
Abstract
Long non-coding RNAs (lncRNAs) have garnered significant attention in biomedical research due to their pivotal roles in gene expression regulation and their association with various human diseases. Among these lncRNAs, ArfGAP With RhoGAP Domain, Ankyrin Repeat, And PH Domain 1 - Antisense RNA 1 (ARAP1-AS1) has recently emerged as an novel oncogenic player. ARAP1-AS1 is prominently overexpressed in numerous solid tumors and wields influence by modulating gene expression and signaling pathways. This regulatory impact is realized through dual mechanisms, involving both competitive interactions with microRNAs and direct protein binding. ARAP1-AS1 assumes an important role in driving tumorigenesis and malignant tumor progression, affecting biological characteristics such as tumor expansion and metastasis. This paper provides a concise review of the regulatory role of ARAP1-AS1 in malignant tumors and discuss its potential clinical applications as a biomarker and therapeutic target. We also address existing knowledge gaps and suggest avenues for future research. ARAP1-AS1 serves as a prototypical example within the burgeoning field of lncRNA studies, offering insights into the broader landscape of non-coding RNA molecules. This investigation enhances our comprehension of the complex mechanisms that govern the progression of cancer.
Similar content being viewed by others


Introduction
Cancer is a multifaceted and diverse malignant disease, ranking among the foremost global causes of mortality [1,2,3,4]. Genome sequencing projects have shown that while approximately 98% of the human genome is non-coding, resulting in a substantial abundance of non-coding transcripts [5,6,7]. With the advancement of sequencing technology, a large number of long non-coding RNAs (lncRNAs) have been discovered and identified [8,9,10], and research has also demonstrated the significant involvement of lncRNAs in the onset and progression of human ailments [11,12,13,14,15,16], especially tumors [17,18,19,20,21]. As oncogenic or anticancer genes, lncRNAs regulate neoplastic properties such as proliferation, migration, and metastasis, thereby participating in tumor initiation and progression [22,23,24,25,26,27,28].
ARAP1-AS1, officially fully known as the ArfGAP With RhoGAP Domain, Ankyrin Repeat, And PH Domain 1 - Antisense RNA 1, exemplifies an RNA gene that falls under the category of lncRNAs. This gene is located on the 11nd chromosome of the human genome, specifically at q13.4. There is only one splice variant that derived from the ARAP1-AS1 gene with 388 bp (https://www.ensembl.org/Homo_sapiens/Gene/Summary?g=ENSG00000256007;r=11:72685075-72693808;t=ENST00000542022). Furthermore, when queried in the NONCODE database (http://www.noncode.org/) [29], it was observed that ARAP1-AS1 exhibits expression exclusively within specific normal human tissues, including adrenal glands, testicles, breasts, colon, and kidneys. However, the expression levels are notably low or undetectable in other tissues. According to data from the lncLocator database (http://www.csbio.sjtu.edu.cn/bioinf/lncLocator/) [30] and Lnc2Cancer 3.0 (http://www.bio-bigdata.net/lnc2cancer or http://bio-bigdata.hrbmu.edu.cn/lnc2cancer) database [31], ARAP1-AS1 is mainly located in the cytoplasm.
Recently, ARAP1-AS1 has been reported to exhibit upregulation in diverse tumor tissues, including lung cancer [32,33,34], lymphoma [35], thyroid cancer [36], bladder cancer [37,38,39], cervical cancer [40,41,42], ovarian cancer [43], kidney cancer [44], breast cancer [45], gastric cancer [46], and colorectal cancer [47]. ARAP1-AS1 is also implicated in the initiation and advancement of tumorigenesis. However, the precise molecular mode of action by which ARAP1-AS1 exerts its oncogenic potential remains elusive. This article reviews the current evidence on abnormal expression of ARAP1-AS1 and its clinical significance, and further summarizes the regulatory mechanism of ARAP1-AS1 and its role in human carcinogenesis.
Elevated expression of ARAP1-AS1 across tumor tissues and cell lines
The Cancer Genome Atlas (TCGA) is an important research initiative focused on comprehensively elucidating genomic alterations in various cancer types [48], including alterations associated with ncRNAs, such as lncRNAs. By collecting diverse cancer samples and employing high-throughput genomics technologies, including microarrays and second-generation sequencing, TCGA helps advance detailed studies in cancer research and lncRNA exploration [49, 50]. This approach enhances our understanding of cancer biology and provides insights for personalized cancer treatment [48].
In 2019, Teng et al. [39] conducted an analysis utilizing data from the TCGA database. Their study revealed a significant upregulation of the novel lncRNA, ARAP1-AS1, in bladder cancer samples. This finding was further substantiated through quantitative reverse transcription polymerase chain reaction (qRT-PCR) analysis performed on 88 pairs of bladder cancer tissues and their corresponding non-tumorous tissues. Notably, this study marked the initial documentation of ARAP1-AS1’s relevance in bladder cancer. Subsequently, in the same year, Xu et al. [38] conducted a comprehensive characterization of differentially expressed RNA profiles in bladder cancer using TCGA data. They also identified ARAP1-AS1 as one of the prominently regulated lncRNAs. These observations underscore its potential involvement in the oncogenic processes associated with bladder cancer.
