
Abstract
Under arid and semi-arid climates, adopting the appropriate tools for alleviating water deficit impacts is a critical factor that affects the physiological characteristics and yield of sunflower. Therefore, in order to find promising field practices in sunflower cultivation, the strip plots design in randomized complete block arrangement was used to examine the effects of two irrigation regimes as 100% (FI) and 85% (DI) of crop evapotranspiration and five antioxidant treatments on physiological and agronomic traits, yield response factor, and irrigation water use efficiency (IWUE) of sunflower. The antioxidant treatments involved two rates of ascorbic acid (150 and 300 mg L−1) and two rates of citric acid (250 and 500 mg L−1), in addition to the check treatment (tap water). The study was conducted for two growing seasons of 2019 and 2020 at the Experimental Farm of Ain Shams University, Egypt, located in a semi-arid environment. Findings showed that exogenous application of higher rate of ascorbic acid, i.e. 300 mg L−1 with FI exhibited the highest increase of chlorophyll a, chlorophyll b and the lowest proline content compared to other interaction treatments. Seed yield was significantly higher with FI plus ascorbic acid 300 mg L−1 and DI plus ascorbic acid 300 mg L−1 treatments than with their counterpart check treatment in both growing seasons. Under DI, IWUE was improved with antioxidant-treated plants compared to untreated plants. Yield response factor as an indicator of crop tolerance to drought was higher than the unit (> 1) under all ascorbic acid and citric acid levels. It could be concluded that ascorbic acid and citric acids partially mitigated the reductions in growth and yield caused by low water supply. However, yield response factor demonstrated that the crop is still sensitive to drought. Thus, other applicable patterns should be adopted to increase the yield potential of sunflower for counteracting the adverse impacts of drought.
Similar content being viewed by others
Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
The predicted global warming and climate changes indubitably has adverse effects such as drought, salinity and nutrient deficiency on growth and productivity of economic crops (Saudy and Mubarak 2015; Abd El-Mageed et al. 2022). Such effects are most pronounced in arid and semi-arid regions, which suffer from water scarcity (Souri and Hatamian 2019). Accordingly, irrigation strategies that ensure the efficient and rational use of water and nutrients must be adopted (Saudy and El-Bagoury 2014; Saudy and El-Metwally 2019, 2022; Saudy et al. 2020a). In this respect, deficit irrigation strategy could save water and increase water use efficiency (WUE), resolving the contradiction of water needs and supply in semi-arid regions (Cui et al. 2009). Moreover, several studies proved that deficit irrigation had significant effects on water saving and increase in WUE, clarifying the priority of deficit water tactic in crop irrigation programs under water limitation in arid environment as in Egypt (El-Bially et al. 2018; El-Metwally and Saudy 2021a; El-Metwally et al. 2022a; Saudy et al. 2022b). However, using water less than normal represents abiotic stress (drought) on crop plants. Decrease in leaf water potential, promoting stomatal closure, decrease in leaf photosynthesis, and weakness in nutrient uptake are the most physiological phenomena associated with water deficit (Bresson et al. 2015; Yan et al. 2016; El-Metwally et al. 2021; Abd-Elrahman et al. 2022; Makhlouf et al. 2022). Decrease in sunflower yield was reported by Saudy et al. (2021a); El-Bially et al. (18,19,a, b) as 22.4–42.8% owing to water deficit stress.
Sunflower (Helianthus annuus L.) is one of the main oil crops in the world. Among oilseed crops, sunflower is the fifth most cultivated annual crop (FAO 2017). Sunflower seed has high oil (36–52%) and protein (28–32%) contents (Rosa et al. 2009) and extracted oil has low cholesterol and high unsaturated fatty acids (Qahar et al. 2010). Moreover, sunflower can adapt to different climatic and soil conditions (Kaleem et al. 2011).
Therefore, in particularly economically valuable plants, many approaches have been employed to induce stress tolerance with enhancing the crop growth and development (Saudy et al. 2018, 2021b). Ascorbic acid (ASC) is a plant nonenzymatic antioxidant and mediates biotic and abiotic stress, since it is the first line of plant defense against oxidative stress (Sharma et al. 2019). Since ASC is mostly a substrate of ascorbate peroxidase, an essential enzyme of the ascorbate-glutathione pathway, it acts as a protector against the oxidative stress by removing several free radicals (Bilska et al. 2019; Sharma et al. 2019).
Citric acid (CA) is a weak organic acid found in many fruits and is used as preservative agent in human food being an antioxidant. Inside the plant cell, citric acid plays an important role in the intermediary metabolism, being a component of the tricarboxylic acid cycle or Krebs cycle (Omar et al. 2018). In some plant species as cotton (Gebaly et al. 2013) and bean (El-Tohamy et al. 2013), CA ameliorated the adverse effect of drought.
The current study hypothysized that sunflower plants have different significant response to both ASC and CA under drought conditions. Therefore, the main objective of this article is to evaluate the influence of different levels of ascorbic and citric acids on physiological and agronomic traits, yield response factor, and irrigation water use efficiency of sunflower under mild water stress compared to full irrigation.
