
Abstract
Recorded knowledge of marine mammals of the Gulf began with commercial whaling of sperm whales, short-finned pilot whales, and Risso’s dolphins in the 1700s and 1800s, progressed to natural history observations and one of the first volunteer stranding organizations, the Texas Marine Mammal Stranding Network, in the 1970s. The vegetarian sirenian, the West Indian manatee (Trichechus manatus), occurs mainly in Florida. All others are cetaceans, members of the whale and dolphin clades. There are no porpoises, sea lions, fur seals, or true seals in the Gulf. The most common cetacean in the Gulf numerically is the pantropical spotted dolphin (Stenella attenuata), and the one with most biomass is the sperm whale (Physeter macrocephalus), common in mid-depth waters off Louisiana and the shelf break off Texas. Major anthropogenic threats exist for all marine mammals but do not appear to be as intensive in the Gulf as in several other oceanic basins. Threats to marine mammals in the Gulf include prey depletion, incidental mortality, injury due to fisheries, intentional and direct takes, vessel strikes, disturbance, acoustic (noise) pollution, chemical contamination, ingestion of solid debris, oil spills, and aspects of ecosystem change.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


Keywords
- Short-finned pilot whale
- Risso’s dolphin
- West Indian manatee
- Pantropical spotted dolphin
- Sperm whale
- Threats
13.1 Introduction
The marine mammals of the Gulf of Mexico consist of whales, dolphins and one species of coastal sirenian, the West Indian manatee (Trichechus manatus). There are no seals, sea lions, fur seals, or sea-going otters as are present in many other parts of the world. One tropical seal, the Caribbean monk seal (Monachus tropicalis), which was apparently never abundant in the Gulf, became extinct in the early part of the twentieth century. At about the same time, California sea lions (Zalophus californianus) escaped from small zoos and, for a while, appeared to be reproducing and establishing a feral population, but that population is also gone. There are no porpoises of the cetacean phocoenid family in the Gulf. Only the harbor porpoise (Phocoena phocoena) occurs in the North Atlantic, and waters around southern Florida are too warm for this species to have made a foray into the Gulf.
The cetaceans of the Gulf are diverse and well established, ranging from the ubiquitous, nearshore (and there is an offshore variant) common bottlenose dolphin (Tursiops truncatus), hereafter referred to as bottlenose dolphin unless differentiation is warranted from the Indo-Pacific bottlenose dolphin (Tursiops aduncus), to the sperm whale (Physeter macrocephalus), the largest toothed whale in the oceans and the largest toothed creature. The Gulf of Mexico is home to several species of continental shelf and deep ocean dolphins or whales of the family Delphinidae as well as deepwater beaked whales of the family Ziphiidae. There are baleen whales (infraorder Mysticeti) in the Gulf as well, members of the family Balaenopteridae from the relatively small minke (Balaenoptera acutorostrata) to the giant of the seas, the largest mammal on Earth, the blue whale (Balaenoptera musculus). Of the baleen whales, only the Bryde’s whale (Balaenoptera edeni) is a resident of the northern Gulf.
Each major marine mammal grouping in the Gulf has evolved to make its living in quite a different way. The manatee is one of only four marine mammals (all of the order Sirenia) that feed on seagrasses and other plant material. Its common name, sea cow, is quite appropriate as manatees are indeed related to early ungulates and have a ruminant stomach somewhat similar to their terrestrial forebears. The baleen whales have a structure of keratinous material—the baleen plates—that hang from the upper jaw with finely fringed inner hairs that form a filter mat and allow for batch feeding. Although baleen whales have three different feeding methods (Heithaus and Dill 2009), the Bryde’s whale, which dominates the baleen whale fauna of the Gulf, is a lunge feeder that uses a technique that has aptly been described as the greatest biomechanical action on Earth (Croll et al. 2001). The toothed whales (infraorder Odontoceti) of the Gulf, which range from the giant sperm whale to the small Clymene dolphin (Stenella clymene), have all evolved the ability to echolocate—use high-frequency sound—to sense their conspecifics, potential danger from sharks and killer whales (Orcinus orca), obstructions in the water as well as the surface and bottom, and—important at all times—potential prey. Some of the toothed whales have also evolved extremely large, complex brains, probably due to the communication needs of sophisticated social living (Marino 2004). Large brains may also be thought of as a specialization that has allowed this group to become highly diverse and successful in a generally forbidding sea.
The present description of the cetaceans and one siren of the Gulf is an update of information in the books by Würsig et al. (2000) and Jefferson et al. (2008) as well as articles by Baumgartner et al. (2001), Davis et al. (2002), Mullin and Fulling (2004), Maze-Foley and Mullin (2006), Schmidly and Würsig (2009), Waring et al. (2010), Schick et al. (2011), and others. Few published comprehensive summaries of marine mammals of the Gulf of Mexico have been made in the past 12 years, so this update comes at a particularly opportune time. The estimated numbers of animals per species represented here for the northern (United States [U.S.]) portion of the Gulf are from the official National Oceanic and Atmospheric Administration-National Marine Fisheries Service (NOAA-NMFS) Office of Protected Resources marine mammal stock assessment reports (Waring et al. 2011),Footnote 1 unless stated otherwise. Information in the primary literature has vastly increased present knowledge, especially about bottlenose dolphins and sperm whales. However, aspects of community structure in pelagic cetaceans are only slowly coming to light. Since the publication of Würsig et al. (2000), a PhD dissertation by Ortega-Ortiz (2002) summarized information about cetaceans and cetacean habitats of the Mexican southern waters of the Gulf. This chapter does not cover much of this newer information south of U.S. waters, but a synopsis and comparison with U.S. waters is made. With the exception of several discrete areas for bottlenose dolphins in select bays and estuaries, population estimates for the Mexican and Cuban parts of the Gulf are not available.
This chapter is organized into seven major sections designed to give the reader a flavor of these charismatic megafauna, as even the smallest of dolphins in the oceans is a large—and large brained—social creature among mammals in general. Major chapter sections are as follows: (1) a general introduction; (2) history of research; (3) basic species, habitat, and number descriptions, if available; (4) discussion of anthropogenic impacts; (5) conclusions that summarize present baseline conditions; (6) references; and (7) two appendices. Throughout this chapter, cetaceans and the one sirenian of the Gulf of Mexico are described in context with their distributions and habitat preferences worldwide, as these animals are far ranging; none of the species discussed occurs only in the Gulf (i.e., none is endemic).
The Würsig et al. (2000) book describes 31 species of cetacean plus the one sirenian known or believed to be in the Gulf as of its writing in the late 1990s. It is now quite certain that three species—the more-northerly occurring long-finned pilot whale (Globicephala melas) and the short-beaked and long-beaked common dolphins (Delphinus delphis and Delphinus capensis, respectively)—can be taken off the list of potential Gulf inhabitants, with the recognition that quite a few species might eventually be discovered as rare vagrants. These three species are not included in this chapter. Common dolphins were quite often cited to occur in the Gulf, but apparently all such descriptions were due to confusion with other species, most commonly Clymene dolphins (Jefferson 1995; Jefferson and Schiro 1997). A further seven species have records for the Gulf but also occur so rarely that they are mentioned only in passing. Six of these are baleen whales: Atlantic right (Eubalaena glacialis), blue, fin (Balaenoptera physalus), sei (Balaenoptera borealis), minke (Balaenoptera acutorostrata), and humpback (Megaptera novaeangliae) whales. This leaves only the tropical Bryde’s whale as a reliable baleen whale of the Gulf, which means that with modern evidence it has become apparent that the baleen whale fauna of the Gulf is less rich than was believed even a decade ago. The seventh species is a beaked whale—Sowerby’s beaked whale (Mesoplodon bidens). However, beaked whales are poorly described in the Gulf, and it would not be surprising to have several added to the list with further knowledge.
13.2 History of Research in the Gulf
13.2.1 Whaling
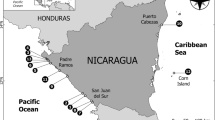
While whaling by itself is not research, much important detail on distribution can be gleaned from whaling records, and the Gulf of Mexico is no exception. The best known records of Gulf whaling come from the worldwide Townsend Charts (Townsend 1935), which illustrate where, when, and how many whales were taken, as recorded from logbook records of U.S. (American or Yankee) whaling ships. These are thus somewhat biased, but they provide a general review. This information has recently been updated and expanded (Reeves et al. 2011), and new insights from this re-evaluation give a modern perspective of at least some species than previously was available for the Gulf (Figure 13.1).

Positions of whaling ships on days when sperm whales were sighted or caught. Dotted lines represent the 100 m (328 ft) and 1,000 m (3,281 ft) isobaths, and the lines between Cuba and Florida (top right) and the Mexican Yucatán Peninsula (bottom left) represent the approximate extent of the Gulf of Mexico to the southeast, as considered for marine mammals in this report. Whaling ship positions are from Reeves et al. (2011), p 44. It is instructive to compare this figure of sperm whale whaling with those of Figures 13.9 and 13.10 of modern sperm whale sightings in the northern and southern Gulf, respectively.
Clark (1887) mentions that right whales were taken in the Gulf by eighteenth century and nineteenth century Yankee whalers, but this claim was not substantiated by Townsend (1935) or by others (Reeves et al. 2011). Instead, it is clear that the largest whaling effort—starting in 1788 and apparently ending in 1877 (with perhaps a few sporadic attempts into the twentieth century)—was on sperm whales, with occasional takes of so-called finbacks (Balaenoptera sp.), so-called porpoises (small delphinids), and killer whales. Besides sperm whales, another deep diving odontocete cetacean, the short-finned pilot whale (Globicephala macrorhynchus), was quite commonly killed. Grampus [probably Risso’s dolphin (Grampus griseus)] was also taken. The whaling records of all three of these major whaled species overlap with what is known of present day distribution in the northern Gulf. Whaling records will be discussed in later detailed sections on the sperm whale, pilot whale, and Risso’s dolphin.
The Gulf of Mexico is one of the few oceanic areas that were exploited only in early U.S. history, predominantly by takes of sperm whales. With the exception of some catches of nearshore bottlenose dolphins for the aquarium trade, there has been little exploitation for about the past 130 years. This remarkable fact allows investigation of these long-lived animals as relatively unperturbed, save for the strong buildup of shipping and industrial activities.
13.2.2 Early Opportunistic Research
Prior to 1977, reports of whales and dolphins came largely from fishermen’s and other boaters’ sightings and publications of strandings. These data were compiled by Gordon Gunter in a long series of papers (Gunter 1954 is but one example) and by J. C. Moore (1953), J. N. Layne (1965), Caldwell and Caldwell (1973), Lowery (1974), and Schmidly and Shane (1978). In 1980, David Schmidly started the Texas Marine Mammal Stranding Network (TMMSN) (Schmidly 1981), the first comprehensive volunteer network to obtain and report detailed biological information on strandings of live and dead marine mammals in the Gulf. Until systematic line-transect efforts from airplanes and boats began, both at about the same time, strandings provided the best information on cetaceans in the northern Gulf (with practically no information for the southern Gulf) and gave the first appreciation that there are many species of deepwater cetaceans.
13.2.3 The Modern Era
The TMMSN continues to this day and is now integrated closely with the U.S.-wide NMFS Stranding Network and specifically the Southeast Region Marine Mammal Stranding Network that includes states from Texas in the southwest to North Carolina on the north-central Atlantic coast as well as Puerto Rico and the U.S. Virgin Islands. The Office of Protected Resources, NOAA Fisheries Service Headquarters, Silver Spring, Maryland, coordinates protected resources and research programs under the auspices of the U.S. Marine Mammal Protection Act (MMPA) of 1972.Footnote 2 Manatees are covered by the U.S. Fish and Wildlife Service (USFWS), and there is some good interface in analyses of habitat use for coastal marine mammals by the NMFS and USFWS.Footnote 3
Even before the TMMSN, there were two studies on local bottlenose dolphins—one begun by Randall Wells and colleagues in the early 1970s in the Sarasota-Bradenton area of west-central Florida (Wells 2003) and another by Susan Shane in the mid-1970s at Aransas Pass, Texas (Shane 1977). Later, Shane compared her Aransas Pass work with the results of a new study in the 1980s off Sanibel Island, Florida, almost precisely east of her former study area in Texas (Shane 2004). The Wells studies have continued and expanded and spawned several 100 publications resulting in one of the best-known cetacean populations in the world. A summary of the major findings from this set of studies is included in the section devoted to bottlenose dolphins in the Gulf.
The first systematic offshore efforts to describe marine mammals in parts of the northern Gulf of Mexico occurred during the Fritts Surveys, 1979–1981 aerial surveys led by T. H. Fritts (Fritts et al. 1983). These surveys established that several dolphins of the genus Stenella occur off continental shelf and slope waters, but these early surveys (originally designed for sea turtle and sea bird assessments) suffered from small cetacean sample size, precluding abundance estimates, and some unfortunate misidentifications. For example, they did not distinguish between the two species of spotted dolphins (Stenella attenuata and Stenella frontalis), now known to have quite different distribution patterns in the Gulf; at times the surveys confused spinner and Clymene dolphins (Stenella longirostris and Stenella clymene, respectively), and sometimes they misidentified either Clymene or pantropical spotted dolphins (Stenella attenuata) as short-beaked common dolphins. Common dolphins do not presently occur in the Gulf, nor are there verified historical records.
The NMFS conducted a series of aerial surveys in continental and coastal waters (continental shelf aerial surveys, 1983–1986; red drum aerial surveys, 1986–1987) and established some estimates of numbers of the predominantly sighted cetacean, the common bottlenose dolphin.
Since 1989, aerial and shipboard surveys have been conducted (albeit not in every year) in both coastal and deeper oceanic U.S. waters, commencing with Keith Mullin’s 1989–1990 continental slope aerial surveys that for the first time properly documented distribution and abundance of oceanic cetaceans in waters off Louisiana and Mississippi (Mullin et al. 1994; Jefferson and Schiro 1997) (Figure 13.2). In the 1990s, NMFS and Texas A&M University coordinated dedicated aerial and boat surveys in concert with descriptions of physical and biological oceanography—the Gulfcet surveys. These Gulfcet surveys provided a more detailed knowledge of how cetaceans are affected by seasonal and interyear climatic conditions, including the presence and positions of eddies and gyres that spin off the Loop Current in the north-central Gulf and the periodic, and at times strong, fresh water incursions from the Mississippi/Atchafalaya River system (Davis et al. 1998, 2002; Baumgartner et al. 2001; Ortega-Ortiz 2002). Such work has continued in the first 10 years of this century, although not always with similar multidisciplinary input as in the 1990s. The sperm whale seismic surveys (SWSS) of the first 6 years of the 2000s revealed much about sperm whale habitat in the north-central Gulf (Jochens et al. 2008), and Keith Mullin has continued to lead NMFS surveys (Maze-Foley and Mullin 2006).

Example of the many surveys conducted in the northern Gulf of Mexico, with 100 and 2,000 m (328 and 6,562 ft) isobaths, and the boundary of the U.S. Exclusive Economic Zone (marked as EEZ). Thinner lines represent NMFS vessel surveys 1991–2001 (but not in all years). Note that there is much more survey effort between 200 and 2,000 m (656 and 6,562 ft) depths than in abyssal waters deeper than 2,000 m (6,562 ft) (from Maze-Foley and Mullin 2006).
Studies, now better integrated year to year with standardized visual (and often acoustic) data gathering and analysis techniques, are finally giving a more complete picture of the cetaceans of the northern Gulf of Mexico. Such long-term information is allowing for assessments of community structure of pelagic cetaceans at large spatial scales, as recently attempted by Schick et al. (2011). Data for the southern Gulf are much less complete, except for an analysis by Ortega-Ortiz (2002); and for western nearshore areas in the state of Veracruz, there are analyses by Galindo et al. (2009), Martínez-Serrano et al. (2011), and Valdes-Arellanes et al. (2011). The shores of Cuba are poorly represented, except for a thesis by Pérez-Cao (2004) from the Camagüey Archipelago, NE Cuba, and outside of the Gulf.
Studies of bottlenose dolphins in nearshore and offshore waters deserve special attention, since bottlenose dolphins of the Sarasota-Bradenton area of west Florida are arguably the longest continuously studied marine mammal (Wells and Scott 2009), vying for this distinction only with killer whales of several pods in the coastal Pacific near the U.S.–Canadian border (Ford 2009). Other studies also have been carried out, most notably by Shane off south Texas and southwest Florida (Shane 1977, 2004; Shane et al. 1986), Mullin (1988) in the north-central Gulf, and students of Würsig (e.g., Bräger et al. 1994; Maze 1997; Moreno 2005) off the northwest Gulf, near Galveston Texas. Most of these studies have relied heavily on photo-identification for mark-recapture numbers analyses and society descriptions, and more is now known about the generally open, fluid social systems of bottlenose dolphins than of the social systems of most other cetaceans.
The study of Caribbean (West Indian) manatee, one of only four species of the taxonomic order Sirenia (Figure 13.3) and the only vegetarians among the 122 or so species of marine mammals, had a similar early introduction by Gordon Gunter in his work with cetaceans (Gunter 1941). The monograph by Daniel Hartman (1979) on ecology and behavior of manatees in Florida (work carried out in the late 1960s) finally gave detailed information on their preferred habitats, life-history strategies, foraging techniques, and social/sexual and other aspects of behavior and behavioral ecology. While there has been much updated work (e.g., Reynolds and Odell 1991; O’Shea et al. 1995; Marsh et al. 2011), this early monograph still stands as a hallmark of marine mammal studies.

Manatees of the world. There are three species, and the Caribbean (or West Indian) manatee has the largest distribution, divided into two subspecies, shown in blue in the north and in red south of this. Both subspecies occur in the Gulf. The Florida subspecies range shown in this figure includes outlying areas, and only occasionally in summer do a few manatees occur as far north as Long Island, New York, and as far west as Louisiana, as shown here (adapted from Gonzalez-Socoloske and Olivera-Gomez 2012, after an International Union for Conservation of Nature (IUCN) species map).
Since Hartman (1979), it is now known that there are two subspecies of Caribbean manatee—the Florida manatee (Trichechus manatus latirostris) and the Antillean manatee (Trichechus manatus manatus) (Figure 13.3). The Florida manatee regularly occurs on both sides of Florida (i.e., into the eastern Gulf of Mexico and north to about Tampa), although more rare excursions to northwest Florida, Alabama, Mississippi, and Louisiana occur, while the Antillean manatee occurs in the southern Mexican part of the Gulf and all the way south to (and even south of) the estuary of the Amazon River, where it is believed to hybridize with the much smaller-bodied Amazon manatee (Trichechus inunguis). Marsh et al. (2011) provide excellent up-to-date information on all Sirenia.
In the Gulf, Florida manatees rely in large part on eight major warm-water refugia, caused by both natural warm-water springs and industrial power plants. This is a worry for wildlife managers especially in the northern part of that range such as the Tampa Bay Apollo Beach (or Big Bend) power plant (27° 48′ N). Manatees are warm-water creatures with low metabolic rates, and they quickly become cold-stressed in temperatures at or less than 17 °C; (63° Fahrenheit [F]), a common occurrence in winter off the central Florida coast and a cataclysmic event for manatees when there is a need to shut down a power plant (Laist and Reynolds 2005).
13.3 Species Summaries, Habitat Use, and Numbers
13.3.1 Overview of Species
Except for coastal bottlenose dolphins and Florida manatees, concepts of numbers, group sizes, seasonal and interyear distributions, and habitat use of ocean-going cetaceans in the Gulf are far from complete, and little is known about the Gulf-inhabiting Antillean manatee in Mexican waters. Nevertheless, in large part due to multidisciplinary studies carried out especially by Texas A&M University and the U.S. NMFS in the 1990s, the Gulfcet studies, and subsequent aerial and behavioral censuses by NMFS since then, some basics are known. Much of the NMFS work was conducted from research vessels and was usually in conjunction with ichthyoplankton research (e.g., Maze-Foley and Mullin 2006). However, manatee and cetacean stock assessments contain a great deal of uncertainty, and present knowledge is quite incomplete regarding what is a separate stock (i.e., population) and how many animals are contained therein.
Costly aerial and shipboard surveys in open ocean waters can rarely provide the kind of fine-scale survey lines and repetitions of surveys (in all seasons and with variabilities from year to year) needed to provide numbers estimates with low variances. Thus, surveys of the same areas in different years can provide quite different number estimates for certain species. All that can be done is present the recent ones for which the authors express greatest confidence.
While Würsig et al. (2000) described 32 potential and known species for the Gulf, several of these are likely not found at all, and several others are so rare or vagrant that they need only cursory treatment. The potential species are listed in Table 13.1 with bold highlights for those known to occur in the northern Gulf regularly enough for there to be reliable information on basic group size, sea surface temperature, and depth of occurrence (see Table 13.2 for the latter).
The North Atlantic right, blue, fin, humpback, minke, and sei whales are species that were historically present in very low numbers and are only occasionally sighted or stranded. In other words, the only baleen whale found regularly during surveys in the northern Gulf is the Bryde’s whale. This is a strong departure from what was understood in the 1990s when the Würsig et al. (2000) book was being prepared, which means that the Gulf is less rich in baleen whale diversity than originally supposed. Additionally, while there are often unidentified beaked whales sighted in the Gulf, there are reliable records only for Cuvier’s, Blainville’s, and Gervais’ beaked whales (Ziphius cavirostris, Mesoplodon densirostris, and Mesoplodon europaeus, respectively). Sowerby’s beaked whales and several others are possibly greater in number than modern records indicate, but Sowerby’s are dropped from the list during this treatment due to paucity of confirmed sightings. The colder-water long-finned pilot whale also does not seem to be present and neither do short-beaked or long-beaked common dolphins. As was mentioned before, the quite copious previous records of the genus Delphinus were due to misinterpretation of sightings and strandings, and they may not have occurred in the Gulf in recent (or even earlier) history.
Three species habitually occur in waters less than 200 m (656 ft) deep: the inshore/coastal manatee, inshore/coastal and shelf bottlenose dolphin, and shelf Atlantic spotted dolphin (Stenella frontalis); another 19 species—all but the Bryde’s whale being toothed cetaceans—occur over the continental slope and into deep oceanic waters. No species in the Gulf of Mexico are endemic to the Gulf, and all but the warm-water North Atlantic endemic Caribbean manatee and Clymene and Atlantic spotted dolphins occur in other oceans. Of those that occur beyond Atlantic Ocean waters, the bottlenose dolphin and sperm and killer whales are considered cosmopolitan as they occur in tropical to quite cold waters; all others are tropical, occurring in Atlantic, Pacific, and Indian Ocean waters. While the sperm whale is considered endangered by the United States, it likely is not endangered globally. This leaves the fortunate situation that in the Gulf there are no truly threatened or endangered marine mammal species except for the manatee. Since bottlenose dolphins exist as rather discrete populations in separate nearshore systems and the sperm whale is a separate population in the Gulf, these species are of concern at the population level.
13.3.2 Species, Habitats, and Numbers
To orient the reader to how the marine mammals tend to occur worldwide and in the Gulf, a species-by-species description is in order with approximate numbers in the northern Gulf and related details. Population estimates often are approximate, even for the better-known species such as bottlenose dolphins and sperm whales. Because they exist in a huge area, there are strong sighting variations by season and sighting conditions. Line-transect sampling from airplanes and surface vessels is not an exact science, and mark-recapture population estimates can be made for only some special small and well-studied areas, such as the Sarasota-Bradenton area of west Florida for bottlenose dolphins. Most of the information for this summary comes from Würsig et al. (2000), Maze-Foley and Mullin (2006), Jefferson et al. (2008), Perrin et al. (2009), Schmidly and Würsig (2009), Marsh et al. (2011), and Waring et al. (2011), but not for data beyond March 2010; other data sources will also be cited.
It is here emphasized that population discreteness and number estimates for cetaceans and the one sirenian of the area are quite incomplete—different years yield different results, often quite widely so. While acoustic censuses along with visual ones have been touted (e.g., Davis et al. 2002), there are presently not enough data on frequency and other parameter types of sounds to say much about approximate numbers of animals beyond the possible exception of sperm whales. Recent correlations of select cetacean sounds and visual sightings are promising for improving the accuracy of future descriptions of cetacean presence (Baumann-Pickering et al. 2010).
Since about 65 % of the Gulf of Mexico is part of the nation of Mexico, it is appropriate that Spanish names are linked with English ones. The common Mexican names are in Table 13.3.
13.3.2.1 Bryde’s Whale
The Bryde’s whale (Balaenoptera edeni) is a small member of the Family Balaenopteridae, also called rorquals, the latter a Norwegian term referring to their throat grooves (ventral pleats) that allow them to lunge forwards into water with concentrated prey, open up the mouth/throat to prodigious volume, push the water out through a filter mat formed inside the mouth by finely fringed baleen, and swallow the euphausiid crustaceans (krill) or fish that were engulfed. Bryde’s whales are approximately 13 m (43 ft) long and weigh about 12,000 kg or 12 metric tons (26,455 lbs) or 12 metric tons (13 t). As is the case for other balaenopterids and indeed baleen whales in general, females tend to be slightly larger than males. The female’s larger size probably aids in gestating a 4 m (13 ft) long calf for 11 months and nursing it intensively for about 6 months; the average calf is 7 m (23 ft) long at weaning.
Bryde’s whales are bluish/black above and whitish below and have a small dorsal fin that rises abruptly in front and is falcate (back-curved). The Bryde’s whale’s dorsal fin has a more abrupt rise and is of course taller than the dorsal fin of most delphinids. It reaches about 46 centimeters (cm) (18 in.) high in an adult. The Bryde’s whale has 40–70 throat grooves that extend all the way back to the navel and three dorsal head ridges—one in the middle of the upper head and two about halfway between midline and jaw line. These ridges are a special distinguishing visual diagnostic feature, as other balaenopterids have only one. Bryde’s whales are often confused with the slightly larger sei whale, and a good view of the head dorsal ridges may be needed for positive identification (Figures 13.4 and 13.5).