Subsequently, multiple research groups have consistently reported a substantial upregulation of ARAP1-AS1 expression in solid tumor specimens originating from a diverse spectrum of cancer types. These encompass colorectal cancer [47], gastric cancer [46], breast cancer [45], cervical cancer [40, 41], lung cancer [32, 34], ovarian cancer [43, 51], renal cancer [44], thyroid cancer [36], and lymphoma [35]. Furthermore, heightened expression of ARAP1-AS1 has been consistently identified in various cancer cell lines pertaining to the aforementioned tumor types. For instance, Ye et al. [47] conducted a study involving 82 pairs of colorectal cancer (CRC) tissues and adjacent non-tumor tissues, revealing frequent overexpression of ARAP1-AS1 in CRC samples. They also observed higher ARAP1-AS1 expression in five CRC cell lines when compared to normal colorectal epithelial cells HCoEpiC. In cervical cancer, Zhang et al. [42] reported a significant upregulation of ARAP1-AS1 in cancer tissues and tumor cell lines. Remarkably, they extended their investigation to peripheral blood samples from both cervical cancer patients and healthy controls. Their findings revealed an eightfold increase in ARAP1-AS1 expression in cervical cancer serum in comparison to serum from healthy controls. In addition, we also comprehensively evaluate the expression of ARAP1-AS1 in pan-cancer using the GEPIA 2 online database tool (http://gepia2.cancer-pku.cn/#index) [52]. As shown in Fig. 1, the expression of ARAP1-AS is significantly up-regulated in most cancer tissues, while the expression of ARAP1-AS is relatively very low in majority of normal tissues. These collective observations underscore the consistent upregulation of ARAP1-AS1 expression in malignant tumors originating from diverse tissues and organs. Consequently, ARAP1-AS1 emerges as a novel oncogenic candidate, with the potential to serve as a tumor biomarker applicable to various cancer types.

Expression analysis of ARAP1-AS1 in matched TCGA cancer and normal tissues using GEPIA2 tool. The expression of ARAP1-AS is significantly up-regulated in most cancer tissues, while the expression of ARAP1-AS is relatively very low in majority of normal tissues. The abbreviations in this figure can be found in Supplementary Table 1 for reference
ARAP1-AS1 as a novel tumor-related clinical marker
ARAP1-AS1 emerges as a promising molecular marker with clinical value across various solid tumors. Numerous research groups have explored the relationship between ARAP1-AS1 expression, clinical parameters, and prognosis in cancer, as summarized in Table 1. For instance, in colorectal cancer (CRC) tissues, ARAP1-AS1 expression exhibited significant correlations with TNM stage and lymph node metastasis [47]. Additionally, a preliminary investigation by Jiang et al. [46] unveiled a positive association between ARAP1-AS1 expression and TNM stage and lymphatic metastasis, with higher ARAP1-AS1 levels predicting poorer overall survival (OS) and disease-free survival (DFS) in gastric cancer cases. Furthermore, elevated ARAP1-AS1 levels were indicative of advanced pathological characteristics, including higher FIGO stage and larger tumor size in cervical cancer. Notably, overexpression of ARAP1-AS1 was associated with diminished OS in ovarian cancer, cervical cancer, bladder cancer, and lymphoma. Intriguingly, serum ARAP1-AS1 demonstrated good diagnostic potential for cervical cancer patients, with an area under the curve (AUC) value of 0.8953. ARAP1-AS1 harbors significant promise for tumor diagnostic detection and prognostic assessment; however, further clinical investigations remain imperative to solidify its applications in the clinical setting.
Functional role and regulatory mechanisms of ARAP1-AS1 in tumor development
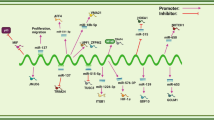
ARAP1-AS1 consistently demonstrates upregulation in a multitude of tumor tissues and cell lines, collectively emphasizing its potential oncogenic role across various tumor types. Studies have explored the biological role of ARAP1-AS1 in different tumors in vivo and/or in vitro experimental approaches, as documented in Table 2. ARAP1-AS1 is involved in a series of tumor-related biological processes (Fig. 2). Specifically, ARAP1-AS1 acts as a potent promoter of epithelial-mesenchymal transition (EMT), an enhancer of cellular proliferation and viability, a facilitator of cell migration and invasion, a catalyst for tumor growth, and a potent contributor to metastasis. Conversely, ARAP1-AS1 exerts inhibitory effects on apoptosis and hinders cell cycle arrest in tumor cells. These findings collectively highlight the multifaceted role of ARAP1-AS1 as a key oncogenic regulator with significant impact on tumor initiation and progression.