Method and Materials
Experimental Site
The current work was conducted for two years during two growing summer seasons of 2019 and 2020, at the Experimental Farm, Faculty of Agriculture, Ain Shams University, El Nubaria region, El Behaira Governorate, Egypt (30° 30′N, 30° 20′E). According to the aridity categorization (Ponce et al. 2000), the experimental site was in a semi-arid environment with no rainfall in summer (beginning of April to late October). The averages of minimum air temperature were 21.2–20.3 oC, maximum air temperature were 33.2–32.4 oC, relative humidity were 54.3–55.8%, wind speed were 3.4–3.5 m sec−1, solar radiation were 28.3–28.6 MJ m−2 day−1, and mean class “A” pan evaporation was 3.90 and 3.92 mm d−1 for 2019 and 2020 seasons, respectively. The soil of the experimental site is classified as sandy-loam. In the root zone, soil water contents at the field capacity and permanent wilting point were 12.3–12.6% and 5.4–4.9% in both seasons, respectively. The properties of soil in the experimental site are shown in Table 1.
Experimentation and Procedures
This experiment was implemented in a strip-plots in randomized complete blocks design with three replicates. In the vertical plots, two levels of irrigation as 85 and 100% of full irrigation representing drought (deficit irrigation, DI) and well-watered (full irrigation, FI) treatments, respectively, were applied. The experimental unit had an area of 14 m2 (4 m × 3.5 m) with five ridges. The distance between plots was 1.0 m to prevent the overlapping of irrigation water of the nearby treatments. Sunflower seeds (cv. Sakha 53) were planted by hand in hills at 20-cm distance on the ridge at the depth of 3 cm on the 19th and 21st of May in 2019 and 2020, respectively. At 15 days after sowing (DAS), plants were thinned to secure one plant per hill. The recommended doses of mineral fertilizers were applied as follows: 21.2 kg ha−1 P was applied during the soil preparation as calcium super phosphate 15.5% P2O5, while N (107.1 kg ha−1) was added as ammonium nitrate (33.5% N) into five equal doses, @ 10, 20, 30, 40 and 50 DAS. Additionally, K (47.4 kg ha−1) was applied as potassium sulfate (48% K2O) in two equal doses at sowing and 50 DAS. Plants were irrigated through drip irrigation system using drippers of 2 L h−1 capacity.
Five treatments of antioxidant solutions occupied the horizontal plots involving two rates of ascorbic acid, ASC (150 and 300 mg L−1 namely: ASC150 and ASC 300, respectively) and two rates of citric acid, CA (250 and 500 mg L−1 namely: CA250 and CA500, respectively), in addition to the check treatment (tap water). The spray solutions were applied two twice 35 and 50 DAS, syncorinizing the plant heights of 40–50 and 90–100 cm, respectively, using a knapsack sprayer having a single nozzle with volume of 500 L ha−1.
Irrigation Requirements
Reference evapotranspiration (ETo; mm day−1) was calculated from the recorded meteorological data of the study area using the FAO-56 Penman-Monteith equation (Allen et al. 1998).
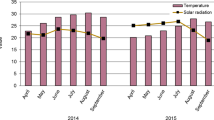
The obtained reference evapotranspiration (ETo) values during sunflower growth stages are shown in Fig. 1. Moreover, irrigation water requirement for sunflower was calculated by determining the daily crop evapotranspiration (ETc). The depth of applied irrigation water was calculated as described by Vermeirer and Jopling (1984).

Changes in reference evaporation (ETo) during 2019 and 2020 growing seasons of sunflower at El Nubaria region, Egypt
The same amount of water was applied to all treatments until 30 DAS, the time representing elongation stage, the deficit irrigation treatment (DI) was started under each antioxidant treatment. Deficit irrigation continued till maturity stage (harvesting), therefore plants exposed to drought for about 53 and 57 days along the life cycle in the first and second seasons, respectively. Irrigation was applied at 2‑day interval. Moreover, sunflower plants received irrigation water amounts of 4736.0 and 4650.8 m3 ha−1 in 2019 season as well as 5312.0 and 5226.2 m3 ha−1 in 2020 season, with irrigation by DI and FI, respectively.
Sampling and Assessments
Physiological Traits
The 4th leaves from the top were taken at 65 DAS and used for measuring both chlorophyll a and b (mg g−1 fresh wt.) according to Wettstein (1957), and free proline content (µg g−1 fresh wt.) according to Bates et al. (1973).
Growth Traits
At 65 DAS, five guarded plants were chosen randomly from each plot to estimate plant height, stem diameter and leaf area index (LAI) according to Beadle (1993).
Yield
At harvest dates (on the 4th and 10th August in 2019 and 2020, respectively), whole plants of each plot were harvested to estimate head weight plant−1, seed yield ha−1. A representative sample of seeds was obtained for estimating oil percentage using Soxhlet Apparatus with hexane as organic solvent according to AOAC (2012). Then, oil yield ha−1 was calculated by multiplying seed oil content by seed yield ha−1.
Irrigation Water Use Efficieny (IWUE)
According to Kirda et al. (2005), IWUE (kg m−3) was calculated by dividing seed yield (kg ha−1) by the total amount of irrigation water applied (m3 ha−1) during each growing season.
Yield Response Factor (Ky)
The yield response factor (Ky) was computed from the seed yield for each antioxidant treatment using the pooled data across the two experimental years (Doorenbos and Kassam 1979), using Eq 1:
where Ya is the seed yield (kg ha−1) obtained from DI treatment, Yx is the seed yield (kg ha−1) obtained from the FI treatment, ETa is the crop water consumption (m3 ha−1) under deficit irrigation (DI), and ETx is the crop water consumption (m3 ha−1) under full irrigation (FI).