Bryde’s whale (Balaenoptera edeni). The dorsal fin front rises abruptly from the back, allowing for quick distinction from the fin whale, whose dorsal fin rises more gently (photo by Thomas A. Jefferson, with permission).

Bryde’s whale (Balaenoptera edeni). The top of the head has two ridges to either side of the center ridge, distinguishing it from other rorquals at close range (photo by Thomas A. Jefferson, with permission).
Bryde’s whales are also termed the tropical whale, for they (and their recently named generic counterpart, Omura’s whale (Balaenoptera omurai), of the tropical western Pacific and Indian Ocean) are the only baleen whale species not to be found in colder temperate waters at least part of the year. Probably as a result of their tropical nature, they tend to breed and calve year-round and do not engage in long migrations. They tend not to be highly social, and when several Bryde’s whales are seen in proximity, they are likely to be in feeding, not social, aggregations. They tend to feed not on krill but on shoals of small fishes and are capable of engulfing an entire school of fish—1 m (3 ft) or more in diameter—at or below the surface.
The worldwide population in the Atlantic, Pacific, and Indian Oceans is estimated at about 30–50 thousand whales; they are not considered endangered worldwide and are not listed as endangered under the U.S. Endangered Species Act. Until recently, best estimate for the northern Gulf was about 40, with 95 % confidence interval (CI) of 13–129 (Mullin and Fulling 2004; NMFS Stock Assessment Reports 2006–2011), but a smaller best estimate of 15 (coefficient of variation [CV] 1.98) whales (Mullin 2007) from more recent data is presently published by NMFS as the official estimate (Table 13.4). This does not necessarily mean that the population in the Gulf has declined. It simply may reflect the vagaries of sighting members of a small population depending on chance in particular line-transect surveys (Table 13.4). There is not enough information on Bryde’s whales in the Gulf to determine population trend or whether there is fisheries-related or other human-caused mortality (Figure 13.6).

Distribution of Bryde’s whale (Balaenoptera edeni) sightings from the Southeast Fisheries Science Center (SEFSC) spring vessel surveys during 1996 to 2001 and from summer 2003 and spring 2004 surveys. All the on-effort sightings are shown, although not all were used to estimate abundance. Solid lines indicate the 100 m (328 ft) and 1,000 m (3,281 ft) isobaths and the offshore extent of the U.S. EEZ (Waring et al. 2011 for these and subsequent figures of cetacean distribution in the northern Gulf of Mexico).
13.3.2.2 Sperm Whale
The sperm whale (Physeter macrocephalus), medium gray above and light gray to white below, is the largest toothed whale and, indeed, the largest toothed creature on Earth. It is highly sexually dimorphic. Males average 15 m (49 ft) in length and a prodigious 36,000 kg or 36 metric tons (79,350 lb), and females average 11 m (36 ft) and 20,000 kg or 20 metric tons (44,100 lb). The maximum size of male sperm whales is around 20 m (66 ft), although due to last mid-century’s intensive worldwide whaling, there are probably few of these giants around at present. The heads of the male sperm whales grow disproportionately rapidly as they age. The male’s head takes up about one-fifth to one-quarter of the body’s length in young ones and up to one-third of the body’s length in older males. It is obviously a secondary sexual characteristic, and males use the head for intrasex fighting and probably acoustic displays. The terrestrial analog might be deer stags with their antlers (the sperm whale male head) and roars (special male-only loud sounds that sperm whales emit). The head houses a giant structure of waxy oil—the spermaceti organ. The blowhole, placed differently from that of any other cetacean, is at the upper front of the mighty head, not along the mid-line but somewhat to the left, which results in a very distinctive, forward-tilted exhalation blow to the animal. Teeth are displayed in the lower jaw only and fit neatly into corresponding sockets in the upper jaw. The back has a dorsal ridge but no dorsal fin.
Sperm whales have a matriarchal society. Females and their female young tend to stay in one or adjacent groups for many years or for life. Males leave the group as they become sexually mature, at about age 10. The matriarchy, which tends to stay in tropical and subtropical waters, allows for related animals to help each other (e.g., take turns patrolling for danger to their nondiving neonates at the surface while others dive to depth). Indeed, deep diving for food (squid and fishes) seems to have driven the evolution towards high sociality in this species. Young males tend to stay together in groups of a dozen or so animals and travel to somewhat higher latitudes than the matriarchies they have left. As they mature—males do not seem to reach social maturity for re-inserting themselves briefly into matriarchies to mate until about age 25—older males tend to be alone (probably to avoid or minimize competition for access to females), and they travel to high near-Arctic and Antarctic latitudes to feed in very deep, productive waters. The general pattern of matriarchy, maturing males, older lone males, etc., has a close analog in the matriarchal systems of the largest land mammals—African and Asian elephants (Loxodonta sp. and Elephas maximus, respectively)—in what has been termed the collosal convergence of social/sexual strategies coupled with gigantism, long lives, and extended caregiving to young (Weilgart et al. 1996; Whitehead 2003) (Figures 13.7 and 13.8).

Sperm whales (Physeter macrocephalus) are the largest toothed whale and, indeed, the largest toothed creature on Earth. They are highly social, and all but older males are found in tight societies. There is a resident population in the northern Gulf of Mexico (photo by Thomas A. Jefferson, with permission).

Satellite tracks of seven sperm whales (Physeter macrocephalus) tagged July 3, 2002, and tracked for as long as early June, 2003 (two whales) (for details see Ortega-Ortiz et al. 2012 from which this figure was taken).

Distribution of sperm whale (Physeter macrocephalus) sightings from SEFSC spring vessel surveys during 1996–2001 and from summer 2003 and spring 2004 surveys. All on-effort sightings are shown, though not all were used to estimate abundance. Solid lines indicate the 100 m (328 ft) and 1,000 m (3,281 ft) isobaths, and the southern line represents the U.S. EEZ (from Waring et al. 2011).
Sperm whales occur throughout the world’s oceans but generally in waters deeper than about 500 m (1,640 ft) because of their habit of seeking largely deep-diving squid and fishes. Sperm whales in the Gulf are on average 1.5–2.0 m (4.9–6.6 ft) smaller than those found elsewhere (Richter et al. 2008; Jaquet and Gendron 2009). This size difference was noted by whalers 150 years ago (Reeves et al. 2011) and strongly suggests a different population from the sperm whales of the North Atlantic, a verification of which was provided by Engelhaupt et al. (2009) from genetic analysis. Mitochondrial DNA (inherited only from the mother) shows significant differences between Gulf sperm whales and sperm whales in other parts of the North Atlantic, while nuclear (bi-parentally inherited) DNA shows no difference. This indicates that females stay within the Gulf but that at least some males travel and breed in both the Gulf and North Atlantic. Indeed, recent satellite tracking of sperm whales showed that matriarchies stayed in waters about 200–3,499 m (656–11,480 ft) deep, generally in the area south and southwest of the Mississippi/Atchafalaya mouths, while males traveled south to Mexico’s Campeche area, and one male left the Gulf but returned after about 2 months (Ortega-Ortiz et al. 2012).
Typical group size of Gulf sperm whales in the north, which is almost always of presumed matriarchies, is 8–11 animals (Richter et al. 2008), often with calves less than 3–5 years old. This is smaller than groups (24–31) in the Pacific (Coakes and Whitehead 2004), but similar to groups (about six) in the adjacent Caribbean (Gero 2005). Statistical lagged association rates (Whitehead 2009) indicate that Gulf sperm whale groups are stable for longer (about 62 days) than in the Pacific (7–19 days) (Coakes and Whitehead 2004) but similar (about 80 days) to another enclosed body of water, the Gulf of California (Jaquet and Gendron 2009). It is possible that group sizes and association rates are ecologically related and that food or other ocean-basin physical/biological variables help to define social patterns (Richter et al. 2008).
Only recently have more accurate estimates of sperm whale numbers in the northern Gulf emerged. The latest estimate is about 1,665 (CV 0.20) animals (Table 13.5) (Mullin and Fulling 2004). Sperm whales overlap strongly with shipping lanes between New Orleans and Houston, industrial seismic activities, and deepwater oil/gas rigs (Azzara 2012). They were the only large whales to be hunted in the Gulf (although apparently not into the twentieth century), and their population characteristics may still be influenced by this earlier depredation (Reeves et al. 2011). There is not enough precision to estimate population trends and current productivity rates.
Sperm whales also occur in the southern Gulf and were hunted there in the past (Reeves et al. 2011). Most sperm whales encountered during cruises in the south appear to be concentrated on the continental slope (Figure 13.10).

Records of sperm whales (Physeter macrocephalus) in the Mexican waters of the southern Gulf of Mexico and Caribbean. Display for distribution is as follows: solid symbol, strandings; hollow symbols, confirmed sightings; triangles, spring (Mar–May); squares, summer (Jun–Aug); circles, autumn (Sep–Nov); diamonds, winter (Dec–Feb); crosses, unknown dates. Thin contour lines show the 200 m, 1,000 m, 2,000 m, and 3,000 m (656 ft, 3,281 ft, 6,562 ft, and 9,843 ft) isobaths (from Ortega-Ortiz 2002).
13.3.2.3 Pygmy and Dwarf Sperm Whales
The pygmy and dwarf sperm whales (Kogia breviceps and Kogia sima, respectively) of the family Kogiidae (which are much smaller than the sperm whale but most closely related to it) are not found together. However, they will be treated together here as they appear to have quite similar habitats and habits and are often not identified to species during surveys. Like the sperm whale, pygmy and dwarf sperm whales have a spermaceti organ of waxy oil in their heads and teeth only in the lower jaw. Again like sperm whales, the kogiids have a blunt and squarish head and an underslung lower jaw, but this is thicker than and not as long as that of the sperm whale. However, unlike the sperm whale, their blowhole is in the center top of the head, like that of dolphins, and they have a dorsal fin, also like dolphins. Both species are a steel blue/gray above and lighter below. Both species have a light colored, false gill mark just behind the eye (Figures 13.11 and 13.12).

Pygmy sperm whale (Kogia breviceps). They are shy and difficult to photograph, and not many good photos exist (photo by Robert L. Pitman, with permission).

Dwarf sperm whales (Kogia sima), probably adult and young. This adult’s dorsal fin is deformed. Normally the dorsal fin is not so strongly curved (photo by Robin W. Baird, with permission).
The pygmy sperm whale is the larger of the two at about 2.7–3.7 m (8.8–12.1 ft) and 317–410 kg (699–904 lb). The dwarf sperm whale is the size of smaller delphinids at about 2.1–2.7 m (6.9–8.9 ft) and 136–212 kg (300–467 lb). The dorsal fin of the dwarf sperm whale is larger, relative to body size, than that of the pygmy one and is set just a bit further forward on the body. Both species leap or are active at the surface only rarely, thus surfacing quite low and cryptically. It is difficult to distinguish the two species from the vantage point of a ship or an airplane except at close range and by the most expert of observers. As a result, most observations of individuals have been lumped as Kogiids. Both species dive to at least several 100 m and feed largely on squid.
Kogiids are likely much more numerous than present estimates suggest and occur in most oceans, generally in warmer waters. The slightly larger pygmy sperm whale moves to slightly higher latitudes, up to about Nova Scotia, Canada, in the western North Atlantic, as compared to about Virginia for the dwarf sperm whale. Worldwide, there are many strandings throughout their known ranges, again suggesting that the animals are more abundant than sighting records indicate, probably due to the difficulties in seeing them. Between 2003 and 2007, there were six pygmy sperm whale strandings on Florida beaches and four on Texas beaches. During the same period, seven dwarf sperm whales were stranded on Florida beaches and four on Texas beaches (with none reported for the other U.S. Gulf States). In the northern Gulf, the best estimate for both species combined is 453 (CV 0.35) (Figure 13.13 and Table 13.6) (Mullin 2007).

Distribution of dwarf (Kogia sima) and pygmy sperm whale (Kogia breviceps) sightings from SEFSC spring vessel surveys during 1996–2001 and from summer 2003 and spring 2004 surveys. All the on-effort sightings are shown, though not all were used to estimate abundance. Solid lines indicate the 100 m (328 ft) and 1,000 m (3,281 ft) isobaths and the offshore extent of the U.S. EEZ (from Waring et al. 2011).
13.3.2.4 Beaked Whales
Beaked whales consist of 22 species worldwide. They are almost invariably in deep waters and feed on deepwater squid. Only recently has knowledge been gained about some of these species, with northern bottlenose whales (Hyperoodon ampullatus) studied in The Gully off Nova Scotia (e.g., Gowans et al. 2001) and tagging of Cuvier’s beaked whales with short-term data tags that give details of depths, three-dimensional dive pattern, speeds (including accelerations and decelerations), and simultaneous recordings of their own click vocalizations and those of their conspecifics (Zimmer et al. 2005). This is exciting science; beaked whales are no longer ecologically unknown. The Gulf of Mexico has no known beaked whale hot spots, but detailed work has not been carried out there.
Beaked whales are often lumped during surveys in the Gulf, but there are enough data on Cuvier’s, Blainville’s, and Gervais’ beaked whale (most beaked whales of the Ziphiidae family have traditionally been named after the person who first described them) to make some overall statements. They all occur in the open ocean, in the tropic to temperate zones, and generally in small groups, and they feed on deepwater fishes and squid. Males have only one pair of erupted lower jaw teeth, and females have none. They therefore appear to be suction feeders that can inhale their prey without needing teeth to bite or pierce.
Cuvier’s beaked whales reach a size of 5.8 m (19 ft) for females and 5.5 m (18 ft) for males (females are larger than males in this species), and coloration can be a dark brown to slate gray. Cuvier’s beaked whale heads can be quite light in color, and the erupted tooth pair of males is set far to the front of the jaw. Blainville’s beaked whale males have their erupted jaw teeth on the midpoint of the jaw at a prominent upward jutting part of the lower jaw making them easy to distinguish in the field. Both males and females reach a size of about 4.7 m (15.4 ft). They occur in all tropical and temperate oceans, but apparently not in large numbers anywhere that has yet been discovered. Finally, Gervais’ beaked whale, at about 4.2–5.7 m (13.8–18.7 ft) for females and about 4.2–4.6 m (13.8–15.1 ft) for the smaller males, is endemic to the tropical- and cool-temperate waters of the Atlantic (i.e., as far north as western Scotland in the East Atlantic). In the northern tropical Gulf, unidentified beaked whales (which could also be of other species not mentioned here) are estimated at 337 (CV 0.40), Cuvier’s at 654 (CV 0.67), Blainville’s and Gervais’ beaked whale estimates are combined as Mesoplodon sp., and the combined best estimate is 57 (CV 1.40). Of course, it is also possible that some of these latter two species are represented in the “unidentified” category of 337 animals mentioned above. Figure 13.20 is a beaked whale in general sighting map, and Tables 13.7 and 13.8 summarize Cuvier’s beaked whale and combined Blainville’s/Gervais’ beaked whales estimates, respectively (Figures 13.14, 13.15, 13.16, 13.17, 13.18 and 13.19).

Cuvier’s beaked whale (Ziphius cavirostris), with likely conspecific tooth rake markings (photo by Thomas A. Jefferson, with permission).

Cuvier’s beaked whale (Ziphius cavirostris), with likely conspecific tooth rake markings. The small front teeth mark this individual as a male (photo by Charlotte Dunn, with permission).

Blainville’s beaked whale (Mesoplodon densirostris), adult female (photo by Robin W. Baird, with permission).

Blainville’s beaked whale (Mesoplodon densirostris), adult male. The mid-jaw erupted teeth are crowned by barnacles (photo by Robin W. Baird, with permission).

Blainville’s beaked whale (Mesoplodon densirostris), adult female and a large calf or subadult (photo by Robin W. Baird, with permission).

Gervais’ beaked whale (Mesoplodon europaeus), probable mother and young (photo by Charlotte Dunn, with permission).

Distribution of beaked whales. Sightings from SEFSC shipboard spring vessel surveys during 1996–2001 and from summer 2003 and spring 2004 surveys. All the on-effort sightings are shown, though not all were used to estimate abundance. Solid lines indicate the 100 m (328 ft) and 1,000 m (3,281 ft) isobaths and the offshore extent of the U.S. EEZ (from Waring et al. 2011).
13.3.2.5 Killer Whale
The cosmopolitan killer whales occur in all oceans from the tropics to the Arctic and Antarctic ice, although what was for many years thought to be just one species may be classified as several species in the future. They are the largest of the delphinids and have high sexual dimorphism, with adult males reaching about 9.8 m (32.2 ft) and females about 8.5 m (27.9 ft). The distinctive male dorsal fin grows throughout life, and it becomes a high erect, pointed structure in a fully mature male. Killer whales are strikingly colored, black above and white below, with a white oblong eye spot just above and behind the eye, a variably shaped saddle blaze behind and below the dorsal fin, and white undersides of the flukes, at times extending to a part or all of the fluke dorsum. Within a population, individuals have slightly different white marks and blaze patterns, and populations can generally be distinguished by common coloration factors within them as well. Individuals can be recognized by the distinctive natural marks, along with the pattern of scars and nicks, particularly on the dorsal fin (Figure 13.21).

Killer whales (Orcinus orca) are the largest of the delphinids and certainly one of the most charismatic for humans (photo by Thomas A. Jefferson, used with permission).
Killer whale social structure is varied and complex. In the North Pacific along the shores of North America, for example, there are nearshore forms (termed residents) that travel little and eat salmon almost exclusively, other nearshore forms (termed transients) that travel over several 100 km and feed almost exclusively on marine mammals, and offshore forms that feed largely on fish but take other prey as well; the latter can move over 1,000 km (621 mi) in short time periods (days to weeks). All forms appear to be matriarchal, with youngsters staying within the mothers’ group or pod for long periods or for life, but details of this vary by social grouping (Bigg et al. 1990). For example, one salmon-eating pod off Vancouver Island, British Columbia, Canada, is socially closed, with both female and male young staying with their mothers (but mating with others during occasional superpod congregations, which probably serve to avoid inbreeding). Such closed intergenerational social living allows for a complicated culture to be developed, due to transmission of foraging, vocal, and other information through generations (Whitehead 1998). For example, some societies of killer whales have individuals beaching themselves to take pinnipeds on land or in the turbulent surf zone; they cooperate in these hunts and even appear to teach youngsters the tricky business of temporarily stranding without becoming stranded forever (Guinet and Bouvier 1995).
Killer whales of the Gulf are only sporadically sighted (see Figure 13.22), and limited photo-identifications (presently 32 individuals) indicate that they travel for at least up to 1,100 km (684 mi) (O’Sullivan and Mullin 1997). It is presently unknown whether killer whales of the Gulf form a stock or population separate from those in the adjacent North Atlantic, and photo-identification comparisons and genetic data are needed. Presently, the best estimate for the northern Gulf is about 49 (CV 0.77) animals, but an earlier estimate of 133 (CV 0.49), based on data from 1996 to 2001 (Mullin and Fulling 2004), may be more appropriate, given the identification record of 32 from limited work. There are persistent reports from sport fishers that killer whales feed on tuna in the Gulf, but these have not been verified. On May 17, 2008, a killer whale became entangled in a fishing longline (Garrison et al. 2009), which suggests that killer whales take fish off longlines in the Gulf at times, as they are known to do in some other areas (Table 13.9).

Distribution of killer whale (Orcinus orca) sightings from SEFSC spring vessel surveys during 1996–2001 and from summer 2003 and spring 2004. All the on-effort sightings are shown, though not all were used to estimate abundance. Solid lines indicate the 100 m (328 ft) and 1,000 m (3,281 ft) isobaths and the offshore extent of the U.S. EEZ (from Waring et al. 2011).
13.3.2.6 Short-Finned Pilot Whales
There are two species of pilot whales. The long-finned pilot whale (Globicephala melas) occurs in the North Atlantic (including the western part of the Mediterranean Sea) and in the southern hemisphere but not in the North Pacific. It is a relatively cold-water species and does not frequent waters south of the U.S. state of Georgia, and thus does not make it (at least not regularly) into the Gulf of Mexico. The short-finned pilot whale (Globicephala macrorhynchus), on the other hand, occurs worldwide in the tropics and subtropics and overlaps with its congener in fringe habitats of both, including off the U.S. eastern seaboard. It can occur in groups as small as one dozen or so animals but also occurs in schools of hundreds, and before major hunting in most of its range, it even occurred in groups over a thousand. Pilot whales tend to feed on squid, but fish are also taken. Short-finned pilot whales in the Gulf at times harass groups of sperm whale matriarchies with young in them. But whether this is an attempt to feed on sperm whale newborns or perhaps to get sperm whales to regurgitate food, as is believed to have been seen, is not known (Weller et al. 1996) (Figure 13.23).

Short-finned pilot whales (Globicephala macrorhynchus) occur in apparent matriarchal long-term tight societal bonds (photo by Thomas A. Jefferson, with permission).
Female short-finned pilot whales become sexually mature at about age nine and are about 5.5 m (18.0 ft) in length. Male short-finned pilot whales become sexually mature at about age 15 and are about 6.0 m (19.9 ft) in length, with the male dorsal fin growing disproportionately larger and more strongly curved. The male head (or melon) becomes more bulbous and squarish than the female head as seen from the side and in older males, may even overhang the front of the jaw under the melon. Pilot whales are quite dark to black above with a light belly patch of variable shape below. They have a light chin, a grayish to white stripe or chevron dorsally behind the eye and pointing towards the dorsal fin (useful for distinguishing the species from the air), and a variably light saddle patch pattern on both sides and just behind the dorsal fin.
Data from drive fisheries off Japan indicate that social maturity of males may take considerably longer to achieve than does sexual maturity, as younger males may not be able to mate effectively with females in a generally tight matriarchal society. Generally, pilot whale females nurse their young for at least 3 years, and some evidence points to them nursing into their offsprings’ teens! Females often continue to nurse their offspring even when they are no longer reproductively active (beyond about age 40). This indicates that nursing is important beyond meeting nutritional needs; it is a part of social bonding and may also extend to alo-nursing (i.e., nursing offspring that are not their own) (Kasuya and Marsh 1984). These data come from dead animals, and unfortunately long-term studies of living populations are scarce. Heimlich-Boran (1993) studied the species off Tenerife and reported that pilot whales there live in matriarchal societies that include adult males (presumably the offspring of mothers in the society) that mate outside of their immediate groupings. This system may therefore be quite similar to that of resident killer whales.
Short-finned pilot whales tend to occur in deep waters, as they feed on mesopelagic fishes and squid, but are more common over continental slopes than over the abyssal plain, and this is true for the Gulf as well (see Figure 13.24). Although there is no good, overall, worldwide population estimate, short-finned pilot whales of the eastern tropical Pacific are estimated at about 590,000 (CV 0.26) (Gerrodette and Forcada 2002) and in the northern Gulf at 716 (CV 0.34) (Table 13.10). This recent lower best estimate may be too low. Table 13.10 shows that a previous estimate was more than 2,000 animals in the same area. Short-finned pilot whales often mass strand, but only two strandings have been reported in the Gulf since the 1990s: one in 1999 (two animals) and one in 2001 (nine animals), and both strandings were off Florida; these numbers are much lower than the mass strandings that occur in many other places. Short-finned pilot whales also occur in Mexican waters and have been sighted in waters up to about 3,000 m (9,843 ft) deep (Figure 13.25).

