
Abstract
In deep-sea hydrothermal vent fields, faunal distribution is associated with geochemical environments generated by hydrothermal vent activity. The Okinawa Trough is located between the Eurasian Continent and the Ryukyu Arc, and is characterized by sediment-associated fauna associated with vents. In this chapter, the faunal distribution in hydrothermal vent fields in the Okinawa Trough is summarized at inter- and intra-field levels, and its relationship with geochemical environments and species-specific ecologies is discussed. Although the zonation of sediment-associated fauna is not distinct, the fauna on rocky seabed can be categorized into four zones based on thermal conditions. Genetic connectivity among conspecific populations in the Okinawa Trough appears to be common; further, a few faunal groups, such as tubeworms, show connectivity to regions outside of the Okinawa Trough. The faunal composition of vent communities in the Okinawa Trough has been categorized into three groups: the Minami-Ensei Knoll, Yoron Hole, and middle-southern Okinawa Trough. To determine more precisely the relationships between faunal composition and environmental factors in the Okinawa Trough vent fields, both generalized and vent-specific environmental factors should be measured simultaneously with quantitative faunal sampling for analyses.
You have full access to this open access chapter, Download chapter PDF
Similar content being viewed by others


Keywords
1 Background
Deep-sea hydrothermal vents are among the most flourishing deep-sea environments, due to the associated chemosynthetic bacterial productivity. The Okinawa Trough is a backarc basin located between the Eurasian continent and the Ryukyu island arc. The first hydrothermal vent field in the Okinawa Trough was discovered in the 1980s at the JADE site, Izena Hole (Halbach et al. 1989), and geological and geochemical knowledge has been accumulated since then. In total, eight hydrothermal vent fields, i.e. the Minami-Ensei Knoll, the Yoron Hole, the Iheya Ridge, the Iheya North Knoll, the Izena Hole, Irabu Knoll, the Hatoma Knoll and the Daiyon-Yonaguni Knoll, have been discovered to date in the Okinawa Trough, and exploration is still in progress (see Fukuba et al. Chaps. 38 and 39; Yanamaka et al. Chap. 40). Rifting of the Okinawa Trough appears to have started from the southern end. In terms of water columns, the Kuroshio Current flows from the southern end of the Okinawa Trough to the Kerama Gap or to the Japan Sea. The details of the vent fields in the Okinawa Trough are summarized by Ishibashi et al. (Chap. 29).
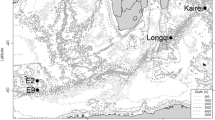
The faunal assemblages associated with hydrothermal vents in the Okinawa Trough were first summarized for the Iheya Ridge (Ohta 1991; Kim and Ohta 1991) and updated by Hashimoto et al. (1995) with extensive descriptions of the vent fauna in the Minami-Ensei Knoll and comparisons with vent fauna in the north (Minami-Ensei Knoll) and middle (Iheya Ridge) parts of the Okinawa Trough (Fig. 34.1). In the early 2000s, reports described vent fauna in the southern part of the Okinawa Trough (Tsuchida et al. 2000; Fujikura et al. 2001). On the basis of these descriptions, the fauna associated with hydrothermal vent fields in the Okinawa Trough appear to show similarities to those in methane seep sites in Sagami Bay due to the dense sedimentation (Fujikura et al. 1995; Watanabe et al. 2010).

Distribution of deep-sea hydrothermal vent fields in the Okinawa Trough. Me Minami-Ensei Knoll (D = 600–800 m), Yr Yoron Hole (D = 580 m), Ik Iheya North Knoll (D = 900–1,000 m), Ih Iheya Ridge (D = 1,300–1,500 m), Iz Izena Hole (D = 1,300–1,600 m), Ir Irabu Knoll (D = 1,650 m), Ht Hatoma Knoll (D = 1,400–1,530 m), Yg Daiyon-Yonaguni Knoll (D = 1,320–1,387 m)
In this paper, we summarize the previous research and recent results from the TAIGA project on vent fauna in the Okinawa Trough, to focus on (1) species distribution and population connectivity among the vent fields, (2) the zonation of fauna around vents, and (3) faunal similarities among vent fields in the Okinawa Trough.
