
Abstract
Platelets control hemostasis and play a key role in inflammation and immunity. However, platelet function may change during aging, and a role for these versatile cells in many age-related pathological processes is emerging. In addition to a well-known role in cardiovascular disease, platelet activity is now thought to contribute to cancer cell metastasis and tumor-associated venous thromboembolism (VTE) development. Worldwide, the great majority of all patients with cardiovascular disease and some with cancer receive anti-platelet therapy to reduce the risk of thrombosis. However, not only do thrombotic diseases remain a leading cause of morbidity and mortality, cancer, especially metastasis, is still the second cause of death worldwide. Understanding how platelets change during aging and how they may contribute to aging-related diseases such as cancer may contribute to steps taken along the road towards a “healthy aging” strategy. Here, we review the changes that occur in platelets during aging, and investigate how these versatile blood components contribute to cancer progression.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
1 Introduction
Physiological changes occur in all organ systems during aging, and are a reflection of changes that occur on a molecular level in individual cells. Diverse animal and yeast models have shown that aging is associated with tissue-specific changes in transcriptomes as well as intra- and extracellular metabolite changes [1]. Cellular senescence, a block in cellular proliferation as a result of (amongst others) telomere shortening and loss of DNA damage repair, plays an important role in the process of aging [2]. In addition to telomere attrition, genomic instability, and cellular senescence, other hallmarks of cellular aging include stem cell exhaustion, epigenetic alterations, loss of proteostasis, deregulated nutrient sensing, mitochondrial dysfunction, and altered intercellular communication [3]. Not all cells become senescent, and removal of senescent cells may reduce aging on an organismal level [4]. However, cellular communication is mediated in part via the release of vesicles known as exosomes, which can carry cellular components from one cell to another across large distances. Senescent cells also release such exosomes and these have been speculated to play a significant role in age-related phenotypes including age-related diseases [5]. Connecting all known cellular alterations to biological aging remains challenging, and finding ways to promote “healthy aging” remains a holy grail [3].
Thus far, aging is often studied in the context of stem cell capacity and longevity, but cellular changes in individual cell types have also been investigated for neurons, skin fibroblasts and keratinocytes, bone and bone marrow (bone-proximal osteoblastic niche), and many other tissues and cell types [6,7,8]. One more cellular component to be added to this mix are platelets, as a role for these blood constituents in aging and age-related diseases is now emerging [9]. Like many systems in cellular metabolism and catabolism, the biology/function of platelets appears to be altered in the elderly. In addition, altered platelet function and clinical conditions such as cancer create a complex chain of cause and effect, which can culminate in systemic responses responsible for the main causes of death in the world, namely, (1) inappropriate blood clot formation known as thrombosis and (2) cancer metastasis, responsible for more than 90% of cancer-related deaths [9, 10]. Thrombotic risk in the elderly is associated with genetic factors, but also with lifestyle, obesity, and diseases such as cancer [11, 12], creating a complex feedback loop. Other examples of the interrelationship between platelet function and pathological conditions can be seen in the acquisition of bleeding disorders such as hemophilia or Von Willebrand syndrome [13], or the involvement of platelets to neurological disorders such as Alzheimer disease (for review, see [14]). In this latter condition, the microenvironment sensitizes platelets to activation and renders them less sensitive to inhibition, most likely due to increased sensitivity to some platelet activation agonists, such as thrombin and collagen, leading to an increase in β-amyloid production by platelets [15, 16]. Large-scale omics studies have demonstrated age-specific proteomic changes in platelets from childhood to adulthood [17], and miRNA patterns associated with age in individuals ranging from 18 to 46 years old [18]. It is conceivable that such cellular changes may predispose an individual to aging-related diseases. In this review, we summarize the impact of aging on platelet function, and investigate how such altered platelet functionality can contribute to aging-related diseases, with particular emphasis on cancer.
2 Aging-associated changes in platelet phenotype and function(s)
Since the lifespan of platelets is around 7 to 10 days in the bloodstream, changes in platelet functions may be correlated with megakaryocyte maturation, adhesion, and thrombopoiesis, as changes in megakaryocyte maturation during aging lead to altered proplatelet formation and release of platelets with an altered content [19]. Some of these events appear to be driven by β-adrenergic signals coming from a senescent microenvironment [19,20,21]. As such, megakaryocyte aging, aging of platelets in the circulation, and cues from an aged microenvironment to megakaryocytes and nascent platelets during organismal aging can all contribute to changes in platelet biology in elderly individuals. Under normal conditions, there is a gradual loss of RNA content over the course of a platelet lifespan, while in aged organisms, distribution of megakaryocyte content to platelets is altered. However, there are also clear differences between “aged platelets” and “platelets in aged individuals.” Hepatic clearance of senescent platelets from the circulation of adult organisms is dependent on the loss of sialic acid residues of glycoproteins in the cell membrane. Activation of the pro-apoptotic BAX–BAK pathway in aged platelets results in caspase-dependent surface exposure of phosphatidylserine, which serves as a recognition signal for phagocytic cells. In terms of functionality, senescent platelets have impaired adhesion and aggregation responses. On the other hand, platelets in senescent organism might be primed to increase their responsiveness to agonists (hyper-reactive platelets) [22, 23].
Several recent studies have investigated the effect of aging on platelet morphology and function. During the course of life, platelet size increases [24], which directly affects platelet content, including granules and pro-coagulation factors. Other morphological changes seen in platelets from older individuals include an irregular, less smooth plasma membrane with more frequent ruptures, and an increase of slender pseudopodia [25]. The number of circulating platelets is thought to decrease with advanced age. While a study of over 5000 participants suggested that platelet count in individuals of > 65 years is not affected by subsequent age differences [26], two large studies investigating over 25,000 and 40,000 individuals, respectively, showed that platelet numbers drop from early childhood, are relatively stable in adulthood, and drop again over the age of 60 years old, irrespective of gender and ethnicity [27, 28]. Careful consideration of the age groups studied is essential, and for the purpose of this review, we therefore aimed to compare young adults (18–39 years), middle-aged (40–59 years), old-aged (60–79 years), and very-old-aged (> 80 years) groups, where possible (Figs. 1 and 2). While the cause of reduced platelet numbers during aging remains to be clarified, some studies have suggested changes in hematopoietic stem cells as a pivotal cause of lower platelet counts in advanced age [59,60,61].

Age-associated changes in platelet function. Platelet function of aggregation, tissue repair, and remodeling changes discriminated on age groups. The concept of age groups is based on young adults (18–39 years), middle-aged (40–59 years), old-aged (60–79 years), very-old-aged group (> 80 years) [27, 29]

Age-associated changed in platelet markers. Platelets present several changes during the aging process on their content (cytosolic and membrane) and release thereof. The concept of age groups is based on young adults (18–39 years), middle-aged (40–59 years), old-aged (60–79 years), very-old-aged group (>80 years) [40,41,42,43,44,45,46,47, 31, 32, 48, 58]
Despite a lower platelet count in older individuals, bleeding times are reduced during aging, which is thought to contribute to an increased risk of blood clot formation [62]. Bleeding time (i.e., time before efficient blood clotting occurs) is dependent on platelet count and vessel contractibility, as well as platelet function, and platelets in the elderly are indeed hyper-reactivated, especially in subjects with associated comorbidities (for review, see [61, 63]). For instance, spontaneous platelet aggregation is higher in very old subjects as compared with old adults [30, 64], and a higher sensitivity to ADP stimulation [10, 65, 66] and thrombin receptor–activating protein (TRAP6) [67] is seen. Several other platelet agonists, including ristocetin, thrombin, and collagen, have received attention but whether responsiveness of platelets towards these agonists is increased or decreased during aging remains disputed (Fig. 1).
Whether overactivation of platelets is a failed compensation mechanisms to make up for the loss of platelet count remains speculative. The mechanisms contributing to higher platelet activity in elderly individuals are still under investigation. It has been suggested that age-related inflammatory and metabolic changes contribute to an increased platelet function in the elderly [66]. Mouse models have shown an increase of hydrogen peroxide concentration in blood, which directly increases platelet activity during aging [67]. In humans, oxidative stress markers in platelets increase from young to middle-aged individuals [30, 68]. Hydrogen peroxide accumulation in platelets could be the result of NADPH oxidase and superoxide dismutase activity, which are associated with an increased integrin αIIbβ3 activity in platelets [68]. Indeed, the expression of surface markers such as integrin αIIb and αIIbβ3 is increased during the course of aging [69, 70]. Thus, overall increased oxidative stress is generally seen during the aging process, contributing to the concept that platelet alterations in aging are associated with an increasing inflammatory state. The oxidative burst triggers activation of the signaling molecule mTOR, a key regulator of lifespan and aging [69]. mTOR activation in turn results in an increased platelet production by megakaryocytes [70]. Moreover, mTOR hyper-activation during aging is associated with increased platelet aggregability and aging-related venous thrombosis risk in mice [59]. Thus, mTOR plays a dual role in platelet hyper-aggregability by increasing the activity of platelets, while oxidative stress further increases platelet reactivity, resulting in an enhanced risk of thrombi formation in the elderly (Fig. 2).
Association between activated platelets and monocytes, as would occur during blood clotting, enhances the formation of aggregates. While there is no impact of age on platelet-monocyte aggregation per se in healthy adults [71], higher levels of platelet-monocytes aggregates were seen in patients with acute coronary syndrome [72], and platelet hyper-activation may thus be further exacerbated in disease states. Others have shown that the age-related increases of platelet-derived β-2-microglobulin levels in the serum cause monocyte differentiation towards a less regenerative phenotype, providing a further link between platelet changes during aging and the aging process [73].
A clear association between platelet hyper-reactivity and the occurrence of thromboembolic events exists and may contribute to cardiovascular comorbidities in the elderly [74]. In addition to the direct effect of aging on platelet aggregation described above, this phenomenon has also been attributed to the fact that the production of anti-coagulation factors does not follow the increasing pro-coagulation factor production during aging [11]. Gleerup and Winther showed that, in addition to an enhancement of platelet aggregability, aging provokes a decrease of fibrinolytic activity, further reinforcing the association between lower fibrinolytic activity forming stable thrombus formation and accumulation, an imbalance between thrombotic versus fibrinolytic events [75]. The same research group described that adrenaline and sub-concentration ADP-induced canonical platelet activation is enhanced in old and very old individuals, as is the synergistic effect of serotonin on adrenaline-/ADP-induced platelet activation. Adrenaline levels were also augmented in the old and very old groups [76, 77]. This might be a compensatory mechanism for the fact that β-adrenoreceptors from older individuals show higher ligand affinity. This receptor reduces platelet aggregation through the production of cAMP, and a reduced signaling capacity through this receptor may thus contribute to an enhanced platelet aggregation in the elderly; however, the levels of cAMP in plasma did not change significantly during aging [76, 77]. Endothelial dysfunction during aging may further increase platelet responsiveness [75]. For instance, it has been speculated that platelet activation and aggregation caused by dysfunctional lung epithelium in virally infected individuals may cause depletion of thrombocytes, and contribute to the thrombocytopenia observed in COVID-19 patients infected with SARS-CoV-2 [76, 77].
In addition to blood clotting, it is increasingly recognized that platelets play an important role in wound healing. While wound healing is not absolutely impaired, delayed closure rates and weaker wound repair are commonly seen in subjects of advanced age [78]. During wound healing, many different cell types, including fibroblasts and immune cells such as macrophages and lymphocytes, cooperate to restore tissue architecture. Activated platelets trapped in the blood clot release mediators to attract these cells and express P-selectin which acts as cell adhesion molecule for passing lymphocytes [79]. Furthermore, the secretion of several growth factors, such VEGF, PDGF, EGF, and TGFβ, may modulate T cells to induce keratinocyte regenerative capacity and enhance proliferation of regenerative cells such as fibroblasts [80, 81]. However, while reduced serum levels of these platelet-derived factors could theoretically contribute to decreased wound healing rates, age-related variations in cytokine levels appear most pronounced in early adulthood, disputing their relevance for wound healing delay in the very old individuals [25, 82].