The functions of oncogenic ARAP1-AS1 in different cancers. This figure offers a detailed overview of the roles of ARAP1-AS1 in a variety of tumor types. It underscores the wide-ranging functions of ARAP1-AS1 in diverse cancers, including lung cancer, lymphoma, thyroid cancer, bladder cancer, among others. These functions encompass the regulation of crucial cellular processes such as proliferation, adhesion, invasion, tumor metastasis, and more
Mounting evidence suggests that lncRNAs can be triggered into action by the transcription factors situated upstream of them. Notably, the promoter region of ARAP1-AS1 contains binding sites for a number of transcription factors, such as c-Myc and YY1 (as depicted in Fig. 3). To delve deeper, the study by Ye et al. [47] revealed that YY1 has the capability to enhance ARAP1-AS1 expression by promoting its transcription. Furthermore, it was uncovered that the Wnt/β-catenin signaling pathway operates downstream of ARAP1-AS1. This YY1/ARAP1-AS1 axis was found to drive colorectal cancer cell migration by activating the Wnt/b-catenin signaling pathway. Another investigation by Zhang et al. [42] documented that c-Myc can activate ARAP1-AS1 at the transcriptional level by directly binding to the E-box motif located within the ARAP1-AS1 promoter. This discovery led to the identification of a positive feedback loop between ARAP1-AS1 and c-Myc, amplifying the carcinogenic impact of ARAP1-AS1.

Mechanism responsible for the upregulated expression of ARAP1-AS1 in tumors. Certain transcription factors, including YY1 and c-Myc, attach themselves to the ARAP1-AS1 promoter, thereby initiating the transcription of ARAP1-AS1
The regulatory mechanisms associated with ARAP1-AS1 in tumor development are complex and multifaceted. In addition to its involvement in ceRNA networks and protein binding (acting as a modular scaffold or protein decoy), ARAP1-AS1 can also regulate gene expression through multiple mechanisms, affecting the behavior of cancer cells. The specific details of these mechanisms are complex and require further exploration. For instance, in the context of breast cancer [45], ARAP1-AS1 was observed to enhance the expression of HDAC2 by binding to miR-2110. Subsequently, HDAC2 led to the transcriptional silencing of PLIN1 through deacetylation of the PLIN1 promoter. Moreover, it was confirmed that ARAP1-AS1 exerts a negative regulatory effect on PLIN1 expression. Thus, ARAP1-AS1 exacerbates the development of breast cancer by inducing the transcriptional suppression of PLIN1 through the deacetylation activity of HDAC2 on the PLIN1 promoter. Furthermore, Min et al. [34] observed that the increased expression of ARAP1-AS1 led to G0/G1 cell cycle arrest in lung cancer cells by suppressing cyclin D1 expression. Nevertheless, the precise mechanism through which ARAP1-AS1 modulates cyclin D1 expression, whether via miRNA or other genetic proteins, warrants additional investigation. All these findings underscore the multifaceted regulatory roles of ARAP1-AS1 in influencing gene expression, consequently, contributing regulate various biological processes in different cancer types.
LncRNA ARAP1-AS1’s participation in the competing endogenous RNA network
ARAP1-AS1 exerts control over the expression of downstream genes by engaging in competitive interactions with miRNAs, a fundamental component of the competing endogenous RNA (ceRNA) regulatory mechanism, as previously explained in earlier studies [53,54,55]. The ceRNA hypothesis proposes that extended RNA molecules, including both lncRNAs and mRNAs, which share common miRNA binding sites, participate in dynamic competitions for miRNA binding, thereby mutually influencing each other’s expression [56,57,58].
ARAP1-AS1, as an exemplar of lncRNAs, harbors numerous potential miRNA binding sites. This attribute endows ARAP1-AS1 with the capacity to regulate the expression of diverse mRNAs by competitively vying for multiple miRNAs. In turn, this regulation cascades downstream, impacting gene expression and culminating in the modulation of malignant phenotypes in tumors. Across different tumor types, ARAP1-AS1 exhibits the versatility to competitively bind various miRNAs, including miR-8068, miR-516b-5p, miR-3918, miR-4735-3p, miR-149-3p, miR-361-3p, miR-2110, and more (Fig. 4). Consequently, this competition leads to the upregulation of corresponding mRNA expression, involving genes such as CEACAM5, PDE5A, KIF20A, NOTCH2, POU2F2, PLAGL2, PGF, and HDAC2 (Fig. 4). The ensuing synthesis of these proteins contributes to the orchestration of malignant phenotypes, encompassing tumor proliferation, invasion, migration, thereby fueling the initiation and progression of tumors. It is noteworthy that ARAP1-AS1’s potential to bind additional miRNAs for the modulation of tumor development remains an intriguing avenue that warrants further exploration.

