
Abstract
In youth with posttraumatic stress disorder (PTSD) non-response rates after treatment are often high. Epigenetic mechanisms such as DNA methylation (DNAm) have previously been linked to PTSD pathogenesis, additionally DNAm may affect response to (psychological) therapies. Besides investigating the direct link between DNAm and treatment response, it might be helpful to investigate the link between DNAm and previously associated biological mechanisms with treatment outcome. Thereby gaining a deeper molecular understanding of how psychotherapy (reflecting a change in the environment) relates to epigenetic changes and the adaptability of individuals. To date, limited research is done in clinical samples and no studies have been conducted in youth. Therefore we conducted a study in a Dutch cohort of youth with and without PTSD (n = 87, age 8–18 years). We examined the cross-sectional and longitudinal changes of saliva-based genome-wide DNA methylation (DNAm) levels, and salivary cortisol secretion. The last might reflect possible abbreviations on the hypothalamic–pituitary– adrenal (HPA) axis. The HPA-axis is previously linked to DNAm and the development and recovery of PTSD. Youth were treated with 8 sessions of either Eye Movement Reprocessing Therapy (EMDR) or Trauma Focused Cognitive behavioral Therapy (TF-CBT). Our epigenome wide approach showed distinct methylation between treatment responders and non-responders on C18orf63 gene post-treatment. This genomic region is related to the PAX5 gene, involved in neurodevelopment and inflammation response. Additionally, our targeted approach indicated that there were longitudinal DNAm changes in successfully treated youth at the CRHR2 gene. Methylation at this gene was further correlated with cortisol secretion pre- and post-treatment. Awaiting replication, findings of this first study in youth point to molecular pathways involved in stress response and neuroplasticity to be associated with treatment response.
Similar content being viewed by others


Introduction
Posttraumatic stress disorder (PTSD) is a common mental health disorder observed in approximately 16% of youth exposed to traumatic events [1]. Youth with PTSD are troubled by frequent re-experiencing of the traumatic event, persistent avoidance, hyper arousal and negative alterations in cognition and mood [2]. If left untreated, these symptoms can interfere with social functioning and school performance, and have ongoing negative effects on the quality of life of the affected youth [3]. Furthermore, they are considered a crucial factor in shaping the vulnerability to depression and suicidality later in life [4]. This emphasizes the importance of effective treatments for youth with PTSD. Randomized controlled trials (RCTs) have demonstrated the efficacy of trauma-focused psychotherapies in youth with PTSD [5]. But response varies considerably among individuals, with high rates of heterogeneity in response and 20-50% of youth not benefiting sufficiently [6,7,8,9]. Several mechanisms have been associated with differential responses to treatment in youth, amongst them are biological factors, such as; epigenetic, endocrinological and neurological factors [10,11,12]. Besides the potential role of these biological factors as a predictor for treatment outcome, related studies have shown that symptomatic change is likely mediated by underlying biological mechanisms, such as epigenetic and endocrinological change [13]. DNA methylation (DNAm) is an important epigenetic mechanism which reflects epigenetic change and affects endocrine functioning [14]. DNAm represents the transcriptional status of a particular gene and can be influenced by both genetic and environmental factors [15, 16]. Provoked by early exposure to traumatic events, DNAm is assumed to be associated with altered hypothalamic-pituitary-adrenal (HPA) axis functioning, and it is related to the development and recovery of PTSD [17,18,19]. It is assumed that DNAm affects glucocorticoid functioning, in particular the release of stress hormones such as cortisol, the end product of HPA-axis activation, which is pivotal in several central mechanisms involved in PTSD, and trauma-focused treatment, such as the primary stress response, (emotional) memory consolidation, memory retrieval, reconsolidation and extinction learning [20,21,22,23,24]. In our prior study, we reported that higher pretreatment basal cortisol secretion was a potential indicator of treatment response in youth with PTSD [25]. Despite growing evidence showing that DNAm and endocrine mechanisms are important for a successful adaptation to stressful events, and play an important role in development and persistence of PTSD, translational studies in clinical practice are still rare, especially in youth. To the best of our knowledge only four studies [11,12,13, 26] have investigated changes in DNAm in relation to symptomatic response in adults with PTSD. The most recent study [13] examined the relation between DNAm and hydrocortisone treatment. This study identified epigenetic markers, previously linked to startle reaction and fear learning and memory processes, predicting both symptom change and PTSD recovery [11,12,13, 26]. Confirming the evidence from animal models showing that epigenetic changes can be dynamic and potentially reversible as a consequence of environmental programming [27, 28].
Objective and reliable predictive biological markers of treatment response and/or symptomatic change may have the potential to guide treatment selection and improve treatment efficacy. However, despite the promising results of the studies described above, there remains a considerable knowledge gap. First, it is important to recognize that translational studies in humans are still scarce, and so far have only been conducted in adults. These results cannot be automatically translated to youth, because other biological pathways and mechanisms are likely involved as both epigenetic and endocrinological regulation undergoes considerable developmental change [29,30,31,32]. Secondly, it remains insufficiently clear if changes in DNAm related to PTSD, adapt in reaction to trauma-focused psychotherapy, and how these treatment-related changes in DNAm are associated with longitudinal changes in the endocrine system, such as cortisol secretion. Third, the predictive value of baseline DNAm for treatment response is still insufficiently clear. Therefore, in this study we aim to address these knowledge gaps by investigating the cross-sectional and longitudinal changes in DNAm and cortisol secretion in relation to the treatment response of youth with and without PTSD. Youth with PTSD received trauma-focused psychotherapy. We used pre-and post-treatment exploratory methylome wide analysis (MWAS) and targeted analysis on differently methylated positions (DMPs) and regions (DMRs). In addition, we measured salivary cortisol to compare changes in cortisol secretion with findings from the methylation analysis.
