
Abstract
Understanding the targets and mechanisms of human immunity to malaria caused by Plasmodium falciparum is crucial for advancing effective vaccines and developing tools for measuring immunity and exposure in populations. Acquired immunity to malaria predominantly targets the blood stage of infection when merozoites of Plasmodium spp. infect erythrocytes and replicate within them. During the intra-erythrocytic development of P. falciparum, numerous parasite-derived antigens are expressed on the surface of infected erythrocytes (IEs). These antigens enable P. falciparum-IEs to adhere in the vasculature and accumulate in multiple organs, which is a key process in the pathogenesis of disease. IE surface antigens, often referred to as variant surface antigens, are important targets of acquired protective immunity and include PfEMP1, RIFIN, STEVOR and SURFIN. These antigens are highly polymorphic and encoded by multigene families, which generate substantial antigenic diversity to mediate immune evasion. The most important immune target appears to be PfEMP1, which is a major ligand for vascular adhesion and sequestration of IEs. Studies are beginning to identify specific variants of PfEMP1 linked to disease pathogenesis that may be suitable for vaccine development, but overcoming antigenic diversity in PfEMP1 remains a major challenge. Much less is known about other surface antigens, or antigens on the surface of gametocyte-IEs, the effector mechanisms that mediate immunity, and how immunity is acquired and maintained over time; these are important topics for future research.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
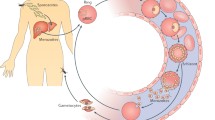
Plasmodium falciparum is the most virulent form of human malaria and is a leading cause of mortality among children under 5 years [1]. Plasmodium falciparum has a complex lifecycle involving a mosquito vector and a human host. The on-going asexual reproduction during the blood stage leads to clinical symptoms of malaria [2]. The pathogenesis of human malaria stems from various host and parasite factors that concurrently influence the severity and outcome of disease. Key pathophysiological features include the sequestration of P. falciparum-infected erythrocytes (IEs) in the microvasculature, the induction of proinflammatory cytokines and anemia resulting from the suppression of erythropoiesis [2, 3]. The destruction of uninfected erythrocytes and IEs further compromises oxygen delivery and exacerbates disease pathogenesis [4]. An important virulence property of P. falciparum is the expression of parasite-derived antigens on the surface of IEs, generally known as variant surface antigens (VSAs; Fig. 1), and its strong propensity to adhere in the vasculature. VSAs are comprised of novel parasite-derived proteins and include P. falciparum erythrocyte membrane protein 1 (PfEMP1) [5], repetitive interspersed family (RIFIN) proteins [6–8], sub-telomeric variable open reading frame (STEVOR) proteins [9–11], surface-associated interspersed gene family (SURFIN) proteins [12] and possibly others such as P. falciparum Maurer’s cleft two transmembrane (PfMC-2TM) proteins [13, 14]. Parasite-modified erythrocyte band 3 has also been proposed as a surface antigen or ligand for IE sequestration [15, 16]. These IE surface proteins are antigenically diverse and undergo clonal antigenic variation because of the selective pressure exerted by human immunity. The significance of VSAs as targets of naturally acquired immunity and their potential as vaccine candidates is the focus of this review. Acquired immunity to blood stage P. falciparum will be addressed, followed by a summary of the VSAs expressed on the IE surface and finally human antibodies to different VSA families.

Parasite-induced modifications to P. falciparum-infected erythrocytes. A. During intra-erythrocytic development, P. falciparum expresses knob structures and VSAs on the surface of pigmented trophozoite IEs. PfEMP1, P. falciparum erythrocyte membrane protein 1; RIFIN, repetitive interspersed family; STEVOR, subtelomeric variable open reading frame; SURFIN, surface-associated interspersed gene family; KAHRP, knob-associated histidine-rich protein. B. Scanning (left) and transmission (right) electron microscopy (EM) shows the ultrastructural features of the IE membrane. The IE membrane is distorted by surface knob protrusions (arrows) that present the major virulence factor, PfEMP1
Plasmodium falciparum sequestration and cytoadhesion
The virulence of P. falciparum malaria is attributed to the adhesion of IEs to the vascular endothelium or to uninfected erythrocytes to form rosettes [17–19]. Mature P. falciparum disappear from the peripheral circulation and are sequestered in various organs throughout the body. The importance of splenic clearance of IEs in controlling disease severity has been demonstrated by numerous studies. For example, a study conducted with Aotus monkeys showed that splenectomised animals developed virulent infections, presumably because of enhanced accumulation of IEs in the microvasculature [20, 21]. IE sequestration contributes to the pathogenesis of severe disease syndromes such as cerebral [17, 22, 23] and placental complications [24, 25]. An important feature of IEs that enables P. falciparum to sequester is the expression of knob structures on the IE membrane [26–28]. The major structural component of knobs is the knob-associated histidine-rich protein (KAHRP) [27, 29–32]. Other parasite-encoded proteins such as P. falciparum erythrocyte membrane protein 3 (PfEMP3) [33] and mature IE surface antigen (MESA; also known as PfEMP2) [34, 35] also contribute to knob assembly. KAHRP interacts with cytoskeletal components of the erythrocyte such as spectrin and actin [36–38], resulting in reduced membrane deformability [39]. Knobs present the major virulence factor, PfEMP1 [5], on the external surface of the IE membrane, where it mediates IE cytoadhesion to the host endothelium under physiological flow conditions [40, 41]. Disruption of the kahrp gene impairs proper knob formation and leads to a decrease in surface-exposed PfEMP1 and reduced cytoadhesion [42]. However, the presence of knobs may not necessarily result in sequestration [43]; P. malariae has knob structures but does not sequester, while P. chabaudi sequesters without knobs [16, 43].
A diverse range of host receptors that mediate IE cytoadhesion has been identified [44–46]. The main parasite ligand responsible for cytoadhesion is PfEMP1 and it binds to a range of endothelial and erythrocyte molecules including CD36 [47], ICAM-1 [48], chondroitin sulphate A (CSA) [49, 50], complement receptor 1 (CR1) [51], heparan sulfate (HS) [52] and others. IEs are capable of binding via multiple receptors [53] therefore creating a synergistic effect on IE adhesion [54]. Most P. falciparum isolates adhere to both ICAM-1 and CD36, which are widely distributed in the vasculature [53, 55, 56], but parasites isolated from infected placentas mainly adhere to specific receptors expressed by the syncytiotrophoblasts of the infected placenta [57, 58], particularly CSA [56, 59, 60], and possibly secondary receptors such as hyaluronic acid (HA) [61–63] and non-immune IgM [64–66] and IgG [67]. The differential expression of endothelial cell receptors in various tissues leads to the preferential binding of IEs. For example, it is proposed that ICAM-1-binding parasites are more likely to sequester in the brain [46, 68] as the brain endothelium expresses ICAM-1. While it has been speculated that receptor-specific adhesion (e.g. ICAM-1) predisposes to a particular pattern of disease (e.g. cerebral malaria), studies to date have been inconclusive. In one study, cerebral malaria patients did not show a significant association between disease and ICAM-1 binding [69], and another study reported that ICAM-1-binding was lowest in children with severe malaria [70]. In contrast, post-mortem histopathological analyses of infected cerebral vessels proposed a role for ICAM-1 in the manifestation of severe disease [68], and another study demonstrated that ICAM-1-binding was greater in cerebral malaria patients compared to patients with uncomplicated malaria [71]. This may propose a role for ICAM-1 in the pathogenesis of cerebral malaria but additional studies are necessary to further validate this association. A recent study identified endothelial protein receptor C as a likely mediator of cerebral sequestration [72]. Other host receptors implicated in IE cytoadherence include thrombospondin (TSP) [73], platelet/endothelial cell adhesion molecule (PECAM/CD31) [74], P-selectin [75] and vascular cell adhesion molecule-1 (VCAM-1) [53], but the significance of these receptors in disease pathogenesis remains unclear.
The clustering of mature IEs to uninfected erythrocytes, known as rosetting [76], is also thought to contribute to excessive microvasculature obstruction [77, 78]. Rosetting is associated with severe malaria in African children, suggesting that it contributes to disease pathogenesis [79–83]. However, a study with Malawian [70] and PNG [84] children reported that the rosetting occurred at a similar rate between children with severe and uncomplicated malaria, suggesting that rosette formation is not always associated with severe clinical outcomes. The parasite ligand for rosetting in P. falciparum has been identified as a specific PfEMP1 variant that binds to CR1 [51] or HS proteoglycans [85] expressed by the host erythrocyte.
Plasmodium falciparum also causes vascular obstruction through the clumping of IEs, a feature that was first reported as autoagglutination [86]. This adhesive phenotype is distinct from rosetting, as autoagglutinating parasites do not form rosettes and rosetting parasites do not autoagglutinate [86]. Autoagglutination was a common feature of infection, although more autoagglutinates were observed in children with severe malaria compared to those with mild malaria. This suggests that autoagglutination is more frequently observed in, but not restricted to, severe disease [87]. It was later reported that autoagglutination is mediated by platelets and required the expression of the platelet glycoprotein CD36 [88]. Scanning electron microscopy of platelet-mediated clumping of IEs showed that this interaction occurred at the IE knob structures [88]. Studies with Kenyan children [88] and patients from Thailand [89] established that platelet-mediated clumping of IEs was associated with severe disease, presumably through local disruptions of blood flow. Conversely, binding of platelets to IEs has also been implicated in protection against P. falciparum by directly inhibiting intra-erythrocytic parasite growth [90, 91].
VSAs of P. falciparum
The most extensively studied VSA is the major virulence factor PfEMP1, an important target of naturally acquired immunity [92, 93]. The var genes that encode PfEMP1 appear to be unique to P. falciparum, but the P. knowlesi schizont-infected cell agglutination (SICA) antigens encoded by the SICAvar multigene family [94, 95] have been described as conceptually similar to PfEMP1 [96]. Orthologs of rif and stevor genes have been identified in other Plasmodium species, known collectively as the pir multigene family (Plasmodium interspersed repeats). These include the vir multigene family in P. vivax [97], kir multigene family in P. knowlesi, yir multigene family in P. yoelii, bir multigene family in P. berghei and cir multigene family in P. chabaudi [98, 99].
Currently, little is known regarding the mechanisms that regulate gene transcription in P. falciparum other than the involvement of specific transcription factors and promoter interactions. Exploiting the mutually exclusive transcription of var genes has allowed for the suppression of the entire endogenous var multigene family [100, 101], thus enabling the specific study of PfEMP1 [102]. Conversely, disrupting up to 150 genes such as in the rif family is not currently feasible, and knockdowns of rif, stevor and pfmc2tm gene families have not been achieved. These multigene families share a common activation factor necessary for gene expression, and it has been proposed that the downregulation of one multigene family may affect the expression of members of other multigene families in some conditions [103]. A transcriptionally active rif promoter co-localised with an active var promoter and the downregulation of members in the stevor multigene family appeared to increase transcription of the pfmc-2tm multigene family [103]. Further studies must be employed to dissect the functional roles of these multigene families.
It is hypothesised that a large proportion of parasite proteins exported into the host erythrocyte supports the correct trafficking and surface display of PfEMP1 and other erythrocyte surface proteins. This also includes alterations in the spectrin network and knob protrusions at IE membrane [104]. Many exported proteins contain a pentameric sequence, known as the Plasmodium export element (PEXEL) [105] or vacuolar translocation signal [106], required for the translocation of proteins across the PV membrane. Exported parasite proteins such as PfEMP1 are trafficked via the translocon complex (PTEX; Plasmodium translocon of exported proteins) located at the parasitophorous vacuole (PV) [107, reviewed in 108]. Most parasite proteins are destined for the IE cytosol, and only a small portion is exposed on the IE surface. Interestingly, recent studies have showed that exported parasite proteins may play a role in cellular communication between IEs through microvesicles [109] and exosome-like vesicles [110]. Microvesicles lack components of the knob structure like KAHRP and PfEMP1, suggesting that they originate from MC structures or regions of the erythrocyte membrane that exclude knobs [109]. It was further demonstrated that PfEMP1 is not required for efficient intercellular communication as modified parasites with inhibited PfEMP1 expression were still able to receive exosome-like vesicles [110].
PfEMP1 and var genes
PfEMP1 was first identified by immunoprecipitation with immune sera from infected Aotus monkeys [5] and is encoded by the highly polymorphic var multigene family (~60 genes per genome) [40, 111, 112]. Through mutually exclusive transcription of var genes, only a single PfEMP1 variant is generally expressed on the IE surface at a given time [52, 113]. However, a recent study reported the potential expression of more than one PfEMP1 variant on the IE surface as demonstrated by live confocal microscopy, in vitro adhesion assays and cell sorting by flow cytometry [114]. PfEMP1 is a high-molecular-weight protein (200–350 kDa) and is highly sensitive to cleavage by mild trypsin treatment of intact IEs (10 μg/ml) [5]. The biochemical properties of PfEMP1 (Triton X-100-insoluble and SDS-soluble) demonstrate its anchorage to the IE membrane [115].
The export of PfEMP1 is a highly complex process due to its large size, number of membranes to traverse before reaching the IE surface and the involvement of various chaperone proteins (reviewed in [116, 117]). PfEMP1 molecules are associated with the Maurer’s clefts (MCs) and are ultimately presented by the knob structures at the IE surface at approximately 18 h post-invasion [5, 32, 115, 118–120]. The mechanism of transport of PfEMP1 from the parasite to the IE surface as well as its partner proteins involved remains a process that is poorly defined.
An important MC-resident protein essential for PfEMP1 trafficking is the 48 kDa P. falciparum skeleton binding protein 1 (SBP1) [121–123]. Disruption of the pfsbp1 gene impaired the loading of PfEMP1 molecules into the MCs and resulted in the loss of surface-exposed PfEMP1, but the trafficking of other MC proteins such as KAHRP, MAHRP1 and REX1 was unaffected in the transgenic parasites [122, 123]. A large-scale gene knockout screen further identified MC proteins termed P. falciparum PfEMP1 trafficking protein (PfPTP) involved in the trafficking of PfEMP1 to the IE surface [124]. Of the 83 P. falciparum genes that were disrupted, 6 genes were specifically found to affect the export and surface display of PfEMP1, and 2 were found to disrupt proper knob formation. In these transgenic parasites, PfEMP1 export was arrested either at the PVM or the MC structures in the IE cytosol [124]. Among the proteins involved in the trafficking of PfEMP1 are members of the HSP40/DNAJ and PHIST family [124]. Recently, an exported parasite-encoded HSP70 known as PfHSP70-x was found to complex with HSP40s and colocalised with PfEMP1 in the IE cytosol [125]. Other MC-associated proteins that have been proposed to play a role in the trafficking of PfEMP1 include MAHRP [126], REX1 [127] and Pf332 [128].
Transcription of var genes is epigenetically regulated by the SIR complex as gene disruption of PfSIR2 results in activation of multiple members of this multigene family [129–131]. The rapid switching rate of var genes, of up to 2 % per generation [86], was demonstrated to correlate with changes in IE adhesive and antigenic phenotypes [112, 132]. The typical structure of PfEMP1 includes the variable N-terminal segment (NTS) exposed on the IE surface to interact with host receptors [133], a transmembrane domain and a conserved acidic terminal segment (ATS) [48]. The cytoplasmic ATS domain interacts with KAHRP [134–136], thus anchoring PfEMP1 to the IE membrane. The extracellular portion of PfEMP1 consists of Duffy binding-like (DBL) adhesive domains, the C2 domain and the cysteine-rich interdomain regions (CIDR) [48]. DBL domains are grouped as five sequence classes (α, β, γ, δ, ε), and CIDR domains are grouped as three distinct classes (α, β, γ). While the number of CIDR and DBL domains may vary between different PfEMP1 variants, certain domain architectures such as DBLαCIDRα or DBLδCIDRβ are preferred. This conservation may reflect the biological function of PfEMP1 [48]. The binding site of ICAM-1 resides within the DBL2β and c2 regions of PfEMP1 [137, 138], while the binding site of CD36 is mapped to the CIDRα region [47, 139, 140]. The binding site of CSA in pregnancy-associated parasites lies within the DBL1-DBL3 domain of PfEMP1 [141].