Distribution of short-finned pilot whale (Globicephala macrorhynchus) sightings from SEFSC spring vessel surveys during 1996–2001 and from summer 2003 and spring 2004 surveys. All the on-effort sightings are shown, though not all were used to estimate abundance. Solid lines indicate the 100 m (328 ft) and 1,000 m (3,281 ft) isobaths and the offshore extent of the U.S. EEZ (from Waring et al. 2011).

Records of short-finned pilot whales (Globicephala macrorhynchus) in the Mexican Waters of the Southern Gulf of Mexico and Caribbean. Display for distribution is as follows: solid symbol, strandings; hollow symbols, confirmed sightings; triangles, spring (Mar–May); squares, summer (Jun–Aug); circles, autumn (Sep–Nov); diamonds, winter (Dec–Feb); crosses, unknown dates. Thin contour lines show the 200 m, 1,000 m, 2,000 m, and 3,000 m (656 ft, 3,281 ft, 6,562 ft, and 9,843 ft) isobaths (from Ortega-Ortiz 2002).
13.3.2.7 False Killer Whale
Along with pilot, pygmy killer (Feresa attenuata), and melon-headed (Peponocephala electra) whales, the false killer whale (Pseudorca crassidens) species was termed a blackfish by whalers and fishermen, because of its generally very dark coloration. All of these oceanic blackfish are known for their frequent mass strandings, probably because they have tight long-term (generally matriarchal) societies, and when several animals make a navigational mistake near a shoal or headland, the integrity of the group has all others following. The false killer whale is about the same size as pilot whales, with males slightly larger than females, but its body form is considerably more slender than that of pilot whales. Group size can be just a few animals or into the hundreds. As with pilot whales, males and females travel together in apparently tight bonds. Unlike pilot whales, however, details of genetic relationships, length of maternal care, and other life history and behavioral characteristics are not yet known.
False killer whales feed on squid and fishes and also at times attack sperm whales and humpback whales. In the latter cases, it is presumed that they are attempting to isolate more vulnerable animals (i.e., old, infirm, or newborn animals) from the more robust animals, but details are unknown (Figure 13.26).

False killer whales (Pseudorca crassidens) occur in all tropical oceans (photo by Robin W. Baird, with permission).
False killer whales occur in tropical and warm temperate oceans and are usually found in deep water (but not in or near the center of oceans). They may be quite close to shore where deep waters occur close to oceanic islands and atolls, such as the Hawaiian Islands. They may occur in cooler temperate waters into 50°N latitude, as well as south of the equator, probably due to their large body size (they are the third largest delphinid cetacean, after killer and long-finned pilot whales). Worldwide numbers are not available. One older estimate of about 40,000 (CV 0.64) (Wade and Gerrodette 1993) has been made for the eastern tropical Pacific. In the northern oceanic Gulf, where false killer whales occur in deep waters and not normally on the slope, the estimate is 777 (CV 0.56) (Figure 13.27 and Table 13.11).

Distribution of false killer whale (Pseudorca crassidens) sightings from SEFSC spring vessel surveys during 1996–2001 and from summer 2003 and spring 2004 surveys. All the on-effort sightings are shown, though not all were used to estimate abundance. Solid lines indicate the 100 m (328 ft) and 1,000 m (3,281 ft) isobaths and the offshore extent of the U.S. EEZ (from Waring et al. 2011).
13.3.2.8 Pygmy Killer Whale
The pygmy killer whale (Feresa attenuata) is slender like the false killer whale but substantially smaller, with males around 2.3 m (7.5 ft) and females 2.1 m (6.9 ft), or about dolphin size. Without adequate size reference, this species can easily be confused with false killer whales and (see below) melon-headed whales. However, Pygmy killer whales have a white patch (or goatee) at the front of their lower chin, which is more pronounced than the goatee on the melon-headed whale.
Pygmy killer whales feed largely on fishes, but squid are also taken. They can be quite aggressive, and attacks on smaller as well as similar-sized delphinids, such as spotted and spinner dolphins, have been witnessed in and near tuna nets in the eastern tropical Pacific.
Pygmy killer whales occur in tropical waters worldwide in groups of about 12–50 animals, although somewhat larger groups also occur. In the eastern tropical Pacific, the population estimate is similar to that of false killer whales, slightly less than 40,000 (CV 0.64) (Wade and Gerrodette 1993). In the oceanic northern Gulf, best estimate is 323 (CV 0.60), with sightings both on the slope and in abyssal plain waters (Figures 13.28 and 13.29 and Table 13.12).

Pygmy killer whales (Feresa attenuata) are very small members of the blackfish group of small cetaceans (photo by Robert L. Pitman, with permission).

Distribution of pygmy killer whale (Feresa attenuata) sightings from SEFSC spring vessel surveys during 1996–2001 and from summer 2003 and spring 2004 surveys. All the on-effort sightings are shown, though not all were used to estimate abundance. Solid lines indicate the 100 m (328 ft) and 1,000 m (3,281 ft) isobaths and the offshore extent of the U.S. EEZ (from Waring et al. 2011).
13.3.2.9 Melon-Headed Whale
The final blackfish—the melon-headed whale—is a bit larger than the diminutive pygmy killer whale, at about 2.7 m (8.9 ft) for males and 2.6 m (8.5 ft) for females. As mentioned above, chin coloration is not quite as white as that of the pygmy killer whale, but it too can have white lips. Both species (as well as the larger false killer whale) have a rounded head that is more pointed than the blunt rounded heads of pilot whales and killer whales. Melon-headed whales feed on fishes and squid (Figure 13.30).

Melon-headed whales (Peponocephala electra) are about half way in size between pygmy and false killer whales (photo by Robin W. Baird, with permission).
Melon-headed whales occur throughout warm waters of the tropics and near-tropics (to about 40°N latitude and 30°S latitude), and their estimated numbers of about 45,000 (CV 0.47) are similar to those of pygmy and dwarf killer whales in the eastern tropical Pacific (Wade and Gerrodette 1993). They can occur in much larger schools (100–1,500 animals) than the false and pygmy killer whales (but not this large in the Gulf), and they are often found in multispecies aggregations with Fraser’s (Lagenodelphis hosei) and spinner dolphins. In the Gulf, an estimated 2,283 (CV 0.76) melon-headed whales can occur in the northern oceanic area, but apparently they are more often in the western part rather than the eastern part of the Gulf (Figure 13.31 and Table 13.13).

Distribution of melon-headed whale (Peponocephala electra) sightings from SEFSC spring vessel surveys during 1996–2001 and from summer 2003 and spring 2004 surveys. All the on-effort sightings are shown, though not all were used to estimate abundance. Solid lines indicate the 100 m (328 ft) and 1,000 m (3,281 ft) isobaths and the offshore extent of the U.S. EEZ (from Waring et al. 2011).
13.3.2.10 Rough-Toothed Dolphin
The rough-toothed dolphin (Steno bredanensis) is a small delphinid with a beak that tapers from the head, but is not as sharply demarked as it is in the abrupt beak of the bottlenose dolphins. This taper also gives the rough-toothed dolphin (which, indeed, has fine lateral ridges on its teeth, giving them a rough feeling) the nickname lizard dolphin. Males are about 2.7 m (8.9 ft) at maturity and females 2.3 m (7.5 ft). They are dark above and lightish below, but often with a bluish/purplish tinge of coloration, and with yellowish/white dots along the sides.
Rough-toothed dolphins do not appear to be very deep divers, preferring to feed on fishes, squid, octopuses, and often even large fishes such as mahi-mahi (Coryphaena hippurus) that are found in deep waters but within 100 m (328 ft) or so of the ocean’s surface. Off Hawaii, groups of rough-toothed dolphins have gotten into the habit of taking large fishes off long-lines set by humans, and this has put them at odds with the local fishing industry.
Rough-toothed dolphins occur in tropical waters worldwide. They can be confused with bottlenose dolphins from a distance, due to similar size and general morphology, but the beak and spots should distinguish them upon closer inspection. They tend to occur in groups of ten or so animals, but larger groups of more than 100 have been seen (Figure 13.32).

Rough-toothed dolphins (Steno bredanensis) generally occur in small groups. They superficially resemble bottlenose dolphins in shape and size, but their rostrum is more curved from tip to rise of head, while that of the bottlenose dolphin is very abrupt (photo by Thomas A. Jefferson, with permission).
We know practically nothing about the social order of rough-toothed dolphins. We know that they have a very large brain-to-body ratio, and the few that have been kept in captivity have been noted to be extremely flexible behaviorally (i.e., intelligent), with evidence for sophisticated second order learning (also called deutero-learning), which implies thought (Pryor et al. 1969). Recent work (Kuczaj and Yeater 2007) indicates that they have tight social bonds with long-term relationships.
No reliable estimates of numbers of the species worldwide are available. However, there are about 146,000 (CV 0.32) estimated for the eastern tropical Pacific (Wade and Gerrodette 1993) and 2,653 (CV 0.42) for the northern oceanic Gulf (Figure 13.33 and Table 13.14).

Distribution of rough-toothed dolphin (Steno bredanensis) sightings from SEFSC spring and fall vessel surveys during 1996–2001 and from summer 2003 and spring 2004 surveys. All on-effort sightings are shown, although not all were used to estimate abundance. Solid lines indicate the 100 m (328 ft) and 1,000 m (3,281 ft) isobaths and the offshore extent of the U.S. EEZ (from Waring et al. 2011).
13.3.2.11 Risso’s Dolphin
Risso’s dolphin (Grampus griseus) is often called grampus by fishermen, and whaling records indicate that it was hunted for oil and meat in the Gulf in the 1700s and 1800s (Reeves et al. 2011). Males and females are about the same size, at a bit over 3 m (10 ft) in length, with no hint of sexual dimorphism. They have a prominent dorsal fin and, consequently, at a distance are sometimes confused with killer whales. However, Risso’s dolphins are quite differently colored. While young, they are all gray. As they age they receive more linear scars on their bodies, until older individuals are almost entirely white. Apparently, all scrapes of their skins—presumably usually caused by intraspecific interactions of tooth rakes—disrupt dermal melanin pigments, which do not regrow or reinvade damaged skin in this species (Figure 13.34).

A Risso’s dolphin (Grampus griseus) showing evidence of loss of melanin pigmentation and conspecific rake marks that stay for life (photo by Thomas A. Jefferson, with permission).
Risso’s dolphins occur in tropical to cool temperate waters worldwide, and in the North Atlantic they are found as far north as Newfoundland. They are quite cold-water adapted (down to about 10 °C [50 °F]). Most occurrences are on the high seas in deep water, and numbers have generally been underestimated, it is now believed, due to the difficulty of surveying the open ocean habitat. Risso’s dolphins have been described as very common cetaceans in the southern California Channel Island area, where groups can vary from 1 to approximately 100. Because they are so light colored as seen from above, individuals can be described from a circling airplane even when the animals have dived to twice their own lengths, making their social study a recently recognized plausibility. They feed on squid, but fishes and crustaceans are taken as well.
Risso’s dolphins occur quite close to shore off California, Oregon, and Washington and have been designated as a subpopulation there, estimated at 16,000 (CV 0.28) (Barlow 2003). Before the intensive 1983–1984 El Niño event of the eastern Pacific, Risso’s dolphins were uncommon off southern California. During the event, however, pilot whales all but disappeared (presumably because of a lack of squid), and Risso’s dolphins came into the area. They are still present there in rather large numbers (Shane 1994). In the northern oceanic Gulf, they occur in some abundance both off the slope and abyssal plain, with the best estimate currently at 1,589 (CV 0.27) (Figure 13.35 and Table 13.15).

Distribution of Risso’s dolphin (Grampus griseus) sightings from SEFSC vessel surveys during 1996–2001 and from summer 2003 and spring 2004 surveys. All the on-effort sightings are shown, though not all were used to estimate abundance. Solid lines indicate the 100 m (328 ft) and 1,000 m (3,281 ft) isobaths and the offshore extent of the U.S. EEZ (from Waring et al. 2011).
13.3.2.12 Fraser’s Dolphin
Fraser’s dolphin (Lagenodelphis hosei) is a physically robust species, with a short beak and small flippers, flukes, and dorsal fin. As adults, Fraser’s dolphins have a dark tie-width stripe running along the side, demarcating the darker dorsum from the lighter ventrum. This variably appears to be much stronger in Pacific than in Atlantic animals, and it is stronger in adult males than in adult females. It is not present at all in calves and other immature animals. Males are about 2.7 m (8.9 ft) long, and females are slightly shorter, but there is no pronounced sexual dimorphism.
Fraser’s dolphins feed largely on mid-water fishes and squid and may be able to dive as deep as about 600 m (1,969 ft). They are often active at the surface, frequently splashing and leaping in low arcs that create whitewater that can make their presence known from several kilometers. They occur worldwide in tropical and warm temperate waters, to about 30° north and south of the equator, but exact ranges and numbers are poorly known. There are an estimated 290,000 (CV 0.34) Fraser’s dolphins in the eastern tropical Pacific (Wade and Gerrodette 1993). In the northern oceanic Gulf, no best estimate is given by NOAA because no sightings were made during the most recent surveys conducted in 2003–2004, and it is reported that sometimes none are seen for several years. Nevertheless, the most recent estimate, made from sightings in 1996–2001, is 726 dolphins (CV 0.70) (Figures 13.36 and 13.37 and Table 13.16).

Blunt-beaked, small bodied with very small flippers and dorsal fins, Fraser’s dolphins (Lagenodelphis hosei) occur throughout warm waters in variable color morphs (photo by Thomas A. Jefferson, with permission).

Distribution of Fraser’s dolphin (Lagenodelphis hosei) sightings from SEFSC spring vessel surveys during 1996–2001 and from summer 2003 and spring 2004 surveys. All the on-effort sightings are shown, although not all were used to estimate abundance. Solid lines indicate the 100 m (328 ft) and 1,000 m (3,281 ft) isobaths and the offshore extent of the U.S. EEZ (from Waring et al. 2011).
13.3.2.13 Stenella Dolphins: Spinner, Clymene, Striped, Pantropical and Atlantic Spotted Dolphins
These dolphins of the Stenella genus are all thin-bodied and none are deep divers. They prefer to feed within the top several 100 m of the surface, although often on mesopelagic prey that comes towards the surface at night with the diurnally migrating prey of the deep scattering layer (DSL) in open oceans. While group sizes can vary greatly, stenellids are highly social animals and often occur in groups comprised of hundreds to several thousand animals. Stenellids also often occur in interspecies aggregations (e.g., spinner and pantropical spotted dolphins in the eastern tropical Pacific).
13.3.2.13.1 Spinner Dolphin
Spinner dolphins (Stenella longirostris) are the most numerous of the tropical oceanic cetaceans worldwide. They are thin, extremely long-beaked stenellids that occur as different morphologies (and as four different subspecies) in different parts of the tropics. Oceanic eastern tropical spinners show marked stripes along their flanks and reduced sexual dimorphism, and they live in huge herds of up to several thousand animals. The eastern spinner of the far eastern Pacific is almost uniformly dark gray and highly sexually dimorphic, with males having a pronounced post-anal keel and a high forward-curved dorsal fin; the eastern spinner morphology indicates a polygynous mating system, unlike the usual polygynandry (multimate or promiscuous) system of most delphinids, but detailed behavioral observations have not been carried out.
Some populations of spinner dolphins exist in the open ocean. Many others rely on daytime resting in or near island bays or in atolls and move offshore to feed at night on myctophid and squid prey of the DSL (Norris and Dohl 1980; Karczmarski et al. 2005). Despite considerable variation in size and morphology in different areas, overall spinner dolphin length is about 1.8 m (5.9 ft), making it a rather small dolphin. It is the only dolphin that spins around its axis extremely rapidly and with up to six revolutions, either in a horizontal or vertical position above the surface of the water. These spins appear to have to do with social facilitation as animals move from a resting to an alert (often highly social/sexual) state. Island spinner dolphins were studied intensively on the Kona Coast of the Big Island, Hawaii, in the late 1970s to early 1980s. Norris et al. (1994) is a detailed book of their behaviors and life-history strategies.
While there are no estimates of worldwide numbers, spinner dolphins of the eastern tropical Pacific are estimated at about 1.4 million animals for two subspecies (Gerrodette et al. 2005) and at 1,989 (CV 0.48) in the northern oceanic Gulf. However, note the large variations from other sets of surveys (Table 13.17). Almost all survey sightings of spinner dolphins in the Gulf of Mexico have been in the central and eastern Gulf, but not western Gulf, and largely, but not exclusively, in slope waters (Figures 13.38 and 13.39).

Spinner dolphins (Stenella longirostris) occur in warm waters worldwide and in quite a few different color and body morphs (photo by Thomas A. Jefferson, with permission).

Distribution of spinner dolphin sightings from SEFSC spring vessel surveys during 1996–2001 and from summer 2003 and spring 2004 surveys. All the on-effort sightings are shown, though not all were used to estimate abundance. Solid lines indicate the 100 m (328 ft) and 1,000 m (3,281 ft) isobaths and the offshore extent of the U.S. EEZ (from Waring et al. 2011).
13.3.2.13.2 Clymene Dolphin
The Clymene dolphin (Stenella clymene) is about the same size as the spinner dolphin and has been confused with it, and also the short-beaked common dolphin. It is a bit more robust animal than the spinner dolphin, however, and has a shorter beak, black lips and a pronounced black beak tip.
Not much is known about Clymene dolphins. Most of what is known was aggregated by Thomas Jefferson, a world authority on marine mammals and author of the guide The Marine Mammals of the World (Jefferson et al. 2008). Clymene dolphins feed largely on mesopelagic fishes and squid and take advantage of the ecological cascade of the DSL. Clymene dolphins often associate with spinner dolphins, and as the two are difficult to distinguish from a distance, this association further clouds counts of the species (Figure 13.40).

Clymene dolphins (Stenella clymene) have often been confused with spinner dolphins. They occur only in the tropical Atlantic (photo by Robert L. Pitman, with permission).
Clymene dolphins occur only in the tropical and warm temperate Atlantic from about New Jersey (in summer) down to Brazil in the west Atlantic. In the Gulf of Mexico, group sizes average 42 animals; however, group size is highly variable, and some groups contain several hundred individuals (Mullin et al. 1994). They are estimated at a minimum of about 100,000 in the Atlantic Basin, including an estimate of 6,575 (CV 0.36) in the northern oceanic Gulf; but note that previous estimates have been as high as 17,000 (Table 13.18). Unlike spinner dolphins that use mainly the eastern portion of the Gulf, Clymene dolphins largely use the abyssal part of the western section (Figure 13.41) with some overlap.

Distribution of Clymene dolphin (Stenella clymene) sightings from SEFSC shipboard spring surveys during 1996–2001 and from summer 2003 and spring 2004 surveys. All the on-effort sightings are shown, though not all were used to estimate abundance. Solid lines indicate the 100 m (328 ft) and 1,000 m (3,281 ft) isobaths and the offshore extent of the U.S. EEZ (from Waring et al. 2011).
13.3.2.13.3 Striped Dolphin
Striped dolphins (Stenella coeruleoalba) are about 2.4 m (7.9 ft) long, with little sexual dimorphism. They are a bit more robust in body form than spinner, pantropical and Atlantic spotted dolphins, but are nevertheless generally slender. They are strikingly marked with stripes along their sides, highlighted by a stripe that begins at the eye and swoops dorsally, ending just below the dorsal fin. Striped dolphins feed largely on mesopelagic fishes and squid.
This stenellid is distributed in tropical and warm temperate waters worldwide, between about 50°N latitude and 40°S latitude. In the North Atlantic, the species occurs as far north as Nova Scotia (in summer) and throughout the tropics to the southern hemisphere of Brazil and Africa. It occurs in the Mediterranean Sea and was the subject of ancient Greek frescoes. Group sizes vary from dozens of animals to hundreds and may have numbered in the thousands historically (Figure 13.42).

Striped dolphins (Stenella coeruleoalba) are animals of the open ocean, often found in the deepest waters, including in the Gulf of Mexico (photo by Thomas A. Jefferson, with permission).
In the eastern tropical Pacific, the most recent population estimate of striped dolphins, which was derived from results of a 2003 line-transect survey, was about 1.5 million animals (Gerrodette et al. 2005). Abundance estimates within about 500 km (310 mi) of the U.S. West Coast have averaged about 19,000 (CV 0.28) between 1991 and 2005.
While global estimates are questionable, there are surely several million worldwide. Striped dolphins in the northern oceanic Gulf are estimated at about 3,325 animals (CV 0.48), with some sightings on the eastern Gulf slope but most in deep ocean waters (Figure 13.43 and Table 13.19).

Distribution of striped dolphin (Stenella coeruleoalba) sightings from SEFSC spring vessel surveys during 1996–2001 and from summer 2003 and spring 2004 surveys. All the on-effort sightings are shown, although not all were used to estimate abundance. Solid lines indicate the 100 m (328 ft) and 1,000 m (3,281 ft) isobaths and the offshore extent of the U.S. EEZ (from Waring et al. 2011).
13.3.2.13.4 Pantropical Spotted Dolphin
The pantropical spotted dolphin (Stenella attenuata) is marked by a rather long beak, slender body, and quite strongly falcate dorsal fin. While a muted gray above and light below, it develops spots along the sides as it ages, and rough categories of age can be determined by the amount of spotting. There is much geographic variation in coloration and size by area. This dolphin occurs in all tropical and subtropical waters, worldwide, from the equator to about 40°N latitude and 30°S latitude. Sexually mature individuals are from about 1.7 m (5.6 ft) to 2.6 m (8.5 ft), with males only slightly larger than females.
Pantropical spotted dolphins occur in rather large numbers in deep waters of the world’s oceans, where they feed on mesopelagic and epipelagic fishes, crustaceans, and squid often related to the DSL, but they also feed on surface-dwelling flying fishes in some areas. An estimated 640,000 still exist in the eastern tropical Pacific, but this represents probably only about 20 % of the original population(s) before intensive killing as bycatch in the tuna fishing industry during the 1950s through early 1990s (related estimates also, Gerrodette et al. 2005). In the Gulf of Mexico, pantropical spotted dolphins are the most numerous cetacean, with estimates in the northern oceanic Gulf ranging from about 34,000 (CV 0.18) to 91,000 (CV 0.16) (Figures 13.44 and 13.45 and Table 13.20). They occur on the upper slope in waters of about 100 m (328 ft), as well as their primary habitat—waters deeper than 100 m (328 ft) and into the open abyssal zone of the Gulf, including in the southern Gulf (Figure 13.46).

Pantropical spotted dolphins (Stenella attenuata) are likely the most numerous of the genus Stenella in the world’s oceans and the most numerous marine mammal in the Gulf of Mexico as well (photo by Thomas A. Jefferson, with permission).

Distribution of pantropical spotted dolphin (Stenella attenuata) sightings from SEFSC spring vessel surveys during 1996–2001 and from summer 2003 and spring 2004 surveys. All the on-effort sightings are shown, though not all were used to estimate abundance. Solid lines indicate the 100 m (328 ft) and 1,000 m (3,281 ft) isobaths and the offshore extent of the U.S. EEZ (from Waring et al. 2011).