Results
Participant characteristics and clinical outcomes
A summary of participant characteristics is shown in Table 1. Responders and non-responders did not differ in baseline sociodemographic, trauma, and clinical characteristics, apart from ethnicity and type of index trauma. At baseline 93.48% of participants met the full DSM-IV diagnostic criteria for PTSD, the remaining 6.52% met criteria for a partial PTSD diagnosis. The most common index trauma in the PTSD group was interpersonal violence, followed by sexual abuse. Youth with PTSD did not differ from trauma exposed controls (TEC) (N = 41) in gender and age, but did differ at baseline in ethnicity, and type of index trauma. The average baseline CAPS-CA score in treatment responders was M = 51.14 points, SD = 23.0, and in non-responders M = 47.75 points, SD = 22.6 which is indicative of moderately severe PTSD. Post treatment mean total CAPS-CA scores improved at T2 (M = 32.51 points, SD = 23.04 t(46) = 9.67, p < .000), and T3 (M = 23.5 points, SD = 22.59 t(18) = 4.41, p < 0.000). See Table 1.
Longitudinal epigenetic trajectories related to treatment outcome in youth with PTSD
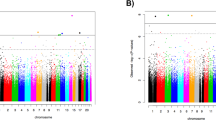
To examine the relation between successful treatment and DNAm we examined the effect of time of measurement (pre- (T1) and post-treatment (T2)) and follow-up treatment (T3) with the diagnosis of PTSD after treatment (responders versus non-responders). The results of the MWAS show no significant differences in treatment responders at T1 vs T2 (see Table 2a, b). Our targeted approach, showed one significant finding in treatment responders (T1 vs T2), at the CRHR2 gene (p = 0.0003). Treatment responders showed increased methylation at T2, see Table 3, Fig. 1. A more in depth examination of this gene related to the glucocorticoid system, showed that increased methylation on the CRHR2 gene in responders, was further associated with lower levels of basal cortisol at T2 (see Table 4). Additionally, our data indicates, however non-significant that in non-responders CRHR2 methylation decreases from T1 to T2 (β = −0.013), this points to an opposite relation with cortisol secretion (decrease of methylation vs. increase of cortisol post-treatment), see Table 4. Furthermore CRHR2 methylation is correlated with cluster D symptoms (hyperarrousal) before treatment, in non-responders (see Table 4). We didn’t detect any longitudinal differences between T1 and T3 in treatment responders. In treatment non-responders we didn’t detect any longitudinal differences between the T1 and T2, and T1 and T3.

Treatment responders at the CRHR2 gene before and after treatment.
Cross-sectional epigenetic differences and clinical outcomes in PTSD patients and TEC
Additionally, we performed a MWAS comparing the treatment responders, non-responders and trauma exposed controls (TEC) across the three different time points. Our DMP analysis revealed significant differences between responders and non-responders at T3, located on the C18orf63 gene (cg15154763, FDR 0.05), indicating a decrease in C18orf63 methylation in treatment responders (see Table 5). Our targeted approach yielded no additional significant findings. Despite not surviving our stringent multiple testing correction in our MWAS and targeted approach, of interest are our top 5 findings from our EWAS DMR analysis. These DMRs identified in the treatment responders vs non responders at T2 and T3 located at RNF39, ALOX12, DUSP22, DIP2C and HOXA4, MUC4 genes, were all previously related to treatment outcome and/or development of PTSD, see Table 6 [11, 26, 33].
To control for the effect of trauma exposure and time, we performed an additional analysis between the responders [22] and non-responders [24] and the TEC. We detected no significant DMR’s or DMP’s between the clinical group and the controls at T1 and T2. At T3 we detected one significant finding between the clinical groups and the control group at one DMR located on chr2: NUP35 (FWER 0.05) Clinical groups showed hypermethylation compared to controls.
Discussion
To the best of our knowledge, this is the first study to detect DNAm in youth with PTSD in relation to trauma-focused psychotherapy response.
Our epigenome-wide cross-sectional analysis showed significant differences between responders and non-responders at T3 (follow-up), located on the C18orf63 gene. Indicating a decrease in C18orf63 methylation in treatment responders. A prior epigenome-wide methylation study found a relationship between C18orf63 methylation and socioeconomic status (SES) in placentas from preterm infants [34]. We consider these findings relevant to our findings since variance in SES is associated with many health outcomes. Including the development of neurodevelopmental and neurobehavioral disorders in children, as well as poorer adult health status and shorter life expectancy [34,35,36]. The association with DNAm might imply biological embedding of SES adversity within critical developmental periods, which in turn could affect long-term child health outcomes [34]. On a molecular level C18orf63 is related to the PAX5 gene (C18orf63 Gene - GeneCards | CR063 Protein | CR063 Antibody). PAX5 was previously found to be related to PTSD and depression [37, 38]. Thompson and colleagues suggested that the perceived capacity of PAX genes to respond to stress is relative high and PAX genes seem to respond within the central nervous system (CNS) as well as interact with a damaged or regenerating environment [39]. In addition the PAX5 gene plays an important role in inflammatory responses (by regulating B-cell differentiation). Multiple studies observed the associated between elevated levels of inflammation in PTSD [40, 41]. Additionally, DNA methylation in genes related to the inflammatory responses are observed as well in PTSD [42,43,44]. These findings indicate that alterations of specific peripheral inflammatory markers maybe related (or induced by) alterations on specific DNAm sites. In addition to these outcomes our MWAS DMR analysis showed a subsequent amount (nominal significant) of overlap with DMR’s reported in the first treatment related studies in adults with PTSD, annotated to RNF39, DUSP22, DIP2C and HOXA4, MUC4 genes [11, 13, 26, 45]. We detected one locus of interest regarding responsivity to treatment located on the ALOX12 genomic region. ALOX12 is the predominant LOX enzyme in the brain and previously this location was related to cortical thickness in PTSD, responsivity to oxidative stress en elevated inflammatory responses [46,47,48]. Interestingly, the few structural neuroimaging studies of trauma-focused psychotherapy in adults that found evidence for pre-to post treatment changes within several regions of the cortex, including the prefrontal cortex, cingulate cortex and insula [46, 49,50,51]. However, imaging data from the cohort used in this study did not replicate the relationship between treatment response and prefrontal cortex volume change, yet we did show that non-response to trauma-focused psychotherapy was characterized by longitudinal bilateral volume decrease in both the posterior and anterior insula [52]. The results from our longitudinal analysis in treatment responders and non-responders did not indicate any significant effects that survived our multiple testing correction. Only at T3 we detected one significant finding between the clinical groups and the control group at one DMR annotated to the NUP35 gene. Clinical groups showed hypermethylation compared to controls. Differential methylation on NUP35 was mentioned before in relation to variation in cognitive function between twins [53].