The var genes can be classified into three main subgroups based on their upstream promoter regions (upsA, upsB and upsC) and the single-copy conserved intergenomic genes var1 and var2csa [142–145]. The var gene repertoires of clinical isolates can also be classified according to short sequence tags amplified from the DBLα domain [146, 147]. Analyses of the DBLα sequences from Kenya showed the presence of two or four cysteine residues, with a minority containing one, three, five or six cysteines; therefore, var genes were sub-grouped according to cys2, cys4 and cysX, respectively [147]. These DBLα sequences were further classified according to the amino acid motifs occurring at four fixed positions within the sequenced region, known as “positions of limited variations” (PoLV1-4) [147]. Expression of var genes from the cys2 group was associated with severe malaria in young children [148, 149]. The differential transcription of var gene subgroups has been linked to clinical disease. Transcription of group A var genes was also associated with rosetting parasites [150] and severe malaria in African children [151–153]. Furthermore, the elevated expression of group A-like var genes was associated with impaired consciousness, a key feature of severe disease [154]. Recent studies also reported the expression of a restricted subset of var genes encoding PfEMP1 variants that bind human brain endothelial cells [155, 156]. These var genes belong to group B/A genes that are expressed in early childhood infections [148, 157] and are associated with more severe infections [149, 151]. PfEMP1 variants from group B and C var genes are also associated with autoagglutination and ICAM-1-binding, features that contribute to severe disease [133, 138]. The lack of association between transcription of var group C and clinical presentation suggests that perhaps these var genes are involved in the establishment of chronic infections [158]. These findings support the correlation between var gene expression patterns and clinical presentations, thus suggesting that protective immunity could be conferred by antibodies to key var gene subgroups [147, 158].
A specific var gene, var2csa, is relatively conserved in sequence and is present as a single-copy gene in most isolates. However, some isolates have more than one copy of var2csa (e.g. HB3 has two copies of var2csa) [159–161]. This gene is upregulated in placental isolates and mediates IE adhesion to CSA and other placental receptors such as HA and immunoglobulins [162–165]. Polyclonal antibodies generated against recombinant domains of VAR2CSA recognised the IE surface of parasites isolated from infected placental tissue [166]. Furthermore, sera from pregnant women recognised the IE surface of VAR2CSA-expressing parasites in a parity-dependent manner [164, 167]. Pregnant women with elevated levels of these antibodies had a reduced risk of delivering low-birth-weight babies [164]. Targeted gene disruption of var2csa in the isolates FCR3 [168] and 3D7 [169] inhibited CSA adhesion, suggesting the central role of var2csa in mediating placental adhesion. In contrast, disruption of var2csa in CS2 parasites also ablated CSA binding but repeated selection on CSA restored their binding ability [169] presumably through the expression of other PfEMP1 variants proposed to bind CSA [49]. The use of different parasite lines may reflect the discrepancies between these studies and that the FCR3 or 3D7 isolates lack the PfEMP1 variants thought to rescue the CSA-binding ability.
The var transcripts have also been detected by RT-PCR in both immature (I–II) and mature stage gametocyte-IEs (IIB–V). Initial studies suggested that the var genes transcribed in gametocyte-IEs were similar or identical to those expressed by asexual parasites [170]. However, it was later discovered that the var transcript profile was unlinked to their asexual progenitors. Furthermore, it appears that the most abundant var transcripts found in gametocyte-IEs (generated in vitro) belong to the non-subtelomeric group C var genes [171]. Data regarding the pattern of PfEMP1 expression in gametocyte-IEs are conflicting. Early studies reported the expression of PfEMP1 in all five stages of gametocyte development but with stage-specific patterns. PfEMP1 staining was visualised at the IE membrane of immature gametocyte-IEs (stages I–IIA) but not of mature gametocyte-IEs (IIB–V) [170]. A recent study reported low levels of PfEMP1 expression on the surface of immature gametocyte-IEs, which was absent in mature gametocyte-IEs [172]. Thus, it is thought that sequestration of immature gametocyte-IEs is mediated by PfEMP1, after which its role is replaced by an alternative ligand present on the surface of mature gametocyte-IEs or through mechanical effects [173].
RIFIN proteins
The rif multigene family (150–200 genes per genome) encoding a group of clonally variant RIFIN proteins represents the largest multigene family identified in P. falciparum [6–8]. Transcription of rif genes occurs approximately 12 h post-invasion, but RIFIN proteins are thought to appear on the IE surface at the same time as PfEMP1 [174]. In contrast to var genes, a single parasite simultaneously transcribes several rif genes, resulting in the expression of multiple RIFIN variants on the IE surface [8]. All RIFIN sequences contain the PEXEL motif required for correct export [105]. Surface exposure of RIFIN was evident from immunoprecipitation and Western blot analyses. Bands of expected size corresponding to RIFIN (30–45 kDa) were absent after IEs were treated with high concentrations of trypsin (>100 μg/ml), a concentration much greater than that needed to cleave the highly trypsin-sensitive PfEMP1 [7, 8]. Some variants of the large RIFIN family are also expressed in other developmental stages such as merozoites, sporozoites and gametocytes [175–177]. Bioinformatic analyses of RIFIN sequences revealed two major subgroups of the RIFIN family [178] that are simultaneously expressed in a single parasite. A-type RIFINs associate with the MCs and are destined for the IE surface, whereas B-type RIFINs remain confined within the parasite [176]. Although the biological function of RIFIN remains unknown, the exposure of their highly polymorphic V2 epitope on the IE surface suggests they contribute to antigenic variation of P. falciparum [6, 179]. Although direct evidence is lacking, RIFIN was proposed to play a role in rosetting [7, 8].
STEVOR proteins
The stevor multigene family, encoding STEVOR proteins (~30–40 kDa), is the third largest identified in P. falciparum (reviewed in [10]). First described as 7h8, stevor was detected by a monoclonal antibody as an expressed sequence [6]. Each parasite genome is predicted to contain approximately 30–40 copies of stevor genes. Like var and rif, stevor genes are located at the telomeres of most P. falciparum chromosomes [6]. Similar to rif, multiple stevor transcripts were detected in a single parasite. Peak stevor transcription occurs at 28 h post-invasion during late trophozoites and early schizonts, where they appear to localise in the IE cytosol. As the parasite matures, STEVOR co-localises with PfSBP1 and PfEMP3 at the MCs [9, 180] in immunofluorescence microscopy with fixed IEs [10, 11]. Immunofluorescence microscopy with live, intact schizont stage IEs suggests the surface localisation of STEVOR, which was removed upon IE trypsinisation [11, 181]. A recent study demonstrated that stevor overexpression contributes to increased IE rigidity [182], together with other IE cytoskeletal members such as RESA [183]. It is proposed that the STEVOR-increased stiffness of IEs enhances PfEMP1-mediated IE sequestration [182]. The biological function of STEVOR remains unclear. Because STEVOR is clonally variant [13, 184], it may be involved in immune evasion concurrently with PfEMP1 and RIFIN [10, 11, 13]. In addition, STEVOR has been proposed to play a role in parasite invasion [185]. STEVOR proteins are also expressed in merozoites [10, 186, 187], sporozoites [175] and gametocytes [9]. Interestingly, the same STEVOR variants are transcribed in gametocytes and their asexual progenitors, suggesting that perhaps STEVOR plays a similar role in these lifecycle stages [171].
SURFIN proteins
Little is known about the surf multigene family (10 genes), which encodes high-molecular-weight antigens (~280–300 kDa) known as SURFIN proteins [12]. The expression of surf genes is differentially transcribed according to different stages of the intra-erythrocytic parasite. The expression of surf 1.3, surf 4.2 and surf 8.3 genes was detected throughout parasite development while other surf genes were either not detected or restricted to later developmental stages [188]. A variant expressed by 3D7 and FCR3 parasites, SURFIN4.2 was identified by mass spectrometric analysis of proteins cleaved off the surface of intact IEs by trypsin [12]. SURFIN4.2 was only detected in a subpopulation of cultured IEs (~25 %) with increasing protein expression during mature developmental stages (24–44 h post-invasion). Immunoelectron microscopy showed the presence of SURFIN4.2 at the knob structures suggesting its co-localisation with PfEMP1 at the IE surface [12]. However, attempts to verify the surface localisation of SURFIN4.2 proteins with live, intact IEs were inconclusive. Another variant, SURFIN4.1, localised to the parasitophorous vacuole (PV) but not within the erythrocyte cytosol in mature IEs (>30 h post-invasion) [188]. SURFIN antibodies did not agglutinate mature IEs and no fluorescence was observed with live IEs, suggesting that SURFIN4.1 is not exposed on the IE surface [188]. Whether SURFIN proteins potentially elicit humoral immunity or mediate immune evasion has not been determined.
Other membrane proteins
Plasmodium falciparum Maurer’s clefts two-transmembrane protein (PfMC-2TM) is encoded by a novel gene family (~13 members) located at the subtelomeric regions of several P. falciparum chromosomes [14, 142]. PfMC-2TM is highly conserved within the N-terminus, both transmembrane domains and the C-terminus. The short loop between the transmembrane domains is highly polymorphic, similar to that proposed for RIFIN and STEVOR [13, 184]. The diversity within this loop region proposed the inclusion of pfmc-2tm as a variant multigene family together with var, rif and stevor [13, 14]. It has not been determined whether PfMC-2TM is associated with the IE membrane or exposed on the IE surface.
Another IE membrane protein is modified erythrocyte band 3, which has been proposed as a ligand for IE adhesion to CD36 [189] and thrombospondin [190, 191]. Chemical modifications of band 3 led to a reduction in CD36 binding but not thrombospondin, thus supporting its role in CD36 adhesion [189]. Synthetic peptides based on the exofacial loops of band 3 and antibodies generated against these peptides were capable of inhibiting IE adhesion to C32 amelanotic melanoma cells. Additionally, intravenous infusion of these peptides into Aotus and Saimiri monkeys infected with P. falciparum isolates prevented IE sequestration [192]. However, its significance in relation to PfEMP1 as an adhesive ligand remains unclear. In our recent study, parasites with suppressed PfEMP1 expression were found to retain a substantial proportion of CD36 binding, but not ICAM-1, thus raising the possibility that additional surface antigens contribute to CD36 adhesion [102].
Naturally acquired immunity to malaria
Protective immunity to malaria is elicited through complex interactions between both humoral and cell-mediated responses [193–195]. This protection against symptomatic malaria in humans develops gradually after repeated exposure to P. falciparum infections (reviewed in [196]). In malaria-endemic areas, the risk of severe disease is greatest during the first few years of life, after which the risk rapidly declines as children begin to acquire natural immunity. A study of young African children reported that immunity to non-cerebral severe malaria may develop after several infections and is almost complete by the age of 5 [197]. Adolescents and adults eventually develop protection from severe illness and death, although sterile immunity is rarely or perhaps never achieved [198]. Maternal antibodies transferred across the placenta are also thought to confer protection in young infants [199, 200]. It is becoming increasingly clear that effective immunity to malaria involves immune responses to multiple antigens expressed at different parasite stages and requires multiple immune effector mechanisms [195]. It is likely that the development of a highly effective malaria vaccine will require the inclusion of multiple antigens and that single-antigen vaccines will not be optimally efficacious.
Cell-mediated immunity to VSAs
While the significance of humoral immunity to P. falciparum is well established in humans, the role of cell-mediated immunity (CMI) remains poorly understood. CMI acts through complex interactions with the innate and adaptive immune response (reviewed in [195, 201]). Most studies of CMI have been based on murine models of malaria (reviewed in [202]). Early studies showed that mice incapable of making B cells have the ability to control infection [203], suggesting the importance of CMI in protection against malaria. Antigen-presenting cells process parasite antigens for display on major histocompatibility complex molecules to recruit antigen-specific CD4+ T cells. Th1 cells produce proinflammatory cytokines such as TNF-α and IFN-γ, which lead to monocyte activation and the release of toxic mediators that limit P. falciparum growth [204]. In a study of Gabonese children, IFN-γ responses to erythrocytic antigens were associated with lower rates of P. falciparum reinfection [205]. Despite this protective potential, CMI responses have also been implicated in disease pathogenesis and the development of severe malaria (reviewed in [206]).
Although data on CMI responses to VSAs are limited, studies have reported that parasites may modulate CMI to evade host immune responses. For example, the maturation of dendritic cells cultured in vitro was suppressed following exposure to erythrocytes infected with P. falciparum [207]. It was later reported that the modulation of dendritic cells was not dependent on the interaction between PfEMP1 and CD36 [208]. Interaction between IEs and natural killer (NK) cells leads to their activation, including production of IFN-γ [209]. A recent study using a humanised mouse model has reported that NK cell binding of IEs leads to the activation of NK cells and the elimination of IEs [210]. Using parasites with modified PfEMP1 expression, others reported that PfEMP1 appeared to suppress innate IFN-γ production by naïve CD4+ T cells and NK cells [211]. The CIDR1α domain of PfEMP1 was found to induce polyclonal B cell activation that contributes to the evasion of host immune responses [212–214] and stimulates CD4+ T cells from both malaria-exposed and non-exposed individuals [215]. The addition of recombinant CIDR1α to naïve human peripheral blood mononuclear cells resulted in the activation of CD4+ T cells and NK cells, leading to IFN-γ production [216]. It appears that a fine balance between protective immunity and immunopathology must be achieved in CMI. The lack of CMI-related studies in human malaria and the difficulty of inferring results from murine models are continuing obstacles in our understanding of the role of CMI and a priority topic for further research.
Human antibodies to VSAs
The development of protective immunity to P. falciparum is characterised by a decrease in disease severity over several years after repeated infections [217]. Sterile immunity to P. falciparum is rarely achieved, as adults living in malaria-endemic regions remain susceptible to asymptomatic infection and often experience persistent low levels of parasitaemia without clinical disease [196, 198]. The passive transfer of gamma-globulin from immune individuals to P. falciparum-infected individuals confers protection against malaria infection [218]. Antibodies to both merozoite antigens [219–221] and VSAs appear to play an important role in mediating acquired immunity. The focus of this review is on VSAs, and the significance of merozoite antigens as immune targets is reviewed in detail elsewhere [222]. In brief, numerous antigens on the surface of merozoites (e.g. merozoite surface protein 1, 2, and 3) and erythrocyte invasion ligands (e.g. erythrocyte-binding antigens, PfRh invasion ligands and apical membrane antigen 1) have been identified as important targets of acquired immunity and promising vaccine candidates [219–221, 223, 224]. Although it is highly likely that antibodies to VSA and merozoite antigens contribute to immunity and a strong response to both types of antigens may be essential for highly effective immunity, there are few reports on the relationship between these responses and how they may interact to mediate immunity [225]; this is an important question for further research.