Records of pantropical spotted dolphins (Stenella attenuata) in the Mexican waters of the southern Gulf of Mexico and Caribbean. Display for distribution is as follows: solid symbol, strandings; hollow symbols, confirmed sightings; triangles, spring (Mar–May); squares, summer (Jun–Aug); circles, autumn (Sep–Nov); diamonds, winter (Dec–Feb); crosses, unknown dates. Thin contour lines show the 200 m, 1,000 m, 2,000 m, and 3,000 m (656 ft, 3,281 ft, 6,562 ft, and 9,843 ft) isobaths (from Ortega-Ortiz 2002).
13.3.2.13.5 Atlantic Spotted Dolphin
The Atlantic spotted dolphin and the Clymene dolphin are the only species of cetaceans found in the Gulf that are endemic to the Atlantic Ocean, with the Atlantic spotted dolphin occurring as far north as 50°N latitude (although more commonly only to about 40°N latitude) and about 25°S latitude. It occurs—and has been studied intensively (e.g., Herzing 1997)—on the shallows of the Bahama banks, where it socializes during daytime and (presumably) feeds on epipelagic and mesopelagic fishes and squid in the drop-off oceanic zones, feeding relative to DSL organisms in deeper waters, at night. In the Gulf, Atlantic spotted dolphins generally occur within the 200-m (656-ft) depth contour and are thus animals of the shallower waters of the oceanic and near-oceanic zones.
Atlantic spotted dolphins are often confused with the pantropical species, as they also develop spots along their sides (and the rest of the body, depending on geographic area/population) as they mature. Male Atlantic spotted dolphins at full maturity are about 2.7 m (8.9 ft) in length, and females are about 2.5 m (8.2 ft). In the Atlantic Ocean, best estimate is unknown, but in the northern Gulf of Mexico, there are an estimated 37,611 (CV 0.28) animals; this estimate is not accepted as being current by the NMFS, because it is from data greater than 8 years old (Figure 13.47 and Table 13.21). Almost all of these sightings are from within the 100-m (328-ft) depth contours, especially off the Florida shelf, while a few range into the 100-m to 200-m (328-ft to 656-ft) depth area. Atlantic spotted dolphins in the Gulf do not seem to be found in deeper oceanic waters in the northern (Figure 13.48) or southern (Figure 13.49) Gulf. They co-occur in habitat with continental-slope bottlenose dolphins.

This underwater photo is of Atlantic spotted dolphins (Stenella frontalis) in the Bahamas, but they look very similar in the Gulf, developing spots as they age (photo by Bernd Würsig).

Distribution of Atlantic spotted dolphin (Stenella frontalis) sightings from SEFSC spring and fall vessel surveys during 1996–2001 and from summer 2003 and spring 2004 surveys. All the on-effort sightings are shown, though not all were used to estimate abundance. Solid lines indicate the 100 m (328 ft) and 1,000 m (3,281 ft) isobaths and the offshore extent of the U.S. EEZ (from Waring et al. 2011).

Records of Atlantic spotted dolphins (Stenella frontalis) in the Mexican waters of the southern Gulf of Mexico and Caribbean. Display for distribution is as follows: solid symbol, strandings; hollow symbols, confirmed sightings; triangles, spring (Mar–May); squares, summer (Jun–Aug); circles, autumn (Sep–Nov); diamonds, winter (Dec–Feb); crosses, unknown dates. Thin contour lines show the 200 m, 1,000 m, 2,000 m, and 3,000 m (656 ft, 3,281 ft, 6,561 ft, and 9,843 ft) isobaths (from Ortega-Ortiz 2002).
13.3.2.13.6 Common Bottlenose Dolphin
As the name implies, the common bottlenose dolphin (Tursiops truncatus) is a very common animal near shore in most tropical, temperate, and even cooler waters of all oceans, occurring as far north as northern Scotland in the Atlantic, as far south as mid-Patagonia in South America, and also as far south as the cold water (in winter at times slightly iced over) fjords of the South Island of New Zealand. It also occurs as separate populations in inshore bays and estuaries, alongshore barrier islands and other geographic situations, and in oceanic waters, often in quite disparate morphs of coloration and size. It is ubiquitous in nearshore areas of the Gulf of Mexico, both northern and southern, and along the continental shelf, to and beyond 200-m (656-ft) depths.
The bottlenose dolphin has been subdivided into over one dozen species and/or subspecies during its taxonomic history. Presently, its congener, the Indo-Pacific bottlenose dolphin (Tursiops aduncus), is accepted as the only other bottlenose dolphin. This designation will likely be refined with further genetic analyses. The Indo-Pacific bottlenose dolphin occurs from Cape Agulhas, South Africa, to the main island of Japan (i.e., in the Indian Ocean and the western Pacific, including most of Australia). This distribution overlaps strongly with that of the common bottlenose dolphin, and there is some confusion on species designations in certain overlap areas (Figure 13.50).

The ubiquitous worldwide dolphin that is most often envisioned by the nonmarine mammalogist when dolphins are mentioned. It is common in all waters except over the abyssal plain in the Gulf of Mexico (photo by Giovanni Bearzi, with permission).
In the Atlantic, there is no argument as to species, as the Indo-Pacific bottlenose dolphin does not reach there. But there is considerable debate on population and subpopulation designations. Bottlenose dolphins typically occur in groups of about one dozen animals, as mixed age and sex groups, all-female and youngster nursery groups, and those of immature or mature males (Wells et al. 1987). However, groupings as large as 1,000 animals in the open ocean (but not in the Gulf) have been reported.
Bottlenose dolphins (Tursiops truncatus) occur throughout most bays, sounds, and estuaries of the Gulf of Mexico, often into quite brackish water, with salinities of less than ten parts per thousand (ppt). However, the definitions of populations or stocks is complicated by the fact that bay animals also travel outside of their major habitats and interact and mate with animals outside of these bays as well. Most populations or subpopulations have not been well studied, with the most notable exceptions being those of the Sarasota-Bradenton, Florida, area (Wells 2003) and Sanibel Island (Shane 2004) of west Florida. Similarly, dolphin populations are known in the north-central Gulf (Mullin 1988) and off Texas (Shane 1977; Moreno 2005) as well, but none has been followed for as long or as thoroughly as the subject of the Wells (2003) study.
Bottlenose dolphins of the inshore areas of the northern Gulf of Mexico number about 28–32 separate stocks (that are not necessarily distinguished as genetic populations), with an estimated total of 5,355 (no reliable CV available) animals (Table 13.22). Three coastal stocks outside of bays and estuaries and up to 20 m (66 ft) deep, total approximately 13,600 animals (see details, below). The northern continental shelf stock between 20 m and 200 m (66 ft and 656 ft) totals about 17,777 (CV 0.32). The northern oceanic stock deeper than 200 m (656 ft) totals about 3,708 (CV 0.42). Details of these groupings are given below. Anthropogenic influences on this species will be discussed, including toxins, noises, and other aspects, since more is known about these dolphins and their ecology than is known about other cetaceans in the Gulf, largely due to work by Randall Wells and colleagues, which is summarized in Reynolds et al. (2000) and Wells and Scott (2009) (Figure 13.51).

Northern Gulf of Mexico Bays and Sounds. Each of the alpha-numerically designated blocks corresponds to one of the NMFS SFSC logistical aerial survey areas listed in Table 13.1. The bottlenose dolphins (Tursiops truncatus) inhabiting each bay and sound are considered to comprise a unique stock for purposes of this assessment (after Waring et al. 2010). See also Table 13.22.
13.3.2.13.6.1 Northern Gulf of Mexico Bay, Sound, and Estuarine Stock (Often Divided into Communities)
The bottlenose dolphins seen by people inside bays, estuaries, and channels tend to be of the 28–32 stocks mentioned above. The fidelity of these animals to particular areas appears to be quite strong, as evidenced by the well-studied communities (summarized by Reynolds et al. 2000). Nevertheless, as has been noted by many researchers, there is behavioral and genetic interaction between the resident communities and members of the next category (see below) of the three NMFS-designated coastal stocks in U.S. Gulf waters to about the 20 m (66 ft) depth.
Analyses of mitochondrial DNA (inherited only from the mother) variations between communities and along shore indicate clinal variations among areas (Duffield and Wells 2002), and differences in the seasonality of reproduction among sites also suggest genetic differences among communities (Urian et al. 1996).
Studies by Randall S. Wells and colleagues describe the long-term structure and stability of bottlenose dolphin residents of greater Sarasota Bay, Florida, since 1970 (Irvine and Wells 1972; Scott et al. 1990; Wells 1991, 2003). By photo-identification and periodic captures for age, reproductive, and health data, presently five generations have been tracked in the area, including several first seen in the 1970s that are still living. Estimated immigration and emigration rates are about 2–3 % (Wells and Scott 1990). However, while it is rather stable and almost all individuals remain, this is not a wholly isolated, genetically closed population, and at least some calves were sired by nonresidents (Duffield and Wells 2002). While year-round residents occur in other areas as well, at least some animals can move quite long distances, as nearshore animals have been identified up to several 100 km away in Texas waters (Lynn and Würsig 2002). There is some aspect of seasonality as well. In smaller bays such as Sarasota, Florida, and San Luis Pass, Texas, some residents move into Gulf coastal waters during fall and winter and return inshore in spring and summer (Irvine et al. 1981; Maze and Würsig 1999, respectively). In larger bays, there may be even more seasonal migrations, as there is a tendency for greater numbers in northerly bays in summer and southerly bays in winter (e.g., Tampa Bay, Florida, Scott et al. 1989; and Galveston Bay, Texas, Bräger et al. 1994).
The above data must not obscure the fact that most bottlenose dolphin populations or communities of the Gulf are not thoroughly described, and much more information is needed for proper identifications and numbers. Only four populations are sufficiently well known with data from the past 8 years or less for reliable numbers estimates: Sarasota Bay (160 animals, direct count), Choctawhatchee Bay (179 animals, best estimate), Apalachicola Bay (537 animals, best estimate), and St. Joseph Bay (146 animals, best estimate). In total, an estimated 5,355 inshore/nearshore bottlenose dolphins reside in the U.S. waters of the Gulf in 28–32 bays and estuaries from the Laguna Madre, south Texas, to the Florida Keys, south Florida. Unfortunately, data are not good enough for overall trend analysis of numbers, although the one area with good, long-term data—Sarasota Bay—shows a rather constant number since the early 1970s. Maximum net productivity rate is also unknown but has been assumed to be around 0.04 (=4 %) per year (Wade 1998), based on theoretical modeling showing that cetacean populations may not grow at rates much greater than 4 % due to their typical large-mammalian pattern of low pregnancy rate and the production with each pregnancy of only a single calf that exhibits slow growth and requires a long time to achieve sexual/social maturity.
13.3.2.13.6.2 Coastal Stocks
The Gulf of Mexico coastal stocks are divided into eastern, northern, and western bottlenose dolphins that generally occur outside of bays and estuaries, but in Gulf waters less than 20 m (66 ft) deep. There is much contact between these and inshore animals, and contact between these and the greater than 20 m (66 ft) depth dolphins as well. Thus, these should be considered stocks for management purposes, not as separate or distinct behavioral or genetic entities. As mentioned above, genetic clinal-like variations exist, with animals further apart showing greater genetic dissimilarity (e.g., Duffield and Wells 2002). The three coastal stocks have approximate numbers of 7,702 (CV 0.19), 2,473 (CV 0.25), and 3,499 (CV 0.21) for eastern, northern, and western stocks, respectively. Maps of sightings per stock are provided below (Figures 13.52, 13.53, and 13.54).
13.3.2.13.6.3 Gulf of Mexico Eastern Coastal Stock

Locations (circles) of bottlenose dolphin (Tursiops truncatus) groups sighted in coastal waters during aerial surveys conducted in the western coastal stock area in 1992 and 1996 and in the northern coastal stock and eastern coastal stock areas in 2007. Dark circles indicate groups within the boundaries of the Eastern Coastal stock. The 20 and 200 m (66 and 656 ft) isobaths are shown (from Waring et al. 2013).
13.3.2.13.6.4 Gulf of Mexico Northern Coastal Stock

Locations (circles) of bottlenose dolphin (Tursiops truncatus) groups sighted in coastal waters during aerial surveys conducted in the western coastal stock area in 1992 and 1996, and in the northern coastal stock and eastern coastal stock areas in 2007. Dark circles indicate groups within the boundaries of the Northern Coastal Stock. The 20 and 200 m (66 and 656 ft) isobaths are shown (from Waring et al. 2013).
13.3.2.13.6.5 Gulf of Mexico Western Coastal Stock

Locations (circles) of bottlenose dolphin (Tursiops truncatus) groups sighted in coastal waters during aerial surveys conducted in the western coastal stock area in 1992 and 1996, and in the northern coastal stock and eastern coastal stock areas in 2007. Dark circles indicate groups within the boundaries of the western coastal stock. The 20 and 200 m (66 and 656 ft) isobaths are shown. Apparent gaps between stock areas are likely due to inadequate aerial survey coverage (from Waring et al. 2013).
13.3.2.13.6.6 Continental and Oceanic Stocks
Again, there is no clear stock delineation between the shelf and more oceanic animals, with management estimates of the northern continental shelf stock at 17,777 (CV 0.32) between 20 and 200 m (66 and 656 ft); and the northern oceanic stock deeper than 200 m (656 ft) at 3,708 (CV 0.42) (Figures 13.55 and 13.56). Note that bottlenose dolphins overlap on the shelf strongly with Atlantic spotted dolphins (Figure 13.48) and neither species frequents waters deeper than about 1,000 m (Table 13.23).
13.3.2.13.6.7 Northern Gulf of Mexico Continental Shelf Stock

Distribution of bottlenose dolphin (Tursiops truncatus) sightings from SEFSC fall vessel surveys during 1998–2001. All the on-effort sightings are shown, though not all were used to estimate abundance. Solid lines indicate the 100 m (328 ft) and 1,000 m (3,281 ft) isobaths and the offshore extent of the U.S. EEZ (from Waring et al. 2011).
13.3.2.13.6.8 Northern Gulf of Mexico Oceanic Stock

Distribution of bottlenose dolphin (Tursiops truncatus) sightings from SEFSC shipboard surveys during spring 1996–2001 and from summer 2003 and spring 2004 surveys. All the on-effort sightings are shown, though not all were used to estimate abundance. Solid lines indicate the 100 m (328 ft) and 1,000 m (3,281 ft) isobaths and the offshore extent of the U.S. EEZ (from Waring et al. 2011).

Records of bottlenose dolphins (Tursiops truncatus) in the Mexican waters of the southern Gulf of Mexico and Caribbean. Display for distribution is as follows: solid symbol, strandings; hollow symbols, confirmed sightings; triangles, spring (Mar–May); squares, summer (Jun–Aug); circles, autumn (Sep–Nov); diamonds, winter (Dec–Feb); crosses, unknown dates. Thin contour lines show the 200 m, 1,000 m, 2,000 m, and 3,000 m (656 ft, 3,281 ft, 6,562 ft, and 9,843 ft) isobaths (from Ortega-Ortiz 2002).
13.3.2.14 West Indian Manatee
The West Indian manatee (Trichechus manatus) is but one of three manatees, which along with the dugong (Dugong dugon) make up the small mammalian order Sirenia. It is the largest of the manatees, at about 4.6 m (15.1 ft) in length and approximately 600–1,000 kg (1,323–2,205 lb) in weight. It has a paddle-shaped flattened dorsal fin, somewhat like the tail of a beaver (Castor canadensis). Manatees and the dugong are the only true vegetarians of the entire grouping of more than 122 marine mammals, feeding on tropical seagrasses, water hyacinth, and even fruits that drop from vegetation above.
Manatees have been hunted for centuries for meat and hide and continue to be hunted in Central and South America. However, collisions with speeding motorboats, especially in Florida, are the most constant source of manatee fatalities in U.S. waters. In west Florida, deaths due to cold spells are also a major problem, as manatees use natural springs and (more often) warm power plant outfalls as refugia, and the latter can become death traps if a power plant is accidentally or purposefully shut down in winter (Laist and Reynolds 2005).
Two subspecies are currently recognized: the Florida manatee and the Antillean manatee. Both species occur in the Gulf of Mexico, with the Florida manatee in the northeast and the Antillean manatee in Mexican waters in the south (Figure 13.3). The Florida manatee subspecies is protected in Gulf waters by the U.S. Endangered Species Act, Convention on International Trade in Endangered Species of Wild Fauna and Flora (CITES) Appendix 1, and the International Union for Conservation of Nature (IUCN), and they all list it as endangered. Furthermore, the Florida Manatee Sanctuary Act of 1978, the Manatee Recovery Plan, and the Save the Manatee Club all help to create awareness of problems to manatees in U.S. waters and ways to mitigate these problems. While the population worldwide (that is, from northern Florida south to Brazil) is estimated at somewhat fewer than 10,000 mature individuals, about 2,800 occur off east Florida and 2,300 off west Florida in the Gulf (Marsh et al. 2011). No good estimate exists for the Antillean manatee of Mexico in the southern Gulf (Figure 13.58).

West Indian manatee. Note the rounded flippers and tail, large nostrils and small eyes. The light speckling on the back is the reflection of wavelets on the surface (photo by Christopher Marshall, with permission).
In April 2007, the USFWS announced that the West Indian manatee population of Florida was doing well and advised that the species be reclassified as threatened rather than endangered. However, computer models by a federal study showed a 50 % chance that the statewide manatee population, estimated then as about 3,300, could dwindle over the next 50 years to just 500 on either coast if further depredations such as habitat degradation and vessel strikes continued or increased. Presently, there is some disagreement as to how well manatees are doing in U.S. (and other) waters.
During winter months, manatees often congregate near warm-water outflows of power plants along the coast of Florida instead of migrating south as they once did, causing biologists to worry that manatees may have become too reliant on these human-made, warm-water refugia. Laws restricting temporary closures of power plants during cold spells have been put in place (Figure 13.59). Some manatees also move into the northern Gulf of Mexico and are sporadically seen in Alabama, Mississippi, Louisiana, and even Texas (Fertl et al. 2005).

Warm-water refugia of Florida manatees (modified drawing by Adella Edwards from Marsh et al. 2011).
13.3.3 Multispecies Aggregations
Mixed species groups are relatively common among mammals (Stensland et al. 2003) and often have been described among cetaceans as ranging from closely related species or species of similar size to species from different orders or having remarkably different body sizes. Mixed species groups of cetaceans occur in a number of habitats, oceanic as well as coastal, and vary greatly in their structure, frequency, duration, and activity, depending on the species involved and the habitat. As happens more generally among mammals, mixed species groups tend to occur because of foraging advantages, predator avoidance, or both. However, there could be additional social or reproductive benefits that contribute to group formation and stability. These advantages do not need to be equal among the participating species and can vary over time (Stensland et al. 2003).
Although most cetacean groups are monospecific, several species often or regularly associate with other species for variable periods of time. For instance, bottlenose dolphins have been recorded to associate with more than 20 different cetaceans (Ballance 2009), including much larger species such as the humpback whale (Rossi-Santos et al. 2009). Spotted and spinner dolphins occur regularly in mixed schools, especially in the eastern tropical Pacific. Risso’s, Pacific white-sided (Lagenorhynchus obliquidens), and northern right whale (Lissodelphis borealis) dolphins also commonly occur in association (Ballance 2009). Somewhat surprisingly, associations between dolphins and their natural cetacean predators may also take place. For instance, rough-toothed dolphins and bottlenose dolphins sometimes associated with false killer whales off the Hawaiian Islands (Baird et al. 2008).
Mixed species groups also occur in the Gulf of Mexico, but the percentage of such groups in the northern portion of the Gulf has reportedly been low (Maze-Foley and Mullin 2006). For instance, of 736 cetacean groups observed between 1992 and 1998, only 9 (1.4 %) were of mixed species (Mullin and Hoggard 2000; Mullin et al. 2004). No pantropical spotted and spinner dolphin mixed species associations were documented for the northern Gulf, despite the fact that this is a common occurrence in other ocean basins.
Association with birds and fishes is also not as common in the Gulf as it is in some other areas of the world. This may be in part because much of the information has been gathered on deep water cetacean species that tend to feed nocturnally on DSL-related organisms such as myctophids and squid. Nevertheless, Maze-Foley and Foley (2006) summarized the available data and found that about 2.4 % of their cetacean sightings in water deeper than 200 m (656 ft) were associated with birds and 1.1 % were with birds and fish, including surface-dwelling tunas. The most abundant of the oceanic birds in the northern Gulf are terns, the smaller shearwaters, and storm petrels (Hess and Ribic 2000), and while these make up most of the cetacean/bird associations, these associations are also not as prevalent in the Gulf as in much of the Pacific, Atlantic, and Indian oceans (Maze-Foley and Mullin 2006). While it is not certain why this is so, perhaps cetacean feeding is less often at or near the surface in the Gulf and therefore does not as often attract marine birds.
13.3.4 Cetacean Occurrences Relative to Oceanographic Features
The Gulf of Mexico consists of about 1.5 million km2 (579,153 mi2) in waters of the United States, Mexico (whose borders incorporate about 65 % of the Gulf), and the western coast of Cuba. While almost totally enclosed, the Gulf is open to the Caribbean through the Yucatán Channel (about 2,000 m [6,562 ft] deep) and the shallower (about 800 m [2,625 ft] deep) Florida Straits. The Gulf has extensive continental shelf areas less than 180 m (591 ft) deep, with the shelf about 160–240 km (99–149 mi) wide off central and southern Texas, along the Florida west coast, and the Campeche Bank north of the Yucatán Peninsula, covering about 35 % of the Gulf. However, this shelf is only about 32–48 km (20–30 mi) wide south of the Mississippi River and is very narrow off Tampico, Mexico. The intermediate area between slope and abyss, at depths of 180–3,000 m (590–9,843 ft), covers about 40 % of the Gulf. The abyssal plain, at depths greater than 3,000 m (9,843 ft)—Sigsbee Plain in the west and parts of the Lower Mississippi Fan—makes up the remaining 25 %. Since cetacean habitats are in large part influenced by basic depth characteristics, it is no surprise that bottlenose dolphins are the only cetaceans normally seen off the shallow coast of Texas, out to many dozens and even more than 100 km (62 mi) from shore, while a host of more deepwater species, including sperm whales, are found within tens of kilometers of shore south of Louisiana (Davis and Fargion 1996).
Warm water from the Caribbean Sea flows into the Gulf through the Yucatán Channel, forms a Loop Current in the mid-eastern part of the Gulf, and flows out of the Gulf through the Florida Straits into the Atlantic Ocean. In the central and western Gulf, warm anticyclonic eddies that have shed off the Loop Current move slowly towards the west with adjacent cold cyclonic eddies (Sturges and Leben 2000). Upwelling cold eddies and interfaces between cold and warm eddies are areas where elevated chlorophyll levels and higher productivity occur, such as in estimated mean biomass, EMB, for example, Wormuth et al. (2000). These areas provide for a more rich, near-surface flora and fauna than outside of these zones, and therefore at least some cetaceans are attracted to them. Also, the Mississippi/Atchafalaya River complex and other rivers provide much nutrient-rich fresh water to the Gulf, draining about two-thirds of the continental U.S. watershed and one-half that of Mexico. Besides nutrients, associated land-runoff pollutants and sediments also influence the slope waters in the northern Gulf. River discharge is quite seasonal—highest flows March–May, and the lowest flows August–October—and provides rich shelf areas for spawning and juvenile fishes. For example, much Gulf menhaden (Brevoortia patronus) spawning occurs off the Mississippi Delta (Christmas and Waller 1975).
While a thorough analysis of physical and biological features of the northern Gulf relative to primary/upper level productivity is beyond the scope of this chapter, some interesting comparisons can be made of the dynamic nature of oceanography as related to cetacean occurrence patterns. For that discussion, Davis et al. (2002) separate (1) sperm whales as squid feeders; (2) smaller squid feeders (i.e., dwarf and pygmy sperm whales, melon-headed whales, pilot whales, pygmy killer whales, Risso’s dolphins, rough-toothed dolphins, and beaked whales of the taxonomic family Ziphiidae); (3) oceanic dolphins of the genus Stenella (i.e., Clymene, pantropical spotted, spinner, and striped dolphins); and (4) upper slope bottlenose and Atlantic spotted dolphins. A final grouping is that of the nearshore and inshore living bottlenose dolphins and West Indian manatees. Killer whales were not seen during many surveys used to generate this information, but they are known to travel widely through shelf and deeper waters (Ortega-Ortiz 2002).
Sperm whale sightings in the 1990s Gulfcet studies (Davis et al. 1998, 2002) and in subsequent work (for example, Ortega-Ortiz et al. 2012) show that they are consistently present in lower slope waters south and west-south-west of the Mississippi River outfall, in a mean depth of 1,580 m (5,184 ft), and with high EMB, in colder gyre and interface cyclonic/anticyclonic waters. It is likely that the river outflows from Louisiana are especially important given that productivity is high directly south of the Delta. Although sperm whales are present in these lower slope waters year-round, they move according to productivity as measured by remote sensing, with an average time lag of about 2 weeks from primary productivity to sperm whale presence (O’Hern and Biggs 2009). Sperm whales have been in this lower slope area for a long time, as indicated by whaling records of the 1700s and 1800s (Reeves et al. 2011).
Squid feeders of the slope and deeper parts of the Gulf tend to be associated with higher salinity (less riverine influence) waters over waters of the lower, deeper slope, and in conjunction with cold-core (cyclonic) eddies or confluence zones but not in the warm anticyclonic areas.
Oceanic Stenella dolphins also occur most often in cold-core cyclonic rather than anticyclonic zones. They do not generally occur on the upper slope or in the abyssal, greater than 3,000 m (9,843 ft) deep zone. The pantropical spotted dolphin is the most numerous of any cetacean in the Gulf of Mexico and presents a striking feature of large group size—often more than 100 dolphins per group—and general abundance.
The more important use of cold-core rather than warm-core eddies/gyres can be found in numerous examples of data presented by Davis et al. (2002) and Ortega-Ortiz (2002) as well as subsequent studies (O’Hern and Biggs 2009). One snapshot of this general situation for the northwest central Gulf is presented in Figure 13.60.

