
Abstract
Doubled haploids (DH) have become a standard tool in breeding and genetic studies of many crops and in most cases androgenesis is the only available route of their production. It has been recently observed that some populations of DH lines obtained via androgenesis contain high proportions of clones. This seriously reduces the efficiency of breeding and may jeopardize genetic studies. This study was designed to determine at which stage of androgenesis these clones are created, using samples set aside during routine production of DH lines in breeding of hexaploid triticale. The fate of each androgenic structure was carefully followed through the entire regeneration process, and all obtained plants were genotyped using DNA markers. Overall, 189 plants were regenerated forming 33 families, each originating from a single original androgenic structure (callus, polyembryos). In ca. 80 % of cases all members of a family were genetically identical. However, in about 20 % of cases the families of regenerants were genetically heterogeneous, showing that not all androgenic structures originate from single microspores. The evidence shown here demonstrates that retention of single plants from each original structure guarantees the production of only unique genotypes but it reduces the total output of plants. If maximum output is desired, multiple regenerants from single callus can be retained but must be genotyped using at least 10 polymorphic markers to identify clones.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Production of doubled haploids (DH) for crop breeding and genetics has become routine in many species (Maluszynski et al. 2003; Forster et al. 2007). Recent review articles (Croser et al. 2006; Ferrie 2007; Dunwell 2010) list dozens of species in which DH have been produced and dozens more in which the technology has been tried. The list of species keeps growing rapidly as the technology improves. DH lines are developed from cells of the male or female gametophytic pathways, and later have their chromosome numbers doubled. Doubling can be either by natural (spontaneous) or artificial means, creating plants that are perfectly homozygous at every locus, with the possible exception of post-doubling mutation. As such, populations of DH lines are invaluable both to breeders and geneticists. For breeders, they provide genetically stable material that can be quickly evaluated in replicated trials, eliminating the long process of reaching homozygosity by self-pollination or sib mating. For geneticists, they offer a unique set of materials for detailed studies of many plant characteristics, especially quantitative traits controlled by genes with small additive effects.
In any approach to the DH production, the efficiency of the process is of paramount importance. The term “efficiency” is understood here as the number of unique, useful genotypes produced from the available source, suitable for the purpose for which they are developed, obtained with a specific investment, whether financial, labor, or both. Because each DH line is expected to be derived from a single cell of the gametophytic pathway, each line is expected to be unique. Therefore, implicit is the assumption that the number of recovered plants directly indicates the number of unique genotypes available for testing. This number in turn determines the experiment’s resolution, or the probability of success in finding the desired allele combination in a breeding effort. Anything that distorts the 1:1 relationship between the number of lines and the number of unique genotypes reduces the efficiency of the process. If chromosome elimination is used to generate DH, such as wheat × maize (Laurie and Bennett 1986) or barley × Hordeum bulbosum (Kasha and Kao 1970) crosses, chances for a distortion of the 1:1 ratio appear remote. Not so in androgenesis, where microspores are induced to switch from the gametophytic to sporophytic development pathway. Chances for the generation of clones appear high; how high they really are is not entirely clear as the problem is only sporadically mentioned in the literature. Perhaps it is marginal and confined to only some laboratories, or perhaps it is more widespread. From anecdotal evidence and personal communication we have reasons to suspect the latter. In some experiments large proportions of DH turned out to be clones, reducing the effective population sizes to below acceptable resolution levels. Such incidents are not widely publicized, which only makes the situation more serious as proportions of clones in populations of DH may in fact be much larger than published data suggest. Tenhola-Roininen et al. (2006) had to reject 32 % of a DH mapping population in rye because of their clonal nature. Genotyping with DArT (Diversity Arrays Technology) markers among several populations of DH lines of hexaploid triticale created for QTL (Quantitative Trait Loci) mapping revealed up to 60 % of clones in one population, with the largest clone numbering 11 presumed DH lines (Oleszczuk et al. unpublished). Such a high proportion of duplication reduces the number of useful lines thereby limiting the resolution of the mapping effort and seriously increases the cost of the operation. In breeding populations, high proportions of clones limit chances of finding useful recombinants because the effective population size is much smaller than sheer numbers imply. In a way of consolation, clones can be viewed as a measure of experiment replication already during the first field seed increase. However, the identity of clones may never be known because in breeding, extensive (and expensive) genotyping is rarely performed. If genotyping is done, any benefit from unexpected replication is more than offset by reduced resolution of the experiment. In this sense, the clones never assist or speed up the process of line evaluation; they only use up valuable resources.