2 Distribution and Connectivity of Vent Fauna in the Okinawa Trough
Vent fauna in the Okinawa Trough have been summarized previously (Kojima 2002; Fujikura et al. 2008; Watanabe et al. 2010); however, the community structure is not yet fully understood due to a lack of quantitative information. Recent comprehensive DNA barcoding analyses have revealed the existence of cryptic species, or morphologically similar species, particularly among the polychaetes and small gastropods. On the other hand, the possibility of synonymy is arising. Therefore, verification of the distributional records of vent fauna in the Okinawa Trough is still in progress. To date, a total of 109 species has been reported in and near vent fields in the Okinawa Trough (summarized in Table 34.1). In this section, previous reports are briefly summarized for the three major taxa (Annelida, Arthropoda, and Mollusca), together with recent distributional remarks and results of genetic analyses from the TAIGA project, which infer population connectivity.
Annelida
In total, 36 species have been reported in the Okinawa Trough hydrothermal vent fields, but as yet no species strictly endemic to the region have been recognized. Vestimenteiferan tubeworms are among the conspicuous fauna in the vent fields. Three species have been separately reported in the Okinawa Trough vent fields, but these are also found in the methane seep faunas: Lamellibrachia sp. in the Iheya Ridge and the Iheya North Knoll; Paraescarpia echinospica in the Iheya Ridge; and two lineages of Alaysia, A2 in the Daiyon-Yonaguni Knoll and A4 in the Iheya Ridge, the Iheya North Knoll and the Izena Hole (Fujikura et al. 2008). Two unidentified species of tubeworm were reported in Depression B of the Minami-Ensei Knoll (Hashimoto et al. 1995; Fig. 34.2a). The tubeworm clump possibly consisted of Alaysia and Lamellibrachia; however, identification to the species level has been impossible. Population genetic analyses of tubeworms showed no differences among the populations in the Okinawa Trough, or from those of populations outside of the Okinawa Trough (Watanabe et al. 2010).

Typical vent fauna in the Okinawa Trough. (A) tubeworm clump in the Minami-Ensei Knoll, (B) typical rocky fauna in the Minami-Ensei Knoll, (C) vent associated fauna in the Yoron Hole, (D) peripheral vent fauna in the Izena Hole, (E) high-temperature vent fauna in the Izena Hole, (F) barnacle-dominated zone in the Irabu Knoll, (G) typical rocky fauna in the Irabu Knoll, (H) typical rocky fauna in the Hatoma Knoll
Other than tubeworms, the connectivity among populations of annelids has yet to be examined. For the polychaetes, classification of the collected animals is not sufficient for a comparison of the faunal composition among the vent fields. Ampharetid polychaetes are, however, abundant in the middle and southern parts of the Okinawa Trough, and are frequently associated with a squat lobster, Shinkaia crosnieri (Fig. 34.2). Recent DNA barcoding indicates the existence of at least three different clusters ampharetid polychaetes in the Okinawa Trough (data not shown), probably including the “ampharetid sp.” reported by Hashimoto et al. (1995). Two Eunice species have been described (E. masudai and E. northioidea), and the existence of further species in this genus has been indicated (Hashimoto et al. 1995). As most of the species-level composition of polychaete fauna in the Okinawa Trough is still incomplete, we have only identified them to the genus level. However, it is apparent that the diversity of vent polychaetes in the Okinawa Trough is underestimated, and a detailed re-examination is accordingly required.
Arthropoda
In total, 23 species have been reported in the Okinawa Trough hydrothermal vent fields, and three dominant species, Shinkaia crosnieri; a shrimp with a pair of unfused dorsal eyes, Shinkaicaris leurokolos; and a shrimp with stalked eyes, Lebbeus shinkaiae, are treated as endemic to this region. No bythograeid crabs have been reported.
Shinkaia crosnieri has been observed in most vent fields and is among the representative animals of the Okinawa Trough, although its abundance decreases in shallower vent fields such as the Minami-Ensei Knoll and it is absent from the Yoron Hole. S. crosnieri harbors bacterial symbionts in its ventral side (Watsuji et al. 2010) and the ecological characteristics are similar to those of Kiwa (Roterman et al. 2013). Genetic diversity is relatively high (Kumagai et al., Chap. 5), probably due to its high abundance caused by frequent reproduction to sustain a huge biomass.