Cetacean (all identified species, summarized) sightings (+) during the mid-summer 1997 Gulfcet cruise. Thin dashed lines represent isobaths. Bold solid (positive) and bold dashed (negative) lines are sea surface dynamic height anomaly (DHA), relative to the 105-cm (41-in.) mean. Contour intervals are 5 cm (2 in.). The cyclone is between 0 and 13 cm (0 and 5 in.), the confluence between 0 and 24 cm (0 and 9 in.), and the anticyclone between 25 and 71 cm (9 and 28 in.). The color contours indicate nighttime EMB—estimated mean biomass (cm3m−2) (see scale above to the right) in the interval 10–50 m (33–164 ft) (data and figure legend from Davis et al. 2002).
Atlantic spotted dolphins and the offshore ecotypes of the bottlenose dolphin prefer upper-slope, continental shelf waters, and they frequent waters with a mean depth of about 200 m (656 ft). Bottlenose dolphins of nearshore and inshore areas form a separate population grouping and prefer productive, river outflow-influenced waters less than 20 m (66 ft) deep. Besides manatees (largely of west Florida), bottlenose dolphins are the only marine mammal species to occur in very shallow waters and the only one to do so in the western Gulf. Bottlenose dolphins outside of bays and inlets and within the about 20 m (66 ft) depth contour also engage in a partial migration, tending to move further north in summer and further south in winter. However, these movements are not likely to be much greater than about 100 km (62 mi) in most cases and are presently poorly understood (Shane et al. 1986).
Manatees prefer riverine and shallow nearshore waters where temperatures are above 17 °C (63 °F) and where seagrasses, water hyacinth, and aquatic weeds are abundant (Marsh et al. 2011). Along the western Florida coast, especially north of Warm Mineral Springs (Figure 13.59), warm-water refugia provided by waste cooling water from power plants are of special importance.
13.4 Anthropogenic Impacts
After a summary of the effects of physical and biological factors that influence numbers and habitat use of marine mammals in the Gulf, it is appropriate to consider human influences (i.e., anthropogenic impacts). Major anthropogenic impacts with the potential to affect cetacean and sirenian individuals and populations defined below have been modified and are based on Bearzi et al. (2011). Not all have been documented for the Gulf of Mexico. Data for each are summarized in this section:
-
Prey depletion—Depletion of food resources caused directly or indirectly by fishing
-
Incidental mortality and injury in fisheries (bycatch)—Mortality or injury from accidental entanglement in gear of various types including passive and active nets, longlines, traps and discarded or lost nets and lines, and illegal fishing practices (e.g., use of high explosives such as dynamite)
-
Intentional and direct takes—Killing or capture to obtain products for animal or human consumption, live capture for public display facilities, acts of retaliation for actual or perceived damage to fish catches or gear, and shooting for sport
-
Vessel strikes—Accidental mortality or injury from contact with a vessel, particularly the hull or propeller
-
Disturbance—Behavioral disruption through intentional or unintentional approaches, with the potential to induce long-term effects on dolphin populations
-
Acoustic pollution (noise)—Mortality, injury, or chronic disturbance from exposure to human-made sounds
-
Chemical contamination—Accumulation in the body tissues, mostly through the food web, of chemicals known to adversely affect mammalian functions and health, in particular, persistent organic pollutants (POPs)
-
Ingestion of solid debris—Mortality or injury from the ingestion of foreign objects and materials (e.g., plastic, wood, textiles) obstructing part of the digestive tract
-
Oil pollution—Health problems or mortality deriving from contamination, contact or ingestion of hydrocarbons derived from oil spills and oil derivatives at sea
-
Ecosystem change—Reduced habitat quality due to effects of coastal or other development (e.g., eutrophication, harmful algal blooms, alien species invasions)
-
Climate change—Changes, potentially due to natural or human-caused climate change in, for example, prey abundance or distribution, shifts in distribution of competitors and exposure to novel diseases
13.4.1 Prey Depletion
Excessive fishing pressure and the resulting decline in fish stocks and loss of marine biodiversity is a growing concern worldwide (Pauly et al. 1998, 2002; Jackson et al. 2001; Worm et al. 2006, 2009; Swartz et al. 2010; Anticamara et al. 2011). Jackson et al. (2001) argue that “ecological extinction caused by overfishing precedes [i.e., is more important than] all other pervasive human disturbance to coastal ecosystems, including pollution, degradation of water quality, and anthropogenic climate change.” Overfishing may contribute to the decline of marine mammal populations by affecting the availability of key prey (Bearzi et al. 2008). Several marine mammal populations around the world have declined rapidly, and overfishing has been suggested as one of the reasons behind their collapse (Crowder et al. 2008; Heithaus et al. 2008; Read 2008).
Pauly and Palomares (2005) analyzed landings data from fisheries in the Gulf of Mexico (as well as in the U.S. Atlantic Ocean south of Chesapeake Bay) and conclude that these regions were severely overfished and had badly degraded food webs, as evidenced by a low and declining mean trophic index, which assesses the trophic levels at which fisheries are operating. However, de Mutsert et al. (2008) subsequently point out that fisheries landings in these regions historically would be expected to have low indices because the fisheries have been dominated by menhaden and shrimp, both of which feed at low trophic levels. These authors argue that low indices derived from landings data are driven by large landings of commercial species of low trophic level, particularly Gulf menhaden and penaeid shrimp species. De Mutsert et al. (2008) also question the predictions of near-future collapses of fish populations in the Gulf of Mexico made by Worm et al. (2006), and show that—although several taxa of fish and shellfish, as well as several fisheries, have indeed collapsed in this region—the overall scenario is less dramatic than depicted in the earlier article by Worm et al. (2006). However, because the conclusions cited above are based on different datasets (i.e., the ones by the Food and Agriculture Organization of the United Nations, FAO, and NMFS), which also refer to different geographic areas, it is presently difficult to endorse either scenario.
Little information is available to permit an assessment of the effects of possible past and current impacts on marine mammals resulting from prey depletion caused by fishing in the Gulf of Mexico. A study of the potential effects of hurricane Katrina suggests that calving by bottlenose dolphins in the years following the storm may have increased for reasons including higher resource availability resulting from reduced fishing pressure, since much of the fishing fleet was destroyed by the hurricane. Calving also might have increased because many young calves were likely lost due to the hurricane, and therefore pregnancies increased afterwards. A combination of these or other factors may have been involved (Miller et al. 2010).
13.4.2 Incidental Mortality and Injury in Fisheries (Bycatch)
Fishing can deplete populations of marine mammals and other endangered megafauna, such as sharks and marine birds, in many parts of the globe through incidental bycatch in fishing gear (Lewison et al. 2004; Read 2008). Most of the following information applies primarily to bottlenose dolphins, the species that is closest to shore and has been most studied in the Gulf of Mexico. Relatively little is known even about bottlenose dolphins and the importance of impacts from fisheries; potential issues with other species are even less clear.
According to the total analyzed records from the southeastern U.S. Atlantic, including the Gulf, from 1990 to 2008, 112 (2.8 %) of the 4,029 animals stranded bore signs of fishery interaction, defined as “wounds related to fishing gear, or fishing gear attached to the animal” (Southeast U.S. Marine Mammal Stranding Network 2011). In addition to the animals reported as bearing signs of fishery interaction, a further 123 animals (3.1 %) were reported to have borne signs of human interaction, defined as “ingested plastic, debris entanglement, wounds from other weapons besides firearms (arrows, harpoons, etc.).” Side notes and specifications added to the records show that a large part of these human interactions also included signs of fishery bycatch: entanglement, amputations, cuts, and other signs that are often related to bycatch, but that at times may also be due to vandalism after stranding (Kuiken 1996; Read and Murray 2000).
Because of body decomposition at the time of inspections and other difficulties implicit in such assessments as the one described above (e.g., parts of the body not visible during the inspection, lack of necessary expertise, etc.), the numbers reported should be considered a minimum indicative estimate of the occurrence of fishery interaction among stranded animals. For example, the Southeast U.S. Marine Mammal Stranding Network (2011) reports that in 2,949 of 4,029 records, the occurrence of human interaction could not be determined due to decomposition or other problems. Many of these stranded animals are likely to have succumbed due to fisheries or other human-related interactions, but exact numbers are unknown.
Wells and Scott (1994) found that of 146 bottlenose dolphins handled during scientific catch and release live captures, about 11 % showed signs of previous gear entanglement (rope cuts, marks, etc.). However, evidence of mortality in the Sarasota-Bradenton area of west Florida was extremely low. A detailed review of their study animals yielded the conclusions that while many dolphins survive human interactions, swallowing of hooks and body constrictions by lines more often led to mortality. However, no clear numbers were available (Wells et al. 1998, 2008). Garrison (2007) describes incidents of pilot whales and Risso’s dolphins becoming entangled in pelagic longlines and being released by fishermen with lines and hooks still embedded, with the supposition that many of these animals were subject to eventual mortality due to the line and hook interactions.
There is also a scarcity of information in the southern Gulf, with sporadic reports of confirmed bycatch problems. For example, of 15 records of stranded cetaceans inspected on the Veracruz coast, southwestern Gulf of Mexico, two pygmy sperm whales and one bottlenose dolphin died as a result of entanglement in gillnets (Ortega-Argueta et al. 2005). Vidal et al. (1994) surmise from limited data that especially bottlenose, Atlantic spotted, and spinner dolphins are at risk of entanglement due to gillnets in the southern Gulf. Bycatch of bottlenose dolphins is reported to be “practically zero” by the Cuban CITES administrative authority but is likely to occur in unknown numbers (Van Waerebeek et al. 2006).
Commercial fisheries that may interact with bottlenose dolphins are shrimp trawling, blue crab trap/pot fishing, stone crab trap/pot fishing, menhaden purse seining, gillnetting, and shark bottom longline fishing (Waring et al. 2010). Lack of (complete) observer program data for some of these fisheries means that the information reported below, which is limited to bottlenose dolphins, should be treated as indicative.
-
Shark Bottom Longline Fishery—Three interactions with bottlenose dolphins have been recorded since the fishery started being observed in 1994: one mortality in 2003 and two hooked animals in 1999 and 2002 (Burgess and Morgan 2003a, b). No interactions were observed between 2004 and 2008 (Hale and Carlson 2007; Hale et al. 2007, 2009; Richards 2007). Bottlenose dolphin mortalities were estimated at 58 (CV 0.99) for 2003, but none for 2004–2008 (Richards 2007).
-
Shrimp Trawl Fishery—Information recorded since 1992 shows that a few dozen animals have died in this fishing gear or have been caught in turtle excluder device or trawl line (Waring et al. 2010).
-
Blue and Stone Crab Trap/Pot Fisheries—A few stranded bottlenose dolphins had polypropylene rope around their flukes (Waring et al. 2010) suggesting possible entanglement with crab pot lines.
-
Menhaden Purse Seine Fishery—Bottlenose dolphins have died incidentally in this fishery (Reynolds 1985) with numerous self-reported kills in northern Gulf coastal and estuarine waters from the 1970s to the 1990s. The fishery was observed to take (in this sense, take means caught, including animals released) nine bottlenose dolphins, with three killed, between 1992 and 1995. Extrapolation of takes from 1992 through 1995 that considered the total number of sets indicates that up to about 172 bottlenose dolphins could have been harmed and up to 57 animals could have been killed by menhaden purse seining (Waring et al. 2010).
-
Gillnet Fishery—Stranding data for this fishery suggest that there is probably a low frequency of takes that occur. For example, five research-related gillnet mortalities were documented between 2003 and 2008 in Texas and Louisiana (Waring et al. 2010). This is suggestive of a potential for incidental mortality in this fishery.
13.4.3 Intentional and Unintentional Direct Takes
The capture of animals from a wild population removes them from that population, and in terms of recruitment, population dynamics, and conservation value, they are effectively dead. This loss is exacerbated if certain animals are preferentially removed, such as young females, as is often the case (Van Waerebeek et al. 2006).
Since captures ceased in U.S. waters in 1989, intentional takes of bottlenose dolphins off the northeast coast of Cuba are an isolated case of removal in the Gulf of Mexico area. Nevertheless, social group effects could still be present in nature, as these dolphins can live more than 50 years (Urian et al. 2009). Bottlenose dolphins have been targets of live-capture fishery off Cuba since at least 1982. Removals occur off Sabana-Camagüey Archipelago, and 238 animals were exported from Cuba between 1986 and 2004 for the global captive dolphin industry. Twenty-eight animals were captured in 2002 alone (Van Waerebeek et al. 2006); this may be an ongoing serious problem, since global demand for aquarium dolphins is also increasing over time (Fisher and Reeves 2005). Van Waerebeek et al. (2006) recommend that the Cuban live trade in bottlenose dolphins cease until evidence of no detriment can be substantiated, but numbers within communities/populations have not been well documented, with the regional exception of Pérez-Cao’s (2004) master’s thesis regarding northeast Cuban waters.
Until such takes stopped in 1989, bottlenose dolphins were live-captured from several northern Gulf bays and sounds, to supply the U.S. Navy and aquarium trade (Waring et al. 2010). Between 1972 and 1989, 490 dolphins, at an average of 29 per year, were taken from several bays in the north-central Gulf as well as Tampa Bay, Charlotte Harbor to the south, and the Florida Keys. Many captures occurred in Mississippi Sound, with 202 dolphins taken. Of the dolphins captured from 1982 to 1988, 73 % were females (Waring et al. 2010), and because these animals are long-lived and slow in reproducing, population and social effects from those removals could still be present to this day.
Intentional and direct takes, such as purposeful wounding or killing of animals that are perceived to be in conflict with fisheries activities, may also take place by illegal means. Before 1988, fishermen were permitted to use almost any method, including lethal means, to protect their gear and catch. But in 1988, Congress amended the Marine Mammal Protection Act (MMPA) to forbid the lethal taking of cetaceans. Despite their protected status, cetaceans are still being shot. The Southeast U.S. Marine Mammal Stranding Network (2011) indicates that at least some strandings are of bottlenose dolphins, and in one instance, a dwarf sperm whale had been shot or otherwise wounded or killed. While this kind of interaction is presumably infrequent, actual numbers are not known. Fishermen occasionally kill or harm dolphins in retaliation for depredation of recreational and commercial fishing gear. Three cases were documented between 2006 and 2008 (Waring et al. 2010).
Two dolphin research-related mortalities occurred in western Florida in 2002 and 2006. Four others resulted from entanglements in gillnet fisheries research gear off Louisiana and Texas from 2003 through 2007. Five incidents—four of which were mortalities—involving bottlenose dolphins occurred during sea turtle relocation trawling activities by the Army Corps of Engineers. Overall, intentional or otherwise unintentional but direct takes occur now and then. However, they are not perceived to be a large problem that would endanger populations or species in the Gulf of Mexico.
13.4.4 Vessel Strikes
Vessel strikes can injure or kill a variety of marine mammals including large whales (Laist et al. 2001; Panigada et al. 2006) and dolphins (Wells and Scott 1997; Van Waerebeek et al. 2007; Wells et al. 2008). In the coastal waters of the Gulf of Mexico, marine mammals share habitat with large and increasing numbers of boats. Bottlenose dolphins suffer boat-related injuries (e.g., Wells and Scott 1997), and injuries and deaths may be high in areas of high boat traffic. Of 637 total bottlenose dolphin strandings during 2004–2008, seven showed signs of boat collision (Waring et al. 2010). Bottlenose dolphins often survive propeller strikes if these involve only soft tissue and not bone (Wells et al. 2008). Extrapolation from studies of bottlenose dolphins to other dolphin species must be done with caution, however, and Wells et al. (2008) recommend against it, especially since the very nearshore interactions tend to be with small pleasure craft, whereas offshore interactions are more likely to involve medium-size and large vessels.
A recent study by Azzara (2012) compared the known occurrence of sperm whales off New Orleans with the pattern of major shipping lanes related to the ports of New Orleans and Houston and found a high overlap between critical sperm whale habitat and vessel traffic. Because extremely high shipping traffic occurs in the areas that are most often used by sperm whales, the potential for collision exists, but these have not been reported. It is known that ship strikes increase mortalities with vessels longer than 80 m (262 ft) and traveling at speeds greater than 26 km/h or 14 knots (kts; 16 mi/h) (Laist et al. 2001).
Most living Florida manatees bear scars from vessel collisions, and it is estimated that about 30 % of all mortalities in U.S. waters are caused by such collisions (Wright et al. 1995). Manatees are generally aware of approaching vessels and attempt to evade the vessel by orienting into deeper waters of channels, and by swimming faster. However, since a rapid boat is likely to be in deepest water close to the center of channels, this behavior does not always decrease the collision potential (Nowacek et al. 2004). It is well established that slower boat speeds in manatee habitat can greatly reduce injuries and mortalities, and some progress has been made on stricter regulations, enforcement, and public awareness in recent years (Marsh et al. 2011).
13.4.5 Disturbance and Acoustic Pollution (Noise)
Sound is probably the most important sensory modality for cetaceans. All species communicate by sound. Baleen whales communicate at great distances, with fin and blue whales communicating across many tens of kilometers (Payne and Webb 1971); and toothed-whales echolocate in sophisticated fashion (Au 1993). Prolonged direct (or physical) disturbance caused by boat traffic can affect the behavior and habitat use of cetaceans. Alterations of surfacing patterns, swimming speed, directionality, group cohesion, and group fluidity have been related to boat disturbance in a number of cetacean species—several of which live in the Gulf of Mexico. Vessel traffic can also affect habitat use, which can include displacement for periods of hours to days and longer-term avoidance of areas, and reduce reproductive success (Nowacek et al. 2007).
Some of the short-term effects of boat disturbance on coastal dolphins in the Gulf of Mexico were investigated in the waters of Sarasota Bay, where dolphin whistle rates were found to increase at the onset of vessel approaches. This could be because of heightened arousal, increased motivation of animals to come close together, or to compensate for signal masking due to noise (Buckstaff 2004). Behavioral responses including changes in grouping patterns and headings as well as increased swimming speed occurred more often during experimental vessel approaches than during control (no disturbance) periods (Nowacek et al. 2001). Another study in the Mississippi Sound found short-term changes in dolphin behavior, including a decrease in feeding behavior, following the passing of speedboats (Miller et al. 2008). The long-term effects of such disturbances and the possible impacts on populations remain unknown. However, the Sarasota Bay population is known to have remained rather stable in size over the past four decades (Wells and Scott 2009).
The Gulf of Mexico is home to two of the world’s ten busiest ports by cargo volume: New Orleans and Houston. In 2008, these ports hosted a combined total of 14,000 ships. An indication of behavioral disruption in sperm whale communication caused by shipping noise was found in a recent study by Azzara (2012). Azzara used recordings from one hydrophone recording buoy situated at a depth of approximately 1,000 m (3,281 ft), close by a junction of both the New Orleans and Houston waterways for major shipping. The study found significant differences in sperm whale vocalization patterns before, during, and after the passing of a ship. While this study does not show that shipping noise can negatively affect survival or reproduction, the findings are suggestive of potentially important behavioral alterations that can alter communication and foraging success and potentially cause stress. Industrial noises have been shown to change calls of several species of whales; a recent example is of blue whales (Melcón et al. 2012). Physiological stress (measured from stress hormones contained in scat samples) has been indicated relative to shipping noise and North Atlantic right whales off the U.S. Atlantic seaboard (Rolland et al. 2012).
The oceans have become much louder during the industrial (propeller-driven) age than before, especially in the lower frequencies to which baleen whales are most sensitive (McCarthy 2004). Of these noises, industrial seismic airguns for oil and gas exploration and military sonars are especially loud, and have increased ocean noise in several areas. An experimental study investigating the impact of airguns on sperm whales suggested that sperm whales in a highly exposed area of the Gulf of Mexico may not exhibit avoidance reactions to airguns, but the animals may be affected at ranges well beyond those currently regulated due to more subtle effects on their foraging behaviors (Miller et al. 2009). The high intensity of Navy-produced surveillance echolocation pings—low frequency active sonar (commonly referred to as LFA)—is known to cause death in especially deep-diving cetaceans such as some beaked whales and pilot whales and are therefore also a worry for the Gulf of Mexico (Southall et al. 2007).
A special form of disturbance is the intentional feeding of dolphins for purposes of tourism. Feeding wild dolphins is considered a form of take under the MMPA, since it changes natural behavior and has the potential for increasing injury or death or even creating dependency. Frequent provisioning was observed near Panama City Beach (Samuels and Bejder 2004), south of Sarasota Bay (Cunningham-Smith et al. 2006; Powell and Wells 2011), and in Texas near Corpus Christi (Bryant 1994). Swimming with wild bottlenose dolphins—illegal under the MMPA—has been documented near Panama City Beach, and it is likely that dolphins were attracted by swimmers due to provisioning (Samuels and Bejder 2004).
13.4.6 Chemical Contamination
Toxic contaminants are a major concern in marine mammal populations because of their environmental persistence and the potential effects on reproduction and health (Gauthier et al. 1999; O’Shea et al. 1999; O’Hara and O’Shea 2001; Newman and Smith 2006). Because cetaceans are long-lived apex predators with extensive fat stores, they accumulate persistent organic pollutants (POPs)—including brominated flame retardants, dichlorodiphenyltrichloroethane (DDT) and associated compounds, polychlorinated biphenyls (PCBs), hexachlorocyclohexanes (HCHs), chlorobenzenes, and chlordanes—from lower trophic level prey (Ross 2006; Ross and Birnbaum 2003; Yordy et al. 2010). Causal links have been described between POPs exposure and immunological, endocrine, and reproductive disorders in cetaceans (Aguilar and Borrell 1994; Lahvis et al. 1995; Jepson et al. 1999; Schwacke et al. 2002; Hall et al. 2006).
Coastal stocks of bottlenose dolphins live in highly populated areas, of which some—Tampa Bay, Florida; Galveston Bay, Texas; and Mobile Bay, Alabama, for example—have much industry. Around the periphery of Galveston Bay, more than 50 % of all U.S. chemical products are manufactured and about 17 % of oil produced in the Gulf of Mexico is refined (Henningsen and Würsig 1991). Concentrations of anthropogenic chemicals and their metabolites vary from site to site and can reach levels of concern in bottlenose dolphins (Schwacke et al. 2002). Most studies conducted in the Gulf of Mexico focus on common bottlenose dolphins, particularly in western Florida (Wells et al. 2004). Levels of POPs in dolphins sampled in the northwestern Gulf of Mexico are known to be relatively high (Kucklick et al. 2011), variable by sex and age class, and have negative effects on health and reproduction. Similar levels of POPs have been found in melon-headed whales that stranded in the Gulf of Mexico (Davis 1993).
The high POPs burden carried by bottlenose dolphins in the Gulf of Mexico may increase susceptibility to parasitic microorganisms (Kuehl and Haebler 1995) and suppress immune function and increase recovery time postinfection in marine mammals (Kendall et al. 1992). There is a probable relationship between high concentrations of organochlorines (such as PCB and DDT metabolites) in the blood and male immune dysfunction (Lahvis et al. 1995). Increasing PCB concentrations may induce vitamin A deficiencies in marine mammals that can lead to reproductive disorders and susceptibility to infection (Brouwer et al. 1989).
In Sarasota Bay, Florida, Wells et al. (2001, 2003) found that transfer of organochlorines through placental and mother’s milk was implicated in mortality of first-borns to (generally) young mothers (Vedder 1996; Wells et al. 2004). First-born bottlenose dolphin calves are estimated to accumulate approximately 80 % of their mother’s organochlorines during the first 7 weeks of lactation (Cockcroft et al. 1989). Dead young dolphins had high PCB and chlorinated pesticide concentrations; these concentrations increased as males aged but declined to lower concentrations as females reared offspring (Küss 1998; Wells et al. 2004). Schwacke et al. (2010) and Wells et al. (2003, 2005) described a similar pattern based on samples of blubber and blood.
Organochlorines were higher in females whose calves died within the first 6 months compared to females whose calves survived (Reddy et al. 2001). POPs concentrations in adult female bottlenose dolphins in Sarasota, Florida, are generally greater in blubber than milk; however, there is congener-specific variation in mobilization of POPs from blubber to milk (Yordy et al. 2010). Deceased suckling calves collected throughout the Gulf of Mexico had nearly ten times higher concentrations of triphenylphosphate (TTP) in their blubber than adult males (Kuehl and Haebler 1995). Young suckling dolphins may be at higher risk for POPs-related health effects as their bodies undergo rapid development.
Based on probabilistic risk assessment, bottlenose dolphins sampled off Sarasota, Florida, and Matagorda Bay, Texas, indicated a high likelihood that reproductive success—primarily in primiparous females—is severely impaired by chronic exposure to PCBs. Excess risk of reproductive failure for primiparous females, measured in terms of stillbirth or neonatal mortality, was estimated as 79 % for the Sarasota sample, and 78 % for the Matagorda Bay sample (Schwacke et al. 2002). High levels of infertility were also found among common dolphins with the highest PCB burdens and most ovarian scars, which suggest that ovulation was occurring without the reproduction of a viable calf (Murphy et al. 2010). High POPs burdens also correlated with few ovarian scars in harbor porpoises, suggesting inhibition of ovulation cycles (Murphy et al. 2010).
Levels of trace elements may also be high in the Gulf of Mexico and increase with age class (e.g. Kuehl and Haebler 1995; Bryan et al. 2007). While the link between the concentration of these elements and cetacean population health and status needs to be further clarified, Rawson et al. (1993, 1995) related indicators of liver disease to high mercury concentrations in stranded dolphins from western Florida.
The findings summarized here for cetaceans highlight the importance of considering indirect anthropogenic stressors such as contaminant pollution in U.S. management schemes (Wells et al. 2005). Fine-scale spatial variation in POPs suggests that individual patterns of habitat usage can influence individual toxin burden profiles (Litz et al. 2007; Pulster and Maruya 2008). Concentrations of DDT decreased in bottlenose dolphins in the Gulf of Mexico following a ban on commercial use of DDT in the United States (Salata et al. 1995), suggesting that dolphins can recover from high contamination loads if exposure decreases.
Manatees tend to have low levels of organochlorine residues in their bodies, possibly because of their low position in the food web, the lowered recent pesticide levels in Gulf waters than during earlier studies such as by O’Shea et al. (1984), and possibly a more effective mechanism for metabolizing toxic compounds than is possessed by most terrestrial mammals (Ames and Van Fleet 1996).
13.4.7 Ingestion of Solid Debris
The world’s oceans contain much plastic debris (Wolfe 1987; Laist et al. 1999; Derraik 2002) and obstruction of the alimentary canal or stomach/intestine due to ingested plastic is a known cause of cetacean mortality (e.g. Tarpley and Marwitz 1993). Much of plastic ingestion is probably due to investigating and testing items in the environment, especially for young calves recently weaned, although starvation or disease could also exacerbate such inappropriate attempts at feeding (Kastelein and Lavaleije 1992; Baird and Hooker 2000; Poncelet et al. 2000).
In 2006, three well-known dolphins of Sarasota Bay died from ingesting fishing gear, an unprecedented mortality level (Powell and Wells 2011). Additional scattered information for the Gulf of Mexico comes from the Southeast U.S. Marine Mammal Stranding Network (2011), where several animals are reported to have ingested plastic and other debris. The threat is generally considered as relatively minor, but potentially important for some teuthophagous cetacean species, such as Risso’s dolphins (Bearzi et al. 2011).
13.4.8 Oil Pollution
Oil pollution can impact many parts of an ecosystem, from primary production to fishes and of course birds, sea turtles, and marine mammals (Loughlin 1994). Oil fouling of furred marine mammals has been studied most and is thought to be most dangerous, especially in sea otters that rely almost exclusively on aerated fur for thermoregulation, often in very cold environments. Williams and Davis (1995) provide a quite thorough review; however, no furred marine mammals exist in the Gulf of Mexico, so this is not a problem there.
Cetaceans feed at various trophic levels. The balaenid whales, such as North Atlantic right whales, feed on tiny calanoid copepods; blue whales feed on somewhat larger euphausiid crustaceans; other baleen whales (such as the Bryde’s whales of the Gulf) feed largely on fishes; some odontocetes feed on small to medium fishes (bottlenose dolphins of nearshore waters); and other odontocetes feed on fishes and squid in the deeper ocean. A host of benthic organisms concentrate petroleum hydrocarbon residues in their tissues, but most mid and surface water crustaceans and teleost fishes metabolize and excrete them rapidly, and thus do not tend to become heavily (or for a long time) contaminated (Neff 1990). The cytochrome P450 system can be used as a biomarker to indicate exposure to polycyclic aromatic hydrocarbons (PAHs), as well as to a variety of other chemicals and stressors. However, this biomarker does not show a strong link to toxicity, lesions, or reproductive failure (Lee and Anderson 2005). A major problem for baleen whales that feed on the surface or gray whales (Eschrichtius robustus) that feed largely on in-benthic fauna is baleen fouling; feeding in oil slicks is therefore detrimental to these filter feeders (Geraci 1990). However, neither surface- nor bottom-foraging baleen whales regularly occur in the Gulf, and therefore, baleen fouling is not likely to be a problem there.
The Gulf of Mexico has many natural oil seeps, so it is likely that cetaceans are quite used to dealing with them. Natural seeps have been occurring not only throughout the evolutionary history of cetaceans but also before. Approximately 47 % of crude oil that is introduced into the marine environment occurs via natural seeps (Kvenvolden and Cooper 2003); however, these estimates involve broad extrapolations based on little data (NRC 2003). The Gulf of Mexico has more than 600 natural oil seeps, and one recent study identified at least 164 of these in the northern Gulf of Mexico (NASA 2000; Hu et al. 2009). Natural oil seeps have established meiofaunal communities comprised of deposit-feeding taxa capable of handling toxic environments with low oxygen levels (Steichen et al. 1996). Nevertheless, a major oil spill presents a larger and more intensive footprint than natural seeps, and potential large-scale deaths of cetacean prey may have strong, but at this time unknown, detrimental effects. Bottlenose dolphins studied behaviorally during the Mega Borg oil spill off Galveston, Texas, in 1990 showed no ability to travel out of volatile new oil. They showed some behavioral reactions to surface oil while in it, but no deaths or other signs of long-term damage were evident (Smultea and Würsig 1995). There are no data to indicate that oil spills to early 2010 in the Gulf of Mexico substantially impacted cetaceans.
13.4.9 Unusual Mortality Events
Several major unusual mortality events (UMEs) occurring in recent years have raised concern for the health of bottlenose dolphin populations along the coasts of the eastern and northern Gulf of Mexico, particularly in Florida and Texas. Since 1991 and through March 2010, 13 bottlenose dolphin and seven manatee UMEs were declared in the Gulf of Mexico. A more recent UME (January–March 2010) in the northern Gulf included a combination of bottlenose dolphins, Atlantic spotted dolphins, and pygmy and dwarf sperm whales (Table 13.24).
Mortality events can occur as a result of algal blooms and red tides involving the release of neurotoxins into the marine food web. For instance, a bloom of the dinoflagellate (Karenia brevis) occurred in western Florida in 2004, which resulted in the death of 107 bottlenose dolphins and 34 Florida manatees (Waring et al. 2010; Flewelling et al. 2005). A similar major UME occurred in 2008 in the coastal waters of Texas, resulting in over 100 bottlenose dolphin deaths. This second mortality event overlapped spatially and temporally with an algal bloom of the toxin-producing Dinophysis spp. and Prorocentrum spp. (Fire et al. 2011). Fish and seagrass can accumulate high concentrations of the brevetoxins produced by dinoflagellates and act as toxin vectors to marine mammals (Flewelling et al. 2005; Fire et al. 2007, 2008, 2011).
Red tide blooms occur in the Gulf of Mexico on a nearly annual basis. A recent study showed that bottlenose dolphins in western Florida were consistently exposed to brevetoxin and/or domoic acid over a 10-year study period (2000–2009), and 36 % of all animals tested positive for brevetoxin (n = 118) and 53 % tested positive for domoic acid (n = 83) (Twiner et al. 2011).
Harmful algal blooms are also known to alter dolphin behavior. A study in the coastal waters of western Florida found that bottlenose dolphins displayed a suite of behavioral changes associated with red tide blooms, including altered activity budgets, increased sociality, and expanded ranging behavior. These behavioral changes may result in more widespread population impacts and increased susceptibility to disease outbreaks. While the mechanisms behind red tide-associated behavioral effects are not well understood, they are most likely linked to changes in resource availability and distribution (McHugh et al. 2010).
13.4.10 Climate Change
Climate change may cause large-scale and long-lasting changes in physical and biological systems (Pollack et al. 1998; Barnett et al. 2001; Peñuelas et al. 2002; Parmesan and Yohe 2003; Díaz-Almela et al. 2007; IPCC 2007). Climate change effects on cetaceans are at present largely unknown, but it is believed that effects would be mediated mainly through alteration of prey distribution and abundance and could shift cetacean habitat use, foraging strategies and grouping patterns (Lusseau et al. 2004; Learmonth et al. 2006; MacLeod 2009; Simmonds and Eliott 2009). An increase of carbon dioxide levels that results in ocean acidification has perhaps the most important effect on marine biodiversity, but it is not understood how or in what timeframe (Orr et al. 2005; Whitehead et al. 2008).
Of the several (and for the most part poorly understood) ways in which climate change may negatively affect marine mammal populations, perhaps most relevant to the Gulf of Mexico is a potential link between climate change and hurricane occurrence. The occurrence of hurricanes in the Gulf of Mexico is known to have increased over time, and this increase has been suggested to be one of the consequences of global warming (Emanuel 2005). While the impacts of hurricanes on marine mammals are poorly understood, they may range from death, injury, short-term displacements, habitat degradation, changes in prey occurrence, and health effects (medium and long term). For instance, hurricanes Rita, Katrina and Wilma reportedly resulted in the displacement of seven bottlenose dolphins into inland areas where they do not normally occur (Rosel and Watts 2008), and Florida manatees had lower survival probabilities in years with intense coastal storms (Langtimm and Beck 2003). However, hurricanes might also benefit marine mammal populations in ways that may balance the negative effects (e.g., the dramatically reduced fishing effort following hurricanes may increase prey availability) (Miller et al. 2010). An increase of bottlenose dolphin calves was recorded approximately 1.5 years after hurricane Katrina. This increase has been suggested to have been both a consequence of the decreased fishing effort by humans and the increase in the number of females that lost calves, and hence became reproductively receptive following the storm (Miller et al. 2010).
13.4.11 Strandings
Most strandings of single cetaceans in the Gulf are by bottlenose dolphins. It is not always possible to tell whether the live or dead animals come from the bay/estuary, nearshore, or offshore stocks. Most strandings are for undetermined reasons, although emaciation, disease, possible cold exposure, and other reasons not necessarily related to human interactions are documented (Southeast U.S. Marine Mammal Stranding Network 2011). Note in Table 13.25 below that the overwhelming number of single strandings did not have a determination of cause of death, due to decomposition, vague or absent indicators, or inexperience of personnel.
The number of total stranded marine mammals (1,392) does not equal the sum of the five categories listed above (totaling 1,475), because a number of strandings were included in more than one category (e.g., Human Interaction and Fisheries).
13.4.12 Global Status and U.S. Population Trends
Manatee causes of mortality are listed as a special case in Figure 13.61. Mortalities from speeding watercraft have been the highest overall source of mortality, but there is some indication that with greater education and enforcement, these are on the wane. Cold stress due to periodic shutdown of power plants used as refuges by manatees has also decreased in recent years, due to concerted human action. Many carcasses are too decomposed for accurate assessments of cause of death, and human causes are probably larger than indicated in Figure 13.61 and Table 13.26.

