
Abstract
Sap-sucker insects are distinguished by their specialized mouthparts, adapted to penetrate and suck fluids from plant tissues. Many sap-sucking insect species are of major economic importance for forests, forest nurseries, and urban trees, causing plant stress, distortion, shoot stunting, and gall formation, or transmitting plant pathogens. In this chapter, we give an overview of the diversity and biology of sap-sucker insects of forest trees, with an emphasis on the two major groups, aphids, and scale insects. We further present their ecology highlighting their biotic interactions with other organisms. Finally, we discuss sap-sucking forest pests and their management.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


13.1 Introduction
Piercing-and-sucking insects are distinguished by their specialized mouthparts, with stylets adapted to penetrate and suck fluids from plant or animal tissues. These insects are primarily hemipteroids (e.g. Psocodea, Hemiptera, and Thysanoptera), Siphonaptera (fleas), and nematocerous Diptera. Among the piercing-and-sucking hemipteroids, the sucking lice (Psocodea, Anoplura) are obligate ecotoparasites, feeding on mammal or bird blood. Most of the hemipteran species are phytophagous. The non-phytophagous species all belong to the sub-order Heteroptera and are predators, scavengers, a few are blood-feeders and some are necrophages. Thrips (Thysanoptera) include mycetophagous species (about 50%), phytophagous and a few predatory species. Non-hemipteroid piercing-and-sucking insects are mostly insectivores or blood-feeders, and less commonly feed on fungi or algae (Gullan and Cranston 2014; Labandeira and Phillips 1996; Morse and Hoddle 2006).
Two feeding strategies exist within the phytophagous hemipterans: (1) salivary sheath feeding, whereby individuals feed on contents of plant vascular tissue, i.e. phloem or xylem (most sap feeders); or (2) cell rupture feeding, i.e. mesophyll feeders. Sap-sucking insects are salivary sheath feeders and can be further differentiated in two feeding guilds: (1) phloem-feeders, most of the species of the suborder Sternorrhyncha, including aphids (Aphidoidea), whiteflies (Aleyrodoidea), scale insects (Coccoidea), and psyllids (Psylloidea); many Auchenorrhyncha, including treehoppers and many leafhoppers (Membracoidea: Membracidae, most Cicadellidae), and planthoppers (e.g. Fulgoroidea: Cixiidae, Delphacidae, Flatidae, Fulgoridae), and some Heteroptera; and (2) xylem-feeders, such as cicadas (Cicadoidea), spittlebugs (Cercopoidea), and sharpshooter leafhoppers (Cicadellinae), all belonging to the suborder Auchenorrhyncha. Phytophagous thrips feed on the content of individual epidermal or mesophyll cells (i.e. mesophyll feeders) (Bennett and Moran 2013; Chuche et al. 2017a; Douglas 2006; Labandeira and Phillips 1996; Redak et al. 2004).
Many sap-sucking insect species are of major economic importance, because they often cause plant stress, distortion, shoot stunting, and gall formation, or transmit plant pathogens (Baumann 2005; Gullan and Cranston 2014). In this chapter, we address the evolution and diversity of sap-sucking insects of forest trees, with an emphasis on the two major groups, aphids and scale insects. We present their biology and ecology. Particular emphasis is given to their highly specialized feeding mode and biotic interactions. Finally, we discuss sap-sucking forest pests and their management. In Figs. 13.1a–l and 13.2a–k, we provide images of example species and some of the aspects mentioned in the chapter.


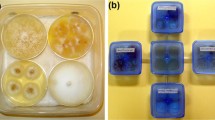
Photo credits: 13.1a, b, f, j © Alex Protasov; 13.1d, i © Zvi Mendel; 13.1e, h © Vera Zina; 13.1c, g, k, l © Manuela Branco
a Paleococcus fuscipennis mating; b Thaumastocoris peregrinus development stages; c Matsucoccus feytaudi male; d Mouth parts of T. peregrinus; e and h Ants tending lerps of G. brimblecombei, Crematogaster scutellaris and Plagiolepis pygmaea, respectively; f Elatophilus hebraicus (Anthocoridae) feeding on Matsucoccus jospehi while mating; g Larviform nymphs of M. feytaudi eclosing from egg mass. i A whitefly (Aleyrodidae); j A soft scale, Kermes echinatus; k and l Adult and larvae of Iberorhyzobius rondensis (Coccinelidae), monophagous predator of M. feytaudi.

Photo credits: 13.2a, e, h, i, j, k © Zvi Mendel; 13.2d, f © Vera Zina: 13.2b, c, g © Manuela Branco
a and b Predators (Anthocoridae and Hemerobiidae) and males of Matsucoccus feytaudi attracted to lures with the sex pheromone of the bast scale; c Band trap to collect ovipositing females of M. feytaudi; d Honeydew exploited by ants; e Soft scales (Coccidae) and sooty mold on Ficus sycamorus; f Infestation of Glycaspis brimblecombei on Eucalyptus camaldulensis; g Ultracoelostoma sp. canal tube with a drop of honeydew and sooty mold on Nothofagus; h Honey production in Crete from Marchalina helenica; i Forest damage caused by Pineus pini on Pinus pinea; j Crystallized honeydew excreted by Cinara palestina; k Large colony of Cinara cedri.
13.2 Diversity and Biology of Sap-Sucking Insects with Emphasis on Importance for Forestry
13.2.1 Background
Insects evolved on land in the Ordovician, about 480 million years ago (Misof et al. 2014). Approximately 80 million years later, in the Devonian period, one lineage of insects evolved flight. Hemipteran insects, with their defining trait of piercing-sucking stylet mouthparts (Fig. 13.1d), probably arose in the Carboniferous period, about 300 million years ago. The order Hemiptera is divided into several monophyletic branches, including the Auchenorrhyncha (cicadas, spittlebugs, leafhoppers, treehoppers), the Sternorrhyncha (scale insects, psyllids, whiteflies, aphids) and the Heteroptera (true bugs) (Song et al. 2012). The oldest fossils of aphids, coccoids and Heteroptera are from the Triassic period, about 220 million years ago (Hong et al. 2009). Table 13.1 displays the different major groups of Hemiptera and the significance of their members as forest pests.
13.2.2 Aphids: Aphidomorpha
The entire Aphidomorpha infraorder has now been fully catalogued: 5,218 valid extant and 314 valid extinct species (Favret et al. 2016). Aphids form a distinctive insect clade that features considerable variability in their biological traits. For example there are many distinct, yet genetically identical, forms of females during the life cycle (polyphenism), alternation of sexual and asexual reproduction and seasonal alternation between unrelated groups of host plants. These traits vary among species, reflecting evolutionary histories of biogeographical expansions and contractions and co-diversification with plant hosts. Many aspects of phylogenetic relationships among aphids remain unresolved, and several evolutionary questions unanswered (Nováková et al. 2013).
Extant Aphidomorpha are divided in two superfamilies. The superfamily Aphidoidea includes the true aphids (Aphididae). The superfamily Phylloxeroidea includes the adelgids (Adelgidae), feeding on conifers, and phylloxerids (Phylloxeridae), whose members develop on broadleaf trees. Adelgidae, unlike true aphids, have no tail-like cauda and no cornicles. All three families have winged and wingless forms. Winged forms are produced for dispersal in asexual generations, and for migration between the secondary and primary host plants (see Sect. 13.3.1). In these species, the sexual morphs mate on the primary host, producing overwintering eggs.
Adelgids are often known as “woolly conifer aphids” or “woolly adelgids”. The family is composed of species associated with pine, spruce and other Pinaceae genera (Table 13.2, Fig. 13.2i). The most common classification system recognizes 8 genera (Adelges, Aphrastasia, Cholodkovskya, Dreyfusia, Eopineus, Gilletteella, Pineus and Sacchiphantes), including about 50 species. All of them are native to the northern hemisphere, although some have been spread to the southern hemisphere as invasive species. Adelgids exhibit cyclical parthenogenesis and are oviparous (Havill and Foottit 2007). They exhibit a two-year life cycle, with some species alternating hosts between spruce (Picea) and other Pinaceae (Abies, Larix, Pinus, Pseudotsuga, Tsuga).
The phylloxerids include 75 described species within two subfamilies (Phylloxerininae and Phylloxerinae) and 11 genera (Blackman and Eastop 1994). They have a worldwide distribution but are more diverse in temperate climates where they likely originated. Their adaptation to tropical habitats is probably secondary. They feed on leaves and roots and induce galls at their feeding sites. Most phylloxerids feed on Juglandaceae or Fagaceae. Still, they are not considered significant pests of forest trees. In Israel, Phylloxera quercus occurs on oak trees (mainly Quercus calliprinos) in very low densities and may be often found on shoots developed after a major fire (Table 13.2). However, like the grape phylloxera Daktulsphaira vitifoliae, the majority of the population occurs on the roots.
The true aphids are a very large insect family, including several thousand species, many of which are known as serious plant pests (Table 13.2). The oldest aphid fossils are from the Triassic (at least 220–210 mya) but aphids may have originated in the Permian. Phylogenetic analysis of molecular data suggests that aphids underwent a rapid radiation into the current tribes after switching from gymnosperms to angiosperms, sometime during the Upper Cretaceous. Furthermore, the ancestral aphid probably had a simple life cycle with host alternation evolving independently in each of the subfamilies (Gullan and Martin 2009).
Two genera of the subfamily Lachninae, i.e. Cinara, the conifer aphids or giant conifer aphids, and Eulachnus, are well known as forest pests. Cinara species (≈150) are widespread in the Northern Hemisphere. Many species are native to North America, but there are also 55 species found in Europe and Asia. They are specialized on members of the families Pinaceae and Cupressaceae. Cinara spp. do not alternate hosts. The genus Eulachnus comprises about 17 species, all of which live on pine needles. They are cryptic when feeding, but very active when disturbed. The best-known species show preferences for certain Pinus spp., but none is strictly confined to one species. Several introduced Cinara spp. have become serious pests of forest plantations. For example, Cinara cedri (Fig. 13.2k) and Cinara laportai attack true cedars. Cinara cupressi causes damage to cypress trees. This aphid of unclear origin is an invasive species in Africa and Europe, South America and in the Middle East.
13.2.3 Jumping Plant Lice: Psylloidea
Psylloidea is a superfamily of true bugs, including the jumping plant lice, recently classified in eight families (Aphalaridae, Calophyidae, Carsidaridae, Homotomidae, Liviidae, Phacopteronidae, Psyllidae and Triozidae). There are about 3,000–3,500 described species. They are common worldwide, but most diverse in tropical and subtropical areas. Most Australian psyllids belong to the subfamilies Spondyliaspidinae (Aphalaridae) and Acizziinae (Psyllidae). The former is largely associated with eucalypts and the latter with acacias (Carver et al. 1991). Members of the Psylloidea also include many gall-inducing species, which are narrowly host-specific, and are most species-rich in the tropics and south temperate regions (Burckhardt 2005).
Psyllids reproduce sexually and mature through five nymphal stages. Unlike aphids, psyllids insert their eggs into host plant tissue (Hodkinson 1974). They are phloem feeders and produce honeydew. Many are known vectors of plant diseases. Of major economic importance are those vectoring Liberibacter and Phytoplasma species, the causal agents of serious plant diseases. They generally have narrow host ranges and are restricted almost exclusively to perennial dicotyledonous plants. Within species, nymphs usually have a more restricted host range than adults. At low densities, nymphal survival is enhanced by group feeding similar to many Heteroptera. Many species occur in dry or semi dry areas and the immature stages exhibit morphological and behavioral adaptations to resist desiccation. This is for example seen in the circular lerp of the red gum lerp psyllid, Glycaspis brimblecombei, which may also protect them from predators (Fig. 13.1e, h).
Psyllids are not known from conifers, but may overwinter on them (e.g. Čermák and Lauterer 2008). Also, they are not common on oak trees and none are known as pests of oaks. Conversely, psyllids feeding on eucalypts are among the most devastating insect pest groups in Australia, affecting both native forests and plantations.
