
Abstract
The present study was aimed at investigating the coexistence and interactions between free living amoebae of Naegleria and Hartmannella genera and pathogenic Legionella pneumophila bacteria in thermal saline baths used in balneotherapy in central Poland. Water samples were collected from November 2010 to May 2011 at intervals longer than 1 month. The microorganisms were detected with the use of a very sensitive fluorescence in situ hybridisation method. In addition, the morphology of the amoebae was studied. Despite relatively high salinity level, ranging from 1.5 to 5.0 %, L. pneumophila were found in all investigated baths, although their number never exceeded 106 cells dm−3. Hartmannella were not detected, while Naegleria fowleri were found in one bath. The observation that N. fowleri and L. pneumophila may coexist in thermal saline baths is the first observation emphasising potential threat from these microorganisms in balneotherapy.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
First identified in 1976 (Saint and Ho 1999; Huang et al. 2011), Legionella sp. bacteria are one of the main groups of pathogenic bacteria transmitted via water (Papciak and Zamorska 2005) and belong to the gamma Proteobacteria (Heuner and Steinert 2003). Approximately half of 48 species of Legionella cause a disease called legionellosis (Legionnaires’ disease), most notably Legionella pneumophila, responsible for more than 80 % of all known cases (Yaňez et al. 2005). Human infection by L. pneumophila usually results from inhaling aerosol droplets of water, which contain bacterial cells (Turetgen et al. 2005).
Free living amoebae (FLA), including Naegleria fowleri, Acanthamoeba sp., Balamuthia mandrillaris, Sappinia sp. and Hartmannella sp., responsible for dangerous infections in humans and animals, enter the human body in a similar way (Martinez 1996; Schuster and Visvesvara 2004; Dykova and Lom 2004; Daft et al. 2005; Karanis et al. 2007; Visvesvara et al. 2005, 2007). N. fowleri causes primary amoebic meningoencephalitis, a disease of the central nervous system, resulting in death of the infected people (Song et al. 2008; Edagawa et al. 2009; Jamerson et al. 2009).
FLA, also referred to as deadly amoebae or “brain-eating” amoebae inhabit natural and anthropogenic aquatic environments (Behets et al. 2007; Schuster and Visvesvara 2004; Sheehan et al. 2003a; De Jonckheere 2002; Tyndall et al. 1989). The relationship between pathogenic species of Legionella and FLA has a unique character. Legionella sp. are parasites of Naegleria and Hartmannella amoebae, within which they multiply, acquiring new ways of spreading in the environment (Fields et al. 2002; Molmeret et al. 2008; Fields 1996; Abu Kwaik 1998; Heuner and Steinert, 2003; Ettinger et al. 2003; Huang et al. 2011). The above-mentioned ubiquitous species of FLA are frequently found in ecosystems contaminated with Legionella (Grimm et al. 2001; Greub and Raoult 2004; Suzan-Monti et al. 2006), providing food and shelter for the pathogenic bacteria. Owing to the fact that a range of other pathogenic organisms including Mycobacterium avium, viruses and fungi may also develop inside FLA, in scientific literature, they are referred to as “Trojan horses” (Edgawa et al. 2009) and represent a potential source of pathogens dangerous for humans (Horn and Wagner 2004).
Successful coexistence of Legionella and FLA is based mainly on their shared ability to grow in biofilms which form at the solid–liquid interfaces or at the liquid–air interface (Flemming et al. 2000; Huws et al. 2005; Hoffman and Michel 2001) as well as on their tolerance to elevated temperatures. L. pneumophila can survive in water at temperatures ranging from 0 to 70 °C, and their optimum temperature (32–35 °C) (Declerck et al. 2007) partially overlaps with the temperature conditions preferred by Naegleria and Hartmannella amoebae (30–42 °C) (Jamerson et al. 2009; Mazur 1984; Lorenzo-Morales et al. 2007; Pelandakis et al. 2000). Both the pathogenic bacteria and their host amoebae, which display tolerance to elevated temperatures, find favourable conditions for growth not only in water bodies in regions characterised by hot climate but also in heated water of baths, pools and other facilities used in recreation and balneotherapy. Taking into consideration the high pathogenicity of Legionella and several FLA amoebae, the study focused on establishing whether L. pneumophila can coexist with Naegleria and Hartmannella amoebae in thermal saline baths used in balneotherapy.