However, our targeted approach showed interesting longitudinal outcomes. We observed differential DNAm before and after treatment in treatment responders at the CRHR2 genomic region (cg18090898). In response to acute stress, the hypothalamic corticotrophin releasing hormone (CRH) interacts with CRHR2 (a corticotrophin releasing hormone receptor), and it stimulates the anterior pituitary to release adrenocorticotropic hormone. In turn this hormone stimulates the adrenal cortex to release glucocorticoid hormone (cortisol) [54, 55]. Interestingly, in our sample substantial correlations between DNAm at this site and cortisol secretion before and after treatment were observed. Increased methylation on CRHR2 genomic region in treatment responders was associated with an observed increase of cortisol during trauma-related psychotherapy. An observed increase of cortisol during trauma-related psychotherapy, as shown in our sample in treatment responders, may enhance the consolidation of the memories of fear and other strong emotions to facilitate coping with re-exposure [56].
The direction of effect of these findings seems in line with the results of previous studies. These studies showed that lower levels of pretreatment cortisol and lower cortisol change during treatment are related to poorer treatment outcome [25, 57]. Despite that we could not confirm other previous DNAm findings in relation to treatment response previously reported in specific glucocorticoid related regions, such as differential methylation on the FKBP5 and NR3C1 genes. Our findings on the CRHR2 genomic region do support the hypothesis that DNAm at stress-related genes are related with glucocorticoid signaling and with trauma-focused psychotherapy response. This in turn could be possibly related to emotional reactivity during treatment, previously identified as a risk factor for ongoing disbalance after exposure to traumatic events early in life [32]. Only limited information is available on the relation with gene expression within this area. Mainly because of non-matching tissues in common used gene expression databases and the absence of information about mQTLs. However, since cg18090898 is located on first exon of the gene, in general it can be assumed that hypermethylation (similar as to gene promotor hypermethylation) is related to less gene expression [58].
This study has several strengths and limitations. The major strengths of our study are the longitudinal design and standardized PTSD and biological assessments in a unique sample of youth with PTSD, before and after treatment. Furthermore, we used an unbiased methylome-wide approach, extended with a literature based targeted approach, and we included a trauma exposed reference group to control for effect of trauma. We used stringent exclusion criteria and a strict multiple testing correction. However there are also several limitations that have to be acknowledged. Firstly, the relative small size of the groups included in this study, limited us to detect significant epigenome wide results [59]. This also limited us in our ability to examine differences between treatment responders and non-responders for both treatment conditions separately. Despite that both are equally effective trauma-focused psychotherapies in youth [60]. And both therapies share multiple common elements such as exposure to and reprocessing of traumatic memories [61], we acknowledge that there is need for additional research. Information from cohorts with larger sample-sizes, or combined meta-analyses might provide a deeper insight between possible differential effects of both therapies in relation to epigenetic predictors of treatment response and/or biological changes during treatment. Furthermore, the considerable drop-out rate of randomized patients lost at follow-up, restricted us further in our longitudinal analysis. Although, dropout rates in our study reflect routine clinical practice, there is a possibility that drop-out could have influenced our main findings through attrition bias.
Secondly, there are several limitations regarding our post-hoc cortisol measures, such as the reaction on our script driven imagery procedure and the lack of an additional circadian cortisol secretion measure. These are described in more detail in our previous paper [25]. Furthermore, despite that we’ve tried to account for most confounding factors, we weren’t able to include all known factors to influence endocrine function, for example we omitted inquiry on menstrual and pubertal stage. Lastly, another possible limitation is the relevance of methylation in saliva to other tissues such as the brain, given that methylation differences across tissues are substantial. Despite that consistent effects of various methylation quantitative trait loci (mQTLs) are found across tissues [62], partial evidence exists on cross-tissue consistent findings [63]. Therefore a cautious approach to the interpretation of findings obtained from single tissue analyses is needed. In future studies we suggest to link PTSD and treatment outcome and DNAm with specific brain and endocrine related endophenotypes. Given the expected age and time dependent differences, especially considering HPA-axis and brain plasticity, during critical periods it would be helpful to increase sample sizes and include youth already early in life thereby increasing feasibility to differentiate across the developmental stages. In addition the use of continuous outcomes (using symptom dimensions scores instead of a dichotomous diagnosis) reflecting the different symptoms of PTSD and associated emotional and behavioural problems might be considered in future research.