Alvinocaridid shrimps are the second most dominant crustaceans in the Okinawa Trough vent fields; however, the two reported genera, Alvinocaris and Shinkaicaris, have different distributions (Komai and Segonzac 2005). The Alvinocaris shrimps have been reported in a wide area in the Okinawa Trough: A. dissimilis in the Minami-Ensei Knoll, and A. longirostris in all the other vent fields except for the Yoron Hole (Fujikura et al. 2008). On the other hand, Shinkaicaris leurokolos, whose distribution was thought to be restricted to the Minami-Ensei Knoll, now appears to be distributed in all the known vent fields in the Okinawa Trough (Yahagi per. com).
Lebbeus shinkaiae, which is a recently described shrimp, but which has also been reported as Lebbeus washingtonius or Lebbeus sp. (Komai et al. 2012), is also widely distributed, but is only reported from the Okinawa Trough. This shrimp is not aggregated and is commonly distributed near peripheral areas of vent fields.
A barnacle of the genus Neoverruca is patchily but abundantly distributed in all the Okinawa Trough vent fields except for the Minami-Ensei Knoll and Yoron Hole. Among the local populations in the Okinawa Trough, no genetic differences have been detected and their population expansion date has been estimated to be earlier than that in the Izu-Ogasawara Arc vent fields (Watanabe et al. 2005). Rearing experiments on larvae of this barnacle revealed that high temperature surface water may act as a dispersal barrier for this species (Watanabe et al. 2006).
Mollusca
In total, 34 species have been reported in the Okinawa Trough hydrothermal vent fields, and most vent gastropod species are thought to be endemic to this region.
Large clams of the genus Calyptogena s.l. are distributed only in sedimentary fields (Kim and Ohta 1991) in the hydrothermal vent fields of Minami-Ensei Knoll and Iheya Ridge and Iheya North Knoll in the Okinawa Trough. Hitherto, three species have been reported, and the population genetic structure has been examined for C. kawamurai and C. okutanii (Watanabe et al. 2010, based on the data in Kojima et al. 2001, 2006). In both cases, populations in the Okinawa Trough were significantly different from those outside of the Okinawa Trough. No differences were detected between C. okutanii populations in vent fields of the Iheya Ridge and Iheya North Knoll.
Bathymodiolus mussels are among the representative animals of the Okinawa Trough, and three species are distributed in the different ranges: B. japonicus in Minami-Ensei Knoll (Fig. 34.2b), B. aduloides in the Minami-Ensei Knoll and Iheya Ridge, and B. platifrons in all the vent fields except for the Minami-Ensei Knoll and Yoron Hole (Fig. 34.2, Table 34.1). The population connectivity of B. platifrons was examined using partial sequences of mitochondrial NADH dehydrogenase 4 (ND4), and the most dominant haplotype was shared between populations inside and outside of the Okinawa Trough (Kyuno et al. 2009), inferring connectivity between the populations.
Provanna snails are the most abundant macrofauna in the Okinawa Trough vent fields. They are associated with both hard and soft substrata, both in vents and seeps (Fujikura et al. 2008). The Provanna gastropods in vent fields in the Okinawa Trough were first referred to as P. glabra based on the populations in Sagami Bay, but certain morphological differences were identified; therefore, they have been referred to as P. aff. glabra for more than 10 years (Fujikura et al. 2008). Morphometric and DNA barcoding analyses is now in progress and it shows the possibility that more than 2 lineages of Provanna gastropods are distributed in the Okinawa Trough.
A small limpet, Lepetodrilus nux, is a species endemic to the Okinawa Trough and has been found in all of the vent fields except for the Yoron Hole. It is sometimes associated with S. crosnieri and B. platifrons, but can also be found on inorganic hard substrata. Lepetodrilus japonicus was described from a vent field in the Minami-Ensei Knoll, but a recent investigation could not determine its distribution in Depression C of the knoll. On the other hand, L. nux appears to be distributed in the Minami-Ensei Knoll in addition to the previously known vent fields (Nakamura et al. 2014).
