
Abstract
The phylogenetic relationship among Geosmithia argillacea, Talaromyces emersonii, Talaromyces byssochlamydoides and other members of the Trichocomaceae was studied using partial RPB2 (RNA polymerase II gene, encoding the second largest protein subunit), Tsr1 (putative ribosome biogenesis protein) and Cct8 (putative chaperonin complex component TCP-1) gene sequences. The results showed that these species form a distinct clade within the Trichocomaceae and Trichocoma paradoxa is phylogenetically most closely related. Based on phenotypic and physiological characters and molecular data, we propose Rasamsonia gen. nov. to accommodate these species. This new genus is distinct from other genera of the Trichocomaceae in being thermotolerant or thermophilic and having conidiophores with distinctly rough walled stipes, olive-brown conidia and ascomata, if present, with a scanty covering. Species within the genus Rasamsonia were distinguished using a combination of phenotypic characters, extrolite patterns, ITS and partial calmodulin and β-tubulin sequences. Rasamsonia brevistipitata sp. nov. is described and five new combinations are proposed.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Thermophilia and thermotolerance have been variously defined (Apinis 1963; Cooney and Emerson 1964; Craveri et al. 1964; Crisan 1973). The commonly accepted definition is that of Cooney and Emerson (1964), who defined thermophilic species as those which can grow at or above 50°C, but not below 20°C. The definition of thermotolerance is similar except that the maximum growth temperature is at or above 45°C, having a maximum near 50°C, and the minimum growth temperature is below 20°C. Thermophilic fungi have gained much attention in biotechnology the last decades, primarily because of their capacity to produce thermostable enzymes, which are stable at room temperature and can be purified by heat treatment (Maheshwari et al. 2000; Turner et al. 2007). For example, the thermophilic species Talaromyces emersonii produces multiple β-glucosidase enzymes, a number of which are components of a potent cellulytic enzyme system and are used in commercial applications (Collins et al. 2007; Coughlan et al. 1993; McHale and Coughlan 1981; Murray et al. 2004).
Thermophilic fungi are not restricted to a specific taxonomic order and occur all over the fungal tree of life. Examples are Rhizomucor in the Mucorales, Myceliophthora in Sordariales and Talaromyces, Thermomyces and Thermoascus in the Eurotiales (Mouchacca 1997). The majority of Talaromyces species are mesophiles but exceptions are species within sections Emersonii and Thermophila. Section Emersonii includes Talaromyces emersonii, T. byssochlamydoides, T. bacillisporus and T. leycettanus, all of which grow well at 40°C. Talaromyces bacillisporus is thermotolerant, T. leycettanus is thermotolerant to thermophilic, and T. emersonii and T. byssochlamydoides are truly thermophilic (Stolk and Samson 1972). The sole member of Talaromyces section Thermophila, T. thermophilus, grows rapidly at 50°C (Evans and Stolk 1971; Evans 1971; Stolk and Samson 1972). The current classification of these thermophilic Talaromyces species is mainly based on phenotypic and physiological characters, such as their ability to grow above 40°C, ascospore color, the structure of ascomatal covering and the formation of a certain type of anamorph. Stolk and Samson (1972) stated that the members of the section Emersonii have anamorphs of either Paecilomyces (T. byssochlamydoides and T. leycettanus) or Penicillium cylindrosporum series (T. emersonii and T. bacillisporus). Later, Pitt (1979) transferred the species belonging to the Penicillium cylindrosporum series to the genus Geosmithia, based on various characters such as the formation of conidia from terminal pores instead of on collula (necks), a character of Penicillium and Paecilomyces. Within the genus Geosmithia, only G. argillacea is thermotolerant, and Stolk et al. (1969) and Evans (1971) proposed a connection with members of Talaromyces sect. Emersonii. The phylogenetic relationship of the thermophilic Talaromyces species within Talaromyces and the Trichocomaceae is unknown.
In this study, the phylogenetic relationship among Geosmithia argillacea, T. emersonii, T. byssochlamydoides and other thermotolerant/thermophilic members of the Trichocomaceae is studied using partial RPB2 (RNA polymerase II gene, encoding the second largest protein subunit), Tsr1 (putative ribosome biogenesis protein) and Cct8 (putative chaperonin complex component TCP-1) gene sequences. These data indicate that these species are closely related and phenotypic and physiological characters show that they represent a new genus. The taxonomy of this new genus is studied here using a combination of phenotypic characters, extrolite patterns, ITS and partial calmodulin and β-tubulin sequences. One new species is described and five new combinations are proposed.
Materials and methods
Strains
The strains used in the study on the phylogeny of the Trichocomaceae are listed in Supplementary Table 1. An overview of strains used for phenotypic analysis and phylogeny is shown in Table 1. All cultures are maintained in the culture collection of the CBS-KNAW Fungal Biodiversity Centre, Utrecht, The Netherlands.
Morphological characterization
Cultures for morphological observation were inoculated in a three point position onto the following agar media: Czapek yeast extract agar (CYA), CYA supplemented with 5% NaCl (CYAS), yeast extract sucrose agar (YES), creatine sucrose agar (CREA), dichloran 18% glycerol agar (DG18), oatmeal agar (OA) and malt extract agar (Oxoid) (MEA). All media were prepared as described by Samson et al. (2010). Two sets were inoculated per strain: one set was incubated at 25°C, the other at 37°C. Colony diameter and other macroscopical features were recorded after 7 days of incubation. Microscopical features were studied by light microscopy (Olympus BH2 and Zeiss Axioskop 2 Plus) and microscopic mounts were made from MEA or OA.
Molecular analysis
Strains were grown on MEA for 7–14 days prior to DNA extraction. Genomic DNA was extracted using the UltracleanTM Microbial DNA isolation Kit (MoBio, Solana Beach, U.S.A.) according to the manufacturer’s instructions. For the phylogenetic analysis of the Eurotialean taxa, parts of the RPB2, Tsr1 and Cct8 genes were sequenced. The latter two genes were chosen, since they were among the best-performing genes for recovering robust phylogenies in the study by Aguileta et al. (2008). The partial RPB2, Tsr1 and Cct8 sequences consists of exon data only and the alignment of the data set was performed using the Muscle software on the translated amino acid data. The ITS (incl. 5.8S rDNA), β-tubulin and calmodulin loci were sequenced in order to study the phylogenetic relationship among T. emersonii, T. byssochlamydoides, T. eburneus, G. argillacea and G. cylindrospora. The obtained sequences were aligned using the software Muscle on the EMBL-EBI website and subsequently combined. Primers used for amplification and sequencing and annealing temperatures are shown in Table 2. Maximum likelihood analysis was performed on the two data sets using RAxML version 7.2.8. Each locus was treated as a separate partition. The number of bootstrap replicates was set on 1,000 replicates (Stamatakis et al. 2008). A second measure of branch support was conducted using a Bayesian tree inference (BI) analysis. Prior to analysis, the best nucleotide substitution models were determined for each partition with MrModeltest v2.2 (Nylander 2004). A BI analysis was performed with MrBayes v3.1.2 (Huelsenbeck and Ronquist 2001). One tree was saved per 100 generations, and the run was automatically ended when the standard deviation of split frequencies was below 0.01. To avoid suboptimal trees being taken into account for the consensus tree, a burn-in of 25% of the saved trees was used. Coccidioides immitis sequences were downloaded from the full genome data set (Sharpton et al. 2009) and used as an outgroup in the study of the phylogeny of selected Trichocomaceae. Trichocoma paradoxa CBS 247.57 and CBS 103.73 were used as an outgroup in the phylogenetic analysis.