Conclusion
In conclusion, this is the first study in youth with PTSD, that shows the association between successful trauma-focused psychotherapy with specific DNAm changes Overall our results do support and extend previous outcomes presenting DNAm change in relation to treatment response in youth with PTSD, presenting DNAm change in specific genes related to exposure to trauma, the development of PTSD and its related psychological treatments [11, 12, 64]. The study provides further insight in underlying biological mechanisms, and indicates how biological mechanisms might interact with symptomatic change in youth with PTSD. Our results provide novel insights that may contribute to the discovery of the epigenetic mechanisms underlying a successful treatment of PTSD, especially related to HPA-axis related endophenotypes. We expect that the findings may help to better understand how psychological and biological systems interact in order to improve and individualize treatment outcomes, since ideally we aim to prevent ongoing PTSD symptoms in youth and its severe consequences by improving interventions, that are better tailored to each individual patient.
Method
Cohort and study design
In the present study a MWAS, targeted epigenetic approach, cortisol and clinical assessments were performed in a cohort of youth (N = 87, 54.1% female, aged 8-18), Youth with PTSD (N = 46) were matched for age and sex with a control group of trauma exposed controls (TEC) without PTSD (N = 41). Youth with PTSD were recruited between April 2011 and September 2018 at the outpatient child psycho-trauma center of the department of child and adolescent psychiatry, de Bascule in Amsterdam, The Netherlands. The participants were part of a larger RCT comparing trauma-focused cognitive behavioural therapy (TF-CBT) and eye movement desensitization and reprocessing (EMDR) [65]. They were referred by child welfare services, physicians or their general practitioner. TEC were recruited between June 2011 and September 2018 through local elementary- and high schools by researchers JBZ, RodK and JBME. Exclusion criteria for both groups included imminent suicidality, history of psychotic disorder, substance abuse or dependence; IQ < 70; unstable medical condition; recent use of psychotropic medication (past 4 weeks; 6 weeks for fluoxetine); and possibility of pregnancy in females. All participants received a monetary incentive for participation (€5 for each assessment). In both groups written parental and youth assent were obtained for all participants. All procedures were approved by the Medical Ethical Committee of the University Medical Center.
Procedures and measures
Diagnosis for PTSD in the clinical group were established clinically by an experienced child and adolescent psychiatrist or psychologists according to the DSM-IV-TR criteria using both child reports on the Clinician-Administered PTSD Scale for Children and Adolescents (CAPS-CA), which is a reliable semi-structured interview [66]. In addition caregiver information was obtained from the PTSD scale of the Anxiety Disorders Interview Schedule – Parent Version (ADIS-P) [67]. (Partial) PTSD diagnosis was determined using joint-child and caregiver reports on individual symptoms. A symptom was established as present, if either child or caregiver reported its presence. All participants were required to have a CAPS-CA total score indicating at least mild PTSD symptom severity ( > 20 points). Clinical evaluations were performed pre-treatment (T1), post-treatment (8 sessions of psychotherapy) and follow-up (6 months post-treatment). Based on the psychometric properties of the CAPS (-CA) and previous treatment outcome studies using the CAPS-CA, we used ≥ 30% reduction of CAPS-CA total score as response criterion for clinically meaningful improvement [68, 69]. In the TEC exposure to traumatic events were validated according to A1 and A2 criteria of DSM-IV-TR (American Psychiatric Association, 2000) using the Children’s Revised Impact of Event Scale (CRIES) [70, 71]. Information about additional internalizing and externalizing symptoms were measured using youth and caregiver reports, with the Revised Children’s Anxiety and Depression Scale (RCADS) and the Child Behavioral Checklist (CBCL) and Youth Self Report (YSR) [72,73,74,75,76]. Clinical characteristics of both groups are shown in Table 1.
Treatment
After study entry all patients were randomized to receive either protocolled sessions of trauma-focused cognitive behavioral therapy (TF-CBT) or eye movement desensitization and reprocessing (EMDR) by trained and experienced therapists. Supervision by experts on TF-CBT and EMDR was provided throughout the study. Treatment protocols, training and supervision of therapists, as well as treatment fidelity have been described in detail previously [25, 65].
DNA methylation
Three milliliters of saliva were collected and stored in Oragene DNA sample collection kits (DNA Genotek, Canada). DNA was extracted using a Gentra autopure LS system following manufacturers protocol. Genomic DNA samples were resolved on a 1% agarose gel to verify that the DNA was of high molecular integrity. Quantification of the DNA was determined using Qubit (Qiagen, USA). Five hundred nanograms of genomic DNA was sodium bisulfite–treated for unmethylated cytosine (C) to thymine (T) conversion using the EZ DNA Methylation-Gold kit (Zymo Research). Prior to DNA methylation profiling, cases and controls were randomized across the 96 well plates. Technical replicates (n = 8) were included for quality control of array, monitoring potential batch effects. Briefly, converted DNA was amplified, fragmented, hybridized, and scanned using the Illumina Methylation EPIC 850k Beadchip, following the manufacturer’s guidelines.
Cortisol
Information about SDI and cortisol collection is published previously [25]. In short, all participants performed a standardized protocol for script driven imagery (SDI) [77], in combination with the collection of five samples, 10 min and 1 min prior to trauma script imagery as well as 10 min, 20 min, and 30 min after trauma script imagery. Next, For each participant we determined if SDI induced a cortisol stress response [78]. This was defined as an increase of at least 1.5 nmol/l compared to baseline levels. Area under the curve with respect to ground (AUCg) and increase (AUCi) were derived using the trapezoidal formulas [79]. As the SDI failed to induce a cortisol stress response in all but two participants, we did not use the AUCi as measure of cortisol responsivity in further analyses. Therefore, the AUCg was used as a measure of total cortisol output which captures basal cortisol secretion as opposed to stressor reactivity. AUCg was examined for normality of distribution within each group. Non normally distributed data were log-transformed. See also for more detailed information our previous paper for additional information about the data handling [25].