Materials and methods
The object of the investigation
Water samples were collected from thermal saline baths, supplied with thermal saline waters (type Cl–Na), containing mainly iodides and iron, components with pharmacodynamic properties (Table 1). Found at great depths (700–1,700 m), they are well isolated from surface waters and appear to contain almost no organic compounds. The temperature of the water in the intake ranges from 32 to 40 °C. There is a continuous water flow from the intake into the pipes.
Sampling
Water samples were collected from November 2010 to May 2011 (five sampling cycles) from three thermal baths: bath 1, water salinity 5 %; bath 2, water salinity 4 %; bath 3, water salinity 1.5 %. Baths 1 and 2 are used for balneotherapy; bath 3 is used for recreation only.
Each sampling operation involved collecting one litre of “open water” taken from the bath into sterile glass bottles from each bath and measuring the following physicochemical parameters of water: its temperature, redox potential, pH value (with the Elmetron pH meter) and oxygen saturation (with the Hanna Instruments oximetre). The samples were then transported to the laboratory in 7 °C.
Fluorescence in situ hybridisation method
The numbers of bacteria belonging to different phylogenetic groups (Eubacteria, Legionella sp. and L. pneumophila) and the investigated eukaryotic organisms (Eukariota, Naegleria sp., N. fowleri and Hartmannella sp.) were determined with the use of a molecular fluorescence in situ hybridisation (FISH) method. After the end of water uptake, the water samples were fixed with formamide. Later, the water samples were filtered through polycarbonate membrane filters with a 0.22-μm pore size in order to capture particles bigger than 0.22 μm. The hybridisation was performed according to Grimm et al. 2001. Bacterial cells retained on the surface of the membrane were hybridised using Legionella-specific fluorescence-labelled (with dye CY3) oligonucleotide probes EUB338, LEG705, LEGPNE1 and fluorescein-labelled, eukaryote-specific probes EUK516, HART498, NAEG1088 and NAE1041.
The probes were suspended in the hybridisation buffer consisting of formamide [whose concentration depended on the probe sequence (in volumes per volume)], NaCl 0.9 M, sodium dodecyl sulphate 0.01 % and Tris/HCl (20 mM), pH 7.6. This solution was applied to the surface of the filter with captured cells.
For probes LEG705, LEGPNE1 and EUK516, formamide concentration was 25 %; for probes NAEG1088 and NAE1041, formamide concentration was 30 %; for probe EUB338, formamide concentration was 35 %, and for probe HART498, formamide concentration was 40 %. The probe concentration in the hybridisation buffer was 30 ng of probes for prokaryotic organisms and 150 ng of probes for eukaryotic organisms.
The filters were then placed for 2 h in a hybridisation chamber and in ultrathermostat at 46 °C. After that, in order to remove the unbound probes, the filters were placed for 15 min in the washing buffer having the temperature of 48 °C (the composition of the buffer, 20 mM Tris/HCl, pH 7.6; 0.01 % sodium dodecyl sulphate; 5 mM ethylene diamine triacetic acid (EDTA)-160 mM NaCl for probes LEG705, LEGPNE1 and EUK516; 5 mM EDTA; 56 mM NaCl for probe HART498; 5 mM EDTA; 112 mM NaCl for probes NAEG1088 and NAE1041; 80 mM NaCl for probe EUB338), rinsed with the distilled water and dried.
Subsequently, the filters were covered with the mixture of immersion oil and Citifluor AF2 (Citifluor Ltd., London, UK). Fluorescence was detected using Olympus BX50 microscope equipped for epifluorescence microscopy with a 50-W mercury high-pressure bulb and the appropriate filter set 00 and 10. The slides with hybridised prokaryotic cells were analysed under the lens at × 100 magnification, while the slides with eukaryotic cells were analysed at × 10 and 40 magnifications. Colour micrographs were taken with digital image processing (Olympus XC50) using the software package (CellB v. 3.1.). The number of bacteria in the investigated slides was evaluated using MultiScan-Based programme.
