germanica (pp. 1–301). Berlin: Salfeld.
Insole, A., & Daley, B. (1985). A revision of the lithostratigraphical nomenclature of the late Eocene and early Oligocene strata of the Hampshire Basin, southern England. Tertiary Research, 7, 67–100.
Kimura, Y., Casanovas-Vilar, I., Maridet, O., Kalthoff, D. C., Mörs, T., & Tomida, Y. (2020). The Eomyidae in Asia: Biogeography, diversity and dispersals. Fossil Imprint, 76, 181–200.
Lartet, E. (1869). Sur le Trechomys bonduelli et deux autres rongeurs fossiles de l’Eocène parisien. Annales des Sciences Naturelles, Paris, 12, 151–166.
Lavocat, R. (1951). Révision de la faune des mammifères oligocènes d'Auvergne et du Velay (pp. 1–153). Thèse présentée à la Faculté des Sciences de l’Université de Paris, Série A, No. 2445.
Linnaeus, C. (1758). Systema Naturae per regna tria naturae, secundum classes, ordines, genera, species, cum characteribus, differentiis, synonymis, locis (pp. 1–824). Tomus I. Editio decima, reformata. Holmiae: Laurentius Salvius.
Major, C. I. F. (1873). Nagerüberreste aus Bohnerzen Süddeutschlands und der Schweiz. Palaeontographica, 22, 75–130.
Maridet, O., Daxner-Höck, G., Badamgarav, D., & Göhlich, U. B. (2015). The eomyid rodents (Mammalia) from the Oligocene and Miocene of the Valley of Lakes (Central Mongolia). Paläontologische Zeitschrift, 89, 207–228.
Maridet, O., Hugueney, M., & Heissig, K. (2010). New data about the diversity of Early Oligocene eomyids (Mammalia, Rodentia) in Western Europe. Geodiversitas, 32, 221–254.
Maridet, O., Wu, W., Ye, J., Ni, X., & Meng, J. (2011). New discoveries of glirids and eomyids (Mammalia, Rodentia) in the Early Miocene of the Junggar basin (Northern Xinjiang province, China). Swiss Journal of Palaeontology, 130, 315–323.
Misonne, X. (1957). Mammifères oligocènes de Hoogbutsel et de Hoeleden. I. Rongeurs et ongulés. Bulletin de l’Institut royal des Sciences naturelles de Belgique, 33, 1–16.
Muirhead, L. (1819). Mazology. In D. Brewster (Ed.), The Edinburgh Encyclopaedia, 4th edition, volume 13, 393–480. Edinburgh: Blackwood.
Pictet, F. J. (1853). Traité de paléontologie (tome I), seconde édition (pp. 1–584). Paris: J. B. Baillière.
Pomel, N. A. (1853). Catalogue méthodique et descriptif des vertébrés fossiles découverts dans le bassin hydrographique supérieur de la Loire, et surtout dans la vallée de son affluent principal, l’Allier (pp. 1–193). Paris: J. B. Baillière.
Schlosser, M. (1884). Die Nager des europäischen Tertiärs nebst Betrachtungen über die Organisation und die geschichtliche Entwicklung der Nager überhaupt. Palaeontographica, Abteilung A, 31, 19–161.
Schmidt-Kittler, N., Godinot, M., Franzen, J. L., Hooker, J. J., Legendre, S., Brunet, M., & Vianey-Liaud, M. (1987). European reference levels and correlation tables. Münchner Geowissenschaftliche Abhandlungen A, 10, 13–31.
Schmidt-Kittler, N., & Vianey-Liaud, M. (1979). Evolution des Aplodontidae oligocènes européens. Palaeovertebrata, 9, 33–82.
Smith, R. (2003). Les vertébrés terrestres de l’Oligocène inférieur de Belgique (Formation de Borgloon, MP 21): inventaire et interprétation des données actuelles. Coloquios de Paleontología, Volumen Extraordinario, 1, 647–657.
Stehlin, H. G. (1909). Remarques sur les faunules de mammifères des couches éocènes et oligocènes du Bassin de Paris. Bulletin de la Société géologique de France, série, 4, 9, 488–520.
Stehlin, H. G., & Schaub, S. (1951). Die Trigonodontie der simplicidentaten Nager. Schweizerische Palaeontologische Abhandlungen, 67, 1–385.