Causes of mortalities of Florida manatees (Trichechus manatus latirostris) (data from the Florida Fish and Wildlife Conservation Commission, 2010; drawn by Shane Blowes, and reproduced from Marsh et al. 2011).
13.5 Conclusions: Brief Summary of Present Knowledge
The 22 species of marine mammals of the Gulf of Mexico appear to be doing rather well, with no evidence of strong declines (or rapid increases) for any populations. Manatees and sperm whales are listed as endangered by the United States, but there is no evidence of declines in sperm whales, and manatees—while small in numbers in the Gulf—also appear now to be doing better than they were in the 1990s.
Determination of stocks (or biological populations) and numbers of marine mammals is not an exact science, except for one population of bottlenose dolphins in the Tampa Bay, Florida, area where almost each individual is known; aerial, shipboard, and acoustic censuses provide basic information on minimum numbers. However, there are broad variances (or estimates of error) depending on sighting factors, ease of acoustically identifying recorded animals, and chance of encounters with generally limited survey numbers due to costs involved. Broad variances are a common problem especially for cetacean studies in open oceans.
Much has been learned in the past 20 years about cetacean distribution in the Gulf. Bottlenose dolphins are ubiquitous in most bays that are well flushed by tides as well as along the outer shoreline. They also occur—along with Atlantic spotted dolphins—on the continental shelf in waters generally less than 200 m deep. In waters deeper than 200 m, the most commonly sighted species during multiple surveys, in descending orders of numbers, tend to be pantropical spotted, Clymene, spinner, and striped dolphins (all members of the Genus Stenella). Sperm whales occur generally in waters greater than 800 m deep, as do pygmy and dwarf sperm whales, pilot whales, and other members of the blackfish clade. Sperm whales occur throughout the U.S. EEZ, with a usual concentration within about 100 km south of Louisiana in an area strongly influenced by nutrient rich waters from the Mississippi River. Bryde’s whales, the only common baleen whale in the Gulf, occur mainly in shallower waters (100–200 m deep) of the northeastern Gulf, largely south of Alabama and the western part of the Florida Panhandle. There is evidence that deepwater cetaceans tend to be more numerous in or near cold-core cyclones and the confluence of cyclone/anticyclone eddy pairs where primary productivity and zooplankton to higher trophic levels also tend to be more abundant than in warm-core anticyclonic eddies.
While quite a few potential anthropogenic impacts have been identified for the Florida manatee—especially cold spells and boat collisions—and cetaceans, the major impacts are probably from chemical contamination, especially POPs that are known to decrease immune responses and increase reproductive disorders in mammals in general, with building evidence for cetaceans.
Open Access This chapter is distributed under the terms of the Creative Commons Attribution-Noncommercial 2.5 License (http://creativecommons.org/licenses/by-nc/2.5/) which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
The images or other third party material in this chapter are included in the work’s Creative Commons license, unless indicated otherwise in the credit line; if such material is not included in the work’s Creative Commons license and the respective action is not permitted by statutory regulation, users will need to obtain permission from the license holder to duplicate, adapt or reproduce the material.
References
Aguilar A, Borrell A (1994) Abnormally high polychlorinated biphenyl levels in striped dolphins (Stenella coeruleoalba) affected by the 1990–1992 Mediterranean epizootic. Sci Total Environ 154:237–247
Ames AL, Van Fleet ES (1996) Organochlorine residues in the Florida manatee, Trichechus manatus latirostris. Mar Pollut Bull 32:374–377
Anticamara JA, Watson R, Gelchu A, Pauly D (2011) Global fishing effort (1950–2010): Trends, gaps, and implications. Fish Res 10:131–136
Au WWL (1993) The sonar of dolphins. Springer-Verlag, New York, NY, USA. 277 p
Azzara A (2012) Impacts of vessel noise perturbations on the resident sperm whale population in the Gulf of Mexico. PhD Thesis. Texas A&M University, College Station, TX, USA. 104 p
Baird RW, Hooker SK (2000) Ingestion of plastic and unusual prey by a juvenile harbour porpoise. Mar Pollut Bull 40:719–720
Baird RW, Gorgone AM, McSweeney DJ, Webster DL, Salden DR, Deakos MH, Ligon AD, Schorr GS, Barlow J, Mahaffy SD (2008) False killer whales (Pseudorca crassidens) around the main Hawaiian Islands: Long-term site fidelity, inter-island movements, and association patterns. Mar Mamm Sci 24:591–612
Ballance LT (2009) Cetacean ecology. In: Perrin WF, Würsig B, Thewissen JGM (eds) Encyclopedia of marine mammals, 2nd edn. Academic/Elsevier, San Diego, CA, USA, pp 196–201
Balmer BC, Wells RS, Nowacek SM, Nowacek DP, Schwacke LH, McLellan WA, Scharf FS (2008) Seasonal abundance and distribution patterns of common bottlenose dolphins (Tursiops truncatus) near St. Joseph Bay, Florida, USA. J Cetacean Res Manag 10:157–167
Barlow J (2003) Preliminary estimates of the abundance of Cetaceans along the U.S. West Coast: 1991–2001. Southwest Fisheries Science Center Administrative Report LJ-03-03. La Jolla, CA, USA. 31 p
Barnett TP, Pierce DW, Schnur R (2001) Detection of anthropogenic climate change in the world's oceans. Science 292:270–274
Baumann-Pickering S, Roch MA, Wiggins SM, Schnitzler H-U, Hildebrand JA (2010) Classification of echolocation signals of melon-headed whales (Peponocephala electra), bottlenose dolphins (Tursiops truncatus), and Gray’s spinner dolphins (Stenella longirostris longirostris). J Acoust Soc Am 128:2212–2224
Baumgartner MF, Mullen KD, May LN, Leming TD (2001) Cetacean habitats in the northern Gulf of Mexico. Fish Bull 99:219–239
Bearzi G, Agazzi S, Gonzalvo J, Costa M, Bonizzoni S, Politi E, Piroddi C, Reeves RR (2008) Overfishing and the disappearance of short-beaked common dolphins from western Greece. Endang Species Res 5:1–12
Bearzi G, Reeves RR, Remonato E, Pierantonio N, Airoldi S (2011) Risso’s dolphin Grampus griseus in the Mediterranean Sea. Mamm Biol 76:385–400
Bigg MA, Olesiuk PF, Ellis GM, Ford JKB, Balcomb KC (1990) Social organization and genealogy of resident killer whales (Orcinus orca) in the coastal waters of British Columbia and Washington State. In: Hammond PS, Mizroch SA, Donovan GP (eds) Individual recognition of Cetaceans: Use of photoidentification and other techniques to estimate population parameters. Report of the International Whaling Commission Cambridge Special Issue 12:383–405
Blaylock RA, Hoggard W (1994) Preliminary estimates of bottlenose dolphin abundance in southern U.S. Atlantic and Gulf of Mexico continental shelf waters. NOAA Technical Memorandum NMFS-SEFSC-356. U.S. Department of Commerce, Washington, DC, USA. 10 p
Bräger S, Würsig B, Acevedo A, Henningsen T (1994) Association patterns of bottlenose dolphins (Tursiops truncatus) in Galveston Bay, Texas. J Mammal 75:431–437
Brouwer A, Reijnders PJH, Koeman JH (1989) Polychlorinated biphenyl (PCB) contaminated fish induces vitamin A and thyroid hormone deficiency in the common seal (Phoca vitulina). Aquat Toxicol 15:99–106
Bryan CE, Christopher SJ, Balmer BC, Wells RS (2007) Establishing baseline levels of trace elements in blood and skin of bottlenose dolphins in Sarasota Bay, Florida: Implications for noninvasive monitoring. Sci Total Environ 388:325–342
Bryant L (1994) Report to Congress on results of feeding wild dolphins: 1989–1994. NMFS (National Marine Fisheries Service), Office of Protected Resources, Washington, DC, USA. 22 p
Buckstaff KC (2004) Effects of watercraft noise on the acoustic behavior of bottlenose dolphins, Tursiops truncatus, in Sarasota Bay, Florida. Mar Mamm Sci 20:709–725
Burgess G, Morgan A (2003a) Commercial shark fishery observer program. Renewal of an observer program to monitor the directed commercial shark fishery in the Gulf of Mexico and South Atlantic: 1999 fishing season. Final Report. Highly Migratory Species Management Division Award NA97FF0041. U.S. National Marine Fisheries Service, Washington, DC, USA
Burgess G, Morgan A (2003b) Commercial shark fishery observer program. Renewal of an observer program to monitor the directed commercial shark fishery in the Gulf of Mexico and the South Atlantic: 2002(2) and 2003(1) fishing seasons. Final Report. Highly Migratory Species Management Division Award NA16FM0598. U.S. National Marine Fisheries Service, Washington, DC, USA
Caldwell D, Caldwell M (1973) Marine mammals of the eastern Gulf of Mexico. In: Jones JI, Ring RE, Rinkel MO, Smith RE III (eds) A summary of knowledge of the Eastern Gulf of Mexico. State University System of Florida, Gainesville, FL, USA, pp 1–3
Christmas JY, Waller RS (1975) Location and time of menhaden spawning in the Gulf of Mexico. National Marine Fisheries Report, Gulf Coast Research Laboratory, Ocean Springs, MS, USA. 20 p
Clark AH (1887) History and present condition of the fishery. In: Goode GB (ed) The fisheries and fishery industries of the United States, vol 2, Section V. History and methods of the fisheries, Part XV, The Whale fishery. U.S. Government Printing Office, Washington, DC, USA, pp 3–218
Coakes AK, Whitehead H (2004) Social structure and mating system of sperm whales off northern Chile. Can J Zool 82:1360–1369
Cockcroft VG, De Kock AC, Lord DA, Ross GJB (1989) Organochlorines in bottlenose dolphins Tursiops truncatus from the east coast of South Africa. S Afr J Mar Sci 8:207–217
Croll DA, Clark CW, Calambokidis J, Ellison WT, Tershy BR (2001) Effect of anthropogenic low-frequency noise on the foraging ecology of Balaenoptera whales. Anim Conserv 4:13–27
Crowder L, Hazen E, Avissar N, Bjorkland R, Latanich C, Ogburn M (2008) The impacts of fisheries on marine ecosystems and the transition to ecosystem-based management. Annu Rev Ecol Evol Syst 39:259–278
Cunningham-Smith P, Colbert DE, Wells RS, Speakman T (2006) Evaluation of human interactions with a provisioned wild bottlenose dolphin (Tursiops truncatus) near Sarasota Bay, Florida and efforts to curtail the interactions. Aquat Mamm 32:346–356
Davis JW (1993) An analysis of tissues for total PCB and planar PCB concentrations in marine mammals stranded along the Gulf of Mexico. MS Thesis. Texas A&M University, College Station, TX, USA. 139 p
Davis RW, Fargion GS (eds) (1996) Distribution and abundance of cetaceans in the north-central and western Gulf of Mexico. Final Report. MMS (Minerals Management Service), Vol 2. U.S. Department of the Interior, MMS, Gulf of Mexico OCS Region, New Orleans, LA, USA. 375 p
Davis RW, Fargion GS, May N, Leming TD, Baumgartner M, Evans WE, Hansen LJ, Mullin K (1998) Physical habitats of cetaceans along the Continental Slope in the northcentral and western Gulf of Mexico. Mar Mamm Sci 14:490–507
Davis RW, Ortega-Ortiz JG, Ribic CA, Evans WE, Biggs DC, Ressler PH, Cady RB, Leben RR, Mullin KD, Würsig B (2002) Cetacean habitat in the northern oceanic Gulf of Mexico. Deep-Sea Res Pt I 49:121–142
De Mutsert K, Cowan JH Jr, Essington TE, Hilborn R (2008) Reanalyses of Gulf of Mexico fisheries data: Landings can be misleading in assessments of fisheries and fisheries ecosystems. Proc Natl Acad Sci USA 105:2740–2744
Derraik JGB (2002) The pollution of the marine environment by plastic debris: A review. Mar Pollut Bull 44:842–852
Díaz-Almela E, Marbà N, Duarte CM (2007) Consequences of Mediterranean warming events in seagrass (Posidonia oceanica) flowering records. Global Change Biol 13:224–235
Duffield DA, Wells RS (2002) The molecular profile of a resident community of bottlenose dolphins, Tursiops truncatus. In: Pfeiffer CJ (ed) Cell and molecular biology of marine mammals. Krieger Publishing, Melbourne, FL, USA, pp 3–11
Emanuel K (2005) Increasing destructiveness of tropical cyclones over the past 30 years. Nature 436:686–688
Engelhaupt D, Hoelzel AR, Nicholson C, Frantzis A, Mesnick S, Gero S, Whitehead H, Rendell L, Miller P, De Stefanis R, Cañadas A, Airoldi S, Mignucci-Giannoni AA (2009) Female philopatry in coastal basins and male dispersion across the North Atlantic in a highly mobile marine species, the sperm whale (Physeter macrocephalus). Mol Ecol 18:4193–4205
Fertl D, Schiro AJ, Regan GT, Beck CA, Adimey N, Price-May L, Amos A, Worthy GAJ, Crossland R (2005) Manatee occurrence in the northern Gulf of Mexico west of Florida. Gulf Caribb Res 17:69–94
Fire SE, Fauquier D, Flewelling LJ, Henry M, Naar J, Pierce R, Wells RS (2007) Brevetoxin exposure in bottlenose dolphins (Tursiops truncatus) associated with Karenia brevis blooms in Sarasota Bay, Florida. Mar Biol 152:827–834
Fire SE, Flewelling LJ, Wang Z, Naar J, Henry MS, Pierce RH, Wells RS (2008) Florida red tide and brevetoxins: Association and exposure in live resident bottlenose dolphins (Tursiops truncatus) in the eastern Gulf of Mexico, USA. Mar Mamm Sci 24:831–844
Fire SE, Wang Z, Byrd M, Whitehead HR, Paternoster J, Morton SL (2011) Co-occurrence of multiple classes of harmful algal toxins in bottlenose dolphins (Tursiops truncatus) stranding during an unusual mortality event in Texas, USA. Harmful Algae 10:330–336
Fisher SJ, Reeves RR (2005) The global trade in live cetaceans: Implications for conservation. J Int Wildl Law Pol 8:315–340
Flewelling LJ, Naar JP, Abbott JP, Baden DG, Barros NB, Bossart GD, Bottein MJD, Hammond DG, Haubold EM, Heil CA, Henry MS, Jacocks HJ, Leighfield TA, Pierce RH, Pitchford TD, Rommel SA, Scott PS, Steidinger KA, Truby EW, Van Dolah FM, Landsberg JH (2005) Red tides and marine mammal mortalities. Nature 435:755–756
Ford JKB (2009) Killer whale, Orcinus orca. In: Perrin WF, Würsig B, Thewissen JGM (eds) Encyclopedia of marine mammals, 2nd edn. Academic/Elsevier Press, Amsterdam, The Netherlands, pp 650–657
Fritts TH, Irvine AB, Jennings RD, Collum LA, Hoffman W, McGehee MA (1983) Turtles, birds, and mammals in the northern Gulf of Mexico and nearby Atlantic waters. Final Report. U.S. Fish and Wildlife Service, Washington, DC, USA. 455 p
Galindo JA, Serrano A, Vásquez-Castán L, González-Gándara C, López-Ortega M (2009) Cetacean diversity, distribution, and abundance in Northern Veracruz, Mexico. Aquat Mamm 35:12–18
Garrison LP (2007) Interactions between marine mammals and pelagic longline fishing gear in the U.S. Atlantic Ocean between 1992 and 2004. Fish Bull 105:408–417
Garrison LP, Stokes L, Fairfield C (2009) Estimated bycatch of marine mammals and sea turtles in the U.S. Atlantic pelagic longline fleet during 2008. NOAA (National Oceanic and Atmospheric Administration) Technical Memorandum NMFS—SEFSC-591. Southeast Fisheries Science Center, NOAA, Washington, DC, USA. 63 p
Gauthier JM, Dubeau H, Rassart E, Jarman WM, Wells RS (1999) Biomarkers of DNA damage in marine mammals. Mutat Res 444:427–439
Geraci JR (1990) Physiologic and toxic effects on cetaceans. In: Geraci JR, St. Aubin DJ (eds) Sea mammals and oil: Confronting the risks. Academic Press, New York, NY, USA, pp 167–197
Gero S (2005) Fundamentals of sperm whale societies: Care for calves. MS Thesis, Dalhousie University, Halifax, Nova Scotia, Canada. 99 p
Gerrodette T, Forcada J (2002). Estimates of abundance of striped and common dolphins, and pilot, sperm and Bryde’s whales in the eastern tropical Pacific Ocean. SWFSC (Southwest Fisheries Science Center) Administrative Report, LJ-02-20. La Jolla, CA, USA. 24 p
Gerrodette T, Watters G, Forcada J (2005) Preliminary estimates of 2003 dolphin abundance in the eastern tropical Pacific. SWFSC Administrative Report, La Jolla, CA, USA. http://swfsc.nmfs.noaa.gov. Accessed 19 Feb 2013
Gonzalez-Socoloske D, Olivera-Gomez LD (2012) Gentle giants in dark waters: Using side-scan sonar for manatee research. Open Remote Sens J 5:1–14
Gowans S, Whitehead H, Hooker SK (2001) Social organization in northern bottlenose whales (Hyperoodon ampullatus): Not driven by deep water foraging? Anim Behav 62:369–377
Guinet C, Bouvier J (1995) Development of intentional stranding hunting techniques in killer whale Orcinus orca calves at Crozet Archipelago. Can J Zool 73:27–33
Gunter G (1941) Occurrence of the manatee in the United States, with records from Texas. J Mammal 22:60–64
Gunter G (1954) Mammals of the Gulf of Mexico. Fish Bull 55:543–551
Hale LF, Carlson JK (2007) Characterization of the shark bottom longline fishery: 2005–2006. Technical Memorandum NMFS-SEFSC-554. NOAA, Washington, DC, USA. 28 p
Hale LF, Hollensead LD, Carlson JK (2007) Characterization of the shark bottom longline fishery: 2007. Technical Memorandum NMFS-SEFSC-564. NOAA, Washington, DC, USA. 25 p
Hale LF, Gulak SJB, Carlson JK (2009) Characterization of the shark bottom longline fishery, 2008. Technical Memorandum NMFS-SEFSC-586. NOAA, Washington, DC, USA. 23 p
Hall AJ, McConnell BJ, Rowles TK, Aguilar A, Borrell A, Schwacke L, Reijnders PJ, Wells RS (2006) Individual-based model framework to assess population consequences of polychlorinated biphenyl exposure in bottlenose dolphins. Environ Health Perspect 114:60–64
Hartman DS (1979) Ecology and behavior of the manatee (Trichechus manatus) in Florida. Special Publication 5. American Society of Mammalogists, Lawrence, KA, USA. 153 p
Heimlich-Boran JR (1993) Social organization of the short-finned Pilot Whale, Globicephala macrorhynchus, with special reference to the comparative social ecology of Delphinids. PhD Thesis. University of Cambridge, Cambridge, UK. 197 p
Heithaus MR, Dill LM (2009) Feeding strategies and tactics. In: Perrin WF, Würsig B, Thewissen JGM (eds) Encyclopedia of marine mammals, 2nd edn. Elsevier Press, San Diego, CA, USA, pp 414–423
