

Abstract
Intravital 2P-microscopy enables the longitudinal study of brain tumor biology in superficial mouse cortex layers. Intravital microscopy of the white matter, an important route of glioblastoma invasion and recurrence, has not been feasible, due to low signal-to-noise ratios and insufficient spatiotemporal resolution. Here, we present an intravital microscopy and artificial intelligence-based analysis workflow (Deep3P) that enables longitudinal deep imaging of glioblastoma up to a depth of 1.2 mm. We find that perivascular invasion is the preferred invasion route into the corpus callosum and uncover two vascular mechanisms of glioblastoma migration in the white matter. Furthermore, we observe morphological changes after white matter infiltration, a potential basis of an imaging biomarker during early glioblastoma colonization. Taken together, Deep3P allows for a non-invasive intravital investigation of brain tumor biology and its tumor microenvironment at subcortical depths explored, opening up opportunities for studying the neuroscience of brain tumors and other model systems.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Introduction
Glioblastomas (GB) are the most common malignant, primary brain tumors characterized by their infiltrative growth and colonization of the entire normal brain1,2. Furthermore, these tumors are cellularly and molecularly heterogeneous3,4,5,6 with a notorious therapeutic resistance towards standard-of-care treatment with radio- and chemotherapy as well as surgical resection7. It has long been known that GB are predominantly a disease of the white matter8. GB frequently occur in the white matter and can invade into the contralateral hemisphere along the corpus callosum9. Furthermore, a majority of glioblastoma recurrence is detected within the white matter9. Recently, a large autopsy series further highlighted the importance of white matter tracts as an invasion route to invade and colonize the brainstem1. However, intravital studies of glioblastoma in the white matter have so far been lacking as technologies for deep, microscopic intravital and longitudinal monitoring were missing. So far, experimental results were entirely restricted to ex vivo analyzes10,11,12. This highlights the need and importance of studying this highly dynamic disease in vivo in the microenvironmental niche of the white matter. Such an approach would allow dissecting the underlying principles of glioblastoma biology including the yet elusive mechanisms of invasion in and into the corpus callosum.
Intravital imaging of brain tumors including gliomas as well as brain metastases has so far been restricted to superficial cortex layers, dictated by two-photon microscopy (2PM) which is fundamentally limited to an effective penetration of ~300-700 μm due to scattering and out-of-focus fluorescence background3,6,13,14,15,16,17. Deep in vivo brain imaging with 2PM in the mouse brain is currently only possible using highly invasive methods such as gradient index lens implantation or cortical aspiration. These methods have not yet been used to investigate intravital brain tumor biology because of the associated invasiveness.
In this respect, three-photon microscopy (3PM) has shown potential for deeper imaging beyond 1 mm in the brain due to an increased signal-to-background ratio and longer wavelength excitation which reduces tissue scattering18,19. So far, 3PM has been used to investigate the static morphology of brain vasculature and neurons18,20, and to perform calcium imaging of neurons21 and astrocytes22. Yet, their application to investigate spatiotemporally dynamic brain tumor biology poses additional technical challenges. In particular, tumor masses are opaque and dense structures, leading to a significant scattering of light23. Furthermore, the fluorescent membrane labeling of tumor cells required to visualize fine processes3 leads to intrinsically low fluorescence signals. This is exacerbated by the relatively high density of the labeled cells within a given imaging volume, which makes distinguishing and tracking of individual tumor cells and their fine cellular processes difficult. With this comes the requirement of long-term stability of the microscopy system retaining high spatial resolution during extended volumetric time-lapse imaging and the ability to track the same image regions over days and weeks, all while retaining high spatial resolution. Lastly, photodamage- and toxicity need to be prevented by diligent optimization of laser power while keeping a reasonable signal-to-noise ratio (SNR), and an efficient screening system needs to be established that allows to identify both appropriate tumor regions and the white matter niche within a minimal time, in order to utilize imaging time and laser power diligently.
As outlined above, a predominant challenge in deep tissue imaging, including 3PM, is the low SNR of the raw image data. Here, computational methods using deep learning-based image restoration24,25 have the potential to substantially increase the low SNR of images typically acquired in deep tissue conditions. While now increasingly applied to images obtained with confocal, light-sheet, or two-photon microscopy setups3,25, these methods have not yet been adapted and customized to the peculiar detector noise and background sources typically encountered in 3PM. Another opportunity of 3PM lies in the label-free imaging of blood vessels and myelinated axonal tracts in the brain using the third-harmonic generation signal. As these are both important structures of the tumor and brain microenvironment, it would be desirable to unequivocally demix and classify the THG signal into both anatomically distinct structures.
To overcome all of the above-mentioned fundamental technical limitations, we further adapted and advanced state-of-the-art 3PM and image analysis methods to enable the study of glioblastoma biology in the deep gray and white matter. In particular, we developed a minimally invasive intravital imaging and analysis workflow for modified patient-derived glioblastoma xenograft models stably transduced with a membrane-bound green fluorescent protein. Using bespoke 3PM, adaptive optics, and AI-enhanced post-processing and analysis tools, we demonstrate the capability to investigate glioblastoma biology and its tumor microenvironment in the deep white and gray matter of the living mouse, up to a depth of 1.2 mm and at near diffraction-limited spatial resolution. Deep3P time-lapse imaging allowed us to uncover distinct behavioral differences of glioblastoma in white matter tracts as compared to the gray matter of the cortex. Furthermore, we were able to track myelinated axonal fibers over weeks and characterize how they change in the course of early white matter glioblastoma colonization. In particular, Deep3P allowed us to compare invasion patterns in the whole cortex and subcortical white matter, which revealed an enrichment of a vascular invasion route from the cortex into the corpus callosum as compared to intracortical invasion. Within the corpus callosum, we found that glioblastoma cells and their neurite-like processes predominantly align with white matter tracts and follow the anatomical structure of myelinated axons. Surprisingly, we uncovered two additional vascular invasion mechanisms within the corpus callosum allowing invasion orthogonal to the fiber tracts similar to patterns from oligodendrocyte and astrocytic precursor cells during neurodevelopment26,27. Lastly, the third harmonic generation (THG) signal of our Deep3P methodology permitted to dynamically investigate white matter disruptions during glioma infiltration of the corpus callosum. Here, morphometric characterization of the THG signal indicated a potential imaging biomarker of white matter disruption during early glioblastoma colonization that extends the applicability of our workflow to investigate axonal degeneration in other tumor and disease models.
In this work, we introduce Deep3P, a specialized intravital microscopy and artificial intelligence-based methodology that facilitates routine deep imaging of glioblastoma over extended periods. This approach combines three-photon microscopy and adaptive optics with deep learning-based denoising and machine-learning segmentation, to allow detailed investigation of tumor biology up to 1.2 mm depth. Through Deep3P, we reveal the predominance of perivascular routes for glioblastoma invasion into the corpus callosum and identify two distinct vascular mechanisms of tumor migration within the white matter, enhancing our understanding of glioblastoma invasion and potential diagnostic markers. Overall, Deep3P enables an efficient and non-invasive exploration of brain tumor biology and its microenvironment within the deep white and gray matter of a living mouse, presenting further possibilities for advancing the neuroscience of brain tumors and other related model systems.
Results
Infiltration of the corpus callosum as a hallmark of glioma
Previous research has suggested that infiltration into the white matter tracts may be an important route for glioblastoma to invade the contralateral cortex12,28. However, it was not clear how prevalent the infiltration of the corpus callosum is in a human patient glioma cohort.
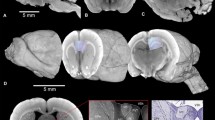
To address this question, we analyzed a large brain tumor patient autopsy cohort (n = 50 patients) and found that 84% of all glioma patients showed at least microscopic infiltration into the corpus callosum irrespective of the location of their primary clinical manifestation (Fig. 1a, Supplementary Fig. 1a–e, Supplementary Data 1). Further analysis showed that 82% of patients with isocitrate dehydrogenase (IDH)-wild-type (n = 42 patients) and 100% of patients with an IDH-mutant glioma showed corpus callosum infiltration. In addition, we studied 20 patient-derived xenograft as well as patient-derived organoid xenograft glioma models and found that all of the analyzed models demonstrated a microscopically visible, infiltrative behavior of the corpus callosum after 40–60 days of implantation, regardless of whether the tumor was implanted into the cortex or striatum (Fig. 1a, Supplementary Fig. 1f, Supplementary Data 2). These findings indicate that white matter infiltration of the corpus callosum is a hallmark of glioblastoma growth, highlighting the need for intravital imaging of glioblastoma invasion and colonization inside the distinct microenvironment of the corpus callosum (Supplementary Fig. 1g), which is yet unattainable with current light microscopy technologies.

a Left: Autopsy brain slice of a patient with IDH-WT glioblastoma. The corpus callosum (CC) is labeled with a dashed line. Arrowhead indicates main tumor mass; arrow indicates tumor infiltration in the CC as seen in the inset. Middle: Tumor infiltrated CC in S24 PDX mouse model. The CC is labeled with a dashed line, fluorescently labeled glioblastoma cells (GBMCs) are shown in green. Arrow indicates glioma infiltration of the CC. Right: Percentage of CC infiltration in human autopsies (n = 43 patients) and in PDX models (n = 23 PDX models). b Workflow of Deep3P. Upper left: Establishment of in vivo patient-derived glioma xenograft model. GBM cell lines are derived from patients and stably transduced with mGFP and injected into mice brain. Upper right: Combination of 2PM and 3PM to identify target regions for deep brain imaging. Bottom: Deep brain imaging in the CC and subsequent deep and machine learning based post-processing to allow simultaneous myelin, vessel and tumor analysis. Created with BioRender.com released under a Creative Commons Attribution-NonCommercial-NoDerivs 4.0 International license (https://creativecommons.org/licenses/by-nc-nd/4.0/deed.en).
Deep3P as a workflow to investigate deep brain tumor biology
To investigate glioblastoma biology in the cortex and subcortical white matter tracts, we used state-of-the-art patient-derived xenograft models that accurately reflect glioblastoma molecular characteristics as analyzed with methylation array analyzes and the histological growth patterns of glioma patients (Supplementary Fig. 1f, Supplementary Data 2). To visualize the fine processes of glioblastoma cells (GBMCs) in vivo, we stably transduced them with lentivirally packaged membrane-bound GFP (mGFP) and injected them into the cortex of adult mice at a depth of approximately 500 µm (Fig. 1b). We used two-photon microscopy (2PM) to longitudinally screen the injected cells up to a depth of 600-700 µm in large diagonal field of views of up to 4.3 mm, taking advantage of the higher imaging speed of 2PM for mapping dynamic glioblastoma growth and its tumor microenvironment in superficial layers to select appropriate regions for deep imaging (Fig. 1b).