Data Analysis
Differences in the physiological, growth, yield traits, and IWUE among treatments were statistically analyzed by two-way analysis of variance (ANOVA) for each season using Costat software program version 6.303 (2004). For comparison among means, Duncan’s multiple range test was used at 0.05 probability level (p ≤ 0.05).
Results
Physiological Traits
Results illustrated in Table 2 showed that deficit irrigation (DI) lowered chlorophyll a by 22.08 and 24.17% and chlorophyll b by 38.14 and 42.88% while increased proline content by 116.49 and 105.90% as comparing to full irrigation (FI) in 2019 and 2020 seasons respectively. ASC300 gave the highest values of pigments concentration in leaves, with increases of 18.33 and 20.72% in chlorophyll a as well as 27.69 and 35.48% in chlorophyll b, while proline content reduced by 17.12 and 16.22% compared to the check treatment (Tap water) in 2019 and 2020 seasons, respectively (Table 2). In full irrigated sunflower plots, exogenous application of higher rate of ascorbic acid (FI plus ASC300) exhibited the highest increase of chlorophyll a, chlorophyll b and the lowest proline content compared to other interaction treatments (Table 2). While, the lowest contents of chlorophyll a and the highest value of proline were obtained in leaves of plants grown under DI plus check treatment in both seasons. DI × CA250 or check treatment showed the minimal chlorophyll b.
Growth Traits
The irrigation regime significantly affected the plant height, stem diameter and LAI, with the FI regime resulting in 9.29, 22.95 and 35.38% increases, respectively, in the 2019 season and 9.11, 24.13 and 36.91% increases, respectively, in the 2020 season higher than the DI regime (Table 3). Compared to the check treatment ASC300 treatment caused increases of 4.96 and 4.29%, in plant height, 7.96 and 7.77% in stem diameter, and 16.21 and 16.77% in LAI in both growing seasons, respectively. The promotive effects of CA500 or ASC150 on plant height and stem diameter as well as CA500 on LAI came in the second order. Overall, interaction revealed the potential of ASC300 to improve plant growth either with FI or DI (Table 3). Specifically for each irrigation pattern, plant height, stem diameter and LAI with ASC300 in both seasons as well as LAI with CA500 in 2019 season showed the maximum values under FI. Similar trent was also obtained under DI.
Yield
The irrigation regime significantly influenced head weight, seed yield and oil yield, with the DI regime resulting in 35.88, 22.42, and 29.95% lower values, respectively, in 2019 season and 36.76, 23.47, and 31.19% lower values, respectively, in 2020 season than the FI regime (Table 4). ASC300 foliar application was the most efficient treatment during both seasons, with increasing the head weight by 18.24 and 17.54%, seed yield by 6.84 and 8.84%, and oil yield by 11.83 and 14.20% in 2019 and 2020 seasons, respectively, as compared to the check treatment (Table 4).
Sunflower sprayed with ASC300 and irrigated with FI produced the highest values of all yield traits surpassing the other combinations. Moreover, it should be noted that the seed yield was significantly higher with FI plus ASC300 and DI plus ASC300 treatments than their counterpart check treatment achieving 13.93 and 8.89% increases in 2019 season as well as 16.23 and 11.32% increases in 2020 season, respectively.
Irrigation Water Use Efficiency
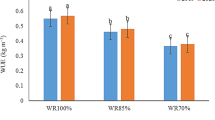
As dipeted in Fig. 2, ASC300 was the most effective practice for enhancing IWUE of sunflower in 2019 and 2020 seasons, surpassing the other practices either under FI or DI. With DI, IWUE was improved with antioxidant-treated plants compared to untreated plants.

Irrigation water use efficiency (IWUE) of sunflower response to irrigation regime and antioxidant applications in 2019 and 2020 seasons. (FI and DI full and deficit irrigation, representing 100 and 85 of crop evapotranspiration, respectively, ASC150 and ASC300 ascorbic acid at rates of 150 and 300 mg L−1, CA250 and CA500 citric acid at rates of 250 and 500 mg L−1, respectively, Check tap water treatment). (Values are the mean of 3 replicates ± standard errors. Different letters between bars indicate that there are significant differences at p ≤ 0.05 based on Duncan’s multiple range test)
Yield Response Factor
For providing an indication of the tolerance level of sunflower crop to water deficit stress, Ky was estimated for antioxidant treatment with pooled data from the two growing seasons (Fig. 3). Ky factor values were > 1 under any antioxidant treatment. The values of Ky factor were noted as follows: ASC300 > CA500 > CA250 > ASC150 > check treatment.