Heithaus M, Frid A, Wirsing A, Worm B (2008) Predicting ecological consequences of marine top predator declines. Trends Ecol Evol 23:202–210
Henningsen T, Würsig B (1991) Bottlenose dolphins in Galveston Bay, Texas: Numbers and activities. In: Proceedings of the European Society Conference, Cambridge University Press, Cambridge, UK, pp 36–38
Herzing DL (1997) The life history of free-ranging Atlantic spotted dolphins (Stenella frontalis): Age classes, color phases, and female reproduction. Mar Mamm Sci 13:576–595
Hess NA, Ribic CA (2000) Seabird ecology. In: Davis RW, Evans WE, Würsig B (eds) Cetaceans, Sea Turtles and Seabirds in the Northern Gulf of Mexico: Distribution, abundance and habitat associations, Vol 2, Technical Report. OCS Study MMS 2000-003. MMS, Gulf of Mexico OCS Region, New Orleans, LA, USA, pp 275–315
Hu C, Xiaofeng L, Pichel WG, Muller-Karger FE (2009) Detection of natural oil slicks in the NW Gulf of Mexico using MODIS imagery. Geophys Res Lett 36:1–5
IPCC (Intergovernmental Panel on Climate Change) (2007) Climate change 2007: The physical science basis. Summary for policy makers. WMO/UNEP Intergovernmental Panel on Climate Change. http://www.ipcc.ch. Accessed 19 Feb 2013
Irvine AB, Wells RS (1972) Results of attempts to tag Atlantic bottlenose dolphins (Tursiops truncatus). Cetology 13:1–5
Irvine AB, Scott MD, Wells RS, Kaufmann JH (1981) Movements and activities of the Atlantic bottlenose dolphin, Tursiops truncatus, near Sarasota, Florida. Fish Bull 79:671–688
Irwin LJ, Würsig B (2004) A small resident community of bottlenose dolphins, Tursiops truncatus, in Texas: Monitoring recommendations. Gulf Mex Sci 22:13–21
Jackson JBC, Kirby MX, Berger WH, Bjorndal KA, Botsford LW, Bourque BJ, Bradbury RH, Cooke R, Erlandson J, Estes JA, Hughes TP, Kidwell S, Lange CB, Lenihan HS, Pandolfi JM, Peterson CH, Steneck RS, Tegner MJ, Warner RR (2001) Historical overfishing and the recent collapse of coastal ecosystems. Science 293:629–638
Jaquet N, Gendron D (2009) The social organization of sperm whales in the Gulf of California and comparisons with other populations. J Mar Biol Assoc UK 89:975–983
Jefferson TA (1995) Distribution, abundance, and some aspects of the biology of cetaceans in the offshore Gulf of Mexico. PhD Thesis. Texas A&M University, College Station, TX, USA. 232 p
Jefferson TA, Schiro AJ (1997) Distribution of cetaceans in the offshore Gulf of Mexico. Mamm Rev 27:27–50
Jefferson TA, Webber M, Pitman R (2008) Marine mammals of the world: A comprehensive guide to their identification. Academic Press, London, UK. 573 p
Jepson PD, Bennett PM, Allchin CR, Law RJ, Kuiken T, Baker JR, Rogan E, Kirkwood JK (1999) Investigating potential associations between chronic exposure to polychlorinated biphenyls and infectious disease mortality in harbour porpoises from England and Wales. Sci Total Environ 243–244:339–348
Jochens A, Biggs D, Benoit-Bird K, Engelhaupt D, Gordon J, Hu C, Jaquet N, Johnson M, Leben R, Mate B, Miller P, Ortega-Ortiz J, Thode A, Tyack P, Würsig B (2008) Sperm whale seismic study in the Gulf of Mexico: Synthesis Report. OCS Study MMS 2008-006. U.S. Department of the Interior, New Orleans, LA, USA. 323 p
Karczmarski L, Würsig B, Gailey GA, Larson KW, Vanderlip C (2005) Spinner dolphins in a remote Hawaiian atoll: Social grouping and population structure. Behav Ecol 16:675–685
Kastelein RA, Lavaleije MSS (1992) Foreign bodies in the stomach of a female harbour porpoise (Phocoena phocoena) from the North Sea. Aquat Mamm 18:40–46
Kasuya T, Marsh H (1984) Life history and reproductive biology of the short–finned pilot whale, Globicephala macrorhynchus, off the coast of Japan. Rep Int Whal Comm Sp Issue 6:259–310
Kendall MD, Safieh B, Harwood J, Pomeroy PP (1992) Plasma thymulin concentrations, the thymus and organochlorine contamination levels in seals infected with phocine distemper virus. Sci Total Environ 115:133–144
Kucklick J, Schwacke L, Wells R, Hohn A, Guichard A, Yordy J, Hansen L, Zolman E, Wilson R, Litz J, Nowacek D, Rowles T, Pugh R, Balmer B, Sinclair C, Rosel P (2011) Bottlenose dolphins as indicators of persistent organic pollutants in the western North Atlantic Ocean and Northern Gulf of Mexico. Environ Sci Technol 45:4270–4277
Kuczaj SA II, Yeater DB (2007) Observations of rough-toothed dolphins (Steno bredanensis) off the coast of Utila, Honduras. J Mar Biol Assoc UK 87:141–148
Kuehl DW, Haebler R (1995) Organochlorine, organobromine, metal, and selenium residues in bottlenose dolphins (Tursiops truncatus) collected during an unusual mortality event in the Gulf of Mexico, 1990. Arch Environ Contam Toxicol 28:494–499
Kuiken T (1996) Diagnosis of by-catch in cetaceans. In: Proceedings, 2nd European Cetacean Society Workshop on Cetacean Pathology (European Cetacean Society Special Issue 26). Montpellier, France, 2 March 1994, pp 1–43
Küss KM (1998) The occurrence of PCBs and chlorinated pesticide contaminants in bottlenose dolphins in a resident community: comparison with age, gender and birth order. MS Thesis. Nova Southeastern University, Fort Lauderdale, FL, USA
Kvenvolden KA, Cooper CK (2003) Natural seepage of crude oil into the marine environment. Geo Mar Lett 23:140–146
Lahvis GP, Wells RS, Kuehl DW, Stewart JL, Rhinehart HL, Via CS (1995) Decreased lymphocyte responses in free-ranging bottlenose dolphins (Tursiops truncatus) are associated with increased concentrations of PCBs and DDT in peripheral blood. Environ Health Perspect 103:67–72
Laist DW, Reynolds JE III (2005) Influence of power plants and other warm-water refuges on Florida manatees. Mar Mamm Sci 21:739–764
Laist DW, Coe JM, O’Hara KJ (1999) Marine debris pollution. In: Twiss JR, Reeves RR (eds) Conservation and management of marine mammals. Smithsonian Institution Press, Washington, DC, USA, pp 342–366
Laist DW, Knowlton AR, Mead JG, Collet AS, Podestà M (2001) Collisions between ships and whales. Mar Mamm Sci 17:35–75
Langtimm C, Beck C (2003) Lower survival probabilities for adult Florida manatees in years with intense coastal storms. Ecol Appl 13:257–268
Layne JN (1965) Observations on marine mammals in Florida waters. Bull Florida State Mus 9:131–181
Learmonth JA, MacLeod CD, Santos MB, Pierce GJ, Crick HQP, Robinson RA (2006) Potential effects of climate change on marine mammals. Oceanogr Mar Biol Ann Rev 44:431–464
Lee RF, Anderson JW (2005) Significance of cytochrome P450 system responses and levels of bile fluorescent aromatic compounds in marine wildlife following oil spills. Mar Pollut Bull 50:705–723
Lewison RL, Crowder LB, Read AJ, Freeman SA (2004) Understanding impacts of fisheries bycatch on marine megafauna. Trends Ecol Evol 19:598–604
Litz JA, Garrison LP, Fieber LA, Martinez A, Contillo JP, Kucklick JR (2007) Fine-scale spatial variation of persistent organic pollutants in bottlenose dolphins (Tursiops truncatus) in Biscayne Bay, FL. Environ Sci Technol 41:7222–7228
Loughlin TR (1994) Marine mammals and the Exxon Valdez. Academic Press, San Diego, CA, USA. 395 p
Lowery GH (1974) The mammals of Louisiana and its adjacent waters. Louisiana State University Press, Baton Rouge, LA, USA. 565 p
Lusseau D, Williams R, Wilson B, Grellier K, Barton TR, Hammond PS, Thompson PM (2004) Parallel influence of climate on the behaviour of Pacific killer whales and Atlantic bottlenose dolphins. Ecol Lett 7:1068–1076
Lynn SK, Würsig B (2002) Summer movement patterns of bottlenose dolphins in a Texas Bay. Gulf Mex Sci 20:25–37
MacLeod CD (2009) Global climate change, range changes and potential implications for the conservation of marine cetaceans: A review and synthesis. Endang Spec Res 7:125–136
Marino L (2004) Cetacean brain evolution: Multiplication generates complexity. Int J Comp Psychol 17:1–16
Marsh H, O’Shea TJ, Reynolds JE III (2011) Ecology and conservation of the Sirenia. Dugongs and Manatees. Cambridge University Press, Cambridge, UK. 536 p
Martínez-Serrano I, Serrano A, Heckel G, Schramm Y (2011) Distribution and home range of bottlenose dolphins (Tursiops truncatus) off Veracruz, Mexico. Cienc Mar 37(4A):379–392
Maze KS (1997) Bottlenose dolphins of San Luis Pass, Texas: Occurrence patterns, site fidelity, and habitat use. MS Thesis. Texas A&M University, College Station, TX, USA. 79 p
Maze KS, Würsig B (1999) Bottlenose dolphins of San Luis Pass, Texas: Occurrence patterns, site fidelity, and habitat use. Aquat Mamm 25:91–103
Maze-Foley K, Mullin KD (2006) Cetaceans of the oceanic northern Gulf of Mexico: Distributions, group sizes and interspecific associations. J Cetacean Res Manage 8:203–213
McCarthy E (2004) International regulation of underwater sound: Establishing rules and standards to address ocean noise pollution. Kluwer Academic Press, Dordrecht, The Netherlands. 287 p
McHugh KA, Allen JB, Barleycorn AA, Wells RS (2010) Severe Karenia brevis red tides influence juvenile bottlenose dolphins (Tursiops truncatus) behavior in Sarasota Bay, Florida. Mar Mamm Sci 27:622–643
Melcón ML, Cummins AJ, Kerosky SM, Roche LK, Wiggins SM, Hildebrand JA (2012) Blue whales respond to anthropogenic noise. PLoS One 7(2):e32681
Miller C (2003) Abundance trends and environmental habitat usage patterns of bottlenose dolphins (Tursiops truncatus) in lower Barataria and Caminada Bays, Louisiana. PhD Thesis. Louisiana State University, Baton Rouge, LA, USA. 125 p
Miller LJ, Solangi M, Kuczaj SA II (2008) Immediate response of Atlantic bottlenose dolphins to high speed personal watercraft in the Mississippi Sound. J Mar Biol Assoc UK 88:1139–1143
Miller PJO, Johnson MP, Madsen PT, Biassoni N, Quero M, Tyack PL (2009) Using at-sea experiments to study the effects of airguns on the foraging behavior of sperm whales in the Gulf of Mexico. Deep-Sea Res 56:1168–1181
Miller LJ, Mackey AD, Hoffland T, Solangi M, Kuczaj SA II (2010) Potential effects of a major hurricane on Atlantic bottlenose dolphins (Tursiops truncatus) reproduction in the Mississippi Sound. Mar Mamm Sci 26:707–715
Moore JC (1953) Distribution of marine mammals to Florida waters. Am Midl Nat 49:117–158
Moreno MPT (2005) Environmental predictors of bottlenose dolphin distribution and core feeding densities in Galveston Bay, Texas. PhD Thesis. Texas A&M University, College Station, TX, USA. 88 p
Mullin KD (1988) Comparative seasonal abundance and ecology of bottlenose dolphins (Tursiops truncatus) in three habitats of the north-central Gulf of Mexico. PhD Thesis. Mississippi State University, Starkville, MS, USA. 135 p
Mullin KD (2007) Abundance of cetaceans in the oceanic Gulf of Mexico based on 2003–2004 ship surveys. NMFS, SEFSC, Pascagoula, LA, USA. 26 p
Mullin KD, Fulling GL (2004) Abundance of cetaceans in the oceanic northern Gulf of Mexico. Mar Mamm Sci 20:787–807
Mullin KD, Hoggard W (2000) Visual surveys of cetaceans and sea turtles from aircraft and ships. In: Davis RW, Evans WE, Würsig B (eds) Cetaceans, Sea Turtles, and Seabirds in the Northern Gulf of Mexico: Distribution, abundance and habitat associations, vol 2. Technical Report. OCS Study MMS 96-0027. MMS, Gulf of Mexico OCS Region, New Orleans, LA, USA, pp 111–172
Mullin K, Hoggard W, Roden C, Lohoefener R, Rogers C, Taggart B (1994) Cetaceans on the upper continental slope in the north-central Gulf of Mexico. Fish Bull 92:773–786
Mullin KD, Hoggard W, Hansen LJ (2004) Abundance and seasonal occurrence of cetaceans in outer continental shelf and slope waters of the north-central and northwestern Gulf of Mexico. Gulf Mex Sci 22:62–73
Murphy S, Pierce GJ, Law RL, Bersuder P, Jepson PD, Learmonth JA, Addink M, Dabin W, Santos MB, Deaville R, Zegers BN, Mets A, Rogan E, Ridoux V, Reid RJ, Smeenk C, Jauniaux T, López A, Alonso Farré JM, González AF, Guerra A, García-Hartmann M, Lockyer C, Boon JP (2010) Assessing the effect of persistent organic pollutants on reproductive activity in common dolphins and harbour porpoises. J Northw Atl Fish Sci 42:153–173
NASA/Goddard Space Flight Center–EOS Project Science Office (2000) Scientists find that tons of oil seep into the Gulf of Mexico each year. ScienceDaily. http://www.sciencedaily.com/releases/2000/01/000127082228.htm. Accessed 3 Apr 2013
National Research Council (NRC) (2003) Oil in the Sea III: Inputs, fates, and effects. The National Academies Press, Washington, DC, USA. 280 p
Neff JM (1990) Composition and fate of petroleum and spill treating agents in the marine environment. In: Geraci JR, St. Aubin DJ (eds) Sea mammals and oil: confronting the risks. Academic Press, New York, NY, USA. 259 p
Newman SJ, Smith SA (2006) Marine mammal neoplasia: A review. Vet Pathol 43:865–880
NOAA (National Oceanic and Atmospheric Administration) (2004) Interim report on the bottlenose dolphin (Tursiops truncatus) unusual mortality event along the panhandle of Florida March–April 2004. NOAA Fisheries Office of Protected Resources, Washington, DC, USA. 36 p
Norris KS, Dohl TP (1980) Behavior of the Hawaiian spinner dolphin, Stenella longirostris. Fish Bull 77:821–849
Norris KS, Würsig B, Wells RS, Würsig M (1994) The Hawaiian Spinner Dolphin. University of California Press, Berkeley, CA, USA. 436 p
Nowacek SM, Wells RS, Solow AR (2001) Short-term effects of boat traffic on bottlenose dolphins, Tursiops truncatus, in Sarasota Bay, Florida. Mar Mamm Sci 17:673–688
Nowacek SM, Wells RS, Owen ECG, Speakman TR, Flamm RO, Nowacek DP (2004) Florida Manatees, Trichechus manatus latirostris, respond to approaching vessels. Biol Conserv 119:517–523
Nowacek DP, Thorne LH, Johnston DW, Tyack PL (2007) Responses of cetaceans to anthropogenic noise. Mamm Rev 37:81–111
O'Hara TM, O'Shea TJ (2001) Toxicology. In: Dierauf LS, Gulland FMD (eds) CRC handbook of marine mammal medicine, 2nd edn. CRC Press, Boca Raton, FL, USA, pp 471–520
O’Hern J, Biggs DC (2009) Sperm whale (Physeter macrocephalus) habitat in the northern Gulf of Mexico: Satellite observed ocean color and altimetry applied to small-scale variability in distribution. Aquat Mamm 35:358–366
O’Shea TJ, Moore JF, Kochman HI (1984) Contaminant concentrations in manatees in Florida. J Wildl Manage 48:741–748
O’Shea TJ, Ackerman BB, Percival HF (1995) Population biology of the Florida manatee: An overview. In: O’Shea TJ, Ackerman BB, Percival HF (eds) Population biology of the Florida manatee. National Biological Service Information and Technology Report I. Department of the Interior, Washington, DC, USA, pp 280–287
O’Shea TJ, Reeves RR, Long AK (1999) Marine mammals and persistent ocean contaminants. In: Proceedings of the Marine Mammal Commission Workshop, Keystone, 12–15 October 1998. Marine Mammal Commission, Bethesda, MD, USA. 150 p
O’Sullivan S, Mullin KD (1997) Killer whales (Orcinus orca) in the northern Gulf of Mexico. Mar Mamm Sci 13:141–147
Orr JC, Fabry VJ, Aumont O, Bopp L, Doney SC, Feely RA, Gnanadesikan A, Gruber N, Ishida A, Joos F, Key RM, Lindsay K, Maier-Reimer E, Matear R, Monfray P, Mouchet A, Najjar RG, Plattner G-K, Rodgers KB, Sabine CL, Sarmiento JL, Schlitzer R, Slater RD, Totterdell IJ, Weirig M-F, Yamanaka Y, Yool A (2005) Anthropogenic ocean acidification over the twenty-first century and its impact on calcifying organisms. Nature 437:681–686
Ortega-Argueta A, Perez-Sanchez CE, Gordillo-Morales G, Gordillo OG, Perez DG, Alafita H (2005) Cetacean strandings on the southwestern coast of the Gulf of Mexico. Gulf Mex Sci 23:179–185
Ortega-Ortiz JG (2002) Multiscale analysis of cetacean distribution in the Gulf of Mexico. PhD Thesis. Texas A&M University, College Station, TX, USA. 170 p
Ortega-Ortiz JG, Engelhaupt D, Winsor M, Mate BR, Hoelzel AR (2012) Kinship of long-term associates in the highly social sperm whale. Mol Ecol 21:732–744
Panigada S, Pesante G, Zanardelli M, Capoulade F, Gannier A, Weinrich M (2006) Mediterranean fin whales at risk from fatal ship strikes. Mar Pollut Bull 52:1287–1298
Parmesan C, Yohe G (2003) A globally coherent fingerprint of climate change impacts across natural systems. Nature 421:37–42
Pauly D, Palomares ML (2005) Fishing down marine food web: It is far more pervasive then we thought. Bull Mar Sci 76:197–211
Pauly D, Christensen V, Dalsgaard J, Froese R, Torres F Jr (1998) Fishing down marine food webs. Science 279:860–863
Pauly D, Christensen V, Guénette S, Pitcher TJ, Sumaila UR, Walters CJ (2002) Towards sustainability in world fisheries. Nature 418:689–695
Payne R, Webb D (1971) Orientation by means of long range acoustic signaling in baleen whales. Ann NY Acad Sci 188:110–142
Peñuelas J, Filella I, Comas PE (2002) Changed plant and animal life cycles from 1952 to 2000 in the Mediterranean region. Global Change Biol 8:531–544
Pérez-Cao H (2004) Abundance and distribution of the bottlenose dolphin Tursiops truncatus (Montagu, 1821) in two areas of the Sabana-Camagüey Archipelago, Cuba. MS Thesis. National Aquarium of Cuba, Havana, Cuba. 67 p. (English translation)
Perrin WF, Würsig B, Thewissen JGM (2009) Encyclopedia of marine mammals, 2nd edn. Academic Press, Amsterdam, The Netherlands. 1352 p
Pollack HN, Huang S, Shen PY (1998) Climate change record in subsurface temperatures: A global perspective. Science 282:279–281
Poncelet E, Van Canneyt O, Boubert JJ (2000) Considerable amount of plastic debris in the stomach of a Cuvier's beaked whale (Ziphius cavirostris) washed ashore on the French Atlantic coast. Eur Res Cetaceans 14:44–47
Powell JR, Wells RS (2011) Recreational fishing depredation and associated behaviors involving common bottlenose dolphins (Tursiops truncatus) in Sarasota Bay, Florida. Mar Mamm Sci 27:111–129
Pryor KW, Haag R, O’Reilly J (1969) The creative porpoise: Training for novel behavior. J Exp Anal Behav 12:653–661
Pulster EL, Maruya KA (2008) Geographic specificity of Aroclor 1268 in bottlenose dolphins (Tursiops truncatus) frequenting the Turtle/Brunswick River Estuary, Georgia (USA). Sci Total Environ 393:367–375