ARAP1-AS1 engages in competitive binding with miRNAs, thereby orchestrating the modulation of various mRNA expressions that drive tumor progression. ARAP1-AS1 competes for miRNAs such as miR-8068, miR-516b-5p, miR-3918, among others, resulting in the upregulation of mRNAs such as CEACAM5, PDE5A, KIF20A, and others. This molecular interplay promotes malignant characteristics like proliferation, invasion, and migration, ultimately fueling the initiation of tumorigenesis
LncRNA ARAP1-AS1 modulates downstream gene expression via direct protein binding
Apart from competing with miRNAs and regulating mRNA stability, lncRNA ARAP1-AS1 could form complexes with specific proteins, acting at the epigenetic and transcriptional levels to control gene expression (Fig. 5). In a study by Liu et al. [33], it was uncovered that ARAP1-AS1 silences ARAP1 expression by recruiting EZH2 to its promoter region, leading to increased enrichment of H3K27me on the promoter. Consequently, the suppression of ARAP1 expression promotes cell proliferation, migration, and invasion in lung adenocarcinoma. In the context of cervical cancer, ARAP1-AS1 recruits EZH2 to the DUSP5 promoter, resulting in epigenetic silencing of DUSP5. The suppression of DUSP5 through EZH2-mediated methylation further enhances cervical cancer cell proliferation and migration [41]. Additionally, Zhang et al. [42]reported that ARAP1-AS1 directly interacts with PSF to release PTB, which accelerates the translation of c-Myc, ultimately facilitating the development of progression of cervical cancer.
The ARAP1-AS1 RNA function is intricately tied to its secondary structure. Through an examination of the secondary structure of ARAP1-AS1, we have the potential to unveil additional proteins associated with its role in tumor regulation. Additionally, it’s worth noting that ARAP1-AS1 has been documented to reside both in the cytoplasm and the nucleus. Conducting subcellular localization analyses of ARAP1-AS1 in various tissues provides yet another avenue for delving into its mechanisms of tumor development regulation.

ARAP1-AS1 promotes tumor development by binding to proteins. ARAP1-AS1 epigenetically silences ARAP1 or DUSP5 by recruiting EZH2 to promote tumorigenesis of lung adenocarcinoma and cervical cancer, respectively, and ARAP1-AS1 can directly interact with PSF to release PTB, thereby accelerating c-Myc translation, and promoting occurrence of cervical cancer
Summary and future perspectives
The realm of cancer research has witnessed significant advancements with the discovery and study of lncRNAs, which play crucial roles in the regulation of gene expression and the development of various malignancies [59,60,61]. Among these, ARAP1-AS1, a recently identified lncRNA, has garnered attention for its involvement in tumor progression and potential as a biomarker for cancer diagnosis and prognosis. This review summarized the current understanding of ARAP1-AS1’s mechanisms of action, the limitations of existing studies, and propose directions for future research.
ARAP1-AS1 plays a multifaceted role in the regulation of tumorigenesis through its complex interactions at the molecular level. It primarily acts as a miRNA sponge, effectively sequestering miRNAs and preventing them from inhibiting their target oncogenes. Additionally, ARAP1-AS1 serves as a decoy for proteins, modulating the activity of RNA-binding proteins crucial for post-transcriptional regulation. These strategic interactions demonstrate that ARAP1-AS1 regulates gene expression at the epigenetic and transcriptional level, contributing further to the modulation of related signaling pathways. ARAP1-AS1 has been established as a carcinogenic lncRNA in numerous tumor types. Elevated ARAP1-AS1 expression significantly promotes malignant traits like cell proliferation, invasion, and migration in various tumors. Conversely, downregulating ARAP1-AS1 has opposing effects. These findings underscore the potential of ARAP1-AS1 as a vital anti-tumor target, and inhibiting its expression may enhance therapeutic efficacy in cancer patients. Increasingly, studies have reported the use of small molecule drugs derived from natural products to inhibit the expression of oncogenic lncRNAs, such as cucurbitacin B [62] and oridonin [63] targeting AFAP1-AS1. Thus, the exploration of small molecule compounds, particularly those from natural sources, that can inhibit ARAP1-AS1 expression holds promise as a meaningful avenue of research.
Despite these insights, there are significant gaps in our understanding of ARAP1-AS1. Many studies to date have relied heavily on in vitro assays and limited clinical investigations, highlighting the necessity for comprehensive in vivo studies and more extensive, diverse clinical trials. These studies are crucial for validating the molecular interactions and functions related to ARAP1-AS1 in a broader range of cancers, not only on human solid tumors but also in hematological malignancies. Additionally, the exploration of ARAP1-AS1’s role in larger ceRNA networks and additional tumor-related signaling pathways in different cancers remains an underexplored area with considerable potential for future research. Nevertheless, it is imperative to delve into the intricate upstream and downstream molecular mechanisms of ARAP1-AS1 to elucidate its precise role in tumor regulation. Moreover, the retrospective nature of some studies and the use of small sample sizes for evaluating ARAP1-AS1’s diagnostic and prognostic value are significant limitations. These issues underscore the need for larger, prospective studies to establish the clinical utility of ARAP1-AS1 more firmly.