The change in yield response factor (Ky) of sunflowr with application of ascorbic and citric acids. (ASC150 and ASC300 ascorbic acid at rates of 150 and 300 mg L−1, CA250 and CA500 citric acid at rates of 250 and 500 mg L−1, respectively, Check tap water treatment)
Discussion
Drought as abiotic stress is considered as one of the most important consequences of climate change, which is an obstacle to crop productivity. Drought stress has tremendous impacts on plants, which could involve imbalance, stress damage, growth delay, and low availability of nutrients (Mubarak et al. 2021). Several chemical compounds are exogenously applied to improve grwoth and health of plants under both normal or stress conditions (Noureldin et al. 2013; Saudy et al. 2020b, 2022a; El-Metwally and Saudy 2021b; Elgala et al. 2022; El-Metwally et al. 2022b). The current study investigated some drought response mechanisms in sunflower under application of ASC and CA. As presented in Table 2, decreasing irrigation water by 15% than normal led to reduction in chlorophyll content and increase in proline content. Such findings are in accordance with those found by El-Bially et al. (2018) and Mostafa (2020) who reported that photosynthetic pigments of sunflower leaf (chlorophyll a and b) had large decline, but proline content increased due to drought stress. It has been reported that reactive oxygen species (ROS) overproduced in plant cells exposed to environmental stresses (Hatamian et al. 2020; Souri et al. 2019) can damage membrane and other essential macromolecules like photosynthetic pigments, proteins, DNA and lipids. Biochemical damage was measured due to production of ROS which eventually led to poor growth and metabolic damage of the plant (Zafar et al. 2015). High ROS levels cause serious dysfunction in many processes such as hormonal equilibrium, gene expression, pathways of signaling, photosynthetic efficiency, protein inactivation, inhibit the action of multiple enzymes involved in metabolic pathways and decrease grain yield, resulting in lipid and DNA oxidation (Huang et al. 2012; Choudhury et al. 2017). It has been proved that plant growth regulators and osmoprotectants like free amino acids, sugars, and polyamines have protective roles against drought (Chan et al. 2013). In this regrad, proline accumulation in stressed plants has been well established to play a key role as osmoregulation defense mechanism, leading to prevent the cell osmotic pressure and survive in the extreme conditions (Souri and Bakhtiarizade 2019; Souri and Tohidloo 2019). In this connection, Manivannan et al. (2007) reported that water stress caused reduction in activity of proline oxidase so, proline content increased. Moreover, a large proportion of the drought-responsive proteins are involved in photosynthesis. Rubisco (Ribulose bisphosphate carboxylase) is a vital enzyme associated with carbon fixation (Feller et al. 2008). Rubisco similarly showed decreasing abundance under drought stress, indicating that drought negatively affects the key protein of the photosynthetic apparatus (Wang et al. 2017).
Since drought affects plant physiology as well as nutrient availability and accumulation (Mubarak et al. 2021; Salem et al. 2021, 2022), crop traits and its potentiality to expliot irrigation water were influenced (Tables 2 and 3). Drought stress caused significant reduction in growth, yield traits as well as IWUE because irrigation is the key factor for obtaining high yield (FAO 2010). Herein, deficit water significantly reduced root length, stem length, total leaf area, fresh and dry weight of sunflower plants (Manivannan et al. 2007). Thus, drought stress has been shown to significantly decrease plant height (Sincik et al. 2013), stem diameter (Saeed et al. 2015), leaf area index (Furtado et al. 2016), head weight (Ibrahim et al. 2016), seed yield (Soleimanzadeh 2012), oil yield (Kassab et al. 2012) and water use efficiency (El-Bially et al. 2018). These reductions could be due to lowering content of chlorophyll in leaves (Table 2), which leads to a decrease in photosynthesis rate and the dry matter accumulation.
On the other site, plant cells have many effective defense mechanisms to remove the harmful effect of ROS (Birben et al. 2012). ASC and CA are non-enzymatic antioxidants that play an important role to protect plants from oxidative damage by scavenging and sweep of ROS (Prasad and Upadhyay 2011; Tahjib-ul-Arif et al. 2021). Therefore, enhancements in leaves content of chlorophylls a and b were obtained owing to the application of antioxidants (Table 2). Previous studies have shown that foliar application of ASC or CA significantly increased leaf pigmints of stressed plants (Amin and Ismail 2015; El-Mantawy 2017; Farid et al. 2017, 2019). On the other hand, proline content in leaves was reduced when ASC and CA were sprayed under drought stress as compared to control (see Table 3),. Proline stabilizes subcellular structures and molecules experiencing osmotic stress conditions by working as a molecular chaperone, maintaining the integrity of proteins. Previous studied have shown that foliar application of ASC or CA significantly decreased proline content in leaves (El-Bially et al. 2018; Mostafa 2020). Since antioxidants alleviate the effects of stress, ASC and CA significantly decrease leaves proline content as compared to control treatment (Mostafa 2020). CA foliar application improved the germination rate and root weight of sunflower plant by improving the activities of several antioxidants enzymes including superoxide dismutase, catalase, peroxidase, and ascorbate peroxidase (Ondrasek et al. 2019). Using ASC and CA as foliar applications, especiallly with higher rates (ASC300 and CA500), counteraceted the harmful effect of drought stress and significantly increased growth and yield attributes. These results are in consistance with the findings of El-Mantawy (2017). Since ASC is a preventive agent and the important compound working nonenzymatic system reinforcement against ROS in plant (Qian et al. 2014), foliar application of ASC enhanced physiological and biochemical traits, productivity and water use efficiency of sunflower under abiotic stress (Saudy et al. 2021a).
Despite ASC and CA alleviated partially the adverse impacts of deficit water, sunflower plants still sensetive to low water supply. In this regard, Fig. 3 showed that Ky values exceeded the unit (higher than 1.0) clarifing the sensetivity of sunflower to drought. Ky indicates the relationship between relative yield and relative crop water consumption (Lovelli et al. 2007; Singh et al. 2010), with values > 1 indicating that the crop is very sensitive to water stress, values < 1 indicating that the crop is more tolerant to water stress, and values of 1 indicating that the relative yield reduction is equal to the relative water use reduction (Steduto et al. 2012). Accordingly, our findings refer to that sunflower plants require additional practices to increase their tolerance to deficit water stress. The value of Ky was affected by climate and soil conditions, irrigation method and applied amount of irrigation water (Aydinsakir et al. 2021). Candogan and Yazgan (2016) reported that Ky value was greater than 1 (1.21) and stated that soybean was sensitive to water stress.