Naturally acquired antibodies against VSAs typically demonstrate a high degree of strain specificity [132, 226]. Antigenic diversity by P. falciparum enables repeated infections to occur over time, as new infections appear to exploit gaps in the repertoire of previously acquired variant-specific antibodies [226]. Antibodies to polymorphic VSAs expressed on the IE surface, such as PfEMP1, have been proposed to play a key role in mediating protective immunity [93, 226–228]. Most published studies used agglutination assays to describe antigenic variation in PfEMP1 as switches in the agglutination phenotype are correlated with switches in var gene expression [112] or PfEMP1 [132]. However, it is difficult to determine PfEMP1-specific antibody responses because of the number of antigens expressed on the IE surface; therefore, antibodies measured to the IE surface are hereafter classified as antibodies to all VSAs (studies summarised in Table S1).
Early studies that measured agglutination antibody responses to P. falciparum infections in Pakistan [229], Papua New Guinea [230] and Africa [226, 231] reported that children developed isolate-specific antibodies to VSAs after infection. Mixed agglutination assays that allowed the determination of shared epitopes on the IE surface also showed that acquired human antibodies are predominantly variant specific while cross-reactive antibodies are rare [231]. However, antibodies from convalescent sera from adults were capable of agglutinating diverse P. falciparum isolates [232], and antibodies acquired towards placental-binding parasites expressing VAR2CSA have a significant amount of cross-reactivity against different isolates, despite polymorphisms in VAR2CSA [159, 233]. Furthermore, an acute P. falciparum infection in returned travellers was sufficient to induce broadly cross-reactive antibodies that were relatively long lived (>20 weeks post-infection) [208], thus suggesting that cross-reactive antibodies are prevalent following infection. The molecular basis for this remains unclear but may be due to extensive sharing of polymorphic epitopes between PfEMP1 variants [159, 234]. In an early study, parasite isolates from ten Gambian children were tested in a checkerboard manner with the acute and convalescent sera collected from each child [226]. Most of the acute sera were not reactive towards the parasite isolate from that same child, whereas each of the serum samples collected during convalescence were highly reactive to the IE surface of the isolate from the same child but not from other children [226]. This study suggested that the VSAs expressed on the IE surface are highly diverse and children tend to acquire antibodies towards the variants expressed by the parasite causing that particular episode. Moreover, hyperimmune sera from Gambian adults agglutinated IEs from those ten children, suggesting that by adulthood most individuals have acquired a broad range of antibodies that protect against numerous parasite variants [226], and the presence of cross-reactive antibodies was also proposed. A large prospective study of young Kenyan children further showed that parasite variants expressed during episodes of clinical infection were less likely to be recognised by homologous sera collected before infection. This suggests that the variants are exploiting gaps in the pre-existing antibody repertoire in order to cause subsequent infections [227]. Consistent with that observation, antibodies from Sudanese children measured after the malaria season could agglutinate a broader range of isolates tested compared with antibodies measured before the season [235]. This suggests that natural P. falciparum infections are capable of inducing high antibody titres directed towards VSAs of the infecting parasite. In a cross-sectional study of Kenyan children during the low transmission season, antibodies to VSAs were higher in parasitaemic individuals, suggesting that these antibodies were induced by current infections [236]. Furthermore, plasma antibody levels were positively correlated with age, indicating an age-related and exposure-related component in the acquisition of VSA-specific antibodies [237].
The importance of antibodies to VSAs is evident through their role in mediating protective immunity to malaria (studies are summarised in Table 1). For example, in a longitudinal study conducted with young Gambian children, acquired antibodies to VSAs from several different isolates were consistently associated with protection from clinical malaria [228]. In Gabonese children, higher levels of IgG to VSAs were associated with lower rates of malaria [238]. A study of Ghanian children demonstrated that those with pre-existing antibodies before the malaria season were less likely to contract malaria than those with low levels of antibodies [239]. Furthermore, the presence of antibodies to a Ghanian isolate was significantly associated with protection from malaria [240]. However, the ability of sera from Kenyan children to recognise VSAs expressed by a Kenyan isolate was not associated with protection from malaria [227]. It appears that anti-VSA antibodies to some but not all parasite isolates are associated with protection [236, 239, 240], and presumably this depends on the prevalence of the parasite variant and its virulence properties. Nonetheless, evidence from all of the studies presented above supports the important contribution of anti-VSA antibodies to protection against malaria.
Despite the apparent role of PfEMP1 antibodies in mediating protection against malaria, the immense diversity of PfEMP1 limits its potential as a vaccine candidate. Although the repertoire of PfEMP1 variants is large, studies have suggested the expression of a dominant subset of variants that are restricted by their biological function in clinical disease. Immune sera from distinct geographic regions agglutinated IEs from other populations, suggesting that antibodies targeted cross-reactive epitopes expressed by many isolates [241]. Another study demonstrated that plasma antibodies were capable of recognising various parasite isolates regardless of the geographic origin of those IEs, suggesting that the repertoire of VSA-specific antibodies may be conserved over different populations [242]. A study in Kenya demonstrated that parasites isolated from children presenting with severe malaria were recognised by heterologous plasma antibodies, suggesting the expression of common PfEMP1 variants in this population [92]. Similarly, IEs from young Ghanian children with severe malaria were more commonly recognised by plasma antibodies from other children than those with uncomplicated malaria [243]. This suggests that a restricted subset of variants is expressed during severe disease to which antibodies are rapidly acquired [197]. These studies propose that the infecting parasites causing severe disease may be expressing a commonly expressed subset of VSA variants [236, 237]. A limitation of these studies is that they were only able to measure antibodies directed towards all VSAs expressed on the IE surface and not the proportion of antibodies to individual VSAs such as PfEMP1.
Little is known about antibody responses directed at antigens expressed on the surface of erythrocytes infected with gametocytes during their development in the human host. Such antigens could potentially elicit immune responses, similar to those of asexual parasites, which may result in the clearance of gametocytes in the host. Sera from Gambian children were reported to be highly reactive towards the surface of mature stage gametocyte-IEs but not towards immature stage gametocyte-IEs [244]. This may suggest that the antigens recognised by the serum antibodies in mature gametocyte-IEs are distinct from those expressed by immature gametocyte-IEs [244]. However, another study showed that sera from children in Papua New Guinea (PNG) were highly reactive to the surface of immature gametocyte-IE, similar to that observed with asexual trophozoite IEs, but not towards the surface of mature gametocyte-IEs [245]. These conflicting results demonstrate that further work is needed to better understand the antibody response directed towards antigens on the surface of gametocyte-IEs. Antibodies targeting these surface antigens represent potential vaccine candidates as they may mediate gametocyte clearance from the circulation, thus leading to reduced malaria transmission. More importantly, understanding the humoral response elicited by antigens on the surface of gametocyte-IEs will shed light on how these antibodies potentially act in synchrony with antibodies to other parasite stages to clear parasitaemia and reduce transmission.
Human antibodies to PfEMP1
Epidemiological data have demonstrated that naturally acquired antibodies predominantly target variant-specific epitopes on the IE surface and PfEMP1 is thought to be a major antibody target. Lacking the molecular tools required to evaluate the significance of PfEMP1 independently of other VSAs, most studies have relied on the use of recombinant purified PfEMP1 domains to study human antibody responses to PfEMP1 (studies summarised in Table S2). A recent study in PNG used a DBLα protein microarray to demonstrate that the magnitude of the anti-PfEMP1 response was limited and variant specific in young children (<3 years of age), after which a broader spectrum of antibody recognition was achieved. By adulthood, serum antibodies were capable of recognising at least 20 different variants indicating an expansion of the PfEMP1 antibody repertoire [246]. Consistent with the preferential expression of PfEMP1 variants in severe disease [151, 152], the acquisition of anti-PfEMP1 antibodies by Tanzanian children was reported to be highly structured. Antibodies to different recombinant PfEMP1 domains were sequentially acquired, with children first acquiring antibodies to particular variants encoded by group A var genes [157, 247]. These findings were supported by another study whereby PfEMP1 DBLα domains were linked to young host age, disease severity and low levels of immunity [148]. Furthermore, they complement the finding that immunity to severe disease may be rapidly acquired after several infections [197].
Others have reported that while sera from Gabonese adults recognised most of the recombinant PfEMP1 based on the DBLα region, sera from children were less reactive to the different variants [248]. Interestingly, they also showed that antibodies from highly reactive adults chosen from previous assays were capable of recognising synthetic peptides based on conserved regions of DBLα [248], suggesting that antibodies to both variant-specific and conserved regions of PfEMP1 are co-acquired. Similarly, serum antibodies from Ghanian [239] and Sudanese [249] children recognised a recombinant peptide derived from a conserved epitope of the DBLα domain, with higher antibody levels observed in asymptomatic individuals compared to those with febrile malaria suggesting that antibodies against conserved epitopes of PfEMP1 may play a role in protective immunity. However, there was no association observed between these antibodies and protection from malaria in Ghanian children, because the recombinant peptide originated from a domain that is inaccessible to antibodies [239]. Since conserved epitopes are not consistent with being key antibody targets, the role of these antibodies in protective immunity remains unclear. Few studies have evaluated the protective effect of anti-PfEMP1 antibodies (studies summarised in Table 2) and results have been inconsistent. A longitudinal study with Ghanian children did not find a correlation between protection and antibodies to the DBLα domain of PfEMP1 [239]. No protective association was observed with antibodies to the recombinant PfEMP1 domains derived from the A4 parasite line [250].
Quantifying the importance of different VSAs as targets of human antibodies is important for understanding immunity to malaria, but has been challenging to achieve. Recently, we developed a novel approach using genetically modified P. falciparum with inhibited PfEMP1 expression to evaluate the significance of PfEMP1 and other antigens as targets of acquired antibodies [102]. Suppressed PfEMP1 surface expression was achieved by the transfection of P. falciparum with a construct that encodes a var promoter without a downstream var gene [100, 101]. During in vitro culture with drug-selectable markers, the var promoter is expressed, which causes the silencing of all the other endogenous var promoters and thus suppresses PfEMP1 expression [100, 101] (Fig. 2). This approach was applied to human studies in Kenya to quantify serum antibodies to VSAs. We found that among malaria-exposed individuals, IgG binding to the surface of erythrocytes infected with the transgenic parasites was markedly reduced compared to that seen with parental parasites expressing PfEMP1. This suggests that the majority of the acquired human antibody response to the IE surface targets PfEMP1, while other VSAs appear to play a minor role as antibody targets. Our longitudinal studies further showed that individuals with PfEMP1-specific antibodies had a reduced risk of symptomatic disease while antibodies to other VSAs were not associated with protective immunity. Together, our findings demonstrate the significance of PfEMP1 as a major target of humoral immunity to malaria [102].

Evaluating the antibody response to PfEMP1 using transgenic P. falciparum P. falciparum-infected erythrocytes transfected with a construct that inhibits PfEMP1 expression but does not appear to have an impact on the expression of other VSAs (referred to as ‘var promoter knockdown’). This provides a novel approach to quantify antibodies to PfEMP1 and assess its importance as an immune target. The figure shows a representative selection of serum samples that were tested for IgG binding to parental and transgenic parasites [102]. Samples were from malaria-exposed Kenyan adults (K2-K16) and non-exposed Melbourne residents (Control). IgG binding to the surface of erythrocytes infected with the transgenic parasites was markedly reduced compared to parental parasites as previously reported [102]. The horizontal dotted line represents the mean level of IgG binding to parental parasites (n = 8); bars represent mean and range of samples tested in duplicate; IgG levels are expressed as geometric mean fluorescence intensity for both graphs
Human antibodies to RIFIN and STEVOR
Data suggest that RIFIN and STEVOR may play significant roles as targets of malaria immunity; however, they have been little studied compared to PfEMP1. In an area of intense malaria transmission in Gabon, high levels of antibodies to recombinant RIFIN were detected in a majority of the adult population. Although RIFIN antibodies were also detected in children, the prevalence of these antibodies was much lower [179]. Despite the high copy number of rif genes, most adult sera were capable of recognising more than one RIFIN variant suggesting the generation of a large anti-RIFIN repertoire. In addition, elevated levels of RIFIN antibodies were associated with rapid parasite clearance in children [251]. Longitudinal studies with these children showed that although RIFIN antibodies were not correlated with a reduced rate of reinfection, RIFIN antibodies were long lived (~2 years) [251]. Furthermore, higher levels of RIFIN antibodies were detected in asymptomatic children than in those with severe disease, suggesting a protective effect of these antibodies [251]. The preadsorption of immune sera on recombinant RIFIN resulted in a marked reduction in the overall antibody reactivity to the IE surface [252]. This study proposed that in addition to PfEMP1, RIFIN is a key contributor to the overall anti-VSA response [252]. Others have shown that severe malaria patients in Ghana had substantially higher antibody levels to recombinant RIFIN than asymptomatic controls, suggesting the effect of antibody boosting during a malaria episode [253].
Little has been done on naturally acquired antibodies to STEVOR. Adult plasma had elevated levels of STEVOR antibodies suggesting the immunogenicity of STEVOR during a natural infection [254]. A longitudinal study with 9-month-old infants found no correlation between STEVOR antibodies and protective immunity, but revealed an increase in the frequency of parasitaemic episodes in those with high levels of antibodies [254]. The explanation for this observation remains unclear, but it is speculated that STEVOR is not involved in mediating immunity and acts as a marker of malaria exposure instead [254]. Further studies are necessary to elucidate the importance of antibodies against native STEVOR to fully understand its biological role. As noted above, our study using parasites with suppressed PfEMP1 expression demonstrated that PfEMP1 is the dominant target of human antibodies. However, a proportion of antibody reactivity to the transgenic parasites was observed, suggesting that antibodies to other VSAs, such as RIFIN and STEVOR, may still play an important role in immunity [102].
Function of antibodies to VSAs
The mechanism by which antibodies to VSAs mediate protective immunity is only partially understood. Antibodies targeting VSAs are also thought to confer protection by interfering with IE sequestration or rosetting, features that contribute to malaria pathogenesis [79, 255]. Immune sera from infected Aotus monkeys blocked the binding of IEs to endothelial cells [18, 256]. Serum samples from pregnant women were capable of inhibiting IE adhesion to CSA [156, 257–259] and these antibodies were associated with improved birth outcomes in some studies [58, 260, 261]. In contrast, few studies have addressed adhesion inhibition in non-pregnant individuals. Convalescent serum from PNG children with symptomatic malaria inhibited the binding of homologous isolates to melanoma cells [230]. Antibodies from immune African adults inhibited the binding of a recombinant PfEMP1 domain to ICAM-1 [262]. Taken together, these results suggest the importance of antibodies that inhibit adhesion. Plasma antibodies from children presenting with mild malaria were capable of disrupting rosette formation in vitro, whereas those from children with severe malaria could not, suggesting that acquired antibodies are protective through rosette inhibition [79, 80, 150]. Furthermore, polyclonal antibodies against recombinant PfEMP1 domains disrupted existing rosettes and inhibited the formation of new rosettes [263].
Antibodies to VSAs also play a role in opsonising IEs for phagocytosis, an important mechanism of parasite clearance [264, 265]. A study with pregnant Malawian women showed that high levels of opsonising antibodies targeting VSAs correlated with parasite clearance and a decreased risk of maternal anaemia [266]. Immunisation of rabbits with recombinant PfEMP1 domains generated antibodies capable of inducing the opsonic phagocytosis of IEs [263]. Studies have also found that co-infection with HIV impaired the opsonic activity of antibodies for phagocytosis, thus leading to an increased risk of clinical malaria [267, 268]. Our recent data further identified PfEMP1 as a major target of naturally acquired antibodies that function to opsonise IEs for phagocytic clearance [102]. Individuals with high levels of antibodies to native PfEMP1 expressed on the IE surface promoted opsonic phagocytosis activity compared to transgenic parasites with inhibited PfEMP1 expression [102].