Psyllids of several different families have become economically important invasive pests. Several Aphalaridae species, originating from Australia, have been introduced into other continents, where they become important pests, causing severe damage in eucalypt plantations (Hurley et al. 2016). Most of these species infest the river red gum, Eucalyptus camaldulensis. One of the most widely distributed psyllid species is the red gum lerp psyllid G. brimblecombei (Figs. 13.1e and 13.2f). The psyllid Acizzia jamatonica, native to China, has been reported from Europe and North America as a pest of Albizia julibrissin. Acizzia uncatoides, native to Australia, develops on many ornamental Acacia and Albizia species outside Australia. In Hawaii, it occurs on the native Acacia koa. Calophya schini (Calophyidae) is a leaf galling psyllid that feeds exclusively on Peruvian pepper tree, Schinus molle (Anacardiaceae). This psyllid is now present in California, Mexico, Portugal, South Africa, Ethiopia and Kenya (e,g. Overholt et al. 2013; Zina et al. 2012). The psyllid Macrohomotoma gladiata (Homotomidae) is a new insect pest of Ficus microcarpa originating from Asia, which has recently been found in Spain (Alicante) on urban trees (Mifsud and Porcelli 2012) and is now widely spread in East Mediterranean.
13.2.4 Scale Insects: Coccoidea
The Coccoidea is one of the four superfamilies of the monophyletic suborder Sternorrhyncha (e.g. Gullan and Martin 2009), with 49 families presently recognized (Ben-Dov et al. 2014). The early evolution of Coccoidea must have occurred during the early to mid-Mesozoic, as a sister group of the Aphidoidea (Hennig 1981). Almost all the main lineages of modern coccoids have been identified from Tertiary amber, but relatively few earlier fossils are known (Koteja 1986, 1990). The morphology of these early fossils suggests that some groups of the plesiomorphic Margarodidae sensu lato had reached their contemporary organization by the Lower Cretaceous (Koteja 1990). Miller (1984) used the aphids as outgroup and showed that the earliest scale insects were margarodid-like and that Margarodidae and then the Ortheziidae are successively sister to the remainder of the Coccoidea.
Traditionally, based on the possession of abdominal spiracles, the scale insects were separated between Archaeococcoids and Neococcoids, with the latter characterized by features such as the loss of abdominal spiracles (Koteja 1990). The Archaeococcoids comprise several families, such as Monophlebidae, Margarodidae, Orthezidae and Matsucoccidae. The Neococcoids comprise most of the currently recognized families and species of scale insects (e.g. Kosztarab 1982). Its major groups probably evolved in conjunction with the angiosperms. Almost all neococcoid fossils are from Eocene or younger deposits. Yet, the neococcoid radiation must have begun much earlier because all of the major families (Coccidae, Diaspididae, Eriococcidae and Pseudococcidae) were already present in the Eocene. The soft scales (Coccidae) consist of four major subgroups, the Ceroplastinae, Coccini, Pulvinariini and Saissetiini (Miller and Hodgson 1997). The armored scales (Diaspididae) are divided in several subfamilies and tribes with the most important being Aspidiotini and the Diaspidini. The felt scales (Eriococcidae) are not a single monophyletic group, but a complex of several different groups and several families (Cook et al. 2002). The mealybugs (Pseudococcidae) are currently separated in two subfamilies, Phenacoccinae and Pseudococcinae (Danzig and Gavrilov-Zimin 2015).
There is a very marked sexual dimorphism of adult male and female scale insects (Fig. 13.1a, c). This sexual dimorphism is established by divergent postembryonic developmental pathways after the first-instar nymph, possibly regulated by growth hormones (Vea et al. 2016). Danzig (1980) suggested that neoteny shortens female development time on coccids, resulting in nymphal morphology in adult females. This is not unique to scale insects. The maintenance of juvenile features in adults, through neoteny, has evolved independently at least six times in insects. Mature adult female scale insects generally have a large body relative to nymphs or teneral females (Ben-Dov 1990) and often have very high fecundity (McKenzie 1967).
The inability to fly has been suggested to be adaptive for female, but not male scale insects, because more resources can be allocated to egg production (Roff 1990). The adult scale insect male morphology is adapted for flight and finding females. The male mesothoracic wings have reduced venation, whereas the metathoracic ones are lost or reduced enabling the male to control its equilibrium in flight. The mouthparts in males have become nonfunctional (e.g. Afifi 1968; Gullan and Kosztarab 1997; Kawecki 1964). Waxy caudal filaments in males are known in several coccoid families (Fig. 13.1c) (Afifi 1968; Giliomee 1967). Duelli (1985) suggested that these filaments assist in stabilizing flight. The elongation of the wax caudal filaments was shown to be correlated with sexual maturation of adult males in mealybugs (Mendel et al. 2011). Because of extreme sexual dimorphism in scale insect, the conspicuous and longer-lived adult females are used in taxonomy. For many coccoid genera and species, the male is unknown.
Modern scale insects are all plant feeders, using their stylet-like mouthparts to suck sap from the phloem or parenchyma cells. The first scale insects probably fed on proto-angiosperms, gymnosperms or lower plants, or on fungi and bacteria (Koteja 1986). Vea and Grimaldi (2016) suggested that most major lineages of coccoids shifted from gymnosperms onto angiosperms, when the latter became diverse and abundant in the mid- to late Cretaceous. Alternatively, the ancestral scale insects may have fed on the contents of individual cells from roots, rotting plants, or fungal hyphae. Koteja (1986) has hypothesized that the leaf-litter layer is the primary habitat of coccoids and that feeding on above-ground plant parts is a secondary adaptation.
All forest tree species are infested by scale insects. Figure 13.3 compares the frequency of occurrence of species belonging to eight families, on four tree genera from different botanical families: Pinus (with ~111 tree species and 182 scale species) and Quercus (~600 tree species and 227 scale species), of Laurasia origin; Eucalyptus (~700 tree species and 273 scale species) and Acacia (~980 tree species and 308 scale species), of Gondwanaland origin. These tree genera are naturally distributed over large areas and comprise several tree species, which are among the most economically and environmentally important globally.

Species frequency distribution of eight scale insect families among four genera of forest trees. The data were retrieved from Scale Net (García Morales et al. 2016)
It is interesting to note that some scale insects are primarily associated with specific tree genera. For example, Eriococcidae is the largest family of scale insects on Eucalyptus (~49%), but this family has limited representation on other tree genera (~2–6%); members of the family Kermesidae are only able to develop on Fagaceae (Fig. 13.1i), predominantly the genus Quercus (e.g. Spodek et al. 2013), while Matsucoccidae are associated only with the genus Pinus (e.g. Foldi 2004). Conversely, other families are evenly distributed across tree taxa. In particular Diaspididae, the largest family of scale insects, is the dominant scale insect family of Pinus, Quercus and Acacia (~38–51% of the species), and the second most species-rich family in Eucalyptus. Coccidae is also present in all the four tree genera, but is most common on Acacia (Fig. 13.3).
13.2.5 Other Hemipteran Superfamilies and Their Importance in Forestry
13.2.5.1 Auchenorrhyncha: Cicadomorpha
Frog hoppers and spittlebugs (Cercopoidea) are common insects on eucalypts. Frog hopper nymphs construct tubes which are attached to stems and twigs. The nymphs live inside these tubes where they are protected from desiccation and also to some extent from parasites and predators. Spittlebug nymphs live within a white, frothy secretion that resembles spittle (hence the name). As with the froghopper tubes, the spittle protects the nymphs from desiccation and from parasitoids and predators. The nymphs take shelter and feed beneath this froth. Members of the genus Aphrophora are known as minor pests of several trees in North America and Europe (Floren and Schmidl 2009).
Generally, cicadas are not significant forest pests. Nymphs live in the soil and feed on roots while adults feed on above-ground parts of plants, but this seems to have little effect on plant growth. During oviposition, females pierce plant tissues with their ovipositor to lay eggs and this can result in structural damage. For example, the cuts made by Amphipsalta spp. during oviposition may weaken twigs and branches sufficiently that they break in high winds. Such broken branches on conifers show up as reddish “flags” in the canopy when the foliage dies (Kay 1980). Open cuts also provide points of entry for pathogens and wood-boring insects. Often the cuts heal over making the twigs gnarled in appearance.
Leaf hoppers, family Cicadellidae (whose hind legs are modified for jumping) and treehoppers and thorn bugs, family Membracidae, are common in various types of forest all over the world, but not considered important forest pests. For example, aggregations of Oxyrhachis versicolor are conspicuous on Tamarix trees in Israel, but result in no apparent damage to the trees. Leafhoppers are commonly associated with broadleaved forest trees (Beirne 1956). In Costa Rica, some species have been reported to cause minor damage to native broadleaf trees, as well as to introduced eucalypts (e.g. Macumolla ventralis and Graphocephala coccinea) (Gamboa 2007). Leafhoppers gained particular attention as vectors of the plant pathogenic bacterium Xylella fastidiosa. In particular, sharpshooter leafhoppers (Cicadellinae) are the best-studied vectors of X. fastidiosa (Cornara et al. 2019). For example, the glassy-winged sharpshooter, Homalodisca vitripennis, an invasive species in California affecting a wide host range of trees, is an efficient vector of X. fastidiosa (Almeida and Nuney 2015).
13.2.5.2 Auchenorrhyncha: Fulgoroidea
The planthopper superfamily Fulgoroidea comprises approximately 20 described insect families, depending on which classification is followed, and includes a diverse group of phytophagous or fungivorous insects, exceeding 12,500 species. At least 160 species in 16 families are recorded as pests, including some of major economic importance, such as the brown planthopper, Nilaparvata lugens on rice. Planthoppers are vectors of viral and bacterial (including phytoplasma) agents causing plant diseases. The Tropiduchidae comprises 652 described species, about 4.9% of all Fulgoromorpha. Most species feed on shrubs and trees, and some are crop pests. Their association with host plants is quite diverse, including 21 plant orders. Still, few or no species in most of the families cause economic damage to forests.
13.2.5.3 Sternorrhyncha: Aleyrodoidea
Whiteflies are almost entirely leaf feeders (Fig. 13.1i). In recent years, whitefly pests have become a major problem in agriculture, almost worldwide, but as forest insects are of little concern (Nair 2001). In recent years the Ficus whitefly, Singhiella simplex (Aleyrodidae) has become a pest of Ficus trees in North America and the Mediterranean. Primarily a tropical group, pest species are found in the warmer parts of the world. In temperate areas, several species are serious pests in glasshouses, but they do not pose any risk to forest trees (Martin et al. 2000).
13.2.5.4 True Bugs: Heteroptera
This suborder is highly diverse, although few species are considered as important pests of forest trees. It is interesting to mention two invasive species in Europe and the Mediterranean. One is the bronze bug Thaumastocoris peregrinus (Thaumastocoridae), native in Western Australia affecting Eucalyptus (Fig. 13.1b). Infestations are noted by the reddening of eucalypt canopy leaves and loss of leaves, leading to canopy thinning and occasionally branch dieback or tree mortality (Nadel et al. 2010). Another pest is the Sycamore Lace Bug Corythucha ciliata (Heteroptera: Tingidae). This North American species, was introduced in Europe in the 1960s and first found in China in 2006, where it has major impacts on Platanus tree health in urban parks and street trees (Maceljski 1986; Ju et al. 2009).
13.3 Biology and Ecology of Sap-Sucking Insects
13.3.1 General Models of Life History and Seasonal History
The life cycle of sap-sucking insects may include two or three nymphal stages (e.g. female scale insects), four (e.g. male scale insects, aphids, whiteflies) or five (e.g. jumping plant lice, Auchenorrhyncha and Heteroptera (Fig. 13.1b) (Dietrich 2003; Gullan and Martin 2009). In some cases, such as in whiteflies and scales insects, the immature stages are larviform, much different from adults (Fig. 13.1g), followed by a non-feeding pupal instar. This developmental pattern is similar to holometabolic insects (Gullan and Cranston 2014). The number of complete generations within a year varies among species and with climatic conditions. Univoltine species (one generation per year) are common in cold temperate regions and multivoltine species (more than two generations) in warmer climates (Dietrich 2003; Gullan and Martin 2009). Some species (e.g. the psyllid Strophingia ericae) may take more than one year to complete one generation (Hodkinson 2009). Magicicada (Cicadidade) species have periodical life cycles that last 13 or 17 years (Grant 2005).