A small snail, Iheyaspira lequios, is a species only found in hydrothermal vent fields in the Okinawa Trough. Recently, a second species of the genus Iheyaspira, I. bathycodon, was reported from the Mid-Cayman Rise in the Caribbean Sea (Nye et al. 2013). This patchy worldwide distribution of a vent-endemic genus is quite rare, as there are many vent fields located between the Okinawa Trough and the Caribbean Sea. Iheyaspira gastropods are usually smaller than 5 mm in shell width, in the Okinawa Trough, and may be missed or misidentified due to their small size. A detailed examination of both morphology and genetics is required to understand the distributional pattern of Iheyaspira gastropods, not only in the Okinawa Trough but also on a global scale.
Other Phyla
Poriferans, anthozoans, echinoderms, and fishes are common marine fauna but not well described for the areas near vent fields in the Okinawa Trough. Poriferans are abundant in the Minami-Ensei Knoll (Fig. 34.2a), but to date no species have been identified. Some echinoderms such as Ceramaster asteroids and unidentified ophiuroids have been reported in Depression B in the Minami-Ensei Knoll and the CLAM site in the Iheya Ridge (Hashimoto et al. 1995; Kim and Ohta 1991). Recently, the reproduction of a liparid fish, Careproctus rhodomelas, in the Hatoma Knoll was reported (Takemura et al. 2010); however, for the other fishes, almost nothing has been reported due to the difficulty of sampling. These fauna may be important as predators in such ecosystems, and detailed observations are required for elucidating trophic interactions.
3 Faunal Zonation in a Vent Field
Vent fauna show variation along steep environmental gradients, or patchiness, which is referred to “zonation.” In the Okinawa Trough, hydrothermal vents host two types of fauna: rocky seabed fauna associated with direct vent activity (rock-associated fauna), and epi- and infauna indirectly supported by vents (sediment-associated fauna). Generally, faunal composition differs among substrata (Tunnicliffe et al. 2003). These differences in the vent fields in the Okinawa Trough were first mentioned by Kim and Ohta (1991) based on the observation of vent fauna in the CLAM (sedimentary) and PYRAMID (rocky) sites of the Iheya Ridge. Subsequently, two neighboring vent sites with differing fauna were reported in Depression B (sedimentary) and C (rocky) in the Minami-Ensei Knoll (Hashimoto et al. 1995). This section summarizes zonations in the rocky and sedimentary habitats in the vent fields in the Okinawa Trough.
Rock-Associated Fauna
Rocky fauna are generally dominant in deep-sea hydrothermal vent fields. The zonation sequence of epifauna on the rocky substrata has been categorized into 4 zones in the Iheya North Knoll: Zone 1 (<0.2 m from vent), no fauna except for occasional invasion by shrimps; Zone 2 (0.2–0.8 m from vent), Shinkaia crosnieri aggregation with shrimps; Zone 3 (0.8–2.5 m from vent), co-occurrence of S. crosnieri and Bathymodiolus platifrons with two limpets (Bathyacmaea secunda and Lepetodrilus nux) and shrimps; and Zone 4 (>2.5 m from vent), B. platifrons without S. crosnieri (Tokeshi 2011). Additional observations have updated the faunal composition of each zone (Figs. 34.2 and 34.3). Zone 1, nearest to hydrothermal venting, is inhabited by a dense assemblage of Paralvinella aff. hessleri with white tubes (Fig. 34.2e), although congeneric P. aff. unidentata without tubes are scattered near vents. The surrounding area (Zone 2) is inhabited by Shinkaicaris leurokolos and S. crosnieri. S. crosnieri is sometimes associated with L. nux and ampharetid polychaetes and red copepods. Outside of the S. leurokolos–S. crosnieri assemblage (Zone 4), B. platifrons-hosted fauna are distributed. Alvinocaris longirostris, a shrimp without a dorsal eye, and the limpets B. secunda and L. nux are associated with this fauna. The transitional zone from Zone 2 to Zone 4 is Zone 3, which shares fauna with both zones.