Vaccine studies on PfEMP1
The importance of PfEMP1 as an immune target strongly supports the development of PfEMP1 as a major vaccine candidate. However, a major challenge to its development as a vaccine is substantial antigenic diversity. Studies with animal models have provided evidence that recombinant PfEMP1 is capable of mounting a protective immune response. Immunisation of Aotus monkeys with the CIDRα domain of PfEMP1 protected against a lethal parasite challenge with the homologous, but not the heterologous parasite strain [269]. To overcome the variant-specific limitations of PfEMP1 antibodies, studies have used different combinations of PfEMP1 domains to elicit a broader antibody response. Mice immunised with a combination of CIDRα domains developed antibodies capable of agglutinating IEs using various parasite lines [270, 271]. Using an in vivo model of P. falciparum-IE sequestration, rats immunised with diverse NTS-DBLα domains induced protective antibodies that reduced IE sequestration [272]. This was supported by a recent study in The Netherlands where naïve volunteers, who were infected with P. falciparum, generated cross-reactive antibodies that recognised PfEMP1 from different parasite genomes [273]. Taken together, these studies suggest that it may be possible to induce sufficient cross-reactive antibodies to protect against several PfEMP1 variants, provided that the specific combinations of domains are known. A recent study further demonstrated that rabbits immunised with different recombinant proteins based on the extracellular domains of PfEMP1 recognised native PfEMP1 on intact IEs [263]. These antibodies were also capable of inhibiting rosette formation and promoting the opsonic phagocytosis of IEs [263], suggesting that the inclusion of multiple domains is necessary for effective immunity.
Research efforts on PfEMP1-specific vaccines have centred on the DBLα domain because it is one of the most conserved domains of PfEMP1 and is involved in rosetting [274]. Immunisation of rats with recombinant protein based on the DBLα domain induced antibodies that recognised conserved PfEMP1 peptides [275]. However, these antibodies were not reactive towards the IE surface of intact, mature trophozoites or towards full-length PfEMP1 from different laboratory strains [275]. They were also unable to agglutinate different parasite lines or disrupt rosette formation [275]. Antibodies against recombinant DBLα were reported to inhibit rosette formation in another study [276]. The discrepancy between these two studies [275, 276] may be reflected by different methods of protein expression as the latter, but not the former, utilised protein refolding techniques to obtain conformational-dependent epitopes that may be necessary for antibody recognition [276]. Moreover, antibodies induced by a recombinant mini-PfEMP1 (DBLα-TM-ATS) disrupted preformed rosettes and prevented in vivo sequestration [277]. The importance of the DBLα domain was further supported by the marked reduction in IE sequestration in DBLα-immunised animal models [278].
The PfEMP1 variant, VAR2CSA, is a vaccine candidate for protection against malaria in pregnancy. High levels of antibodies to multiple VAR2CSA domains in acquired pregnant women through natural exposure were associated with reduced placental infection with P. falciparum [279, 280] in some studies. Furthermore, antibodies generated against full-length [160, 281] and single domains of VAR2CSA [282] by immunisation inhibited adhesion of IEs to CSA, suggesting that vaccine-induced antibodies may have a protective function in vivo.
A major obstacle in the development of PfEMP1 as a vaccine against P. falciparum malaria is its substantial level of antigenic diversity. Several strategies can be pursued as an approach to overcome antigenic diversity (reviewed in [283]; Fig. 3). One approach would be to develop a multivalent PfEMP1 vaccine that can induce a broad repertoire of antibodies against most variants. A priority of this approach would be to determine the extent of diversity in PfEMP1 and define a combination of PfEMP1 variants that is needed to generate a broad immune response (Table 3). This approach has been used successfully with the merozoite protein apical membrane antigen 1 to overcome antigenic diversity [284, 285]. Another approach would be to target conserved epitopes of PfEMP1 such that induced antibodies may recognise most PfEMP1 variants expressed. However, identifying conserved epitopes exposed on the IE surface and understanding the tertiary/quaternary structure of PfEMP1 remains highly challenging. Further studies and innovative approaches to target antibody responses towards conserved PfEMP1 epitopes are needed. Studies have demonstrated that naturally acquired cross-reactive antibodies do occur [232, 233] and can be induced by immunisation [271]. Additionally, defining effector mechanisms of PfEMP1 immunity and creating a reference panel of parasite isolates for the evaluation of vaccine candidates must be a priority. Of further importance is a detailed knowledge of the acquisition, boosting and maintenance of antibodies to PfEMP1, as this will impact on vaccine efficacy and durability, but only limited data are currently available. Ideally, malaria vaccines would induce long-lived protection via immune responses that were sustained for an extended period after vaccination and boosted after exposure. One study suggested that antibodies to some VSAs may be short-lived, whereas other responses are sustained [286]. A recent study in pregnant women suggested that antibodies to VAR2CSA may be maintained for several decades, whereas antibodies to merozoite antigens declined more quickly [287]. The durability of vaccine-induced immune responses is not well known, but is an important issue for the development of highly efficacious vaccines against malaria (Table 3).

Approaches to overcome antigenic diversity of PfEMP1 in vaccine development. Antigenic diversity is the major challenge to developing PfEMP1 as a vaccine against malaria. The flow chart provides an overview of the two broad approaches to overcoming antigenic diversity in PfEMP1 and the steps involved in progressing vaccine candidates to the clinical trial stage. One approach is to develop a multivalent vaccine comprised of a mixture of common PfEMP1 variants that induces a broad repertoire of antibodies. A second approach is to identify conserved epitopes on PfEMP1 and develop a vaccine that targets these epitopes to induce broadly cross-reactive antibodies. As discussed in the text, there are significant challenges to overcome for each approach. It is likely that any PfEMP1 candidate vaccine antigen(s) would be included in a multi-antigen approach that includes antigens from other parasite life stages to ensure the development of a highly effective vaccine
It is likely that a highly effective malaria vaccine will require a multi-antigen, multi-stage approach. Therefore, it is anticipated that any PfEMP1-based vaccine antigens would have to be included as part of a vaccine containing antigens from other stages of the parasite life cycle, such as merozoite antigens to enhance blood-stage immunity and circumsporozoite protein for induction of pre-erythrocytic immunity, and the inclusion of gametocyte antigens for transmission-blocking immunity. We suggest that after lead PfEMP1-based vaccine antigens have been identified and prioritised, they will then need to be evaluated in combination with other vaccine antigens before proceeding further to clinical trials.
Conclusion
Understanding the targets and mechanisms of human immunity is crucial for informing and advancing the development of highly effective malaria vaccines and for developing tools for measuring immunity and exposure in populations to help evaluate the impact of malaria control interventions and identify populations at risk of malaria. Multiple studies in different populations now provide strong evidence that IE surface antigens, or VSAs, are important targets of acquired protective immunity. The most important of these antigens is PfEMP1, which is a major virulence factor enabling vascular adhesion and sequestration of IEs. Studies are beginning to identify specific variants of PfEMP1 that may be common in populations or linked to disease pathogenesis and that may be targeted in vaccine development. However, there are still major gaps in our knowledge on this topic, and these are important questions for future research. Little is known about other known or proposed surface antigens and their significance as targets of immunity and new strategies, and approaches are needed to clearly define their significance in immunity. Similarly, knowledge on surface antigens of IEs with P. vivax, the second major cause of malaria, is very limited. The role of surface antigens on gametocyte-IEs needs to be determined, as antibodies to these antigens may help clear gametocyte-IEs and thereby reduce malaria transmission; currently there is great interest globally in transmission-blocking vaccines, but there are few strong candidates in development. A greater understanding of effector mechanisms that mediate immunity is needed, including both humoral and cell-mediated responses, and additional assays to measure antibody functional activity in studies of acquired immunity and in vaccine trials would be valuable. Finally, strategies to overcome antigenic diversity in PfEMP1 would provide an exciting new opportunity in malaria vaccine development.
Abbreviations
- ATS:
-
Acidic terminal segment of PfEMP1
- CD36:
-
Cluster of differentiation 36 receptor
- CIDR:
-
Cysteine-rich interdomain regions
- CMI:
-
Cell-mediated immunity
- CR1:
-
Complement receptor 1
- CSA:
-
Chondroitin sulphate A
- DBL:
-
Duffy binding-like domain
- HA:
-
Hyaluronic acid
- ICAM-1:
-
Intercellular adhesion molecule 1
- IE:
-
P. falciparum infected erythrocyte
- KAHRP:
-
Knob-associated histidine-rich protein
- MAHRP1:
-
Membrane-associated histidine-rich protein 1
- MC:
-
Maurer’s clefts
- MESA:
-
Mature infected erythrocyte surface antigen
- PfEMP1:
-
P. falciparum erythrocyte membrane protein 1
- PfMC-2TM:
-
P. falciparum Maurer’s clefts two transmembrane protein
- REX:
-
Ring exported proteins
- RIFIN:
-
Repetitive interspersed family proteins
- STEVOR:
-
Subtelomeric variable open reading frame proteins
- SURFIN:
-
Surface-associated interspersed gene family proteins
- VSA:
-
Variant surface antigen
References
Elliott SR, Beeson JG (2008) Estimating the burden of global mortality in children aged < 5 years by pathogen-specific causes. Clin Infect Dis 46:1794–1795
Miller LH, Baruch DI, Marsh K, Doumbo OK (2002) The pathogenic basis of malaria. Nature 415:673–679. doi:10.1038/415673a
Beeson JG, Brown GV (2002) Pathogenesis of Plasmodium falciparum malaria: the roles of parasite adhesion and antigenic variation. Cell Mol Life Sci 59:258–271
Chang K-H, Stevenson MM (2004) Malarial anaemia: mechanisms and implications of insufficient erythropoiesis during blood-stage malaria. Int J Parasitol 34:1501–1516. doi:10.1016/j.ijpara.2004.10.008
Leech JH, Barnwell JW, Miller LH, Howard RJ (1984) Identification of a strain-specific malarial antigen exposed on the surface of Plasmodium falciparum-infected erythrocytes. J Exp Med 159:1567–1575
Cheng Q, Cloonan N, Fischer K et al (1998) stevor and rif are Plasmodium falciparum multicopy gene families which potentially encode variant antigens. Mol Biochem Parasitol 97:161–176
Kyes SA, Rowe JA, Kriek N, Newbold CI (1999) Rifins: a second family of clonally variant proteins expressed on the surface of red cells infected with Plasmodium falciparum. Proc Natl Acad Sci USA 96:9333–9338
Fernandez V, Hommel M, Chen Q et al (1999) Small, clonally variant antigens expressed on the surface of the Plasmodium falciparum-infected erythrocyte are encoded by the rif gene family and are the target of human immune responses. J Exp Med 190:1393–1404
Kaviratne M, Khan SM, Jarra W, Preiser PR (2002) Small variant STEVOR antigen is uniquely located within Maurer’s clefts in Plasmodium falciparum-infected red blood cells. Eukaryot Cell 1:926–935
Blythe JE, Yan Yam X, Kuss C et al (2008) Plasmodium falciparum STEVOR proteins are highly expressed in patient isolates and located in the surface membranes of infected red blood cells and the apical tips of merozoites. Infect Immun 76:3329–3336. doi:10.1128/IAI.01460-07
Niang M, Yam XY, Preiser PR (2009) The Plasmodium falciparum STEVOR multigene family mediates antigenic variation of the infected erythrocyte. PLoS Pathog 5:e1000307
Winter G, Kawai S, Haeggström M et al (2005) SURFIN is a polymorphic antigen expressed on Plasmodium falciparum merozoites and infected erythrocytes. J Exp Med 201:1853–1863. doi:10.1084/jem.20041392
Lavazec C, Sanyal S, Templeton TJ (2006) Hypervariability within the Rifin, Stevor and Pfmc-2TM superfamilies in Plasmodium falciparum. Nucleic Acids Res 34:6696–6707. doi:10.1093/nar/gkl942
Sam-Yellowe TY, Florens L, Johnson JR et al (2004) A Plasmodium gene family encoding Maurer’s cleft membrane proteins: structural properties and expression profiling. Genome Res 14:1052–1059
Winograd E, Sherman IW (2004) Malaria infection induces a conformational change in erythrocyte band 3 protein. Mol Biochem Parasitol 138:83–87. doi:10.1016/j.molbiopara.2004.07.008
Sherman IW, Crandall IE, Guthrie N, Land KM (1995) The sticky secrets of sequestration. Parasitol Today (Regul Ed) 11:378–384
MacPherson GG, Warrell MJ, White NJ et al (1985) Human cerebral malaria. A quantitative ultrastructural analysis of parasitized erythrocyte sequestration. Am J Pathol 119:385–401
Udeinya IJ, Schmidt JA, Aikawa M et al (1981) Falciparum malaria-infected erythrocytes specifically bind to cultured human endothelial cells. Science 213:555–557
Looareesuwan S, Merry AH, Phillips RE et al (1987) Reduced erythrocyte survival following clearance of malarial parasitaemia in Thai patients. Br J Haematol 67:473–478
Langreth SG, Peterson E (1985) Pathogenicity, stability, and immunogenicity of a knobless clone of Plasmodium falciparum in Colombian owl monkeys. Infect Immun 47:760–766
Pye D, O’Brien CM, Franchina P et al (1994) Plasmodium falciparum infection of splenectomized and intact Guyanan Saimiri monkeys. J Parasitol 80:558–562