Polyphenism, i.e. phenotypic differences determined by environmental conditions, occurs in some sap feeders. Some aphids show very complex polyphenisms. Parthenogenetic females may have up to eight different phenotypes, and sexual forms are polymorphic. Phenotypic differences may include morphological and physiological aspects, as well as fecundity, timing and size of progeny, developmental time, longevity, and host-plant selection. Photoperiod, temperature and maternal effects are among the environmental cues triggering the development of different aphid morphs (Gullan and Cranston 2014). Adelgids, phylloxerids, and aphids present a complex, polymorphic life cycle with cyclical parthenogenesis and host alternation. Some species are holocyclic, meaning they produce both asexual and sexual generations, while others are anholocyclic, producing only asexual generations. The typical adelgid holocycle takes two years to complete and involves five generations: three are completed on the primary host (spruce, Picea spp.), with sexual reproduction and gall formation; and the other two are asexual generations and occur on the secondary host (Abies, Larix, Pseudotsuga, Tsuga or Pinus) (Havill and Foottit 2007).
13.3.2 Feeding Ecology
Hemipteran insects modified mouth parts include a slender beak-like labium, within which there are two pairs of long stylets (two outer mandibles and two inner maxillae), forming a bundle (Fig. 13.1d). Only the stylets penetrate into the plant tissues for piercing-and-sucking. The maxillary stylets form in their inner surface both the salivary canal and the food canal. Through these channels the insect injects saliva into the plant, or sucks up plant sap into their gut, respectively. Commonly, in sap-feeding insects, the stylets pathway up to the vascular tissue is mostly intercellular, following an apoplastic transit, i.e. following a route from cell wall to cell wall, not entering the cytoplasm. Eventually, some intracellular punctures occur to assess cell content, for host acceptance and setting the position of the stylets inside the plant.
As salivary sheath-feeding insects, sap feeders produce two types of saliva. A gelling lipoproteinaceous saliva is secreted during the penetration process of stylets, which forms a lubricating and hardening sheath around them. This gelling saliva remains within plant tissues after stylet withdrawal. Additionally, a watery saliva, is directly injected in the vascular tissue before sap uptake. This watery saliva may interfere with host-plant defense responses, for example, through proteins that are involved in the detoxification of phenols or interact directly with plant defense signaling (Giordanengo et al. 2010; Gullan and Cranston 2014; Kingsolver and Daniel 1995; Will et al. 2013).
Phloem and xylem tissues have very different characteristics and consequently sap-feeding insects usually specialize on one or the other (Labandeira and Phillips 1996; see Table 13.3). However, phloem-feeding insects may occasionally ingest xylem-sap, possibly for regulating osmotic potential (Pompon et al. 2011). Phloem sap is a rich source of carbon and energy (i.e. sugars), also providing nitrogen (mostly as free amino acids). Also phloem sap usually has no toxins and feeding deterrents. However, sap-sucking insects need to overcome two major nutritional problems to feed on phloem-sap: (1) nitrogen quality in phloem-sap is low (the ratio between essential and non-essential amino acids in phloem-sap is 1:4–1:20); and (2) phloem sap has a very high concentration of sugars (i.e. an osmotic pressure 2–5 times higher than that in insect’s hemolymph) (Douglas 2006).
Phloem feeders excrete the excess carbohydrates from their unbalanced diet in the form of a sugary fluid, honeydew (Baumann 2005) (Fig. 13.2d, g). Xylem sap is often less nutritional than phloem sap, containing low concentrations of carbon and nitrogen compounds and is under negative pressure. Xylem-feeders compensate this constraint with high ingestion rates and generally have larger bodies than phloem-feeders. Also due to the relatively higher metabolic costs of xylem-sap extraction (Chuche et al. 2017b). In fact, the suction pressure needed for sucking xylem sap decreases with food canal width, which is directly correlated to body size of the insect (Novotny and Wilson 1997).
As both phloem and xylem are unbalanced food sources, sap-feeding insects rely on symbiotic bacteria to provide them the essential nutrients lacking in their diet (Bennett and Moran 2013). Primary endosymbionts (P-endosymbionts) are obligate mutualistic bacteria, localized within hemipteran-host polyploid cells (the bacteriocytes), which normally aggregate into a specialized organ, the bacteriome. P-endosymbionts are essential for host survival and reproduction, and are present in all individuals of the host population (Table 13.4). They present an extreme genomic reduction as a result of vertical transmission, from reproductive females to progeny, and living inside bacteriocytes.
In addition to these nutritional primary symbionts, sap-sucking insects may also contain one or more facultative or secondary symbionts (S-symbionts). In general these symbionts are not necessary for host development and reproduction. S-symbionts may inhabit a variety of tissues other than bacteriocytes, often do not infect all individuals within host populations, and can be horizontally transmitted among hosts. These symbionts are known to manipulate host reproduction and provide their hosts with a range of adaptive ecological traits. These include increased host-plant range, efficiency of plant pathogen transmission, and greater resistance to biotic (e.g. parasitoids) or abiotic (e.g. temperature, insecticides) environmental stress (Baumann 2005; Chuche et al. 2017a; López-Madrigal and Gil 2017; Oliver et al. 2010).
Examples of secondary symbionts found in aphids include Serratia (47% of the studied aphid species), Wolbachia (43%), Hamiltonella (34%), Regiella (33%), Rickettsia (29%), X-type (14%), Spiroplasma (13%), and Arsenophonus (9%) (Zytynska and Weisser 2016). A peculiar symbiotic organization was observed in the citrus mealybug Planococcus citri, in which each cell of the P-endosymbiont Tremblaya princeps harbors several cells of the S-symbiont Moranella endobia, representing the first known case of prokaryote-prokaryote endocelullar symbiosis (López-Madrigal et al. 2013).
13.3.3 Reproductive Strategies
Sexual reproduction and oviparity are the most common modes of reproduction in sap-sucking insects (Dietrich 2003; Gullan and Martin 2009). However, other reproductive strategies can be observed in this insect guild (Table 13.5). Different types of parthenogenesis, i.e. apomixis, automixis, and pseudogamy, are known for example in aphids, whiteflies, scale insects and plant hoppers. Examples of mixed systems, including different types of alternation between sexual reproduction and parthenogenesis (e.g. facultative and cyclic parthenogenesis), have been described in adelgids, phylloxerans, aphids and other hemipterans. Few species, such as Icerya spp. are hermaphrodite (Gullan and Martin 2009; Ross et al. 2010).
13.3.4 Insect-Plant Interactions
Sap sucking insects may be classified in different feeding groups, based on the part of the host plant they feed on: (1) shoots and tips, e.g. the Cooley spruce gall adelgid, Adelges cooley; (2) foliage, e.g. the green spruce aphid, Elatobium abietinum; (3) trunk and branches, e.g. maritime pine bast scale, Matsucoccus feytaudi, Beech scale insect Ultracoelostoma spp. (Fig. 13.2g), and (4) roots, e.g. spruce root aphid, Pachypappa termulae (Foldi 2004; Wood and Storer 2003).
Host range is variable among sap sucking insects. Most Auchenorrhyncha are apparently very specific, feeding in one single plant genus or species. However, many Auchenorrhyncha, especially xylem-feeders can feed and develop on several alternate plant species if the preferred host is not present. Phloem- and mesophyll-feeders tend to be more host-specific than xylem-feeders, with many species limited to host plants from a single family, genus, or species (Dietrich 2003).
In the Sternorrhyncha, host range varies among taxa. Adelgids are host specific, as each species survives and reproduces only on trees from a single genus, for both primary and secondary hosts (Havill and Foottit 2007). Most species of plant jumping lice are also host specialists as nymphs, with many restricted to a single plant genus or species, and often to certain host parts (e.g. leaves, young shoots), or growth stages. In the case of whiteflies, most are apparently oligophagous, with a few monophagous. Host plant specificity in scale insects ranges from monophagous to polyphagous. Most aphids are monoecious, meaning that development occurs on one or a few closely related host plants; however, about 10% of species are heteroecious, i.e. with host alternation. As a result, most aphids are host specific. This property is conspicuous in aphids developing on forest trees. An aphid genus is usually associated with a single host-plant family and species with a plant genus or species. The primary and secondary host of heteroecious aphids are usually unrelated and host specificity is higher in the case of the primary than in the secondary host (Gullan and Martin 2009).
Several lineages of sap sucking insects induce the formation of plant galls. Plant galls, or cecidia, are abnormal growths of plant tissue, involving cell proliferation (hyperplasy) and enlargement (hypertrophy). This abnormal tissue growth results in the development of characteristic gall structures, which are specific to a certain gall making organism (Schick and Dahlsten 2003). Gall makers evolved independently in the Hemiptera, primarily within the Sternorrhyncha, including the aphids, adelgids, phylloxerids, woolly aphids (Eriosomatidae) (Wool 2005), scale insects (Asterolecaniidae, Coccidae, Diaspididae, Eriococcidae, Kermidae) (Gullan et al. 2005), psyllids (Psyllidae), and few Auchenorrhyncha (Cercopidae, Cicadellidae) (Burckhardt 2005). Galls behave as physiological sinks in the host plant, sequestering nutrients used by developing insects inside them, as well as a defensive refuge against the natural enemies of gall makers (Schick and Dahlsten 2003; see also Chapter 14).
13.4 Associated Organisms
13.4.1 Natural Enemies
Although sap sucking insects are prey of some insectivorous vertebrates (e.g. birds and lizards), they are primarily predated by invertebrates (Table 13.6). This diverse assemblage of predators includes spiders, and insects from different orders [e.g. assassin and minute pirate bugs (Hemiptera: Reduviidae and Anthocoridae) (Fig. 13.1f), ladybirds (Coleoptera: Coccinellidae) (Fig. 13.1k), green and brown lacewings (Neuroptera: Chrysopidae and Hemerobiidae), ants (Hymenoptera: Formicidae) (but see Sect. 13.4.2), wasps (Hymenoptera: Vespidae), and predatory flies (Diptera: Cecidomyiidae, Chamaemyiidae and Asilidae)]. Parasitoids in the families Dryinidae (Hymenoptera, Chrysidoidea), Encyrtidae, Eulophidae, Aphelinidae (Hymenoptera, Chalcidoicdea), and Braconidae (Hymenoptera, Ichneumonoidea), are also natural enemies of sap suckers.
Due to their feeding habits, sap feeders are usually not affected by entomopathogenic viruses, bacteria, protozoa, or nematodes, as these entomopathogens are not common in the plant vascular system. Entomopathogenic fungi are their most important pathogens. Unlike other entomopathogens, fungi usually actively penetrate the insect cuticle and do not need to be ingested by insects for infection to occur. Populations of sap sucking insects, particularly aphids and leafhoppers, may suffer epizootics caused by fungi (Dietrich 2003; Federici 2003; Gullan and Martin 2009).
13.4.2 Interaction with Ants
Honeydew produced by many Sternorrhyncha is a food source for different animal species, including many insects (e.g. flies, wasps, bees, ants, beetles, lacewings, butterflies, and moths), and nectar feeding birds and bats. Ant-tending behavior, which consists of collecting honeydew droplets directly from the anus of sap-sucking insects, is common among ants, especially in Dolichoderinae and Formicinae (Douglas 2006) (Figs. 13.1e, h and 13.2d). This ant behavior is linked to a food-for-protection type of mutualism (Styrsky and Eubanks 2007). A food resource, the sugar-rich honeydew excreted by tended sap-sucking insects, is traded for a service delivered by ants, the protection of hemipterans from predators and parasitoids (Way 1963). In the presence of honeydew-producing hemipterans, increased ant predation of other herbivores may indirectly benefit host plants if the amount of damage originated by those herbivores is greater than that inflicted by ant-tended hemipterans (Fig. 13.4). Conversely, tending ants may enhance the negative effects of sap-sucking insects on plants (e.g. reduced plant growth, transmission of plant pathogens), by protecting them from their natural enemies, and by increasing their feeding rate, fecundity and dispersal (Styrsky and Eubanks 2007; Vandegehuchtea et al. 2017 and references therein).

Redrawn from Styrsky and Eubanks (2007)
Interactions among honeydew-producing hemipterans, ants and host-plants. Arrows indicate the direction of effects, positive (+) or negative (−), whereas solid arrows indicate direct effects, and dashed ones indirect effects.
Styrsky and Eubanks (2007) reviewed the literature on the influence of ant–hemipteran interactions on arthropod communities and their host plants and reported that these interactions have mostly negative effects on the abundance and species richness of different herbivore and predator guilds. These authors also observed that in about 73% of the studies plants indirectly benefited from those interactions, as a result of increased predation of other more damaging herbivores by hemipteran-tending ants.