We then used our Deep3P methodology (Fig. 1b) to study the previously identified glioblastoma infiltration zone and investigate glioblastoma invasion, proliferation and colonization in the corpus callosum up to a depth of 1200 µm, which is far outside the reach of 2PM (Fig. 2a). In particular, 3 PM results in significantly enhanced signal-to-noise (SNR) ratios at all imaging depths (Fig. 2b). Deep3P leverages recent technological advances in 3PM and the addition of modal-based, indirect adaptive optics22 to optimize fluorescence signal and ensure near diffraction-limited performance at large imaging depths. The system is based on a custom-built 3P laser scanning microscope that was specifically optimized for 1,300 nm excitation and the use of short-pulsed (~70 fs), low repetition-rate (<1 MHz) lasers that are critical to obtain large imaging depths20,22. To improve the 3PM signal and imaging resolution, we employ a custom, modal-based indirect adaptive optics (AO) approach to correct for wavefront aberrations occurring due to the refractive index (RI) differences between the water-immersion, cranial window and brain, as well as brain tissue intrinsic RI inhomogeneities22. To successfully apply this approach to the requirements of intravital and longitudinal monitoring of cellular dynamics and white matter structural changes, the following challenges had to be met: (1) The entire imaging system and workflow were optimized for non-invasiveness, so that deep-brain time-lapse imaging over multiple hours did not induce photo-damage in terms of bleaching and/or toxicity. This also included adaptations to the cranial window surgery, mounting to minimize light loss due to reflections and absorption, as well as the establishment of an optimized imaging workflow (see Methods, Supplementary Methods and Supplementary Note 1). (2) The fast AO correction measurement speed, as well as robustness of the modal-based indirect AO approach22, was paramount to enable time-lapse volumetric recordings with little time overhead, while minimizing the overall light-exposure (Fig. 2c). Here, AO led to an average improvement in the effective resolution of 1.9 ± 0.2-fold (n = 5 image volumes, see Methods) and a 3-5-fold enhancement of fluorescence signals, as evidenced by intensity line plots and spectral power map analysis of lateral resolution (Fig. 2d–f). As a result, fine biological structures such as neurite-like processes of glioblastoma cells called tumor microtubes (TMs), blood vessels and cell nuclei can be resolved (Fig. 2g).

a 3D renderings showing 2PM and 3PM imaging down to approximately 1000 µm. Gamma values were adjusted for 3D visualization. b SNR of 2PM and 3PM along depths, normalized to the SNR at the brain surface. The dashed red line in (a, b) indicates the imaging depth at which biological structures cannot be clearly discerned anymore in 2PM in contrast to 3PM (approximately 450 µm). c Scheme of 3PM and adaptive optics setup. Modified from Fig. 1a in Ref. 22. d Exemplary GBMC imaged without (top left) and with (bottom left) AO optimization and the corresponding images on the THG channel (top and bottom right). The line indicates the line segment averaged over to produce the line profiles in (e), (n = 6 experiments with similar results) showing the effect of uncorrected optical aberration on the visibility of fine cellular structures. The inset on each panel shows the frequency domain power spectrum of the image, with the ring indicating 1 µm length scale. e Line profile comparisons for both mGFP and THG channels showing intensity enhancement. f The averaged radial profile of the frequency maps is shown, allowing easier estimation of the respective frequency cut-offs. g Exemplary 3PM images with and without AO optimization on the mGFP channel at different depths in deep cortex and CC (left and middle) and on the THG channel within the CC (right). Zoom-ins are shown on the right. Arrowheads show TMs (left), a shadow of a cell nucleus that is caused by the membrane-bound GFP labeling (middle), and a blood vessel (right) on AO on and off images. TMs, the cell nucleus, and the blood vessel are not clearly visible without AO (n = 6 experiments with similar results). Source data are provided as a Source Data file.
AI-based image denoising and analysis of Deep3P data
To further improve the signal-to-noise ratio of the raw images during time-lapse imaging of brain tumor cells while keeping 3PM excitation power minimal, we implemented a customized deep learning-based denoising workflow that can also account for the peculiar, structured noise often encountered in 3PM images (Fig. 3a–c, Supplementary Fig. 2a–d).

a Top left: 3D rendering of a stack going from the surface down to the CC. Red: blood vessels, blue: CC, green: GBMCs. Based on probability maps. Top right: Comparison of raw (left) and denoised (right) images within the CC (dashed lines on the left indicate imaging depth). Arrowheads in the mGFP image point to a GBMC soma that can be barely seen without denoising. Arrowheads in the THG signal point to fibrous structures that can be clearly identified after denoising. Bottom: signal-to-noise ratio (SNR) comparison of raw and denoised in THG and mGFP signal (two-sided Mann-Whitney test, n = 114 slices for each mGFP and THG signal, shown as median +/- quartile, whiskers: min/max within 1.5 IQR). b Left: Exemplary raw and denoised images of GBMCs at different depths. Right: SNR in raw and denoised images across entire image stack. (n = 6 experiments with similar results) c Comparison of the denoised 3PM-N2V image (left) and its version with additional application of the PerStruc-Denoiser (middle) showing the qualitative improvement corresponding to a 3 dB increase in SNR allowing a clearer identification of TMs (arrowhead). Arrows point to structured noise. Right: Averaged line power spectrum of the images depicting the PerStruc-Denoiser’s suppression of the main components of the periodic structured noise (see arrows pointing to its main components). d 3D renderings based on raw images, denoised images, probability maps based on raw images and probability maps based on denoised images. e Close-up 3D renderings of single GBMCs based on probability maps. The arrow heads on the zoom-ins point at small processes (top images and bottom left image) and a TM branching point (bottom right image). Gamma values were adjusted for 3D visualization in (a, d, e). Source data are provided as a Source Data file.
In particular, we developed a custom 3D variant of the Noise2Void approach24 for the denoising of 3PM data, which utilizes a 3D U-Net architecture (Supplementary Fig. 2a). Our method, referred to as 3PM-Noise2Void, effectively improves the image quality by taking into account the 3D nature of the data. This workflow increased the SNR by an additional ~10-15 dB across all imaging depths (Fig. 3a, b, Supplementary Movie 1). Furthermore, to counteract periodic structured noise patterns that cannot be addressed by Noise2Void, we implemented an additional post-processing technique, called PerStruc-Denoiser, to further reduce this structured noise effect by ~3 dB (Fig. 3c, Supplementary Fig. 2). Lastly, we implemented a machine learning-based segmentation using the denoised data as a prediction mask for further biological analysis (Fig. 3d). Furthermore, we highlight that denoising on 3PM images without AO resulted in qualitatively and quantitatively worse image quality compared to 3PM images with AO correction (Supplementary Fig. 3a–d). The effective increase in image SNR due to our bespoke AO and AI-based denoising is key for our Deep3P workflow. It allowed to keep excitation power and hence photodamage low during the extended, up to 4 h long, time-lapse image acquisitions. In effect, this maintained and ensured the non-invasiveness of our imaging approach. Overall, the improvements in resolution and SNR of Deep3P across all imaging depths were critical to resolve the fine, neurite-like, cellular protrusions of the glioblastoma cells (Fig. 3e, Supplementary Fig. 4a–c) and to qualitatively as well as quantitatively distinguish between different cellular migration and invasion patterns in vivo.
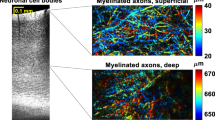
Our Deep3P imaging system permits acquisition of two separate channels, which were used for mGFP to visualize tumor cells, and to record the THG signal. The latter yields label-free image contrast that visualizes both myelinated white matter tracts in densely and sparsely myelinated regions (Fig. 4, Supplementary Fig. 5a–c), as well as blood vessels in the brain29,30. To further confirm that the THG signal captures myelin, we performed correlation analyzes between myelin fibers labeled via MBP and THG signal in regions differing in myelin fiber density (Supplementary Fig. 5c). Since white matter tracts and blood vessels constitute important tumor microenvironmental niches8,28, we aimed at unambiguously differentiating the THG signal.

a Scheme for customized machine learning based classification of THG signal into myelin and vessel signal. b Distribution of uncertainty level with machine learning compared with customized machine learning (left) and statistical comparison of uncertainty levels (n = 239568 THG pixels, two-sided Mann–Whitney test, shown as median +/- quartile, whiskers: min/max within 1.5 IQR). c Exemplary image of THG signal without (top) and with (bottom) predicted labels of blood vessels and myelin fibers. Arrowheads indicate blood vessels; arrow indicates myelin fibers (n = 3 experiments with similar results). d 3D rendering within the CC, illustrating the results of the machine learning-based classification. Arrowhead indicates vessels, arrow indicates myelin fibers. e Validation of ML-classification for blood vessels with FITC as fluorescent dye colored in green. The arrowheads point at vessels (n = 3 experiments with similar results). f Comparison of high and low FITC signal with THG signal (n = 62244 pixels, two-sided Mann–Whitney test, shown as median +/- quartile, whiskers: min/max within 1.5 IQR). g Histogram of measured blood vessels based on their diameter and colored based on their identification from THG signal (blue: visible with FITC and in THG signal, green: visible only with FITC, n = 68 vessels). h UMAP based on pixel features that are different between background, myelin, and vessels based on the machine learning prediction (n = 100 features). Pixels are colored based on the local frequency of pixels in the dimensionality-reduced space (n = 239568 pixels). i Close-up 3D rendering of a single GBMC (green) and its surrounding microenvironment (vessel in red, myelin in blue). The asterisk points at a vessel branching points, the arrow at a TM branching point and the arrowhead at a glioblastoma small process. Gamma values were adjusted for 3D-visualization in (d, i). Source data are provided as a Source Data file.
To this end, we trained a customized, interactive machine-learning algorithm based on ilastik31 (see Methods) to distinguish between the background, blood vessels, and myelinated axonal tracts (Fig. 4). This enabled us to investigate the bidirectional relationship between brain tumor cells and the blood vessel and white matter microenvironmental compartments (Fig. 4a–d, Supplementary Movie 1–3). We validated our AI-classifier by intravenous injection of FITC-dextran as ground truth signal for blood vessels (see Fig. 4e–g, Supplementary Fig. 5d and Methods). This workflow allowed to clearly distinguish between blood vessels and myelinated axonal tracts both in 2D (Fig. 4c) and 3D rendering (Fig. 4d).
We further corroborated that the THG signal carries distinct information in its pixel distribution of blood vessels and myelinated white matter tracts using a dimensionality reduction analysis using uniform manifold approximation and projection (UMAP) (Fig. 4h, see Methods). Comparing our customized machine-learning workflow to the standard machine learning pipeline, we could observe that the amount of pixels classified with high uncertainty significantly decreased (Fig. 4b). Lastly, we validated the classifier with human annotation (Supplementary Fig. 5a). Taken together, this workflow enables us to analyze both tumor cells and their microenvironment of blood vessels and myelinated axonal tracts at the same time using a single fluorescence detection channel (Fig. 4f, Supplementary Fig. 5c). The near diffraction-limited resolution of Deep3P allows to clearly identify TMs and the even finer class of neurite-like structures called small processes3 in the corpus callosum (Fig. 4i, Supplementary Fig. 4c).
Dynamic deep brain investigation of glioblastoma invasion
Our Deep3P imaging workflow and analysis revealed a number of distinct glioblastoma invasion and colonization patterns within the deep cortex and white matter as well as microenvironmental changes in the white matter in vivo that were enabled by the near diffraction-limited resolution and deep imaging depth.