Aikawa M (1988) Human cerebral malaria. Am J Trop Med Hyg 39:3–10
Pongponratn E, Riganti M, Punpoowong B, Aikawa M (1991) Microvascular sequestration of parasitized erythrocytes in human falciparum malaria: a pathological study. Am J Trop Med Hyg 44:168–175
Beeson JG, Amin N, Kanjala M, Rogerson SJ (2002) Selective accumulation of mature asexual stages of Plasmodium falciparum-infected erythrocytes in the placenta. Infect Immun 70:5412–5415
Walter PR, Garin Y, Blot P (1982) Placental pathologic changes in malaria. A histologic and ultrastructural study. Am J Pathol 109:330–342
Trager W, Rudzinska MA, Bradbury PC (1966) The fine structure of Plasmodium falciparum and its host erythrocytes in natural malarial infections in man. Bull World Health Organ 35:883–885
Kilejian A (1979) Characterization of a protein correlated with the production of knob-like protrusions on membranes of erythrocytes infected with Plasmodium falciparum. Proc Natl Acad Sci USA 76:4650–4653
Gruenberg J, Allred DR, Sherman IW (1983) Scanning electron microscope-analysis of the protrusions (knobs) present on the surface of Plasmodium falciparum-infected erythrocytes. J Cell Biol 97:795–802
Pologe LG, Pavlovec A, Shio H, Ravetch JV (1987) Primary structure and subcellular localization of the knob-associated histidine-rich protein of Plasmodium falciparum. Proc Natl Acad Sci USA 84:7139–7143
Culvenor JG, Langford CJ, Crewther PE et al (1987) Plasmodium falciparum: identification and localization of a knob protein antigen expressed by a cDNA clone. Exp Parasitol 63:58–67
Hadley TJ, Leech JH, Green TJ et al (1983) A comparison of knobby (K+) and knobless (K−) parasites from two strains of Plasmodium falciparum. Mol Biochem Parasitol 9:271–278
Leech JH, Barnwell JW, Aikawa M et al (1984) Plasmodium falciparum malaria: association of knobs on the surface of infected erythrocytes with a histidine-rich protein and the erythrocyte skeleton. J Cell Biol 98:1256–1264
Pasloske BL, Baruch DI, Van Schravendijk MR et al (1993) Cloning and characterization of a Plasmodium falciparum gene encoding a novel high-molecular weight host membrane-associated protein, PfEMP3. Mol Biochem Parasitol 59:59–72
Coppel RL, Culvenor JG, Bianco AE et al (1986) Variable antigen associated with the surface of erythrocytes infected with mature stages of Plasmodium falciparum. Mol Biochem Parasitol 20:265–277
Howard RJ, Barnwell JW, Rock EP et al (1988) Two approximately 300 kilodalton Plasmodium falciparum proteins at the surface membrane of infected erythrocytes. Mol Biochem Parasitol 27:207–223
Kilejian A, Rashid MA, Parra M, Yang YF (1991) Sequence of the knob protein of Plasmodium falciparum recognized by a monoclonal antibody. Mol Biochem Parasitol 48:231–233
Pei X, An X, Guo X et al (2005) Structural and functional studies of interaction between Plasmodium falciparum knob-associated histidine-rich protein (KAHRP) and erythrocyte spectrin. J Biol Chem 280:31166–31171. doi:10.1074/jbc.M505298200
Deitsch KW, Wellems TE (1996) Membrane modifications in erythrocytes parasitized by Plasmodium falciparum. Mol Biochem Parasitol 76:1–10
Glenister FK, Coppel RL, Cowman AF et al (2002) Contribution of parasite proteins to altered mechanical properties of malaria-infected red blood cells. Blood 99:1060–1063
Baruch DI, Pasloske BL, Singh HB et al (1995) Cloning the P. falciparum gene encoding PfEMP1, a malarial variant antigen and adherence receptor on the surface of parasitized human erythrocytes. Cell 82:77–87
Cooke BM, Glenister FK, Mohandas N, Coppel RL (2002) Assignment of functional roles to parasite proteins in malaria-infected red blood cells by competitive flow-based adhesion assay. Br J Haematol 117:203–211
Crabb BS, Cooke BM, Reeder JC et al (1997) Targeted gene disruption shows that knobs enable malaria-infected red cells to cytoadhere under physiological shear stress. Cell 89:287–296
Sherman IW, Eda S, Winograd E (2003) Cytoadherence and sequestration in Plasmodium falciparum: defining the ties that bind. Microbes Infect 5:897–909
Oquendo P, Hundt E, Lawler J, Seed B (1989) CD36 directly mediates cytoadherence of Plasmodium falciparum parasitized erythrocytes. Cell 58:95–101
Barnwell JW, Asch AS, Nachman RL et al (1989) A human 88-kD membrane glycoprotein (CD36) functions in vitro as a receptor for a cytoadherence ligand on Plasmodium falciparum-infected erythrocytes. J Clin Invest 84:765–772. doi:10.1172/JCI114234
Berendt AR, Simmons DL, Tansey J et al (1989) Intercellular adhesion molecule-1 is an endothelial cell adhesion receptor for Plasmodium falciparum. Nature 341:57–59. doi:10.1038/341057a0
Baruch DI, Ma XC, Singh HB et al (1997) Identification of a region of PfEMP1 that mediates adherence of Plasmodium falciparum infected erythrocytes to CD36: conserved function with variant sequence. Blood 90:3766–3775
Smith JD, Craig AG, Kriek N et al (2000) Identification of a Plasmodium falciparum intercellular adhesion molecule-1 binding domain: a parasite adhesion trait implicated in cerebral malaria. Proc Natl Acad Sci USA 97:1766–1771. doi:10.1073/pnas.040545897
Reeder JC, Cowman AF, Davern KM et al (1999) The adhesion of Plasmodium falciparum-infected erythrocytes to chondroitin sulfate A is mediated by P. falciparum erythrocyte membrane protein 1. Proc Natl Acad Sci USA 96:5198–5202
Buffet PA, Gamain B, Scheidig C et al (1999) Plasmodium falciparum domain mediating adhesion to chondroitin sulfate A: a receptor for human placental infection. Proc Natl Acad Sci USA 96:12743–12748
Rowe JA, Moulds JM, Newbold CI, Miller LH (1997) P. falciparum resetting mediated by a parasite-variant erythrocyte membrane protein and complement receptor 1. Nature 388:292–295. doi:10.1038/40888
Chen Q, Fernandez V, Sundström A et al (1998) Developmental selection of var gene expression in Plasmodium falciparum. Nature 394:392–395. doi:10.1038/28660
Ockenhouse CF, Tegoshi T, Maeno Y et al (1992) Human vascular endothelial cell adhesion receptors for Plasmodium falciparum-infected erythrocytes: roles for endothelial leukocyte adhesion molecule 1 and vascular cell adhesion molecule 1. J Exp Med 176:1183–1189
McCormick CJ, Craig A, Roberts D et al (1997) Intercellular adhesion molecule-1 and CD36 synergize to mediate adherence of Plasmodium falciparum-infected erythrocytes to cultured human microvascular endothelial cells. J Clin Invest 100:2521–2529. doi:10.1172/JCI119794
Rowe JA, Claessens A, Corrigan RA, Arman M (2009) Adhesion of Plasmodium falciparum-infected erythrocytes to human cells: molecular mechanisms and therapeutic implications. Expert Rev Mol Med 11:e16. doi:10.1017/S1462399409001082
Beeson JG, Brown GV, Molyneux ME et al (1999) Plasmodium falciparum isolates from infected pregnant women and children are associated with distinct adhesive and antigenic properties. J Infect Dis 180:464–472. doi:10.1086/314899
Beeson JG, Rogerson SJ, Elliott SR, Duffy MF (2005) Targets of protective antibodies to malaria during pregnancy. J Infect Dis 192:1647–1650. doi:10.1086/496895
Beeson JG, Duffy PE (2005) The immunology and pathogenesis of malaria during pregnancy. Curr Top Microbiol Immunol 297:187–227
Rogerson SJ, Chaiyaroj SC, Ng K et al (1995) Chondroitin sulfate A is a cell surface receptor for Plasmodium falciparum-infected erythrocytes. J Exp Med 182:15–20
Fried M, Duffy PE (1996) Adherence of Plasmodium falciparum to chondroitin sulfate A in the human placenta. Science 272:1502–1504
Beeson JG, Rogerson SJ, Brown GV (2002) Evaluating specific adhesion of Plasmodium falciparum-infected erythrocytes to immobilised hyaluronic acid with comparison to binding of mammalian cells. Int J Parasitol 32:1245–1252
Beeson JG, Rogerson SJ, Cooke BM et al (2000) Adhesion of Plasmodium falciparum-infected erythrocytes to hyaluronic acid in placental malaria. Nat Med 6:86–90. doi:10.1038/71582
Beeson JG, Brown GV (2004) Plasmodium falciparum-infected erythrocytes demonstrate dual specificity for adhesion to hyaluronic acid and chondroitin sulfate A and have distinct adhesive properties. J Infect Dis 189:169–179. doi:10.1086/380975
Barfod L, Dalgaard MB, Pleman ST et al (2011) Evasion of immunity to Plasmodium falciparum malaria by IgM masking of protective IgG epitopes in infected erythrocyte surface-exposed PfEMP1. Proc Natl Acad Sci 108:12485–12490. doi:10.1073/pnas.1103708108
Rowe JA, Shafi J, Kai OK et al (2002) Nonimmune IgM, but not IgG binds to the surface of Plasmodium falciparum-infected erythrocytes and correlates with rosetting and severe malaria. Am J Trop Med Hyg 66:692–699
Creasey AM, Staalsoe T, Raza A et al (2003) Nonspecific immunoglobulin M binding and chondroitin sulfate A binding are linked phenotypes of Plasmodium falciparum isolates implicated in malaria during pregnancy. Infect Immun 71:4767–4771. doi:10.1128/IAI.71.8.4767-4771.2003
Flick K, Scholander C, Chen Q et al (2001) Role of nonimmune IgG bound to PfEMP1 in placental malaria. Science 293:2098–2100. doi:10.1126/science.1062891
Turner GD, Morrison H, Jones M et al (1994) An immunohistochemical study of the pathology of fatal malaria. Evidence for widespread endothelial activation and a potential role for intercellular adhesion molecule-1 in cerebral sequestration. Am J Pathol 145:1057–1069
Newbold CI, Craig AG, Kyes S et al (1997) PfEMP1, polymorphism and pathogenesis. Ann Trop Med Parasitol 91:551–557
Rogerson SJ, Tembenu R, Dobaño C et al (1999) Cytoadherence characteristics of Plasmodium falciparum-infected erythrocytes from Malawian children with severe and uncomplicated malaria. Am J Trop Med Hyg 61:467–472
Ochola LB, Siddondo BR, Ocholla H et al (2011) Specific receptor usage in Plasmodium falciparum cytoadherence is associated with disease outcome. PLoS ONE 6:e14741. doi:10.1371/journal.pone.0014741
Turner L, Lavstsen T, Berger SS et al (2013) Severe malaria is associated with parasite binding to endothelial protein C receptor. Nature. doi:10.1038/nature12216
Roberts DD, Sherwood JA, Spitalnik SL et al (1985) Thrombospondin binds falciparum malaria parasitized erythrocytes and may mediate cytoadherence. Nature 318:64–66
Treutiger CJ, Heddini A, Fernandez V et al (1997) PECAM-1/CD31, an endothelial receptor for binding Plasmodium falciparum-infected erythrocytes. Nat Med 3:1405–1408
Ho M, Schollaardt T, Niu X et al (1998) Characterization of Plasmodium falciparum-infected erythrocyte and P-selectin interaction under flow conditions. Blood 91:4803–4809
David PH, Handunnetti SM, Leech JH et al (1988) Rosetting: a new cytoadherence property of malaria-infected erythrocytes. Am J Trop Med Hyg 38:289–297
Kaul DK, Roth EF, Nagel RL et al (1991) Rosetting of Plasmodium falciparum-infected red blood cells with uninfected red blood cells enhances microvascular obstruction under flow conditions. Blood 78:812–819
Wahlgren M, Fernandez V, Scholander C, Carlson J (1994) Rosetting. Parasitol Today (Regul Ed) 10:73–79
Carlson J, Helmby H, Hill AV et al (1990) Human cerebral malaria: association with erythrocyte rosetting and lack of anti-rosetting antibodies. Lancet 336:1457–1460
Treutiger CJ, Hedlund I, Helmby H et al (1992) Rosette formation in Plasmodium falciparum isolates and anti-rosette activity of sera from Gambians with cerebral or uncomplicated malaria. Am J Trop Med Hyg 46:503–510
Rowe A, Obeiro J, Newbold CI, Marsh K (1995) Plasmodium falciparum rosetting is associated with malaria severity in Kenya. Infect Immun 63:2323–2326
Rowe JA, Kyes SA, Rogerson SJ et al (2002) Identification of a conserved Plasmodium falciparum var gene implicated in malaria in pregnancy. J Infect Dis 185:1207–1211. doi:10.1086/339684
Doumbo OK, Thera MA, Koné AK et al (2009) High levels of Plasmodium falciparum rosetting in all clinical forms of severe malaria in African children. Am J Trop Med Hyg 81:987–993. doi:10.4269/ajtmh.2009.09-0406
al-Yaman F, Genton B, Mokela D et al (1995) Human cerebral malaria: lack of significant association between erythrocyte rosetting and disease severity. Trans R Soc Trop Med Hyg 89:55–58
Chen Q, Barragan A, Fernandez V et al (1998) Identification of Plasmodium falciparum erythrocyte membrane protein 1 (PfEMP1) as the rosetting ligand of the malaria parasite P. falciparum. J Exp Med 187:15–23
Roberts DJ, Craig AG, Berendt AR et al (1992) Rapid switching to multiple antigenic and adhesive phenotypes in malaria. Nature 357:689–692. doi:10.1038/357689a0
Roberts DJ, Pain A, Kai O et al (2000) Autoagglutination of malaria-infected red blood cells and malaria severity. Lancet 355:1427–1428. doi:10.1016/S0140-6736(00)02143-7
Pain A, Ferguson DJ, Kai O et al (2001) Platelet-mediated clumping of Plasmodium falciparum-infected erythrocytes is a common adhesive phenotype and is associated with severe malaria. Proc Natl Acad Sci USA 98:1805–1810. doi:10.1073/pnas.98.4.1805
Chotivanich K, Sritabal J, Udomsangpetch R et al (2004) Platelet-induced autoagglutination of Plasmodium falciparum-infected red blood cells and disease severity in Thailand. J Infect Dis 189:1052–1055. doi:10.1086/381900
McMorran BJ, Marshall VM, de Graaf C et al (2009) Platelets kill intraerythrocytic malarial parasites and mediate survival to infection. Science 323:797–800
McMorran BJ, Wieczorski L, Drysdale KE et al (2012) Platelet factor 4 and Duffy antigen required for platelet killing of Plasmodium falciparum. Science 338:1348–1351. doi:10.1126/science.1228892
Bull PC, Lowe BS, Kortok M, Marsh K (1999) Antibody recognition of Plasmodium falciparum erythrocyte surface antigens in Kenya: evidence for rare and prevalent variants. Infect Immun 67:733–739
Piper KP, Roberts DJ, Day KP (1999) Plasmodium falciparum: analysis of the antibody specificity to the surface of the trophozoite-infected erythrocyte. Exp Parasitol 91:161–169. doi:10.1006/expr.1998.4368
Howard RJ, Barnwell JW, Kao V (1983) Antigenic variation of Plasmodium knowlesi malaria: identification of the variant antigen on infected erythrocytes. Proc Natl Acad Sci USA 80:4129–4133
al-Khedery B, Barnwell JW, Galinski MR (1999) Antigenic variation in malaria: a 3′ genomic alteration associated with the expression of a P. knowlesi variant antigen. Mol Cell 3:131–141