Data collection on platelet function during aging is complicated by several issues. For one thing, platelet aging may be gender-specific, as studies have indicated that aging-related loss of interaction with the adhesion molecule von Willebrand factor (vWF) is more pronounced in women as compared to men [28, 83]. Thus, hormonal changes may contribute to platelet alterations in older subjects [84]. Levels of steroids such as testosterone and dihydrotestosterone in older individuals are negatively associated with platelet activation markers, and these steroids can directly inhibit collagen-induced aggregation in vitro [85]. Secondly, recent data suggest that changes that occur during aging are complicated and were not always found to be continuous during aging. Spontaneous aggregation was increased in elderly individuals compared with younger subjects, while ristocetin or collagen-induced aggregation was decreased (pointing towards platelet exhaustion) [30]. However, these trends did not follow linear relationships with changes most pronounced in the very old (80+ years) [30]. Other platelet activation markers (soluble P-selectin, integrin αIIb, caspase 3, oxidative stress) were shown to increase from young to old individuals, but decrease again in the very old [68]. However, it should be noted that others found no differences in basal membrane-bound P-selectin between individuals < 45 years and > 65 years old [34, 35], while the percentage of platelets expressing P-selectin upon stimulation with TRAP-6 was actually higher in younger individuals [67]. Differences in age groups, methods, and stimuli used vary per study and may account for conflicting results. It should further be noted that the effects observed are sometimes small, and small group sizes may hamper interpretation of results. While many studies point towards disturbances in platelet functionality during aging, the direct consequences on coagulation in healthy aging may not always be clear [85, 86], and may be more pronounced under pathological conditions.
2.1 Platelet bioactive lipids in aging
A detailed study on platelet lipid production and aging was reported in 1986 [49]. This study investigated platelet cholesterol and phospholipids content, and observed a slight increase of cholesterol/phospholipids molar ratio upon aging within a range of 20 to 69 years old [87]. It is important to highlight that platelets are not able to produce their own cholesterol, which must be obtained during their genesis (from megakaryocytes) or derived from plasma. The cholesterol/phospholipid molar ratio is important to maintain platelet membrane fluidity, and, consequently, the platelet capacity to change its shape during activation. In addition, activation of platelets via agonist-receptor activation in many cases requires localization of receptors and downstream signaling molecules in cholesterol-rich lipid rafts [88]. The lipid composition is also affected by aging [89], with increased fatty acids 16:0 phosphatidylcholine and sphingomyelin, and a decrease of linoleic acids 18:2, 20:4, and 20:3 in older subjects [49]. It is important to note that lipid oxidation occurs on platelet LDL, and this phenomenon may have severe consequences for cardiovascular diseases. One study showed that older males at risk for coronary heart disease due to dietary habits (55–73 years old) showed higher platelet aggregation in response to epinephrine as compared with younger individuals (28–54 years old) and males at lower risk for heart disease, indicating that age-related platelet changes associated with phospholipid content may be a risk factor for cardiovascular diseases [90].
Besides the platelet membrane lipid composition, the most important bioactive lipids relevant to platelet function are the signaling lipids derived from the eicosanoid pathway. Briefly, upon stimulation of cells, membrane-anchored arachidonic acids (AA) are released from the membrane phospholipids by phospholipases (phospholipase A2), after which they are enzymatically converted to prostanoids by COX1/2 enzymes. This process results in production of platelet stimulatory thromboxane (TxA2, mainly produced via COX1 [91]) or platelet antagonistic prostaglandins (PG), PGI2, prostacyclin), PGD2, and PGE2 (mainly via COX2) [92, 93]. Alternatively, AA can be converted to leukotrienes through lipoxygenases activity. Eicosanoids are important mediators of inflammation, and, indeed, eicosanoid biosynthesis is higher on advanced age [77, 94, 95], which in turn may contribute to enhanced inflammatory state during aging [92, 94, 96]. Platelet interaction with peripheral blood mononuclear cells directly modulates inflammatory responses, potentially through their production of PGE2 [79, 80]. In this case, PGE2 decreases the effectiveness of myeloid cell differentiation and affects their responses [97].
However, both increased TxA2 as well as PGE2 and prostacyclin excretion were seen in older humans or rats, which begs the question of how this balance would affect platelet activity [77, 98, 99]. While TxA2 is produced by platelets, the major source of prostacyclins is endothelial cells. While some studies showed no differences in prostacyclin secretion by arterial endothelial cells for donors of different ages [97], others demonstrated reduced prostacyclin expression in aorta endothelia from older individuals, suggesting that perhaps the TxA2 effect wins out during aging. It is of interest to know that dietary restriction, known to prolong healthy aging, is associated with an enhanced prostacyclin/TxA2 ratio in rats [100, 101]. Indeed, increased TxA2 excretion appears to be associated not only with age-related diseases such as atherothrombosis but also with metabolic disease [102, 103]. Obesity and decompensated glucose metabolism increase not only platelet activation but also inflammation (for review, see [104]). In this case, the persistent TxA2-dependent platelet activation increases systemic inflammation [103, 105]. Inflammation-induced endothelial events may play a major role in aging comorbidities. For instance, glycemia-mediated TxA2-receptor activation was associated to disturbed blood-brain barrier integrity in diabetes [106]. Furthermore, TxA2 is a P2X1 ion channel agonist and both platelets and P2X1 are required to maintain vascular integrity in a mouse colitis model [107, 108].
Taken together, a clear change in platelet morphology and function is seen during aging, which may have severe consequences for aging-related physiology. The most relevant changes in platelet biology were highlighted in Figs. 1 and 2.
3 Platelets in cancer—“double-edged sword”?
As described above, platelet hyper-reactivity during aging is associated with an increased risk of formation of embolisms. Nevertheless, despite cancer being an age-related disease, thrombocytopenia is a common event in these patients. The risk of bleeding in thrombocytopenic cancer patients is difficult to predict [109], and platelet counts must be carefully monitored. In particular, cancers of the bone marrow (platelet production from megakaryocytes) or spleen (platelet clearance), where hematopoiesis is affected, are prone to lead to loss of platelet counts. For instance, thrombocytopenia in patients with bone dyscrasias is directly related to bleeding events [110]. However, the most common cause of bleeding due to platelet loss in cancer patients arises as a result of myeloablative chemotherapy [111] and cytopenia may therefore be a bystander effect rather than a pathogenic event. In fact, the role of platelets in cancer appears to be ambiguous, as enhanced blood clotting represents a major risk factor in cancer patients.
Patients with cancer (but also those with cardiovascular diseases including diabetes, hyper-cholesterolemia, and hypertension) can develop an increased platelet activity, which may be either age-related or disease-specific. The hyper-aggregability observed in these diseases appears to be related to higher platelet reactivity towards agonists or increased circulation of these agonists (such as thrombin and factor Xa), and is a primary cause of thrombotic events, in particular venous thromboembolism events (VTE) and arterial thrombosis (AT) [112, 113]. These events partially overlap, with shared risk factors, and similar incidence in cancer patients [114, 115].
The first report of a platelet-related disorder in cancer came from Armand Trousseau, who described a higher risk of thrombotic events in cancer patients [116], which has subsequently been termed Trousseau syndrome. As the second cause of death, VTE poses a significant comorbidity in cancer patients, and a common cause of hospitalizations, thereby significantly contributing to cancer-associated health care costs [117]. Several cancers are associated with increased VTE risk, including renal carcinoma [118]; hepatocellular carcinoma [119]; lung cancer [120]; and esophageal and stomach cancer [112]. Moreover, VTE in esophageal or gastric cancer patients has been associated with decreased survival: patient survival without VTE is 18 months compared with 13.9 months with VTE [121]. While the risk of VTE appears to be especially high in patients suffering from stomach and pancreatic cancer, up to 20% of all cancer patients may develop thromboembolisms, including pulmonary and venous events. For AT, the overall incidence of events in patients with cancer is increased 2-fold [115].
Enhanced platelet activation as determined by mean platelet volume (MPV) is seen in cancer patients, and may correlate with tumor stage [122, 123]. Both MPV and increased soluble P-selectin levels correlate with VTE development in cancer patients [124,111,126]. Age does not predict VTE risk for all cancer types, suggesting that at least for some cancer types, tumor cells themselves increase platelet reactivity and VTE risk [127]. Indeed, higher platelet P-selectin expression was found in mouse models of breast cancer, which in turn was associated to lung metastasis [128]. In addition, MPV, which is enhanced in malignant tumors, drops upon treatment [129], enforcing the direct link between tumor burden and platelet activation. Thus, cancer cell–mediated platelet hyper-reactivity contributes to increased VTE risk. While to date, there is no method available and validated to monitor the clinical implication of platelet hyper-aggregability in cancer patients; this may be a promising avenue of investigation [130].
Multiple mechanisms may underlie the tendency of platelets from cancer patients to aggregate. Tumor cells can stimulate platelet aggregation through direct interaction via adhesion molecules or via the delivery of extracellular vesicles and/or secreted factors. This phenomenon, described as tumor cell–induced platelet activation (TCIPA), was already identified decades ago [130]. It has now been shown that single tumor cells are capable of attracting and activating platelets to form fibrin clots [131]. Furthermore, platelets from cancer patients differ from platelets from healthy controls in their mRNA profiles, with mRNA transcripts undergoing alternative splicing under influence of tumor-derived stimuli [132, 133]. Platelets are also capable of taking up tumor content, as determined by the fact that tumor-specific mutations can be identified in platelets upon co-culture with tumor cells. This process appears to be regulated by extracellular vesicles released by the tumor cells, which are subsequently taken up by co-cultured platelets [134]. This alteration of platelets by tumor cells, i.e., tumor education, was shown to contribute to an increased adhesive propensity of platelets [135,122,137]. Furthermore, cancer cells shed extracellular vesicles containing the adhesion molecule tissue factor (TF), which may contribute to VTE at sites of vessel damage [134, 138].
4 Platelets drive tumor growth, angiogenesis, and metastasis in cancer
Specifically in solid tumors, the interaction of tumor cells and platelets leads to a condition called paraneoplastic thrombocytosis, in which malignant tumors not only hijack or mimic platelet functions but can also increase their production. A cyclic picture emerges, which contributes to the most feared outcome of a malignant neoplasm: metastasis [139]. Metastasis is the principal cause of death in cancer patients and investigation of the molecular mechanisms that coordinate this process is therefore crucial. The process of metastasis requires several steps: invasion of cells in the surrounding matrix, intravasation to the blood circulation, survival at the circulation, extravasation at the secondary site (tissue or organ), micrometastasis formation and colonization [140]. The primary tumor can shed many cells during the growth phase; however, only a few cells are able to colonize a secondary site [135]. Much depends on the survival of these tumor cells in the blood circulation, survival of detachment, and the hemodynamic flux force, as well as escaping the immune system. One of the principal strategies of cancer cells to survive in the circulation is interaction with platelets, and nearly all processes of cancer metastasis appear to be facilitated by interaction of tumor cells with platelets.
Platelets can stimulate expression of metalloproteinases in tumor cells, which in turn contributes to tumor cell invasion by facilitating extracellular matrix degradation [141, 142]. Tumor cell metastasis often requires the acquisition of a different phenotype, termed epithelial-to-mesenchymal transition (EMT). This process is characterized by upregulation of several molecular markers (e.g., expression of SNAIL, vimentin cadherin, and MMPs), and platelet-released TGFβ can significantly enhance the upregulation of these markers in cancer cells [143, 144]. In addition, direct contact between cancer cells and platelets contributes to TGFβ/Smad and NFκB pathway activation, culminating in EMT stimulation. Adherence of cells to the extracellular matrix provides survival signals, which are disrupted upon detachment of cells, thereby leading to anoikis: detachment-induced apoptosis. While cancer cells have several mechanisms to overcome anoikis, it has been demonstrated that interaction of cancer cells with platelets further induces tumor cell resistance against anoikis [129]. Thus, platelet-induced alteration of cancer cell intracellular programs contributes to tumor invasiveness and metastasis [135, 144, 145].