Summary of zonation of vent fauna in the Okinawa Trough. ND not determined
For the zonation associated with hydrothermal vents, temperature must be an important factor. There is some evidence to show that temperature contributes to controlling faunal zonation. For instance, in the Okinawa Trough, the difference in temperature effects was also observed for two vent barnacles; Ashinkailepas seepiophila was found on a dead chimney and Neoverruca sp. on an active chimney, as the effects of rearing temperature on larval duration of these two species were different (Yorisue et al. 2013). Other environmental factors, such as salinity, dissolved oxygen (DO), and the concentration of H2S, may contribute to the zonation, but there is no supportive evidence for this from the northwestern Pacific. In vent fields in the southwestern Pacific, sulfide concentration and DO, in addition to temperature, appear to contribute to the gradient distribution of vent fauna (Podowski et al. 2010).
Sediment-Associated Fauna
Sedimentary fauna associated with vents is found in the northern and middle part of the Okinawa Trough. This fauna differentiate the vent fauna in the Okinawa Trough from that of other vent fields in the northwestern Pacific and contribute to the species diversity of the Okinawa Trough. The zonation sequence of sediment-associated fauna is still unclear as it is difficult to stratify sediment with infauna to determine the relationships between environmental gradients and faunal distribution in a vertical spectrum. On the other hand, horizontal zonation has been observed in some methane seep sites. The relationships between the concentration of methane and faunal distribution have been discussed both from the perspective of a toxin for animals and as a substrate for chemosynthetic production (e.g. Barry et al. 1997). However, in general, faunal distribution in marine environments is explained by more generalized factors, such as temperature, salinity, and the concentration of DO. Indeed, the distribution of Calyptogena s.l. assemblages associated with hydrothermal vents and methane seeps in the Okinawa Trough and Sagami Bay could be explained by temperature and salinity (Watanabe et al. 2013). Although the vertical distribution of infauna is still unclear, the importance of infauna as a disturber of sediment has been noted in deep-sea reducing environments (Seike et al. 2012).
Vesicomyid or Calyptogena s.l. clams and solemiyd clams, along with tubeworms, are dominant in the sedimentary fauna in the Okinawa Trough vent fields. Recently, Solemya flava was described from a sedimentary vent field in the Iheya Ridge (Sato et al. 2013). The main difference from rocky fauna is that most sedimentary fauna are shared between methane seep sites in Sagami Bay and Nankai Trough (Watanabe et al. 2010). This can be attributed to the similarity of vent fauna in the Okinawa Trough to the methane seep fauna, in addition to the high concentration of methane and CO2 in vent fluids (Sakai et al. 1990).
4 Faunal Resemblance in the Okinawa Trough
To show the overall similarity among the vent fauna in the Okinawa Trough, Jaccard’s similarity index was calculated based on the species occurrence data (Table 34.1), and the results were presented using group-average clustering analysis (Fig. 34.4). Faunal similarity was lower than 60 % between all the vent fields, but the vent fauna was divisible into three groups at the 20 % similarity index: the Minami-Ensei Knoll, the Yoron Hole, and the others locations in the middle to the southern Okinawa Trough. Recent research has suggested that, although some fauna appear to be shared between the vent faunas in the Minami-Ensei Knoll and the other vent fields in the Okinawa Trough (e.g., Shinkaicaris leurokolos and Lepetodrilus nux), the fauna in the Minami-Ensei Knoll still retains high endemicity. However, the information on the sediment-associated fauna in the Minami-Ensei Knoll (Depression B) was not sufficient to examine the faunal resemblance. In the middle to southern Okinawa Trough, the faunal composition was relatively similar to the neighboring vent fields. The cluster of the Iheya Ridge and Iheya North Knoll vent fauna is distinguished from those in the Izena Hole and Irabu, Hatoma and Daiyon Yonaguni Knolls because it included sediment-associated fauna.