Korir CC, Galinski MR (2006) Proteomic studies of Plasmodium knowlesi SICA variant antigens demonstrate their relationship with P. falciparum EMP1. Infect Genet Evol 6:75–79. doi:10.1016/j.meegid.2005.01.003
del Portillo HA, Fernandez-Becerra C, Bowman S et al (2001) A superfamily of variant genes encoded in the subtelomeric region of Plasmodium vivax. Nature 410:839–842. doi:10.1038/35071118
Janssen CS, Barrett MP, Turner CMR, Phillips RS (2002) A large gene family for putative variant antigens shared by human and rodent malaria parasites. Proc Biol Sci 269:431–436. doi:10.1098/rspb.2001.1903
Janssen CS, Phillips RS, Turner CMR, Barrett MP (2004) Plasmodium interspersed repeats: the major multigene superfamily of malaria parasites. Nucleic Acids Res 32:5712–5720. doi:10.1093/nar/gkh907
Voss TS, Healer J, Marty AJ et al (2006) A var gene promoter controls allelic exclusion of virulence genes in Plasmodium falciparum malaria. Nature 439:1004–1008. doi:10.1038/nature04407
Dzikowski R, Frank M, Deitsch K (2006) Mutually exclusive expression of virulence genes by malaria parasites is regulated independently of antigen production. PLoS Pathog 2:e22. doi:10.1371/journal.ppat.0020022
Chan J-A, Howell KB, Reiling L et al (2012) Targets of antibodies against Plasmodium falciparum-infected erythrocytes in malaria immunity. J Clin Invest. doi:10.1172/JCI62182
Howitt CA, Wilinski D, Llinás M et al (2009) Clonally variant gene families in Plasmodium falciparum share a common activation factor. Mol Microbiol 73:1171–1185. doi:10.1111/j.1365-2958.2009.06846.x
Garcia CR, Takeuschi M, Yoshioka K, Miyamoto H (1997) Imaging Plasmodium falciparum-infected ghost and parasite by atomic force microscopy. J Struct Biol 119:92–98. doi:10.1006/jsbi.1997.3886
Marti M (2004) Targeting malaria virulence and remodeling proteins to the host erythrocyte. Science 306:1930–1933. doi:10.11269/science.1102452
Hiller NL, Bhattacharjee S, van Ooij C et al (2004) A host-targeting signal in virulence proteins reveals a secretome in malarial infection. Science 306:1934–1937. doi:10.1126/science.1102737
de Koning-Ward TF, Gilson PR, Boddey JA et al (2009) A newly discovered protein export machine in malaria parasites. Nature 459:945–949. doi:10.1038/nature08104
Crabb BS, de Koning-Ward TF, Gilson PR (2010) Protein export in Plasmodium parasites: from the endoplasmic reticulum to the vacuolar export machine. Int J Parasitol 40:509–513. doi:10.1016/j.ijpara.2010.02.002
Mantel P-Y, Hoang AN, Goldowitz I et al (2013) Malaria-infected erythrocyte-derived microvesicles mediate cellular communication within the parasite population and with the host immune system. Cell Host Microbe 13:521–534. doi:10.1016/j.chom.2013.04.009
Regev-Rudzki N, Wilson DW, Carvalho TG et al (2013) Cell–cell communication between malaria-infected red blood cells via exosome-like vesicles. Cell. doi:10.1016/j.cell.2013.04.029
Su XZ, Heatwole VM, Wertheimer SP et al (1995) The large diverse gene family var encodes proteins involved in cytoadherence and antigenic variation of Plasmodium falciparum-infected erythrocytes. Cell 82:89–100
Smith JD, Chitnis CE, Craig AG et al (1995) Switches in expression of Plasmodium falciparum var genes correlate with changes in antigenic and cytoadherent phenotypes of infected erythrocytes. Cell 82:101–110
Scherf A, Hernandez-Rivas R, Buffet P et al (1998) Antigenic variation in malaria: in situ switching, relaxed and mutually exclusive transcription of var genes during intra-erythrocytic development in Plasmodium falciparum. EMBO J 17:5418–5426. doi:10.1093/emboj/17.18.5418
Joergensen L, Bengtsson DC, Bengtsson A et al (2010) Surface co-expression of two different PfEMP1 antigens on single Plasmodium falciparum-infected erythrocytes facilitates binding to ICAM1 and PECAM1. PLoS Pathog 6:e1001083. doi:10.1371/journal.ppat.1001083
Kriek N, Tilley L, Horrocks P et al (2003) Characterization of the pathway for transport of the cytoadherence-mediating protein, PfEMP1, to the host cell surface in malaria parasite-infected erythrocytes. Mol Microbiol 50:1215–1227
Boddey JA, Cowman AF (2013) Plasmodium nesting: remaking the erythrocyte from the inside out. Annu Rev Microbiol 67:243–269. doi:10.1146/annurev-micro-092412-155730
Elsworth B, Crabb BS, Gilson PR (2014) Protein export in malaria parasites: an update. Cell Microbiol 16:355–363. doi:10.1111/cmi.12261
Haeggström M, Kironde F, Berzins K et al (2004) Common trafficking pathway for variant antigens destined for the surface of the Plasmodium falciparum-infected erythrocyte. Mol Biochem Parasitol 133:1–14
Horrocks P, Pinches RA, Chakravorty SJ et al (2005) PfEMP1 expression is reduced on the surface of knobless Plasmodium falciparum infected erythrocytes. J Cell Sci 118:2507–2518. doi:10.1242/jcs.02381
Gardner JP, Pinches RA, Roberts DJ, Newbold CI (1996) Variant antigens and endothelial receptor adhesion in Plasmodium falciparum. Proc Natl Acad Sci USA 93:3503–3508
Blisnick T, Morales Betoulle ME, Barale JC et al (2000) Pfsbp1, a Maurer’s cleft Plasmodium falciparum protein, is associated with the erythrocyte skeleton. Mol Biochem Parasitol 111:107–121
Cooke BM, Buckingham DW, Glenister FK et al (2006) A Maurer’s cleft-associated protein is essential for expression of the major malaria virulence antigen on the surface of infected red blood cells. J Cell Biol 172:899–908. doi:10.1083/jcb.200509122
Maier AG, Rug M, O’Neill MT et al (2007) Skeleton-binding protein 1 functions at the parasitophorous vacuole membrane to traffic PfEMP1 to the Plasmodium falciparum-infected erythrocyte surface. Blood 109:1289–1297. doi:10.1182/blood-2006-08-043364
Maier AG, Rug M, O’Neill MT et al (2008) Exported proteins required for virulence and rigidity of Plasmodium falciparum-infected human erythrocytes. Cell 134:48–61. doi:10.1016/j.cell.2008.04.051
Külzer S, Charnaud S, Dagan T et al (2012) Plasmodium falciparum-encoded exported hsp70/hsp40 chaperone/co-chaperone complexes within the host erythrocyte. Cell Microbiol 14:1784–1795. doi:10.1111/j.1462-5822.2012.01840.x
Spycher C, Rug M, Pachlatko E et al (2008) The Maurer’s cleft protein MAHRP1 is essential for trafficking of PfEMP1 to the surface of Plasmodium falciparum-infected erythrocytes. Mol Microbiol 68:1300–1314. doi:10.1111/j.1365-2958.2008.06235.x
Dixon MWA, Kenny S, McMillan PJ et al (2011) Genetic ablation of a Maurer’s cleft protein prevents assembly of the Plasmodium falciparum virulence complex. Mol Microbiol 81:982–993. doi:10.1111/j.1365-2958.2011.07740.x
Glenister FK, Fernandez KM, Kats LM et al (2009) Functional alteration of red blood cells by a megadalton protein of Plasmodium falciparum. Blood 113:919–928. doi:10.1182/blood-2008-05-157735
Duraisingh MT, Cowman AF (2005) Contribution of the pfmdr1 gene to antimalarial drug-resistance. Acta Trop 94:181–190. doi:10.1016/j.actatropica.2005.04.008
Freitas-Junior LH, Hernández-Rivas R, Ralph SA et al (2005) Telomeric heterochromatin propagation and histone acetylation control mutually exclusive expression of antigenic variation genes in malaria parasites. Cell 121:25–36. doi:10.1016/j.cell.2005.01.037
Tonkin CJ, Carret CK, Duraisingh MT et al (2009) Sir2 paralogues cooperate to regulate virulence genes and antigenic variation in Plasmodium falciparum. PLoS Biol 7:e84. doi:10.1371/journal.pbio.1000084.st002
Biggs BA, Goozé L, Wycherley K et al (1991) Antigenic variation in Plasmodium falciparum. Proc Natl Acad Sci USA 88:9171–9174
Chen Q, Heddini A, Barragan A et al (2000) The semiconserved head structure of Plasmodium falciparum erythrocyte membrane protein 1 mediates binding to multiple independent host receptors. J Exp Med 192:1–10
Voigt S, Hanspal M, LeRoy PJ et al (2000) The cytoadherence ligand Plasmodium falciparum erythrocyte membrane protein 1 (PfEMP1) binds to the P. falciparum knob-associated histidine-rich protein (KAHRP) by electrostatic interactions. Mol Biochem Parasitol 110:423–428
Waller KL, Cooke BM, Nunomura W et al (1999) Mapping the binding domains involved in the interaction between the Plasmodium falciparum knob-associated histidine-rich protein (KAHRP) and the cytoadherence ligand P. falciparum erythrocyte membrane protein 1 (PfEMP1). J Biol Chem 274:23808–23813
Rug M, Prescott SW, Fernandez KM et al (2006) The role of KAHRP domains in knob formation and cytoadherence of P. falciparum-infected human erythrocytes. Blood 108:370–378. doi:10.1182/blood-2005-11-4624
Chattopadhyay R, Taneja T, Chakrabarti K et al (2004) Molecular analysis of the cytoadherence phenotype of a Plasmodium falciparum field isolate that binds intercellular adhesion molecule-1. Mol Biochem Parasitol 133:255–265
Springer AL, Smith LM, Mackay DQ et al (2004) Functional interdependence of the DBLbeta domain and c2 region for binding of the Plasmodium falciparum variant antigen to ICAM-1. Mol Biochem Parasitol 137:55–64. doi:10.1016/j.molbiopara.2004.03.019
Robinson BA, Welch TL, Smith JD (2003) Widespread functional specialization of Plasmodium falciparum erythrocyte membrane protein 1 family members to bind CD36 analysed across a parasite genome. Mol Microbiol 47:1265–1278
Smith JD, Kyes S, Craig AG et al (1998) Analysis of adhesive domains from the A4VAR Plasmodium falciparum erythrocyte membrane protein-1 identifies a CD36 binding domain. Mol Biochem Parasitol 97:133–148
Srivastava A, Gangnard S, Dechavanne S et al (2011) Var2CSA minimal CSA binding region is located within the N-terminal region. PLoS ONE 6:e20270. doi:10.1371/journal.pone.0020270
Gardner MJ, Hall N, Fung E et al (2002) Genome sequence of the human malaria parasite Plasmodium falciparum. Nature 419:498–511. doi:10.1038/nature01097
Kraemer SM, Smith JD (2003) Evidence for the importance of genetic structuring to the structural and functional specialization of the Plasmodium falciparum var gene family. Mol Microbiol 50:1527–1538
Lavstsen T, Salanti A, Jensen ATR et al (2003) Sub-grouping of Plasmodium falciparum 3D7 var genes based on sequence analysis of coding and non-coding regions. Malar J 2:27. doi:10.1186/1475-2875-2-27
Voss TS, Thompson JK, Waterkeyn J et al (2000) Genomic distribution and functional characterisation of two distinct and conserved Plasmodium falciparum var gene 5′ flanking sequences. Mol Biochem Parasitol 107:103–115
Bull PC, Kyes S, Buckee CO et al (2007) An approach to classifying sequence tags sampled from Plasmodium falciparum var genes. Mol Biochem Parasitol 154:98–102. doi:10.1016/j.molbiopara.2007.03.011
Bull PC, Berriman M, Kyes S et al (2005) Plasmodium falciparum variant surface antigen expression patterns during malaria. PLoS Pathog 1:e26. doi:10.1371/journal.ppat.0010026.st003
Warimwe GM, Keane TM, Fegan G et al (2009) Plasmodium falciparum var gene expression is modified by host immunity. Proc Natl Acad Sci 106:21801–21806. doi:10.1073/pnas.0907590106
Kyriacou HM, Stone GN, Challis RJ et al (2006) Differential var gene transcription in Plasmodium falciparum isolates from patients with cerebral malaria compared to hyperparasitaemia. Mol Biochem Parasitol 150:211–218. doi:10.1016/j.molbiopara.2006.08.005
Falk N, Kaestli M, Qi W et al (2009) Analysis of Plasmodium falciparum var genes expressed in children from Papua New Guinea. J Infect Dis 200:347–356. doi:10.1086/600071
Rottmann M, Lavstsen T, Mugasa JP et al (2006) Differential expression of var gene groups is associated with morbidity caused by Plasmodium falciparum infection in Tanzanian children. Infect Immun 74:3904–3911. doi:10.1128/IAI.02073-05
Jensen ATR, Magistrado P, Sharp S et al (2004) Plasmodium falciparum associated with severe childhood malaria preferentially expresses PfEMP1 encoded by group A var genes. J Exp Med 199:1179–1190. doi:10.1084/jem.20040274
Lavstsen T, Turner L, Saguti F et al (2012) Plasmodium falciparum erythrocyte membrane protein 1 domain cassettes 8 and 13 are associated with severe malaria in children. Proc Natl Acad Sci. doi:10.1073/pnas.1120455109
Warimwe GM, Fegan G, Musyoki JN et al (2012) Prognostic indicators of life-threatening malaria are associated with distinct parasite variant antigen profiles. Sci Transl Med 4:129ra45. doi:10.1126/scitranslmed.3003247
Avril M, Tripathi AK, Brazier AJ et al (2012) A restricted subset of var genes mediates adherence of Plasmodium falciparum-infected erythrocytes to brain endothelial cells. Proc Natl Acad Sci. doi:10.1073/pnas.1120534109
Claessens A, Adams Y, Ghumra A et al (2012) A subset of group A-like var genes encodes the malaria parasite ligands for binding to human brain endothelial cells. Proc Natl Acad Sci. doi:10.1073/pnas.1120461109
Cham GKK, Turner L, Lusingu J et al (2009) Sequential, ordered acquisition of antibodies to Plasmodium falciparum erythrocyte membrane protein 1 domains. J Immunol 183:3356–3363. doi:10.4049/jimmunol.0901331
Kaestli M, Cockburn IA, Cortés A et al (2006) Virulence of malaria is associated with differential expression of Plasmodium falciparum var gene subgroups in a case-control study. J Infect Dis 193:1567–1574. doi:10.1086/503776
Hommel M, Elliott SR et al (2010) Evaluation of the antigenic diversity of placenta-binding Plasmodium falciparum variants and the antibody repertoire among pregnant women. Infect Immun 78:1963–1978. doi:10.1128/IAI.01365-09
Avril M, Hathaway MJ, Srivastava A et al (2011) Antibodies to a full-length VAR2CSA immunogen are broadly strain-transcendent but do not cross-inhibit different placental-type parasite isolates. PLoS ONE 6:e16622. doi:10.1371/journal.pone.0016622.t001
Kraemer SM, Kyes SA, Aggarwal G et al (2007) Patterns of gene recombination shape var gene repertoires in Plasmodium falciparum: comparisons of geographically diverse isolates. BMC Genomics 8:45. doi:10.1186/1471-2164-8-45
Duffy MF, Byrne TJ, Elliott SR et al (2005) Broad analysis reveals a consistent pattern of var gene transcription in Plasmodium falciparum repeatedly selected for a defined adhesion phenotype. Mol Microbiol 56:774–788. doi:10.1111/j.1365-2958.2005.04577.x
Salanti A, Staalsoe T, Lavstsen T et al (2003) Selective upregulation of a single distinctly structured var gene in chondroitin sulphate A-adhering Plasmodium falciparum involved in pregnancy-associated malaria. Mol Microbiol 49:179–191. doi:10.1046/j.1365-2958.2003.03570.x
Salanti A, Dahlbäck M, Turner L et al (2004) Evidence for the involvement of VAR2CSA in pregnancy-associated malaria. J Exp Med 200:1197–1203. doi:10.1084/jem.20041579