Extrolite analysis
All strains, except Trichocoma paradoxa isolates, (Table 1) were grown for 7 and 14 days at 37°C on the solid media YES, CYA, OA, malt extract agar (MEA-B) (Blakeslee formula, Difco malt extract) (Raper and Thom 1949, p. 67), MEA, Wickerham’s Antibiotic Test Medium (WATM) (Raper and Thom 1949, pp. 69–70) and Raulin-Thom (Smith 1960, p. 259) with added oat meal (RTO agar) (using 2.67 g ammonium tartrate and 30 g oatmeal/l) prior to extrolite extraction. Trichocoma paradoxa was grown on the above mentioned agar media for 4 weeks at 25°C. All media contained a trace metal mixture of 5 ppm CuSO4·5H2O and 10 ppm ZnSO4·7H2O (Smith 1949). Five agar plugs taken along a diameter of the fungal colony were used for extraction. The agar plugs were transferred in ethyl acetate/dichloromethane/methanol (3:2:1, v/v/v) with 1% (v/v) formic acid and ultrasonicated for 50 min. The extracts were transferred to a 1.5 ml autosampler screw-cap vials, evaporated to dryness and re-dissolved in 400 μl methanol by ultrasonication in 10 min. Subsequently, the extracts were filtered through 0.45 μm filter (Minisart RC4, Sartorius, Germany) and kept at −18°C prior to analysis. The extracts were analyzed by ultra high performance liquid chromatography (U-HPLC) using alkylphenone retention indices and diode array UV–VIS detection as described by Frisvad and Thrane (1987). The U-HPLC used was Dionex Ultimate 3000 with two pumps. The extrolites were separated on a Kinetex C18 (150 × 2.1 mm) column with 2.6 μm particles. The column temperature was held at 60°C. The extract injection volume was 1 μl. A gradient based on water with 50 ppm trifluoroacetic acid (TFA) and acetonitrile (AcCN) with 50 ppm TFA, at a flow rate of 0.8 ml/min, was used: Start conditions 85% H2O with TFA and 15% AcCN with TFA. Gradient: 15–25% AcCN in ½ min, from 25 to 65% AcCN in 5½ min, from 60 to 100% AcCN in 1 min, at 100% AcCH in 1 min, back to 15% AcCN in 1 min and then 1 min constant at 15% AcCN. The run-time was therefore 10 min. UV chromatograms were recorded at 210 and 280 nm, but diode array detection was carried out in a range from 190 to 600 nm. Identification of extrolites was performed by comparison of the UV–Visible spectra and retention times of the extrolites with those present in the collection at Department of Systems Biology, Kgs. Lyngby, Denmark.
Results
Generic delimitation
Macro- and microscopical analysis of Talaromyces emersonii, T. byssochlamydoides, T. eburneus, Geosmithia argillacea and G. cylindrospora showed that these species share various phenotypic and physiological characters. Shared phenotypic characters include condiophores with distinct rough-walled stipes and ellipsoidal, ovoid or cylindrical conidia, which are in shades of olive-brown. The ascomata of T. emersonii, T. byssochlamydoides, T. eburneus have a scanty covering and the ascospores are smooth walled, globose, subglobose or ovoidal. Furthermore, these species are thermophilic or thermotolerant and have optimal growth at 37°C or higher (see Supplementary Fig. 1).
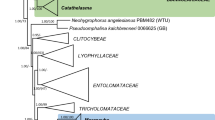
The phylogenetic position of T. emersonii, T. byssochlamydoides, G. argillacea and G. cylindrospora was studied using partial RPB2, Cct8 and Tsr1 sequences. The total length of the combined data sets was 2343 bp (exon data only). The structure of the partitions was as follows: the Cct8 partition was 726 bp long and had 368 variable sites; the Tsr1 data set included 639 characters, and 438 of those were variable and the RPB2 gene partition was 978 bp long, 502 were found to be variable. The SYM+I+G model was found to be optimal for the Tsr1 partition, whereas the best substitution model for the RPB2 and Cct8 sequence matrix was determined to be GTR+I+G. The thermophilic species T. emersonii and T. byssochlamydoides from section Emersonii and the thermotolerant species G. argillacea and G. cylindrospora group together, and the mesophile Trichocoma paradoxa is basal to these species. The two other species of section Emersonii, T. bacillisporus and T. leycettanus, are accommodated in other clades. Talaromyces bacillisporus belongs to the genus Talaromyces, and T. leycettanus groups with Hamigera avellanea and Warcupiella spinulosa. The Eurotialean Geosmithia species and related teleomorphs are polyphyletic (Fig. 1). G. argillacea and G. cylindrospora are closely related to T. emersonii and T. byssochlamydoides, G. namyslowskii is placed within Penicillium and Eupenicillium, and G. viridis and T. bacillisporus (anamorph G. swiftii) are accommodated in a clade comprising species mainly belonging to Penicillium subgenus Biverticillium and Talaromyces. Based on phenotypic, physiological and molecular data, we propose to transfer the species T. emersonii, T. byssochlamydoides, T. eburneus, G. argillacea and G. cylindrospora to Rasamsonia gen. nov.

Best-scoring Maximum Likelihood tree using RAxML based on combined data set of partial RPB2, Cct8 and Tsr1 sequences. The thermophiles are accommodated in three different clades: Rasamsonia, Thermomyces and Thermoascus. Rasamsonia species form a distinct clade in the Trichocomaceae and are closely related to Trichocoma. The bootstrap percentages of the RAxML analysis (first number) and BI Posterior Probabilities (second number) analysis are given at the nodes. Only bootstrap values above 70% or posterior probability values higher than 0.9 are shown. Branches supported less than 70% bootstrap value or 0.9 posterior probability are indicated with a hyphen and branches with 95% bootstrap or more are thickened. The tree is rooted with Coccidioides immitis (strain “RS”, Sharpton et al. 2009)
Species delimitation in Rasamsonia
Species belonging to the genus Rasamsonia can be distinguished by phenotype (Table 3). The optimum and maximum growth temperature on MEA, the growth rate on CYA and YES and the reverse color on MEA at 36°C proved to be valuable characters for differentiation. Supplementary Fig. 1 compares temperature growth relations among Rasamsonia species and with the phylogenetically related species Trichocoma paradoxa. Rasamsonia emersonii and R. byssochlamydoides are thermophilic and have an optimum growth temperature around 45°C. The other Rasamsonia species are thermotolerant and have optimal growth around 33°C (R. brevistipitata) or 36°C (R. argillacea, R. eburnea and R. cylindrospora). Trichocoma paradoxa is mesophilic and has optimum between 21 and 24°C and a maximum between 30 and 33°C. The grouping of thermotolerant and thermophilic Rasamsonia species, although not supported by high bootstrap values, is also reflected in the phylogeny: R. emersonii and R. byssochlamydoides form a clade, and the other four species form another cluster. Growth of Rasamsonia species on CYA is poor with no or weak sporulation except for R. argillacea and R. eburnea, which grow well producing colony diameters larger than 30 mm. Rasamsonia species also differ in micro-morphology. The size and shape of the conidia, branching pattern of the conidiophores and the formation of ascomata are useful characters for differentiation. Rasamsonia byssochlamydoides deviates from the other species in having irregularly branched conidiophores (Paecilomyces-type). Ascomata were observed only in cultures of R. emersonii and R. byssochlamydoides. Although ascomata and ascospores were reported in R. eburnea (Yaguchi et al. 1994), these were not detected in the present study after prolonged incubation of 8 weeks on OA and MEA at 33, 36 and 40°C. Rasamsonia argillacea and R. eburnea are phenotypically and phylogenetically closely related. The major phenotypic difference between these species is the blackish brown reverse of R. eburnea when grown on MEA at 36°C.
The results of the chemical analyses of the extracts of the different Rasamsonia species are listed in Table 3. Chemotaxonomically, the species of Rasamsonia can be divided into four groups: (1) the naphthalenone producers, R. emersonii and R. cylindrospora, which produce most extrolites on RTO agar, (2) the emodin, emodin anthron, and emodin bisanthron producer R. byssochlamydoides, which also produce most extrolites on RTO, (3) the producers of what was tentatively identified as Raistrick phenols and other small polar polyketides R. eburnea and R. argillacea, best produced on YES agar, and (4) R. brevistipitata, a producer of only a few extrolites including a compound tentatively identified as brefeldin A. Media with nitrate, CYA and WATM, were highly inhibitory to extrolite production. Trichocoma paradoxa IFM H-53524 (= DTO 78H2 = CBS 788.83) and CBS 103.73 (= IBT 31160) were also analyzed for extrolites and found to produce rugulovasine A, corymbiferone (Overy et al. 2005a, b), corymbiferane lactones (Overy and Blunt 2004), and compounds related to naphthalic anhydride, atrovenetin and funalenone (Turner 1971), in addition to several unique extrolites. The latter three chromophore families were also detected in R. cylindrospora and R. emersonii. One of the corymbiferan lactones was also found in R. byssochlamydoides.