Very high frequencies of clones in some mapping populations generated via androgenesis were intriguing and deserving closer examination. Although the formation of clones is at times discussed in the literature, especially in the context of micropropagation, this phenomenon has not been investigated in detail for androgenesis of a crop species. This article attempts to shed light on their origin in the process of androgenic line development using standard breeding materials of hexaploid triticale. Triticale (× Triticosecale Wittmack) is a man-made crop created by hybridization between wheat (Triticum sp.) and rye (Secale cereale L.) developed to combine grain quality, productivity, and disease resistance of wheat with rye’s excellent environmental adaptability and tolerance. It is amenable to androgenesis (Immonen and Robinson 2000; Oleszczuk et al. 2004; Ponitka and Slusarkiewicz-Jarzina 2007; Würschum et al. 2012) and registered cultivars have been created via the doubled haploid approach (Dr. Z. Banaszak—Plant Breeding Danko Ltd Poland, Dr. H. Wos—Plant Breeding Strzelce Ltd Poland, pers. comm.).
In this study, we handled experimental material in the process of androgenesis on solid and liquid media in standard ways, yet carefully tracking the history of every selected androgenic structure, so that each regenerated plant had its complete pedigree, to a specific head from which it was derived. We then verified the genetic status of each regenerant with locus-specific co-dominant markers (i.e. microsatellites) randomly selected to represent loci from the A, B and R genomes present.
Materials and methods
Androgenesis and plant regeneration
Androgenic structures used in the experiments here were diverted from a routine production of DH lines for triticale breeding. Twelve F1 hybrids of winter hexaploid triticale were used (Table 1); all were from standard breeding crosses kindly provided by Drs. M. Pojmaj from Plant Breeding Danko Ltd Poland, and H. Wos and. G. Budzianowski from Plant Breeding Strzelce Ltd Poland.
Androgenesis protocols were the same as routinely used for microspore culture (Oleszczuk et al. 2004) and anther culture (Warzecha et al. 2005) of triticale. Tillers were cold-treated in water for 4 weeks. For the solid culture approach, modified 190-2 induction medium (Zhuang and Xu 1983) supplemented with 90 g/l maltose, 438 mg/l glutamine, 2 mg/l 2.4-D and 0.5 mg/l kinetin was used; liquid medium was the same but without growth regulators.
Individual androgenic structures were manually selected under a dissecting microscope at their early stages of development and at the appropriate stage, transferred onto the solidified 190-2 regeneration medium (Zhuang and Xu 1983) supplemented with growth regulators according to Pauk et al. (1991) and sub-cultured every 2 weeks. The cultures were kept under a 16/8-h (day/night) photoperiod. Plants with well-developed roots and shoots from glass tubes were potted, transferred into a cold chamber for acclimatization, vernalization and subsequently to a greenhouse.
The number of originally selected early androgenic structures was not noted. Given the high rate of albinism in triticale androgenesis, this number was immaterial. However, 33 independent original structures eventually produced viable green progeny. “Independent original structures” are understood here as the first identifiable structures, whether appearing as embryos or calli, emerging from bursting anthers or formed by the microspores in suspension. All derivatives of these original structures with their pedigrees noted down at every step of the procedure were maintained independently through sub-culturing, whether those were calli or polyembryos or plantlets. All plants derived from a single original androgenic structure (callus, polyembryo) were treated as families. In this way, every green plant regenerated during the experiment could be traced back through its pedigree to an individual original structure diverted to this experiment. Genotyping with DNA markers was done on plants from original, separately cultured androgenic structures producing at least three regenerants each. Evident twins were not analyzed as their clonal nature was obvious.