Extravasation of tumor cells from tissue to bloodstream is facilitated by platelet-derived ADP stimulation of P2Y2 receptors on endothelial cells [146]. Once the cancer cell enters the blood circulation, the dissemination efficiency also depends on the interaction with platelets, with many studies showing that platelets facilitate the metastatic process via hematogenous dissemination [143, 147]. Survival of tumor cells in the blood stream is not only enhanced by platelets through mechanic protection from shear force but also by protecting the cancer cells from circulating immune cells, which may target neoantigens, expressed by tumor cells. Interestingly, it has been demonstrated that cancer cells may mimic platelets by expressing megakaryocytic genes and expressing platelet surface markers, including adhesion molecules such as integrins and selectins [139, 148]. Additionally, coating of tumor cells with platelets allows transferring their major histocompatibility complex (MHC) class I to tumor cells, thereby giving these cells a false “pseudonormal” exterior, and allowing escape from immunosurveillence by natural killer cells [149]. TGFβ released by platelets also downregulates the NK receptor NKG2D on tumor cells, further shielding them from immunosurveillence [150, 151]. Lastly, extravasation of the tumor cells from the blood stream is facilitated by platelets, and appears to require binding of platelets to Integrin ανβ3 expressed on tumor cells [152].
As a solid tumor grows and its oxygen and nutrient demands increase, angiogenesis, the formation of new blood vessels, is essential for its survival. Tumor-induced angiogenesis often results in an abnormal vasculature with suboptimal perfusion. Nevertheless, tumor cells may benefit from this, as this may reduce delivery of therapies and tumor-targeted immune cells [150]. Furthermore, tumor cells may adapt to such ineffective vascularization, and the ensuing hypoxia may favor tumorigenesis by selecting for aggressive and metastatic clones [153]. Supplementation of platelets or their released products stimulates angiogenesis induced by breast tumor cells in vitro [136, 154]. In glioblastoma patients, release of VEGF by platelets was shown to contribute to vessel formation [155], although other studies indicated that platelet-induced angiogenesis was independent of VEGF but most likely relied on release of several other factors, including IL6, thrombopoietin, and angiopoietin [156, 157]. Furthermore, animal models indicate that tumor-educated platelets are more efficient at inducing angiogenesis than healthy platelets, suggesting a more efficient delivery of pro-angiogenic factors by tumor-educated platelets [158]. This appears to be supported by findings in humans, showing that levels of VEGF are increased in platelets from prostate, breast, and colorectal cancer patients [159, 160]. It is of interest to note that vasculogenic mimicry, where tumor cells themselves rather than endothelial cells form vessels, is inhibited by platelets. While counterintuitive, this process is thought to promote metastasis [161]. Thus, platelets tightly coordinate the vascularization process in the context of cancer, and may thereby potentiate malignancies.
Thus far, platelet participation in cancer progression has been associated with vascularization, delivery of growth factors, and hematogenous dissemination [143]. In addition, platelets may directly stimulate cancer cell proliferation through upregulation of oncogenic genes, as was demonstrated for colorectal cancer cells [131]. Thus, platelets play a role in all aspects of cancer progression, something we may do well to take into account when addressing these diseases.
Taking the above into account, it is perhaps surprising to realize that fibrinolysis, the process of dissolving a blood clot, can also play a tumor-promoting role [162]. The main enzyme promoting fibrinolysis is plasmin, while the platelet-derived plasminogen activator inhibitor (PAI) is the main suppressor of this system. Elevated PAI-1 levels are associated with VTE [163], and may explain VTE in pancreatic and glioma cancer patients [164, 165]. As such, inhibition of fibrinolysis is detrimental to cancer patients. On the other hand, plasminogen itself contributes to metastasis by degradation of the extracellular matrix surrounding tumor cells. In addition, the fibrinolytic system contributes to inflammation, angiogenesis, the release of tumor growth factors, and other tumor-promoting functions [162]. Thus, coagulation and fibrinolysis play double roles in cancer, highlighting platelet performance as double-edged sword [166].
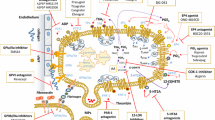
In order to target these interactions in healthy aging as well as age-related diseases, detailed knowledge regarding the molecular mechanisms involved may prove essential (Fig. 3). Many of the molecular interactions between cancer cells and platelets depend on their molecular cell surface composition. Platelets can interact with cancer cells via tissue factor (TF), selectins, integrins, and glycoproteins receptors, all of which may activate signaling pathways leading to platelet activation. Thus, platelet membrane components have multiple functions: they contribute directly to hemostasis during thrombus formation, but can also contribute to multifactorial cancer dissemination. TF expressed by cancer cells stimulates platelet activation and initiation of the coagulation cascade. The fibrin produced by platelets subsequently interacts with integrins from cancer cells as well as platelets themselves, inducing formation of cancer cell–fibrin–platelet clusters, which may enter the circulation [167, 168]. Overexpression of TF on breast cancer cells has been reported, and appears to be linked to the release of TGFβ from activated platelets [169]. Furthermore, in ovarian cancer, platelet-induced increase in TF acts as a metastasis initiator [170].

The cross talk between cancer cells and platelets support metastasis, angiogenesis, and tumor growth. Platelets release factors such as TGFβ and VEGF that stimulate epithelial-to-mesenchymal transition (EMT) and angiogenesis. Additionally, platelets contribute to escape from immunosurveillance by covering cancer cells and shielding them from the immune system
The contribution of integrins to cancer cell–platelet interactions is broad and bidirectional. Platelets express integrins αIIbβ3, αvβ3, α2β1, α5β1, and α6β1, which bind preferentially fibrinogen, vitronectin, collagen, fibronectin, and laminins, respectively, all of which have been described to have adhesive proprieties [150]. Mammadova-Bach and colleagues described that integrin α6β1 from platelets directly binds ADAM9 from tumor cells, a member of the disintegrin and metalloproteinase family. As a consequence of this interaction, platelets are activated and support hematogenous dissemination of cancer cells [171]. Conversely, as already mentioned above, interaction of αvβ3 on platelets was associated with extravasation in aggressive breast cancer [152]. A last class of molecules facilitating the interaction between cancer cells and platelets are selectins, membrane-localized glycoproteins that bind carbohydrates from glycoproteins, glycolipids, and glycosaminoglycan/proteoglycans. Of the selectin family, P-selectin is expressed on platelets and endothelial cells and has already been mentioned above. Platelet dysfunction as a result of P-selectin deficiency limits colon carcinoma and metastasis progression [172, 173]. E-selectin, which is produced by endothelial cells, binds to sialyl-Lewis-x/an, otherwise known as CA19-9, a common tumor marker. The ensuing interaction promotes hematogenous dissemination of colorectal cancer cells [174].
Platelet bioactive lipids are also associated to cancer metastasis (for review, see [175]), and prostanoid synthesis inhibition as a strategy for cancer treatment has been suggested since 1972 [176]. Leukemic cell–induced platelet aggregation is associated with increased TxA2 and decreased leukotriene B4 (LTB-4) production by platelets [177]. TxA2 in turn promotes metastasis of various tumor models by increasing TCIPA, endothelial cell activation, and recruitment of innate immune cells, all contributing to creating a pre-metastatic niche [178]. Targeting COX1/TxA2 appears efficient to reduce tumor cell metastasis [179, 180]. Conversely, prostacyclin, one of the most potent platelet inhibitors, prevents metastasis in a melanoma model [176, 178]. Endothelial function, essential to tumor cell intravasation/extravasation, is also modulated by prostacyclins. Interestingly, endothelial dysfunction, as characterized (amongst others) by decreased prostacyclin and increased P-selectin levels, was associated with more severe lung cancer stage, but also to patient age [181]. PGD2 can also decrease tumor MMP-2 expression, inhibit EMT inhibition, and reduce tumor cell proliferation [182, 183]. While these latter functions appear to be independent of platelets, some of the prostacyclin-mediated anti-tumor effects may come from inactivation of platelet hyper-reactivity in response to cancer cells, as was shown for melanoma, lung cancer, and breast cancer [179]. However, the anti-tumorigenic effects of prostacyclin and PGD2 may be specific to these prostanoids, as PGE2 did not reduce TCIPA, and COX2 and PGE2 have been associated with enhanced rather than reduced cancer metastasis [184, 185]. Thus, while COX2 inhibitors have been advocated as anti-cancer treatments in the context of inflammation (i.e., prostaglandins are important mediators of inflammation, which in turn may have carcinogenic effects), caution should be taken [186, 187]. Complicating matters further is the fact that platelets and their products may actually protect endothelial cells, in particular under inflamed conditions (e.g., platelet dysfunction has been suggested to contribute to endothelial dysfunction in COVID-19 patients) [188]. By strengthening the endothelial barrier, platelets may prevent intra/extravasation of tumor cells, thereby limiting tumor metastasis (reviewed in [189]).
All in all, many different molecular associations underlie platelet–cancer cell interactions and a better insight into these pathways may provide targets for treatment of both cancer and its associated VTE risk in elderly patients. With platelets playing multiple roles in cancer progression, care needs to be taken when using platelet inhibitors [189].
5 Conclusions
It is becoming increasingly clear that aging is associated with changes in platelet ontogenesis/biogenesis and function, and that this may have consequences for physiological aging. With the (relatively late) recognition of the importance of platelets, it has also become evident that age-related diseases such as cancer and cardiovascular disease are associated with platelet alterations (Fig. 4). However, to what extent this is driven by age-related changes or whether these alterations are disease-specific is perhaps unclear and age-matching in platelet investigation is imperative. Nevertheless, evidence showing that tumor cells directly modulate platelet content and functions suggests that while aging may predispose towards platelet dysfunction, specific disease states may further exacerbate platelet dysfunction to a pathological extent. Finding ways to break this pathological interaction while maintaining the balance of hemostasis may prove an important step towards healthy aging.

Aging-related changes in platelet function and their association with aging-related diseases (e.g., cancer). As a cross-link between aging and cancer, oxidative stress, wound healing disturbed, inflammation, lower platelet count, and senescent cells delivery factors are highlighted. Platelets support metastasis by augmentation of integrin activity, increasing expression of metalloproteinases, and the release of growth factors, which also augment angiogenesis. Furthermore, kinase activation, including mTOR pathways, increase platelet activation. Production of reactive oxygen species enhances platelet production
References
Zhang, S., Ratliff, E. P., Molina, B., El-Mecharrafie, N., Mastroianni, J., Kotzebue, R. W., Achal, M., Mauntz, R. E., Gonzalez, A., Barekat, A., Bray, W. A., Macias, A. M., Daugherty, D., Harris, G. L., Edwards, R. A., & Finley, K. D. (2018). Aging and intermittent fasting impact on transcriptional regulation and physiological responses of adult drosophila neuronal and muscle tissues. International journal of molecular sciences, 19(4), 1140. https://doi.org/10.3390/ijms19041140.
Victorelli, S., & Passos, J. F. (2017). Telomeres and cell senescence - size matters not. EBioMedicine, 21, 14–20. https://doi.org/10.1016/j.ebiom.2017.03.027.
López-Otín, C., Blasco, M. A., Partridge, L., Serrano, M., & Kroemer, G. (2013). The hallmarks of aging. Cell, 153(6), 1194–1217. https://doi.org/10.1016/j.cell.2013.05.039.
Baker, D. J., Wijshake, T., Tchkonia, T., LeBrasseur, N. K., Childs, B. G., van de Sluis, B., Kirkland, J. L., & van Deursen, J. M. (2011). Clearance of p16Ink4a-positive senescent cells delays ageing-associated disorders. Nature, 479(7372), 232–236. https://doi.org/10.1038/nature10600.
Jakhar, R., & Crasta, K. (2019). Exosomes as emerging pro-tumorigenic mediators of the senescence-associated secretory phenotype. International journal of molecular sciences, 20(10), 2547. https://doi.org/10.3390/ijms20102547.
Thon, J.N., & Italiano, J.E. Platelets: production, morphology and ultrastructure. Handb Exp Pharmacol. 2012;(210):3–22. https://doi.org/10.1007/978-3-642-29423-5_1 In: Gresele P, Born G, Patrono C, Page C. (eds) Antiplatelet Agents. Handbook of Experimental Pharmacology, vol 210. Springer, Berlin, Heidelberg
Böhnke, L., Traxler, L., Herdy, J. R., & Mertens, J. (2018). Human neurons to model aging: a dish best served old. Drug discovery today. Disease models, 27, 43–49. https://doi.org/10.1016/j.ddmod.2019.01.001.