In the corpus callosum, a majority of glioblastoma cells were aligned with myelinated axonal tracts, both with respect to their cell somata and their TMs (Fig. 5a–d) indicating distinct morphological adaptations of glioblastoma cells in the cortex and corpus callosum, depending on the tumor microenvironment. These adaptions are similar to those by physiologically resident cells in the corpus callosum (Supplementary Fig. 5e–g). Statistical comparisons of glioblastoma cell and TM directionality analyzes revealed a significant correlation of tumor cell directionality with its microenvironment (Fig. 5d). Cell polarity in the cortex showed no clearly structured direction, while in the corpus callosum more than 60 percent of cells grew parallel to the myelin fibers in an angle smaller than 30° (Fig. 5d), without a correlation between myelin and vessel orientation (Supplementary Fig. 5h–j, Supplementary Fig. 6a–e). Interestingly, a subpopulation of glioblastoma cells was found to be not aligned with the corpus callosum fibers (Fig. 5a–d). Next, we used Deep3P and its stable time-lapse deep brain imaging ability to analyze how glioblastoma cells could invade from the deep cortex into the distinct microenvironment and white matter-rich structure of the corpus callosum (Supplementary Fig. 1a). We found that there is a significant enrichment of perivascular invasion of glioblastoma cells to enter the corpus callosum as compared to perivascular invasion prevalent within the cortex (Fig. 5e, f). An average of 60 percent of glioblastoma cells use vessels as a track to enter the corpus callosum while less than 40 percent of glioblastoma cells in the cortex show a perivascular migration pattern, despite the lower vascular density in the corpus callosum (Supplementary Fig. 7a, b). This illustrates how tumor cells are able to adapt their invasion strategy making use of blood vessels to invade the white matter. Furthermore, Deep3P revealed two vascular mechanisms within the corpus callosum that allowed an invasion that is orthogonal to the myelinated axonal tract direction. First, we discern that glioblastoma cells could extend their TMs to attach to vessels, revealed by the high spatial resolution afforded by the use of our AO (Fig. 5g–i). Using this fixed point, glioblastoma cell somata can then translocate their soma towards the vessel as an invasion mechanism. Furthermore, we could even more often observe perivascular invasion as an alternative vascular route (Fig. 5g–i). Most vessels to which tumor cells attached to had vessel diameters typical of capillaries32 (Fig. 5j). We confirmed the structural relationship between TMs, glioblastoma cell somata, and vessels with three-dimensional electron microscopy reconstructions in a patient-derived xenograft model, validating the ability of Deep3P to uncover biology deep in the brain at high resolution (Supplementary Fig. 7c). In the patient-derived glioblastoma xenograft models used in this study to investigate the early stages of brain tumor colonization, no blood-brain barrier disruption was observed (Supplementary Fig. 7d, e) and blood vessel architecture stayed stable over weeks as evidenced by intravital imaging (Supplementary Fig. 7f–h).

a Top: Maximum intensity projections (MIP) of regional overviews of GBMCs in the CC and cortex. Arrowheads: exemplary cells parallel to the myelin fibers (angle <30°), asterisks: exemplary non-parallelly oriented GBMCs. Bottom left: THG signal only. Dashed: myelin fiber direction, white symbols: exemplary GBMCs aligned with white matter tracts, red symbols: exemplary non-parallelly orientated cells. Bottom right: red and white symbols represent exemplary GBMCs in the region shown above (vessel channel). b Directionality analysis of tumor cell regions in CC and cortex (n = 355 GBMCs, n = 12 experiments, n = 10 mice, 2 PDX models). Dashed: myelinated fiber direction within CC. c Rose plot of tumor cell directionality of regions in (a), in CC and cortex (n = 67 GBMCs, n = 2 experiments). d Comparison of predominant angle direction of cell polarity (n = 211 and n = 164 GBMCs in the CC and cortex, respectively, n = 13 experiments, n = 11 mice, 2 PDX models; two-sided Mann-Whitney, shown as median +/- quartile, whiskers: min/max within 1.5 IQR). e Schematic and MIP of a GBMC (soma encircled in white) using vessels to invade into the CC. Dashed: invasion direction along blood vessel. Vessels, tumor cells and myelinated fiber shown as probability maps. f Percentage of GBMCs showing perivascular invasion into the CC as compared to within the cortex (n = 206 GBMCs, n = 14 experiments, two-sided Mann-Whitney test, shown as median +/- quartile, whiskers: min/max within 1.5 IQR). g Schematic drawing and h example of tumor cells using vessel related invasion mechanisms. Dashed: invasion direction. Vessels and tumor cells shown as probability maps. CC signal was post-processed by denoising. i Histogram of angles between perivascular cells and myelin fiber orientation. (n = 29 GBMCs, n = 8 experiments, n = 6 mice, 2 PDX models). j Histogram of blood vessel diameter of perivascular cells in CC (n = 29 GBMCs, n = 8 experiments, n = 6 mice, 2 PDX models) and cortex (n = 54 GBMCs, n = 7 experiments, n = 7 mice). PDX models: S24 and T269. Source data provided as Source Data file.
Interestingly, these invasion patterns resemble migration patterns of oligodendrocytic and astrocytic precursor cells during neurodevelopment26,27. This shows that not only cell-intrinsic mechanisms of neural precursor cells are hijacked by glioblastoma cells for invasion but also that their microenvironmental dependencies are phenocopied.
Subcellular dynamic behavior of TMs including branching, protrusion, and retraction, previously only described in the superficial cortex3, were observed in both deep gray and white matter (Fig. 6a, b, Supplementary Fig. 8a). Interestingly, branching of TMs was reduced within the corpus callosum, potentially indicative of a more directed movement pattern of TMs along white matter tracts as compared to a scanning behavior of TMs prevalent in the superficial layers of the cortex3. Furthermore, three distinct invasion patterns of locomotion, branching migration and translocation could be detected within the corpus callosum, thanks to the superior spatial resolution and depth penetration of Deep3P (Fig. 6c, d, Supplementary Movie 4). These migration mechanisms reflect conserved neuronal mechanisms of invasion that can be seen during neuronal development3,33. Interestingly, this is congruent with previous observations in the superficial layers of the cortex3. However, in contrast to invasion within the cortex branching migration within the white matter is significantly reduced in line with reduced branching behavior of the TMs (Fig. 6d–f). Between PDX models, the invasive patterns were similar (Supplementary Fig. 8b-e) on a cellular and subcellular level. It will be important to characterize the distinct molecular mechanisms of these three invasion phenotypes in their microenvironmental niches of the gray and white matter which is now possible via Deep3P. While these mechanisms are phenotypically distinct, their invasion speed did not significantly differ (Fig. 6e).

a Representative MIP time-lapse images showing TMs in the CC that use branching, protrusion, or retraction. Asterisks point at the GBMC somata, arrowheads point towards the tips of the TMs of interest. Dashed arrows: direction of the TM dynamic. b Distribution of TM dynamics in the CC and comparison with cortex using branching, protrusion or retraction (n = 163 cells from n = 14 datasets in 12 mice in 2 PDX models (S24 and T269)). c Representative MIP time-lapse images of invasion phenotypes in CC showing locomotion, translocation and branching migration. Dashed arrows: invasion direction. The straight line in the first translocation image indicates the stable location of the TM tip throughout all images. d Distribution of invasion phenotypes in the CC compared to the cortex (n = 99 invasive cells from n = 14 experiments in 13 mice, two-sided Mann-Whitney test, shown as median +/- quartile, whiskers: min/max within 1.5 IQR). e Speed comparison of invasion phenotypes in CC (n = 66 GBMCs from n = 7 experiments in 6 mice, two-sided Mann-Whitney test, shown as median +/- quartile, whiskers: min/max within 1.5 IQR). f Mean squared displacement is shown over time (n = 108 GBMCs, error bars indicate mean +/-s.e.m). Images were post-processed as probability maps and using the “smooth” function in ImageJ/Fiji in (a, c). Source data are provided as a Source Data file.
Glioblastoma network formation and tumor cell proliferation in the white matter
In addition to gap junction-coupled tumor-tumor networks that were so far exclusively described in the gray matter3,6,15,34,35,36, we could also observe how tumor-tumor networks in the white matter are formed (Fig. 7a–c, Supplementary Fig. 9a, Supplementary Movie 5). In contrast to tumor-tumor networks in the gray matter, these tumor-tumor networks are to a majority aligned with the white matter fiber direction of the corpus callosum as compared to the cortex (Fig. 7b, c). This illustrates how tumor network formation respects the anatomical boundaries and integrates into the peculiar microenvironment of the corpus callosum (Fig. 7a–c). In addition to investigating invasion and tumor network formation, Deep3P could also be used to characterize the specialized patterns of glioblastoma cell division qualitatively and quantitatively within the deep, native microenvironment of the corpus callosum (Fig. 7d, Supplementary Fig 9b, Supplementary Movie 6).

a Brain tumor networks in the CC (left) and in the cortex (right) shown as 3D renderings in green. CC and cortex imaging were performed with 3PM and 2PM, respectively. Each network is visualized as network orientation in the bottom right corner. The network is colored based on the local orientation. CC imaging depth: z = 840-950 µm. b Rose plot of overall orientation of the tumor cell network in the CC (n = 271985 local network orientation values from n = 6 brain tumor networks from n = 5 mice). c The standard deviation of tumor network orientation is compared in the CC to the cortex. (n = 692 slices from n = 6 brain tumor networks from n = 5 mice, Mann-Whitney test, shown as median +/- quartile, whiskers: min/max within 1.5 IQR). d Top: MIP time-lapse imaging of GBMC division in the CC. White arrowhead: GBMC before cell division. Yellow and purple arrowhead: Daughter GBMCs after cell division. The asterisk points at a newly grown TM after cell division. Post-processed with denoising and “clear outside” function in ImageJ/Fiji. Bottom: 3D rendering of another cell in the CC before and after cell division. The arrowheads point to the somata of the cell before division and the two daughter cells after division, the arrows point to the TMs (n = 18 cell divisions from n = 7 experiments in 6 mice in 2 PDX models (S24 and T269)). Gamma values were adjusted for 3D-visualization in (a, d). Source data are provided as a Source Data file.
White matter disruptions during early glioblastoma colonization
Lastly, we investigated the role of glioblastoma invasion and early colonization in the corpus callosum on myelinated fiber tracts. Evaluating white matter integrity during glioma infiltration with clinical imaging such as magnetic resonance imaging (MRI) would be a useful bioimaging marker for estimating whole-brain colonization of glioma. It has been proposed that diffusion tensor imaging (DTI) enables imaging of early tumor infiltration into the corpus callosum37. However, a comparison to the ground truth of tumor invasion and an exact characterization of tumor density was not possible in these studies. Here, we first examined a patient cohort of eight IDH-wildtype glioblastoma patients that showed macroscopic affection of the corpus callosum on clinical MRI. We quantified the apparent diffusion coefficient (ADC) of the affected ipsi- and non-affected contralateral corpus callosum. Here, we could not detect any significant differences in the ADC values in the different parts of the corpus callosum (Fig. 8a, b, Supplementary Fig. 10a). This suggests that standard clinical diffusion imaging at 3 Tesla is not able to reveal the early glioblastoma white matter colonization. To investigate this phenomenon during early glioblastoma invasion and colonization, we longitudinally performed MRI scans of tumor-infiltrated mouse brains (Fig. 8c) and analyzed the fractional anisotropy of different regions in the corpus callosum. We started with a semi-automatic segmentation of the corpus callosum using tractography (Fig. 8d). Over weeks after tumor infiltration, no changes in fractional anisotropy, mean diffusivity and axial diffusivity were observed (Fig. 8e). Further, no differences in these measures between ipsi- and contralateral corpus callosum were detected at any time point up to 73 days after tumor injection.