Genotyping
Total genomic DNA was extracted from fresh young leaves of regenerated plants using the Nucleo®Spin Food kit according to the Macherey–Nagel GmbH & Co. KG protocol. DNA was suspended in 50 μl of PE buffer (5 mM Tris–HCl, pH 8.5) and further diluted to 20 ng/μl. Microsatellite markers were selected from published lists with known chromosome locations (Table 2). Genotyping was done in two stages. Originally, 10 SSR (Simple Sequence Repeat) markers targeting random chromosome locations were tested on 33 families of regenerants. Samples suspected of heterogeneity were further genotyped with additional markers. Information on the primer sequences of the survey of microsatellite markers (REMS) and S. cereale microsatellite markers (SCM) was kindly provided by Dr. V. Korzun from Lochow–Petkus GmbH (Germany). The wms wheat marker series was developed by Röder et al. (1998); wmc and barc markers were developed within the Wheat Microsatellite Consortium and are available from the GrainGenes Database (wheat.pw.usda.gov). DNA amplification and polymorphism identification were performed as previously (Tyrka et al. 2008).
Genetic distances were calculated according to Nei (1972) and the unweighted pair-group method using the arithmetic means (UPGMA) algorithm was used for grouping. The bootstrap procedure was applied to test the reliability of clustering using 100 random samples of molecular markers. PHYLIP 3.69 Software (Felsenstein 1989) was applied for bootstrapping, genetic distance calculations, UPGMA grouping, and construction of consensus tree. The dendrogram was visualized with TreeView (Page 1996).
Results
In the course of this study, microspore-derived plants of 12 F1 hybrids were regenerated from individual, selected androgenic structures such as callus, polyembryo, or twin-embryo (Fig. 1). Overall, 189 regenerants were obtained from 33 individual androgenic structures forming 33 families, grouped based on their origin. A family (bulk) is understood as a group of plants originating from a single androgenic structure identified at some point in standard production of triticale androgenic regenerants. Of these 189 plants, 20 families totaling 102 plants originated from polyembryos and 13 families totaling 87 plants originated from calli (Table 1). The numbers of green plants eventually derived from an original structure ranged from one to as many as ten from a single callus (B22) and eight plants from a polyembryo (B21) (Table 1). These numbers are by no means indicative of the upper limit of culture capability; the purpose of the experiment was to identify the stage at which clones are formed and maximum sizes of clones were of no interest.

Multiembryo formation and the production of clones in androgenic triticale regenerants: embryos developed on microspore-derived callus on solid medium (a); twin embryo development—two connate scutella each with an embryo axis (b); initiating a cycle of secondary embryo proliferation (c); direct secondary embryos formation on a primary embryo in liquid medium (d); multicellular microspore-derived connate structures producing heterogenous microcallus (e); secondary embryos developing on the suspensor of a primary embryo in liquid medium multicellular (f); mature androgenic twin embryos (g) and germination of plantlet-clones from such twin embryos (h); polyembryos (i) and plantlets-clones derived after embryos conversion (j)
The SSR markers used in this study were selected from 252 combinations of primers previously tested on a set of wheat, rye and triticale genotypes (Tyrka et al. unpublished). All were polymorphic across the genotypes tested. Mean numbers of polymorphic markers among pairs of parents ranged from nine in DD 436/07 × MAH 31938-5 to two in DD 298/06 × DD 466/07 and CD 13469/02 × DD 436/07, with the average number of 4.2 polymorphic loci per cross combination (Table 1). This offers sufficient resolution to reveal heterogeneity within groups of plants sharing common ancestry in culture.
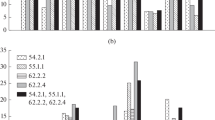
Of 33 families of regenerants tested 27 (81.8 %) were homogenous for the sets of alleles they carry (Table 1). Given the average number of polymorphic markers in each hybrid, the level of confidence is 93.75 % that those were clones. The remaining six families were heterogeneous: four of these were derived via callus (B2, B5, B20, and B25), and two via polyembryos (B6, B17). Among the callus derived families, B2 consisted of five plants and was heterogeneous at three loci—wms275-2A, wmc24-1A and barc324-3A. Among these five plants there were three different homozygous genotypes. Family B5, also regenerated from callus, was also heterogeneous at three loci: (wmc24-1A, barc324-3A and rems1194-2R; Fig. 2) and it consisted of two distinct genotypes, one with four and the other with one plant. Another callus-derived family B20 from was heterogeneous at loci wmc537-5B, wmc24-1A and rems1194-2R and also consisted of two homozygous genotypes: one clone of four and a single genetically distinct plant. However, in one plant two alleles were present at the wmc24 locus. This might have been a mutation or a case of disomy for the chromosome carrying this marker. Finally, B25 consisted of two genetically different clones represented by two and six plants each, and it included a single heterogeneous locus. The experience with the materials tested here suggests that with conservative approach of retaining only single plants from each original callus structure would have reduced the total output of regenerants by ca. 30 % but each line would be unique. If more than a single plant is to be retained from each callus, at least 10 polymorphic SSR markers should be used to detect possible additional unique genotypes.