Qin, W., & Dallas, S. L. (2019). Exosomes and extracellular RNA in muscle and bone aging and crosstalk. Current osteoporosis reports, 17(6), 548–559. https://doi.org/10.1007/s11914-019-00537-7.
Le Blanc, J., & Lordkipanidzé, M. (2019). Platelet function in aging. Frontiers in cardiovascular medicine, 6, 109. https://doi.org/10.3389/fcvm.2019.00109.
Valenzuela, C. A., Quintanilla, R., Olate-Briones, A., Venturini, W., Mancilla, D., Cayo, A., Moore-Carrasco, R., & Brown, N. E. (2019). SASP-dependent interactions between senescent cells and platelets modulate migration and invasion of cancer cells. International journal of molecular sciences, 20(21), 5292. https://doi.org/10.3390/ijms20215292.
Wilkerson, W. R., & Sane, D. C. (2002). Aging and thrombosis. Seminars in thrombosis and hemostasis, 28(6), 555–568. https://doi.org/10.1055/s-2002-36700.
Montagnana, M., Favaloro, E. J., Franchini, M., Guidi, G. C., & Lippi, G. (2010). The role of ethnicity, age and gender in venous thromboembolism. Journal of thrombosis and thrombolysis, 29(4), 489–496. https://doi.org/10.1007/s11239-009-0365-8.
Kruse-Jarres, R. (2015). Acquired bleeding disorders in the elderly. Hematology. American Society of Hematology. Education Program, 2015, 231–236. https://doi.org/10.1182/asheducation-2015.1.231.
Leiter, O., & Walker, T. L. (2019). Platelets: the missing link between the blood and brain? Progress in neurobiology, 183, 101695. https://doi.org/10.1016/j.pneurobio.2019.101695.
Skovronsky, D. M., Lee, V. M., & Praticò, D. (2001). Amyloid precursor protein and amyloid beta peptide in human platelets. Role of cyclooxygenase and protein kinase C. The Journal of biological chemistry, 276(20), 17036–17043. https://doi.org/10.1074/jbc.M006285200.
Pluta, R., & Ułamek-Kozioł, M. (2019). Lymphocytes, platelets, erythrocytes, and exosomes as possible biomarkers for Alzheimer’s disease clinical diagnosis. Advances in experimental medicine and biology, 1118, 71–82. https://doi.org/10.1007/978-3-030-05542-4_4.
Cini, C., Yip, C., Attard, C., Karlaftis, V., Monagle, P., Linden, M., & Ignjatovic, V. (2015). Differences in the resting platelet proteome and platelet releasate between healthy children and adults. Journal of proteomics, 123, 78–88. https://doi.org/10.1016/j.jprot.2015.04.003.
Simon, L. M., Edelstein, L. C., Nagalla, S., Woodley, A. B., Chen, E. S., Kong, X., Ma, L., Fortina, P., Kunapuli, S., Holinstat, M., McKenzie, S. E., Dong, J. F., Shaw, C. A., & Bray, P. F. (2014). Human platelet microRNA-mRNA networks associated with age and gender revealed by integrated plateletomics. Blood, 123(16), e37–e45. https://doi.org/10.1182/blood-2013-12-544692.
Chen, C. H., Lo, R. W., Urban, D., Pluthero, F. G., & Kahr, W. H. (2017). α-Granule biogenesis: from disease to discovery. Platelets, 28(2), 147–154. https://doi.org/10.1080/09537104.2017.1280599.
Ho, Y. H., Del Toro, R., Rivera-Torres, J., Rak, J., Korn, C., García-García, A., Macías, D., González-Gómez, C., Del Monte, A., Wittner, M., Waller, A. K., Foster, H. R., López-Otín, C., Johnson, R. S., Nerlov, C., Ghevaert, C., Vainchenker, W., Louache, F., Andrés, V., & Méndez-Ferrer, S. (2019). Remodeling of bone marrow hematopoietic stem cell niches promotes myeloid cell expansion during premature or physiological aging. Cell stem cell, 25(3), 407–418.e6. https://doi.org/10.1016/j.stem.2019.06.007.
Price, J., Lord, J. M., & Harrison, P. (2020). Inflammaging and platelet hyperreactivity: a new therapeutic target? Journal of thrombosis and haemostasis : JTH, 18(1), 3–5. https://doi.org/10.1111/jth.14670.
Vogler, M., Hamali, H. A., Sun, X. M., Bampton, E. T., Dinsdale, D., Snowden, R. T., Dyer, M. J., Goodall, A. H., & Cohen, G. M. (2011). BCL2/BCL-X(L) inhibition induces apoptosis, disrupts cellular calcium homeostasis, and prevents platelet activation. Blood, 117(26), 7145–7154. https://doi.org/10.1182/blood-2011-03-344812.
van der Meijden, P., & Heemskerk, J. (2019). Platelet biology and functions: new concepts and clinical perspectives. Nature reviews. Cardiology, 16(3), 166–179. https://doi.org/10.1038/s41569-018-0110-0.
Lippi, G., Meschi, T., & Borghi, L. (2012). Mean platelet volume increases with aging in a large population study. Thrombosis research, 129(4), e159–e160. https://doi.org/10.1016/j.thromres.2011.12.031.
Tian, J., Cheng, L. H., Cui, X., Lei, X. X., Tang, J. B., & Cheng, B. (2019). Investigating the effect of age on platelet ultrastructure using transmission electron microscopy. International wound journal, 16(6), 1457–1463. https://doi.org/10.1111/iwj.13214.
van der Bom, J. G., Heckbert, S. R., Lumley, T., Holmes, C. E., Cushman, M., Folsom, A. R., Rosendaal, F. R., & Psaty, B. M. (2009). Platelet count and the risk for thrombosis and death in the elderly. Journal of thrombosis and haemostasis : JTH, 7(3), 399–405. https://doi.org/10.1111/j.1538-7836.2008.03267.x.
Segal, J. B., & Moliterno, A. R. (2006). Platelet counts differ by sex, ethnicity, and age in the United States. Annals of epidemiology, 16(2), 123–130. https://doi.org/10.1016/j.annepidem.2005.06.052.
Biino, G., Santimone, I., Minelli, C., Sorice, R., Frongia, B., Traglia, M., Ulivi, S., Di Castelnuovo, A., Gögele, M., Nutile, T., Francavilla, M., Sala, C., Pirastu, N., Cerletti, C., Iacoviello, L., Gasparini, P., Toniolo, D., Ciullo, M., Pramstaller, P., Pirastu, M., et al. (2013). Age- and sex-related variations in platelet count in Italy: a proposal of reference ranges based on 40987 subjects’ data. PloS one, 8(1), e54289. https://doi.org/10.1371/journal.pone.0054289.
Jørgensen, K. A., Dyerberg, J., Olesen, A. S., & Stoffersen, E. (1980). Acetylsalicylic acid, bleeding time and age. Thrombosis research, 19(6), 799–805. https://doi.org/10.1016/0049-3848(80)90007-9.
Kuhnla, A., Reinthaler, M., Braune, S., Maier, A., Pindur, G., Lendlein, A., & Jung, F. (2019). Spontaneous and induced platelet aggregation in apparently healthy subjects in relation to age. Clinical hemorheology and microcirculation, 71(4), 425–435. https://doi.org/10.3233/CH-199006.
Gleerup, G., & Winther, K. (1995). The effect of ageing on platelet function and fibrinolytic activity. Angiology, 46(8), 715–718. https://doi.org/10.1177/000331979504600810.
Winther, K., & Naesh, O. (1987). Aging and platelet beta-adrenoceptor function. European journal of pharmacology, 136(2), 219–223. https://doi.org/10.1016/0014-2999(87)90713-8.
Reilly, I. A., & FitzGerald, G. A. (1986). Eicosenoid biosynthesis and platelet function with advancing age. Thrombosis research, 41(4), 545–554. https://doi.org/10.1016/0049-3848(86)91700-7.
Driver, J. A., Djoussé, L., Logroscino, G., Gaziano, J. M., & Kurth, T. (2008). Incidence of cardiovascular disease and cancer in advanced age: prospective cohort study. BMJ (Clinical research ed.), 337, a2467. https://doi.org/10.1136/bmj.a2467.
Vericel, E., Croset, M., Sedivy, P., Courpron, P., Dechavanne, M., & Lagarde, M. (1988). Platelets and aging. I--aggregation, arachidonate metabolism and antioxidant status. Thrombosis research, 49(3), 331–342. https://doi.org/10.1016/0049-3848(88)90313-1.
Gleerup, G., & Winther, K. (1988). The effect of ageing on human platelet sensitivity to serotonin. European journal of clinical investigation, 18(5), 504–506. https://doi.org/10.1111/j.1365-2362.1988.tb01047.x.
Hamilton, P. J., Dawson, A. A., Ogston, D., & Douglas, A. S. (1974). The effect of age on the fibrinolytic enzyme system. Journal of clinical pathology, 27(4), 326–329. https://doi.org/10.1136/jcp.27.4.326.
Yamamoto, K., Takeshita, K., Kojima, T., Takamatsu, J., & Saito, H. (2005). Aging and plasminogen activator inhibitor-1 (PAI-1) regulation: implication in the pathogenesis of thrombotic disorders in the elderly. Cardiovascular research, 66(2), 276–285. https://doi.org/10.1016/j.cardiores.2004.11.013.
Kasjanovová, D., & Baláz, V. (1986). Age-related changes in human platelet function in vitro. Mechanisms of ageing and development, 37(2), 175–182. https://doi.org/10.1016/0047-6374(86)90074-6.
Bastyr 3rd, E. J., Kadrofske, M. M., & Vinik, A. I. (1990). Platelet activity and phosphoinositide turnover increase with advancing age. The American journal of medicine, 88(6), 601–606. https://doi.org/10.1016/0002-9343(90)90525-i.
Hernández, B., Fuentes, E., Palomo, I., & Alarcón, M. (2019). Increased platelet function during frailty. Experimental hematology, 77, 12–25.e2. https://doi.org/10.1016/j.exphem.2019.08.006.
Fuentes, E., & Palomo, I. (2016). Role of oxidative stress on platelet hyperreactivity during aging. Life sciences, 148, 17–23. https://doi.org/10.1016/j.lfs.2016.02.026.
Jain, K., Tyagi, T., Patell, K., Xie, Y., Kadado, A. J., Lee, S. H., Yarovinsky, T., Du, J., Hwang, J., Martin, K. A., Testani, J., Ionescu, C. N., & Hwa, J. (2019). Age associated non-linear regulation of redox homeostasis in the anucleate platelet: implications for CVD risk patients. EBioMedicine, 44, 28–40. https://doi.org/10.1016/j.ebiom.2019.05.022.
Begonja, A. J., Gambaryan, S., Geiger, J., Aktas, B., Pozgajova, M., Nieswandt, B., & Walter, U. (2005). Platelet NAD(P)H-oxidase-generated ROS production regulates alphaIIbbeta3-integrin activation independent of the NO/cGMP pathway. Blood, 106(8), 2757–2760. https://doi.org/10.1182/blood-2005-03-1047.
Gekas, C., & Graf, T. (2013). CD41 expression marks myeloid-biased adult hematopoietic stem cells and increases with age. Blood, 121(22), 4463–4472. https://doi.org/10.1182/blood-2012-09-457929.
Shih, L., Sanders, N., Rondina, S., Weyrich, A., Carlisle, M., Men, S., Supiano, M., Zimmerman, G., & Rondina, M. (2015). Activated platelet integrin alpha IIb Beta3 is increased in older adults with frailty but not in healthy aging. Journal of the American Geriatrics Society, 63, 110-S.
Yang, J., Zhou, X., Fan, X., Xiao, M., Yang, D., Liang, B., Dai, M., Shan, L., Lu, J., Lin, Z., Liu, R., Liu, J., Wang, L., Zhong, M., Jiang, Y., & Bai, X. (2016). mTORC1 promotes aging-related venous thrombosis in mice via elevation of platelet volume and activation. Blood, 128(5), 615–624. https://doi.org/10.1182/blood-2015-10-672964.