Rawson AJ, Patton GW, Hofmann S, Pietra GG (1993) Liver abnormalities associated with chronic mercury accumulation in stranded Atlantic bottlenose dolphins. Ecotoxicol Environ Saf 25:41–47
Rawson AJ, Bradley JP, Teetsov A, Rice S, Haller EM, Patton GW (1995) A role for airborne particulate in high mercury levels of some cetaceans. Ecotoxicol Environ Saf 30:309–314
Read A (2008) The looming crisis: Interactions between marine mammals and fisheries. J Mammal 89:541–548
Read AJ, Murray KT (2000) Gross evidence of human-induced mortality in small cetaceans. NOAA Technical Memorandum NMFS-OPR-15. NOAA, Washington, DC, USA. 20 p
Reddy ML, Reif JS, Bachand A, Ridgway SH (2001) Opportunities for using navy marine mammals to explore associations between organochlorine contaminants and unfavorable effects on reproduction. Sci Total Environ 274:171–182
Reeves RR, Lund JN, Smith TD, Josephson EA (2011) Insights from whaling logbooks on whales, dolphins, and whaling in the Gulf of Mexico. Gulf Mex Sci 29:41–67
Reynolds JE III (1985) Evaluation of the nature and magnitude of interactions between bottlenose dolphins, Tursiops truncatus, and fisheries and other human activities in areas of the southeastern United States. PB86-162203. National Technical Information Service, Springfield, VA, USA
Reynolds JE III, Odell DK (1991) Manatees and Dugongs. Facts on file, New York, NY, USA. 192 p
Reynolds JE III, Wells RS, Eide SD (2000) The bottlenose dolphin: Biology and conservation. University Press of Florida, Gainesville, FL, USA. 304 p
Richards PM (2007) Estimated takes of protected species in the commercial directed shark bottom longline fishery 2003, 2004, and 2005. NMFS/SEFSC Contribution PRD-06/07-08. U.S. National Marine Fisheries Service, Washington, DC, USA. 21 p
Richter C, Gordon J, Jaquet N, Würsig B (2008) Social structure of sperm whales in the northern Gulf of Mexico. Gulf Mex Sci 26:118–123
Rolland RM, Parks SE, Hunt KE, Castellote M, Corkeron PJ, Nowacek DP, Wasser SK, Kraus SD (2012) Evidence that ship noise increases stress in right whales. Proc R Soc B 279:2363–2368
Rosel PE, Watts H (2008) Hurricane impacts on bottlenose dolphins in the northern Gulf of Mexico. Gulf Mex Sci 25:88–94
Ross PS (2006) Fireproof killer whales (Orcinus orca): Flame-retardant chemicals and the conservation imperative in the charismatic icon of British Columbia, Canada. Can J Fish Aquat Sci 63:224–234
Ross PS, Birnbaum LS (2003) Integrated human and ecological risk assessment: A case study of persistent organic pollutants (POPs) in humans and wildlife. Hum Ecol Risk Assess 9:303–324
Rossi-Santos MR, Santos-Neto E, Baracho CG (2009) Interspecific cetacean interactions during the breeding season of humpback whale (Megaptera novaeangliae) on the north coast of Bahia State, Brazil. J Mar Biol Assoc UK 89:961–966
Salata G, Wade TL, Sericano JL, Davis JW, Brooks JM (1995) Analysis of Gulf of Mexico bottlenose dolphins for organochlorine pesticides and PCBs. Environ Pollut 88:167–175
Samuels A, Bejder L (2004) Chronic interaction between humans and free-ranging bottlenose dolphins near Panama City Beach, Florida, USA. J Cetacean Res Manage 6:69–77
Schick RS, Halpin PN, Read AJ, Urban DL, Best BD, Good CP, Roberts JJ, LaBrecque EA, Dunn C, Garrison LP, Hyrenbach KD, McLellan WA, Pabst DA, Palka DL, Stevick P (2011) Community structure in pelagic marine mammals at large spatial scales. Mar Ecol Prog Ser 434:165–181
Schmidly DJ (1981) Marine mammals of the southeastern United States and the Gulf of Mexico. FWS/OBS-80/41. U.S. Fish and Wildlife Service, Washington, DC, USA. 165 p
Schmidly DJ, Shane SH (1978) A biological assessment of the cetacean fauna of the Texas coast. Nat Tech Inf Ser PB-281-763. National Technical Information Service, Springfield, VA, USA. 38 p
Schmidly DJ, Würsig B (2009) Mammals (Vertebrata: Mammalia) of the Gulf of Mexico. In: Felder DL, Camp DK (eds) Gulf of Mexico—origins, waters, and biota biodiversity. Texas A&M Press, College Station, TX, USA, pp 1343–1353
Schwacke LH, Voit EO, Hansen LJ, Wells RS, Mitchum GB, Hohn AA, Fair PA (2002) Probabilistic risk assessment of reproductive effects of polychlorinated biphenyls on bottlenose dolphins (Tursiops truncatus) from the southeast United States coast. Environ Toxicol Chem 21:2752–2764
Schwacke LH, Twiner MJ, De Guise S, Balmer BC, Wells RS, Townsend FI, Rotstein DC, Varela RA, Hansen LJ, Zolman ES, Spradlin TR, Levin M, Leibrecht H, Wang Z, Rowles TK (2010) Eosinophilia and biotoxin exposure in bottlenose dolphins (Tursiops truncatus) from a coastal area impacted by repeated mortality events. Environ Res 110:548–555
Scott GP, Burn DM, Hansen LJ, Owen RE (1989) Estimates of bottlenose dolphin abundance in the Gulf of Mexico from regional aerial surveys. National Marine Fishery Service, Southeast Fisheries Science Center, Miami, FL, USA. 24 p
Scott MD, Wells RS, Irvine AB (1990) A long-term study of bottlenose dolphins on the west coast of Florida. In: Leatherwood S, Reeves RR (eds) The bottlenose dolphin. Academic Press, San Diego, CA, USA, pp 235–244
Shane SH (1977) The population biology of the Atlantic bottlenose dolphin, Tursiops truncatus, in the Aransas Pass area of Texas. MS Thesis. Texas A&M University, College Station, TX, USA. 238 p
Shane SH (1994) Occurrence and habitats use of marine mammals at Santa Catalina Island, California from 1983–91. Bull Calif Acad Sci 93:13–29
Shane SH (2004) Residence patterns, group characteristics, and association patterns of bottlenose dolphins near Sanibel Island, Florida. Gulf Mex Sci 2004:1–12
Shane SHR, Wells RS, Würsig B (1986) Ecology, behavior, and social organization of the bottlenose dolphin: A review. Mar Mamm Sci 2:34–63
Simmonds MP, Eliott WJ (2009) Climate change and cetaceans: Concerns and recent developments. J Mar Biol Assoc UK 89:203–210
Smultea MA, Würsig B (1995) Behavioral reactions of bottlenose dolphins to the Mega Borg II oil spill, Gulf of Mexico 1990. Aquat Mamm 21:171–181
Southall BL, Bowles AE, Ellison WT, Finneran JJ, Gentry RL, Greene CR Jr, Kastak D, Ketten DR, Miller JH, Nachtigall PE, Richardson WJ, Thomas JT, Tyack PL (2007) Marine mammal noise exposure criteria: Initial scientific recommendations. Aquat Mamm 33:411–521
Southeast U.S. Marine Mammal Stranding Network—NMFS Southeast Region Stranding Database (2011) Office of Protected Resources, NOAA. http://www.nmfs.noaa.gov/pr/health/networks.htm#southeast. Accessed 3 Apr 2013
Steichen DJ Jr, Holbrook SJ, Osenberg CW (1996) Distribution and abundance of benthic and demersal macrofauna within a natural hydrocarbon seep. Mar Ecol Prog Ser 138:71–82
Stensland E, Angerbjörn A, Berggren P (2003) Mixed species groups in mammals. Mamm Rev 33:205–223
Sturges W, Leben R (2000) Frequency of ring separations from the loop current in the Gulf of Mexico: A revised estimate. J Phys Oceanogr 30:1814–1819
Swartz W, Sala E, Tracey S, Watson R, Pauly D (2010) The spatial expansion and ecological footprint of fisheries (1950 to present). PLoS One 5(12):e15143
Tarpley RJ, Marwitz S (1993) Plastic debris ingestion by cetaceans along the Texas coast: Two case reports. Aquat Mamm 19:93–98
Townsend CH (1935) The distribution of certain whales as shown by logbook records of American whale ships. Zoologica 19:1–50
Twiner MJ, Fire S, Schwacke L, Davidson L, Wang Z, Morton S, Roth S, Balmer B, Rowles TK, Wells RS (2011) Concurrent exposure of bottlenose dolphins (Tursiops truncatus) to multiple algal toxins in Sarasota Bay, Florida, USA. PLoS One 6:1–15
Tyson RB (2008) Abundance of bottlenose dolphins (Tursiops truncatus) in the Big Bend of Florida, St. Vincent Sound to Alligator Harbor. MS Thesis. Florida State University, Tallahassee, FL, USA. 74 p
Urian KW, Duffield DA, Read AJ, Wells RS, Shell ED (1996) Seasonality of reproduction in bottlenose dolphins, Tursiops truncatus. J Mammal 77:394–403
Urian KW, Hofmann S, Wells RS, Read AJ (2009) Fine-scale population structure of bottlenose dolphins (Tursiops truncatus) in Tampa Bay, Florida. Mar Mamm Sci 25:619–638
Valdes-Arellanes MP, Serrano A, Heckel G, Schramm Y, Martínez-Serrano I (2011) Abundancia de dos poblaciones de toninas (Tursiops truncatus) en el norte de Veracruz, México. Rev Mex Biodiv 82:227–235
Van Waerebeek K, Sequeira M, Williamson C, Sanino GP, Gallego P, Carmo P (2006) Live-captures of common bottlenose dolphins Tursiops truncatus and unassessed bycatch in Cuban waters: evidence of sustainability found wanting. Latin Am J Aquat Mamm 5:39–48
Van Waerebeek KA, Baker N, Félix F, Gedamke J, Iñiguez M, Sanino GP, Secchi E, Sutaria D, Van Helden A, Wang Y (2007) Vessel collisions with small cetaceans worldwide and with large whales in the southern hemisphere, an initial assessment. Latin Am J Aquat Mamm 6:43–69
Vedder JM (1996) Levels of organochlorine contaminants in milk relative to health of bottlenose dolphins (Tursiops truncatus) from Sarasota, Florida. MS Thesis. University of California, Santa Cruz, CA, USA
Vidal O, Van Waerebeek K, Fidley LT (1994) Cetaceans and gillnet fisheries in Mexico, Central America, and the wider Caribbean: A preliminary review. In: Perrin WF, Donovan GP, Barlow J (eds) Gillnets and cetaceans, Report of the International Whaling Commission Special Issue, 15:221–233
Wade PR (1998) Calculating limits to the allowable human-coast mortality of cetaceans and pinnipeds. Mar Mamm Sci 14:1–37
Wade PR, Gerrodette T (1993) Estimates of cetacean abundance and distribution in the Eastern Tropical Pacific. Rep Int Whal Comm 43:477–493
Waring GT, Josephson E, Maze-Foley K, Rosel PE (2010) U.S. Atlantic and Gulf of Mexico Marine Mammal Stock Assessments 2010. NOAA Technical Memorandum NMFS-NE-219. http://www.nefsc.noaa.gov/nefsc/publications/ (original publication Dec. 2010). Accessed 3 Apr 2013
Waring GT, Josephson E, Maze-Foley K, Rosel PE (2011) U.S. Atlantic and Gulf of Mexico Marine Mammal Stock Assessments, 2011. NOAA Technical Memorandum NMFS-NE-221. http://www.nefsc.noaa.gov/nefsc/publications/ (Original publication Dec. 2011). Accessed 3 Apr 2012
Waring GT, Josephson E, Maze-Foley K, Rosel PE (eds) (2013) U.S. Atlantic and Gulf of Mexico Marine Mammal Stock Assessments—2012. NOAA Tech Memo NMFS NE 223, National Marine Fisheries Service, Woods Hole, MA, USA. 419 p. http://www.nefsc.noaa.gov/nefsc/publications/. Accessed 3 Jun 2013
Weilgart L, Whitehead H, Payne K (1996) A colossal convergence. Am Sci 84:278–287
Weller D, Würsig B, Whitehead H, Norris J, Lynn S, Davis R, Clauss N, Brown P (1996) Observations of an interaction between sperm whales and short-finned pilot whales in the Gulf of Mexico. Mar Mamm Sci 12:588–593
Wells RS (1991) The role of long-term study in understanding the social structure of a bottlenose dolphin community. In: Pryor K, Norris KS (eds) Dolphin societies: Discoveries and puzzles. University of California Press, Berkeley, CA, USA, pp 199–225
Wells RS (2003) Dolphin social complexity: Lessons from long-term study and life history. In: de Waal FBM, Tyack PL (eds) Animal social complexity: Intelligence, culture, and individualized societies. Harvard University Press, Cambridge, MA, USA, pp 32–56
Wells RS, Scott MD (1990) Estimating bottlenose dolphin population parameters from individual identification and capture-release techniques. Rep Int Whal Comm Sp Issue 14:407–415
Wells RS, Scott MD (1994) Incidence of gear entanglement for resident inshore bottlenose dolphins near Sarasota, FL. In: Perrin WF, Donovan GP, Barlow J (eds) Gillnets and Cetaceans. Report of the International Whaling Commission Special Issue 15. IWC Press, Cambridge, UK. 629 p
Wells RS, Scott MD (1997) Seasonal incidence of boat strikes on bottlenose dolphins near Sarasota, Florida. Mar Mamm Sci 13:475–480
Wells RS, Scott MD (2009) Common bottlenose dolphin Tursiops truncatus. In: Perrin WF, Würsig B, Thewissen JGM (eds) Encyclopedia of marine mammals, 2nd edn. Elsevier, Amsterdam, The Netherlands, pp 249–255
Wells RS, Scott MD, Irvine AB (1987) The social structure of free-ranging bottlenose dolphins. In: Genoways HH (ed) Current mammalogy, vol 1. Plenum Press, New York, NY, USA, pp 247–305
Wells RS, Hofmann S, Moors TL (1998) Entanglement and mortality of bottlenose dolphins, Tursiops truncatus, in recreational fishing gear in Florida. Fish Bull 96:647–650
Wells RS, Duffield D, Hohn AA (2001) Reproductive success of free ranging bottlenose dolphins: Experience and size make a difference. In: 14th Biennial Conference on the Biology of Marine Mammals, Vancouver, Canada, 28 November–3 December 2001. 231 p
Wells RS, Rowles TK, Borrell A, Aguilar A, Rhinehart HL, Jarman WM, Hofmann S, Hohn AA, Duffield DA, Mitchum G, Stott J, Hall A, Sweeney JC (2003) Integrating data on life history, health, and reproductive success to examine potential effects of POPs on bottlenose dolphins (Tursiops truncatus) in Sarasota Bay, FL. Organohalogen Compd 62:208–211
Wells RS, Rhinehart HL, Hansen LJ, Sweeney JC, Townsend FI, Stone R, Casper DR, Scott MD, Hohn AA, Rowles TK (2004) Bottlenose dolphins as marine ecosystem sentinels: Developing a health monitoring system. Ecosyst Health 1:246–254
Wells RS, Tornero V, Borrell A, Aguilar A, Rowles TK, Rhinehart HL, Hofmann S, Jarman WM, Hohn AA, Sweeney JC (2005) Integrating life-history and reproductive success data to examine potential relationships with organochlorine compounds for bottlenose dolphins (Tursiops truncatus) in Sarasota Bay, Florida. Sci Total Environ 349:106–119
Wells RS, Allen JB, Hofmann S, Bassos-Hill K, Fauquier DA, Barros NB, DeLynn RE, Sutton G, Socha V, Scott MD (2008) Consequences of injuries on survival and reproduction of common bottlenose dolphins (Tursiops truncatus) along the west coast of Florida. Mar Mamm Sci 24:774–794
Whitehead H (1998) Cultural selection and genetic diversity in matrilineal whales. Science 282:1708–1711
Whitehead H (2003) Sperm whales: Social evolution in the ocean. University of Chicago Press, Chicago, IL, USA. 431 p
Whitehead H (2009) Estimating abundance from one-dimensional passive acoustic surveys. J Wildl Manage 73:1000–1009
Whitehead H, McGill B, Worm B (2008) Diversity of deep-water cetaceans in relation to temperature: Implications for ocean warming. Ecol Lett 11:1198–1207
Williams M, Davis RW (1995) Emergency care and rehabilitation of oiled sea otters: A guide for oil spills involving fur-bearing marine mammals. University of Alaska Press, Fairbanks, AK, USA. 300 p
Wolfe DA (1987) Persistent plastics and debris in the ocean: An international problem of ocean disposal. Mar Pollut Bull 18:303–305
Worm B, Barbier EB, Beaumont N, Duffy JE, Folke C, Halpern BS, Jackson JBC, Lotze HK, Micheli F, Palumbi SR, Sala E, Selkoe KA, Stachowicz JJ, Watson R (2006) Impacts of biodiversity loss on ocean ecosystem services. Science 314:787–790
Worm B, Hilborn R, Baum JK, Branch TA, Collie JS, Costello C, Fogarty MJ, Fulton EA, Hutchings JA, Jennings S, Jensen OP, Lotze HK, Mace PM, McClanahan TR, Minto C, Palumbi SR, Parma AM, Ricard D, Rosenberg AA, Watson R, Zeller D (2009) Rebuilding global fisheries. Science 325:578–584
Wormuth JH, Ressler PH, Cady RB, Harris EJ (2000) Zooplankton and micronekton in cyclones and anticyclones in the northeast Gulf of Mexico. Gulf Mex Sci 18:23–34
Wright SD, Ackermarn BB, Bondec K, Beckand A, Banowetz DJ (1995) Analysis of watercraft-related mortality of manatees in Florida, 1979–1991. In: O’Shea TJ, Ackermarn BB, Percival HF (eds) Population biology of the Florida manatee, National Biological Service Information and Technology Report 1. National Technical Information Service, Springfield, VA, USA, pp 259–268
Würsig B, Jefferson TA, Schmidly D (2000) Marine mammals of the Gulf of Mexico. Texas A&M University Press, College Station, TX, USA. 232 p
Yordy JE, Wells RS, Balmer BC, Schwacke LH, Rowles TK, Kucklick JR (2010) Life history as a source of variation for persistent organic pollutant (POP) patterns in a community of common bottlenose dolphins (Tursiops truncatus) resident to Sarasota Bay, FL. Sci Total Environ 408:2163–2172
Zimmer WMX, Johnson MP, Madsen PT, Tyack PL (2005) Echolocation clicks of free-ranging Cuvier’s beaked whales (Ziphius cavirostris). J Acoust Soc Am 117:3920–3927
Useful Marine Mammal Websites
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Appendices
Appendix A
Marine mammal strandings recorded with human interaction in the Gulf of Mexico, 2000–March 2010 (human interactions document the presence or absence of signs of human interaction, not necessarily the cause of death).
There is overlap in the database under Human Interactions. Entries listed in more than one category include:
-
Other human interaction + Fishery interaction = 19 entries
-
Other human interaction + Shot = 1 entry
-
Other human interaction + Boat collision = 8 entries
-
Fishery and boat collision = 1 entry
Appendix B
Marine mammal unusual mortality events in the Gulf of Mexico, 2004–March 2010.
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution-NonCommercial 2.5 International License (http://creativecommons.org/licenses/by-nc/2.5/), which permits any noncommercial use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter’s Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter’s Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2017 The Author(s)
About this chapter
Cite this chapter
Würsig, B. (2017). Marine Mammals of the Gulf of Mexico. In: Ward, C. (eds) Habitats and Biota of the Gulf of Mexico: Before the Deepwater Horizon Oil Spill. Springer, New York, NY. https://doi.org/10.1007/978-1-4939-3456-0_5
Download citation
DOI: https://doi.org/10.1007/978-1-4939-3456-0_5
Published:
Publisher Name: Springer, New York, NY
Print ISBN: 978-1-4939-3454-6
Online ISBN: 978-1-4939-3456-0
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)