Polymorphism of selected SSR markers reveals heterogeneity among the 33 families of regenerants. Lane numbers (multichannel loading) correspond to number of bulk (Table 1). Examples of amplification from two loci are in lanes 2 and 5 for marker barc324-3A (a), lanes 2, 5, 20 and 25 of marker wmc24-1A (b), and lanes 13 and 17 of polymorphism revealed with marker scm126-1R (c)
Among the two heterogeneous bulks of polyembryo regenerants, in B6 a single plant was heterozygous at wms537-5B with different signal intensity of the two alleles. This may be an instance of chimaerism, non-specific fragment amplification or perhaps aneuploidy (disomy) for a specific chromosome. B17 consisted of three plants with five polymorphic loci. Two markers (scm126-1R, and rems1259-6R) were used to test individual plants. Marker scm126 indicated the presence of two clones, however, the alleles were atypical and it is not possible at this level of analysis to postulate the exact nature of heterozygosity. It could be duplication or other mutation in primer binding sites. All three plants in the bulk were homogenous for rems1259. This shows that in case of regeneration via polyembryos, retention of a single plant from each polyembryo gives full representation of distinct genotypes present. In contrast to the callus derived DH lines, screening with molecular markers does not reveal any additional unique DH genotypes because all multiple lines regenerated from each original polyembryos are clones.
Discussion
In many crops, androgenesis is currently the only viable option for DH production. Even in species that are yet to be tested, androgenesis seems to offer a better hope for success than potentially open-ended search of a suitable pollinator whose chromosome may be susceptible to elimination in early zygote/embryo divisions. The list of species in which androgenesis is already actively used or is at the stage of testing, is long and growing (Maluszynski et al. 2003; Dunwell 2010). The main tenet of androgenesis is that each derived line is unique because each one originates from a single microspore. For this reason, the number of individual plants should correspond to the number of individual lines. However, clones have been identified among androgenic regenerants on several occasions (Zwierzykowski et al. 1999; Tenhola-Roininen et al. 2006; Czembor et al. 2007; Głowacka et al. 2012) and at times their presence may jeopardize the goals, such as when a population used for phenotyping is eventually found out to contain too few unique genotypes for an acceptable resolution level.
The presence of clones among androgenic regenerants can only be demonstrated with the aid of markers, and larger numbers of markers provide higher confidence in analyses. The use of DNA markers for the identification of clones has been reported for many species (Rivard et al. 1994; Veilleux et al. 1995; Zwierzykowski et al. 1999; Kopecky et al. 2005). Here, SSR analysis was used for its simplicity, low cost and ability to identify multiple alleles at given loci. It has been used for large scale analyses to identify homozygous regenerants in maize (Belicuas et al. 2007), potato (Chani et al. 2000), cucumber (Diao et al. 2009) and tomato (Corral-Martínez et al. 2011).
The primary motivation for this study was an observation of clones among DH lines of triticale genotyped by as many as 1500 polymorphic DArT markers per combination (Oleszczuk et al. unpublished). With the presence of clones among triticale DH lines generated with standard protocols already a known fact, the focus was on their origin and lower resolution levels in clone identification appeared justified. A simple marker system is used to genotype a relatively small sample of regenerants. Still, the approach clearly shows a high frequency of clones among regenerated plants, and the SSR markers fail to resolve only some minor issues.
This study focused on the origin of clones generated during production of haploid and doubled haploid material in triticale. The androgenic structures used and the 189 regenerated plants were a portion of a routine production run of DH triticale lines for breeding, using standard protocols. The regeneration rates were typical, with the 12 F1 hybrids yielding a total of 5,580 green plants passed on to the breeders, the 189 plants held for this study, as well as over 6,700 albino plants.