a Representative MR images of two patients with IDH-wildtype glioma with affection of the CC (T1CE and ADC). b ROI-based quantification of apparent diffusion coefficient (ADC) on diffusion imaging (n = 8 glioma patients) in ipsi- and contralateral CC, (two-sided Mann-Whitney-Test, shown as median +/- quartile, whiskers: min/max within 1.5 IQR and data points). c Axial MR image showing fractional anisotropy map in a glioma-infiltrated mouse brain (left) showing the CC as a region of interest (red) which is transformed out of the tract (d). Right: Zoom in CC (PDX model: S24). d Tracts of the body of CC, extracted from respective DTI image stacks and transformed into ROIs. e Mean fractional anisotropy (FA), mean diffusivity (MD) and axial diffusivity (AD), in mice MRI scans at multiple time points in CC. (n = 42 MR acquisitions (26 days (n = 5), 33 days (n = 7), 60 days (n = 12) and 73 days (n = 13) after injection)). f Regions with and without tumor infiltration analyzed in the CC. mGFP and source THG signal post-processed with denoising. g Morphological parameter (circularity) of tumor-infiltrated holes and non-infiltrated holes (n = 27 cells from n = 3 experiments in 3 mice in 2 PDX models (S24 and T269), two-sided Mann-Whitney test, shown as median +/- quartile, whiskers: min/max within 1.5 IQR). h Left: CC fibers in tumor-infiltrated regions compared to non-infiltrated regions. Images post-processed with denoising. Right: The orientation plot of fibers is shown. i Comparison of orientation of brain tumor free tissue with tumor-infiltrated regions. (n = 29 sample regions from n = 8 mice (S24 and T269), two-sided Mann-Whitney test, shown as median +/- quartile, whiskers: min/max within 1.5 IQR). j White matter displacing (left) and non-displacing (right) GBMC somata and TMs. Arrowheads show tumor-infiltrated-holes. Asterisks show a soma and a TM and their location in the corresponding THG signal showing no displacement. Images post-processed with denoising. k Comparison of a portion of displacing TMs to displacing GBMC somata (n = 33 GBMC somata and n = 22 TMs from n = 4 mice in 2 PDX models (S24 and T269), two-sided Mann–Whitney test, shown as median +/- quartile, whiskers: min/max within 1.5 IQR). Source data provided as Source Data file.
We were able to quantify changes in the white matter tracts that were disrupted by glioblastoma cell invasion in the early stages of glioblastoma colonization using our customized machine learning approach of white matter classification (Fig. 8). We found that the shape of THG discontinuities could potentially serve as an imaging biomarker of early glioblastoma cell invasion into white matter tracts with potential clinical-translational implications. While tumor glioblastoma cell-infiltrated holes in the white matter showed a significantly increased roundness, the holes caused by resident cells of the tumor microenvironment were more oval-shaped in the corpus callosum (Fig. 8f, g), with similar overall ranges of circularities (Supplementary Fig. 10b). However, the white matter tract directionality did not change significantly in the early stages of glioblastoma colonization (Fig. 8h, i), potentially also explaining difficulties of modern clinical imaging to delineate white matter glioma infiltration38. Additionally, we observed that TMs, the fine neurite-like protrusions, displaced white matter tracts significantly less than their glioblastoma cell soma (Fig. 8j, k). This might indicate that the TMs are used to scan and invade the microenvironment without causing major disruptions of its myelinated microenvironment. Taken together, these observations could serve as ground truth for further development of microscopy approaches and high-resolution clinical imaging to detect early glioblastoma invasion.
Discussion
In summary, we developed a tailored intravital imaging workflow that uses 2PM and 3PM together with AO that enabled longitudinal, deep brain tumor imaging up to a depth of 1.2 mm in the living mouse brain. This methodology opens up avenues to study the cancer neuroscience of brain tumors beyond superficial cortex layers in the pathophysiologically highly relevant white matter.
Compared to previous proof-of-principle technical work in 3PM and AO18,20,22,39,40, we optimized and advanced these technologies for longitudinal intravital measurements at the sub-cellular scale in >1 mm depth and over several hours for samples with low SNR. Instrumental to this was the diligent optimization of imaging parameters, conditions, and overall workflow (see Supplementary Note 1 and Supplementary Methods), as well as the addition of an artificial intelligence-driven image restoration workflow adapted to the distinct noise sources of 3 PM to substantially increase image SNR and thereby keep overall excitation light photoburden minimal. Here, we note that, generally speaking, all machine learning methods need to be properly validated and – if needed - retrained or fine-tuned on new experimental data. As similar 3P- AO systems are not yet widely available elsewhere, we could not test or apply our tailored denoising and segmentation pipelines on 3PM data outside our own setups. It is important to note that performance for the same imaging modality can vary depending on multiple parameters, including the model system, tissue properties, and acquisition parameters. As the denoising part of our pipeline is self-supervised and segmentation is supervised weakly, we believe this not to be a barrier for wider adoption.
Overall, this workflow significantly increased the robustness of 3PM ensuring to study white matter glioblastoma biology non-invasively in situ. Lastly, we used customized machine learning to separate the THG signal and thereby clearly distinguish most blood vessels and white matter tracts, enabling simultaneous volumetric time-lapse imaging of tumor cells and their microenvironment. At the moment, the main disadvantage of our Deep3P methodology is the relatively slow acquisition speed, due to constraints associated with laser power, pulse energy, and sample physiology20, as well as AO optimization time overheads, which limits the overall volumetric FOV that can be monitored over time. Yet, the introduction of efficient two-photon microscopy pre-screening as well as the comparably fast modal-based AO procedure22 ensure that biologically interesting brain regions can be effectively and non-invasively imaged over time.
The effectiveness of our Deep3P approach is demonstrated by its ability to track highly dynamic subcellular and cellular processes of glioblastoma in white and gray matter. This approach uncovered different adaptations of glioblastoma cell morphology, directionality, and invasion phenotypes in the corpus callosum as compared to the cortex. For these findings, the superior depth penetration, spatial resolution as well as image SNR was of paramount importance. Our approach also allows for the intravital investigation of tumor biology including proliferation, invasion, and therapeutic resistance across different brain regions and brain tumor entities. Further, the machine learning-based separation of the THG signal allowed the discrimination of blood vessels and the myelinated axons from the raw image data. This analysis approach can be in principle also generalized and adapted to other non-linear microscopy techniques that capture label-free image contrasts of different biological structures.
The understanding of invasion mechanisms deep in the brain and their relationship to the microenvironmental niches of blood vessels and myelinated fibers are fundamental to glioblastoma biology and potential therapies. We find not only a mechanism of vascular translocation that is mechanistically distinct to the perivascular invasion route. In addition, we observe an enrichment of migration along the vascular route into the corpus callosum with different invasion mechanisms prevalent within the corpus callosum. While the route of perivascular migration has been previously described16,17 and was associated with a breakdown of the blood-brain barrier and altered neurovascular coupling, neither vascular translocation at all nor perivascular migration in the corpus callosum was described yet. Interestingly, we observe these phenomena in the very early stages of glioblastoma colonization where the blood-brain barrier is still intact with respect to leakage of Evans Blue. Whether the leakage of smaller molecules remains possible, will be a question for future research. Lastly, we investigated the clinical-translational important question of why standard clinical imaging and even high-field MRI is not able to reliably capture early glioblastoma invasion and colonization with an exploratory study of a human IDH-wildtype glioblastoma patient cohort, and quasi-correlative imaging between high-field MRI and Deep3P in our patient-derived xenograft models. Within the confines of a limited sample size, our investigation of white matter disruptions revealed only minor changes of the overall structure in myelinated white matter tracts. These near diffraction-limited microscopical findings further explain difficulties of clinical imaging and even sophisticated modalities such as diffusion tensor imaging38 to diagnose the earliest stages of glioblastoma colonization in clinical settings. In addition, this quasi-correlative approach can be further used to improve MRI by comparing experimental sequences with the intravital, microscopic ground truth. To overcome hurdles on the way to clinical application, multimodal data integration of preclinical knowledge, microscopic pathology, and high-resolution MRI imaging, potentially with the help of deep learning, will be crucial to understand both the clinical and biological meaning of these observations. Taken together, these are fundamental in our understanding of glioblastoma biology.
In addition to its utility in brain tumor imaging, we envision that our newly developed integrated microscopy and analysis workflow allows for the integration of fluorescent cell state indicators and other subcellular fluorescent reporters with low SNR to simultaneously characterize biological behavior as well as cellular and molecular cell states in the future. In principle, Deep3P can be implemented with multiple fluorescent marker proteins to visualize different cell populations, provided they fall within the 3P excitation wavelength regions41. Furthermore, this method enables the development of correlative technologies deep in the brain, as previously demonstrated for correlative light and electron microscopy, allowing for in-depth characterization of intravital cellular behavior and ultrastructure in a non-invasive manner.
Apart from patient-derived brain tumor models, we believe that our 3PM and analysis workflow can be straightforwardly applied to other disease models such as other brain tumors, extracranial tumors, demyelinating diseases, including imaging of the spinal cord and, as well as across various model organisms to investigate (sub)cellular structure and (patho)physiology with minimal invasiveness and samples with low SNR. Investigating glioma infiltration in model systems with fully functioning immune systems will be a valuable research avenue in the future. Further red-shifted 3PM and/or its combination with wavefront-shaping approaches could lead to even increased imaging depth beyond the corpus callosum to truly enable whole-brain imaging of glioblastoma colonization. Integrating feedback microscopy, also utilizing AI to identify regions of interest, might provide a powerful solution for faster and more automated imaging in the future.
Overall, our approach allows for the investigation of spatiotemporally dynamic brain tumor biology in vivo across the gray and white matter, with the potential to further uncover cellular and subcellular mechanisms in cancer and its microenvironment. This has important implications for our understanding of brain tumors, opportunities to study these devastating diseases, and the development of clinical therapies.
Methods
Experimental models and subject details
All animal studies were performed with adult male NMRI nude mice older than six weeks in accordance with the European Directive on animal experimentation (2010/63/EU) and institutional laboratory animal research guidelines after approval of the Regierungspräsidium Karlsruhe, Germany, the EMBL IACUC (protocol 22004_HD_RP) and the Animal Welfare Structure of the Luxembourg Institute of Health (protocol LRNO-2017-01). The 3 R principles for reducing the number of animals were strictly followed and efforts were made to minimize animal suffering. Animals were scored daily and experiments were terminated in case of weight loss exceeding 10-20%, neurological deficits and signs of pain, tumor size was no termination criterion. For all human tissues, patients have given informed consent and local regulatory authorities have approved (Ethic Committees at the Mannheim and Heidelberg Medical Faculty of the University Heidelberg, protocols (S-206/2005, S-207/2005, S-306/2019, 2018-614N-MA, 2018-843R-MA), the National Committee for Ethics in Research (CNER) Luxembourg (201201/06) and the regionale komiteer for medisinsk og helsefaglig forskningsetikk at the Helse Bergen (protocol 2013/720/REK vest). Informed consent was obtained for all patients who donated their brain post-mortem to the Nervous System Tumor Bank. The protocol was reviewed and approved by the Northwestern University Institutional Review Board (IRB) under study STU00095863. Molecular testing with antibodies against the IDH mutation, ATRX staining, and the Illumina 850k methylation array (Department of Neuropathology, University of Heidelberg) for confirmation of diagnosis were performed. Written informed consent was obtained for all IDH- wildtype glioblastoma patients whose MRI scans were retrospectively reviewed. All examinations were in accordance with the declaration of Helsinki and approved by the local ethics committee of Heidelberg University (study permit: S-078/2021).
No sex-based analyzes were performed in this due to low sample size. The two major cell lines analyzed are from a patient with female sex (S24) and male sex (T269).