Statistics
The distribution of baseline clinical, trauma and demographic characteristics across responders, non-responders and TEC was examined using X2-tests for categorical variables, independent sample t-tests for normally distributed continuous variables and Mann-Whitney tests for non-normally distributed continuous variables. Paired sample t-test were used to examine pre- to post-treatment symptom change. These statistical analyses were performed using SPSS version 24.0 (SPSS Inc., Chicago IL, USA).
Raw DNA methylation profiles were imported into the statistical programming environment R (v.3.4.2) using the Bioconductor (v3.13) and the minfi (v1.38.0) package [41]. Data quality control was performed using MethylAid. We used the following MethylAid thresholds for quality control; 10.5 for methylated and unmethylated intensities, 12 for overall quality control, 11.75 for bisulphite control, 12.75 for hybridization control, and a detection P-value of 0.95(v1.26.0) [42]. At this stage we removed one sample, because it did not meet our quality control criteria (Follow-up measurement). Further statistical analysis were based on M-values calculated as log2 (beta/(1-beta). Probe expressing infinite values were removed from the dataset. Visualization of associations were based beta-values, i.e. methylation index, which ranges between 0% (methylated) and 100% (methylated). In the next step principal component analysis (PCA) was applied in order to detect any outliers and to evaluate concordant sexes of samples. Moreover, we performed hierarchical clustering analysis using 11 single nucleotide polymorphisms, known to be present on the Illumina EPIC array, to evaluate the concordance of the genotype within the longitudinal sample sets. Next we normalized the dataset applying the function funnorm and we removed all probes annotated to the allosomes, susceptible for cross-hybridization, and probes known to confounded by genetic variation with a minor allele frequency >1%.
Subsequently we correlated meta-data, i.e. sex, age and technical potential confounders (slide and array position) with the first eight principal components. These first eight principal components explained together most of the variance. The qq-plots (of the expected p values versus the observed p values) had a lambda of >.0.85, <1.15 this indicated absence of type-I error inflation and no artificial differences between groups. From this analysis we defined the following statistical models for paired and unpaired analyses respectively:
Differential methylated positions (DMPs) were obtained using limma. For the detection of DMRs we applied the bumphunter function, wherein we used delta beta difference thresholds of 10% and 5% for paired and unpaired analyses respectively. We analyzed the main (DMPs and DMRs) effects longitudinal and crossectional between the following groups: (1) PTSD vs TC on the three different time points; T1: baseline level, before treatment, T2: directly after 8 sessions of trauma-focused treatment, T3: follow-up 6 months after treatment. We assumed a false discovery rate (FDR) < .05 for DMPs and a familywise error rate (FWER) of 0.05 for DMRs as significant. For our targeted approach we selected a limited number of replication loci (NDMP = 170, NDMR = 30), which were based on a literature search which included published MWAS data in youth with PTSD, an previous published treatment studies until June 2021. We also added several candidate loci associated with glucocorticoid functioning and previously related to PTSD development. See supplementary Table 1. In our targeted approach we applied Bonferroni correction threshold wherein we assumed a p < 0.002 significant for the DMR approach, and a p < 0.0003 significant for the DMP approach. The Bonferroni threshold was the product of dividing critical α = 0.05 by the number of DMR’s and DMPs of interest extracted from previous studies. In total we tested 170 DMP’s and 30 DMR’s, which we considered relevant based on previous studies.
Next, we calculated Pearson’s correlation in SPSS between DNAm and our cortisol data. We measured the relation between DNA methylation at significant DMP’s either derived from our epigenome-wide or our targeted epigenetic analyses (if considered relevant to the glucocorticoid system) and cortisol secretion.
References
Alisic E, Zalta AK, Van Wesel F, Larsen SE, Hafstad GS, Hassanpour K, et al. Rates of post-traumatic stress disorder in trauma-exposed children and adolescents: meta-analysis. Br J Psychiatry. 2014;204:335–40.
Association AP, Association AP. Diagnostic and statistical manual of mental disorders: DSM-5. Arlington, VA. (2013).
Carrion VG, Weems CF, Ray R, Reiss AL. Toward an empirical definition of pediatric PTSD: The phenomenology of PTSD symptoms in youth. J Am Acad Child Adolesc Psychiatry. 2002;41:166–73.
Molnar BE, Berkman LF, Buka SL. Psychopathology, childhood sexual abuse and other childhood adversities: relative links to subsequent suicidal behaviour in the US. Psychol. Med. 2001;31:965–77.
Morina N, Koerssen R, Pollet TV. Interventions for children and adolescents with posttraumatic stress disorder: A meta-analysis of comparative outcome studies. Clin Psychol Rev. 2016;47:41–54.
Mavranezouli I, Megnin‐Viggars O, Daly C, Dias S, Stockton S, Meiser‐Stedman R, et al. Research review: psychological and psychosocial treatments for children and young people with post‐traumatic stress disorder: a network meta‐analysis. J Child Psychol Psychiatry. 2020;61:18–29.
Leenarts LE, Diehle J, Doreleijers TA, Jansma EP, Lindauer RJ. Evidence-based treatments for children with trauma-related psychopathology as a result of childhood maltreatment: a systematic review. Eur Child Adolesc Psychiatry. 2013;22:269–83.
Bennett RS, Denne M, McGuire R, Hiller RM. A systematic review of controlled-trials for PTSD in maltreated children and adolescents. Child Maltreatment. 2020;26.3:325–43.