Of the 33 families analyzed, 27 contained genetically identical plants. This means that in this study, ca. 82 % of families (plants derived from one common androgenic structure along the regeneration process) were clones. This frequency must not be taken as indicative of the true nature of DH populations in triticale: this study was focused on clones. However, populations of triticale DH lines consisting in one half of clones have already been identified among mapping populations derived by various research groups (Oleszczuk et al. unpublished), so the problem is not trivial. The remaining six families of the 33 analyzed here were composed of genetically distinct plants. In some cases, the evidence for heterogeneity might have been generated by mutation as a consequence of somaclonal variation arising from in vitro culture procedures (Machczyńska et al. 2014), or, perhaps, by participation of unreduced gametes. In other cases, the families were clearly of multiple origins. This also means that with some frequency (here ca. 18 %), common derivation of a plant family does not guarantee its homogeneity. In other words, some embryogenic structures do in fact originate from more than a single haploid microspore. They are unlikely to have originated from unreduced gametes. Depending on their origin such unreduced gametes could produce progeny with two parental alleles at some given locus, but could not produce lineages of genetically different plants homozygous for single alleles per locus. The resolution level of genetic analysis employed here was insufficient to discriminate among all groups of androgenic cultures. The mean significance level of the dendrogram based on 100 bootstraps was 22.4 %; values above 40 % are included in the tree (Fig. 3). There were two groups of bulks consisting of genetically indistinguishable individuals. Bulks B22 (Mikado × DD 466/07), B8 and B9 (DD 298/06 × DD 466/07) shared a common father and based on the DNA markers used were genetically identical. Similarly, there were no differences within groups B10 and B12 derived from a polyembryo and callus of the DD 436/07 × MAH 31938-5 hybrid. B8 and B9 originated from the same head hence, they were regenerated on the same plate. However, B22 was from a different cross, (but with the same male partner), and the two most certainly had never been grown together. Excluding a mix-up in material handling downstream from anther plating, this case may illustrate the resolution limit of the approach with the given set of DNA markers. A similar situation was encountered by Kato et al. (2012) where 17 SSR markers were insufficient to discriminate all clones of flowering cherry.

Dendrogram representing the genetic distance (D) generated by 10 SSR markers. Values from bootstrap analysis are provided at nodes when higher than 40
While it is assumed that some clones can be created by inadvertent splitting of plants during handling, it is difficult to imagine that up to eleven copies of the same genotype can be created in this fashion. The root of the problem is probably located in the process/manner of plant regeneration itself. Embryo production in triticale can be indirect (via callus) or direct, without the callus stage, both in anther culture (on solid media) and in microspore culture (in liquid media) (Fig. 1). Formation of callus always introduces ambiguity to the process, and the nature of regenerants derived from this tissue can never be certain. Overgrowth of callus is often associated with production of large number of embryos, and as long as this callus originates from a single microspore, all regenerants should be genetically identical, excepting possible mutations and consequent chimaerism. This may easily lead to formation of clones, especially for callus with high regenerative potential. Multiple plants originating from callus derived from a single microspore have been noted (Birhman et al. 1994; Rivard et al. 1994). On the other hand, if plants derived from the same callus tissue are genetically different, they must have originated from different haploid microspores or from non-haploid microspores. This may occur because at the early stages of development, multicellular structures formed by adjacent microspores may fuse/intertwine, and form genetically heterogeneous callus (Fig. 1e). Such fusions are perhaps less likely to take place in microspore cultures then in anther culture, where early stages of development take place in a closed anther and are inaccessible to observation. Generally, on solid media, it can be difficult to distinguish the origin of individual regenerants from various calli growing from a single anther; at times it may even be impossible to ascertain the type of embryogenesis taking place. Some authors suggested that absence of the callus stage will eliminate duplication (cloning) among androgenic regenerants (Bentolila et al. 1992; Rivard et al. 1994). However, our results show that there are additional pathways of clone production via androgenesis.