In order to study the phylogenetic relationship among Rasamsonia species, the ITS regions and partial calmodulin and tubulin sequences were combined and the total length of this data set was 1,766 nucleotides. The length of each partition was 706, 553 and 507 nucleotides for the ITS regions, and the partial calmodulin and tubulin gene regions, respectively, and number of variable sites were 158, 240 and 206 nucleotides, respectively. The SYM+G model was found to be optimal for the calmodulin partition, GTR+I+G was the best model for ITS, whereas the best substitution model for the tubulin matrix was determined to be K80 + I. Multilocus sequence analysis of the genus Rasamsonia revealed the presence of six species (Fig. 2). Five new combinations are proposed: R. eburnea, R. argillacea, R. cylindrospora, R. emersonii and R. byssochlamydoides, and one new species is described. We propose the name R. brevistipitata sp. nov. for this species. The phylogram supports the sibling relationship of R. eburnea and R. argillacea, however, the relationships between the other species are poorly or not supported (<95% bs).

Best-scoring Maximum Likelihood tree using RAxML based on combined data set of ITS, and partial β-tubulin and calmodulin sequences. Both Rasamsonia brevistipitata strains are on a single branch and positioned within the Rasamsonia clade. The bootstrap percentages of the RAxML analysis (first number) and BI Posterior Probabilities (second number) analysis are given at the nodes. Only bootstrap values above 70% or posterior probability values higher than 0.9 are shown
Taxonomy
Rasamsonia Houbraken & Frisvad gen. nov. Mycobank MB 519868.
Latin diagnosis: Habitatione thermophila et thermotoleranti, culturis circa vel ultra 37°C optime crescentibus. Ascomatibus parce obtectis, ascosporis laevibus, globosis vel ovoideis, stipitibus conidiophorum et/vel hyphis sustinentibus distincte exasperates. Conidiis ellipsoideis, ovoideis vel cylindraceis, diverse olivaceo-brunneis.
Typus: Rasamsonia emersonii (Stolk) Houbraken & Frisvad comb. nov.
Etymology: Named in honor of Robert A. Samson, for his excellent work in fungal taxonomy, and marking his 65th birthday and 40 years of service at the CBS-KNAW Fungal Biodiversity Centre.
Diagnostic features: Ascomata, if present, with a scanty covering, ascospores smooth walled, globose, subglobose or ovoidal. Stipes of conidiophores often distinctly rough walled, conidia ellipsoidal, ovoid or cylindrical, in shades of olive-brown. Thermophilic or thermotolerant, optimum around 37°C or higher.
Notes: Rasamsonia phenotypically resembles Paecilomyces and its teleomorph Byssochlamys and is placed within the order Eurotiales, family Trichocomaceae.
Byssochlamys/Paecilomyces and Rasamsonia both contain thermotolerant species, produce olive-brown conidia and form ascomata with no or scarce ascomatal covering. Rasamsonia differs in having regularly branched conidiophores with distinct rough walled structures and asci are borne in chains, while the conidiophores of Paecilomyces are irregularly branched, smooth walled and asci of Byssochlamys are born from croziers. Furthermore, the phialides of most Rasamsonia species (except R. byssochlamydoides) are cylindrical and taper gradually towards the apices, while the phialides of Paecilomyces have a broad base tapering into a long narrow neck (Paecilomyces-type). The monotypic genus Trichocoma is phylogenetically closely related to Rasamsonia (Peterson 2008, this study). Trichocoma is characterized by asci borne in hyphal masses or tufts, which measure up to 10–20 mm in length (Kominami et al. 1952; Malloch 1985), while the covering of the ascomata of Rasamsonia are scanty. Furthermore, Trichocoma is a mesophile and has a maximum growth temperature of 33°C, while members of Rasamsonia have a maximum above 45°C (see Supplementary Fig. 1). There are also similarities between both genera: both share formation of bi- and terverticillate Penicillium-like structures, conidia are in shades of brown en masse, and both form rough walled hyphae.
Recently, the proposal to revise article 59 of the International Code of Botanical Nomenclature (ICBN) was accepted at the 2011 IBC Nomenclature Section at Melbourne and the principle of ‘one fungus : one name’ was established (Norvell 2011). In light of this decision, we propose a single name nomenclature for Rasamsonia.
Rasamsonia argillacea (Stolk, Evans & Nilsson) Houbraken & Frisvad comb. nov. Mycobank MB 519878. Supplementary Fig. 2.
Basionym: Penicillium argillaceum Stolk, Evans and Nilsson, Trans. Br. Mycol. Soc. 53: 307. 1969.
= Geosmithia argillacea (Stolk, Evans and Nilsson) Pitt, Can. J. Bot. 57: 2026. 1979.
Description: Colony diameters, 7 days, in mm., 25°C: CYA 15–25; CYAS 0–2; MEA 15–25; YES 15–24; DG18 6–12; OA 18–27; CREA: no growth. Colony diameters, 37°C: CYA 30–40; CYAS 9–10; MEA 30–40; YES 35–46; DG18 13–22; OA 36–47; CREA: 14–21, poor growth, no acid and no base production. Optimum growth temperature 36°C, maximum above 50°C.
Good sporulation on CYA at 25 and 37°C, velvety, olive brown or crème-brown conidia, exudates and soluble pigments absent, colonies not or weakly sulcate, reverse pale brown with brown centre at 25°C and crème-brown at 37°C. Good sporulation on YES at 25°C, weak sporulation at 37°C, soluble pigments absent, mycelium inconspicuous, white, olive brown conidia, reverse brown. Colonies on MEA velvety to floccose, olive brown conidia, reverse light brown, occasionally with darker brown centre.
Stipes terminating in bi- or terverticillate structures, 60–350 × 2.0–3.5 μm, verrucose; branches subterminally present and at lower levels of the stipe, monoverticillate and short, suggesting the presence of monoverticillate conidiophores. Metulae in terminal whorls of 2–5, unequal in length, 10–15 (–30) × 1.5–2.5 μm. Phialides, 5–8 (–12) per metula, cylindrical base tapering more or less abruptly to a narrowed collula, 10–14 (–18) × 1.5–2.5 μm. Conidia smooth walled, cylindrical or ovoid, (3–) 3.5–4.5 (–5.0) × 1.5–2.0 (–2.3) μm.
Extrolites: Production better on YES agar than RTO, poor on other media. No extrolites could be identified, but among the chromophore families were several unidentified compounds with characteristic UV spectra (Table 3). Strain IBT 31197 (= DTO 137A4) was the best producer of extrolites, while CBS 102.69 and IBT 31196 (= DTO 137C1) produced few and CBS 101.69 produced none.
Diagnostic features: Thermotolerant, good growth on CYA (30–40 mm) and YES (35–46 mm) at 37°C, reverse on MEA uncolored or light brown, conidia cylindrical or ovoid and measuring (3–) 3.5–4.5 (–5.0) × 1.5–2.0 (–2.3) μm.
Distribution and ecology: Rasamsonia argillacea is a commonly occurring species and was isolated from hot environments, such as mine tips with a very high surface temperature, (indoor) air, clinical specimens and heat treated fruit concentrates. The presence of this species in heat treated food products suggests the potential presence of a teleomorph. Recently, this species was described as the cause of invasive mycosis in patients with chronic granulomatous disease (CGD, De Ravin et al. 2011; Machouart et al. 2011). Furthermore, this emerging pathogen was reported as a chronic colonist of airways of cystic fibrosis patients (Giraud et al. 2010; Barton et al. 2010). Rasamsonia argillacea might be more common than reported in literature. Retrospective identification showed that all stored P. variotii strains from patient with CGD were R. argillacea and similar misidentifications also occur from non-CGD patients (Houbraken et al. 2010; De Ravin et al. 2011).