The images of hybridised amoebae were additionally evaluated for morphological features; identification of the size and morphological features typical of amoebae of Naegleria and Hartmannella genera was particularly significant. Naegleria sp. are quite uniform in shape—a cell reaches 10–15 μm. The cytoplasm is slightly granular and has a clearly visible bright halo with a dense nucleus. Numerous vacuoles are usually seen in the cytoplasm. Trophozoites move by extending and contracting their rounded pseudopodium (lobopodium), which are bright on the edges and filled with granular cytoplasm. The posterior end of the cell, a hyaline uroid, has many small pseudopodia.
The trophozoites of Hartmannella reach the size of 25–40 μm, have an elongated shape and produce monopodial lobopodia. In the cytoplasm of these amoebae, there are numerous bright areas corresponding to vacuolar vesicles. Due to the morphological similarity of pathogenic species of Naegleria and Hartmannella to non-pathogenic species as well as to small but real risk of non-specific binding of the probes to the particles of organic matter after hybridisation, only the combination of the two methods, i.e. molecular (FISH) and morphological, provides an accurate evaluation of the studied FLA. The number of amoebae detected and identified by fluorescence in situ hybridisation and confirmed in the morphological analysis was recalculated per 1 dm3 of the sampled water with the use of the above-mentioned formula.
Results
The temperature in the investigated baths ranged from 30 to 36.5 °C (Table 2), providing optimum conditions for the growth of both L. pneumophila and Naegleria and Hartmannella amoebae. Slight fluctuations of pH values did not affect the growth of the studied microorganisms. However, oxygen saturation varied significantly; the biggest differences between consecutive measurements being recorded in thermal bath 1 (range 19.8–138 %).
The highest number of Eubacteria was recorded in bath 1. The average number in the entire research cycle for this bath was 98 × 106 dm−3 cells. The lowest number of Eubacteria was recorded in bath 3 (Fig. 1). The average number in the entire research cycle for this bath was 39.5 × 106 dm−3 cells.

Number of Eubacteria in the investigated baths. Vertical bars represent standard deviation
Statistical analysis revealed no significant relationships between physicochemical parameters of water in the investigated baths and the number of Eubacteria (P > 0.05). In addition, no significant changes in the number of bacteria of the investigated group were recorded in particular baths in consecutive samples. Differences in the number of Eubacteria recorded in different baths, although substantial, were not significant statistically either (P > 0.05). The figures reveal positive correlation (r = 0.94) between the number of bacteria and the age of the bath.
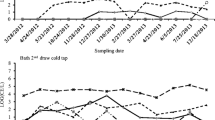
Bacteria belonging to the Legionella sp. and L. pneumophila were identified in all water samples collected from each bath (Fig. 2). Moreover, in the investigated microscopic slides, L. pneumophila were also detected within the biofilm and microcosm (Fig. 3).

Number of Legionella sp. and L. pneumophila in the investigated baths. Vertical bars represent standard deviation

L. pneumophila bacteria in the fragment of biofilm in investigated sample water from bath 1, bar = 20 μm
The highest number of bacteria belonging to the Legionella genus was recorded in bath 2 (12.21 × 106 dm−3 on average). Furthermore, L. pneumophila were also the most abundant in bath 2, where their average number was 2.84 × 106 dm−3 on average (Fig. 2).
The lowest number of Legionella sp. (1.98 × 106 dm−3 on average) was observed in bath 3. L. pneumophila were also the least abundant in bath 3, where their average number was 0.40 × 106 dm−3. In bath 1, the average number of bacteria of the Legionella genus was 5.71 × 106 dm−3, while the average number of L. pneumophila was 2.84 × 106 dm−3.
Amoebae of the Hartmannella genus were not identified in any of the water samples taken from all baths. However, limax amoeba of N. fowleri sp. was identified in one sample taken from bath 1, based on the results of the FISH method (Fig. 4). Morphological features of the hybridised amoeba, including its length (15 μm), monopodial lobopodium, a hyaline uroid and a large kariosom in the nucleus, confirm its affiliation to N. fowleri.