Campbell, R. A., Franks, Z., Bhatnagar, A., Rowley, J. W., Manne, B. K., Supiano, M. A., Schwertz, H., Weyrich, A. S., & Rondina, M. T. (2018). Granzyme A in human platelets regulates the synthesis of proinflammatory cytokines by monocytes in aging. Journal of immunology (Baltimore, Md.:1950), 200(1), 295–304. https://doi.org/10.4049/jimmunol.1700885.
Prisco, D., Rogasi, P. G., Matucci, M., Paniccia, R., Abbate, R., Gensini, G. F., & Serneri, G. G. (1986). Age related changes in platelet lipid composition. Thrombosis research, 44(4), 427–437. https://doi.org/10.1016/0049-3848(86)90321-x.
Bai, M. Y., Chuang, M. H., Lin, M. F., Tang, S. L., Wong, C. C., & Chan, W. P. (2018). Relationships of age and sex with cytokine content and distribution in human platelet fibrin gels. Scientific reports, 8(1), 10642. https://doi.org/10.1038/s41598-018-28376-z.
Evanson, J. R., Guyton, M. K., Oliver, D. L., Hire, J. M., Topolski, R. L., Zumbrun, S. D., McPherson, J. C., & Bojescul, J. A. (2014). Gender and age differences in growth factor concentrations from platelet-rich plasma in adults. Military medicine, 179(7), 799–805. https://doi.org/10.7205/MILMED-D-13-00336.
Marinho, C. F., Costa-Maia, J., Pinto-de-Barros, J., & Oliveira, C. R. (1997). Correlation between human platelet cytoplasmic membrane outer leaflet fluidity, Na+/H+ exchanger activity and aging. European archives of psychiatry and clinical neuroscience, 247(5), 275–277. https://doi.org/10.1007/BF02900306.
Alexandru, N., Constantin, A., & Popov, D. (2008). Carbonylation of platelet proteins occurs as consequence of oxidative stress and thrombin activation, and is stimulated by ageing and type 2 diabetes. Clinical chemistry and laboratory medicine, 46(4), 528–536. https://doi.org/10.1515/CCLM.2008.104.
Michimata, T., Imamura, M., Mizuma, H., Murakami, M., & Iriuchijima, T. (1996). Sex and age differences in soluble guanylate cyclase activity in human platelets. Life sciences, 58(5), 415–419. https://doi.org/10.1016/0024-3205(95)02306-2.
Origlia, C., Pescarmona, G., Capizzi, A., Cogotti, S., Gambino, R., Cassader, M., Benso, A., Granata, R., & Martina, V. (2004). Platelet cGMP inversely correlates with age in healthy subjects. Journal of endocrinological investigation, 27(2), RC1–RC4. https://doi.org/10.1007/BF03346251.
Kawamoto, E. M., Munhoz, C. D., Glezer, I., Bahia, V. S., Caramelli, P., Nitrini, R., Gorjão, R., Curi, R., Scavone, C., & Marcourakis, T. (2005). Oxidative state in platelets and erythrocytes in aging and Alzheimer’s disease. Neurobiology of aging, 26(6), 857–864. https://doi.org/10.1016/j.neurobiolaging.2004.08.011.
Goubareva, I., Gkaliagkousi, E., Shah, A., Queen, L., Ritter, J., & Ferro, A. (2007). Age decreases nitric oxide synthesis and responsiveness in human platelets and increases formation of monocyte-platelet aggregates. Cardiovascular research, 75(4), 793–802. https://doi.org/10.1016/j.cardiores.2007.05.021.
Vericel, E., Rey, C., Calzada, C., Haond, P., Chapuy, P. H., & Lagarde, M. (1992). Age-related changes in arachidonic acid peroxidation and glutathione-peroxidase activity in human platelets. Prostaglandins, 43(1), 75–85. https://doi.org/10.1016/0090-6980(92)90067-4.
Rossi, D. J., Bryder, D., Seita, J., Nussenzweig, A., Hoeijmakers, J., & Weissman, I. L. (2007). Deficiencies in DNA damage repair limit the function of haematopoietic stem cells with age. Nature, 447(7145), 725–729. https://doi.org/10.1038/nature05862.
Rundberg Nilsson, A., Soneji, S., Adolfsson, S., Bryder, D., & Pronk, C. J. (2016). Human and murine hematopoietic stem cell aging is associated with functional impairments and intrinsic megakaryocytic/erythroid bias. PloS one, 11(7), e0158369. https://doi.org/10.1371/journal.pone.0158369.
Montenont, E., Rondina, M. T., & Campbell, R. A. (2019). Altered functions of platelets during aging. Current opinion in hematology, 26(5), 336–342. https://doi.org/10.1097/MOH.0000000000000526.
Macpherson, C. R., & Jacobs, P. (1987). Bleeding time decreases with age. Arch Pathol Lab Med., 111(4), 328–329.
Arauna, D., García, F., Rodríguez-Mañas, L., Marrugat, J., Sáez, C., Alarcón, M., Wehinger, S., Espinosa-Parrilla, Y., Palomo, I., & Fuentes, E. (2020). Older adults with frailty syndrome present an altered platelet function and an increased level of circulating oxidative stress and mitochondrial dysfunction biomarker GDF-15. Free radical biology & medicine, 149, 64–71. https://doi.org/10.1016/j.freeradbiomed.2020.01.007.
Suehiro, A., Uedaa, M., Suehiroh, M., Ohe, Y., & Kakishitaa, E. (1995). Evaluation of platelet hyperfunction in aged subjects using spontaneous platelet aggregation in whole blood. Archives of gerontology and geriatrics, 21(3), 277–283. https://doi.org/10.1016/0167-4943(95)00664-7.
Vilén, L., Jacobsson, S., Wadenvik, H., & Kutti, J. (1989). ADP-induced platelet aggregation as a function of age in healthy humans. Thrombosis and haemostasis, 61(3), 490–492.
Davizon-Castillo, P., McMahon, B., Aguila, S., Bark, D., Ashworth, K., Allawzi, A., Campbell, R. A., Montenont, E., Nemkov, T., D'Alessandro, A., Clendenen, N., Shih, L., Sanders, N. A., Higa, K., Cox, A., Padilla-Romo, Z., Hernandez, G., Wartchow, E., Trahan, G. D., Nozik-Grayck, E., et al. (2019). TNF-α-driven inflammation and mitochondrial dysfunction define the platelet hyperreactivity of aging. Blood, 134(9), 727–740. https://doi.org/10.1182/blood.2019000200.
Dayal, S., Wilson, K. M., Motto, D. G., Miller Jr., F. J., Chauhan, A. K., & Lentz, S. R. (2013). Hydrogen peroxide promotes aging-related platelet hyperactivation and thrombosis. Circulation, 127(12), 1308–1316. https://doi.org/10.1161/CIRCULATIONAHA.112.000966.
Paul, M., Hemshekhar, M., Kemparaju, K., & Girish, K. S. (2019). Berberine mitigates high glucose-potentiated platelet aggregation and apoptosis by modulating aldose reductase and NADPH oxidase activity. Free radical biology & medicine, 130, 196–205. https://doi.org/10.1016/j.freeradbiomed.2018.10.453.
Li, J., Zhao, R., Zhao, H., Chen, G., Jiang, Y., Lyu, X., & Wu, T. (2019). Reduction of aging-induced oxidative stress and activation of autophagy by bilberry anthocyanin supplementation via the AMPK-mTOR signaling pathway in aged female rats. Journal of agricultural and food chemistry, 67(28), 7832–7843. https://doi.org/10.1021/acs.jafc.9b02567.
Fuhler, G. M., Tyl, M. R., Olthof, S. G., Lyndsay Drayer, A., Blom, N., & Vellenga, E. (2009). Distinct roles of the mTOR components Rictor and Raptor in MO7e megakaryocytic cells. European journal of haematology, 83(3), 235–245. https://doi.org/10.1111/j.1600-0609.2009.01263.x.
Haynes, A., Linden, M. D., Robey, E., Naylor, L. H., Cox, K. L., Lautenschlager, N. T., & Green, D. J. (2017). Relationship between monocyte-platelet aggregation and endothelial function in middle-aged and elderly adults. Physiological reports, 5(10), e13189 https://doi.org/10.14814/phy2.13189.
Di Serafino, L., Sarma, J., Dierickx, K., Ntarladimas, I., Pyxaras, S. A., Delrue, L., De Bruyne, B., Wijns, W., Barbato, E., & Bartunek, J. (2014). Monocyte-platelets aggregates as cellular biomarker of endothelium-dependent coronary vasomotor dysfunction in patients with coronary artery disease. Journal of cardiovascular translational research, 7(1), 1–8. https://doi.org/10.1007/s12265-013-9520-x.
Hilt, Z. T., Pariser, D. N., Ture, S. K., Mohan, A., Quijada, P., Asante, A. A., Cameron, S. J., Sterling, J. A., Merkel, A. R., Johanson, A. L., Jenkins, J. L., Small, E. M., McGrath, K. E., Palis, J., Elliott, M. R., & Morrell, C. N. (2019). Platelet-derived β2M regulates monocyte inflammatory responses. JCI insight, 4(5), e122943. https://doi.org/10.1172/jci.insight.122943.
Shi, S., Qin, M., Shen, B., Cai, Y., Liu, T., Yang, F., Gong, W., Liu, X., Liang, J., Zhao, Q., Huang, H., Yang, B., & Huang, C. (2020). Association of cardiac injury with mortality in hospitalized patients with COVID-19 in Wuhan, China. JAMA cardiology, e200950. Advance online publication. https://doi.org/10.1001/jamacardio.2020.0950.
Schäfer, A., Alp, N. J., Cai, S., Lygate, C. A., Neubauer, S., Eigenthaler, M., Bauersachs, J., & Channon, K. M. (2004). Reduced vascular NO bioavailability in diabetes increases platelet activation in vivo. Arteriosclerosis, thrombosis, and vascular biology, 24(9), 1720–1726. https://doi.org/10.1161/01.ATV.0000138072.76902.dd.
Lippi, G., Plebani, M., & Henry, B. M. (2020). Thrombocytopenia is associated with severe coronavirus disease 2019 (COVID-19) infections: a meta-analysis. Clinica chimica acta; international journal of clinical chemistry, 506, 145–148. https://doi.org/10.1016/j.cca.2020.03.022.
Salamanna, F., Maglio, M., Landini, M. P., & Fini, M. (2020). Platelet functions and activities as potential hematologic parameters related to coronavirus disease 2019 (Covid-19). Platelets, 1–6. Advance online publication. https://doi.org/10.1080/09537104.2020.1762852.
Sgonc, R., & Gruber, J. (2013). Age-related aspects of cutaneous wound healing: a mini-review. Gerontology, 59(2), 159–164. https://doi.org/10.1159/000342344.
Waehre, T., Damås, J. K., Yndestad, A., Taskén, K., Pedersen, T. M., Smith, C., Halvorsen, B., Frøland, S. S., Solum, N. O., & Aukrust, P. (2004). Effect of activated platelets on expression of cytokines in peripheral blood mononuclear cells - potential role of prostaglandin E2. Thrombosis and haemostasis, 92(6), 1358–1367. https://doi.org/10.1160/TH04-03-0146.
Nami, N., Feci, L., Napoliello, L., Giordano, A., Lorenzini, S., Galeazzi, M., Rubegni, P., & Fimiani, M. (2016). Crosstalk between platelets and PBMC: new evidence in wound healing. Platelets, 27(2), 143–148. https://doi.org/10.3109/09537104.2015.1048216.
Scopelliti, F., Cattani, C., Dimartino, V., Scarponi, C., Madonna, S., Albanesi, C., Costanzo, G., Mirisola, C., & Cavani, A. (2020). Platelet lysate promotes the expansion of T regulatory cells that favours in vitro wound healing by increasing keratinocyte migration and fibroblast production of extracellular matrix components. European journal of dermatology : EJD, 30(1), 3–11. https://doi.org/10.1684/ejd.2020.3711.