Notes: After comparison of the D1/D2 region of the 28S rDNA, Yaguchi et al. (2005) concluded that R. eburnea (as T. eburneus) is the teleomorph of R. argillacea (as P. argillacea). However, both species are distinct based on phenotype and sequences (see “Results” section).
Rasamsonia brevistipitata Weidner, Houbraken & Frisvad sp. nov. Mycobank MB 519870 (Fig. 3).

Rasamsonia brevistipitata. a–h Colonies 7 days old 37°C: a MEA, b CYA, c YES, d OA, e MEA reverse, f CYA reverse, g YES reverse, h CREA; i–m conidiophores; n conidia. Scale bar 10 μm
Latin diagnosis: Habitatione thermotoleranti, culturis restricte crescentibus in CYA (11–17 mm) et YES (22–30 mm) ad 37°C, coloniis reversis incoloratis vel dilute brunneis in agaro MEA, stipitibus brevibus, conidiis ellipsoideis vel ovoideis, (2.0–) 2.5–3.0 (3.5) × 1.7–2.1 μm.
Etymology: Rasamsonia brevistipitata is referring to short stipes formed by this species.
Typus: Germany, Balingen: isolated from water damaged floor composed of bitumen and cork in a school, 2006, U. Weidner (holotype: CBS H-20546; culture ex type—CBS 128785 = DTO 25H2 = IBT 31187).
Description: Colony diameters, 7 days, in mm, 25°C: CYA 7–13; CYAS 0.5–2; MEA 17–25; YES 11–18; DG18 5–10; OA 20–28; CREA: 2–7, poor growth, no acid and no base production. Colony diameters at 37°C: CYA 11–17; CYAS 0; MEA 24–30; YES 22–30; DG18 8–14; OA 28–32; CREA: 2–7, poor growth, no acid and no base production. Optimum growth temperature 33°C; maximum slightly above 45°C.
Colonies on CYA at 25 and 37°C restricted, no or poor sporulation, velvety, pale olive brown conidia, crème colored mycelium, exudates and soluble pigments absent, colonies not sulcate, reverse crème or light brown. Moderate to good sporulation on YES at 25°C and 37°C, obverse pale olive brown, reverse light brown with dark brown centre. Colonies on MEA velvety, conidia olive brown, reverse uncolored or light brown.
Conidiophores predominantly arising from aerial mycelium, occasionally arising from basal mycelium; stipes terminating in monoverticillate structures when young and becoming biverticillate in older parts of the colony, (50–) 100–250 × 2.0–3.5 μm, branches on subterminally and intercalary positions, suggesting separate conidiophores, short (10–50 (–70) × 2.0–3.5 μm) and often monoverticillate, verrucose. Metulae, when present, in terminal whorls of 2–3 (–5), vesiculate, 12–17 (–20) μm. Phialides Paecilomyces-type, in appressed verticils of 4–10, outer phialides subapically placed on metula or stipe, cylindrical base slowly tapering in a long neck, 10–14 μm. Conidia smooth walled, ellipsoidal or ovoid, (2.0–) 2.5–3.0 (3.5) × 1.7–2.1 μm. Ascomata and sclerotia absent.
Extrolites: Production best on YES and MEA; both isolates produced an extrolite tentatively identified as brefeldin A.
Diagnostic features: Thermotolerant, restricted growth on CYA (11–17 mm) and YES (22–30 mm) at 37°C, reverse on MEA uncolored or light brown, conidia ellipsoidal or ovoid and measuring (2.0–) 2.5–3.0 (3.5) × 1.7–2.1 μm.
Distribution and ecology: This species was isolated from indoor environment in Germany. A BLAST search on GenBank with the ITS sequence of CBS 128785 showed a 100% homology with GenBank accession number FJ820827, a sequence generated during DNA analysis of outdoor air in Mainz, Germany (Fröhlich-Nowoisky et al. 2009).
Rasamsonia byssochlamydoides (Stolk & Samson) Houbraken & Frisvad comb. nov. MB 519877. Supplementary Fig. 3.
Basiomym: Talaromyces byssochlamydoides Stolk & Samson, Stud. Mycol. 2: 45. 1972.
= Paecilomyces byssochlamydoides Stolk & Samson, Stud. Mycol. 2: 45. 1972.
Description: Colony diameters, 7 days, in mm, no growth at 25°C; 37°C: CYA 19–27; CYAS 0; MEA >60; YES 22–52; DG18 20–30; OA >60; CREA: 0–7, no or poor growth, no acid and no base production. Optimum growth temperature between 40 and 45°C, maximum above 50°C.
Poor growth on CYA at 37°C, sporulation, exudate and soluble pigments absent, colonies not sulcate, mycelium inconspicuous and white, reverse pale or pale with a brown centre. Sporulation on YES at 37°C weak or absent, soluble pigments absent, mycelium inconspicuous, obverse olive-brown when sporulation is present, reverse light brown and not influencing the medium color, occasionally with dark brown centre. Colonies on MEA spreading, velvety or with floccose mycelial overgrowth, olive brown conidia, thin layer of ascomata formed near the agar, white when young, orange or pink-reddish in age, reverse uncolored or brown.
Ascomata globose, up to 200 μm in diameter, covering very scanty. Asci eight-spored, subglobose to broadly ellipsoidal, 9–12.5 × 6.5–7.5 μm. Ascospores globose to subglobose, smooth, 3.5–4.5 × 4.0–5.5 μm. Conidiophores arising directly from the agar or in the floccose mycelium, irregularly branched (Paecilomyces-type), terverticillate or quarterverticillate, up to 300 μm in length; stipes broad, 3–6 μm, smooth to verrucose. Metulae in terminal whorls of 2–8, broad, unequal in length, (7) 10–15 × 2.0–5.0 μm. Phialides, cylindrical base tapering in a long narrowed collula, 10–15 (–20) × 2.5–3.5 μm. Conidia smooth walled, cylindrical, 4–8 × 1–2.5 μm. Chlamydospores are rarely produced, globose or subglobose, about 4 μm in diameter (Stolk and Samson 1972).
Extrolites: Production best on RTO, followed by MEA-B, MEA and OAT. Poor on remaining media, except the production of an indole-alkaloid on YES agar by CBS 533.71. All examined strains (CBS 533.71 and CBS 413.71) produced two chromophore families of extrolites (Table 3) each with 4–7 different extrolites. In addition, CBS 533.71 produced extrolites with UV spectra similar to Raistrick phenols, and emodin, emodin anthrone and emodin bisanthrone and emodin bisanthron. This series of anthraquinones and anthrones has also been found to co-occur in Aspergillus wentii (Rabie et al. 1986).
Diagnostic features: Thermophilic, irregularly branched conidiophores (Paecilomyces-type), ascomata produced and ripening within 7–10 days at 40°C, poor to moderate growth on CYA and YES agar.
Distribution and ecology: Rasamsonia byssochlamydoides was isolated from dry soil under Douglas fir (Oregon, USA) and piles of peat (Oldenburg, Germany).
Rasamsonia cylindrospora (G. Smith) Houbraken & Frisvad comb. nov. Mycobank MB 519876. Supplementary Fig. 4.
Basionym: Penicillium cylindrosporum G. Smith—Trans. Br. Mycol. Soc. 40: 483, 1957
= Geosmithia cylindrospora (G. Smith) Pitt—Can. J. Bot. 57(19): 2024, 1979
Description: Colony diameters, 7 days, in mm. 25°C: CYA 3–8; CYAS 0; MEA 10–20; YES 5–15; DG18 5–15; OA 9–17; CREA: no growth. Colony diameters at 37°C: CYA 5–10; CYAS 0; MEA 27–38; YES 10–19; DG18 15–26; OA 24–34; CREA: 1–5, poor growth, no acid and no base production. Optimum growth temperature 36°C, maximum around 50°C.