Patient-derived primary glioblastoma cell lines and Illumina 850k methylation array characterization
Cultivation of patient-derived tumor cell lines from resected glioblastoma was performed6,15,34,42 in DMEM/F-12 under serum-free, non-adherent, ‘stem-like’ conditions with B27 supplement (12587-010, Gibco), insulin, heparin, epidermal growth factor, and fibroblast growth factor, outlined in detail before15. The molecular classification of glioblastoma xenograft models used in this study can be found in Supplementary Data 2. To obtain the DNA methylation status43 at >850,000 CpG sites in all GBMC lines, the Illumina Infinium Methylation EPIC kit was used at the Genomics and Proteomics Core Facility of the German Cancer Research Center in Heidelberg, Germany according to the manufacturer’s instructions. Glioblastoma cell lines kept under stem-like conditions were transduced with lentiviral vectors for membrane-bound GFP with the pLego-T2-mGFP construct based on Dondzillo et al.44.
Regular FACS sorting of transduced cells was performed with FACSAria Fusion 2 Bernhard Shoor or FACSAria Fusion Richard Sweet and the BL530/30 filter was used for FACS- sorting GFP.
Surgical procedures
Surgical procedures were performed as previously described6,15. Cranial window implantation in mice was done similarly to what we had previously described with small modifications, including a custom-made titanium ring for painless head fixation during imaging and an asymmetric placement of the window above the sinus, allowing optimal imaging accessibility of the corpus callosum on one hemisphere (see Supplementary Methods). 50.000-100.000 tumor cells were stereotactically injected at a depth of 500 μm into the mouse cortex.
Histological analysis of organoid-based patient-derived orthotopic xenograft models
Patient-derived orthotopic xenografts were derived by intracranial implantation of glioma organoids as described in Oudin et al.45. PDOXs were assessed at the histopathological and molecular levels as described in Golebiewska et al.46. Invasion of human glioma cells through corpus callosum was assessed by immunohistochemistry with antibodies against human-specific nestin (abcam, ab6320, 1:500) on coronal 4-8 µm sections from paraffin-embedded brains. Primary antibodies were incubated overnight at 4 °C or 3 h at room temperature, followed by 30 min incubation with secondary antibodies. Signal was developed with the Envision+ System/HRP Kit in 5–20 min (K4007, Agilent/Dako).
Spatial transcriptomics analysis of cortex and corpus callosum
Spatial sequencing data as well as imaging data was downloaded from http://molecularatlas.org based on Ortiz et al.47. Data was processed using Seurat48 and clusters were defined using its FindClusters function. Based on the associated histology images, clusters were assigned to either corpus callosum / white matter, cortex / gray matter, or other brain regions. Clusters were then visualized based on the assignment and using a Uniform Manifold Approximation and Projection (UMAP) in Seurat.
Macroscopic and microscopic histological analysis of glioblastoma patients
The postmortem cohort was collected as described in Drumm, et al.1. Briefly, at the time of brain sectioning, portions of key brain and spinal cord regions, as well as extra sampling of tumor, were collected for histologic processing as paraffin-embedded tissue blocks. Tissue sections of corpus callosum were routinely collected whether gross evidence of tumor involvement was present or not. Each section of corpus callosum was then stained with hematoxylin and eosin and examined for the presence of migrating glioma cells.
Standard immunohistochemistry analysis was performed on patient autopsy specimens to visualize IDH1R132H (Dianova, DIA H09, 1:150) and nestin (Abcam, ab22035, 1:750). To achieve this, four-micrometer thick sections of FFPE tissue on charged slides were baked in the oven at 60 C before being deparrafinized and re-hydrated. Antigen retrieval was performed using a pH6 retrieval buffer (Biocare Reveal). Slides were cooled to room temperature and washed in TBS before neutralizing endogenous peroxidase (Biocare Peroxidase 1). Slides were then treated with a serum-free casein background block (Biocare Background Sniper) before incubation in a 10% goat serum block for 60 min at room temperature. Primary antibody was then added to the slides for overnight incubation at 4 C. After incubation, slides were washed well with TBS-T before incubating in HRP polymer (Biocare MACH 4 Universal HRP Polymer). Finally, reaction products were visualized with DAB (Biocare Betazoid DAB Chromogen Kit). Slides were then counterstained with hematoxylin, dehydrated and mounted with xylene-based mounting media.
Two-photon microscopy
For 2PM imaging, chronic cranial window surgeries were performed on adult male NMRI nude mice (Charles River and Janvier) and patient-derived tumor cells were injected into the cortex. We implemented an approach for chronic cranial window implantation that allows a better accessibility of the corpus callosum by transplanting the window asymmetrically above the superior sagittal sinus (see Supplementary Methods). 2PM was first performed three weeks after surgery using a TriM Scope II microscope (LaVision BioTec GmbH) equipped with a pulsed Ti:Sapphire laser (Chameleon II ultra; Coherent). A 16x, 0.8 NA, apochromatic, 3 mm working distance, water immersion objective was used. Before imaging, TRITC dextran (fluorescent conjugated tetramethylrhodamine isothiocyanate-dextran, 500.000 g/mol) was diluted in 0.9% NaCl- solution at 10 mg/ml and 100 µl injected intravenously into the tail vein for vessel signal. GFP and TRITC were imaged using 960 nm wavelengths. Low-noise high-sensitivity photomultiplier tubes were used for fluorescence emission detection. For the anesthesia, isoflurane gas was diluted in 100% O2 to a concentration between 5% (for anesthesia induction) and 0.5-1.5 % (for maintenance of anesthesia). The breathing rate of the mice was monitored and body temperature was kept stable at 37° using a heating pad. Eye cream was applied before anesthesia.
Two-photon microscopy prescreening
After tumor cell injection into the cortex we screened for tumor growth regularly with 2PM, considering the advantage of faster imaging speed. Tumor growth was assessed regarding dissemination and density of tumor cells. 3 P imaging was initiated when tumor cells could be detected up to a z-depth of app. 700 μm. We additionally screened the contralateral hemisphere, where no tumor cells had been injected prior as tumor cell dissemination in the contralateral hemisphere is an indicator for infiltration into the CC. An area of interest is then chosen, taking into account the probable accessibility of CC, probable tumor cell density and presence of superficial landmarks for recognition. To facilitate 2P-3P- correlative imaging, we mapped the macroscopically visible vasculature to the 2PM- tile scans, which later allows faster recognition of the area of interest. We give a more detailed description of the 2PM-3PM workflow in the Supplementary Methods file.
For whole brain tile scan imaging, repetitive stacks of 2PM images were acquired with a field of view of 694 µm x 694 µm. Images were taken down to a depth of approximately 600–700 µm, with a z-step size of 10–20 µm. Subsequently, stacks were stitched together resulting in an overall field of view of up to 4.3 × 4.3 mm × 0.7 mm using the Fiji49-based stitching plugin50.
Three-photon microscopy
The core hardware of the multiphoton 3 P microscope with adaptive optics has been described in detail in previous work22. In the following, a brief summary of the instrumentation is provided emphasizing any differences from the details given in Streich et al.22.
Over the course of the study, two excitation sources were used in this work. One of them is a Spectra-Physics TOPAS tunable non-collinear optical parametric amplifier generated ~60 fs pulses centered at 1300 nm with a repetition rate of 400 kHz, pumped by a 16 W Spectra-Physics Spirit. The near-IR pulses from the TOPAS were pre-compensated for dispersion by a homebuilt single prism (N-SF11) pulse compressor51. The maximum power at this wavelength and repetition rate was ~400 mW, resulting in a maximum available power under the objective of 35 mW. A Pockels cell was used for rapid modulation of the laser power during image acquisition. Furthermore, we also employed a Class 5 Photonics White Dwarf WD-1300-dual laser. The 1300 nm channel used here provided a maximum of over 5 Watts at a repetition rate of 1 MHz. In addition to the Pockels cell, a reflective optical density filter with a static OD = 0.8 attenuation was used to adapt the power range of the White Dwarf, yielding over 100 mW after the objective. Dispersion pre-compensation was done by an internal module in the White Dwarf which yielded 100 fs pulses after the objective. For fluorescence detection in the green channel, we switched to an uncooled H10770PA-40. The adaptive optics module used an ALPAO DM97-15 continuous membrane deformable mirror, relying on the factory calibration of the mirror for the Zernike-to-Control Matrix.
Images acquired with “full AO” optimization are optimized using the same metric and iterative procedure discussed in Streich et al.22. where the fluorescence intensity is measured and optimized directly on mGFP labeled cell in the mouse brain when not specified otherwise. The metric was found to also produce good enhancement when optimized on THG signal, enabling optimization in regions where no labeled cells are available. The typical optimization procedure involved two iterations of Zernike modes 3 through 21, with five amplitudes explored per iteration. With frame rates typically > 1 Hz, the entire process is completed in less than 3 min even in noisy regions. In both AO optimization as well as imaging, a maximum pulse energy at the focus of 2nJ was never exceeded52. Even after a four-hour time-lapse imaging session, only mild bleaching was observed (volume acquisition times range from 6 to 12 min which corresponds to a 30% to 60% duty cycle for light exposure since a volume is acquired every 20 min, though the time spent on each individual plane is at most 5% of that). The correction collar of the objective was set to 1 mm imaging depth for most acquisitions. We intentionally chose to optimize image quality (and hence utilized frame and/or pulse averaging).
In vivo deep brain imaging
For the initial induction of the anesthesia, nude NMRI mice were placed into a closed box and exposed to 6% isoflurane diluted in oxygen. For the maintenance of the anesthesia, a concentration of 1.6-2% isoflurane in oxygen was used, depending on the mouse’s breathing rate. The targeted breathing rate was between 70 and 90 bpm. Once anesthetized, eye cream was applied and mice were positioned on a small animal physiological monitoring system (ST2 75-1500, Harvard Apparatus), which allowed the maintenance of a stable body temperature at 37.5 °C and monitoring of the breathing rate.
For in vivo imaging experiments, animals were head-fixed using a customized headbar and complement holder and the cranial window was aligned so that the objective was placed above the part of the window that is centered slightly next to the sinus (where corpus callosum is anatomically the highest). Acquisition parameters for deep brain imaging are summarized in Supplementary Data 3. Here, we note that imaging depth was reported as raw axial translation of the objective. Taking the refractive index difference between the coverslip, immersion media and various brain tissues into account18, the actual imaging depth is likely around 5–10% larger than the values reported. For validation of the THG vessel signal, FITC dextran (fluorescein isothiocyanate-dextran, 2 M g/mol) was diluted in 0.9% NaCl- solution at 10 mg/ml and 100 µl injected intravenously into the tail vein.
Resolution estimation of 3PM
For estimation of image resolution used in comparing images with and without adaptive optics correction, the decorrelation analysis Fiji plugin was used53. The calculation was performed for all slices of the green channel in 5 different volume stacks. Slices that were clear outliers (order of magnitude off) or where the calculation failed (decorrelation curves had no maxima) were excluded and a mean and standard deviation for the stack resolution was calculated. The corresponding resolution ratio per volume was calculated with a standard deviation using error propagation. A weighted average of five such volumes was calculated with the corresponding standard deviation.
3PM deep learning-based denoising
Network architecture
In our approach, we designed a 3D version of the classic U-Net architecture25,54,55,56,57 (see Supplementary Fig. 2a). The architecture is composed of an encoder-decoder structure connected through skip connections. The encoder network contains 5 hierarchically organized encoder blocks. Each block consists of two consecutive units, including a 3D convolutional layer, a leaky ReLU activation layer, and a group normalization layer. In parallel, the decoder network is made up of 5 blocks with similar layer configurations and a nearest-neighbor upsampling layer at each level. The number of feature maps in the first level of the encoder network is set to 16, and then increases geometrically at each level. The feature maps from the encoder blocks are passed to the corresponding decoder blocks through the skip connections, allowing the combination of high-level features from the encoder with the semantic features from the decoder. This enables the U-Net architecture to effectively incorporate both local and global information in the reconstruction, making it well-suited for image restoration tasks.