13.4.3 Bees and Honey Production from Honeydew
Honeybees are among the insects using the honeydew excreted by sap feeders as a food resource. Honeydew honeys are well known and valued in Europe and New Zealand (Table 13.7), but are also produced in North America (e.g. white cedar honey, from honeydew of the scale insect Xylococculus macrocarpae on Calocedrus decurrens) and South America (e.g. from honeydew of the scale insects Stigmacoccus asper on Quercus humboldtii, and Tachardiella sp. on Mimosa scabrella) (Azevedo et al. 2017; Chamorro et al. 2013). Originated from different plant species, mostly conifers, such as fir, spruce and pines, but also some broadleaf trees, honeydew honeys are highly valued in many European countries and are marketed with specific designations (Oddo et al. 2004). In some countries, such as Greece and Turkey, honeydew honey may represent more than 65% of total honey production, most of it produced from honeydew excreted by Marchalina hellenica (Marchalinidae) in pine forests (Santas 1983) (Fig. 13.2h). This species has been deliberately introduced by beekeepers, for producing honeydew, in pine forests in Crete island, Greece, where it became a serious problem. Similarly, the invasive species in Europe, the citrus flatid planthopper, Metalfa pruinosa, is highly appreciated by beekeepers for the honeydew with high economic value (Preda and Skolka 2011).
13.4.4 Hemiptera as Vectors of Microorganisms
As a result of their feeding habits, sap-sucking insects interact with plant pathogens which colonize the plant vascular system, such as viruses and bacteria, functioning as vectors (Perilla-Henao and Casteel 2016). Vector-borne bacteria are primarily transmitted by Auchenorrhyncha insects, including species from the superfamilies Membracoidea, Cercopoidea, and Fulgoroidea. Although less common, some Sternorrhyncha (e.g. psyllids) are also important vectors of phytopathogenic bacteria. Sap-feeding vectors of plant viruses have been reported in Auchenorrhyncha (Fulgoroidea and Membracoidea), and Sternorrhyncha (Aphidoidea, Aleyrodoidea, and Coccoidea) (Chuche et al. 2017a; Cornara et al. 2019; Perilla-Henao and Casteel 2016).
Four main types of transmission relationship between vectors and plant pathogens have been defined: (1) non-persistent; (2) semi-persistent; (3) persistent, non-propagative; and (4) persistent, propagative (Table 13.8). These types of transmission relationships were defined based on the following parameters: (1) the time needed for the acquisition and inoculation of the pathogen by the vector; (2) the retention time of the pathogen within the vector; (3) whether or not the pathogen circulates within the vector and (4) the ability of the pathogen to reproduce within the vector.
Non-persistent, non-circulative transmission of plant viruses has been only observed among viruses transmitted by aphids, whereas semi-persistent, non-circulative transmission is known in aphid, whitefly and leafhopper-transmitted viruses. Luteoviruses are an example of circulative, non-propagative viruses, which are transmitted by aphid vectors, whereas the genera Fijivirus, Phytoreovirus, and Oryzavirus of Reoviridae are circulative, propagative viruses transmitted by planthoppers or leafhoppers vectors (Whitfield et al. 2015).
The only known xylem-limited pathogenic bacteria, Xylella fastidiosa (class Gammaproteobacteria) is transmitted in a non-circulative mode by different sap-feeding insect vectors from Membracoidea, Cercopoidea, and Cicadoidea. All phloem-limited vector-borne bacteria, including phytoplasmas (class Mollicutes), liberibacters (class Alphaproteobacteria), and spiroplasmas (class Mollicutes), are apparently circulative, propagative, colonizing both the plant host and the insect vector intracellularly. The few phytopathogenic spiroplasmas are transmitted by leafhoppers, which are also the main vectors of the many plant diseases caused by phytoplasmas. The few species of phloem-limited and phytopathogenic bacteria of the genus Liberibacter are vectored by psyllids (Perilla-Henao and Casteel 2016).
Although vectors usually acquire viruses by feeding on infected plants, some propagative viruses can be transmitted transovarially, from female insect vector to the progeny. Vector specificity in plant pathogen transmission may vary greatly among plant pathogens. For example, some viruses are transmitted by only one insect vector, whereas other viruses are less vector specific and may be transmitted by insect species from a single family or subfamily (Purcell 2003).
13.5 Hemipteran Sap Suckers as Forest Pests: Damage, Management and Control
13.5.1 Damage
Despite the frequent occurrence and the high species richness of sap suckers on forest trees, in particular aphids, psyllids and scale insects, they have seldom been considered serious pests in natural or planted forests, in their native ranges. Sap suckers feeding in their native forest areas are usually regulated at low population densities by natural enemies. Invasive hemipteran sap suckers from several families, including Psyllidae, Adelgidae, Coccidae, Pseudococcidae, Diaspididae, Matsucoccidae and Monophlebidae have established on forest trees in all major forest areas globally and several of these species have become key forest pests. These species represent a minority of sap suckers associated with forest trees. For example, 191 scale insect species are associated with pines worldwide (García Morales et al. 2016), but only a few have been reported to cause serious damage on pines, leading to intensive defoliation and tree death. All these species had been introduced outside their native range and became established and spread as invasive species. Effectively, scale insects are frequent invaders in forest and agriculture areas. Illustratively, alien scale insect species represent an important component of the European entomofauna, accounting for about 30% of the total scale fauna in Europe (Pellizzari and Germain 2010). Some examples of well-known invasive scale insects are, the maritime pine blast scale M. feytaudi, which is native to southwestern Europe and invasive in Corsica and Italy (Sciarretta et al. 2016); the Israeli bast scale Matsucoccus josephi is native to Cyprus and invasive in Israel (Mendel et al. 2016); and Matsuccocus matsumarae is native to Japan and invasive in North America and China (McClure 1986). Other examples of important sap suckers on pine are Palaeococcus fuscipennis (Monophlebidae), native to southern Europe and invasive in Israel, and the margarodid M. hellenica, native to Turkey and Greece and invasive in Italy and Australia (Mendel et al. 2016; Nahrung et al. 2016). Similarly, for the genus Eucalyptus a number of sap suckers native to Australia have been established worldwide and cause serious damage. In fact, 40% of invasive species affecting Eucalyptus outside Australia are sap suckers. These include 13 species of psyllids, two scale insects, an Eriococcidae and a Diaspididae, a whitefly and the bronze bug, T. peregrinus (Hurley et al. 2016). In their native range, these species, like many other sap suckers feeding on Eucalyptus, exist mostly at endemic population levels and rarely reach outbreak population levels.
Nevertheless, outbreak of populations in its native range also occur. They are often associated with tree physiological status, weather conditions favorable for sap sucker population growth, or disruption of natural enemy populations. For example, high infestations of eucalyptus psyllids, like the red gum lerp psyllid G. brimblecombei, in its native range in Australia, are mainly reported in urban parks or forests where trees are under some kind of physiological stress (Stone 1996). In Europe, native aphids in conifers, such as Cinara spp. and Pineus pini, or on oaks e.g. Lachnus spp. may build up their populations in years with favorable weather conditions. Such outbreaks occur mostly in spring and summer, facilitated by the high fecundity of females and parthenogenetic generations. Normally, these outbreak populations decline as a result of increases in natural enemy populations or unfavorable weather conditions, such as low temperatures. During outbreaks, some of these species produce honeydew in great abundance, a valuable resource for beekeepers, for the production of high value forest honeys (see Table 13.7). Site conditions and silvicultural treatments that result in poor tree health can also promote outbreaks of sap sucking insects. Intense irrigation and fertilization, as well as tree stress, promote tree physiological conditions that may favor such outbreaks. For these reasons, trees in urban and nursery settings are more likely to suffer from intense attacks of sap suckers than their conspecifics in natural forests. In their native range, the importance of sap suckers also frequently increases when host trees are planted in non-natural habitats (urban parks and street trees) due to tree physiological stress and the lack of natural enemies in these fragmented habitats. For example, psyllid outbreaks on eucalyptus in their native range in Australia were often observed on roadsides, farmlands, grazing areas and sewage-irrigated sites (e.g. Collett 2001). Drought-stressed Eucalyptus and Acacia are often more susceptible to psyllids.
Since individuals are usually small, inconspicuous, frequently hiding in crevices of the bark, they are easily spread unknowingly by human activities and wind. When populations are at low densities, individual insects, particularly eggs and young nymphs, are difficult to detect even for experienced eyes. Nursery trees and seedlings, branches and leaves are all plant materials that may easily transport sap suckers. Dissemination by wind occurs mainly during the early developmental stages. Scale insects typically display wind dispersal behavior during the early non-sedentary young nymphs, termed crawlers. For example, wind dispersal of first instar nymphs of the pine blast scale, Matsucoccus spp. facilitates colonization of new areas. Similarly, the elongate hemlock scale Fiorinia externa (Diaspididae) is often dispersed long distances by wind (McClure 1979). Young nymphs of aphids may also disperse up to several thousand meters by wind, as has been observed in the hemlock woolly adelgid Adelges tsugae (McClure 1990). Sap suckers with winged adult females (e.g. psyllids and aphids) may also disperse by flight. In some species, eggs and nymphs may be spread by animals, such as birds or mammals (McClure 1990).
Whereas at low densities sap suckers cause only minor or no harm to individual trees, at high densities they may cause extensive leaf necrosis, discoloration and defoliation triggering reduced tree growth, and increasing susceptibility to tree pathogens and other insect pests. In some cases, such as in C. laportei infesting cedars, sap suckers may kill twigs and branches of infested trees. Ultimately, the activity of sap suckers may result in tree death and high enough levels of tree mortality, to result in forest decline as observed following M. josephi attacks in Aleppo pine in Israel (Mendel et al. 2016) or A. tsugae declining hemlock stands in eastern North America (Ellison et al. 2018). Some sap suckers become particularly problematic in nurseries or greenhouse conditions. This is the case of the oak phylloxera in Israel.
At high densities sap suckers can pose serious problems in urban settings. This is due to aesthetic concerns caused by foliage discoloration, deformation and defoliation, as well as the intense honeydew production and subsequent growth of sooty mold fungi (Fig. 13.2e, i, j). Under these circumstances, the implementation of control tactics is required.
13.5.2 Pest Management
Sap suckers are often difficult to control. Due to their small size and cryptic feeding habits, they are difficult to detect at low population levels. Their high dispersal capacity facilitates the colonization of new hosts. Furthermore, many of these species are protected by cuticular waxes, rendering contact insecticide treatments less successful or ineffective. The implementation of integrated pest management (IPM) strategies, including monitoring, biological control, cultural practices, the use of behavior modifying chemicals, and the injection of systemic insecticides in urban areas, is essential.
13.5.2.1 Behavior Modifying Signals
Behaviors elicited by chemical stimuli (e.g. host plant volatiles (HPV) or pheromones) have been studied for a few sap sucker species feeding on forest trees. These compounds can be used in insect monitoring and different pest management tactics, such as mating disruption, mass trapping, and lure and kill applications, among others.
Sex pheromones of many insects are very powerful attractants, as males may be attracted from long distances to lures impregnated with few micrograms of pheromone (Fig. 13.2a, b). For example, males of Matsucoccus spp. and P. citri are attracted to the female sex pheromone from distances up to several hundred meters, although the level of captures per trap decreases with the distance between the source population and the lures (Branco et al. 2006b).
Sex pheromones have been identified for 32 scale insect species (Franco et al. 2022) and several aphid species (Dewhirst et al. 2010; Pickett et al. 2013). No sex pheromones have been identified to date for forest psyllids or adelgids. There is evidence that psyllids and other sap suckers, such as Cicadellidae, use vibrational signals in sex communication (Čokl and Millar 2009). Although aphids frequently reproduce asexually, in holocyclic species there are populations that reproduce sexually in part of their life cycle. Sex pheromones have been identified for a number of aphid species, all comprising compounds of the group of cyclopentanoid nepetalactones (Dawson et al. 1996). These compounds can act in synergy with plant volatiles. For example, bird-cherry aphid, Rhopalosiphum padi, male response to nepetalactol is synergized by Prunus padus leaf extract. To date, applications of aphid sex pheromones to control or monitor aphid pests have not been developed.
Among scale insects, diaspidid and pseudococcid pheromones are mostly terpenoid derivatives, whereas those of Matsucoccidae are ketones (Zou and Millar 2015). Only a minority of the sex pheromone compounds identified for sap suckers are commercially available, and with one or two exceptions, their use in pest management is still limited, particularly in forestry.