Positive result of FISH hybridisation for N. fowleri, bar = 200 μm. Amoeba is marked with an arrow: l lobopodium, n nucleus, u uroid. Enlargement in a frame indicates sample 1 from bath 1
Discussion
In the hitherto published research papers, hardly any information is available on the distribution of Legionella sp. and FLA in thermal saline waters applied in balneotherapy. A majority of studies are aimed at identifying Legionellaceae in hot springs and in recreational facilities connected with the springs as well as in drinking and industrial water supply systems (water cooling systems, hydro-electric power stations). The results obtained during this investigation can be therefore regarded as original and innovative.
Taking into consideration the substantial salinity of the investigated thermal saline baths, the number of L. pneumophila seems to be large. However, Barbaree et al. (1983) observed that L. pneumophila serogroups 4 and 5 display the highest resistance to chloride sodium salt of all bacteria belonging to the genus Legionella sp. Besides, Palmer et al. (1993) confirmed the presence of Legionella sp. in 50 % of the collected samples of ocean water and the presence of L. pneumophila in 75 % of the samples.
Although the data suggest a fairly high tolerance of this kind of bacteria to high concentration of NaCl, it should be noted that all attempts to culture Legionella sp. from ocean water proved unsuccessful, which may indicate that only non-culturable cells of these microorganisms are present in saline waters. What is more, Legionella growth is the most rapid within the existing biofilm (Declerck 2010). Since biofilm is known to possess some properties protecting bacterial cells found deep within this formation, professional antiseptics have little or no effect on them. It may be therefore assumed that biofilm effectively protects Legionella against salt.
In this study, the number of Legionella fell within the range 103 to 108 CFU/dm3, which corresponds to the range observed in the natural environment by Stypułkowska-Misiurkiewicz et al. (2001). The results of the investigations conducted in hospitals and spas in Poland (including those focused on examining water from thermal saline baths) show that culturable forms of L. pneumophila were found in 78.7 % of the samples (Matuszewska and Krogulska 2009). In addition, in a majority of positive tests (71.7 % of the samples), the numbers of L. pneumophila were similar to the results obtained in this investigation and ranged from 1.2 × 102 to 1.3 × 105 cfu/100 ml.
Many researchers who applied a real-time PCR method to detect Legionella sp. and L. pneumophila emphasised the fact that these microorganisms are commonly found in anthropogenic water bodies. In their investigations, the presence of Legionella sp. and L. pneumophila was confirmed in all water samples collected from anthropogenic water bodies (Declerck et al. 2007). Huang et al. (2010) also found Legionella sp. in all recreational areas and facilities connected with hot springs in Taiwan. Twenty samples out of 72 collected in the research cycle contained Legionella sp. Considering the prevalence of these bacteria in the environments surrounding humans, a question should be answered whether the epidemics caused by L. pneumophila is a real threat.
According to Exner and Hartemann (2009), an acceptable number of L. pneumophila fall within the range 105–106 cfu/dm3, which may indicate that waters investigated in this study were relatively safe for humans. It must be stressed, however, that there were cases of legionellosis caused by Legionella in water systems when their number was just above 104 cfu/dm3 (Meenhorst et al. 1985; Patterson et al. 1994).
In this study, water samples were also analysed for the presence of amoebae of Naegleria spp. and Hartmannella spp. N. fowleri was identified in one sample (taken from bath 1) (photo 8). Physicochemical and biotic conditions in the investigated baths deviate substantially from the conditions prevailing in the environments inhabited by N. fowleri and may therefore differently (contrarily) affect the growth of these amoebae (Sheehan et al. 2003b; Init et al. 2010). Amoebae are frequently found in fresh water, even strongly acidic or chlorinated (Rivera et al. 1983).