Direct secondary embryogenesis by the polyembryo formation was always suspect in clone regeneration. In this case, the formation of the first (primary) embryo is followed by the formation of additional, secondary embryos (Fig. 1d). This secondary embryogenesis occurs directly at the suspensor of the embryo (Fig. 1f), at times with high efficiency, particularly in liquid media (data not shown). The formation of embryos joined together by a suspensor was previously reported in culture of isolated microspores of triticale cv. Bogo (Oleszczuk et al. 2004). New embryos may be generated from the primary embryo in a cyclic manner (Fig. 1c). However, well developed embryos that are not transferred onto regeneration media at the right time may also de-differentiate into calli on which secondary embryogenesis can proliferate. The scale and extent of clone regeneration depends on the cycle duration and on the number of embryos obtained. Hua et al. (2010) reported three successive cycles of secondary embryogenesis in anther culture of rubber tree. In a wide range of plant species, the efficiency of secondary embryogenesis is much higher than of primary embryogenesis, and an unlimited number of secondary embryos can be generated (Raemakers et al. 1995). In this study, twin (connate) embryo formation, with a somewhat different mechanism of development than polyembryo, was a source of clones (Fig. 1b, g, h). Twin embryos were also observed during androgenesis of wheat and maize (Rybczynski et al. 1991; Wan and Widholm 1992). Finally, secondary embryos can be produced either from calli or from primary embryos, making the identification and tracking of individual embryos difficult, especially under standard production regimes. So, on one hand, the ability to induce secondary embryogenesis among androgenic structures increases the efficiency of plant regeneration, but on the other hand, it is the cause of clones which can dramatically impact the usefulness of the derived material.
Individual microspores may also give rise to chimaeric callus, from which plants with distinct molecular profiles can be produced. Callus may originate from microspore-derived haploid cells, or diploid cells formed as a consequence of nuclear fusion, endoreduplication, endomitosis or restitution (Kasha et al. 2001; Seguí-Simarro and Nuez 2008; Zhang et al. 2011). In this study, 13 families of regenerants were callus-derived and in most cases, each family represented a clone. However, there were also families composed of two different genotypes, indicating that what in early stages of the procedure appeared as single structures, in fact originated from more than a single haploid microspore. Whether this was by fusion or overgrowth of different calli could not be determined. Origination of such families from non-reduced microspores would not produce groups of identical genotypes hemizygous or homozygous for the markers used in genotyping but rather, genetically unique plants heterozygous for two alleles per locus.
When haploids are regenerated in androgenesis process, their microspore origin is essentially certain. There is more ambiguity about any obtained diploids. It is known that in androgenesis of triticale, spontaneous doubling of the chromosome number is relatively common. However, diploids may also originate from somatic tissues of the anther (Corral-Martínez et al. 2011; Smykalova et al. 2012). Genotyping performed here clearly shows that all regenerants were derived from microspores and were homozygous for specific alleles (the source plants were F1 hybrids, heterozygous at many tested loci). This is in line with previous observations on the absence of somatic tissue regenerants in triticale but in contrast to the demonstrated sporophytic origin of some regenerants in other cereals such as rice (Grewal et al. 2011). The few examples of heterozygosity observed here are unconvincing. DNA heterogeneity in sets of callus-mediated regenerants was explained by the presence of several homogenous clones while heterogeneity of the polyembryo regenerants was likely caused by mutations.
Conclusions
The recommendation from this study is that the material in androgenesis must be handled with attention to its final use/destination. If it is to be genetic studies or breeding, the formation of clones must be avoided at all cost, even if it appears to reduce the efficiency of the process. All chances for cyclical embryo formation must be avoided, even though this study shows that not all calli are formed from single microspores and hence, not all cyclical embryos are clones. It would appear that purposeful reduction in the total output (expressed as the number of unique genotypes recovered) is worth the expense of a high proportion of clones. On the other hand, if the purpose is material multiplication, the factors enhancing clone formation would be of considerable benefit. The evidence shown here demonstrates that retention of single plants from each original structure guarantees the production of only unique genotypes but at the same time reduces the total output. If maximum output is desired, multiple regenerants derived via callus may be retained but must be genotyped by at least 10 polymorphic markers to eliminate clones.