Self-supervised denoising
In this study, we employed the Noise2Void strategy for self-supervised denoising of 3PM data, which allows for the training on the same data to be denoised24. This approach relies on the assumption that the noise is independently generated for each pixel, which is valid for the dominant noise sources in 3PM such as Poisson shot noise and Gaussian readout noise. During the training, a random subset of pixels, referred to as “blind spots”, are masked in the 3D input data and the network is optimized to predict the values of these pixels. This masking forces the prediction to be based solely on the surrounding patch. Given the independence of the noise, the network can only learn to determine the locally dependent true signal part of the pixel, effectively learning to denoise the volume data.
For our 3PM-Noise2Void method, we used 2% of the pixels as blind-spots, which were masked with randomly sampled pixel values from the adjacent neighborhood. A combination of L1 and L2 loss metrics was used to measure the difference between the blind-spot values in the prediction and raw volumes. We employed an ADAM optimizer58, with β1 = 0.5, β2 = 0.99, and a learning rate of 1e−3, to minimize this loss value over 300 epochs. The input volumes were randomly cut out into 16x64x64 patches, which were further augmented by rotation and flipping before applying the masking.
3PM data post-processing
The 3PM data also contains periodic structured noise caused by the ripple noise of the photomultiplier tubes (PMTs), which is modulated by the line scanning process into a line-wise periodic signal. This noise violates the assumption of pixel independence and causes the 3PM-Noise2Void approach to restore or even amplify it, leading to errors in downstream analysis (see Fig. 3c). To address this problem, we optimized the hyperparameters of the 3PM-Noise2Void, identifying the z-size parameter as a key factor. We found that a depth of 16 slices minimizes the restored structured noise. However, in cases where this was not sufficient, we developed a post-processing method, referred to as PerStruc-Denoiser.
This method accounts for the origin of the noise by aligning and subsequently extracting the structured noise that occurs misaligned row-by-row and with different patterns due to the acquisition procedure and inconsistent scanning speed. The method begins by flipping each odd row of a z-slice to ensure each line is in the same scanning direction and thus has the same pattern. Next, the function determines the phase differences of the structured noise for all lines of the data to a reference line by calculating the phase shift at the main frequency component of the structured noise in the Fourier domain. This reference line is chosen as the row in the volume with the lowest standard deviation, which presents the clearest structured noise pattern and serves as an accurate reference point for the phase detection. The PerStruc-Denoiser shifts each line under periodic boundary conditions to align the periodic structured noise. By performing a median projection along the row and then along all z-slices, the algorithm extracts the pure structured noise of a single line. This line is subtracted from the modified volume, and the alignment shifts and flips are reversed. Furthermore, a Gaussian filter with a small mask size (σ = 1 pixel) is applied line by line to suppress remaining high-frequency structured noise. Finally, the median value of the data is adjusted back to the original one, effectively reducing the periodic structured noise in the 3PM-Noise2Void denoised data and improving the downstream analysis of biological structures.
Machine learning-based classification of THG signal
The THG-signal channel of 3PM images was denoised as described above. In the custom approach, as opposed to the basic approach, subsequently, both the raw images as well as the denoised images were interpolated in their z-dimension with a factor of 4. The image z stacks were processed with a bilinear interpolation in this step. Semantic segmentation with ilastik was performed on the upsampled stack of raw images, using the denoised images as a prediction mask to limit the amount of false positives caused by the noise. In both approaches, Ilastik “Autocontext” workflow with 2 training stages was chosen for its superior performance on noisy data59. The THG signal was segmented into three classes of either vessel (1), myelin (2), or background (3). All color/intensity features, edge features and texture features were used up to a standard deviation of σ = 6. Additionally, 3D-features were calculated. Pixels were classified using a Random Forest classifier with manually drawn labels31.
To evaluate the effect of our pre-processing, an image stack was partially segmented. Labels were exported and subsequently integrated into the customized ML workflow as described, with the exception only labels from the ML training without pre-processing were used and no additional labeling was performed. With uncertainty, we refer to the classifier uncertainty, which in ilastik is computed as follows: let (p1, … pN) be probabilities predicted for classes 1…N for a pixel in the image. Further, let pMax1 be the highest probability of (p1, …, pN) and pMax2 the second highest. The pixel uncertainty is then computed as 1-(pMax1-pMax2). For example, if we have 2 close class predictions, pMax1 = 0.49 and pMax2 = 0.51, uncertainty = 0.98. If we have 2 far class predictions, pMax1 = 0.98 and pMax2 = 0.02, uncertainty = 0.02.
To visualize the morphological heterogeneity of myelin, vessel, and background, a Uniform Manifold Approximation and Projection (UMAP) was created based on 100 features that were most distinct between groups. Those features were identified using the FindAllMarkers function in Seurat48.
For validation of the THG vessel signal with intravenous FITC injection, FITC images were thresholded and converted to a masked image in Fiji to obtain a segmentation for FITC signal and background. All pixels of a selected region were then analyzed and their intensity for both groups was compared.
Sample preparation, microscopy and analysis for scanning electron microscopy
Tumor-bearing PDX mice were transcardially perfused and tissue was obtained as previously described3,6. For the postfixation, the mouse brain was put into 4% (w/v) PFA overnight and afterwards stored at 4 °C in PBS. The brain was subsequently cut into 200 µm thick sections. To unequivocally identify the glioma cells in scanning electron microscopy, immunolabeling with an antibody against human-specific nestin (abcam, ab22035, 1:500) was conducted3. For this purpose, the brain slices were put into 10% (w/v) sucrose dissolved in PBS for 10 min and for further 10 min in 20% (w/v) sucrose dissolved in PBS. The slice was subsequently incubated in 30% sucrose (w/v) over 12-15 h at 4 °C. Freeze-thaw-cycles were performed in liquid nitrogen twice for 5 min. Afterwards, the slices were incubated in 5% FBS in PBS at RT for 1 h. This was followed by incubation overnight at 4 °C with a human-specific mouse anti-nestin antibody (abcam, ab22035, 1:500) in the blocking solution. It was washed three times with blocking solution afterwards. The slices then were incubated for 12–15 h at 4 °C with a secondary antibody (a biotinylated anti-mouse antibody, abcam, ab6788, 1:500)). Slices were washed in PBS for three times and then incubated in the Vectastain ABC kit for 1 h at RT. Afterwards, the samples were exposed to a solution of glucose and DAB (with glucose at a concentration of 2 mg/ml, and DAB at a concentration of 1.4 mg/ml dissolved in PBS) for 10 min. This was followed by an hour-long incubation in a glucose-DAB-glucose oxidase solution (with glucose oxidase at a concentration of 0.1 mg/ml, from Serva) to generate an electron-dense precipitate. The efficacy of the process was assessed using widefield light microscopy.
The labeled sections were embedded in resin and mounted onto silicon wafers6. For image acquisition, we used a LEO Gemini 1530 scanning electron microscope (Zeiss) combined with an ATLAS scan generator. Potential contacts of brain tumor cells and blood vessels were observed by the identification of blood vessels according to basic ultrastructural features and the DAB (diamniobenzidine)-precipitate in the brain tumor cells respectively. For three-dimensional reconstruction of blood vessel-TM contacts, images were taken at the same position in consecutive layers. A working distance of 2–4 mm was set at an aperture of 20 µm and an acceleration voltage of 2 kV. The pixel sizes were between 3.8 nm and 15 nm in 400–3600 µm2 big images, enabling to create an image stack with a z-resolution of 280 nm. The manual segmentation of the structures was performed in Fiji.
Immunhistochemistry
For immunohistochemistry, xenografted mice were transcardially perfused with 4% (w/v) PFA in PBS (Sigma or Gibco) under deep anesthesia with isoflurane. Brains were collected and stored in 4% (w/v) PFA overnight and subsequently stored in PBS. 80-300 µm thick tissue sections were made using a Leica VT000S vibratome.
Sections were permeabilized with 5% (v/v) fetal bovine serum (FBS) and 1% (v/v) Triton X-100 for 2 h at room temperature, shaking. Primary antibodies were applied at a 1:100 dilution in a 1% FBS and 0,2% Triton X-100 solution and incubated for 24 h at 4 degrees Celsius, shaking. Sections were washed for 3x 15 min at room temperature, shaking. Appropriate secondary antibodies (anti rat antibody, Invitrogen, Cat#A21247; anti mouse antibody, Invtrogen, Cat#A11017 if not stated differently) were incubated at a 1:500 dilution in 1% FBS and 0,2% Triton X-100 for 24 h at 4 degrees Celsius, shaking. They were subsequently washed 3 × 10 min with 1% FBS in PBS and then 3 × 10 min with PBS at room temperature, shaking. Sections were mounted using “SlowFade Gold” solution. Images were acquired with a Leica LSM710 ConfoCor3, Zeiss using a 63x immersion oil objective (NA 1.4).
Evans blue in vivo staining
Evans blue (E2129-10G, Sigma) was diluted in 0.9% sodium chloride to a concentration of 20 mg/ml. Subsequently, 150 µl of the solution was administered via intraperitoneal injection in mice, with a 12-hour interval before perfusion and brain extraction.
Vessel quantification
Maximum intensity projections of field of views measuring 237 × 237 x 25 microns were analyzed. Six regions out of three tumor injected mice on two time points were chosen. 3D registration was performed using a custom written Fiji macro. Vessels were identified manually.
Blood brain barrier analysis
Blood vessels were segmented using ilastik. Then, to measure potential extravasation, the intensity of the extravascular background normalized to the mean region intensity of the images was compared.
Intravital, microscopic blood vessel analysis
For the analyzes of vessel widths, vessel diameter was measured in Fiji. As diameters in the FITC signal were systematically larger than in THG signal, THG vessel diameter was multiplied with 1.248373 (mean factor difference between vessels measured between FITC/THG). Vessel density was assessed using the Fiji plugin “Vessel analysis”60.
Polarity analysis of single glioma cells, myelin fibers and vessels
Tumor cell polarity was investigated analyzing the primary orientation of each tumor cell in Fiji47,49 and calculating the angle between the tumor cell orientation and a horizontal line. In very dense tumor regions, a subset of slices was analyzed to unequivocally identify single tumor cells. Subsequently, for each region in the corpus callosum, the overall direction of fibers was analyzed in the same manner and the calculated angles were subtracted. The angles were recalculated so that all angles were in the range of -90 to 90 degrees. For the analysis of angles between myelin and vessels, denoised THG signal within the corpus callosum was analyzed. Changes of direction in each vessel were considered individually. The angle between each section of a vessel and the nearest myelin fibers was measured using Fiji.
Angle analysis between glioma cells and myelin fibers in the cortex
For the correlation between glioma cells and myelin fibers within the cortex, confocal images of PDX brain slices stained for MBP (Novus Cat# NB600-717,1:100) were analyzed. First, the glioma cells and myelin fibers were segmented in ilastik to get binary masks. Second, boundaries were dilated by 5 µm to define the area close-by to the glioma. Third, to obtain the myelin fibers that are running through this area adjacent to the tumor cell, this area mask was multiplied with the myelin fiber mask, resulting in a mask of myelin-fibers that are adjacent to the tumor cell. Fourth, this mask was then skeletonized in Fiji. Lastly, to obtain single fiber segments that are not touching each other, branching points of the skeleton were removed. Additionally, skeleton segments consisting of only a few pixels (≤ 4) were removed to avoid the influence of artifacts. The major direction was then assessed using the start and endpoint of each segment, and compared to the major axis orientation of the glioma cell.