Storch, G., Engesser, B., & Wuttke, M. (1996). Oldest fossil record of gliding in rodents. Nature, 379, 439–441.
Thaler, L. (1969). Rongeurs nouveaux de l’Oligocène moyen d’Espagne. Palaeovertebrata, 2, 191–207.
Űnay-Bayraktar, E. (1989). Rodents from the Middle Oligocene of Turkish Thrace. Utrecht Micropaleontological Bulletins, Special publication, 5, 1–119.
Vianey-Liaud, M. (1994). La radiation des Gliridae (Rodentia) à l'Eocène supérieur en Europe Occidentale, et sa descendance Oligocène. Münchner Geowissentschaftliche Abhandlungen A, 26, 117–160.
Vianey-Liaud, M. (1989). Parallélisme chez les Theridomyinae (Rodentia) de l’Oligocène: étude de deux nouvelles espèces des genres Theridomys et Blainvillimys. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 178, 203–241.
Vianey-Liaud, M. (2003). Gliridae (Mammalia, Rodentia) de l’Oligocène européen: origine de trois genres miocènes. Coloquios de Paleontología, Volumen Extraordinario, 1, 669–698.
Vianey-Liaud, M. (1991). Les rongeurs de l'Eocène terminal et de l’Oligocène d’Europe comme indicateurs de leur environnement. Palaeogeography, Palaeoclimatology, Palaeoecology, 85, 15–28.
Vianey-Liaud, M., Minwer-Barakat, R., & Marivaux, L. (2019). The rodents from Santpedor-2 (Ebro Basin, NE Spain) confirm the Oligocene age of the latest primates from the Paleogene of Europe. Geobios, 55, 77–88.
Wang, B., & Emry, R. J. (1991). Eomyidae (Rodentia: Mammalia) from the Oligocene of Nei Mongol, China. Journal of Vertebrate Paleontology, 11, 370–377.
Wang, B., & Heissig, K. (1984). Ephemeromys nov. gen., a primitive prosciurine rodent from the Oligocene of Southern Germany. Mitteilungen der Bayerischen Staatssammlung für Paläontologie und historische Geologie, 24, 105–119.
Winge, H. (1887). Jordfundne og nulevende gnavere (Rodentia) fra Lagoa Santa, Minas Geraes, Brasilien. Med udsigt over gnavernes indbyrdes slægtskab (pp. 1–178). Copenhagen: E Museo Lundii, Vol. 1, no. 3.
Wu, W. Y., Meng, J., Ye, J., & Ni, X. J. (2006). The first finds of eomyids (Rodentia) from the Late Oligocene-Early Miocene of the Northern Junggar Basin, China. Beiträge zur Paläontologie, 30, 469–479.
Ziegler, R. (1994). Rodentia (Mammalia) aus den oberoligozänen Spaltenfüllungen Herrlingen 8 und Herrlingen 9 bei Ulm (Baden-Württemberg). Stuttgarter Beiträge zur Naturkunde B, 196, 1–81.
Acknowledgements
Our sincere gratitude goes out to the late Richard L.E. Ford (Yarmouth, IOW, England), who collected the material from Bouldnor Cliff, and placed it at our disposal for scientific study. We dedicate this paper to his memory. Many thanks are due to Dr Gertrud Rössner (Bayerische Staatssammlung für Paläontologie und Geologie, München, Germany) for making the material from the Möhren localities available to us. Other material lent to us comes from the Department of Earth Sciences of the Natural History Museum (London, UK) and Naturalis Biodiversity Center (Leiden, The Netherlands). The SEM pictures were made with the help of Tilly Bouten, and Margot Stoete assisted with the preparation of the figures (both Department of Geosciences, Utrecht University). We are grateful to Dr Jerry Hooker (Natural History Museum, London, UK) for his information on the position of the Eomys bed in the Isle of Wight Paleogene succession. We thank both Jerry Hooker and the other, anonymous, reviewer of this paper for their constructive comments.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflicts of interest
The authors declare that they have no conflict of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Hans de Bruijn passed away on September, 11, 2021
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Bosma, A.A., de Bruijn, H. & Wessels, W. A large Eomys antiquus (Aymard, 1853) (Mammalia, Rodentia) from the early Oligocene sedimentary deposits at Bouldnor Cliff (Isle of Wight, England, UK). Palaeobio Palaeoenv 103, 447–464 (2023). https://doi.org/10.1007/s12549-022-00535-1
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s12549-022-00535-1