Colonies on CYA at 25 and 37°C restricted, velvety, olive brown conidia, exudates and soluble pigments absent, no sulcations in colony, reverse in shades of brown. Good sporulation on YES, soluble pigments absent, mycelium inconspicuous, white, olive brown conidia, reverse brown with dark brown centre. Colonies on MEA velvety to floccose, olive brown conidia, distinct blackish brown reverse at 37°C.
Conidiophores predominantly arising from basal mycelium, predominantly biverticillate, occasionally terverticillate or simple, additional branch both direct under terminal whorl and further down the stipe, (50–) 100–200 × 2.0–3.5 μm, stipes verrucose, Metulae in terminal whorls of 2–4, unequal in length, 10–17 (–25) × 2.5–3.5 μm. Phialides, 5–8 (–12) per metula, cylindrical, with distinct neck, 10–14 × 2.0–2.5 μm. Conidia smooth walled, cylindrical, 4.0–5.0 × 1.6–2.1 μm.
Extrolites: Production best on RTO, followed by MEA, MEA-B and OA; production on remaining media poor. CBS 275.58 produced only one extrolite on MEA, which was not found in CBS 433.62. CBS 433.62 produced a large number (14) of naphthalenes, including some with UV spectra similar to “naphthalic anhydride”, atrovenetin and funalenone (Turner 1971, pp. 150–153; Inokoshi et al. 1999).
Diagnostic features: Thermotolerant, restricted growth on CYA (5–10 mm) and YES (10–19 mm) at 37°C, blackish brown reverse on MEA, conidia cylindrical and measuring 4.0–5.0 × 1.6–2.1 μm.
Distribution and ecology: Two R. cylindrospora isolates are currently present in the CBS culture collection. One strain was isolated from sputum in the Netherlands; the other as a culture contaminant in England, UK.
Notes: Rasamsonia cylindrospora was originally described as Penicillium cylindrosporum by Smith (1957) and this species was later transferred to the genus Geosmithia (Pitt 1979).
Rasamsonia eburnea (Yaguchi, Someya & Udagawa) Houbraken & Frisvad comb. nov. Mycobank MB 519875. Supplementary Fig. 5.
Basionym: Talaromyces eburneus Yaguchi, Someya & Udagawa Mycoscience 35: 249, 1994
= Geosmithia eburnea Yaguchi, Someya & Udagawa Mycoscience 35: 249, 1994
Description: Colony diameters, 7 days, in mm. 25°C: CYA 14–20; CYAS 1–3; MEA 10–20; YES 14–20; DG18 5–10; OA 23–30; CREA: 5–10, poor growth, no acid and base production. Colony diameters at 37°C: CYA 30–40; CYAS1–6; MEA 34–42; YES 48–55; DG18 10–20; OA 36–46; CREA: 12–18, poor growth, no acid and no base production. Optimum growth temperature 36°C, maximum above 50°C.
Colonies on CYA at 37°C spreading, velvety, olive-brown conidia, mycelium cream crème colored, exudates and soluble pigments absent, colonies not sulcate at 25°C, and distinct sulcate at 37°C, reverse (pale) brown. Good sporulation on YES at 37°C, obverse pale olive-brown, mycelium inconspicuous, reverse pale brown and becoming blackish brown in age. Colonies on MEA velvety, olive-brown conidia, reverse distinct dark brown at 37°C (at 25°C often paler brown).
Conidiophores predominantly arising from basal mycelium, young structures monoverticillate, biverticillate in older parts, (50–) 100–350 × 2.0–3.5 μm, occasionally additional branches formed subterminally or at lower levels, suggesting presence of separate conidiophores and these structures are shorter (20–70 μm), stipes verrucose. Metulae, when present, in terminal whorls of 2–3 (–5), unequal in length, 10–20 × 2.5–3.5 μm. Phialides, in appressed verticils of 2–6, cylindrical, 10–14 (–17) μm. Conidia smooth walled, cylindrical at first and later becoming ellipsoidal or ovoid, 2.5–3.5 (–4) × 1.8–2.5 μm. Ascomata not detected in this study, but reported in literature (Yaguchi et al. 1994) as discrete, pale yellow, maturing slowly within 28–35 days, globose to subglobose, (30–) 70–125 μm, soft, scarcely covered by hyaline, delicate mycelium. Asci eight-spored, borne singly, subglobose to ovoid or pyriform, 10.5–13 (15) × 8–9.5 (–11) μm. Ascospores pale yellow, subglobose to more or less ovoid, 4–5 × 4–4.5 μm, thick walled, smooth but occasionally with foveolations, with an equatorial thickening under SEM.
Extrolites: Production best on YES, followed by RTO and MEA, none on other media. CBS 102881 and CBS 124445 (= IBT 31193) were the best producers of extrolites (Table 3).
Diagnostic features: Thermotolerant, good growth on CYA (30–40 mm) and YES (48–55 mm) at 37°C, blackish brown reverse on MEA at 37°C, conidia cylindrical at first and becoming ellipsoidal or ovoid.
Distribution and ecology: The type strain of Rasamsonia eburnea was isolated from Taiwanese soil. Other strains are isolated from clinical specimens, including blood culture, bronchial washing and dialysis fluid. The studies of Houbraken et al. (2010) and Barton et al. (2010) mainly used the same Rasamsonia strains (as Geosmithia). Strains NCPF 7594 and 7596 were isolated from the same patient (respectively from blood culture and peritoneal dialysis fluid) and re-identified here, based on deposited ITS sequences in GenBank, as R. eburnea. An isolate originating from a blood culture (CBS 124447; pseudo-outbreak, UK) was also re-identified as R. eburnea (Houbraken et al. 2010).
Notes: De Ravin et al. (2011) mention that R. eburnea is heterothallic and fails to form a teleomorph without mating appropriate strains. However, no additional details are given for this observation in their publication.
Rasamsonia emersonii (Stolk) Houbraken & Frisvad comb. nov. Mycobank MB 519874. Supplementary Fig. 6.
Basionym: Talaromyces emersonii Stolk, Antonie van Leeuwenhoek 31: 262. 1965.
= Penicillium emersonii Stolk, Antonie van Leeuwenhoek 31: 262. 1965.
Description: Colony diameters, 7 days, in mm., no growth at 25°C; 37°C: CYA 18–30; CYAS 0; MEA >50; YES 16–45; DG18 10–22; OA >60; CREA: 0–5, no or poor growth, no acid and no base production. Optimum growth temperature between 45 and 50°C, maximum above 50°C.
Poor to moderate growth on CYA at 37°C, no or poor sporulation, exudates and soluble pigments absent, colonies not sulcate, mycelium inconspicuous and white, reverse pale to light brown. Sporulation on YES at 37°C weak or absent, soluble pigments absent, mycelium inconspicuous, if sporulation is present, then colony observe pale olive-brown, reverse light brown and not influencing the medium color. Colonies on MEA spreading, velvety to floccose, olive brown conidia; ascomata present and often covered under layer of conidiophores, white when young, becoming yellow-brown or orange-brown within 7–10 days, reverse light brown, occasionally dark brown (CBS 549.92).
Ascomata reddish to orange brown, globose to subglobose, up to 300 μm in diameter; covering scanty. Asci eight-spored, subglobose or broadly ellipsoidal, 8–10.5 × 6.5–8 μm. Ascospores smooth, subglobose to broadly ellipsoidal, 3.5–4 × 2.5–3.5 μm. Conidiophores arising directly from the agar, regularly branched, bi- or terverticillate, 50–150 μm, stipes verrucose, width 2.0–3.5 μm. Metulae in terminal whorls of 3–5, equal in length, 10–15 (–20) × 2.0–3.0 μm. Phialides, 5–10 per metula, cylindrical with a short collula, 8.5–10 × 2–2.5 μm. Conidia smooth walled, cylindrical, 3.5–4.5 (–5.0) × 1.5–3.0 μm.