THG validation
For THG validation, we labeled myelin in brain slices of wild type mice by performing immunohistochemistry using an MBP antibody (invitrogen, Cat#MA1-10837, 1:100).
Orientation analysis of normal brain cells
Histological slides of the mouse brain from the Allen Brain Atlas where analyzed. Coherency of the slide was calculated using OrientationJ61. Segmentation was performed using Ilastik. The coherency analyzes whether image features are oriented or not oriented on a local scale61, with values closer to 1 indicating higher orientation, and values closer to 0 indicating less structuring. Mean coherency was calculated in the regions of interest.
Somatokinesis measurement
Somatokinesis measurements were performed in ImageJ/Fiji49. First, time-lapse images of tumor cell regions were registered using custom written registration scripts. Then, tumor cells were cropped and re-registered with the 3D Drift Correction plugin in ImageJ/Fiji62 when necessary. The glioblastoma cell somata were labeled using a selection tool in Fiji and the center of each cell was measured. The distance between the center points of the first and last time point was calculated and divided by the duration of the experiment to obtain the somatokinetic speed of the glioblastoma cell.
Invasion phenotype classification
Invasion phenotype classification was performed as previously described3. The invasion phenotypes are characterized as shown in Table 1.
Invasion phenotypes for each cell were classified based on their invasive behavior during the time course of up to 4 h.
Tumor microtube and small process identification
A tumor microtube was defined as a small cellular protrusion with a minimum length of 10 µm and a thickness ranging between 0.5–2.5 µm15. Small processes were defined as cellular protrusions smaller than 10 µm in length3.
Directionality analysis of brain tumor networks
Tumor cell regions in the corpus callosum and in the cortex were imaged with 3PM and 2PM, respectively. For each slice, the local orientation was calculated using a local window of σ = 3 pixels and a cubic spline as gradient, as implemented in OrientationJ61. The calculated output was exported as a stack, displaying the local orientation in degrees as well as table. For visualization of local network orientation values using rose plots, calculated orientation values were filtered \(x > (\frac{\sum {Energy}}{n}+\sqrt{\frac{\sum {\left({Energy}-\frac{\sum {Energy}}{n}\right)}^{2}\,}{n}})\) to select only tumor cell associated orientation values. For further visualization, images of tumor cell regions were binarized to generate a mask for the tumor cell network. The absolute values of the stack displaying the local orientation in degrees were multiplied with the tumor cell mask to receive the local orientation of the tumor cell network. The difference to the horizontal line was calculated for each local orientation and visually displayed as a spectrum from 90 degrees (lowest intensity) to 0 degrees (brightest intensity) in the maximum intensity mode of Arivis. This was performed for both cortex as well as corpus callosum brain regions.
White matter reactivity analyzes
Multiple regions with tumor infiltration were analyzed in the white matter-rich areas of the corpus callosum. Regions without THG signal in the corpus callosum were analyzed in Fiji using the polygon tool and measuring their shape descriptors on selected slices. Holes were labeled as either tumor cell-infiltrated or without tumor infiltration. Subsequently, the circularity of both groups was compared.
Orientation analysis of myelinated fibers was analyzed using OrientationJ61. Vector fields were visualized as arrows in R.
To analyze displacement of myelin fibers by TMs or somata, TMs and somata were selected. Then, THG signal was manually labeled as displaced or non-displaced, based on THG signal. The distribution of displacing and non-displacing TMs and somata per group was analyzed.
Human MRI and diffusion imaging analysis
IDH-wildtype glioblastoma patients that received clinical indicated MRI scans at the Department of Neuroradiology (University Hospital Heidelberg) were retrospectively reviewed. Written informed consent was obtained from all patients and all examinations were in accordance with the declaration of Helsinki and approved by the local ethics committee of Heidelberg University (study permit: S-078/2021). Glioma patients were included into the analysis that showed unilateral affection of the corpus callosum (genu or splenium) at baseline based on T1 post Gd contrast MRI (n = 8 patients). Additionally, follow-up investigations 10.3 ± 5.8 months after baseline were assessed. Imaging was performed at 3 T on a Magnetom Skyra or Prisma (Siemens Healthineers, Erlangen Germany). Sequences included T1w after Gd-contrast administration and diffusion imaging.
Sequence parameters were as follows: T1 mpRAGE 3D Sequence: TR: 1.7 ms, TE: 3.3 ms; FOV: 320 × 263 matrix size: 320 × 263 voxel size: 1 mm. Diffusion imaging (Resolve 2D sequence): TR: 3.7 ms, TE: 69 ms, matrix size: 192 × 192 FOV: 220 × 220; slice thickness 5 mm. Regions of interest were selected manually on the ipsi- and contralateral splenium or genu of the corpus callosum and the apparent diffusion coefficient (ADC) was quantified on a PACS RA1000 workstation (GE Healthcare, Chicago, US) by a board-certified neuroradiologist (M.O.B., >10 years of experience in neuroradiology).
High-field animal MRI
MR scans were conducted using a 9.4 Tesla horizontal bore small animal MRI scanner (BioSpec 94/20 USR, Bruker BioSpin GmbH, Ettlingen, Germany) equipped with a gradient strength of 675 mT/m and a 2 × 2 surface array receive-only coil. During the imaging procedure, anesthesia was induced using 4% isoflurane (Baxter, Unterschleißheim, Germany) in 100% O2, and subsequently maintained with 1–1.5% isoflurane in 100% O2 delivered via a nose cone throughout the scanning process. The respiration rate was continuously monitored, and the animals were positioned in a prone position on a Bruker standard MRI bed with an integrated circulating water heating system to ensure body temperature maintenance. A diffusion tensor imaging sequence in axial planes was employed, using the following parameters: echo time = 17.57 ms, repetition time = 1000 ms, acquisition matrix = 75 × 100, field-of-view = 15 × 20 mm2, slice thickness = 0.5 mm.
High-field diffusion tensor imaging analysis
Fractional Anisotropy (FA), mean diffusivity (MD) and axial diffusivity (AD) maps were extracted from the respective DTI image stacks using open-source software MITK-Diffusion (https://github.com/MIC-DKFZ/MITK-Diffusion/)63. Tracts of the body of the corpus callosum (CC) were extracted using the atTRACTive tool in MITK Diffusion64. Tracts were transformed into ROIs by binarizing the tract density image using an empirically chosen threshold of 8. Data emerged from 42 MR acquisitions (26 days (n = 5), 33 days (n = 7), 60 days (n = 12) and 73 days (n = 13) after injection). 5 acquisitions were sorted out because of too small ROIs (ROI < 100 voxels).
3D renderings and visualization
Renderings were created in Arivis Vision 4D for 3D and 4D Image visualization.
Statistical analysis
The results of quantifications were transferred to GraphPad Prism (GraphPad Software) or R to test the statistical significance with the appropriate tests as indicated in the figure legends, normality was tested using the Shapiro-Wilk test. Results were considered statistically significant if the P value was below 0.05. Quantifications were done blinded. Animal group sizes were as low as possible and empirically chosen and longitudinal measurements allowed a reduction of animal numbers by maintaining an adequate power. No statistical methods were used to predetermine the sample size. In quantifications that were depicted as boxplots, the upper and the lower hinges correspond to the third and the first quartiles.
Reporting summary
Further information on research design is available in the Nature Portfolio Reporting Summary linked to this article.
Data availability
The source data of the analyzes are provided in the Source Data file. Spatial transcriptomics data used from reference 47 are available on [https://www.molecularatlas.org/]. Clinical and Pathological characteristics of Tumor cases is available in Supplementary Data 1. Settings and details for reproducing the experiments are provided in the Supplementary Note, Supplementary Methods and in Supplementary Data 3. Source data are provided with this paper.
Code availability
The indirect AO routines developed for integration into ScanImage32 are available at [https://github.com/prevedel-lab/AO.git] as previously described22. AI denoising was performed using a customized approach as described above. Code is available on GitHub at [https://github.com/prevedel-lab/Deep3P-Denoising] and on Zenodo65 [https://doi.org/10.5281/zenodo.11175040].
References
Drumm, M. R. et al. Extensive brainstem infiltration, not mass effect, is a common feature of end-stage cerebral glioblastomas. Neuro Oncol. 22, 470–479 (2020).
Sahm, F. et al. Addressing diffuse glioma as a systemic brain disease with single-cell analysis. Arch. Neurol. 69, 523–526, (2012).
Venkataramani, V. et al. Glioblastoma hijacks neuronal mechanisms for brain invasion. Cell 185, 2899–2917 e2831 (2022).
Garofano, L. et al. Pathway-based classification of glioblastoma uncovers a mitochondrial subtype with therapeutic vulnerabilities. Nat. Cancer 2, 141–156 (2021).
Neftel, C. et al. An integrative model of cellular states, plasticity, and genetics for glioblastoma. Cell 178, 835–849 e821 (2019).
Venkataramani, V. et al. Glutamatergic synaptic input to glioma cells drives brain tumour progression. Nature 573, 532–538 (2019).
Wick, W., Osswald, M., Wick, A. & Winkler, F. Treatment of glioblastoma in adults. Ther. Adv. Neurol. Diso. 11, https://doi.org/10.1177/1756286418790452 (2018).
Scherer, H. J. A critical review: the pathology of cerebral gliomas. J. Neurol. psychiatry 3, 147–177 (1940).
Hide, T. et al. Oligodendrocyte progenitor cells and macrophages/microglia produce glioma stem cell niches at the tumor border. Ebiomedicine 30, 94–104 (2018).
Brooks, L. J. et al. The white matter is a pro-differentiative niche for glioblastoma. Nat. Commun. 12, 2184 (2021).
Liu, C. J., Shamsan, G. A., Akkin, T. & Odde, D. J. Glioma cell migration dynamics in brain tissue assessed by multimodal optical imaging. Biophys. J. 117, 1179–1188 (2019).
Wang, J. et al. Invasion of white matter tracts by glioma stem cells is regulated by a NOTCH1-SOX2 positive-feedback loop. Nat. Neurosci. 22, 91 (2019).
Theer, P. & Denk, W. On the fundamental imaging-depth limit in two-photon microscopy. J. Opt. Soc. Am. A 23, 3139–3149 (2006).
Alieva, M. et al. Intravital imaging of glioma border morphology reveals distinctive cellular dynamics and contribution to tumor cell invasion. Sci. Rep. 9, https://doi.org/10.1038/s41598-019-38625-4 (2019).
Osswald, M. et al. Brain tumour cells interconnect to a functional and resistant network. Nature 528, 93–98 (2015).
Watkins, S. et al. Disruption of astrocyte-vascular coupling and the blood-brain barrier by invading glioma cells. Nat. Commun. 5, 4196 (2014).
Montgomery, M. K. et al. Glioma-induced alterations in neuronal activity and neurovascular coupling during disease progression. Cell Rep. 31, 107500 (2020).
Horton, N. G. et al. In vivo three-photon microscopy of subcortical structures within an intact mouse brain. Nat. Photonics 7, 205–209 (2013).
Choe, K. et al. Intravital three-photon microscopy allows visualization over the entire depth of mouse lymph nodes. Nat. Immunol. 23, 330–340 (2022).
Wang, T. Y. & Xu, C. Three-photon neuronal imaging in deep mouse brain. Optica 7, 947–960, (2020).