Beeson JG, Andrews KT, Boyle M et al (2007) Structural basis for binding of Plasmodium falciparum erythrocyte membrane protein 1 to chondroitin sulfate and placental tissue and the influence of protein polymorphisms on binding specificity. J Biol Chem 282:22426–22436. doi:10.1074/jbc.M700231200
Magistrado P, Salanti A, Tuikue Ndam NG et al (2008) VAR2CSA expression on the surface of placenta-derived Plasmodium falciparum-infected erythrocytes. J Infect Dis 198:1071–1074. doi:10.1086/591502
Beeson JG, Mann EJ, Elliott SR et al (2004) Antibodies to variant surface antigens of Plasmodium falciparum-infected erythrocytes and adhesion inhibitory antibodies are associated with placental malaria and have overlapping and distinct targets. J Infect Dis 189:540–551
Viebig NK, Gamain B, Scheidig C et al (2005) A single member of the Plasmodium falciparum var multigene family determines cytoadhesion to the placental receptor chondroitin sulphate A. EMBO Rep 6:775–781. doi:10.1038/sj.embor.7400466
Duffy MF, Maier AG, Byrne TJ et al (2006) VAR2CSA is the principal ligand for chondroitin sulfate A in two allogeneic isolates of Plasmodium falciparum. Mol Biochem Parasitol 148:117–124. doi:10.1016/j.molbiopara.2006.03.006
Hayward RE, Tiwari B, Piper KP et al (1999) Virulence and transmission success of the malarial parasite Plasmodium falciparum. Proc Natl Acad Sci USA 96:4563–4568
Sharp S, Lavstsen T, Fivelman QL et al (2006) Programmed transcription of the var gene family, but not of stevor, in Plasmodium falciparum gametocytes. Eukaryot Cell 5:1206–1214. doi:10.1128/EC.00029-06
Tibùrcio M, Silvestrini F, Bertuccini L et al (2012) Early gametocytes of the malaria parasite Plasmodium falciparum specifically remodel the adhesive properties of infected erythrocyte surface. Cell Microbiol. doi:10.1111/cmi.12062
Aingaran M, Zhang R, Law SK et al (2012) Host cell deformability is linked to transmission in the human malaria parasite Plasmodium falciparum. Cell Microbiol 14:983–993. doi:10.1111/j.1462-5822.2012.01786.x
Kyes S, Pinches R, Newbold C (2000) A simple RNA analysis method shows var and rif multigene family expression patterns in Plasmodium falciparum. Mol Biochem Parasitol 105:311–315
Florens L, Washburn MP, Raine JD et al (2002) A proteomic view of the Plasmodium falciparum life cycle. Nature 419:520–526. doi:10.1038/nature01107
Petter M, Haeggström M, Khattab A et al (2007) Variant proteins of the Plasmodium falciparum RIFIN family show distinct subcellular localization and developmental expression patterns. Mol Biochem Parasitol 156:51–61. doi:10.1016/j.molbiopara.2007.07.011
Wang CW, Mwakalinga SB, Sutherland CJ et al (2010) Identification of a major rif transcript common to gametocytes and sporozoites of Plasmodium falciparum. Malar J 9:147. doi:10.1186/1475-2875-9-147
Joannin N, Abhiman S, Sonnhammer EL, Wahlgren M (2008) Sub-grouping and sub-functionalization of the RIFIN multi-copy protein family. BMC Genomics 9:19. doi:10.1186/1471-2164-9-19
Abdel-Latif MS, Khattab A et al (2002) Recognition of variant Rifin antigens by human antibodies induced during natural Plasmodium falciparum infections. Infect Immun 70:7013–7021
Przyborski JM, Miller SK, Pfahler JM et al (2005) Trafficking of STEVOR to the Maurer’s clefts in Plasmodium falciparum-infected erythrocytes. EMBO J 24:2306–2317. doi:10.1038/sj.emboj.7600720
McRobert L, Preiser P, Sharp S et al (2004) Distinct trafficking and localization of STEVOR proteins in three stages of the Plasmodium falciparum life cycle. Infect Immun 72:6597–6602. doi:10.1128/IAI.72.11.6597-6602.2004
Sanyal S, Egée S, Bouyer G et al (2012) Plasmodium falciparum STEVOR proteins impact erythrocyte mechanical properties. Blood 119:e1–e8. doi:10.1182/blood-2011-08-370734
Mills JP, Diez-Silva M, Quinn DJ et al (2007) Effect of plasmodial RESA protein on deformability of human red blood cells harboring Plasmodium falciparum. Proc Natl Acad Sci USA 104:9213–9217. doi:10.1073/pnas.0703433104
Lavazec C, Sanyal S, Templeton TJ (2007) Expression switching in the stevor and Pfmc-2TM superfamilies in Plasmodium falciparum. Mol Microbiol 64:1621–1634. doi:10.1111/j.1365-2958.2007.05767.x
García JE, Puentes A, Curtidor H et al (2005) Peptides from the Plasmodium falciparum STEVOR putative protein bind with high affinity to normal human red blood cells. Peptides 26:1133–1143. doi:10.1016/j.peptides.2005.01.013
Khattab A, Meri S (2011) Exposure of the Plasmodium falciparum clonally variant STEVOR proteins on the merozoite surface. Malar J 10:58. doi:10.1186/1475-2875-10-58
Khattab A, Bonow I, Schreiber N et al (2008) Plasmodium falciparum variant STEVOR antigens are expressed in merozoites and possibly associated with erythrocyte invasion. Malar J 7:137. doi:10.1186/1475-2875-7-137
Mphande FA, Ribacke U, Kaneko O et al (2008) SURFIN4.1, a schizont-merozoite associated protein in the SURFIN family of Plasmodium falciparum. Malar J 7:116. doi:10.1186/1475-2875-7-116
Winograd E, Eda S, Sherman IW (2004) Chemical modifications of band 3 protein affect the adhesion of Plasmodium falciparum-infected erythrocytes to CD36. Mol Biochem Parasitol 136:243–248
Eda S, Lawler J, Sherman IW (1999) Plasmodium falciparum-infected erythrocyte adhesion to the type 3 repeat domain of thrombospondin-1 is mediated by a modified band 3 protein. Mol Biochem Parasitol 100:195–205
Lucas JZ, Sherman IW (1998) Plasmodium falciparum: thrombospondin mediates parasitized erythrocyte band 3-related adhesin binding. Exp Parasitol 89:78–85
Crandall I, Collins WE, Gysin J, Sherman IW (1993) Synthetic peptides based on motifs present in human band 3 protein inhibit cytoadherence/sequestration of the malaria parasite Plasmodium falciparum. Proc Natl Acad Sci USA 90:4703–4707
Langhorne J, Ndungu FM, Sponaas A-M, Marsh K (2008) Immunity to malaria: more questions than answers. Nat Immunol 9:725–732. doi:10.1038/ni.f.205
Good MF, Stanisic D, Xu H et al (2004) The immunological challenge to developing a vaccine to the blood stages of malaria parasites. Immunol Rev 201:254–267. doi:10.1111/j.0105-2896.2004.00178.x
Beeson JG, Osier FHA, Engwerda CR (2008) Recent insights into humoral and cellular immune responses against malaria. Trends Parasitol 24:578–584. doi:10.1016/j.pt.2008.08.008
Marsh K, Kinyanjui S (2006) Immune effector mechanisms in malaria. Parasite Immunol 28:51–60. doi:10.1111/j.1365-3024.2006.00808.x
Gupta S, Snow RW, Donnelly CA et al (1999) Immunity to non-cerebral severe malaria is acquired after one or two infections. Nat Med 5:340–343. doi:10.1038/6560
McGregor IA (1964) Studies in the acquisition of immunity of Plasmodium falciparum infections in Africa. Trans R Soc Trop Med Hyg 58:80–92
Snow RW, Nahlen B, Palmer A et al (1998) Risk of severe malaria among African infants: direct evidence of clinical protection during early infancy. J Infect Dis 177:819–822
Kitua AY, Smith T, Alonso PL et al (1996) Plasmodium falciparum malaria in the first year of life in an area of intense and perennial transmission. Trop Med Int Health 1:475–484
Yazdani SS, Mukherjee P, Chauhan VS, Chitnis CE (2006) Immune responses to asexual blood-stages of malaria parasites. Curr Mol Med 6:187–203
Good MF, Xu H, Wykes M, Engwerda CR (2005) Development and regulation of cell-mediated immune responses to the blood stages of malaria: implications for vaccine research. Annu Rev Immunol 23:69–99. doi:10.1146/annurev.immunol.23.021704.115638
van der Heyde HC, Huszar D, Woodhouse C et al (1994) The resolution of acute malaria in a definitive model of B cell deficiency, the JHD mouse. J Immunol 152:4557–4562
Clark IA, Cowden WB (1999) Why is the pathology of falciparum worse than that of vivax malaria? Parasitol Today (Regul Ed) 15:458–461
Luty AJ, Lell B, Schmidt-Ott R et al (1999) Interferon-gamma responses are associated with resistance to reinfection with Plasmodium falciparum in young African children. J Infect Dis 179:980–988. doi:10.1086/314689
Hansen DS, Schofield L (2010) Natural regulatory T cells in malaria: host or parasite allies? PLoS Pathog 6:e1000771. doi:10.1371/journal.ppat.1000771
Urban BC, Ferguson DJ, Pain A et al (1999) Plasmodium falciparum-infected erythrocytes modulate the maturation of dendritic cells. Nature 400:73–77. doi:10.1038/21900
Elliott SR, Spurck TP, Dodin JM et al (2007) Inhibition of dendritic cell maturation by malaria is dose dependent and does not require Plasmodium falciparum erythrocyte membrane protein 1. Infect Immun 75:3621–3632. doi:10.1128/IAI.00095-07
Artavanis-Tsakonas K, Eleme K, McQueen KL et al (2003) Activation of a subset of human NK cells upon contact with Plasmodium falciparum-infected erythrocytes. J Immunol 171:5396–5405
Chen Q, Amaladoss A, Ye W et al (2014) Human natural killer cells control Plasmodium falciparum infection by eliminating infected red blood cells. Proc Natl Acad Sci 111:1479–1484. doi:10.1073/pnas.1323318111
D’Ombrain MC, Voss TS, Maier AG et al (2007) Plasmodium falciparum erythrocyte membrane protein-1 specifically suppresses early production of host interferon-gamma. Cell Host Microbe 2:130–138. doi:10.1016/j.chom.2007.06.012
Donati D, Zhang LP, Chêne A et al (2004) Identification of a polyclonal B-cell activator in Plasmodium falciparum. Infect Immun 72:5412–5418. doi:10.1128/IAI.72.9.5412-5418.2004
Donati D, Mok B, Chêne A et al (2006) Increased B cell survival and preferential activation of the memory compartment by a malaria polyclonal B cell activator. J Immunol 177:3035–3044
Simone O, Bejarano MT, Pierce SK et al (2011) TLRs innate immunereceptors and Plasmodium falciparum erythrocyte membrane protein 1 (PfEMP1) CIDR1α-driven human polyclonal B-cell activation. Acta Trop 119:144–150. doi:10.1016/j.actatropica.2011.05.005
Allsopp CEM, Sanni LA, Reubsaet L et al (2002) CD4 T cell responses to a variant antigen of the malaria parasite Plasmodium falciparum, erythrocyte membrane protein-1, in individuals living in malaria-endemic areas. J Infect Dis 185:812–819. doi:10.1086/339521
Ndungu FM, Sanni L, Urban B et al (2006) CD4 T cells from malaria-nonexposed individuals respond to the CD36-Binding Domain of Plasmodium falciparum erythrocyte membrane protein-1 via an MHC class II-TCR-independent pathway. J Immunol 176:5504–5512
Marsh K (1992) Malaria–a neglected disease? Parasitology 104(Suppl):S53–S69
Cohen S, McGregor IA, Carrington S (1961) Gamma-globulin and acquired immunity to human malaria. Nature 192:733–737
Fowkes FJI, Richards JS, Simpson JA, Beeson JG (2010) The relationship between anti-merozoite antibodies and incidence of Plasmodium falciparum malaria: a systematic review and meta-analysis. Plos Med 7:e1000218. doi:10.1371/journal.pmed.1000218
Osier FHA, Fegan G, Polley SD et al (2008) Breadth and magnitude of antibody responses to multiple Plasmodium falciparum merozoite antigens are associated with protection from clinical malaria. Infect Immun 76:2240–2248
Richards JS, Arumugam TU, Reiling L et al (2013) Identification and prioritization of merozoite antigens as targets of protective human immunity to Plasmodium falciparum malaria for vaccine and biomarker development. J Immunol 191:795–809. doi:10.4049/jimmunol.1300778
Richards JS, Beeson JG (2009) The future for blood-stage vaccines against malaria. Immunol Cell Biol 87:377–390. doi:10.1038/icb.2009.27
Richards JS, Stanisic DI, Fowkes FJI et al (2010) Association between naturally acquired antibodies to erythrocyte-binding antigens of Plasmodium falciparum and protection from malaria and high-density parasitemia. Clin Infect Dis 51:e50–e60. doi:10.1086/656413
Roussilhon C, Oeuvray C, Müller-Graf C et al (2007) Long-term clinical protection from falciparum malaria is strongly associated with IgG3 antibodies to merozoite surface protein 3. Plos Med 4:e320. doi:10.1371/journal.pmed.0040320
Taylor DW, Zhou A, Marsillio LE et al (2004) Antibodies that inhibit binding of Plasmodium falciparum-infected erythrocytes to chondroitin sulfate A and to the C terminus of merozoite surface protein 1 correlate with reduced placental malaria in Cameroonian women. Infect Immun 72:1603–1607
Marsh K, Howard RJ (1986) Antigens induced on erythrocytes by P. falciparum: expression of diverse and conserved determinants. Science 231:150–153
Bull PC, Lowe BS, Kortok M et al (1998) Parasite antigens on the infected red cell surface are targets for naturally acquired immunity to malaria. Nat Med 4:358–360
Marsh K, Otoo L, Hayes RJ et al (1989) Antibodies to blood stage antigens of Plasmodium falciparum in rural Gambians and their relation to protection against infection. Trans R Soc Trop Med Hyg 83:293–303
Iqbal J, Perlmann P, Berzins K (1993) Serological diversity of antigens expressed on the surface of erythrocytes infected with Plasmodium falciparum. Trans R Soc Trop Med Hyg 87:583–588
Forsyth KP, Philip G, Smith T et al (1989) Diversity of antigens expressed on the surface of erythrocytes infected with mature Plasmodium falciparum parasites in Papua New Guinea. Am J Trop Med Hyg 41:259–265
Newbold CI, Pinches R, Roberts DJ, Marsh K (1992) Plasmodium falciparum: the human agglutinating antibody response to the infected red cell surface is predominantly variant specific. Exp Parasitol 75:281–292
Chattopadhyay R, Sharma A, Srivastava VK et al (2003) Plasmodium falciparum infection elicits both variant-specific and cross-reactive antibodies against variant surface antigens. Infect Immun 71:597–604
Beeson JG, Mann EJ, Byrne TJ et al (2006) Antigenic differences and conservation among placental Plasmodium falciparum-infected erythrocytes and acquisition of variant-specific and cross-reactive antibodies. J Infect Dis 193:721–730. doi:10.1086/500145
Bockhorst J, Lu F, Janes JH et al (2007) Structural polymorphism and diversifying selection on the pregnancy malaria vaccine candidate VAR2CSA. Mol Biochem Parasitol 155:103–112. doi:10.1016/j.molbiopara.2007.06.007
Giha HA, Theander TG, Staalsø T et al (1998) Seasonal variation in agglutination of Plasmodium falciparum-infected erythrocytes. Am J Trop Med Hyg 58:399–405
Bull PC, Lowe BS, Kaleli N et al (2002) Plasmodium falciparum infections are associated with agglutinating antibodies to parasite-infected erythrocyte surface antigens among healthy Kenyan children. J Infect Dis 185:1688–1691. doi:10.1086/340420