Extrolites: Production best on OAT, followed by RTO, MEA-B and MEA; no production on other media. CBS 549.92 is the best extrolite-producer followed by CBS 396.64 and CBS 397.64. Weak extrolite production by the ex-type strain CBS 393.64, which produced one naphthalenone and secalonic acid D. CBS 549.92 produced a large number (10) of naphthalenones, including some with UV spectra similar to naphthalic anhydride, atrovenetin and funalenone (Turner 1971, pp. 150–153; Inokoshi et al. 1999) and and other as yet unknown extrolites (Table 3). Strain CBS 549.92 differed from the other strains in additionally producing an indole alkaloid, and an isocoumarin and two partially characterized extrolites (Table 3). It also produced more naphthalenones than the other strains.
Diagnostic features: Thermophilic, regularly branched conidiophores with olive-brown colored conidia, ascomata present and ripening within 7–10 days at 37°C, poor to moderate growth on CYA and YES agar.
Distribution and ecology: Rasamsonia emersonii is a relatively common fungus, but is probably overlooked since it is not able to grow at 25°C. This species was isolated from hot environments such as compost, but also from soil and outdoor air (Fröhlich-Nowoisky et al. 2009; Hultman et al. 2009). Rasamsonia emersonii was also reported to be the causal agent of a chronic airway colonization. Reexamination of the deposited ITS sequences showed that these strains are more close to R. argillacea (Cimon et al. 1999; Giraud et al. 2010).
Notes: The results of the multigene analysis shows that CBS 549.92 (= CBS 814.70) deviates from other strains of R. emersonii. This strain is also phenotypically different and has intensely bright yellow colored mycelium, but this could not be related to any extrolites in the extract of the fungus. The isolate may represent a new species, and more strains should be collected and examined to determine its taxonomic status.
Discussion
The genus Rasamsonia encompasses one new species, three species formally accommodated in Talaromyces and two species previously classified in Geosmithia. Phylogenetic analyses showed that the genus Geosmithia is polyphyletic and forms lineages in the Hypocreales and Eurotiales (Ogawa et al. 1997; Ogawa and Sugiyama 2000; this study). Geosmithia lavendula, the type species of Geosmithia, is accommodated in the Hypocreales and is related to Acremonium alternatum, the type species of Acremonium (Ogawa et al. 1997; Rossman and Seifert 2011). Several new Geosmithia species belonging to the Hypocreales were described in the last decade and the majority of these species are associated with galleries of phloem feeding bark beetles (Kolařík et al. 2004, 2005, 2010). Four of eight Geosmithia species belonging to the Eurotiales are reclassified in Rasamsonia. Geosmithia swiftii (teleomorph Talaromyces bacillisporus) and G. viridis belong to a clade comprising mainly Talaromyces species and members of Penicillium subgenus Biverticillium (Fig. 1), and G. namyslowskii and G. malachitea (teleomorph Chromocleista malachitea) belong to Penicillium s. str. (Ogawa et al. 1997). The transfer of these species to other genera and making Geosmithia a monophyletic genus will be performed in another study. Our results show that the members of the Talaromyces section Emersonii, as defined by Stolk and Samson (1972), belong to different genera. Two species are disposed in Rasamsonia, but T. bacillisporus and T. leycettanus are retained in Talaromyces. Talaromyces bacillisporus is related to Talaromyces flavus, which is the type species of Talaromyces. Stolk and Samson (1972), who showed that the ascomatal coverings of T. bacillisporus are much better developed than those of R. emersonii and R. byssochlamydoides, already had doubts about the relationship between T. bacillisporus and R. emersonii. They suggest that T. bacillisporus resembles an intermediate form between the section Emersonii and section Talaromyces. Pitt (1979) placed the anamorph of T. bacillisporus in Geosmithia (as G. swiftii), suggesting a close relationship with R. argillacea and R. cylindrospora. However, the conidia of T. bacillisporus are cream coloured not olive-brown. Phylogenetic analysis shows that T. leycettanus is not related to Talaromyces but rather to Hamigera and Warcupiella and this confirms earlier findings (Peterson 2008).
Extrolite analysis showed that R. emersonii, R. cylindrospora and R. byssochlamydoides produced naphthalenones or anthraquinones. Many species in Talaromyces sensu stricto also produce naphthalenones and anthraquinones (Frisvad et al. 1990), and these metabolites could act ecologically as antioxidants and thermoprotectants. It seems that these type of compounds have been present in a common ancestor of Talaromyces and Rasamsonia. Rasamsonia eburnea and R. argillacea produced extrolites not seen in any other fungal species yet. Trichocoma paradoxa is phylogenetically related to Rasamsonia and this species produces species specific extrolites, but also naphthalenones, which indicates a relationship with R. cylindrospora and R. emersonii. However, naphthalenones are found in T. macrosporus and P. duclauxii (Frisvad et al. 1990) and also in the phylogenetically more distant species Penicillium atrovenetum, P. raistrickii, P. hordei and P. hirsutum (Turner 1971; Overy et al. 2005a, b) and so these antioxidant metabolites may have evolved more than once.
The Eurotialean genera Talaromyces, Thermoascus, Thermomyces, Coonemeria and Dactylomyces contain thermophilic species (Apinis 1967; Mouchacca 1997; Stolk and Samson 1972; Stolk 1965). The current study shows that no thermophiles are accommodated in Talaromyces. Talaromyces emersonii and T. byssochlamydoides are combined in Rasamsonia and Talaromyces thermophilus is phylogenetically closely related to the type species of Thermomyces, T. lanuginosus, and not to the type of Talaromyces, T. flavus. Talaromyces thermophilus and T. lanuginosus share similar features including thermophilicity, thick-walled chlamydospores or chlamydospore-like conidia. Further research is needed to clarify the taxonomy of Thermomyces and the relationship between T. thermophilus and the other species in Thermomyces. Various taxonomical studies of the thermophilic genus Thermoascus were performed (Apinis 1967; Mouchacca 1997; Pitt et al. 2000; Stolk 1965). Apinis (1967) split this genus in two: Thermoascus was retained for its type species T. aurantiacus, and T. thermophilus and T. crustaceus were transferred to Dactylomyces. Based on anamorph differences, Mouchacca (1997) divided Dactylomyces further in two, creating the genus Coonemeria for T. crustaceus. Although these species have different anamorphs (Paecilomyces/Polypaecilum), our phylogenetic study (Fig. 1) shows that these three species are related and should be retained in the genus Thermoascus. In conclusion, our data shows that eurotialian thermophiles are not present in Talaromyces s. str. but only in Thermomyces, Thermoascus and Rasamsonia.