Osmotic pressure of the environment is one of the factors determining the differentiation of Naegleria amoebae into non-pathogenic flagellated forms (Jeffery and Hawkins 1976; Fulton 2004). Although high temperature, high oxygen saturation (Table 1) and a high number of bacteria (Fig. 1) in the investigated baths constitute favourable conditions for the development of amoebae, high salinity may be a limiting factor, preventing amoebae from rapid spread. Nevertheless, contrary to repeated published information suggesting the absence of pathogenic Naegleria in the marine environment, our observations agree with the observations of Culbertson (1970) and Ettinger et al. (2003), indicating that Naegleria can actually be found in salt water. The authors identified single cells of Naegleria sp. in natural low-salinity water. Water salinity limits the growth of amoebae but does not eliminate them completely; in the Black Sea, amoebae were identified in 30 % of the analysed water samples (Tsvetkova et al. 2004).
The data presented in this study show that the spread of pathogenic amoebae is determined by the combination of several factors. In addition, the data seem to suggest that potentially dangerous FLA and L. pneumophila may coexist in thermal saline baths used in balneotherapy. This fact should be taken into account while planning and introducing actions aimed at eradicating both groups of these potentially dangerous microorganisms.
References
Abu Kwaik Y (1998) Fatal attraction of mammalian cells to Legionella pneumophila. Mol Microbiol 4:689–695
Barbaree JM, Sanchez A, Sanden GN (1983) Tolerance of Legionella species to sodium chloride. Curr Microbiol 9:1–5
Behets J, Declerck P, Delaedt Y, Verelst L, Ollevier F (2007) Survey for the presence of specific free-living amoebae in cooling waters from Belgian power plants. Parasitol Res 100:1249–1256
Culbertson CG (1970) Pathogenic Naegleria and Hartmanella (Acanthamoeba). Ann N Y Acad Sci 74:1018–1022
Daft BM, Visvesvara GS, Read DH, Kinde H, Uzal FA, Manzer MD (2005) Seasonal meningoencephalitis in Holstein cattle caused by Naegleria fowleri. J Vet Diagn Invest 17:605–609
De Jonckheere JF (2002) A century of research on the amoeboflagellate genus Naegleria. Acta Protozool 41:309–342
Declerck P (2010) Biofilms: the environmental playground of Legionella pneumophila. Environ Microbiol 12:557–566
Declerck P, Behets J, Hoef V, Ollevier F (2007) Detection of Legionella spp. and some of their amoeba hosts in floating biofilms from anthropogenic and natural aquatic environments. Water Res 41:3159–3167
Dykova I, Lom J (2004) Advances in the knowledge of amphizoic amoebae infecting fish. Folia Parasitol 51:81–97
Edagawa A, Kimura A, Kawabuchi-Kurata T, Kusuhara Y, Karanis P (2009) Isolation and genotyping of potentially pathogenic Acanthamoeba and Naegleria species from tap-water sources in Osaka, Japan. Parasitol Res 105:1109–1117
Ettinger MR, Webb SR, Harris SA, Macininch SP, Garman GC, Brown BL (2003) Distribution of free-living amoebae in James River, Virginia, USA. Parasitol Res 89:6–15
Exner M, Hartemann P (2009) Summary of the second meeting of the International Forum on Water Hygiene in Buildings (IFOWAHB) from 01 to 02.06.2007 in Stockholm, Meeting Report. Int J Hyg Envir Heal 212:449–458
Fields BS (1996) The molecular ecology of legionellae. Trends Microbiol 4:286–290
Fields BS, Benson RF, Besser RE (2002) Legionella and Legionnaires’ disease: 25 years of investigation. Clin Microbiol Rev 3:506–526
Flemming HC, Wingeneder J, Mayer C, Korstgens V, Borchard W (2000) Cohesiveness in biofilm matrix polymers. In: Allison DG, Lapin-Scott HM, Wilson M (eds) Society for General Microbiology Symposium: community structure and co-operation in biofilms, vol 59. Cambridge University Press, Cambridge, pp 88–106