Blache, D., Gesquière, L., Loreau, N., & Durand, P. (1999). Oxidant stress: the role of nutrients in cell-lipoprotein interactions. The Proceedings of the Nutrition Society, 58(3), 559–563. https://doi.org/10.1017/s0029665199000737.
Cowman, J., Dunne, E., Oglesby, I., Byrne, B., Ralph, A., Voisin, B., Müllers, S., Ricco, A. J., & Kenny, D. (2015). Age-related changes in platelet function are more profound in women than in men. Scientific reports, 5, 12235. https://doi.org/10.1038/srep12235.
Farré, A. L., Modrego, J., & Zamorano-León, J. J. (2014). Effects of hormones on platelet aggregation. Hormone molecular biology and clinical investigation, 18(1), 27–36. https://doi.org/10.1515/hmbci-2013-0055.
Karolczak, K., Konieczna, L., Kostka, T., Witas, P. J., Soltysik, B., Baczek, T., & Watala, C. (2018). Testosterone and dihydrotestosterone reduce platelet activation and reactivity in older men and women. Aging, 10(5), 902–929 https://doi.org/10.18632/aging.101438.
Copplestone, J. A. (1987). Bleeding and coagulation disorders in the elderly. Bailliere’s clinical haematology, 1(2), 559–580. https://doi.org/10.1016/s0950-3536(87)80013-6.
Maleki, A., Roohafza, H., Rashidi, N., Aliyari, F., Ghanavati, R., Foroughi, S., Nabatchi, B., & Torkashvand, M. (2012). Determination of normal range of bleeding time in rural and urban residents of Borujerd, Iran: a pilot study. ARYA atherosclerosis, 8(3), 136–142.
Hossain, M. S., Hashimoto, M., Gamoh, S., & Masumura, S. (1999). Association of age-related decrease in platelet membrane fluidity with platelet lipid peroxide. Life sciences, 64(2), 135–143. https://doi.org/10.1016/s0024-3205(98)00543-8.
Rabani, V., Montange, D., Meneveau, N., & Davani, S. (2018). Impact of ticagrelor on P2Y1 and P2Y12 localization and on cholesterol levels in platelet plasma membrane. Platelets, 29(7), 709–715. https://doi.org/10.1080/09537104.2017.1356453.
Renaud, S., De Backer, G., Thevenon, C., Joossens, J. V., Vermylen, J., Kornitzer, M., & Verstraete, M. (1991). Platelet fatty acids and function in two distinct regions of Belgium: relationship to age and dietary habits. Journal of internal medicine, 229(1), 79–88. https://doi.org/10.1111/j.1365-2796.1991.tb00310.x.
Kanikarla-Marie, P., Kopetz, S., Hawk, E. T., Millward, S. W., Sood, A. K., Gresele, P., Overman, M., Honn, K., & Menter, D. G. (2018). Bioactive lipid metabolism in platelet “first responder” and cancer biology. Cancer metastasis reviews, 37(2-3), 439–454. https://doi.org/10.1007/s10555-018-9755-8.
Friedman, E. A., Ogletree, M. L., Haddad, E. V., & Boutaud, O. (2015). Understanding the role of prostaglandin E2 in regulating human platelet activity in health and disease. Thrombosis research, 136(3), 493–503. https://doi.org/10.1016/j.thromres.2015.05.027.
Brock, T. G., McNish, R. W., & Peters-Golden, M. (1999). Arachidonic acid is preferentially metabolized by cyclooxygenase-2 to prostacyclin and prostaglandin E2. The Journal of biological chemistry, 274(17), 11660–11666. https://doi.org/10.1074/jbc.274.17.11660.
Dennis, E. A., & Norris, P. C. (2015). Eicosanoid storm in infection and inflammation. Nature reviews. Immunology, 15(8), 511–523. https://doi.org/10.1038/nri3859.
Tang, K., Cai, Y., Joshi, S., Tovar, E., Tucker, S. C., Maddipati, K. R., Crissman, J. D., Repaskey, W. T., & Honn, K. V. (2015). Convergence of eicosanoid and integrin biology: 12-lipoxygenase seeks a partner. Molecular cancer, 14, 111. https://doi.org/10.1186/s12943-015-0382-5.
Lau, S., Rangarajan, R., Krüger-Genge, A., Braune, S., Küpper, J. H., Lendlein, A., & Jung, F. (2020). Age-related morphology and function of human arterial endothelial cells. Clinical hemorheology and microcirculation, 74(1), 93–107. https://doi.org/10.3233/CH-199238.
Scott, N. M., Ng, R. L., Gorman, S., Norval, M., Waithman, J., & Hart, P. H. (2014). Prostaglandin E2 imprints a long-lasting effect on dendritic cell progenitors in the bone marrow. Journal of leukocyte biology, 95(2), 225–232. https://doi.org/10.1189/jlb.0513294.
Kim, J. W., Zou, Y., Yoon, S., Lee, J. H., Kim, Y. K., Yu, B. P., & Chung, H. Y. (2004). Vascular aging: molecular modulation of the prostanoid cascade by calorie restriction. The journals of gerontology. Series A, Biological sciences and medical sciences, 59(9), B876–B885. https://doi.org/10.1093/gerona/59.9.b876.
Li, Y., Lei, D., Swindell, W. R., Xia, W., Weng, S., Fu, J., Worthen, C. A., Okubo, T., Johnston, A., Gudjonsson, J. E., Voorhees, J. J., & Fisher, G. J. (2015). Age-associated increase in skin fibroblast-derived prostaglandin E2 contributes to reduced collagen levels in elderly human skin. The Journal of investigative dermatology, 135(9), 2181–2188. https://doi.org/10.1038/jid.2015.157.
Tokunaga, O., Yamada, T., Fan, J. L., & Watanabe, T. (1991). Age-related decline in prostacyclin synthesis by human aortic endothelial cells. Qualitative and quantitative analysis. The American journal of pathology, 138(4), 941–949.
Choi, J. H., & Yu, B. P. (1998). The effects of dietary restriction on age-related changes in rat serum prostaglandins. The journal of nutrition, health & aging, 2(3), 138–142.
Simeone, P., Boccatonda, A., Liani, R., & Santilli, F. (2018). Significance of urinary 11-dehydro-thromboxane B2 in age-related diseases: Focus on atherothrombosis. Ageing research reviews, 48, 51–78. https://doi.org/10.1016/j.arr.2018.09.004.
Santilli, F., Zaccardi, F., Liani, R., Petrucci, G., Simeone, P., Pitocco, D., Tripaldi, R., Rizzi, A., Formoso, G., Pontecorvi, A., Angelucci, E., Pagliaccia, F., Golato, M., De Leva, F., Vitacolonna, E., Rocca, B., Consoli, A., & Patrono, C. (2020). In vivo thromboxane-dependent platelet activation is persistently enhanced in subjects with impaired glucose tolerance. Diabetes/metabolism research and reviews, 36(2), e3232. https://doi.org/10.1002/dmrr.3232.
Samad, F., & Ruf, W. (2013). Inflammation, obesity, and thrombosis. Blood, 122(20), 3415–3422. https://doi.org/10.1182/blood-2013-05-427708.
Simeone, P., Liani, R., Tripaldi, R., Di Castelnuovo, A., Guagnano, M. T., Tartaro, A., Bonadonna, R. C., Federico, V., Cipollone, F., Consoli, A., & Santilli, F. (2018). Thromboxane-dependent platelet activation in obese subjects with prediabetes or early type 2 diabetes: effects of liraglutide- or lifestyle changes-induced weight loss. Nutrients, 10(12), 1872. https://doi.org/10.3390/nu10121872.
Zhao, Z., Hu, J., Gao, X., Liang, H., Yu, H., Liu, S., & Liu, Z. (2017). Hyperglycemia via activation of thromboxane A2 receptor impairs the integrity and function of blood-brain barrier in microvascular endothelial cells. Oncotarget, 8(18), 30030–30038. https://doi.org/10.18632/oncotarget.16273.
Mohanty, I., Singh, J., & Rattan, S. (2019). Downregulation of thromboxane A2 and angiotensin II type 1 receptors associated with aging-related decrease in internal anal sphincter tone. Scientific reports, 9(1), 6759. https://doi.org/10.1038/s41598-019-42894-4.
Wéra, O., Lecut, C., Servais, L., Hego, A., Delierneux, C., Jiang, Z., Keutgens, A., Evans, R. J., Delvenne, P., Lancellotti, P., & Oury, C. (2020). P2X1 ion channel deficiency causes massive bleeding in inflamed intestine and increases thrombosis. Journal of thrombosis and haemostasis: JTH, 18(1), 44–56. https://doi.org/10.1111/jth.14620.
Vinholt, P. J., Alnor, A., Nybo, M., & Hvas, A. M. (2016). Prediction of bleeding and prophylactic platelet transfusions in cancer patients with thrombocytopenia. Platelets, 27(6), 547–554. https://doi.org/10.3109/09537104.2015.1131814.
Fasano, R. M., & Josephson, C. D. (2015). Platelet transfusion goals in oncology patients. Hematology. American Society of Hematology. Education Program, 2015, 462–470. https://doi.org/10.1182/asheducation-2015.1.462.
Liebman, H. A. (2014). Thrombocytopenia in cancer patients. Thrombosis research, 133(Suppl 2), S63–S69. https://doi.org/10.1016/S0049-3848(14)50011-4.
Willoughby, S., Holmes, A., & Loscalzo, J. (2002). Platelets and cardiovascular disease. European journal of cardiovascular nursing : journal of the Working Group on Cardiovascular Nursing of the European Society of Cardiology, 1(4), 273–288. https://doi.org/10.1016/s1474-5151(02)00038-5.
Cooke, N. M., Egan, K., McFadden, S., Grogan, L., Breathnach, O. S., O'Leary, J., Hennessy, B. T., & Kenny, D. (2013). Increased platelet reactivity in patients with late-stage metastatic cancer. Cancer medicine, 2(4), 564–570. https://doi.org/10.1002/cam4.86.
Aronson, D., & Brenner, B. (2018). Arterial thrombosis and cancer. Thrombosis research, 164(Suppl 1), S23–S28. https://doi.org/10.1016/j.thromres.2018.01.003.
Navi, B. B., Reiner, A. S., Kamel, H., Iadecola, C., Okin, P. M., Elkind, M., Panageas, K. S., & DeAngelis, L. M. (2017). Risk of arterial thromboembolism in patients with cancer. Journal of the American College of Cardiology, 70(8), 926–938. https://doi.org/10.1016/j.jacc.2017.06.047.
Trousseau, A. (1865). Phlegmasia alba dolens. Clinique Medicale de l’Hotel-Dieu de Paris (Lectures on Clinical Medicine at the Hotel-Dieu, Paris), 3, 654–712.
Khorana, A. A., Carrier, M., Garcia, D. A., & Lee, A. Y. (2016). Guidance for the prevention and treatment of cancer-associated venous thromboembolism. Journal of thrombosis and thrombolysis, 41(1), 81–91. https://doi.org/10.1007/s11239-015-1313-4.
Park, H., Jeong, C. W., Yuk, H., Ku, J. H., Kim, H. H., Kwak, C., & Kim, I. (2019). Influence of tumor thrombus on occurrence of distant venous thromboembolism and survival in patients with renal cell carcinoma after surgery. Clinical and applied thrombosis/hemostasis : official journal of the International Academy of Clinical and Applied Thrombosis/Hemostasis, 25, 1076029618823288. https://doi.org/10.1177/1076029618823288.
Sahara, K., Paredes, A. Z., Merath, K., Tsilimigras, D. I., Bagante, F., Ratti, F., Marques, H. P., Soubrane, O., Beal, E. W., Lam, V., Poultsides, G. A., Popescu, I., Alexandrescu, S., Martel, G., Aklile, W., Guglielmi, A., Hugh, T., Aldrighetti, L., Endo, I., & Pawlik, T. M. (2020). Evaluation of the ACS NSQIP surgical risk calculator in elderly patients undergoing hepatectomy for hepatocellular carcinoma. Journal of gastrointestinal surgery : official journal of the Society for Surgery of the Alimentary Tract, 24(3), 551–559. https://doi.org/10.1007/s11605-019-04174-4.