To enhance this review, we leveraged multiple online lncRNA databases and high-throughput datasets to uncover the clinical relevance and potential pathway engagement of ARAP1-AS1 in the tumorigenesis and disease progression.
While the diagnostic utility of ARAP1-AS1 expression has been previously documented in cervical cancer [42], we extend this investigation to a broader array of tumor types. We analyzed data from the UCSC XENA platform (https://xenabrowser.net/datapages/), uncovering that ARAP1-AS1 expression possesses the potential to serve as a robust diagnostic biomarker across multiple cancers (Fig. 6). Notably, in pancreatic adenocarcinoma (PAAD) and uterine carcinosarcoma (UCS), ARAP1-AS1 demonstrated an AUC exceeding 0.9, highlighting its significant diagnostic promise (Fig. 6). Our focus then shifted towards evaluating ARAP1-AS1’s potential as a minimally invasive diagnostic instrument. Given the emerging recognition of exosomal lncRNAs as viable tumor diagnostic markers [64,65,66], and considering that ARAP1-AS1’s expression in blood samples has been reported only in cervical cancer [42], we utilized exoRBase 2.0 (http://www.exorbase.org/) [67]—a repository for extracellular vesicles long RNAs derived from RNA-seq data—to assess the expression of lncRNA across various human body fluids, including blood, urine, cerebrospinal fluid (CSF), and bile. Our analysis revealed that ARAP1-AS1 levels vary under different disease conditions, with the highest levels observed in the blood of melanoma (MEL) patients (Fig. 7A). A subsequent comparative analysis of ARAP1-AS1 expression in the blood of cancer patients versus healthy donors revealed significant differences in esophageal squamous cell carcinoma (ESCC), glioblastoma multiforme (GBM), and kidney renal clear cell carcinoma (KIRC) (Fig. 7B). This suggests ARAP1-AS1’s potential as a circulating diagnostic lncRNA in these cancers. However, confirmation of ARAP1-AS1 expression and its diagnostic efficacy is needed in larger clinical cohorts across various types of tumors. These observations underscore ARAP1-AS1’s potential as a promising diagnostic marker in select tumors and pave the way for its exploration as a novel circulating biomarker in future research.
Moreover, we undertook an analysis using NcPath [68], an online platform for the visualization and enrichment analysis of human non-coding RNA and KEGG signaling pathways, to identify the top 10 enriched signaling pathways involving ARAP1-AS1 (Fig. 8). The results illuminated ARAP1-AS1’s association with several cancer-related pathways, notably the TGF-beta signaling pathway, FoxO signaling pathway, and HIF-1 signaling pathway. These pathways are intrinsically linked to the initiation and progression of various tumors [69,70,71,72,73,74,75,76], highlighting the intricate role ARAP1-AS1 in cancer biology and emphasizing its significance in the study of tumorigenesis and cancer progression.

Diagnostic receiver operating characteristic curves for ARAP1-AS1 expression in distinguishing tumors from normal tissues across a broad spectrum of cancers. Notably, ARAP1-AS1 expression demonstrated significant diagnostic potential in PAAD and UCS. The abbreviations in this figure can be found in Supplementary Table 1 for reference

(A) Heat map representation of ARAP1-AS1 expression levels within extracellular vesicles, as determined by RNA sequencing analyses across a variety of human bodily fluids. (B) Comparative analysis of ARAP1-AS1 expression in extracellular vesicles from the blood of individuals with various types of cancer relative to healthy donors. Cancer types demonstrating significantly different ARAP1-AS1 expression levels when compared to those of healthy donors are marked in red. The abbreviations in this figure can be found in Supplementary Table 1 for reference

The top ten significantly enriched signaling pathways targeted by ARAP1-AS1
Conclusion
In summary, ARAP1-AS1, identified as a tumor-promoting lncRNA, exhibits elevated expression levels across various solid tumors and has gained significant attention in non-coding RNA research. It plays a pivotal role in initiating and advancing malignant tumors and holds potential as a molecular marker and therapeutic target for multiple cancer types. Nonetheless, the investigation of ARAP1-AS1 is still in its infancy. Thorough and comprehensive studies into its functions and biological significance are imperative to deepen our understanding of tumor pathogenesis. This expanded knowledge base will prove invaluable in advancing the diagnosis and treatment of cancer.
Data availability
Not applicable.
Change history
16 August 2024
A Correction to this paper has been published: https://doi.org/10.1186/s12935-024-03475-2
References
Hanahan D. Hallmarks of Cancer: New dimensions. Cancer Discov. 2022;12(1):31–46.