Lindebø Knutsen M, Sachser C, Holt T, Goldbeck L, Jensen TK. Trajectories and possible predictors of treatment outcome for youth receiving trauma-focused cognitive behavioral therapy. Psychol Trauma: Theory, Res, Pract, Policy. 2020;12:336.
Bryant R, Felmingham K, Kemp A, Das P, Hughes G, Peduto A, et al. Amygdala and ventral anterior cingulate activation predicts treatment response to cognitive behaviour therapy for post-traumatic stress disorder. Psychol Med. 2008;38:555–61.
Vinkers CH, Geuze E, van Rooij SJ, Kennis M, Schür RR, Nispeling DM, et al. Successful treatment of post-traumatic stress disorder reverses DNA methylation marks. Molecular psychiatry. 2021; 26.4:1264–71.
Yehuda R, Daskalakis NP, Desarnaud F, Makotkine I, Lehrner A, Koch E, et al. Epigenetic biomarkers as predictors and correlates of symptom improvement following psychotherapy in combat veterans with PTSD. Front Psychiatry. 2013;4:118.
Yang R, Xu C, Bierer LM, Flory JD, Gautam A, Bader HN, et al. Longitudinal genome-wide methylation study of PTSD treatment using prolonged exposure and hydrocortisone. Transl Psychiatry. 2021;11:1–7.
García-Carpizo V, Ruiz Llorente L, Fernández Fraga M, Aranda A. The growing role of gene methylation on endocrine function. J Mol Endocrinol. 2011;47:R75–89.
Schübeler D. Function and information content of DNA methylation. Nature 2015;517:321–6.
Feinberg AP. Phenotypic plasticity and the epigenetics of human disease. Nature 2007;447:433–40.
Yehuda R, Bierer LM. The relevance of epigenetics to PTSD: Implications for the DSM‐V. J Trauma Stress. 2009;22:427–34.
McGowan PO, Sasaki A, D’alessio AC, Dymov S, Labonté B, Szyf M, et al. Epigenetic regulation of the glucocorticoid receptor in human brain associates with childhood abuse. Nat Neurosci. 2009;12:342–8.
Labonte B, Yerko V, Gross J, Mechawar N, Meaney MJ, Szyf M, et al. Differential glucocorticoid receptor exon 1B, 1C, and 1H expression and methylation in suicide completers with a history of childhood abuse. Biol Psychiatry. 2012;72:41–8.
Meir Drexler S, Wolf OT. Stress and memory consolidation. Cognitive neuroscience of memory consolidation: Springer. (2017). 285–300.
Dick A, Provencal N. Central neuroepigenetic regulation of the hypothalamic–pituitary–adrenal axis. Prog Mol Biol Transl Sci. 2018;158:105–27.
Houtepen LC, Vinkers CH, Carrillo-Roa T, Hiemstra M, Van Lier PA, Meeus W, et al. Genome-wide DNA methylation levels and altered cortisol stress reactivity following childhood trauma in humans. Nat Commun. 2016;7:1–10.
Roque AD, Craske MG, Treanor M, Rosenfield D, Ritz T, Meuret AE. Stress-induced cortisol reactivity as a predictor of success in treatment for affective dimensions. Psychoneuroendocrinology 2020;116:104646.
Fischer S, Schumacher T, Knaevelsrud C, Ehlert U, Schumacher S. Genes and hormones of the hypothalamic–pituitary–adrenal axis in post-traumatic stress disorder. What is their role symptom Expr Treat response? J Neural Transm. 2021;128:1279–86.
Zantvoord JB, Ensink JB, op den Kelder R, Wessel AM, Lok A, Lindauer RJ. Pretreatment cortisol predicts trauma-focused psychotherapy response in youth with (partial) posttraumatic stress disorder. Psychoneuroendocrinology 2019;109:104380.
Carleial S, Nätt D, Unternährer E, Elbert T, Robjant K, Wilker S, et al. DNA methylation changes following narrative exposure therapy in a randomized controlled trial with female former child soldiers. Sci Rep. 2021;11:1–11.
Meaney MJ, Szyf M. Environmental programming of stress responses through DNA methylation: life at the interface between a dynamic environment and a fixed genome. Dialogues Clin Neurosci. 2005;7:103.
Weaver IC, Meaney MJ, Szyf M. Maternal care effects on the hippocampal transcriptome and anxiety-mediated behaviors in the offspring that are reversible in adulthood. Proc Natl Acad Sci. 2006;103:3480–5.
Cisler JM, Herringa RJ. Posttraumatic stress disorder and the developing adolescent brain. Biol Psychiatry. 2021;89:144–51.
Daskalakis NP, Bagot RC, Parker KJ, Vinkers CH, de Kloet ER. The three-hit concept of vulnerability and resilience: toward understanding adaptation to early-life adversity outcome. Psychoneuroendocrinology 2013;38:1858–73.
Nederhof E, Schmidt MV. Mismatch or cumulative stress: toward an integrated hypothesis of programming effects. Physiol Behav. 2012;106:691–700.
Agorastos A, Pervanidou P, Chrousos GP, Baker DG. Developmental trajectories of early life stress and trauma: a narrative review on neurobiological aspects beyond stress system dysregulation. Front Psychiatry. 2019;10:118.
Rutten BP, Vermetten E, Vinkers CH, Ursini G, Daskalakis NP, Pishva E, et al. Longitudinal analyses of the DNA methylome in deployed military servicemen identify susceptibility loci for post-traumatic stress disorder. Mol Psychiatry. 2018;23:1145–56.
Santos HP Jr, Bhattacharya A, Martin EM, Addo K, Psioda M, Smeester L, et al. Epigenome-wide DNA methylation in placentas from preterm infants: association with maternal socioeconomic status. Epigenetics 2019;14:751–65.