References
Aguileta G, Marthey S, Chiapello H, Lebrun MH, Rodolphe F, Fournier E, Gendrault-Jacquemard A, Giraud T (2008) Assessing the performance of single-copy genes for recovering robust phylogenies. Syst Biol 57:613–627
Apinis AE (1963) Occurrence of thermophilous microfungi in certain alluvial soils near Nottingham. Nova Hedwigia 5:57–78
Apinis AE (1967) Dactylomyces and Thermoascus. Trans Br Mycol Soc 50:573–582
Barton RC, Borman AM, Johnson EM, Houbraken J, Hobson RP, Denton M, Conway SP, Brownlee KG, Peckham D, Lee TW (2010) Isolation of the fungus Geosmithia argillacea in sputum of people with cystic fibrosis. J Clin Microbiol 178(48):2615–2617
Cimon B, Carrere J, Chazalette JP, Vinatier JF, Chabasse D, Bouchara JP (1999) Chronic airway colonization by Penicillium emersonii in a patient with cystic fibrosis. Med Mycol 37:291–293
Collins CM, Murray PG, Denman S, Morrissey JP, Byrnes L, Teeri TT, Tuohy MG (2007) Molecular cloning and expression analysis of two distinct beta-glucosidase genes, bg1 and aven1, with very different biological roles from the thermophilic, saprophytic fungus Talaromyces emersonii. Mycol Res 111:840–849
Cooney DG, Emerson R (1964) Thermophilic fungi. W.H. Freeman and Co., San Francisco
Coughlan MP, Tuohy MG, Filho EXF, Puls J, Claeyssens M, Vrsanska M, Hughes MM (1993) Enymological aspects of microbial hemicellulases with emphasis on fungal systems. In: Coughlan MP, Hazlewood CP (eds) Hemicullulose and hemicellulases. Portland Press, London, pp 53–58
Craveri R, Manachini PL, Craveri A (1964) Eumiceti termofili presenti nel suolo. Ann Microbiol Enzimol 14:13–26
Crisan EV (1973) Current concepts of thermophilism and the thermophilic fungi. Mycologia 65:1171–1198
de Hoog GS, Gerrits van den Ende AHG (1998) Molecular diagnostics of clinical strains of filamentous Basidiomycetes. Mycoses 41:183–189
De Ravin SS, Challipalli M, Anderson V, Shea YR, Marciano B, Hilligoss D, Marquesen M, Decastro R, Liu YC, Sutton DA, Wickes BL, Kammeyer PL, Sigler L, Sullivan K, Kang EM, Malech HL, Holland SM, Zelazny AM (2011) Geosmithia argillacea: an emerging cause of invasive mycosis in human chronic granulomatous disease. Clin Infect Dis 52:e136–e143
Evans HC (1971) Thermophilous fungi of coal spoil tips. II. Occurrence, distribution and temperature relationships. Trans Br Mycol Soc 57:255–266
Evans HC, Stolk AC (1971) Talaromyces leycettanus sp. nov. Trans Br Mycol Soc 56:45–49
Fedorova ND, Khaldi N, Joardar VS et al (2008) Genomic islands in the pathogenic filamentous fungus Aspergillus fumigatus. PLoS Genet 4:e1000046
Frisvad JC, Thrane U (1987) Standardized high-performance liquid chromatography of 182 mycotoxins and other fungal metabolites based on alkylphenone retention indices and UV-VIS spectra (diode array detection). J Chromatogr 404:195–214
Frisvad JC, Filtenborg O, Samson RA, Stolk AC (1990) Chemotaxonomy of the genus Talaromyces. Antonie van Leeuwenhoek 57:179–189
Fröhlich-Nowoisky J, Pickersgill DA, Després VR, Pöschl U (2009) High diversity of fungi in air particulate matter. Proc Natl Acad Sci USA 106:12814–12819
Galagan JE, Calvo SE, Cuomo C et al (2005) Sequencing of Aspergillus nidulans and comparative analysis with A. fumigatus and A. oryzae. Nature 438:1105–1115
Giraud S, Pihet M, Razafimandimby B, Carrère J, Degand N, Mely L, Favennec L, Dannaoui E, Bouchara J-P, Calenda A (2010) Geosmithia argillacea: an emerging pathogen in cystic fibrosis patients? J Clin Microbiol 48:2381–2386
Glass NL, Donaldson GC (1995) Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl Environ Microbiol 61:1323–1330
Hong SB, Cho HS, Shin HD, Frisvad JC, Samson RA (2006) Novel Neosartorya species isolated from soil in Korea. Int J Syst Evol Microbiol 56:477–486
Houbraken J, Verweij PE, Rijs AJ, Borman AM, Samson RA (2010) Identification of Paecilomyces variotii in clinical samples and settings. J Clin Microbiol 48:2754–2761
Huelsenbeck JP, Ronquist F (2001) MrBayes: Bayesian inference of phylogenetic trees. Bioinformatics 17:754–755
Hultman J, Vasara T, Partanen P, Kurola J, Kontro MH, Paulin L, Auvinen P, Romantschuk M (2009) Determination of fungal succession during municipal solid waste composting using a cloning-based analysis. J Appl Microbiol 108:472–487
Inokoshi J, Shiomi K, Masuma R, Tanaka H, Yamada H, Ōmura S (1999) Funalenone, a novel collagenase inhibitor produced by Aspergillus niger. J Antibiot 52:1095–1100
Kolařík M, Kubátová A, Pažoutová S, Šrůtka P (2004) Morphological and molecular characterisation of Geosmithia putterillii, G. pallida comb. nov. and G. flava sp. nov., associated with subcorticolous insects. Mycol Res 108:1053–1069
Kolařík M, Kubátová A, Čepička I, Pažoutová S, Šrůtka P (2005) A complex of three new white-spored, sympatric, and host range limited Geosmithia species. Mycol Res 109:1323–1336
Kolařík M, Freeland E, Utley C, Tisserat N (2010) Geosmithia morbida sp. nov., a new phytopathogenic species living in symbiosis with the walnut twig beetle (Pityophthorus juglandis) on Juglans in USA. Mycologia. doi:10.3852/10-124
Kominami K, Kobayasi Y, Tubaki K (1952) Is Trichocoma paradoxa conspecific with Penicillium luteum? Nagaoa 2:18–23
Liu Y, Whelen S, Hall BD (1999) Phylogenetic relationships among ascomycetes: evidence from an RNA polymerase II subunit. Mol Biol Evol 16:1799–1808
Machouart M, Garcia-Hermoso D, Rivier A, Hassouni N, Catherinot E, Salmon A, Debourgogne A, Coignard H, Lecuit M, Bougnoux M-E, Blanche S, Lortholary O (2011) Emergence of disseminated infections due to Geosmithia argillacea in patients with chronic granulomatous disease receiving long-term azole antifungal prophylaxis. J Clin Microbiol. doi:10.1128/JCM.02456-10
Maheshwari R, Bharadwaj G, Bhat MK (2000) Thermophilic fungi: their physiology and enzymes. Microbiol Mol Biol Rev 64:461–488
Malloch D (1985) The Trichocomaceae: relationships with other Ascomycetes. In: Samson RA, Pitt JI (eds) Advances in Penicillium and Aspergillus systematic. Plenum Press, New York, pp 365–382
Masclaux F, Guého H, de Hoog GS, Christen R (1995) Phylogenetic relationships of human-pathogenic Cladosporium (Xylohypha) species inferred from partial LSU rRNA sequences. J Med Vet Mycol 33:327–338
McHale A, Coughlan MP (1981) The components of the cullulase system of Talaromyces emersonii with emphasis on β-glucosidase. Biochim Biophys Acta 662:145–159
Mouchacca J (1997) Thermophilic fungi: biodiversity and taxonomic status. Crypt Mycol 18:19–69
Murray P, Aro N, Collins C, Grassick A, Panttilä M, Saloheimo M, Tuohy M (2004) Expression in Trichoderma reesei and characterization of a thermostable family 3 beta-glucosidase from the moderately thermophilic fungus Talaromyces emersonii. Protein Expr Purif 38:248–257
Nierman WC, Pain A, Anderson MJ et al (2005) Genomic sequence of the pathogenic and allergenic filamentous fungus Aspergillus fumigatus. Nature 438:1151–1156
Norvell LL (2011) Fungal nomenclature. 1. Melbourne approves a new code. Mycotaxon 116:481–490
Nylander JAA (2004) MrModeltest 2.2. Program distributed by the author. Evolutionary Biology Centre, Uppsala University, Uppsala
O’Donnell K, Cigelnik E (1997) Two divergent intragenomic rDNA ITS2 types within a monophyletic lineage of the fungus Fusarium are nonorthologous. Mol Phylogenet Evol 7:103–116