Coclussion
It could be concluded that reduction in the economic product of sunflower (seed yield) could not completely be compensated by application of antioxidants, i.e. ascorbic and citric. However, the adverse effects of drought were partially alleviated with antioxidant especially ascorbic acid. Also, since sunflower markedly responded to increasing the antioxidant level up to 300 mg L−1 for ascorbic acid and 500 mg L−1 for citric acid, this opens the field to further studies to examine higher rates of each under drought stress in sunflower. Moreover, other agronomic practices should be tested and adopted along application of antioxidant since sunflower still sensetive to drought as proved from measuing of yield response factor.
References
Abd El-Mageed TA, Mekdad AAA, Rady MOA, Abdelbaky AS, Saudy HS, Shaaban A (2022) Physio-biochemical and agronomic changes of two sugar beet cultivars grown in saline soil as influenced by potassium fertilizer. J Soil Sci Plant Nutr. https://doi.org/10.1007/s42729-022-00916-7
Abd-Elrahman SH, Saudy HS, Abd El-Fattah DA, Hashem FA (2022) Effect of irrigation water and organic fertilizer on reducing nitrate accumulation and boosting lettuce productivity. J Soil Sci Plant Nutr 22:2144–2155. https://doi.org/10.1007/s42729-022-00799-8
Allen RG, Pereira LS, Raes D, Smith M (1998) Crop evapotranspiration. Guidelines for computing crop water requirements. FAO irrigation and drainage paper, vol 56. FAO, Rome, p 300
Amin MA, Ismail MA (2015) Effect of indole butyric, arginine, cyanocobalamine (B12), ascorbic acid and their interactions on growth, yield and some metabolic constituents of sunflower plants. Int J Adv Res Biol Sci 2:154–162
AOAC (2012) Official method of analysis: Association of Analytical Chemists, 19th edn. AOAC, Washington
Aydinsakir K, Dinc N, Buyuktas D, Kocaturk M, Ozkan CF, Karaca C (2021) Water productivity of soybeans under regulated surface and subsurface drip irrigation conditions. Irrig Sci 39:773–787. https://doi.org/10.1007/s00271–021–00744–0
Bates LS, Waldren RP, Teare ID (1973) Rapid determination of free proline for water-stress studies. Plant Soil 39:205–207
Beadle CL (1993) Growth analysis. In: Hall DO (ed) Photosynthesis and production in a changing environment. A field and laboratory manual. Chapman and Hall, London, pp 36–46
Bilska K, Wojciechowska N, Alipour S, Kalemba EM (2019) Ascorbic acid—The little-known antioxidant in woody plants. Antioxid 8:645
Birben E, Sahiner UM, Sackesen C, Erzurum S, Kalayci O (2012) Oxidative stress and antioxidant defense. World Allergy Organ J 5:9–19. https://doi.org/10.1097/WOX.0b013e3182439613
Bresson J, Vasseur F, Dauzat M, Koch G, Granier C, Vile D (2015) Quantifying spatial heterogeneity of chlorophyll fluorescence during plant growth and in response to water stress. Plant Method 11:23. https://doi.org/10.1186/s13007–015–0067–5
Candogan BN, Yazgan S (2016) Yield and quality response of soybean to full and deficit irrigation at different growth stages under subhumid climatic conditions. J Agric Sci 22:129–144
Chan KX, Wirtz M, Phua SY, Estavillo GM, Pogson BJ (2013) Balancing metabolites in drought: The sulfur assimilation conundrum. Trends Plant Sci 18:18–29
Choudhury FK, Rivero RM, Blumwald E, Mittler R (2017) Reactive oxygen species, abiotic stress and stress combination. Plant J 90:856–867
Cui NB, Du TS, Kang SZ, Li FS, Hu XT, Wang MX, Li ZJ (2009) Relationship between stable carbon isotope discrimination and water use efficiency under regulated deficit irrigation of pear-jujube tree. Agric Wat Manag 96:1615–1622
Doorenbos J, Kassam AH (1979) Yield response to water. FAO irrigation and drainage, p 33
El-Bially MA, Saudy HS, El-Metwally IM, Shahin MG (2018) Efficacy of ascorbic acid as a cofactor for alleviating water deficit impacts and enhancing sunflower yield and irrigation water-use efficiency. Agric Water Manag 208:32–139. https://doi.org/10.1016/j.agwat.2018.06.016
El-Bially MA, Saudy HS, El-Metwally IM, Shahin MG (2022a) Sunflower response to application of L‑ascorbate under thermal stress associated with different sowing dates. Gesunde Pflan 74:87–96. https://doi.org/10.1007/s10343-021-00590-2
El-Bially MA, Saudy HS, Hashem FA, El-Gabry YA, Shahin MG (2022b) Salicylic acid as a tolerance inducer of drought stress on sunflower grown in sandy soil. Gesunde Pflan. https://doi.org/10.1007/s10343-022-00635-0
El-Mantawy RF (2017) Physiological role of antioxidants in improving growth and productivity of sunflower under different sources of nitrogen fertilizers. Egypt J Agron 39:167–177