Mating disruption attempts to prevent males from locating females by releasing high concentrations of synthetic sex pheromone in the field. Males may then be unable to locate females and females may remain unmated. A revision of the mechanisms of mating disruption is provided by Evenden (2016). Franco et al. (2022) provided an overview of the current knowledge on mating disruption of scale pests. This technique only applies to populations that reproduce sexually. The success of mating disruption depends on the reproductive behavior of the species, the size of the treatment area and the habitat adjacent to the treatment area. For small-scale plots, immigrant gravid females, coming from adjacent habitats or from alternate host plants supporting populations outside the treatment area, can render the method inefficient. Nevertheless, this is not a problem in the case of scale insects, as the females are wingless insects (Franco et al. 2022). Further, the method is usually more efficient at low population density levels for which the probability of mating will be lower. In theory, the method could be used to increase the frequency of Allee effects, facilitating local extinction of non-native species, when populations are recently established and at very low population levels (Tobin et al. 2011; see also Chapter 18).
Whereas mating disruption has been widely investigated in Lepidoptera, only a few attempts have been made to manage hemipteran pests with mating disruption. Based on the data available on Pherobase (El-Sayed 2018), mating disruption has been investigated in about 127 Lepidoptera species, representing 86% of total species in which mating disruption was tested. However, only nine hemipteran sap suckers were targeted for the same purpose. To note, in particular three scale insects, the vine mealybug Planococcus ficus (Walton et al. 2006), the California red scale Aonidiella aurantii (Rice et al. 1997), and the San Jose scale Comstockaspis perniciosa (=Quadraspidiotus perniciosus) (Bar Zakay et al. 1987). Commercial formulations are currently available for mating disruption of the first two species, in vineyards and citrus orchards, respectively (Cocco et al. 2014; Vacas et al. 2014). The scientific, technological and practical developments in mating disruption of scale insects, as well as future prospects were recently reviewed by Franco et al. (2022). There are no examples in the literature of mating disruption to control sap suckers in forest plantations.
Disruption or manipulation of acoustic signals is a new possibility for pest management of sap sucking insects. Using artificial signals to interfere with acoustic communication between male and female sap suckers has been suggested as a mating disruption tactic to control stink bugs (e.g. Nezara viridula, Euschistus heros) (Čokl and Millar 2009), the Asian citrus psyllid Diaphorina citri (Lujo et al. 2016), and the glassy‐winged sharpshooter Homalodisca vitripennis (Mazzoni et al. 2017).
As a pest management tactic, mass trapping attempts to remove a large portion of the target population by capturing individuals in traps, usually baited with kairomones or pheromones, and eliminating them. A weakness of using sex pheromones is that only males are normally attracted. This hinders the efficiency of the method because even a small fraction of surviving males may be enough to ensure that most females are mated, particularly in polygynous species. Mass trapping of mealybugs and scale insects using sex pheromone was tested for P. citri (Franco et al. 2003) and M. feytaudi (Binazzi et al. 2002). In both cases, the results did not prove the efficiency of the method. This was partly attributed to the small study areas and the attraction of males from the surrounding fields. In Italy, a management program against the pine bast scale M. feytaudi, combining the use of mass trapping and silvicultural interventions, supposedly delayed the loss of a P. pinaster forest caused by the pine bast scale (Sciarretta et al. 2016). One consideration is the cost-effectiveness of the technique, even if there is a substantial reduction of the population, the costs may not compensate for the expected benefits.
Mass trapping with aggregation pheromones (which cause the increase of insect density, usually of both males and females, in the vicinity of pheromone source) is theoretically possible, and could be much more powerful than the use of sex pheromones. Few aggregation pheromones are known for forest sap suckers, most from true bugs (Heteroptera). For example, adult males and nymphs of T. peregrinus display an aggregation behavior induced by a male specific pheromone, whose major component is 3-methylbut-2-enyl butanoate (González et al. 2012).
The lure and kill technique is similar to mass trapping, but instead of trapping the insects, individuals are attracted to a semiochemical-baited lure and then killed by exposure to an insecticide. Although limited by the small surface of lure devices, there may be environmental concerns that limit the use of insecticides in forest settings. To date, there are no examples of lure and kill tactics applied in forestry (El-Sayed 2018).
The dispersal behavior induced in aphids by the alarm pheromone (E)-β-farnesene has been commercially exploited for the control of aphids, by combining its application with an insecticide or entomopathogenic fungi. The induction of dispersal behavior by the application of alarm pheromone is expected to increase the probability of the aphids contacting with the insecticide, thus reducing the effective dosage of the required toxicant. Nevertheless, the efficacy of this control tactic, as well as its cost effectiveness are unclear (Čokl and Millar 2009). An innovative application of the alarm pheromone was recently developed for pest management of aphids. A hexaploid variety of wheat was genetically engineered to release (E)-β-farnesene. Laboratory tests showed that three different aphid species were repelled and the foraging behavior of an aphid parasitoid was enhanced (Bruce et al. 2015).
The natural enemies of a number of sap sucker pests are attracted to the pheromones of their prey (i.e. the pheromones act as kairomones for the natural enemies). In these cases, the pheromone could be used to attract natural enemies and potentially increase predation rates and reduce pest population densities and ultimately crop damage. For example, attraction of parasitic wasps of the citrus mealybug (Franco et al. 2008, 2011) and of different predators of pine blast scales, including brown lacewings (Hemerobiidae), flower bugs (Anthocoridae), ladybirds (Coccinellidae), and flower beetles (Dasytidae) (Branco et al. 2006a; Mendel et al. 2003) has been reported. It is interesting that for most of these predators, both the adult and larval stages were found to be attracted to the pheromone of the prey (Branco et al. 2006c).
Volatile cues emitted from plants and attractive to sap suckers may theoretically be used for monitoring or trapping these insect pests. These include several structural categories of volatiles, such as alcohols, aldehydes, ketones, esters and terpenes. Traps baited with these compounds must compete with volatiles released by host plants and consequently often have low capture rates. In some cases, interactions with other organisms vectored by sap suckers may alter the volatile profile of host trees and increase their attractiveness to vectors. For example, apple trees, infected by the pathogen Phytoplasma mali emit higher amounts of the sesquiterpene β-caryophyllene, and are highly attractive to the vector, Cacopsylla picta (Psyllidae) (Mayer et al. 2011). Similarly, citrus trees infected by the bacterial pathogen Liberibacter asiaticus release odors specifically attracting its psyllid vector Diaphorina citri (Mann et al. 2012). These compounds could potentially be used to control vector populations. So far there are no known examples of this phenomenon with forest sap suckers.
13.5.2.2 Monitoring
Monitoring programs normally target a single species as the deployment protocols of monitoring tools are often species-specific (e.g. habitat, part of the plant affected and distinctive symptoms). Visual surveys are recommended for urban trees and plants in nurseries, and for sap sucker species whose symptoms are easily discernible. Band traps may be used to intercept individuals moving along the tree trunk, such as adult females of the pine bast scale, while they are searching for sites suitable for oviposition (Fig. 13.2c). Interception sticky traps may be used to detect crawlers, when dispersing by wind, or winged adults stages, such as for psyllids. For example, sticky traps were used to monitor the blue gum psyllid, Ctenarytaina eucalypti and the red gum psyllid G. brimblecombei in California (Dahlsten et al. 2003). The performance of sticky traps for monitoring can vary depending on trap color and position in the canopy, as observed for the green spruce aphid Elatobium abietinum, in Sitka spruce (Straw et al. 2011). Therefore, trap design and deployment protocol must be optimized for each species-habitat combination.
Monitoring is probably the most frequent use of sex pheromones in IPM. Pheromone-baited traps can catch individuals when populations are at extremely low levels. Therefore, this highly specific method allows detection of population growth before outbreaks or in the early phases of population establishment. Consequently, managers may apply control methods in an early stage. This allow for timely application of treatment, before populations reach hard-to-control proportions and damage has already occurred. It also avoids increased costs of treatments. As for sticky traps, protocols for deployment of pheromone-baited traps need to be optimized for each species, namely regarding, trap design, size and dosage (e.g. Branco et al. 2004). Pheromone-baited traps are often used to follow seasonal activity of flying males of a given species. Among other applications, this allows managers to anticipate treatment periods and follow generations through seasons. For spreading populations, such as invasive species, monitoring by pheromones allows the tracking of their dispersion rate and range.
13.5.2.3 Biological Control
Many sap sucker pests in forest ecosystems are invasive species or native species whose natural enemies have been locally disrupted. Sap suckers are usually well regulated by natural enemies in their native habitats. Therefore, in many cases classical biological control is the optimal solution for invasive pest species. For example, outbreaks of P. fuscipennis (Monophlebidae) in pine stands in Israel came to an end following the introduction of natural enemies from Spain, such as Novius cruentatus (Coleoptera, Coccinellidae) and Cryptochaetum jorgpastori (Diptera, Cryptochaetidae) (Mendel et al. 1998). Similarly, outbreaks of Cinara spp. in the Mediterranean and South Africa were controlled by introduction of specific parasitic wasps. The horse chestnut scale, Pulvinaria regalis (Coccidae), native to Asia and first detected in Europe in 1968 has become a serious pest of horse chestnuts, as well as many other tree species in Europe (Trierweiler and Balder 2005). Efforts are underway to develop a biological control program using the parasitiod Coccophagus lycimnia (Hymenoptera: Aphellinidae), although this parasitoid is already present in Europe and its origin is unknown. Recent invasion of Victoria and South Australia by the giant pine scale M. hellenica has resulted in serious damage to several introduced pine species (Nahrung et al. 2016). Neoleucopis kartliana (Diptera: Chamaemyiidae), which has already been used to control M. hellenica on the Italian island of Ischia, may be the solution for the scale invasion in South Australia (Avtzis et al. 2020).
In some cases, classical biological control alone does not provide adequate control. The Hemlock woolly adelgid (HWA) A. tsugae, native to East Asia, is one of the most damaging agents on hemlock and spruce trees (Tsuga spp., Picea spp.) in North America. It was unintentionally introduced in the 1950s in the eastern states, spreading to nearby states (McClure et al. 2000). The biological control agent Sasajiscymnus tsugae (=Pseudoscymnus tsugae) (Coleoptera, Coccinelidae) was introduced in North America from Japan in the 1990s. In its natural range, the ladybeetle was considered very prey-specific, keeping the adelgid under control. More than 100,000 adult beetles were released in eastern North America in highly affected forests. Although adelgid densities were reduced in treated areas (McClure and Cheah 1998; McClure et al. 2000), 20 years later the Hemlock woolly adelgid was still the most important threat to the native Tsuga species in the eastern USA (Letheren et al. 2017). Additionally, three species of ladybeetles of the genus Scymnus were first found in China and introduced in USA for studies under quarantine conditions. However, due to mass rearing difficulties, only two species were released in limited numbers and to date there have been no field recoveries of these species (Havill et al. 2014). HWA is also invasive in western North America. Species of Laricobius are specialist predators of adelgids and predation by the native beetle Laricobius nigrinus (Coleoptera: Derodontidae) was thought to limit HWA in western North America. Lamb et al. (2006) proposed its release as a biocontrol agent for eastern HWA populations. Additional species of this genus from both North America and Japan are currently under study for the biocontrol of HWA. A difficulty of classical biological control results from finding adequate natural enemies on the insect pest native range. An example is the Beech scale, Cryptococcus fagisuga (Eriococcidae), invasive in North America. Phylogenetic analysis of this eriococcid suggested that its natural range covers the areas of northeastern Greece, the Black Sea drainage basin, the Caucasus Mountains, and northern Iran (Gwiazdowski et al. 2006). But so far efficient natural enemies have not been found.
Another example of the variability of the success of biological control is seen with the Eucalyptus psyllids. Biological control of the blue gum psyllid C. eucalypti by the Australian parasitoid Psyllaephagus pilosus was achieved in the first year after its introduction in Europe (Chauzat et al. 2002). However, the control of red gum lerp psyllid G. brimblecombei, by another Australian parasitoid Psyllaephagus bliteus was only partially successful both in California and Europe (Dahlsten et al. 2003; Boavida et al. 2016).