Fulton C (2004) Early events of cell differentiation in Naegleria gruberi. Synergistic control by electrolytes and a factor from yeast extract. Dev Biol 28:603–619
Greub G, Raoult D (2004) Microorganisms resistant to free-living amoebae. Clin Microbiol Rev 17:413–433
Grimm D, Ludwig W, Brandt BC, Michel R, Schleifer KH, Hacker J, Steinert M (2001) Development of 18S rRNA-targeted oligonucleotide probes for specific detection of Hartmannella and Naegleria in Legionella—positive environmental samples. Syst Appl Microbiol 24:6–82
Heuner K, Steinert M (2003) The flagellum of Legionella pneumophila and its link to the expression of the virulent phenotype. Int J Med Microbiol 293:133–143
Hoffman R, Michel R (2001) Distribution of free-living amoebae (FLA) during preparation and supply of drinking water. Int J Hyg Environ Health 203:215–219
Horn M, Wagner M (2004) Bacterial endosymbionts of free-living amoebae. J Eucaryot Microbiol 51:509–514
Huang SW, Hsu BM, Wu SF, Fan CW, Shih FC, Lin YC, Ji DD (2010) Water quality parameters associated with prevalence of Legionella in hot spring facility water bodies. Water Res 44:4805–4811
Huang L, Boyd D, Amyot W, Hempstead AD, Luo Z, O’Connor TJ, Chen C, Machner M, Montminy T, Isberg RR (2011) The E Block motif is associated with Legionella pneumophila translocated substrates. Cell Microbiol 13:227–245
Huws SA, Mcbain AJ, Gilbert P (2005) Protozoan grazing and its impact upon population dynamics in biofilm communities. J Appl Microbiol 98:238–244
Init I, Lau YL, Arin Fadzlun A, Foead AI, Neilson RS, Nissapatorn V (2010) Detection of free living amoebae, Acanthamoeba and Naegleria, in swimming pools, Malaysia. Trop Biomed 27:566–577
Jamerson M, Remmers K, Cabral G, Marciano-Cabral F (2009) Survey of Naegleria fowleri amebae in lake water used to cool reactors at a nuclear power generating plant. Parasitol Res 104:969–978
Jeffery S, Hawkins SE (1976) Studies on transformation in Naegleria gruberi: effects of ions and bacterial suspensions. Microbios 15:27–36
Karanis P, Kourenti C, Smith H (2007) Waterborne transmission of protozoan parasites: a worldwide review of outbreaks and lessons learnt. J Water Health 5:1–38
Lorenzo-Morales J, Martínez-Carretero E, Batista N, Álvarez-Marín J, Bahaya Y, Walochnik J, Valladares B (2007) Early diagnosis of amoebic keratitis due to a mixed infection with Acanthamoeba and Hartmannella. Parasitol Res 102:167–169
Martinez AJ (1996) Free-living amebas: Naegleria, Acanthamoeba and Balamuthia. Medical Microbiology 4th ed. Galveston (TX): University of Texas Medical Branch at Galveston. http://www.ncbi.nlm.nih.gov/books/NBK7960/
Matuszewska R, Krogulska B (2009) Problemy występowania pałeczek Legionella w instalacjach i urządzeniach wytwarzających aerozol wodno powietrzny w obiektach służby zdrowia w Polsce. Nowa Medycyna 1:56–60
Mazur T (1984) Występowanie Naegleria fowleri w środowisku wolnym i właściwości biologiczne izolowanych szczepów. Wiad Parazytol 30:3–35
Meenhorst PL, Reingold AL, Groothuis DG, Gorman GW, Wilkinson HW, McKinney RM, Feeley JC, Brenner DJ, Van Furth R (1985) Water-related nosocomial pneumonia caused by Legionella pneumophila serogroups 1 and 10. J Infect Dis 152:356–364
Molmeret M, Santic M, Abu Kwaik Y (2008) Interaction of Legionella pneumophila with amoeba. In: Hoffman P, Friedman H, Bendineeli M (eds) Legionella pneumophila: pathogenesis and immunity. Springer, New York, pp 185–196
Palmer CJ, Tsai Y, Paszko-Kolva C, Mayer C, Sangermano LR (1993) Detection of Legionella species in sewage and ocean water by polymerase chain reaction, direct fluorescent-antibody, and plate culture methods. App Env Microbiol 11:3618–3624
Papciak D, Zamorska J (2005) Podstawy biologii i biotechnologii środowiskowej. Oficyna Wydawnicza Politechniki Rzeszowskiej, Rzeszow