El-Metwally IM, Saudy HS (2021a) Interactional impacts of drought and weed stresses on nutritional status of seeds and water use efficiency of peanut plants grown in arid conditions. Gesunde Pflanz 73:407–416. https://doi.org/10.1007/s10343-021-00557-3
El-Metwally IM, Saudy HS (2021b) Interactive application of zinc and herbicides affects broad-leaved weeds, nutrient uptake, and yield in rice. J Soil Sci Plant Nutr 21:238–248. https://doi.org/10.1007/s42729-020-00356-1
El-Metwally IM, Saudy HS, Abdelhamid MT (2021) Efficacy of benzyladenine for compensating the reduction in soybean productivity under low water supply. Ital J Agrometeorol 2:81–90. https://doi.org/10.36253/ijam-872
El-Metwally IM, Geries L, Saudy HS (2022a) Interactive effect of soil mulching and irrigation regime on yield, irrigation water use efficiency and weeds of trickle-irrigated onion. Arch Agron Soil Sci 68:1103–1116. https://doi.org/10.1080/03650340.2020.1869723
El-Metwally IM, Sadak MS, Saudy HS (2022b) Stimulation effects of glutamic and 5‑Aminolevulinic acids on photosynthetic pigments, physio-biochemical constituents, antioxidant activity, and yield of peanut. Gesunde Pflanz. https://doi.org/10.1007/s10343-022-00663-w
El-Tohamy W, El-Abagy H, Badr M, Gruda N (2013) Drought tolerance and water status of bean plants (Phaseolus vulgaris L.) as affected by citric acid application. J Appl Bot Food Qual 86(212):216
Elgala AM, Abd-Elrahman SH, Saudy HS, Nossier MI (2022) Exploiting Eichhornia crassipes shoots extract as a natural source of nutrients for producing healthy tomato plants. Gesunde Pflan 74:457–465. https://doi.org/10.1007/s10343-022-00622-5
FAO (2010) Sunflower crude and refined oils. Viale delle Terme di Caracalla. FAO, Rome
FAO (2017) Food outlook report, June 2017. FAO, Rome
Farid M, Ali S, Rizwan M, Ali Q, Abbas F, Bukhari SAH, Saeed R, Wu L (2017) Citric acid assisted phytoextraction of chromium by sunflower; morpho-physiological and biochemical alterations in plants. Ecotoxicol Environ Saf 145:90–102
Farid M, Ali S, Saeed R, Rizwan M, Bukhari SAH, Abbasi GH, Hussain A, Ali B, Zamir MSI, Ahmad I (2019) Combined application of citric acid and 5‑aminolevulinic acid improved biomass, photosynthesis and gas exchange attributes of sunflower (Helianthus annuus L.) grown on chromium contaminated soil. Int J Phytoremed 21:760–767
Feller U, Anders I, Mae T (2008) Rubiscolytics: fate of rubisco after its enzymatic function in a cell is terminated. J Exp Bot 59:1615–1624. https://doi.org/10.1093/jxb/erm242
Furtado GF, Xavier DA, Andrade EM, Lima GS, Chaves LH, Vasconcelos AC, Wanderley JA (2016) Growth and physiological responses of sunflower grown under levels of water replacement and potassium fertilization. Afr J Agric Res 11:1273–1281
Gebaly SG, Ahmed FM, Namich AA (2013) Effect of spraying some organic, amino acids and potassium citrate on alleviation of drought stress in cotton plant. J Plant Prod 4:1369–1381
Hatamian M, Nejad RA, Kafi M, Souri MK, Shahbazi K (2020) Nitrate improves hackberry seedling growth under cadmium application. Heliyon 6:e3247
Huang GT, Ma SL, Bai LP, Zhang L, Ma H, Jia P, Liu J, Zhong M, Guo ZF (2012) Signal transduction during cold, salt, and drought stresses in plants. Mol Biol Rep 39:969–987
Ibrahim MF, Faisal A, Shehata SA (2016) Calcium chloride alleviates water stress in sunflower plants through modifying some physio-biochemical parameters. Am Eurasia J Agric Environ Sci 16:677–693
Kaleem S, Hassan F, Mahmood I, Ahmad M, Ullah R, Ahmad M (2011) Response of sunflower to environmental disparity. Nat Sci 9:73–81
Kassab OM, Abo Ellil AA, Abo El-Kheir MS (2012) Water use efficiency and productivity of two sunflower cultivars as influenced by three rates of drip irrigation water. J Appl Sci Res 8:3524–3529
Kirda C, Topcu S, Kaman H, Ulge AC, Yazici A, Cetin M, Derici MR (2005) Grain yield response and N‑fertiliser recovery of maize under deficit irrigation. Field Crops Res 93:132–141
Lovelli S, Perniola M, Ferrara A, Di Tommaso T (2007) Yield response factor to water (Ky) and water use efficiency of Carthamus tinctorius L. and Solanum melongena L. Agric Wat Manag 92:73–80
Makhlouf BSI, Khalil Soha RA, Saudy HS (2022) Efficacy of humic acids and chitosan for enhancing yield and sugar quality of sugar beet under moderate and severe drought. J Soil Sci Plant Nutr 22:1676–1691. https://doi.org/10.1007/s42729-022-00762-7
Manivannan P, Jaleel CA, Sankar B, Kishorekumar A, Somasundaram R, Alagu Lakshmanan GM, Panneerselvam R (2007) Growth, biochemical modifications and proline metabolism in Helianthus annuus L. as induced by drought stress. Colloids Surf B Biointerfaces 59:141–149