Nelson CA III. Hazards to early development: the biological embedding of early life adversity. Neuron 2017;96:262–6.
Chin-Lun Hung G, Hahn J, Alamiri B, Buka SL, Goldstein JM, Laird N, et al. Socioeconomic disadvantage and neural development from infancy through early childhood. Int J Epidemiol. 2015;44:1889–99.
Hammamieh R, Chakraborty N, Gautam A, Muhie S, Donohue D, Daigle BJJ, et al. Whole-Genome DNA Methylation Status Associated with Clinical PTSD Measures of OIF/OEF Veterans (Open Access). US Army Center for Environmental Health Research Fort Detrick United States; (2017).
Dunn EC, Wang M-J, Perlis RH. A Summary of Recent Updates on the Genetic Determinants of Depression. Major Depressive Disorder: Elsevier. (2020)1–27.
Thompson JA, Ziman M. Pax genes during neural development and their potential role in neuroregeneration. Prog Neurobiol. 2011;95:334–51.
Speer K, Upton D, Semple S, McKune A. Systemic low-grade inflammation in post-traumatic stress disorder: a systematic review. J Inflamm Res. 2018;11:111.
Kim TD, Lee S, Yoon S. Inflammation in post-traumatic stress disorder (PTSD): a review of potential correlates of PTSD with a neurological perspective. Antioxidants 2020;9:107.
Katrinli S, Maihofer AX, Wani AH, Pfeiffer JR, Ketema E, Ratanatharathorn A, et al. Epigenome-wide meta-analysis of PTSD symptom severity in three military cohorts implicates DNA methylation changes in genes involved in immune system and oxidative stress. Mol Psychiatry. 2022;27:1720–8.
Al Jowf GI, Snijders C, Rutten BP, de Nijs L, Eijssen LM. The Molecular Biology of Susceptibility to Post-Traumatic Stress Disorder: Highlights of Epigenetics and Epigenomics. Int J Mol Sci. 2021;22:10743.
Smith AK, Ratanatharathorn A, Maihofer AX, Naviaux RK, Aiello AE, Amstadter AB, et al. Epigenome-wide meta-analysis of PTSD across 10 military and civilian cohorts identifies methylation changes in AHRR. Nat Commun. 2020;11:1–9.
Kumsta R. The role of epigenetics for understanding mental health difficulties and its implications for psychotherapy research. Psychol Psychotherapy: Theory, Res Pract. 2019;92:190–207.
Miller MW, Wolf EJ, Sadeh N, Logue M, Spielberg JM, Hayes JP, et al. A novel locus in the oxidative stress-related gene ALOX12 moderates the association between PTSD and thickness of the prefrontal cortex. Psychoneuroendocrinology 2015;62:359–65.
Miller GE, Chen E, Sze J, Marin T, Arevalo JM, Doll R, et al. A functional genomic fingerprint of chronic stress in humans: blunted glucocorticoid and increased NF-κB signaling. Biol Psychiatry. 2008;64:266–72.
Løkhammer S, Stavrum A-K, Polushina T, Aas M, Ottesen AA, Andreassen OA, et al. An epigenetic association analysis of childhood trauma in psychosis reveals possible overlap with methylation changes associated with PTSD. Transl Psychiatry. 2022;12:1–9.
Boukezzi S, El Khoury-Malhame M, Auzias G, Reynaud E, Rousseau P-F, Richard E, et al. Grey matter density changes of structures involved in Posttraumatic Stress Disorder (PTSD) after recovery following Eye Movement Desensitization and Reprocessing (EMDR) therapy. Psychiatry Res: Neuroimag. 2017;266:146–52.
Helpman L, Papini S, Chhetry BT, Shvil E, Rubin M, Sullivan GM, et al. PTSD remission after prolonged exposure treatment is associated with anterior cingulate cortex thinning and volume reduction. Depress Anxiety. 2016;33:384–91.
Zantvoord JB, Diehle J, Lindauer RJ. Using neurobiological measures to predict and assess treatment outcome of psychotherapy in posttraumatic stress disorder: systematic review. Psychother Psychosom. 2013;82:142–51.
Zantvoord JB, Zhutovsky P, Ensink JB, den Kelder RO, van Wingen GA, Lindauer RJ. Trauma-focused psychotherapy response in youth with posttraumatic stress disorder is associated with changes in insula volume. J Psychiatr Res. 2021;132:207–14.
Wang W, Li W, Jiang W, Lin H, Wu Y, Wen Y, et al. Genome-wide DNA methylation analysis of cognitive function in middle and old-aged Chinese monozygotic twins. J Psychiatr Res. 2021;136:571–80.
Liaw CW, Lovenberg TW, Barry G, Oltersdorf T, Grigoriadis DE, de Souza EB. Cloning and characterization of the human corticotropin-releasing factor-2 receptor complementary deoxyribonucleic acid. Endocrinology 1996;137:72–7.
Bale TL, Contarino A, Smith GW, Chan R, Gold LH, Sawchenko PE, et al. Mice deficient for corticotropin-releasing hormone receptor-2 display anxiety-like behaviour and are hypersensitive to stress. Nat Genet. 2000;24:410–4.
Roozendaal B, McEwen BS. Chattarji S. Stress, memory and the amygdala. Nat Rev Neurosci. 2009;10:423–33.
Castro-Vale I, Carvalho D, editors. The pathways between cortisol-related regulation genes and PTSD psychotherapy. Healthcare: MDPI; (2020).
Brenet F, Moh M, Funk P, Feierstein E, Viale AJ, Socci ND, et al. DNA methylation of the first exon is tightly linked to transcriptional silencing. PloS one. 2011;6:e14524.