Results of clustering analysis of faunal similarity among the eight vent fields listed in Table 34.1
There are several factors that contribute to inter-field faunal resemblance. In general, the diversity of deep-sea fauna varies according to depth (Rex and Etter 2010). Similar observation was made for deep-sea bivalves in chemosynthetic environments because the species show restricted ranges of vertical distribution (Fujikura et al. 2008). Indeed, the clusters of vent fauna in the Okinawa Trough are correlated with the depth range of each vent field (Fig. 34.4). The depth variation may act as both a dispersal barrier (pre-settlement) and physiological barrier (post-settlement). General oceanographic factors such as ambient temperature, salinity, and DO, are more important than the concentration of reduced chemicals in endmember fluid to field-level similarity, as the latter are quickly diluted by ambient seawater and seem to contribute to the species microdistribution in a vent field. To determine more precisely the relationships between faunal composition and environmental factors in the vent fields, both generalized and vent-specific environmental factors should be measured simultaneously for the analyses.
5 Conclusions
In this chapter, we summarized and updated the information on vent fauna in the Okinawa Trough. A total of 109 species are listed here; however, that number may be either an over- or under-estimate. Detailed morphological and genetic examination of vent fauna, particularly for polychaetes and small gastropods such as Iheyaspira, are required to confirm the existence of synonymous or cryptic species. The composition of sediment-associated fauna is genetically similar to that in methane seep sites outside of the Okinawa Trough, but its zonation is still unclear, as it’s elucidation requires 3-dimensional observation of the sediment. On the other hand, the zonation of rock-associated fauna has been clearly determined, and most endemic species of the Okinawa Trough are found among the rock-associated fauna. The total diversity of vent fauna in the Okinawa Trough is attributable to the associated sediment-associated fauna and the wide depth range causing variation in both ambient temperature and that near hydrothermal vents. Although more than 20 years have passed since the discovery of vent fauna in the Okinawa Trough, further investigation is still required to understand their local biogeography.
References
Amante C, Eakins BW (2009) ETOPO1 1 arc-minute global relief model: procedures, data sources and analysis, NOAA technical memorandum NESDIS NGDC-24, National Geophysical Data Center, Boulder, Colorado, p 19
Barry JP, Kochevar RE, Baxter CH (1997) The influence of porewater chemistry and physiology on the distribution of vesicomyid clams at cold seeps in Monterey Bay: implications for patterns of chemosynthetic community organization. Limnol Oceanogr 42:318–328
Fujikura K, Hashimoto J, Fujiwara Y, Okutani T (1995) Community ecology of the chemosynthetic community at Off Hathushima site, Sagami Bay, Japan. JAMSTEC J Deep Sea Res 11:227–241 (In Japanese with English abstract and captions)
Fujikura K, Fujiwara Y, Ishibashi J, Katayama S, Komatsu T, Maezawa Y, Maki Y, Miyazaki J-I, Miyake H, Okamoto K, Okoshi K, Toth LG, Tsuchida S, Yamaguchi T, Yamanaka T, Watabe H, Watanabe H, Zielinski S, Kato K (2001) Report on investigation of hydrothermal vent ecosystems by the crewed submersible ‘Shinkai 2000’ on the Dai-yon (No. 4) Yonaguni Knoll and the Hatoma Knoll, the Okinawa Trough. JAMSTEC J Deep Sea Res 19:141–154 (In Japanese with English abstract and captions)
Fujikura K, Okutani T, Maruyama T (2008) Deep-sea life – biological observations using research submersibles. Tokai University Press, Kanagawa