Native beneficial organisms may also prey upon non-native forest pests and contribute to regulating their populations (Table 13.6). For example, the predator Elatophilus nigricornis exerts some control upon the non-native pine bast scale M. feytaudi in Corsica. This control is thought to be more effective in mixed forests, where this native natural enemy feeds on a congeneric native scale insect Matsucoccus pini and consequently is expected to have more stable and persistent populations (Jactel et al. 2006). The leaf galling psyllid C. schini feeds on the Peruvian pepper tree, and is invasive in Kenya, where it is heavily parasitized by native eulophid parasitoids, which probably switched from native psyllids developing on Tamarisk (Overholt et al. 2013).
In summary, although biological control is usually a long lasting sustainable solution to control invasive sap suckers it is not always successful. Biological control may not be successful because the agent selected may not be appropriate for the release site. Some natural enemies require several years to establish and build up their populations until effects on target populations are realized. Even after several years of establishment, biocontrol agents may not control pest populations enough to avoid damages. Since landowners wish for inexpensive solutions, quick results and in the short-term, complementary tactics may be needed (e.g. cultural, silvicultural or chemical tactics) until biological control agents are established and providing adequate control.
13.5.2.4 Cultural, Chemical and Physical Methods
Cultural and physical methods may solve problems with temporary outbreaks of sap suckers. Mechanical cleaning of infested trees can reduce populations of sap suckers and reduce damage. Common practices are washing infested branches with soaped water or pruning. These tactics are labor-intensive and costly. Therefore, they are mainly applied in arboretum, parks or urban settings. If these actions are applied regularly, they may reduce sap sucker populations and allow natural enemies to regulate the pest populations. These techniques are mainly applied to individual high-value trees and often the objective is not the control of sap sucker populations but rather protecting the aesthetics of the host plant, or in cases when the scale population spoils the surroundings with honeydew, or causing nuisance high male flight. For example, control of the horse chestnut scale P. regalis in Europe is mostly done by mechanical cleaning in urban areas (Speight et al. 1998). In circumstances where populations are spatially delimited such as nurseries, colored sticky traps may exert some control on psyilld, cicada and leafhopper populations.
Chemical control measures may be used for treating individual street or park trees. Some environmentally friendly methods, such as non-toxic insecticidal soap, plant extracts or horticultural oils are available. In some cases, systemic insecticides may be used with good results to control aphids, psyllids and scale insects. The use of these products in forests is forbidden in many countries and their use is heavily restricted in nurseries. In urban settings, insecticide application might be practiced by soil drenching or trunk injections. The insecticide is transported systemically through the tree vascular system to the foliage or other plant parts where it may kill infesting sap suckers. With soil treatments, care needs to be taken to avoid contamination of water bodies. Of concern is the fact that other insect communities on the tree may be affected by systemic insecticides.
In some cases, host plant resistance plays a major role in tree health. This phenomenon is well known among several Matsucoccus spp., which devastated large pine areas as invasive species, for which pine provenance is a major factor in tree susceptibility (e.g. Mendel 1984; Mendel et al. 2016). Similarly, for HWA host plant resistance plays a major role on population densities of this adelgid, both in the native and introduced areas (Havill et al. 2014). The presence of resistant species or provenances may then help to control the problem. However, lack of resistance is usually a major management challenge and often remains an unresolved situation (e.g. some species of Matsucoccidae, Adelgidae and Eriococcidae). If resistant genetic materials are not available, cultural and silvicultural measures may be suggested as a solution to reduce damage, at least until other, more long-lasting solutions such as biological control, can be developed.
13.6 Conclusions
Sap sucking insects, are characterized by their specialized feeding mode and are quite species diverse in forest trees. By producing honeydew, sap suckers also establish interactions with other forest species, including both vertebrates and invertebrates, which use this sugar resource. The general effect of sucking insects on forest trees is by far much less conspicuous than other major insect groups, like bark beetles and defoliators.
Sucking insects often become major pests under two scenarios: (1) planting of highly susceptible trees outside their native ranges; and (2) introduction or natural spread of these hemipterans outside their native ranges. Invasions by non-native sap suckers often occur without their principal natural enemies. In several cases, the negative effects of these invasive species on tree health have been mitigated by the introduction of natural enemies. On the other hand, range expansion may result in interactions between sap suckers and host trees lacking resistance. These cases may benefit from tree breeding and selection programs. In their native range, damage by sap suckers also increases when the trees are under physiological stress or top-down effects of their natural enemies are disrupted. Understanding the ecology of sap suckers and the factors that promote outbreaks is essential for developing effective control strategies.
References
Afifi S (1968) Morphology and taxonomy of the adult males of the families Pseudococcidae and Eriococcidae (Homoptera: Coccoidea). Bull Br Mus Nat Hist Entomol Suppl 13:1–21
Almeida RP, Nuney LL (2015) How do plant diseases caused by Xylella fastidiosa emerge? Plant Dis 99:1457–1467
Avtzis DN, Lubanga UK, Lefoe GK, Kwong RM, Eleftheriadou N, Andreadi A, Elms S, Shaw R, Kenis M (2020) Prospects for classical biological control of Marchalina hellenica in Australia. Biocontrol 65:413–423
Azevedo MS, Seraglio SKT, Rocha G, Balderas CB, Piovezan M, Gonzaga LV, de Barcellos Falkenberg D, Fett R, de Oliveira MAL, Costa ACO (2017) Free amino acid determination by GC-MS combined with a chemometric approach for geographical classification of bracatinga honeydew honey (Mimosa scabrella Bentham). Food Control 78:383–392
Bar Zakay I, Peleg BA, Hefetz A (1987) Mating disruption of the California red scale Aonidiella aurantii. Hassadeh 69:1228–1231
Baumann P (2005) Biology of bacteriocyte-associated endosymbionts of plant sap-sucking insects. Annu Rev Microbiol 59:155–189
Beirne BP (1956) Leafhoppers (Homoptera: Cicadellidae) of Canada and Alaska. Can Entomol 86:548–553
Ben-Dov Y (1990) The adult female. In: Rosen D (ed) World crop pests, vol. 4A, armored scale insects: their biology, natural enemies and control. Elsevier, Amsterdam, pp 5–20
Ben-Dov Y, Miller DR, Gibson GAP (2014) ScaleNet: a database of the scale insects of the world. http://www.sel.barc.usda.gov/SCALENET/scalenet.htm
Bennett GM, Moran NA (2013) Small, smaller, smallest: the origins and evolution of ancient dual symbioses in a phloem-feeding insect. Genome Biol Evol 5:1675–1688
Binazzi A, Pennacchio F, Francardi V (2002) The use of sex pheromones of Matsucoccus species (Homoptera Margarodidae) for monitoring and mass trapping of M. feytaudi Ducasse and for kairomonal attraction of its natural enemies in Italy. Redia 85:155–171
Blackman RL, Eastop VF (1994) Aphids on the world’s trees. An identification and information guide. CAB International, Wallingford, 1024 pp
Boavida C, Garcia A, Branco M (2016) How effective is Psyllaephagus bliteus (Hymenoptera: Encyrtidae) in controlling Glycaspis brimblecombei (Hemiptera: Psylloidea)? Biol Control 99:1–7
Branco M, Jactel H, Silva EB, Binazzi A, Mendel Z (2004) Effect of trap design, trap size and pheromone dose on male capture of two pine bast scales species (Hemiptera: Matsucoccidae): implications for monitoring and mass-trapping. Agric For Entomol 6:233–239
Branco M, Franco JC, Dunkelblum E, Assael F, Protasov A, Ofer D, Mendel Z (2006a) A common mode of attraction of larvae and adults of insect predators to the sex pheromone of their prey (Hemiptera: Matsucoccidae). Bull Entomol Res 96:179–185
Branco M, Jactel H, Franco JC, Mendel Z (2006b) Modelling response of insect trap captures to pheromone dose. Ecol Model 197:247–257
Branco M, Lettere M, Franco JC, Binazzi A, Jactel H (2006c) Kairomonal response of predators to three pine bast scale sex pheromones. J Chem Ecol 32:1577
Bruce TJA, Aradottir GI, Smart LE, Martin JL, Caulfield JC, Doherty A, Sparks CA, Woodcock CM, Birkett MA, Napier JA, Jones HD, Pickett JA (2015) The first crop plant genetically engineered to release an insect pheromone for defence. Sci Rep 5:11183
Burckhardt D (2005) Biology, ecology, and evolution of gall-inducing psyllids (Hemiptera: Psylloidea). In: Raman A, Schaefer CW, Withers TM (eds) Biology, ecology, and evolution of gall-inducing arthropods. Science Publishers, Enfield, NH and Plymouth, pp 143–157
Carver M, Gross GF, Woodward TE (1991) Hemiptera. In: CSIRO division of entomology, Australia—the insects of Australia, 2nd ed. Melbourne University Press, pp 429–509
Čermák V, Lauterer PC (2008) Overwintering of psyllids in South Moravia (Czech Republic) with respect to the vectors of the apple proliferation cluster phytoplasmas. Bull Insectol 61(1):147–148
Chamorro FJ, Nates-Parra G, Kondo T (2013) Mielato de Stigmacoccus asper (Hemiptera: Stigmacoccidae): Recurso melífero de bosques de roble en Colombia. Rev Colomb Entomol 39:61–70
Chauzat MP, Purvis G, Dunne R (2002) Release and establishment of a biological control agent, Psyllaephagus pilosus for eucalyptus psyllid (Ctenarytaina eucalypti) in Ireland. Ann Appl Biol 141(3):293–304
Chuche J, Auricau-Bouvery N, Danet JL, Thiéry D (2017a) Use the insiders: could insect facultative symbionts control vector-borne plant diseases? J Pest Sci 90:51–68
Chuche J, Sauvion N, Thiéry D (2017b) Mixed xylem and phloem sap ingestion in sheath-feeders as normal dietary behavior: evidence from the leafhopper Scaphoideus titanus. J Insect Physiol 102:62–72
Cocco A, Lentini A, Serra G (2014) Mating disruption of Planococcus ficus (Hemiptera: Pseudococcidae) in vineyards using reservoir pheromone dispensers. J Insect Sci 14:144
Čokl AA, Millar JG (2009) Manipulation of insect signaling for monitoring and control of pest insects. In: Ishaaya I, Horowitz AR (eds) Biorational control of arthropod pests: application and resistance management. Springer, Dordrecht, pp 279–316
Collett N (2001) Biology and control of psyllids, and the possible causes for defoliation of Eucalyptus camaldulensis Dehnh. (river red gum) in south-eastern Australia—a review. J Aust For 64:88–95
Cook L, Gullan P, Trueman H (2002) A preliminary phylogeny of the scale insects (Hemiptera: Sternorrhyncha: Coccoidea) based on nuclear small-subunit ribosomal DNA. Mol Phylogenet Evol 25(1):3–52