Ofori MF, Dodoo D, Staalsoe T et al (2002) Malaria-induced acquisition of antibodies to Plasmodium falciparum variant surface antigens. Infect Immun 70:2982–2988
Tebo AE, Kremsner PG, Piper KP, Luty AJF (2002) Low antibody responses to variant surface antigens of Plasmodium falciparum are associated with severe malaria and increased susceptibility to malaria attacks in Gabonese children. Am J Trop Med Hyg 67:597–603
Dodoo D, Staalsoe T, Giha H et al (2001) Antibodies to variant antigens on the surfaces of infected erythrocytes are associated with protection from malaria in Ghanaian children. Infect Immun 69:3713–3718. doi:10.1128/IAI.69.6.3713-3718.2001
Giha HA, Staalsoe T, Dodoo D et al (2000) Antibodies to variable Plasmodium falciparum-infected erythrocyte surface antigens are associated with protection from novel malaria infections. Immunol Lett 71:117–126
Aguiar J, Albrecht G, Cegielski P (1992) Agglutination of Plasmodium falciparum-infected erythrocytes from east and West African. Am J Trop Med Hyg 47:621–632
Nielsen MA, Vestergaard LS, Lusingu J et al (2004) Geographical and temporal conservation of antibody recognition of Plasmodium falciparum variant surface antigens. Infect Immun 72:3531–3535. doi:10.1128/IAI.72.6.3531-3535.2004
Nielsen MA, Staalsoe T, Kurtzhals JAL et al (2002) Plasmodium falciparum variant surface antigen expression varies between isolates causing severe and nonsevere malaria and is modified by acquired immunity. J Immunol 168:3444–3450
Saeed M, Roeffen W, Alexander N et al (2008) Plasmodium falciparum antigens on the surface of the gametocyte-infected erythrocyte. PLoS ONE 3:e2280. doi:10.1371/journal.pone.0002280
Piper KP, Hayward RE, Cox MJ, Day KP (1999) Malaria transmission and naturally acquired immunity to PfEMP1. Infect Immun 67:6369–6374
Barry AW, Trieu A, Fowkes FJI et al (2011) The stability and complexity of antibody responses to the major surface antigen of Plasmodium falciparum are associated with age in a malaria endemic area. Mol Cell Proteomics 10(M111):008326. doi:10.1074/mcp.M1111.008326
Cham GKK, Turner L, Kurtis JD et al (2010) Hierarchical, domain type-specific acquisition of antibodies to Plasmodium falciparum erythrocyte membrane Protein 1 in Tanzanian children. Infect Immun 78:4653–4659. doi:10.1128/IAI.00593-10
Oguariri RM, Borrmann S, Klinkert MQ et al (2001) High prevalence of human antibodies to recombinant Duffy binding-like alpha domains of the Plasmodium falciparum-infected erythrocyte membrane protein 1 in semi-immune adults compared to that in nonimmune children. Infect Immun 69:7603–7609. doi:10.1128/IAI.69.12.7603-7609.2001
Staalsø T, Khalil EA, Elhassan IM et al (1998) Antibody reactivity to conserved linear epitopes of Plasmodium falciparum erythrocyte membrane protein 1 (PfEMP1). Immunol Lett 60:121–126
Mackintosh CL, Christodoulou Z, Mwangi TW et al (2008) Acquisition of naturally occurring antibody responses to recombinant protein domains of Plasmodium falciparum erythrocyte membrane protein 1. Malar J 7:155. doi:10.1186/1475-2875-7-155
Abdel-Latif MS, Dietz K, Issifou S et al (2003) Antibodies to Plasmodium falciparum rifin proteins are associated with rapid parasite clearance and asymptomatic infections. Infect Immun 71:6229–6233
Abdel-Latif MS, Cabrera G, Köhler C et al (2004) Antibodies to rifin: a component of naturally acquired responses to Plasmodium falciparum variant surface antigens on infected erythrocytes. Am J Trop Med Hyg 71:179–186
Schreiber N, Brattig N, Evans J et al (2006) Cerebral malaria is associated with IgG2 and IgG4 antibody responses to recombinant Plasmodium falciparum RIFIN antigen. Microbes Infect 8:1269–1276. doi:10.1016/j.micinf.2005.12.007
Schreiber N, Khattab A, Petter M et al (2008) Expression of Plasmodium falciparum 3D7 STEVOR proteins for evaluation of antibody responses following malaria infections in naive infants. Parasitology 135:155
Udomsangpetch R, Wahlin B, Carlson J et al (1989) Plasmodium falciparum-infected erythrocytes form spontaneous erythrocyte rosettes. J Exp Med 169:1835–1840
Udeinya IJ, Miller LH, McGregor IA, Jensen JB (1983) Plasmodium falciparum strain-specific antibody blocks binding of infected erythrocytes to amelanotic melanoma cells. Nature 303:429–431
Elliott SR, Brennan AK, Beeson JG et al (2005) Placental malaria induces variant-specific antibodies of the cytophilic subtypes immunoglobulin G1 (IgG1) and IgG3 that correlate with adhesion inhibitory activity. Infect Immun 73:5903–5907. doi:10.1128/IAI.73.9.5903-5907.2005
Ricke CH, Staalsoe T, Koram K et al (2000) Plasma antibodies from malaria-exposed pregnant women recognize variant surface antigens on Plasmodium falciparum-infected erythrocytes in a parity-dependent manner and block parasite adhesion to chondroitin sulfate A. J Immunol 165:3309–3316
Fried M, Nosten F, Brockman A et al (1998) Maternal antibodies block malaria. Nature 395:851–852. doi:10.1038/27570
Duffy PE, Fried M (2003) Antibodies that inhibit Plasmodium falciparum adhesion to chondroitin sulfate A are associated with increased birth weight and the gestational age of newborns. Infect Immun 71:6620–6623
Staalsoe T, Shulman CE, Bulmer JN et al (2004) Variant surface antigen-specific IgG and protection against clinical consequences of pregnancy-associated Plasmodium falciparum malaria. Lancet 363:283–289. doi:10.1016/S0140-6736(03)15386-X
Oleinikov AV, Amos E, Frye IT et al (2009) High throughput functional assays of the variant antigen PfEMP1 reveal a single domain in the 3D7 Plasmodium falciparum genome that binds ICAM1 with high affinity and is targeted by naturally acquired neutralizing antibodies. PLoS Pathog 5:e1000386. doi:10.1371/journal.ppat.1000386.g006
Ghumra A, Khunrae P, Ataíde R et al (2011) Immunisation with recombinant PfEMP1 domains elicits functional rosette-inhibiting and phagocytosis-inducing antibodies to Plasmodium falciparum. PLoS ONE 6:e16414. doi:10.1371/journal.pone.0016414
Celada A, Cruchaud A, Perrin LH (1982) Opsonic activity of human immune serum on in vitro phagocytosis of Plasmodium falciparum infected red blood cells by monocytes. Clin Exp Immunol 47:635–644
Bouharoun-Tayoun H, Attanath P, Sabchareon A et al (1990) Antibodies that protect humans against Plasmodium falciparum blood stages do not on their own inhibit parasite growth and invasion in vitro, but act in cooperation with monocytes. J Exp Med 172:1633–1641
Feng G, Aitken E, Yosaatmadja F et al (2009) Antibodies to variant surface antigens of Plasmodium falciparum-infected erythrocytes are associated with protection from treatment failure and the development of anemia in pregnancy. J Infect Dis 200:299–306. doi:10.1086/599841
Ataíde R, Hasang W, Wilson DW et al (2010) Using an improved phagocytosis assay to evaluate the effect of HIV on specific antibodies to pregnancy-associated malaria. PLoS ONE 5:e10807. doi:10.1371/journal.pone.0010807
Keen J, Serghides L, Ayi K et al (2007) HIV impairs opsonic phagocytic clearance of pregnancy-associated malaria parasites. Plos Med 4:e181. doi:10.1371/journal.pmed.0040181
Baruch DI, Gamain B, Barnwell JW et al (2002) Immunization of Aotus monkeys with a functional domain of the Plasmodium falciparum variant antigen induces protection against a lethal parasite line. Proc Natl Acad Sci USA 99:3860–3865. doi:10.1073/pnas.022018399
Baruch DI, Gamain B, Miller LH (2003) DNA immunization with the cysteine-rich interdomain region 1 of the Plasmodium falciparum variant antigen elicits limited cross-reactive antibody responses. Infect Immun 71:4536–4543
Gratepanche S, Gamain B, Smith JD et al (2003) Induction of crossreactive antibodies against the Plasmodium falciparum variant protein. Proc Natl Acad Sci USA 100:13007–13012. doi:10.1073/pnas.2235588100
Ahuja S, Pettersson F, Moll K et al (2006) Induction of cross-reactive immune responses to NTS-DBL-1alpha/x of PfEMP1 and in vivo protection on challenge with Plasmodium falciparum. Vaccine 24:6140–6154. doi:10.1016/j.vaccine.2006.05.030
Turner L, Wang CW, Lavstsen T et al (2011) Antibodies against PfEMP1, RIFIN, MSP3 and GLURP are acquired during controlled Plasmodium falciparum malaria infections in naïve volunteers. PLoS ONE 6:e29025. doi:10.1371/journal.pone.0029025
Flick K, Chen Q (2004) var genes, PfEMP1 and the human host. Mol Biochem Parasitol 134:3–9
Oguariri RM, Mattei D, Tena-Tomás C et al (2003) Recombinant Duffy binding- like-alpha domains of Plasmodium falciparum erythrocyte membrane protein 1 elicit antibodies in rats that recognise conserved epitopes. Parasitol Res 90:467–472. doi:10.1007/s00436-003-0884-8
Mayor A, Rovira-Vallbona E, Srivastava A et al (2009) Functional and immunological characterization of a Duffy binding-like alpha domain from Plasmodium falciparum erythrocyte membrane protein 1 that mediates rosetting. Infect Immun 77:3857–3863. doi:10.1128/IAI.00049-09
Chen Q, Pettersson F, Vogt AM et al (2004) Immunization with PfEMP1-DBL1alpha generates antibodies that disrupt rosettes and protect against the sequestration of Plasmodium falciparum-infected erythrocytes. Vaccine 22:2701–2712. doi:10.1016/j.vaccine.2004.02.015
Moll K, Pettersson F, Vogt AM et al (2007) Generation of cross-protective antibodies against Plasmodium falciparum sequestration by immunization with an erythrocyte membrane protein 1-duffy binding-like 1 alpha domain. Infect Immun 75:211–219. doi:10.1128/IAI.00749-06
Tutterrow YL, Salanti A, Avril M et al (2012) High avidity antibodies to full-length VAR2CSA correlate with absence of placental malaria. PLoS ONE 7:e40049. doi:10.1371/journal.pone.0040049
Tutterrow YL, Avril M, Singh K et al (2012) High levels of antibodies to multiple domains and strains of VAR2CSA correlate with the absence of placental malaria in Cameroonian women living in an area of high Plasmodium falciparum transmission. Infect Immun 80:1479–1490. doi:10.1128/IAI.00071-12
Bigey P, Gnidehou S, Doritchamou J et al (2011) The NTS-DBL2X region of VAR2CSA induces cross-reactive antibodies that inhibit adhesion of several Plasmodium falciparum isolates to chondroitin sulfate A. J Infect Dis 204:1125–1133. doi:10.1093/infdis/jir499
Nielsen MA, Pinto VV, Resende M et al (2009) Induction of adhesion-inhibitory antibodies against placental Plasmodium falciparum parasites by using single domains of VAR2CSA. Infect Immun 77:2482–2487. doi:10.1128/IAI.00159-09
Beeson JG, Chan J-A, Fowkes FJ (2013) PfEMP1 as a target of human immunity and a vaccine candidate against malaria. Expert Rev Vaccines 12:105–108. doi:10.1586/erv.12.144
Drew DR, Hodder AN, Wilson DW et al (2012) Defining the antigenic diversity of Plasmodium falciparum apical membrane antigen 1 and the requirements for a multi-allele vaccine against malaria. PLoS ONE 7:e51023. doi:10.1371/journal.pone.0051023
Miura K, Herrera R, Diouf A et al (2013) Overcoming allelic specificity by immunization with five allelic forms of Plasmodium falciparum apical membrane antigen 1. Infect Immun 81:1491–1501. doi:10.1128/IAI.01414-12
Kinyanjui SM, Bull P, Newbold CI, Marsh K (2003) Kinetics of antibody responses to Plasmodium falciparum-infected erythrocyte variant surface antigens. J Infect Dis 187:667–674. doi:10.1086/373994
Fowkes FJI, McGready R, Johnstone-Robertson S et al (2012) Antibody boosting and longevity following tetanus immunization during pregnancy. Clin Infect Dis. doi:10.1093/cid/cis979
Yone CLRP, Kremsner PG, Luty AJF (2005) Immunoglobulin G isotype responses to erythrocyte surface-expressed variant antigens of Plasmodium falciparum predict protection from malaria in African children. Infect Immun 73:2281–2287. doi:10.1128/IAI.73.4.2281-2287.2005
Mackintosh CL, Mwangi T, Kinyanjui SM et al (2008) Failure to respond to the surface of Plasmodium falciparum infected erythrocytes predicts susceptibility to clinical malaria amongst African children. Int J Parasitol 38:1445–1454. doi:10.1016/j.ijpara.2008.03.009
Magistrado PA, Lusingu J, Vestergaard LS et al (2007) Immunoglobulin G antibody reactivity to a group A Plasmodium falciparum erythrocyte membrane protein 1 and protection from P. falciparum malaria. Infect Immun 75:2415–2420. doi:10.1128/IAI.00951-06
Bull PC, Kortok M, Kai O et al (2000) Plasmodium falciparum-infected erythrocytes: agglutination by diverse Kenyan plasma is associated with severe disease and young host age. J Infect Dis 182:252–259. doi:10.1086/315652
Cabrera G, Yone C, Tebo AE et al (2004) Immunoglobulin G isotype responses to variant surface antigens of Plasmodium falciparum in healthy Gabonese adults and children during and after successive malaria attacks. Infect Immun 72:284–294
Giha HA, Staalsoe T, Dodoo D et al (1999) Overlapping antigenic repertoires of variant antigens expressed on the surface of erythrocytes infected by Plasmodium falciparum. Parasitology 119(Pt 1):7–17
Reeder JC, Rogerson SJ, al-Yaman F et al (1994) Diversity of agglutinating phenotype, cytoadherence, and rosette-forming characteristics of Plasmodium falciparum isolates from Papua New Guinean children. Am J Trop Med Hyg 51:45–55
Crompton PD, Kayala MA, Traoré B et al (2010) A prospective analysis of the Ab response to Plasmodium falciparum before and after a malaria season by protein microarray. Proc Natl Acad Sci 107:6958–6963. doi:10.1073/pnas.1001323107
Acknowledgments
Thank you to Christine Langer for providing the scanning electron microscopy image of an infected erythrocyte and to Campbell Aitken for helpful comments on the manuscript. The authors gratefully acknowledge funding from the National Health and Medical Research Council of Australia (Fellowship, programme grant and project grant to J. Beeson; infrastructure support scheme), the Australian Research Council (Future Fellowship to J. Beeson), the Australian Government (Australian Postgraduate Award to J. Chan) and the Victorian Operational Infrastructure Support Program.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article
Cite this article
Chan, JA., Fowkes, F.J.I. & Beeson, J.G. Surface antigens of Plasmodium falciparum-infected erythrocytes as immune targets and malaria vaccine candidates. Cell. Mol. Life Sci. 71, 3633–3657 (2014). https://doi.org/10.1007/s00018-014-1614-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00018-014-1614-3