Halbach P, Nakamura K, Wahsner M, Lange J, Sakai H, Käselitz L, Hansen RD, Yamano M, Post J, Prause B, Seifert R, Michaelis W, Teichmann F, Kinoshita M, Märten A, Ishibashi J, Czerwinski S, Blum N (1989) Probably modern analogue of Kuroko-type massive sulphide deposits in the Okinawa Trough back-arc basin. Nature 338:496–499
Hashimoto J, Ohta S, Fujikura K, Miura T (1995) Distribution pattern and biogeography of the hydrothermal vent communities of the Minami-Ensei Knoll in the Mid-Okinawa Trough, Western Pacific. Deep Sea Res I 42:577–598
Kim DS, Ohta S (1991) Submersible observations and comparison of the biological communities of the two hydrothermal vents on the Iheya Ridge of the Mid-Okinawa Trough. JAMSTEC Tech Rep Deep Sea Res 221–253 (In Japanese with English abstract and captions)
Kojima S (2002) Deep-sea chemosynthesis-based communities in the northwestern Pacific. J Oceanogr 58:343–363
Kojima S, Ohta S, Yamamoto T, Miura T, Fujiwara Y, Hashimoto J (2001) Molecular taxonomy of vestimentiferans of the western Pacific and their phylogenetic relationship to species of the eastern Pacific. I. Family Lamellibrachiidae. Mar Biol 139:211–219
Kojima S, Tsuchida E, Numanami H, Fujikura K, Okutani T (2006) Synonymy of Calyptogena solidissima with Calyptogena kawamurai (Bivalvia: Vesicomyidae) and its population structure revealed by mitochondrial DNA sequences. Zool Sci 23:835–842
Komai T, Segonzac M (2005) A revision of the genus Alvinocaris Williams and Chace (Crustacea: Decapoda: Caridea: Alvinocarididae), with descriptions of a new genus and a new species of Alvinocaris. J Nat History 39:1111–1175
Komai T, Tsuchida S, Segonzac M (2012) Records of species of the hippolytid genus Lebbeus White, 1847 (Crustacea: Decapoda: Caridea) from hydrothermal vents in the Pacific Ocean, with descriptions of three new species. Zootaxa 3241:35–63
Kyuno A, Shintaku M, Fujita Y, Matsumoto H, Utsumi M, Watanabe H, Fujiwara Y, Miyazaki J-I (2009) Dispersal and differentiation of deep-sea mussels of the genus Bathymodiolus (Mytilidae, Bathymodiolinae). J Mar Biol. doi:10.1155/2009/625672, Article ID 625672
Nakamura M, Watanabe H, Sasaki T, Ishibashi J, Fujikura K, Mitarai S (2014) Life history traits of Lepetodrilus nux in the Okinawa Trough, based upon gametogenesis, shell size, and gene variability. Mar Ecol Prog Ser 505:109–130
Nye V, Copley J, Linse K, Plouviez S (2013) Iheyaspira bathycodon new species (Vetigastropoda: Trochoidae: Turbinidar: Skeneinae) from the Von Damm vent field, Mid-Cayman spreading center, Caribbean. J Mar Biol Assoc UK 93:1017–1024
Ohta S (1991) Deep-sea submersible survey of the hydrothermal vent community on the northern slope of the Iheya Ridge, the Okinawa Trough. JAMSTEC Tech Rep Deep Sea Res 145–156 (In Japanese with English abstract and captions)
Podowski EL, Ma S, Luther GW III, Wardrop D, Fisher CR (2010) Biotic and abiotic factors affecting distributions of megafauna in diffuse flow on andesite and basalt along the Eastern Lau Spreading Center, Tonga. Mar Ecol Prog Ser 418:25–45
Rex MA, Etter RJ (2010) Deep-sea biodiversity: pattern and scale. Harvard University Press, Cambridge, p 354
Roterman CN, Copley JT, Linse KT, Tyler PA, Rogers AD (2013) The biogeography of the yeti crabs (Kiwaidae) with notes on the phylogeny of the Chirostyloidea (Decapoda: Anomura). Proc Roy Soc B. doi:10.1098/rspb.2013.0718
Sakai H, Gamo T, Ki ES, Tsutsumi M, Tanaka T, Ishibashi J, Wakita H, Yamano M, Oomori T (1990) Venting of carbon dioxide-rich fluid and hydrate formation in mid-Okinawa Trough backarc basin. Science 248:1093–1096
Sato K, Watanabe H, Sasaki T (2013) A new species of Solemya (Bivalvia: Protobranchia: Solemyidae) from a hydrothermal vent in the Iheya Ridge in the mid-Okinawa Trough, Japan. Nautilus 127:93–100
Seike K, Genkins RG, Watanabe H, Nomaki H, Sato K (2012) Novel use of burrow casting as a research tool in deep-sea ecology. Biol Lett 8:648–651