Ogawa H, Sugiyama J (2000) Evolutionary relationships of the cleistothecial genera with Penicillium, Geosmithia, Merimbla and Sarophorum anamorphs as inferred from 18S rDNA sequence divergence. In: Samson RA, Pitt JI (eds) Integration of modern taxonomic methods for Penicillium and Aspergillus classification. Plenum Press, New York, pp 149–161
Ogawa H, Yoshimura A, Sugiyama J (1997) Polyphyletic origins of species of the anamorphic genus Geosmithia and the relationships of the cleistothecial genera: evidence from 18S, 5S and 28S rDNA sequence analyses. Mycologia 89:756–771
Overy DP, Blunt JW (2004) Corymbiferan lactones from Penicillium hordei: stimulation of novel phenolic metabolites using plant tissue media. J Nat Prod 67:1850–1853
Overy DP, Valdez JG, Frisvad JC (2005a) Revisions to Penicillium ser. Corymbifera: agents responsible for blue mould storage rot of various flower and vegetable bulbs. Can J Bot 83:1422–1433
Overy DP, Zidorn C, Petersen BO, Duus JO, Dalsgaard PW, Larsen TO, Phipps RK (2005b) Medium dependent production of corymbiferone a novel products from Pencillium hordei cultures on plant tissue agar. Tetrahedron Lett 46:3225–3228
Pel HJ, de Winde JH, Archer DB et al (2007) Genome sequencing and analysis of the versatile cell factory Aspergillus niger CBS 513.88. Nat Biotechnol 7:221–231
Peterson SW (2008) Phylogenetic analysis of Aspergillus species using DNA sequences from four loci. Mycologia 100:205–226
Pitt J (1979) Geosmithia gen. nov. for Penicillium lavendulum and related species. Can J Bot 57:2021–2030
Pitt JI, Samson RA, Frisvad JC (2000) List of accepted species and their synonyms in the family Trichocomaceae. In: Samson RA, Pitt JI (eds) Integration of modern taxonomic methods for Penicillium and Aspergillus classification. Plenum Press, New York, pp 149–161
Rabie CJ, Steyn PS, van Heerden FR (1986) The isolation and identification of some toxic constituents of Aspergillus wentii Wehmer. Mycotoxin Res 2:19–24
Raper KB, Thom C (1949) Manual of the Penicillia. Williams and Wilkins, Baltimore
Rossman A, Seifert K (2011) Phylogenetic revision of taxonomic concepts in the Hypocreales and other Ascomycota—a tribute to Gary J. Samuels. Stud Mycol 68:1–248
Samson RA, Houbraken J, Thrane U, Frisvad JC, Andersen B (2010) Food and indoor fungi. CBS laboratory manual series 2. CBS-Fungal Biodiversity Centre, Utrecht
Sharpton TJ, Stajich JE, Rounsley SD et al (2009) Comparative genomic analyses of the human fungal pathogens Coccidioides and their relatives. Genome Res 19:1722–1731
Smith G (1949) The effect of adding trace elements to Czapek-Dox culture medium. Trans Br Mycol Soc 32:280–283
Smith G (1957) Some new and interesting species of micro-fungi. Trans Br Mycol Soc 40:81–488
Smith G (1960) Industrial mycology, 5th edn. Edward Arnold Ltd., London
Stamatakis A, Hoover P, Rougemont J (2008) A rapid bootstrap algorithm for the RAxML Web-Servers. Syst Biol 75:758–771
Stolk AC (1965) Thermophilic species of Talaromyces Benjamin and Thermoascus Miehe. Antonie van Leeuwenhoek 31:262–276
Stolk AC, Samson RA (1972) The genus Talaromyces. Studies on Talaromyces and related genera II. Stud Mycol 2:1–67
Stolk AC, Evans HC, Nilsson T (1969) Penicillium argillaceum sp. nov., a thermotolerant Penicillium. Trans Br Mycol Soc 53:307–311
Turner WB (1971) Fungal metabolites. Academic Press, London
Turner P, Mamo G, Nordberg-Karlsson E (2007) Potential and utilization of thermophiles and thermostable enzymes in biorefining. Microb Cell Fact 6:9
van den Berg MA, Albang R, Albermann K et al (2008) Genome sequencing and analysis of the filamentous fungus Penicillium chrysogenum. Nat Biotechnol 26:1161–1168
Yaguchi T, Someya A, Udagawa S (1994) Two new species of Talaromyces from Taiwan and Japan. Mycoscience 35:249–255
Yaguchi T, Udagawa S, Nishimura K (2005) Geosmithia argillacea is the anamorph of Talaromyces eburneus as a heat resistant fungus. Crypt Mycol 26:133–141
Acknowledgments
The excellent technical assistance of Tineke van Doorn, Martin Meijer, Neriman Yilmaz and Ellen Kirstine Lyhne is greatly acknowledged. Uwe Braun is thanked for providing the Latin diagnosis, Ursula Weidner for providing the Rasamsonia brevistipitata isolates, Vanessa Barrs for critically reading the manuscript and all anonymous reviewers for their helpful suggestions.
Open Access
This article is distributed under the terms of the Creative Commons Attribution Noncommercial License which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
Author information
Authors and Affiliations
Corresponding author
Electronic supplementary material
Below is the link to the electronic supplementary material.
10482_2011_9647_MOESM1_ESM.tif
Supplementary Figure 1. Overview of growth rates on MEA of Rasamsonia species and Trichocoma paradoxa at different temperatures, 18, 21, 24, 27, 30, 33, 36, 40, 45 and 50°C. From left to right: R. argillacea DTO 137A5, R. eburnea CBS 124445, R. cylindrospora CBS 275.58NT, R. brevistipitata CBS 128785, R. emersonii CBS 355.92, R. byssochlamydoides CBS 533.71, Trichocoma paradoxa CBS 103.73. (TIFF 26822 kb)
10482_2011_9647_MOESM2_ESM.tif
Supplementary Figure 2. Rasamsonia argillacea. A–H. Colonies 7 days old 37°C. A. MEA; B. CYA; C. YES; D. OA; E. MEA reverse; F. CYA reverse; G. YES reverse; H. CREA. I–M. Conidiophores. N. Conidia. Scale bar = 10 μm. (TIFF 13122 kb)
10482_2011_9647_MOESM3_ESM.tif
Supplementary Figure 3. Rasamsonia byssochlamydoides. A–H. Colonies 7 days old 37°C. A. MEA; B. CYA; C. YES; D. OA; E. MEA reverse; F. CYA reverse; G. YES reverse; H. CREA. I–J. Ascomata. K–N. Conidiophores. O. Conidia. P. Ascospores. Scale bar = 10 μm. (TIFF 13712 kb)
10482_2011_9647_MOESM4_ESM.tif
Supplementary Figure 4. Rasamsonia cylindrospora. A–H. Colonies 7 day old 37°C. A. MEA; B. CYA; C. YES; D. OA; E. MEA reverse; F. CYA reverse; G. YES reverse; H. CREA. I–M. Conidiophores. N. Conidia. Scale bar = 10 μm. (TIFF 13924 kb)
10482_2011_9647_MOESM5_ESM.tif
Supplementary Figure 5. Rasamsonia eburnea. A–H. Colonies 7 days old 37°C. A. MEA; B. CYA; C. YES; D. OA; E. MEA reverse; F. CYA reverse; G. YES reverse; H. CREA. I–M. Conidiophores. N. Conidia. Scale bar = 10 μm. (TIFF 13769 kb)
10482_2011_9647_MOESM6_ESM.tif
Supplementary Figure 6 Rasamsonia emersonii. A–H. Colonies 7 days old 37°C. A. MEA; B. CYA; C. YES; D. OA; E. MEA reverse; F. CYA reverse; G. YES reverse; H. CREA. I–J. Ascomata. K–N. Conidiophores. O. Conidia. P. Ascospores. Scale bar = 10 μm. (TIFF 13406 kb)
Rights and permissions
Open Access This is an open access article distributed under the terms of the Creative Commons Attribution Noncommercial License (https://creativecommons.org/licenses/by-nc/2.0), which permits any noncommercial use, distribution, and reproduction in any medium, provided the original author(s) and source are credited.
About this article
Cite this article
Houbraken, J., Spierenburg, H. & Frisvad, J.C. Rasamsonia, a new genus comprising thermotolerant and thermophilic Talaromyces and Geosmithia species. Antonie van Leeuwenhoek 101, 403–421 (2012). https://doi.org/10.1007/s10482-011-9647-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10482-011-9647-1