Siegel RL, et al. Cancer statistics, 2023. CA Cancer J Clin. 2023;73(1):17–48.
Cronin KA, et al. Annual report to the nation on the status of cancer, part 1: National cancer statistics. Cancer. 2022;128(24):4251–84.
Rumgay H, et al. Global burden of primary liver cancer in 2020 and predictions to 2040. J Hepatol. 2022;77(6):1598–606.
Palazzo AF, Lee ES. Non-coding RNA: what is functional and what is junk? Front Genet. 2015;6:2.
Comfort N. Genetics: we are the 98%. Nature. 2015;520(7549):615–6.
Bodmer W. Human Genome Project, in Brenner’s Encyclopedia of Genetics (Second Edition), S. Maloy and K. Hughes, Editors. 2013, Academic Press: San Diego. pp. 552–554.
Wang HV, Chekanova JA. An overview of methodologies in studying lncRNAs in the high-throughput era: when acronyms ATTACK! Methods Mol Biol. 2019;1933:1–30.
Statello L, et al. Gene regulation by long non-coding RNAs and its biological functions. Nat Rev Mol Cell Biol. 2021;22(2):96–118.
Micheel J, Safrastyan A, Wollny D. Advances in non-coding RNA sequencing. Noncoding RNA, 2021. 7(4).
Okuyan HM, Begen MA. LncRNAs Osteoarthr Clin Chim Acta. 2022;532:145–63.
Huang Y. The novel regulatory role of lncRNA-miRNA-mRNA axis in cardiovascular diseases. J Cell Mol Med. 2018;22(12):5768–75.
Schmitz SU, Grote P, Herrmann BG. Mechanisms of long noncoding RNA function in development and disease. Cell Mol Life Sci. 2016;73(13):2491–509.
Uchida S, Dimmeler S. Long noncoding RNAs in cardiovascular diseases. Circ Res. 2015;116(4):737–50.
Ghasemian M, et al. Long non-coding RNA MIR4435-2HG: a key molecule in progression of cancer and non-cancerous disorders. Cancer Cell Int. 2022;22(1):215.
Ghasemian M, Poodineh J. A review on the biological roles of LncRNA PTCSC3 in cancerous and non-cancerous disorders. Cancer Cell Int. 2023;23(1):184.
Xie W, et al. Emerging roles of long noncoding RNAs in chemoresistance of pancreatic cancer. Semin Cancer Biol. 2022;83:303–18.
Xu Y, et al. The emerging regulatory roles of long non-coding RNAs implicated in cancer metabolism. Mol Ther. 2021;29(7):2209–18.
Bhan A, Soleimani M, Mandal SS. Long noncoding RNA and Cancer: a New Paradigm. Cancer Res. 2017;77(15):3965–81.
Pan X, Li C, Feng J. The role of LncRNAs in tumor immunotherapy. Cancer Cell Int. 2023;23(1):30.
Lv N, et al. Long noncoding RNAs: glycolysis regulators in gynaecologic cancers. Cancer Cell Int. 2023;23(1):4.
Wang B, et al. The functional role of oncogenic LncRNA BCAR4 for Cancer Outcome. Curr Pharm Des. 2021;27(39):4107–13.
Qi Y, et al. Oncogenic LncRNA CASC9 in Cancer Progression. Curr Pharm Des. 2021;27(4):575–82.
Rajagopal T, et al. HOTAIR LncRNA: a novel oncogenic propellant in human cancer. Clin Chim Acta. 2020;503:1–18.
Li S, et al. LncRNA LENGA acts as a tumor suppressor in gastric cancer through BRD7/TP53 signaling. Cell Mol Life Sci. 2022;80(1):5.
Kim J, et al. Long noncoding RNA MALAT1 suppresses breast cancer metastasis. Nat Genet. 2018;50(12):1705–15.
Cho SW, et al. Promoter of lncRNA Gene PVT1 is a tumor-suppressor DNA boundary element. Cell. 2018;173(6):1398–e141222.
Wei GH, Wang X. lncRNA MEG3 inhibit proliferation and metastasis of gastric cancer via p53 signaling pathway Eur Rev Med Pharmacol Sci, 2017. 21(17): pp. 3850–3856.
Xie C, et al. NONCODEv4: exploring the world of long non-coding RNA genes. Nucleic Acids Res. 2014;42(Database issue):D98–103.
Cao Z, et al. The lncLocator: a subcellular localization predictor for long non-coding RNAs based on a stacked ensemble classifier. Bioinformatics. 2018;34(13):2185–94.
Gao Y, et al. Lnc2Cancer 3.0: an updated resource for experimentally supported lncRNA/circRNA cancer associations and web tools based on RNA-seq and scRNA-seq data. Nucleic Acids Res. 2021;49D1:pD1251–d1258.