Chen, Y., Tsang, Y. S., Chou, X., Hu, J., & Xia, Q. (2019). A lung cancer patient with deep vein thrombosis:a case report and literature review. BMC cancer, 19(1), 285. https://doi.org/10.1186/s12885-019-5513-8.
Marshall-Webb, M., Bright, T., Price, T., Thompson, S. K., & Watson, D. I. (2017). Venous thromboembolism in patients with esophageal or gastric cancer undergoing neoadjuvant chemotherapy. Diseases of the esophagus : official journal of the International Society for Diseases of the Esophagus, 30(2), 1–7. https://doi.org/10.1111/dote.12516.
Oge, T., Yalcin, O. T., Ozalp, S. S., & Isikci, T. (2013). Platelet volume as a parameter for platelet activation in patients with endometrial cancer. Journal of obstetrics and gynaecology : the journal of the Institute of Obstetrics and Gynaecology, 33(3), 301–304. https://doi.org/10.3109/01443615.2012.758089.
Pyo, J. S., Sohn, J. H., & Kang, G. (2016). Diagnostic and prognostic roles of the mean platelet volume in malignant tumors: a systematic review and meta-analysis. Platelets, 27(8), 722–728. https://doi.org/10.3109/09537104.2016.1169265.
Riedl, J., Hell, L., Kaider, A., Koder, S., Marosi, C., Zielinski, C., Panzer, S., Pabinger, I., & Ay, C. (2016). Association of platelet activation markers with cancer-associated venous thromboembolism. Platelets, 27(1), 80–85. https://doi.org/10.3109/09537104.2015.1041901.
Díaz, J. M., Boietti, B. R., Vazquez, F. J., Waisman, G. D., Giunta, D. H., Rojas, L. P., Peuchot, V., & Posadas-Martínez, M. L. (2019). Mean platelet volume as a prognostic factor for venous thromboembolic disease. Revista medica de Chile, 147(2), 145–152. https://doi.org/10.4067/s0034-98872019000200145.
van Es, N., Louzada, M., Carrier, M., Tagalakis, V., Gross, P. L., Shivakumar, S., Rodger, M. A., & Wells, P. S. (2018). Predicting the risk of recurrent venous thromboembolism in patients with cancer: a prospective cohort study. Thrombosis research, 163, 41–46. https://doi.org/10.1016/j.thromres.2018.01.009.
Ratib, S., Walker, A. J., Card, T. R., & Grainge, M. J. (2016). Risk of venous thromboembolism in hospitalised cancer patients in England-a cohort study. Journal of hematology & oncology, 9(1), 60. https://doi.org/10.1186/s13045-016-0291-0.
Kassassir, H., Karolczak, K., Siewiera, K. M., Wojkowska, D. W., Braun, M., & Watala, C. W. (2020). Time-dependent interactions of blood platelets and cancer cells, accompanied by extramedullary hematopoiesis, lead to increased platelet activation and reactivity in a mouse orthotopic model of breast cancer - implications for pulmonary and liver metastasis. Aging, 12(6), 5091–5120. https://doi.org/10.18632/aging.102933.
Haemmerle, M., Stone, R. L., Menter, D. G., Afshar-Kharghan, V., & Sood, A. K. (2018). The platelet lifeline to cancer: challenges and opportunities. Cancer cell, 33(6), 965–983. https://doi.org/10.1016/j.ccell.2018.03.002.
Adachi, I., Takahashi, M., Adachi, H., & Horikoshi, I. (1986). EL-4 tumor cell-induced human and rabbit platelet aggregations. Cell structure and function, 11(2), 157–163. https://doi.org/10.1247/csf.11.157.
Mitrugno, A., Tassi Yunga, S., Sylman, J. L., Zilberman-Rudenko, J., Shirai, T., Hebert, J. F., Kayton, R., Zhang, Y., Nan, X., Shatzel, J. J., Esener, S., Duvernay, M. T., Hamm, H. E., Gruber, A., Williams, C. D., Takata, Y., Armstrong, R., Morgan, T. K., & McCarty, O. (2019). The role of coagulation and platelets in colon cancer-associated thrombosis. American journal of physiology. Cell physiology, 316(2), C264–C273. https://doi.org/10.1152/ajpcell.00367.2018.
Sol, N., & Wurdinger, T. (2017). Platelet RNA signatures for the detection of cancer. Cancer metastasis reviews, 36(2), 263–272. https://doi.org/10.1007/s10555-017-9674-0.
Best, M. G., Wesseling, P., & Wurdinger, T. (2018). Tumor-educated platelets as a noninvasive biomarker source for cancer detection and progression monitoring. Cancer research, 78(13), 3407–3412. https://doi.org/10.1158/0008-5472.CAN-18-0887.
Gomes, F. G., Sandim, V., Almeida, V. H., Rondon, A., Succar, B. B., Hottz, E. D., Leal, A. C., Verçoza, B., Rodrigues, J., Bozza, P. T., Zingali, R. B., & Monteiro, R. Q. (2017). Breast-cancer extracellular vesicles induce platelet activation and aggregation by tissue factor-independent and -dependent mechanisms. Thrombosis research, 159, 24–32. https://doi.org/10.1016/j.thromres.2017.09.019.
Labelle, M., Begum, S., & Hynes, R. O. (2014). Platelets guide the formation of early metastatic niches. Proceedings of the National Academy of Sciences of the United States of America, 111(30), E3053–E3061. https://doi.org/10.1073/pnas.1411082111.
Andrade, S. S., Sumikawa, J. T., Castro, E. D., Batista, F. P., Paredes-Gamero, E., Oliveira, L. C., Guerra, I. M., Peres, G. B., Cavalheiro, R. P., Juliano, L., Nazário, A. P., Facina, G., Tsai, S. M., Oliva, M. L., & Girão, M. J. (2017). Interface between breast cancer cells and the tumor microenvironment using platelet-rich plasma to promote tumor angiogenesis - influence of platelets and fibrin bundles on the behavior of breast tumor cells. Oncotarget, 8(10), 16851–16874. https://doi.org/10.18632/oncotarget.15170.
Faria, A., Andrade, S. S., Reijm, A. N., Spaander, M., de Maat, M., Peppelenbosch, M. P., Ferreira-Halder, C. V., & Fuhler, G. M. (2019). Targeting tyrosine phosphatases by 3-bromopyruvate overcomes hyperactivation of platelets from gastrointestinal cancer patients. Journal of clinical medicine, 8(7), 936. https://doi.org/10.3390/jcm8070936.
Mezouar, S., Darbousset, R., Dignat-George, F., Panicot-Dubois, L., & Dubois, C. (2015). Inhibition of platelet activation prevents the P-selectin and integrin-dependent accumulation of cancer cell microparticles and reduces tumor growth and metastasis in vivo. International journal of cancer, 136(2), 462–475. https://doi.org/10.1002/ijc.28997.
Lin, R. J., Afshar-Kharghan, V., & Schafer, A. I. (2014). Paraneoplastic thrombocytosis: the secrets of tumor self-promotion. Blood, 124(2), 184–187. https://doi.org/10.1182/blood-2014-03-562538.
Fidler, I. J. (2003). The pathogenesis of cancer metastasis: the ‘seed and soil’ hypothesis revisited. Nature reviews. Cancer, 3(6), 453–458. https://doi.org/10.1038/nrc1098.
Medina, C., Jurasz, P., Santos-Martinez, M. J., Jeong, S. S., Mitsky, T., Chen, R., & Radomski, M. W. (2006). Platelet aggregation-induced by caco-2 cells: regulation by matrix metalloproteinase-2 and adenosine diphosphate. The Journal of pharmacology and experimental therapeutics, 317(2), 739–745. https://doi.org/10.1124/jpet.105.098384.
Radziwon-Balicka, A., Santos-Martinez, M. J., Corbalan, J. J., O'Sullivan, S., Treumann, A., Gilmer, J. F., Radomski, M. W., & Medina, C. (2014). Mechanisms of platelet-stimulated colon cancer invasion: role of clusterin and thrombospondin 1 in regulation of the P38MAPK-MMP-9 pathway. Carcinogenesis, 35(2), 324–332. https://doi.org/10.1093/carcin/bgt332.
Labelle, M., Begum, S., & Hynes, R. O. (2011). Direct signaling between platelets and cancer cells induces an epithelial-mesenchymal-like transition and promotes metastasis. Cancer cell, 20(5), 576–590. https://doi.org/10.1016/j.ccr.2011.09.009.
Guo, Y., Cui, W., Pei, Y., & Xu, D. (2019). Platelets promote invasion and induce epithelial to mesenchymal transition in ovarian cancer cells by TGF-β signaling pathway. Gynecologic oncology, 153(3), 639–650. https://doi.org/10.1016/j.ygyno.2019.02.026.
Im, J. H., Fu, W., Wang, H., Bhatia, S. K., Hammer, D. A., Kowalska, M. A., & Muschel, R. J. (2004). Coagulation facilitates tumor cell spreading in the pulmonary vasculature during early metastatic colony formation. Cancer research, 64(23), 8613–8619. https://doi.org/10.1158/0008-5472.CAN-04-2078.
Schumacher, D., Strilic, B., Sivaraj, K. K., Wettschureck, N., & Offermanns, S. (2013). Platelet-derived nucleotides promote tumor-cell transendothelial migration and metastasis via P2Y2 receptor. Cancer cell, 24(1), 130–137. https://doi.org/10.1016/j.ccr.2013.05.008.
Leblanc, R., & Peyruchaud, O. (2016). Metastasis: new functional implications of platelets and megakaryocytes. Blood, 128(1), 24–31. https://doi.org/10.1182/blood-2016-01-636399.
Tímár, J., Tóvári, J., Rásó, E., Mészáros, L., Bereczky, B., & Lapis, K. (2005). Platelet-mimicry of cancer cells: epiphenomenon with clinical significance. Oncology, 69(3), 185–201. https://doi.org/10.1159/000088069.
Placke, T., Örgel, M., Schaller, M., Jung, G., Rammensee, H. G., Kopp, H. G., & Salih, H. R. (2012). Platelet-derived MHC class I confers a pseudonormal phenotype to cancer cells that subverts the antitumor reactivity of natural killer immune cells. Cancer research, 72(2), 440–448. https://doi.org/10.1158/0008-5472.CAN-11-1872.
Lavergne, M., Janus-Bell, E., Schaff, M., Gachet, C., & Mangin, P. H. (2017). Platelet integrins in tumor metastasis: do they represent a therapeutic target? Cancers, 9(10), 133. https://doi.org/10.3390/cancers9100133.
Cluxton, C. D., Spillane, C., O'Toole, S. A., Sheils, O., Gardiner, C. M., & O'Leary, J. J. (2019). Suppression of Natural Killer cell NKG2D and CD226 anti-tumour cascades by platelet cloaked cancer cells: implications for the metastatic cascade. PloS one, 14(3), e0211538. https://doi.org/10.1371/journal.pone.0211538.
Weber, M. R., Zuka, M., Lorger, M., Tschan, M., Torbett, B. E., Zijlstra, A., Quigley, J. P., Staflin, K., Eliceiri, B. P., Krueger, J. S., Marchese, P., Ruggeri, Z. M., & Felding, B. H. (2016). Activated tumor cell integrin αvβ3 cooperates with platelets to promote extravasation and metastasis from the blood stream. Thrombosis research, 140 Suppl, 1(Suppl 1), S27–S36. https://doi.org/10.1016/S0049-3848(16)30095-0.
Viallard, C., & Larrivée, B. (2017). Tumor angiogenesis and vascular normalization: alternative therapeutic targets. Angiogenesis, 20(4), 409–426. https://doi.org/10.1007/s10456-017-9562-9.
Jiang, L., Luan, Y., Miao, X., Sun, C., Li, K., Huang, Z., Xu, D., Zhang, M., Kong, F., & Li, N. (2017). Platelet releasate promotes breast cancer growth and angiogenesis via VEGF-integrin cooperative signalling. British journal of cancer, 117(5), 695–703. https://doi.org/10.1038/bjc.2017.214.