Tsai P-C, Bell JT. Power and sample size estimation for epigenome-wide association scans to detect differential DNA methylation. Int J Epidemiol. 2015;44:1429–41.
Hoogsteder LM, Ten Thije L, Schippers EE, Stams GJJ A meta-analysis of the effectiveness of EMDR and TF-CBT in reducing trauma symptoms and externalizing behavior problems in adolescents. Int J Offender Ther Comparat Criminol. 2021; 0306624×211010290.
Kooij LH, van der Pol TM, Daams JG, Hein IM, Lindauer RJ. Common elements of evidence-based trauma therapy for children and adolescents. Eur J Psychotraumatol. 2022;13:2079845.
Hannon E, Knox O, Sugden K, Burrage J, Wong CC, Belsky DW, et al. Characterizing genetic and environmental influences on variable DNA methylation using monozygotic and dizygotic twins. PLoS Genet. 2018;14:e1007544.
Armstrong DA, Lesseur C, Conradt E, Lester BM, Marsit CJ. Global and gene‐specific DNA methylation across multiple tissues in early infancy: Implications for children’s health research. The. FASEB J. 2014;28:2088–97.
Hoye JR, Cheishvili D, Yarger HA, Roth TL, Szyf M, Dozier M. Preliminary indications that the attachment and Biobehavioral catch-up intervention alters DNA methylation in maltreated children. Dev Psychopathol. 2020;32:1486–94.
Diehle J, Opmeer BC, Boer F, Mannarino AP, Lindauer RJ. Trauma-focused cognitive behavioral therapy or eye movement desensitization and reprocessing: What works in children with posttraumatic stress symptoms? A randomized controlled trial. Eur Child Adolesc psychiatry. 2015;24:227–36.
Nader KO, Kriegler J, Blake D, Pynoos R, Newman E, Weathers F. Clinician Administered PTSD scale for children and adolescents. National Center for PTSD. 1996.
Verlinden E, van Laar YL, van Meijel EP, Opmeer BC, Beer R, De Roos C, et al. A parental tool to screen for posttraumatic stress in children: First psychometric results. J Trauma Stress. 2014;27:492–5.
Diehle J, de Roos C, Boer F, Lindauer RJ A cross-cultural validation of the Clinician Administered PTSD Scale for Children and Adolescents in a Dutch population. Eur J Psychotraumatol. 2013;4.
Weathers FW, Keane TM, Davidson JR. Clinician‐Administered PTSD Scale: A review of the first ten years of research. Depress Anxiety. 2001;13:132–56.
Verlinden E, van Meijel EP, Opmeer BC, Beer R, de Roos C, Bicanic IA, et al. Characteristics of the Children’s Revised Impact of Event Scale in a clinically referred Dutch sample. J Trauma Stress. 2014;27:338–44.
Perrin S, Meiser-Stedman R, Smith P. The Children’s Revised Impact of Event Scale (CRIES): Validity as a screening instrument for PTSD. Behav Cogn Psychother. 2005;33:487–98.
Chorpita BF, Yim L, Moffitt C, Umemoto LA, Francis SE. Assessment of symptoms of DSM-IV anxiety and depression in children: A revised child anxiety and depression scale. Behav Res Ther. 2000;38:835–55.
Kösters MP, Chinapaw MJ, Zwaanswijk M, van der Wal MF, Koot HM. Structure, reliability, and validity of the revised child anxiety and depression scale (RCADS) in a multi-ethnic urban sample of Dutch children. BMC Psychiatry. 2015;15:132.
Achenbach TM. Manual for the youth self-report and 1991 profile: Department of Psychiatry, University of Vermont Burlington; 1991.
Achenbach TM, Edelbrock CS. Manual for the child behavior checklist and revised child behavior profile. 1983.
Verhulst FC, van der Ende J, Koot JM. Youth Self-Report (YSR): Afdeling Kinder-en Jeugdpsychiatrie, Sophia Kinderziekenhuis/Academisch Ziekenhuis Rotterdam/Erasmus Universiteit Rotterdam; 1997.
Shalev AY, Orr SP, Pitman RK. Psychophysiologic response during script-driven imagery as an outcome measure in posttraumatic stress disorder. J Clin psychiatry. 1992;53:324–6.
Miller R, Plessow F, Kirschbaum C, Stalder TJPM. Classification criteria for distinguishing cortisol responders from nonresponders to psychosocial stress: evaluation of salivary cortisol pulse detection in panel designs. Psychosom Med. 2013;75:832–40.
Pruessner JC, Kirschbaum C, Meinlschmid G, Hellhammer DH. Two formulas for computation of the area under the curve represent measures of total hormone concentration versus time-dependent change. Psychoneuroendocrinology 2003;28:916–31.
Author information
Authors and Affiliations
Contributions
J.E., R.L., P.H., and M.M. contributed to the study concept and design. Material preparation, data collection and analysis were performed by J.E., R.K., and J.Z. J.E., P.H. and A.V. performed the EWAS and statistical analysis. Interpretation of data was done by all authors. The first draft of the manuscript was written by J.E, all authors critically reviewed and approved the final manuscript.
Corresponding author
Ethics declarations
Competing interests
The authors declare no competing interests.
Consent to publication
All authors consented to publication.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary information
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Ensink, J.B.M., Henneman, P., Venema, A. et al. Distinct saliva DNA methylation profiles in relation to treatment outcome in youth with posttraumatic stress disorder. Transl Psychiatry 14, 309 (2024). https://doi.org/10.1038/s41398-024-02892-1
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41398-024-02892-1
- Springer Nature Limited