Wu Z et al. LncRNA ARAP1-AS1 contributes to lung adenocarcinoma development by targeting miR-8068 to upregulate CEACAM5. Cancer Biomark, 2023.
Liu J, et al. Long noncoding RNA ArfGAP with RhoGAP domain, ankyrin repeat and PH domain 1 antisense RNA 1 recruits enhancer of zeste 2 polycomb repressive complex 2 subunit to promote the proliferation, migration and invasion of lung adenocarcinoma cells. Bioengineered. 2022;13(3):7868–80.
Tao X, et al. Low expression of long non-coding RNA ARAP1-AS1 can inhibit lung cancer proliferation by inducing G0/G1 cell cycle organization. J Thorac Dis. 2020;12(12):7326–36.
Pang B et al. lncRNA ARAP1-AS1 enhances proliferation and impairs apoptosis of lymphoma cells by sponging miR-6867-5p Cancer Biomark, 2023.
Du Q, Huang L, Guo W. LncRNA ARAP1-AS1 targets miR-516b-5p/PDE5A axis to facilitate the progression of thyroid cancer. Anticancer Drugs. 2023;34(6):735–46.
Zhang W, et al. LncRNA ARAP1-AS1 promotes bladder Cancer Development by regulating the miR-3918/KIF20A Axis. Mol Biotechnol. 2022;64(11):1259–69.
Xu Z, et al. Characterization of mRNA expression and endogenous RNA profiles in bladder Cancer based on the Cancer Genome Atlas (TCGA) database. Med Sci Monit. 2019;25:3041–60.
Teng J, et al. Long non-coding RNA ARAP1-AS1 promotes the progression of bladder cancer by regulating miR-4735-3p/NOTCH2 axis. Cancer Biol Ther. 2019;20(4):552–61.
Zhou L, Xu XL. Long non-coding RNA ARAP1-AS1 facilitates the progression of Cervical Cancer by regulating mir-149-3p and POU2F2. Pathobiology. 2021;88(4):301–12.
Min H, He W. Long non-coding RNA ARAP1-AS1 promotes the proliferation and migration in cervical cancer through epigenetic regulation of DUSP5. Cancer Biol Ther. 2020;21(10):907–14.
Zhang Y, Wu D, Wang D. Long non-coding RNA ARAP1-AS1 promotes tumorigenesis and metastasis through facilitating proto-oncogene c-Myc translation via dissociating PSF/PTB dimer in cervical cancer. Cancer Med. 2020;9(5):1855–66.
Li C, et al. LncRNA ARAP1-AS1 aggravates the malignant phenotypes of ovarian cancer cells through sponging mir-4735-3p to enhance PLAGL2 expression. Cytotechnology. 2021;73(3):363–72.
Zhong L, Zhong X. Long non-coding RNA ARAP1-AS1 contributes to cell proliferation and migration in clear cell renal cell carcinoma via the miR-361-3p/placental growth factor axis. Bioengineered. 2021;12(1):6629–42.
Lu C et al. Long non-coding RNA ARAP1-AS1 accelerates cell proliferation and migration in breast cancer through miR-2110/HDAC2/PLIN1 axis. Biosci Rep, 2020. 40(4).
Jiang YH, et al. Increased long non-coding RNA ARAP1-AS1 expression and its prognostic significance in human gastric cancer: a preliminary study. Eur Rev Med Pharmacol Sci. 2020;24(4):1815–20.
Ye Y, et al. YY1-Induced Upregulation of Long Noncoding RNA ARAP1-AS1 promotes Cell Migration and Invasion in Colorectal Cancer through the Wnt/β-Catenin signaling pathway. Cancer Biother Radiopharm. 2019;34(8):519–28.
Tomczak K, Czerwińska P, Wiznerowicz M. The Cancer Genome Atlas (TCGA): an immeasurable source of knowledge. Contemp Oncol (Pozn). 2015;19(1a):A68–77.
Yan X, et al. Comprehensive genomic characterization of long non-coding RNAs across human cancers. Cancer Cell. 2015;28(4):529–40.
Ye B, et al. Advancing pan-cancer gene expression survial analysis by inclusion of non-coding RNA. RNA Biol. 2020;17(11):1666–73.
Li C, et al. Correction to: LncRNA ARAP1-AS1 aggravates the malignant phenotypes of ovarian cancer cells through sponging mir-4735-3p to enhance PLAGL2 expression. Cytotechnology. 2022;74(1):201.
Tang Z, et al. GEPIA2: an enhanced web server for large-scale expression profiling and interactive analysis. Nucleic Acids Res. 2019;47(W1):W556–60.
Kartha RV, Subramanian S. Competing endogenous RNAs (ceRNAs): new entrants to the intricacies of gene regulation. Front Genet. 2014;5:8.
David R. A new layer of regulation. Nat Rev Mol Cell Biol. 2011;12(12):766–766.