Di Vito, C., Navone, S. E., Marfia, G., Abdel Hadi, L., Mancuso, M. E., Pecci, A., Crisà, F. M., Berno, V., Rampini, P., Campanella, R., & Riboni, L. (2017). Platelets from glioblastoma patients promote angiogenesis of tumor endothelial cells and exhibit increased VEGF content and release. Platelets, 28(6), 585–594. https://doi.org/10.1080/09537104.2016.1247208.
Uribesalgo, I., Hoffmann, D., Zhang, Y., Kavirayani, A., Lazovic, J., Berta, J., Novatchkova, M., Pai, T. P., Wimmer, R. A., László, V., Schramek, D., Karim, R., Tortola, L., Deswal, S., Haas, L., Zuber, J., Szűcs, M., Kuba, K., Dome, B., Cao, Y., et al. (2019). Apelin inhibition prevents resistance and metastasis associated with anti-angiogenic therapy. EMBO molecular medicine, 11(8), e9266. https://doi.org/10.15252/emmm.201809266.
Etulain, J., Fondevila, C., Negrotto, S., & Schattner, M. (2013). Platelet-mediated angiogenesis is independent of VEGF and fully inhibited by aspirin. British journal of pharmacology, 170(2), 255–265. https://doi.org/10.1111/bph.12250.
Pietramaggiori, G., Scherer, S. S., Cervi, D., Klement, G., & Orgill, D. P. (2008). Tumors stimulate platelet delivery of angiogenic factors in vivo: an unexpected benefit. The American journal of pathology, 173(6), 1609–1616. https://doi.org/10.2353/ajpath.2008.080474.
Caine, G. J., Lip, G., Zanetto, U., Maheshwari, M., Stonelake, P. S., & Blann, A. D. (2007). A comparison of plasma versus histologic indices of angiogenic markers in breast cancer. Applied immunohistochemistry & molecular morphology : AIMM, 15(4), 382–388. https://doi.org/10.1097/01.pai.0000213137.01536.ca.
Chater, C., Bauters, A., Beugnet, C., M'Ba, L., Rogosnitzky, M., & Zerbib, P. (2018). Intraplatelet vascular endothelial growth factor and platelet-derived growth factor: new biomarkers in carcinoembryonic antigen-negative colorectal cancer? Gastrointestinal tumors, 5(1-2), 32–37. https://doi.org/10.1159/000486894.
Martini, C., Thompson, E. J., Hyslop, S. R., Cockshell, M. P., Dale, B. J., Ebert, L. M., Woods, A. E., Josefsson, E. C., & Bonder, C. S. (2020). Platelets disrupt vasculogenic mimicry by cancer cells. Scientific reports, 10(1), 5869. https://doi.org/10.1038/s41598-020-62648-x.
Lin, H., Xu, L., Yu, S., Hong, W., Huang, M., & Xu, P. (2020). Therapeutics targeting the fibrinolytic system. Experimental & molecular medicine, 52(3), 367–379. https://doi.org/10.1038/s12276-020-0397-x.
Hisada, Y., & Mackman, N. (2017). Cancer-associated pathways and biomarkers of venous thrombosis. Blood, 130(13), 1499–1506. https://doi.org/10.1182/blood-2017-03-743211.
Andrén-Sandberg, A., Lecander, I., Martinsson, G., & Astedt, B. (1992). Peaks in plasma plasminogen activator inhibitor-1 concentration may explain thrombotic events in cases of pancreatic carcinoma. Cancer, 69(12), 2884–2887. https://doi.org/10.1002/1097-0142(19920615)69:12<2884::aid-cncr2820691204>3.0.co;2-s.
Sciacca, F. L., Ciusani, E., Silvani, A., Corsini, E., Frigerio, S., Pogliani, S., Parati, E., Croci, D., Boiardi, A., & Salmaggi, A. (2004). Genetic and plasma markers of venous thromboembolism in patients with high grade glioma. Clinical cancer research : an official journal of the American Association for Cancer Research, 10(4), 1312–1317. https://doi.org/10.1158/1078-0432.ccr-03-0198.
Repetto, O., & De Re, V. (2017). Coagulation and fibrinolysis in gastric cancer. Annals of the New York Academy of Sciences, 1404(1), 27–48. https://doi.org/10.1111/nyas.13454.
Koizume, S., & Miyagi, Y. (2014). Breast cancer phenotypes regulated by tissue factor-factor VII pathway: possible therapeutic targets. World journal of clinical oncology, 5(5), 908–920. https://doi.org/10.5306/wjco.v5.i5.908.
van Es, N., Sturk, A., Middeldorp, S., & Nieuwland, R. (2014). Effects of cancer on platelets. Seminars in oncology, 41(3), 311–318. https://doi.org/10.1053/j.seminoncol.2014.04.015.
Saito, M., Ichikawa, J., Ando, T., Schoenecker, J. G., Ohba, T., Koyama, K., Suzuki-Inoue, K., & Haro, H. (2018). Platelet-derived TGF-β induces tissue factor expression via the Smad3 pathway in osteosarcoma cells. Journal of bone and mineral research : the official journal of the American Society for Bone and Mineral Research, 33(11), 2048–2058. https://doi.org/10.1002/jbmr.3537.
Orellana, R., Kato, S., Erices, R., Bravo, M. L., Gonzalez, P., Oliva, B., Cubillos, S., Valdivia, A., Ibañez, C., Brañes, J., Barriga, M. I., Bravo, E., Alonso, C., Bustamente, E., Castellon, E., Hidalgo, P., Trigo, C., Panes, O., Pereira, J., Mezzano, D., et al. (2015). Platelets enhance tissue factor protein and metastasis initiating cell markers, and act as chemoattractants increasing the migration of ovarian cancer cells. BMC cancer, 15, 290. https://doi.org/10.1186/s12885-015-1304-z.
Mammadova-Bach, E., Zigrino, P., Brucker, C., Bourdon, C., Freund, M., De Arcangelis, A., Abrams, S. I., Orend, G., Gachet, C., & Mangin, P. H. (2016). Platelet integrin α6β1 controls lung metastasis through direct binding to cancer cell-derived ADAM9. JCI insight, 1(14), e88245. https://doi.org/10.1172/jci.insight.88245.
Gil-Bernabé, A. M., Lucotti, S., & Muschel, R. J. (2013). Coagulation and metastasis: what does the experimental literature tell us? British journal of haematology, 162(4), 433–441. https://doi.org/10.1111/bjh.12381.
Coupland, L. A., & Parish, C. R. (2014). Platelets, selectins, and the control of tumor metastasis. Seminars in oncology, 41(3), 422–434. https://doi.org/10.1053/j.seminoncol.2014.04.003.
Sato, H., Usuda, N., Kuroda, M., Hashimoto, S., Maruta, M., & Maeda, K. (2010). Significance of serum concentrations of E-selectin and CA19-9 in the prognosis of colorectal cancer. Japanese journal of clinical oncology, 40(11), 1073–1080. https://doi.org/10.1093/jjco/hyq095.
Honn, K. V., Cicone, B., & Skoff, A. (1981). Prostacyclin: a potent antimetastatic agent. Science, 212(4500), 1270–1272.
Oleksowicz, L., Zuckerman, D., Lomazova, K. D., Buryachkofska, L. E., Astrina, O. C., & Slavina, E. G. (1993). Role of interleukin-2 and mononuclear cells in platelet-tumor interactions. Invasion & metastasis, 13(3), 119–131.
Menter, D. G., Onoda, J. M., Taylor, J. D., & Honn, K. V. (1984). Effects of prostacyclin on tumor cell-induced platelet aggregation. Cancer research, 44(2), 450–456.
Lucotti, S., Cerutti, C., Soyer, M., Gil-Bernabé, A. M., Gomes, A. L., Allen, P. D., Smart, S., Markelc, B., Watson, K., Armstrong, P. C., Mitchell, J. A., Warner, T. D., Ridley, A. J., & Muschel, R. J. (2019). Aspirin blocks formation of metastatic intravascular niches by inhibiting platelet-derived COX-1/thromboxane A2. The Journal of clinical investigation, 129(5), 1845–1862. https://doi.org/10.1172/JCI121985.
Li, H., Lee, M., Liu, K., et al. (2017). Inhibiting breast cancer by targeting the thromboxane A2 pathway. npj Precision Onc, 1, 8. https://doi.org/10.1038/s41698-017-0011-4.
Dumanskiy, Y. V., Stoliarova, O. Y., Syniachenko, O. V., & Iegudina, E. D. (2015). Endothelial dysfunction of vessels at lung cancer. Experimental oncology, 37(4), 277–280.
Ahn, J. H., Lee, K. T., Choi, Y. S., & Choi, J. H. (2018). Iloprost, a prostacyclin analog, inhibits the invasion of ovarian cancer cells by downregulating matrix metallopeptidase-2 (MMP-2) through the IP-dependent pathway. Prostaglandins & other lipid mediators, 134, 47–56. https://doi.org/10.1016/j.prostaglandins.2017.12.002.
Sakai, T., Yamaguchi, N., Shiroko, Y., Sekiguchi, M., Fujii, G., & Nishino, H. (1984). Prostaglandin D2 inhibits the proliferation of human malignant tumor cells. Prostaglandins, 27(1), 17–26. https://doi.org/10.1016/0090-6980(84)90217-x.
Maeda, S., Nakamura, T., & Murata, T. (2014). Tumor suppressor prostaglandin D2. Oncoscience, 1(6), 396–397. https://doi.org/10.18632/oncoscience.53.
Dovizio, M., Alberti, S., Sacco, A., Guillem-Llobat, P., Schiavone, S., Maier, T. J., Steinhilber, D., & Patrignani, P. (2015). Novel insights into the regulation of cyclooxygenase-2 expression by platelet-cancer cell cross-talk. Biochemical Society transactions, 43(4), 707–714. https://doi.org/10.1042/BST20140322.
Fu, S. L., Wu, Y. L., Zhang, Y. P., Qiao, M. M., & Chen, Y. (2004). Anti-cancer effects of COX-2 inhibitors and their correlation with angiogenesis and invasion in gastric cancer. World journal of gastroenterology, 10(13), 1971–1974. https://doi.org/10.3748/wjg.v10.i13.1971.
Xu, X. C. (2002). COX-2 inhibitors in cancer treatment and prevention, a recent development. Anti-cancer drugs, 13(2), 127–137. https://doi.org/10.1097/00001813-200202000-00003.
Gao, C., Wang, H., Wang, T., Luo, C., Wang, Z., Zhang, M., Chen, X., & Tao, L. (2020). Platelet regulates neuroinflammation and restores blood-brain barrier integrity in a mouse model of traumatic brain injury. Journal of neurochemistry, 154(2), 190–204. https://doi.org/10.1111/jnc.14983.
Smeda, M., & Chlopicki, S. (2020). Endothelial barrier integrity in COVID-19-dependent hyperinflammation: does the protective facet of platelet function matter? Cardiovascular research, 116(10), e118–e121. https://doi.org/10.1093/cvr/cvaa190.
Smeda, M., Przyborowski, K., Stojak, M., & Chlopicki, S. (2020). The endothelial barrier and cancer metastasis: Does the protective facet of platelet function matter? Biochemical pharmacology, 176, 113886. https://doi.org/10.1016/j.bcp.2020.113886.
Acknowledgments
The authors would like to thank the Sao Paulo Research Foundation, Coordination for the Improvement of Higher Education Personnel (CAPES) and National Council for Scientific and Technological Development (CNPq).
Funding
The studies related to platelet biology were supported by the Sao Paulo Research Foundation under grants AVSF (2017/08119-8 and 2018/00736-0), SSA (2016/14459-3 and 2017/26317-1), and CVFH (2015/20412-7); National Council for Scientific and Technological Development (CNPq) - Brazil under grant: 303900/2017-2 (CVFH) and the Coordenação de Aperfeiçoamento de Pessoal de Nível Superior - Brasil (CAPES) - Finance Code 001.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflicts of interest.
Additional information
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Faria, A.V.S., Andrade, S.S., Peppelenbosch, M.P. et al. Platelets in aging and cancer—“double-edged sword”. Cancer Metastasis Rev 39, 1205–1221 (2020). https://doi.org/10.1007/s10555-020-09926-2
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10555-020-09926-2