Mostafa N (2020) Effect of foliar application of some vitamins and irrigation intervals on vegetative growth, flowering, and some biochemical constituents of Helianthus annuus L. plants. Sci J Agri Sci 2:1–8
Mubarak M, Salem EMM, Kenawey MKM, Saudy HS (2021) Changes in calcareous soil activity, nutrient availability, and corn productivity due to the integrated effect of straw mulch and irrigation regimes. J Soil Sci Plant Nutr 21:2020–2031. https://doi.org/10.1007/s42729-021-00498-w
Noureldin NA, Saudy HS, Ashmawy F, Saed HM (2013) Grain yield response index of bread wheat cultivars as influenced by nitrogen levels. Ann Agric Sci 58(2):147–152. https://doi.org/10.1016/j.aoas.2013.07.012
Omar MEA, Nermeen MS, Enayat AO, Noha NY (2018) Chapter 16—Citric acid an antioxidant in liver. In: Patel VB, Rajendram R, Preedy VR (eds) The Liver. Academic Press, New York, pp 183–198
Ondrasek GH, Bakić M, Zovko L, Filipović C, Meriño-Gergichevich R, Savić Z, Rengel Z (2019) Biogeochemistry of soil organic matter in agroecosystems and environmental implications. Sci Total Environ 658:1559–1573
Ponce V, Pandey R, Ercan S (2000) Characterization of drought across climatic spectrum. J Hydrol Eng 5:222–224
Prasad V, Upadhyay RS (2011) Ascorbate and glutathione: saviours against oxidative stress. In: Anjum NA, Umar S, Ahmad A (eds) Oxidative stress in plants: causes, consequences and tolerance. I.K. International Publishing House, New Delhi, pp 149–176
Qahar A, Khan ZH, Anwar S, Badshan H, Ullah H (2010) Nitrogen use efficiency, yield and other characteristics of sunflower (Helianthus annuus L.) hybrids as affected by different levels of nitrogen. Biol Diver Conserv 3:121–125
Qian HF, Peng XF, Han X, Ren J, Zhan KY, Zhu M (2014) The stress factor, exogenous ascorbic acid, affects plant growth and the antioxidant system in Arabidopsis thaliana. Russ J Plant Physiol 61:467–475
Rosa PM, Antoniassi R, Freitas SC, Bizzo HR, Zanotto DL, Oliveira MF, Castiglioni VBR (2009) Chemical composition of Brazilian sunflower varieties. Helia 32:145–156
Saeed U, Sher A, Hussain S, Khan A, Ameen A, Jincai L, Shakoor A (2015) Impact of foliar application of boron on growth and yield of sunflower (Helianthus annuus L.) under different irrigation conditions. Acad J Agric Res 3:219–225
Salem EMM, Kenawey MKM, Saudy HS, Mubarak M (2021) Soil mulching and deficit irrigation effect on sustainability of nutrients availability and uptake, and productivity of maize grown in calcareous soils. Commun Soil Sci Plant Anal 52:1745–1761. https://doi.org/10.1080/00103624.2021.1892733
Salem EMM, Kenawey MKM, Saudy HS, Mubarak M (2022) Influence of silicon forms on nutrient accumulation and grain yield of wheat under water deficit conditions. Gesunde Pflanz. https://doi.org/10.1007/s10343-022-00629-y
Saudy HS, El-Bagoury KF (2014) Quixotic coupling between irrigation system and maize-cowpea intercropping for weed suppression and water preserving. Afr Crop Sci J 22:97–108
Saudy HS, El-Metwally IM (2019) Nutrient utilization indices of NPK and drought management in groundnut under sandy soil conditions. Commun Soil Sci Plant Anal 50:1821–1828. https://doi.org/10.1080/00103624.2019.1635147
Saudy HS, El-Metwally IM (2022) Effect of irrigation, nitrogen sources and metribuzin on performance of maize and its weeds. Commun Soil Sci Plant Anal. https://doi.org/10.1080/00103624.2022.2109659
Saudy HS, Mubarak MM (2015) Mitigating the detrimental impacts of nitrogen deficit and fenoxaprop-p-ethyl herbicide on wheat using silicon. Commun Soil Sci Plant Anal 46:913–923. https://doi.org/10.1080/00103624.2015.1011753
Saudy HS, Abd El-Momen WR, El-khouly NS (2018) Diversified nitrogen rates influence nitrogen agronomic efficiency and seed yield response index of sesame (Sesamum indicum, L.) cultivars. Commun Soil Sci Plant Anal 49(19):2387–2395. https://doi.org/10.1080/00103624.2018.1510949
Saudy HS, El-Metwally IM, Abd El-Samad GA (2020a) Physio-biochemical and nutrient constituents of peanut plants under bentazone herbicide for broad-leaved weed control and water regimes in dry land areas. J Arid Land 12:630–639. https://doi.org/10.1007/s40333-020-0020-y
Saudy HS, Hamed MF, Abd El-Momen WR, Hussein H (2020b) Nitrogen use rationalization and boosting wheat productivity by applying packages of humic, amino acids and microorganisms. Commun Soil Sci Plant Anal 51:36–1047. https://doi.org/10.1080/00103624.2020.1744631