Salmena L, et al. A ceRNA hypothesis: the Rosetta Stone of a hidden RNA language? Cell. 2011;146(3):353–8.
Tay Y, Rinn J, Pandolfi PP. The multilayered complexity of ceRNA crosstalk and competition. Nature. 2014;505(7483):344–52.
Karreth FA, Pandolfi PP. ceRNA cross-talk in cancer: when Ce-bling rivalries go awry. Cancer Discov. 2013;3(10):1113–21.
Qi X, et al. ceRNA in cancer: possible functions and clinical implications. J Med Genet. 2015;52(10):710–8.
Bartonicek N, Maag JLV, Dinger ME. Long noncoding RNAs in cancer: mechanisms of action and technological advancements. Mol Cancer. 2016;15(1):43.
Engreitz JM, et al. Local regulation of gene expression by lncRNA promoters, transcription and splicing. Nature. 2016;539(7629):452–5.
Prensner JR, Chinnaiyan AM. The emergence of lncRNAs in cancer biology. Cancer Discov. 2011;1(5):391–407.
Zhou J, et al. Cucurbitacin B suppresses proliferation of pancreatic cancer cells by ceRNA: effect of miR-146b-5p and lncRNA-AFAP1-AS1. J Cell Physiol. 2019;234(4):4655–67.
Lou S, et al. Downregulation of lncRNA AFAP1-AS1 by oridonin inhibits the epithelial-to-mesenchymal transition and proliferation of pancreatic cancer cells. Acta Biochim Biophys Sin (Shanghai). 2019;51(8):814–25.
Zeng Y et al. Exosome cargos as biomarkers for diagnosis and prognosis of Hepatocellular Carcinoma. Pharmaceutics, 2023. 15(9).
Lin H, et al. Exosomal long noncoding RNAs in NSCLC: dysfunctions and clinical potential. J Cancer. 2023;14(10):1736–50.
Tao Y, et al. Exploration of serum exosomal LncRNA TBILA and AGAP2-AS1 as promising biomarkers for diagnosis of Non-small Cell Lung Cancer. Int J Biol Sci. 2020;16(3):471–82.
Lai H, et al. exoRBase 2.0: an atlas of mRNA, lncRNA and circRNA in extracellular vesicles from human biofluids. Nucleic Acids Res. 2021;50(D1):D118–28.
Li Z et al. NcPath: a novel platform for visualization and enrichment analysis of human non-coding RNA and KEGG signaling pathways. Bioinformatics, 2023. 39(1).
Alharbi KS. The ncRNA-TGF-β axis: unveiling new frontiers in colorectal cancer research. Pathol Res Pract. 2024;254:155138.
Ahuja S, Zaheer S. Multifaceted TGF-β signaling, a master regulator: from bench-to-bedside, intricacies, and complexities. Cell Biol Int. 2024;48(2):87–127.
Farhan M, et al. FOXO Signaling Pathways as therapeutic targets in Cancer. Int J Biol Sci. 2017;13(7):815–27.
Dilmac S et al. SIRT1/FOXO signaling pathway in breast Cancer progression and metastasis. Int J Mol Sci, 2022. 23(18).
Yuan J, et al. Long intergenic non-coding RNA DIO3OS promotes osteosarcoma metastasis via activation of the TGF-β signaling pathway: a potential diagnostic and immunotherapeutic target for osteosarcoma. Cancer Cell Int. 2023;23(1):215.
Li YM, et al. Piezo1 promoted hepatocellular carcinoma progression and EMT through activating TGF-β signaling by recruiting Rab5c. Cancer Cell Int. 2022;22(1):162.
Zhou X, et al. THBS1 promotes angiogenesis and accelerates ESCC malignant progression by the HIF-1/VEGF signaling pathway. Cell Biol Int. 2024;48(3):311–24.
Shen J, et al. β-catenin/TCF4-induced SCUBE3 upregulation promotes ovarian cancer development via HIF-1 signaling pathway. Mol Cell Endocrinol. 2024;582:p112127.
Acknowledgements
Not applicable.
Funding
Not applicable.
Author information
Authors and Affiliations
Contributions
All authors made a significant contribution to the work reported, whether that is in the conception, study design, execution, acquisition of data, analysis and interpretation, or in all these areas; took part in drafting, revising or critically reviewing the article; gave final approval of the version to be published; have agreed on the journal to which the article has been submitted; and agree to be accountable for all aspects of the work.
Corresponding author
Ethics declarations
Ethics approval and consent to participate
Not applicable.
Consent for publication
Not applicable.
Competing interests
The authors declare no competing interests.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Wang, J., Luo, H., Yang, L. et al. ARAP1-AS1: a novel long non-coding RNA with a vital regulatory role in human cancer development. Cancer Cell Int 24, 270 (2024). https://doi.org/10.1186/s12935-024-03435-w
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12935-024-03435-w