Patterson WJ, Seal DV, Curran E, Sinclair TM, McLuckie JC (1994) Fatal nosocomial Legionnaires’ disease: relevance of contamination of hospital water supply by temperature-dependent buoyancy-driven flow from spur pipes. Epidemiol Infect 112:513–525
Pelandakis M, Serre S, Pernin P (2000) Analysis of the 5.8S rRNA gene and the internal transcribed spaces in Naegleria spp. and in N. fowleri. J Eucaryot Microbiol 47:116–121
Rivera F, Ramirez P, Vilaclara G, Robles E, Medina F (1983) A survey of pathogenic and free-living amoebae inhabiting swimming pool water in Mexico City. Env Res 32:205–211
Saint CP, Ho L (1999) A PCR test for identification and discrimination of Legionella longbeachae serogroups 1 and 2. J Microbiol Meth 37:245–253
Schuster FL, Visvesvara GS (2004) Free-living amoebae as opportunistic and non-opportunistic pathogens of humans and animals. Int J Parasitol 34:1001–1027
Sheehan KB, Fagg JA, Henson JM (2003a) PCR detection and analysis of the free-living amoeba Naegleria in hot springs in Yellowstone and Grand Teton National Parks. Appl Env Microbiol 69:5914–5918
Sheehan KB, Ferris MJ, Henson JM (2003b) Detection of Naegleria sp. in a thermal, acidic stream in Yellowstone National Park. J Eucaryot Microbiol 50:263–265
Song KJ, Song K, Kim JH, Sohn HJ, Lee YJ, Park CE, Shin HJ (2008) Heat shock protein 70 of Naegleria fowleri is important factor for proliferation and in vitro cytotoxicity. Parasitol Res 103:313–317
Stypułkowska-Misiurkiewicz H, Krogulska B, Pancer K, Matuszewska R (2001) Metodyka wykrywania i oznaczania bakterii z rodzaju Legionella w środowisku wodnym i w materiale klinicznym. Wydawnictwo Medyczne PZH Warszawa 1–47
Suzan-Monti M, La Scola B, Raoult D (2006) Genomic and evolutionary aspects of Mimivirus. Virus Res 117:145–155
Tsvetkova N, Schild M, Panaiotov S, Kurdova-Mintcheva R, Gottstein B, Walochnik J, Aspock H, Lucas MS, Muller N (2004) The identification of free-living environmental isolates of amoebae from Bulgaria. Parasitol Res 92:405–413
Turetgen I, Sungur EI, Cotuk A (2005) Enumeration of Legionella pneumophila in cooling water tower systems. Env Mon Ass 1–3:53–58
Tyndall RL, Ironside KS, Metler PL, Tan EL, Hazen TC, Fliermans CB (1989) Effect of thermal additions on the density and distribution of thermophilic amoebae and pathogenic Naegleria fowleri in a newly created cooling lake. Appl Environ Microbiol 55:722–732
Visvesvara GS, De Jonckheere JF, Arciano-Cabral F, Schuster FL (2005) Morphologic and molecular identification of Naegleria dunnebackei sp. isolated from a water sample. J Eukaryot Microbiol 6:523–531
Visvesvara GS, Moura H, Schuste FL (2007) Pathogenic and opportunistic free-living amoebae: Acanthamoeba spp., Balamuthia mandrillaris, Naegleria fowleri, and Sappinia diploidea. Immunol Med Microbiol 1:1–26
Yaňez M, Carrasco-Serrano AC, Barbera V, Catalan V (2005) Quantitative detection of Legionella pneumophila in water samples by immunomagnetic purification and real-time PCR amplification of the dotA gene. Appl Environ Microbiol 71:3433–3441
Acknowledgments
Experiments comply with the current laws in Poland. This research has been carried out with thanks to the State Committee for Scientific Research, grant no. 3878/B/P01/2010/38.
Open Access
This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 2.0 International License (https://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Żbikowska, E., Walczak, M. & Krawiec, A. Distribution of Legionella pneumophila bacteria and Naegleria and Hartmannella amoebae in thermal saline baths used in balneotherapy. Parasitol Res 112, 77–83 (2013). https://doi.org/10.1007/s00436-012-3106-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00436-012-3106-4