Cornara D, Morente M, Markheiser A, Bodino N, Tsai C-W, Fereres A, Redak RA, Perring TM, Spotti Lopes JR (2019) An overview on the worldwide vectors of Xylella fastidiosa. Entomol Gen 39:157–181
Crane E, Walker P (1985) Important honeydew sources and their honeys. Bee World 66:105–112
Crozier LR (1981) Beech honeydew: forest produce. N Z J For 26:200–209
Dahlsten DL, Rowney DL, Robb KL, Downer JA, Shaw DA, Kabashima JN (2003) Biological control of introduced psyllids on Eucalyptus. In: Proceedings of 1st international symposium biological control of arthropods, pp 356–361
Danzig EM (1980) Coccids of the Far East of the USSR (Homoptera, Coccinea) with an analysis of the phylogeny of coccids of the world fauna. Opredeliteli Po Faune SSR 124:1–367
Danzig EM, Gavrilov-Zimin IA (2015) Palaearctic mealybugs (Homoptera: Coccinea: Pseudococcidae). Part 2. Subfamily Pseudococcinae. ZIN RAS, St. Petersburg, 619 pp [Fauna of Russia and neighbouring countries. New series, No. 149. Insecta: Hemiptera: Arthroidignatha]
Dawson GW, Pickett JA, Smiley DW (1996) The aphid sex pheromone cyclopentanoids: synthesis in the elucidation of structure and biosynthetic pathways. Bioorg Med Chem 4(3):351–361
Dewhirst SY, Pickett JA, Hardie J (2010) Aphid pheromones. In: Litwack G (ed) Vitamins and hormones: pheromones, vol 8. Academic Press, London, pp 551–574
Dietrich CH (2003) Auchenorrhyncha (cicadas, spittlebugs, leafhoppers, treehoppers, and planthoppers). In: Resh VH, Cardé RT (eds) Encyclopedia of insects. Academic Press, Amsterdam, pp 66–74
Dinant S, Bonnemain J, Girousse C, Kehr J (2010) Complexité de la séve phloemienne et impact sur l’alimentation des pucerons. CR Biol 333:504–515
Douglas AE (2006) Phloem-sap feeding by animals: problems and solutions. J Exp Bot 57:747–754
Duelli P (1985) A new functional interpretation of the visual system of male scale insects (Coccida, Homoptera). Experientia 41:1036
Ellison AM, Orwig DA, Fitzpatrick MC, Preisser EL (2018) The past, present, and future of the hemlock woolly adelgid (Adelges tsugae) and its ecological interactions with eastern hemlock (Tsuga canadensis) forests. Insects 9(4):172. https://doi.org/10.3390/insects9040172
El-Sayed AM (2018) The pherobase: database of pheromones and semiochemicals. http://www.pherobase.com. Accessed on 19 Aug 2019
Evenden M (2016) Mating disruption of moth pests in integrated pest management. In: Allison JD, Carde RT (eds) Pheromone communication in moths. Evolution, behavior, and application. University of California Press, Oakland, CA, pp 365–393
Federici BA (2003) Pathogens of insects. In: Resh VH, Cardé RT (eds) Encyclopedia of insects. Academic Press, Amsterdam, pp 856–865
Foldi M (2004) The Matsucoccidae in the Mediterranean basin with a world list of species (Hemiptera: Sternorrhyncha: Coccoidea). Ann Société Entomol Fr 40(2):145–168
Favret C, Blackman RL, Miller GL, Victor B (2016) Catalog of the phylloxerids of the world (Hemiptera, Phylloxeridae). ZooKeys 629:83–101
Floren A, Schmidl J (2009) Canopy arthropod research in Europe: basic and applied studies from the high frontier. 2.4.2 Auchenorrhyncha: Cicadoidea and Membracoidea. Bioform, Germany, 576 pp
Franco JC, Gross S, Silva EB, Suma P, Russo A, Mendel Z (2003) Is mass-trapping a feasible management tactic of the citrus mealybug in citrus orchards? An Do Inst Super Agron 49:353–367
Franco JC, Silva EB, Cortegano E, Campos L, Branco M, Zada A, Mendel Z (2008) Kairomonal response of the parasitoid Anagyrus spec. nov. near pseudococci to the sex pheromone of the vine mealybug. Entomol Exp Appl 126(2):122–130
Franco JC, Silva EB, Fortuna T, Cortegano E, Branco M, Suma P, La Torre I, Russo A, Elyahu M, Protasov A, Levi-Zada, A (2011) Vine mealybug sex pheromone increases citrus mealybug parasitism by Anagyrus sp. near pseudococci (Girault). Biol Control 58(3):230–238
Franco JC, Cocco A, Lucchi A, Mendel Z, Suma P, Vacas S, Mansour R, Navarro-Llopis V (2022) Scientific and technological developments in mating disruption of scale insects. Entomol Gen. https://doi.org/10.1127/entomologia/2021/1220
Gamboa MA (2007) Plagas y enfermedades forestales en Costa Rica. Rev For Mes Kurú 4(11):1–69
García Morales M, Denno BD, Miller DR, Miller GL, Ben-Dov Y, Hardy NB (2016) ScaleNet: a literature-based model of scale insect biology and systematics. Database. http://scalenet.info. Accessed on 14 Oct 2017
Giliomee J (1967) Morphology and taxonomy of the adult males of the family Coccidae (Homoptera: Coccoidea). Bull Br Mus Nat Hist Entomol Suppl 7:1–168
Giordanengo P, Brunissen L, Rusterucci C, Vincent C, Van Bel A, Dinant S, Girousse C, Faucher M, Bonnemain JL (2010) Compatible plant-aphid interactions: how aphids manipulate plant responses. Comptes Rendus - Biol 333:516–523
González A, Calvo MV, Cal V, Hernández V, Doño F, Alves L, Gamenara D, Rossini C, Martínez G (2012) A male aggregation pheromone in the bronze bug, Thaumastocoris peregrinus (Thaumastocoridae). Psyche 2012:1–7
Grant PR (2005) The priming of periodical cicada life cycles. Trends Ecol Evol 20(4):169–174
Gullan PJ, Cranston PS (2014) The insects: an outline of entomology. Wiley, Chichester
Gullan PJ, Kosztarab M (1997) Adaptations in scale insects. Annu Rev Entomol 42:23–50
Gullan PJ, Martin JH (2009) Sternorrhyncha: (jumping plant-lice, whiteflies, aphids, and scale insects). In: Encyclopedia of insects, 2nd ed. Academic Press, San Diego, pp 957–967
Gullan PJ, Miller D, Cook L (2005) Gall-inducing scale insects (Hemiptera: Sternorrhyncha: Coccoidea). In: Raman A, Schaefer C, Withers T (eds) Biology, ecology and evolution of gall-inducing arthropods. Science Publishers, Enfield, NH and Plymouth, pp 159–229
Gwiazdowski RA, Van Driesche RG, Desnoyers A, Lyon S, Wu S-A, Kamata N, Normark BB (2006) Possible geographic origin of beech scale, Cryptococcus fagisuga (Hemiptera: Eriococcidae), an invasive pest in North America. Biol Control 39(1):9–18. ISSN 1049-9644. https://doi.org/10.1016/j.biocontrol.2006.04.009
Havill NP, Foottit RG (2007) Biology and evolution of adelgidae. Annu Rev Entomol 52:325–349
Havill NP, Vieira LC, Salom SM (2014) Biology and control of hemlock woolly adelgid. FHTET-2014-05. US Department of Agriculture, Forest Service, Forest Health Technology Enterprise Team, Morgantown, WV, pp 1–21
Hennig W (1981) Insect phylogeny. Wiley, Chichester, 514 pp
Hodkinson ID (1974) The biology of the Psylloidea (Homoptera): a review. Bull Entomol Res 64(2):325–338
Hodkinson ID (2009) Life cycle variation and adaptation in jumping plant lice (Insecta: Hemiptera: Psylloidea): a global synthesis. J Nat Hist 43:65–179
Honey Traveler (2017) Honeydew honey or forest honeys. http://www.honeytraveler.com/types-of-honey/honeydew-forest-honey/. Accessed on 20 Nov 2017
Hong Y, Zhang Z, Guo X, Heie OE (2009) A new species representing the oldest aphid (Hemiptera, Aphidomorpha) from the Middle Triassic of China. J Paleontol 83(5):826–831
Hurley BP, Garnas J, Wingfield MJ, Branco M, Richardson DM, Slippers B (2016) Increasing numbers and intercontinental spread of invasive insects on eucalypts. Biol Invasions 18(4):921–933
Jactel H, Menassieu P, Vetillard F, Gaulier A, Samalens J, Brockerhoff E (2006) Tree species diversity reduces the invasibility of maritime pine stands by the bast scale, Matsucoccus feytaudi (Homoptera: Margarodidae). Can J For Res 36:314–323
Ju R, Li Y, Wang F, Du Y (2009) Spread of and damage by an exotic lacebug, Corythuca ciliata (Say, 1832) (Hemiptera: Tingidae), in China. Entomol News 120(4):409–414
Kawecki Z (1964) On the suitable term for the second pair of wings in male scale insects (Homoptera—Coccoidea). Frustula Entomol 7:1–4
Kay MK (1980) Amphipsalta zelandica (Boisduval), Amphipsalta cingulata (Fabricius), Amphipsalta strepitans (Kirkaldy) (Hemiptera: Cicadidae). Large cicadas. New Zealand Forest Service, Forest and Timber Insects in New Zealand No. 44
Kehr J (2006) Phloem sap proteins: their identities and potential roles in the interaction between plants and phloem-feeding insects. J Exp Bot 57:767–774
Kingsolver JG, Daniel TL (1995) Mechanisms of food handling by fluid-feeding insects. In: Chapman RF, de Boer G (eds) Regulatory mechanisms in insect feeding. Chapman & Hall, New York, pp 32–73
Kosztarab M (1982) Homoptera. In: Parker SP (ed) Synopsis and classification of living organisms. New York: McGraw-Hill, 1232 pp, pp 447–470
Koteja J (1986) Current state of coccid paleontology. Boll Del Lab Di Entomol Agrar Filippo Silvestri 43(suppl.):29–34
Koteja J (1990) Paleontology. In: Rosen D (ed) World crop pests, vol. 4A, armored scale insects: their biology, natural enemies and control. Elsevier, Amsterdam, 384 pp, pp 149–163
Labandeira CC, Phillips TOML (1996) Insect fluid-feeding on Upper Pennsylvanian tree ferns (Palaeodictyoptera, Marattiales) and the early history of the piercing-and-sucking functional feeding group. Ann Entomol Soc Am 89:157–183
Lamb AB, Salom SM, Kok LT, Mausel DL (2006) Confined field release of Laricobius nigrinus (Coleoptera: Derodontidae), a predator of the hemlock woolly adelgid, Adelges tsugae (Hemiptera: Adelgida) in Virginia. Can J For Res 36(2):369–375
Letheren A, Hill S, Salie J, Parkman J, Chen J (2017) A Little bug with a big bite: impact of hemlock woolly adelgid infestations on forest ecosystems in the eastern USA and potential control strategies. Int J Environ Res Public Health 14(4):438
López-Madrigal S, Gil R (2017) Et tu, brute? Not even intracellular mutualistic symbionts escape horizontal gene transfer. Genes 8:1–16
López-Madrigal S, Latorre A, Porcar M, Moya A, Gil R (2013) Mealybugs nested endosymbiosis: going into the “matryoshka” system in Planococcus citri in depth. BMC Microbiol 13:74
Lucas WJ, Groover A, Lichtenberger R, Furuta K, Yadav SR, Helariutta Y, He XQ, Fukuda H, Kang J, Brady SM, Patrick JW, Sperry J, Yoshida A, López-Millán AF, Grusak MA, Kachroo P (2013) The plant vascular system: evolution, development and functions. J Integr Plant Biol 55:294–388
Lujo S, Hartman E, Norton K, Pregmon EA, Rohde BB, Mankin RW (2016) Disrupting mating behavior of Diaphorina citri (Liviidae). J Econ Entomol 109:2373–2379
Maceljski M (1986) Current status of Corythuca ciliata in Europe. EPPO Bull 16(4):621–624
Mann RS, Ali JG, Hermann SL, Tiwari S, Pelz-Stelinski KS, Alborn HT, Stelinski LL (2012) Induced release of a plant-defense volatile ‘deceptively’ attracts insect vectors to plants infected with a bacterial pathogen. PLoS Pathog 8(3):e1002610
Martin JH, Mifsud D, Rapisarda C (2000) The whiteflies (Hemiptera: Aleyrodidae) of Europe and the Mediterranean Basin. Bull Entomol Res 90:407–448, 407
Mayer CJ, Vilcinskas A, Gross J (2011) Chemically mediated multitrophic interactions in a plant–insect vector−phytoplasma system compared with a partially nonvector species. Agric For Entomol 13:25–35
Mazzoni V, Gordon SD, Nieri R, Krugner R (2017) Design of a candidate vibrational signal for mating disruption against the glassy-winged sharpshooter, Homalodisca Vitripennis. Pest Manag Sci 73(11):2328–2333
McClure MS (1979) Self-regulation in populations of the elongate hemlock scale, Fiorinia externa (Homoptera: Diaspididae). Oecologia 39(1):25–36
McClure MS (1986) Role of predators in regulation of endemic populations of Matsucoccus matsumurae (Homoptera: Margarodidae) in Japan. Environ Entomol 15(4):976–983