Takemura A, Tamotsu S, Miwa T, Yamamoto H (2010) Preliminary results on the reproduction of a deep-sea snailfish Careproctus rhodomelas around the active hydrothermal vent on the Hatoma Knoll, Okinawa, Japan. J Fish Biol 77:1709–1715
Tokeshi M (2011) Spatial structures of hydrothermal vents and vent-associated megafauna in the back-arc basin system of the Okinawa Trough, western Pacific. J Oceanogr 67:651–665
Tsuchida S, Watanabe K, Ishibashi J, Miyake H, Watabe H, Yamaguchi T, Kitajima T, Nakano A, Matsumura M, Watanabe H (2000) Preliminary report of a biological, geological, and geochemical survey on hydrothermalism at the Hatoma and Minna Knolls. JAMSTEC J Deep Sea Res 17:35–42
Tunnicliffe V, Juniper K, Sibuet M (2003) Reducing environments of the deep-sea floor. In: Tyler PA (ed) Ecosystems of the deep oceans, vol 28, Ecosystems of the world. Elsevier, Amsterdam, pp 81–110
Watanabe H, Tsuchida S, Fujikura K, Yamamoto H, Inagaki F, Kyo M, Kojima S (2005) Population history associated with hydrothermal vent activity inferred from genetic structure of neoverrucid barnacles around Japan. Mar Ecol Prog Ser 288:233–240
Watanabe H, Kado R, Kaida M, Tsuchida S, Kojima S (2006) Dispersal of vent-barnacle (genus Neoverruca) in the Western Pacific. Cah Biol Mar 47:353–357
Watanabe H, Fujikura K, Kojima S, Miyazaki J-I, Fujiwara Y (2010) Japan: vents and seeps in close proximity. In: Kiel S (ed) The vent and seep biota. Topics in geobiology, vol 33, pp 379–401
Watanabe H, Seo E, Takahashi Y, Yoshida T, Kojima S, Fujikura K, Miyake H (2013) Spatial distribution of sister species of vesicomyid bivalves Calyptogena okutanii and Calyptogena soyoae along an environmental gradient in chemosynthetic biological communities in Japan. J Oceanogr 69:129–134
Watsuji T, Nakagawa S, Tsuchida S, Toki T, Hirota A, Tsunogai U, Takai K (2010) Diversity and function of epibiotic microbial communities on the galatheid crab, Shinkaia crosnieri. Microbes Environ 25:288–294
Wessel P, Smith WH (1998) New, improved version of the generic mapping tools released. EOS Trans Am Geophys Union 79:579
Yorisue T, Kado R, Watanabe H, Hoeg JT, Inoue K, Kojima S, Chan BKK (2013) Influence of water temperature on the larval development of Neoverruca sp. and Ashinkailepas seepiophila—implications for larval dispersal and settlement in the vent and seep environments. Deep Sea Res I 71:33–37
Acknowledgements
We are grateful to the on-board scientist of the NT11-20 cruise for providing an overview of the faunal assemblages in the Okinawa Trough vent fields, Mr. Naoto Jimi and Ms. Mai Karasawa, for their great help with our polychaete identification, Dr. Hidenori Kumagai kindly provided us the map of Okinawa Trough, using free mapping software, GMT (Wessel and Smith 1998) based on the global data set of topography model, ETOPO1 (Amante and Eakins 2009). This study was supported by the “TAIGA project,” which was funded by a Grant-in-Aid for Scientific Research on Innovative Areas (No. 20109004) from the Ministry of Education, Culture, Sports, Science and Technology (MEXT), Japan, and the Canon Foundation.
Author information
Authors and Affiliations
Corresponding author
Editor information
Editors and Affiliations
Rights and permissions
Open Access This chapter is distributed under the terms of the Creative Commons Attribution Noncommercial License, which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Copyright information
© 2015 The Author(s)
About this chapter
Cite this chapter
Watanabe, H., Kojima, S. (2015). Vent Fauna in the Okinawa Trough. In: Ishibashi, Ji., Okino, K., Sunamura, M. (eds) Subseafloor Biosphere Linked to Hydrothermal Systems. Springer, Tokyo. https://doi.org/10.1007/978-4-431-54865-2_34
Download citation
DOI: https://doi.org/10.1007/978-4-431-54865-2_34
Published:
Publisher Name: Springer, Tokyo
Print ISBN: 978-4-431-54864-5
Online ISBN: 978-4-431-54865-2
eBook Packages: Earth and Environmental ScienceEarth and Environmental Science (R0)