Saudy HS, El-Bially MA, El-Metwally IM, Shahin MG (2021a) Physio-biochemical and agronomic response of ascorbic acid-treated sunflower (Helianthus annuus) grown at different sowing dates and under various irrigation regimes. Gesunde Pflanz 73:169–179. https://doi.org/10.1007/s10343-020-00535-1
Saudy HS, El-Metwally IM, Shahin MG (2021b) Co-application effect of herbicides and micronutrients on weeds and nutrient uptake in flooded irrigated rice: Does it have a synergistic or an antagonistic effect? Crop Prot 149:105755. https://doi.org/10.1016/j.cropro.2021.105755
Saudy HS, Abd El-Samad GA, El-Temsah ME, El-Gabry YA (2022a) Effect of iron, zinc and manganese nano-form mixture on the micronutrient recovery efficiency and seed yield response index of sesame genotypes. J Soil Sci Plant Nutr 22:732–742. https://doi.org/10.1007/s42729-021-00681-z
Saudy HS, Salem EMM, Abd El-Momen WR (2022b) Effect of potassium silicate and irrigation on grain nutrient uptake and water use efficiency of wheat under calcareous soils. Gesunde Pflanz. https://doi.org/10.1007/s10343-022-00729-9
Sharma R, Bhardwaj R, Thukral AK, Al-Huqail AA, Siddiqui MH, Ahmad P (2019) Oxidative stress mitigation and initiation of antioxidant and osmoprotectant responses mediated by ascorbic acid in Brassica juncea L. subjected to copper (II) stress. Ecotoxicol Environ Saf 182:109436
Sincik M, Goksoy AT, Dogan R (2013) Responses of sunflower (Helianthus annuus L.) to irrigation and nitrogen fertilization rates. Zemdirbyste-Agriculture 100:151–158
Singh Y, Rao SS, Regar PL (2010) Deficit irrigation and nitrogen effects on seed cotton yield, water productivity and yield response factor in shallow soils of semi-arid environment. Agric Water Manag 97:965–970
Soleimanzadeh H (2012) Response of sunflower (Helianthus annuus L.) to selenium application under water stress. World Appl Sci J 17:1115–1119
Souri MK, Bakhtiarizade M (2019) Biostimulation effects of rosemary essential oil on growth and nutrient uptake of tomato seedlings. Sci Hortic 243:472–476
Souri MK, Hatamian M (2019) Aminochelates in plant nutrition; a review. J Plant Nutr 42:67–78
Souri MK, Tohidloo G (2019) Effectiveness of different methods of salicylic acid application on growth characteristics of tomato seedlings under salinity. Chem Biol Technol Agric 6:26. https://doi.org/10.1186/s40538-019-0169-9
Souri MK, Hatamian M, Tesfamariam T (2019) Plant growth stage influences heavy metal accumulation in leafy vegetables of garden cress and sweet basil. Chem Biol Technol Agric 26:25. https://doi.org/10.1186/s40538–019–0170–3
Steduto P, Hsiao TC, Fereres E, Raes D (2012) Crop yield response to water. FAO irrigation and drainage, paper, vol 66. FAO, Rome
Tahjib-ul-Arif M, Zahan MI, Karim MM, Imran S, Hunter CT, Islam MS, Mia MA, Hannan MA, Rhaman MS, Hossain MA, Brestic M, Skalicky M, Murata Y (2021) Citric acid-mediated abiotic stress tolerance in plants. Int J Mol Sci 22:7235
Vermeirer L, Jopling GA (1984) Localized irrigation. FAO. Irrigation paper, vol 36. FAO, Rome
Wang Y, Peng C, Zhan Y, Yu L, Li M, Li J, Geng G (2017) Comparative proteomic analysis of two sugar beet cultivars with contrasting drought tolerance. J Plant Growth Regul 36:537–549. https://doi.org/10.1007/s00344–017–9703–9
Wettstein D (1957) Chlorophyll lethal faktoren under submikro skopoch for mvechsel der plastide. Exp Cell Res 12:427–433
Yan W, Zhong Y, Shangguan Z (2016) A meta-analysis of leaf gas exchange and water status responses to drought. Sci Rep 6:1–9
Zafar S, Ashraf MY, Niaz M, Kausar A, Hussain J (2015) Evaluation of wheat genotypes for salinity tolerance using physiological indices as screening tool. Pak J Bot 47:397–405
Acknowledgements
The authors extend their appreciation to Faculty of Agriculture, Ain Shams University and Central Laboratory for Agricultural Climate, Agricultural Research Center, Egypt, for technical support of this study
Funding
Open access funding provided by The Science, Technology & Innovation Funding Authority (STDF) in cooperation with The Egyptian Knowledge Bank (EKB).
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
H.S. Saudy, M.E. El-Bially, F.A. Hashem, M.G. Shahin and Y.A. El-Gabry declare that they have no competing interests.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Saudy, H.S., El-Bially, M.E., Hashem, F.A. et al. The Changes in Yield Response Factor, Water Use Efficiency, and Physiology of Sunflower Owing to Ascorbic and Citric Acids Application Under Mild Deficit Irrigation. Gesunde Pflanzen 75, 899–909 (2023). https://doi.org/10.1007/s10343-022-00736-w
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10343-022-00736-w