McClure MS (1990) Role of wind, birds, deer, and humans in the dispersal of hemlock woolly adelgid (Homoptera: Adelgidae). Environ Entomol 19(1):36–43
McClure MS, Cheah CASJ (1998) Released Japanese ladybugs are multiplying and killing hemlock woolly adelgids. Front Plant Sci 50(2):6–8
McClure MS, Cheah CASJ, Tigner TC (2000) Is Pseudoscymnus tsugae the solution to the hemlock woolly adelgid problem? An early perspective. In: McManus KA, Shields KS, Souto DR (eds) Proceedings: symposium on sustainable management of hemlock ecosystems in eastern North America, Durham, New Hampshire, June 22–24, 1999. General Technical Report 267. USDA, Newtown Square, PA, pp 89–96
McKenzie H (1967) Mealybugs of California with taxonomy, biology and control of North American species. University of California Press, Berkeley and Los Angeles, 526 pp
Mendel Z (1984) Provenance as a factor in susceptibility of Pinus halepensis to Matsucoccus josephi (homoptera: Margarodidae). For Ecol Manage 9(4):259–266
Mendel Z, Assael F, Zeidan S, Zehavi A (1998) Classical Biological Control of Palaeococcus fuscipennis (Burmeister) (Homoptera: Margarodidae) in Israel. Biol Control 12:151–157
Mendel Z, Dunkelblum E, Branco M, Franco JC, Kurosawa S, Mori K (2003) Synthesis and structure-activity relationship of diene modified analogs of Matsucoccus sex pheromones. Naturwissenschaften 90(7):313–317
Mendel Z, Protasov A, Jasrotia P, Silva E, Levi-Zada A, Franco J (2011) Sexual maturation and aging of adult male mealybug (Hemiptera; Pseudococcidae). Bull Entomol Res 15:1–10
Mendel Z, Branco M, Battisti A (2016) Invasive sap-sucker insects in the Mediterranean Basin. In: Insects and diseases of Mediterranean forest systems. Springer, Cham, pp 261–291
Mifsud D, Porcelli F (2012) The psyllid Macrohomotoma gladiata Kuwayama, 1908 (Hemiptera: Psylloidea: Homotomidae): a Ficus pest recently introduced in the EPPO region. EPPO Bull 42:161–164
Miller DR (1984) Phylogeny and classification of the Margarodidae and related groups (Homoptera: Coccoidea). In: Kaszab Z (ed) Proceedings of the 10th International Symposium of Central European Entomofaunistics (SIEEC), Budapest, Hungary, 1983. Muzsak Public-Educ., Budapest, 420 pp, pp 321–324
Miller DR, Hodgson CJ (1997) 1.1.3.7 phylogeny. In: World crop pests, vol. 7. Elsevier, Amsterdam, pp 229–250
Misof B, Liu S, Meusemann K, Peters RS., Donath A et al (2014) Phylogenomics resolves the timing and pattern of insect evolution. Science 346(6210)(7):763–767
Morrow JL, Hall AAG, Riegler M (2017) Symbionts in waiting: the dynamics of incipient endosymbiont complementation and replacement in minimal bacterial communities of psyllids. Microbiome 5:58
Morse JG, Hoddle MS (2006) Invasion biology of thrips. Annu Rev Entomol 51:67–89
Nadel RL, Slippers B, Scholes MC, Lawson SA, Noack AE, Wilcken CF, Bouvet JP, Wingfield MJ (2010) DNA bar-coding reveals source and patterns of Thaumastocoris peregrinus invasions in South Africa and South America. Biol Invasions 12(5):1067–1077
Nahrung HF, Loch AD, Matsuki M (2016) Invasive insects in Mediterranean forest systems: Australia. In: Insects and diseases of Mediterranean forest systems. Springer, Cham, pp 475–498
Nair KSS (2001) Exotic forests pest outbreaks in tropical forest plantations: is there a greater threat. https://books.google.co.il/books?isbn=9798764870
Normark B (2003) The evolution of alternative genetic systems in insects. Annu Rev Entomol 48:397–423
Nováková E, Hypša V, Klein J, Foottit RG, von Dohlen CD, Moran NA (2013) Reconstructing the phylogeny of aphids (Hemiptera:Aphididae) using DNA of the obligate symbiont Buchnera aphidicola. Mol Phylogenet Evol 68:42–54
Novotny V, Wilson MR (1997) Why are there no small species among xylem-sucking insects? Evol Ecol 11:419–437
Oddo LP, Piana L, Bogdanov S, Bentabol A, Gotsiou P, Kerkvliet J, Martin P, Morlot M, Valbuena AO, Ruoff K, Von Der Ohe K (2004) Botanical species giving unifloral honey in Europe. Apidologie 35(Suppl. 1):S82–S93
Oliver KM, Degnan PH, Burke GR, Moran NA (2010) Facultative symbionts in aphids and the horizontal transfer of ecologically important traits. Annu Rev Entomol 55:247–266
Overholt WA, Copeland RS, Halbert SE (2013) First record of Calophya schini (Hemiptera: Calophyidae) in Ethiopia and Kenya. Int J Trop Insect Sci 33(4):291–293
Perilla-Henao LM, Casteel CL (2016) Vector-borne bacterial plant pathogens: interactions with hemipteran insects and plants. Front Plant Sci 7:1–15
Pellizzari G, Germain J-F (2010) Scales (Hemiptera, Superfamily Coccoidea). Chapter 9.3. In: Roques A et al (eds) Alien terrestrial arthropods of Europe. BioRisk 4(1):475–510. https://doi.org/10.3897/biorisk.4.45
Pickett JA, Allemann RK, Birkett MA (2013) The semiochemistry of aphids. Nat Prod Rep 30:1277–1283
Pompon J, Quiring D, Goyer C, Giordanengo P, Pelletier Y (2011) A phloem-sap feeder mixes phloem and xylem sap to regulate osmotic potential. J Insect Physiol 57:1317–1322
Preda C, Skolka M (2011) Range expansion of Metcalfa pruinosa (Homoptera: Fulgoroidea) in southeastern Europe. Ecol Balk 3(1):79–87
Purcell AH (2003) Plant diseases and insects. In: Resh VH, Cardé RT (eds) Encyclopedia of insects. Academic Press, Amsterdam, pp 907–912
Redak RA, Purcell AH, Lopes JRS, Blua MJ, Mizell RF III, Andersen PC (2004) The biology of xylem fluid–feeding insect vectors of Xylella fastidiosa and their relation to disease epidemiology. Annu Rev Entomol 49:243–270
Rice RE, Atterholt CA, Delwiche MJ, Jones RA (1997) Efficacy of mating disruption pheromones in paraffin emulsion dispensers. IOBC WPRS Bull 20:151–162
Roff D (1990) The evolution of flightlessness in insects. Ecol Monogr 60:389–421
Ross L, Pen I, Shuker DM (2010) Genomic conflict in scale insects: the causes and consequences of bizarre genetic systems. Biol Rev 85:807–828
Santas LA (1983) Insects producing honeydew exploited by bees in Greece. Apidologie 14:93–103
Schick KN, Dahlsten DL (2003) Gallmaking insects. In: Resh VH, Cardé RT (eds) Encyclopedia of insects. Academic Press, Amsterdam, pp 464–466
Sciarretta A, Marziali L, Squarcini M, Marianelli L, Benassai D, Logli F, Roversi P (2016) Adaptive management of invasive pests in natural protected areas: the case of Matsucoccus feytaudi in Central Italy. Bull Entomol Res 106(1):9–18
Song N, Liang A-P, Bu C-P (2012) A molecular phylogeny of Hemiptera inferred from mitochondrial genome sequences. PLoS ONE 7(11):e48778
Speight MR, Hails RS, Gilbert M, Foggo A (1998) Horse chestnut scale (Pulvinaria regalis) (Homoptera: Coccidae) and urban host tree environment. Ecology 79:1503–1513
Spodek M, Ben-Dov Y, Mendel Z (2013) The scale insects (Hemiptera: Coccoidea) of oak trees (Fagaceae: Quercus spp.) in Israel. Isr J Entomol 43:95–124
Stone, C (1996) The role of psyllids (Hemiptera: Psyllidae) and bell miners (Manorina melanophrys) in canopy dieback of Sydney blue gum (Eucalyptus saligna Sm.). Austral Ecol 21(4):450–458
Straw NA, Williams DT, Green G (2011) Influence of sticky trap color and height above ground on capture of Alate Elatobium abietinum (Hemiptera: Aphididae) in sitka spruce plantations. Environ Entomol 40(1):120–125
Styrsky JD, Eubanks MD (2007) Ecological consequences of interactions between ants and honeydew-producing insects. Proc R Soc B: Biol Sci 274:151–164
Tobin PC, Berec L, Liebhold AM (2011) Exploiting Allee effects for managing biological invasions. Ecol Lett 14(6):615–624
Trierweiler T, Balder H (2005) Spread of horse chestnut scale (Pulvinaria regalis) in Germany. In: Symposium article. Plant protection and plant health in Europe. Introduction and spread of invasive species. Humboldt University, Berlin, Germany
Vacas S, Alfaro C, Primo J, Navarro-Llopis V (2014) Deployment of mating disruption dispensers before and after first seasonal male flights for the control of Aonidiella aurantii in citrus. J Pest Sci 88:321–329
Vandegehuchtea ML, Wermelingerc B, Fraefeld M, Baltensweilerd A, Düggelind C, Brändlid U-B, Freitage A, Bernasconie C, Cherixf D, Rischa AC (2017) Distribution and habitat requirements of red wood ants in Switzerland: implications for conservation. Biol Conserv 212:366–375
Vea I, Grimaldi D (2016) Putting scales into evolutionary time: the divergence of major scale insect lineages (Hemiptera) predates the radiation of modern angiosperm hosts. Sci Rep 6:23487
Vea I, Tanaka S, Shiotsuki T, Jouraku A, Tanaka T, Minakuchi C (2016) Differential juvenile hormone variations in scale insect extreme sexual dimorphism. PLoS ONE 11(2):e0149459
Walton VM, Daane KM, Bentley WJ, Millar JG, Larsen TE, Malakar-Kuenen R (2006) Pheromone-based mating disruption of Planococcus ficus (Hemiptera: Pseudococcidae) in California vineyards. J Econ Entomol 99(4):1280–1290
Way MJ (1963) Mutualism between ants and honeydew-producing Homoptera. Annu Rev Entomol 8:307–344
Whitfield AE, Falk BW, Rotenberg D (2015) Insect vector-mediated transmission of plant viruses. Virology 479:278–289
Will T, Furch A, Zimmermann M (2013) How phloem-feeding insects face the challenge of phloem-located defenses. Front Plant Sci 4:336
Wool DL (2005) Gall-inducing aphids: biology, ecology, and evolution. In: Raman A, Schaefer C, Withers T (eds) Biology, ecology, evolution of gall-inducing arthropods. Science Publishers, Enfield, NH and Plymouth, pp 73–132
Wood DL, Storer AJ (2003) Forest habitats. In: Resh VH, Cardé RT (eds) Encyclopedia of insects. Academic Press, Amsterdam, pp 442–454
Zina V, Lima A, Caetano F, Silva EB, Ramos AP, Franco JC (2012) First record of the pepper tree psyllid, Calophya schini Tuthill (Hemiptera, Calophyidae), in the Palaearctic region. Phytoparasitica 40:127–130
Zou Y, Millar JG (2015) Chemistry of the pheromones of mealybug and scale insects. Nat Prod Rep 32(7):1067–1113
Zytynska SE, Weisser WW (2016) The natural occurrence of secondary bacterial symbionts in aphids. Ecol Entomol 41:13–26
Acknowledgements
We thank Alex Protasov and Vera Zina for allowing the use of some photos. Thanks are also due to the editors and reviewers for their comments and suggestions that greatly helped us improving the manuscript. This work was funded by the Forest Research Centre (CEF), a research unit funded by Fundação para a Ciência e a Tecnologia I.P. (FCT), Portugal (UIDB/00239/2020), and by the project “Technical cooperation, knowledge sharing and training in the area of forest and agricultural entomology and integrated pest management” funded by KKL – JNF Israel.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is licensed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license and indicate if changes were made.
The images or other third party material in this chapter are included in the chapter's Creative Commons license, unless indicated otherwise in a credit line to the material. If material is not included in the chapter's Creative Commons license and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder.
Copyright information
© 2023 The Author(s)
About this chapter

Cite this chapter
Branco, M., Franco, J.C., Mendel, Z. (2023). Sap-Sucking Forest Pests. In: D. Allison, J., Paine, T.D., Slippers, B., Wingfield, M.J. (eds) Forest Entomology and Pathology. Springer, Cham. https://doi.org/10.1007/978-3-031-11553-0_13
Download citation
DOI: https://doi.org/10.1007/978-3-031-11553-0_13
Published:
Publisher Name: Springer, Cham
Print ISBN: 978-3-031-11552-3
Online ISBN: 978-3-031-11553-0
eBook Packages: Biomedical and Life SciencesBiomedical and Life Sciences (R0)