Ouzounov, D. G. et al. In vivo three-photon imaging of activity of GCaMP6-labeled neurons deep in intact mouse brain. Nat. methods 14, 388 (2017).
Streich, L. et al. High-resolution structural and functional deep brain imaging using adaptive optics three-photon microscopy. Nat. methods 18, 1253–1258 (2021).
Andresen, V. et al. Infrared multiphoton microscopy: subcellular-resolved deep tissue imaging. Curr. Opin. Biotechnol. 20, 54–62 (2009).
Krull, A., Buchholz, T.-O. & Jug, F. in Proceedings of the IEEE/CVF conference on computer vision and pattern recognition. 2129–2137.
Weigert, M. et al. Content-aware image restoration: pushing the limits of fluorescence microscopy. Nat. methods 15, 1090–1097 (2018).
Tsai, H. H. et al. Oligodendrocyte precursors migrate along vasculature in the developing nervous system. Science 351, 379–384 (2016).
Tabata, H. et al. Erratic and blood vessel-guided migration of astrocyte progenitors in the cerebral cortex. Nat. commun. 13, https://doi.org/10.1038/s41467-022-34184-x (2022).
Cuddapah, V. A., Robel, S., Watkins, S. & Sontheimer, H. A neurocentric perspective on glioma invasion. Nat. Rev. Neurosci. 15, 455–465 (2014).
Barad, Y., Eisenberg, H., Horowitz, M. & Silberberg, Y. Nonlinear scanning laser microscopy by third harmonic generation. Appl. Phys. Lett. 70, 922–924 (1997).
Redlich, M. J. & Lim, H. A method to measure myeloarchitecture of the murine cerebral cortex in vivo and ex vivo by intrinsic third-harmonic generation. Front. Neuroanat. 13, 65 (2019).
Berg, S. et al. ilastik: interactive machine learning for (bio)image analysis. Nat. methods 16, 1226–1232 (2019).
Li, S. Z. & Jain, A. (eds) in Capillary Blood Vessel. Encyclopedia of Biometrics (Springer, 2009).
Marin, O., Valiente, M., Ge, X. & Tsai, L. H. Guiding neuronal cell migrations. Cold Spring Harb. Perspect. Biol. 2, a001834 (2010).
Jung, E. et al. Tweety-homolog 1 drives brain colonization of gliomas. J. Neurosci. 37, 6837–6850 (2017).
Jung, E. et al. Tumor cell plasticity, heterogeneity, and resistance in crucial microenvironmental niches in glioma. Nat. Commun. 12, 1014 (2021).
Hausmann, D. et al. Autonomous rhythmic activity in glioma networks drives brain tumour growth. Nature 613, 179–186 (2023).
Kallenberg, K. et al. Glioma infiltration of the corpus callosum: early signs detected by DTI. J. Neuro-Oncol. 112, 217–222 (2013).
Smits, M. & van den Bent, M. J. Imaging correlates of adult glioma genotypes. Radiology 284, 316–331 (2017).
Booth, M. J. Adaptive optical microscopy: the ongoing quest for a perfect image. Light-Sci. Appl. 3, https://doi.org/10.1038/lsa.2014.46 (2014).
Rodriguez, C. et al. An adaptive optics module for deep tissue multiphoton imaging in vivo. Nat. methods 18, 1259 (2021).
Hontani, Y., Xia, F. & Xu, C. Multicolor three-photon fluorescence imaging with single-wavelength excitation deep in mouse brain. Sci. Adv. 7, eabf3531 (2021).
Weil, S. et al. Tumor microtubes convey resistance to surgical lesions and chemotherapy in gliomas. Neuro Oncol. 19, 1316–1326 (2017).
Capper, D. et al. DNA methylation-based classification of central nervous system tumours. Nature 555, 469–474 (2018).
Dondzillo, A. et al. Targeted three-dimensional immunohistochemistry reveals localization of presynaptic proteins Bassoon and Piccolo in the rat calyx of Held before and after the onset of hearing. J. Comp. Neurol. 518, 1008–1029 (2010).
Oudin, A. et al. Protocol for derivation of organoids and patient-derived orthotopic xenografts from glioma patient tumors. STAR Protoc. 2, 100534 (2021).
Golebiewska, A. et al. Patient-derived organoids and orthotopic xenografts of primary and recurrent gliomas represent relevant patient avatars for precision oncology. Acta Neuropathol. 140, 919–949 (2020).
Ortiz, C. et al. Molecular atlas of the adult mouse brain. Sci. Adv. 6, https://doi.org/10.1126/sciadv.abb3446 (2020).
Hao, Y. et al. Integrated analysis of multimodal single-cell data. Cell 184, 3573–3587.e3529 (2021).
Schindelin, J. et al. Fiji: an open-source platform for biological-image analysis. Nat. methods 9, 676–682 (2012).
Preibisch, S., Saalfeld, S. & Tomancak, P. Globally optimal stitching of tiled 3D microscopic image acquisitions. Bioinformatics 25, 1463–1465 (2009).
Kong, L. & Cui, M. A high throughput (> 90%), large compensation range, single-prism femtosecond pulse compressor. Preprint at arXiv preprint arXiv:1306.5011 (2013).
Yildirim, M., Sugihara, H., So, P. T. C. & Sur, M. Functional imaging of visual cortical layers and subplate in awake mice with optimized three-photon microscopy. Nat. Commun. 10, https://doi.org/10.1038/s41467-018-08179-6 (2019).
Descloux, A., Grussmayer, K. S. & Radenovic, A. Parameter-free image resolution estimation based on decorrelation analysis. Nat. methods 16, 918 (2019).
Ronneberger, O., Fischer, P. & Brox, T. 234-241 (Springer International Publishing).
Li, X. et al. Reinforcing neuron extraction and spike inference in calcium imaging using deep self-supervised denoising. Nat. methods 18, 1395–1400 (2021).
Chaudhary, S., Moon, S. & Lu, H. Fast, efficient, and accurate neuro-imaging denoising via supervised deep learning. Nat. Commun. 13, 5165 (2022).
Xu, Y. K. T. et al. Cross-modality supervised image restoration enables nanoscale tracking of synaptic plasticity in living mice. Nat. methods 20, 935–944 (2023).
Kingma, D. P. & Ba, J. Adam: A method for stochastic optimization. Preprint at arXiv preprint arXiv:1412.6980 (2014).
Kreshuk, A. & Zhang, C. Machine learning: advanced image segmentation using ilastik. Methods Mol. Biol. 2040, 449–463 (2019).
Elfarnawany, M. H. E.-K. Signal processing methods for quantitative power doppler microvascular angiography. (The University of Western Ontario (Canada), 2015).
Puspoki, Z., Storath, M., Sage, D. & Unser, M. Transforms and operators for directional bioimage analysis: a survey. Adv. Anat. Embryol. Cell Biol. 219, 69–93 (2016).
Parslow, A., Cardona, A. & Bryson-Richardson, R. J. Sample drift correction following 4D confocal time-lapse imaging. J. Vis. Exp. https://doi.org/10.3791/51086 (2014).
Fritzsche, K. H. et al. MITK diffusion imaging. Methods Inf. Med. 51, 441–448 (2012).
Peretzke, R. et al. atTRACTive: Semi-automatic white matter tract segmentation using active learning. arXiv:2305.18905 https://ui.adsabs.harvard.edu/abs/2023arXiv230518905P (2023).
Maus, E., Prevedel, Robert. Deep3P Denoising: Code of the 3PM-Noise2Void and PerStruc-Denoiser Method. https://doi.org/10.5281/zenodo.11175040 (2024).
Acknowledgements
V.V. received financial support from the German Research Foundation (DFG: VE1373/2-1), the Else Kröner-Fresenius-Stiftung (2020-EKEA.135), the University of Heidelberg (Physician-Scientist-Program, Krebs- und Scharlachstiftung) and the Health + Life Science Alliance Heidelberg Mannheim. M.S., J.S., E.R., and S.T. were supported by the Deutsche Krebshilfe/German Cancer Aid (Mildred-Scheel-Scholarship for MD students). W.W., F.W., M.B., and V.V. were supported by a grant from the German Research Foundation (DFG: SFB 1389). R.P. acknowledges support from the European Commission (grant no. 951991, Brainiaqs), and the Chan Zuckerberg Initiative (Deep Tissue Imaging grant no. 2020-225346). R.P., A.T., and E.M. were supported by the EMBL. The Northwestern Nervous System Tumor Bank is supported by the P50CA221747 SPORE for Translational Approaches to Brain Cancer. M.O.B. was supported by the Emmy Noether program of the German Research Foundation (DFG, BR 6153/1-1) and the Chica and Heinz Schaller Foundation. F.T.K. was supported by a grant from the German Research Foundation (# 507778602). We thank M. Kaiser, Y. Dörflinger, S. Hoppe, M. Schmitt, A. Oudin, M. Fischer, and V. Baus for their technical assistance. We thank F. Blum, T. Rubner, M. Eich, K. Hexel and S. Schmitt from the Flow Cytometry Core Facility of the German Cancer Research center for their assistance in performing FACS experiments. We thank K. Becker for providing support and assistance in animal care and design of animal experiments. We thank Lina Streich and the EMBL animal facilities for help and support. We thank the EM Core Facility of Heidelberg University and the EM and Microscopy Core Facility of the German Cancer Research center for their support. The authors thank the donors and their families who donated through the Nervous System Tumor Bank. Some graphic elements shown were created with Biorender.com.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Contributions
R.P. and V.V. contributed and supervised equally; Conceptualization, R.P., V.V.; Methodology, M.S., S.S., A.T., E.M., A.K., R.P., V.V.; Software Analysis, M.S., Software Denoising: E.M.; Project Administration, R.P., V.V.; Investigation, M.S., S.S., A.T., J.S., N.W., E.R., S.K.T., Y.Y., R.D., M.D., A.H., V.B., A.S, J.W., K.M.; Formal Analysis, M.S., S.S., A.T., J.S., N.W., E.R., S.K.T., Y.Y., R.D., C.B., M.D., R.P., A.H., V.B., F.T.K.; Resources, S.H., A.G., F.T.K., W.W., T.K., F.W., C.H., R.P., V.V.; Visualization, M.S., S.S., J.S., E.R., C.B.; Writing- Original Draft, R.P., V.V.; Writing- Review & Editing, M.S., S.S., A.T., E.M., E.R., S.K.T., N.W., R.D., M.D., M.B., P.N., A.G., C.B., W.W., F.W., A.K., M.B., T.K., C.H., R.P., V.V.; Funding Acquisition, W.W., F.W., T.K., R.P., V.V.
Corresponding authors
Ethics declarations
Competing interests
F.W. and W.W. report the patent (WO2017020982A1) “Agents for use in the treatment of glioma.” F.W. is co-founder of DC Europa Ltd (a company trading under the name Divide & Conquer) that is developing new medicines for the treatment of glioma. Divide & Conquer also provides research funding to F.W.’s lab under a research collaboration agreement. The remaining authors declare no competing interests.
Peer review
Peer review information
Nature Communications thanks the anonymous reviewer(s) for their contribution to the peer review of this work. A peer review file is available.
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Source data
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Schubert, M.C., Soyka, S.J., Tamimi, A. et al. Deep intravital brain tumor imaging enabled by tailored three-photon microscopy and analysis. Nat Commun 15, 7383 (2024). https://doi.org/10.1038/s41467-024-51432-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1038/s41467-024-51432-4
- Springer Nature Limited