References
Belicuas PR, Guimarães CT, Paiva LV, Duarte JM, Maluf WR, Paiva E (2007) Androgenetic haploids and SSR markers as tools for the development of tropical maize hybrids. Euphytica 156:95–102
Bentolila S, Hardy T, Guitton C, Freyssinet G (1992) Comparative genetic analyses of F2 plants and anther culture derived plants of maize. Genome 35:575–582
Birhman RK, Rivard SR, Cappadocia M (1994) Restriction fragment length polymorphism analysis of anther culture-derived Solanum chacoense. HortScience 29:206–208
Chani E, Veilleux RE, Boluarte-Medina T (2000) Improved androgenesis of interspecific potato and efficiency of SSR markers to identify homozygous regenerants. Plant Cell Tiss Organ Cult 60:101–112
Corral-Martínez P, Nuez F, Seguí-Simarro J (2011) Genetic, quantitative and microscopic evidence for fusion of haploid nuclei and growth of somatic calli in cultured ms10 35 tomato anthers. Euphytica 178:215–228
Croser JS, Lulsdorf MM, Davies PA, Clarke HJ, Bayliss KL, Mallikarjuna N, Siddique KHM (2006) Toward doubled haploid production in the Fabaceae: progress, constraints, and opportunities. Crit Rev Plant Sci 25:139–157
Czembor P, Cz Radecka M, Arseniuk E (2007) A molecular map of wheat (Triticum aestivum L.). Biul IHAR 243:279–288
Diao WP, Jia YY, Song H, Zhang XQ, Lou QF, Chen JF (2009) Efficient embryo induction in cucumber ovary culture and homozygous identification of the regenerants using SSR markers. Sci Hortic 119:246–251
Dunwell JM (2010) Haploids in flowering plants: origins and exploitation. Plant Biotech J 8:1–48
Felsenstein J (1989) PHYLIP—phylogeny inference package (version 3.2). Cladistics 5:164–166
Ferrie AMR (2007) Doubled haploid production in nutraceutical species: a review. Euphytica 158:347–357
Forster BP, Heberle-Bors E, Kasha KJ, Touraev A (2007) The resurgence of haploids in higher plants. Trends Plant Sci 12:368–375
Głowacka K, Kaczmarek Z, Jeżowski S (2012) Androgenesis in the bioenergy plant Miscanthus sinensis: from calli induction to plant regeneration. Crop Sci 52:2659–2673
Grewal D, Manito Ch, Bartolome V (2011) Doubled haploids generated through anther culture from crosses of elite Indica and Japonica cultivars and/or lines of rice: large scale production, agronomic performance and molecular characterization. Crop Sci 51:2544–2553
Hua YW, Huang TD, Huang HS (2010) Micropropagation of self-rooting juvenile clones by secondary somatic embryogenesis in Hevea brasiliensis. Plant Breed 129:202–207
Immonen S, Robinson J (2000) Stress treatments and ficoll for improving green plant regeneration in triticale anther culture. Plant Sci 150:77–84
Kasha KJ, Kao KN (1970) High frequency haploid production in barley (Hordeum vulgare L.). Nature 225:874–876
Kasha KJ, Hu TC, Oro R, Simion E, Shim YS (2001) Nuclear fusion leads to chromosome doubling during mannitol pretreatment of barley (Hordeum vulgare L.) microspores. J Exp Bot 52:1227–1238
Kato S, Matsumoto A, Yoshimura K, Katsuki T, Iwamoto K, Tsuda Y, Ishio S, Nakamura K, Moriwaki K, Shiroishi T, Gojobori T, Yoshimaru H (2012) Clone identification in Japanese flowering cherry (Prunus subgenus Cerasus) cultivars using nuclear SSR markers. Breed Sci 62:248–255
Kopecky D, Lukaszewski AJ, Gibeault V (2005) Reduction of ploidy level by androgenesis in intergeneric Lolium–Festuca hybrids for turf grass breeding. Crop Sci 45:274–281
Laurie DA, Bennett MD (1986) Wheat × maize hybridization. Can J Genet Cytol 28:313–316
Machczyńska J, Orłowska R, Zimny J, Bednarek PT (2014) Extended metAFLP approach in studies of the tissue culture induced variation (TCIV) in case of triticale. Mol Breed. doi:10.1007/s11032-014-0079-2
Maluszynski M, Kasha KJ, Forster BP, Szarejko I (2003) Doubled haploid production in crop plants: a manual. Kluwer, Dordrecht
Nagy S, Poczai P, Cernák I, Gorji AM, Hegedűs G, Taller J (2012) PICcalc: an online program to calculate polymorphic information content for molecular genetic studies. Biochem Genet 50:670–672
Nei M (1972) Genetic distance between populations. Am Nat 106:283–292
Oleszczuk S, Sowa S, Zimny J (2004) Direct embryogenesis and green plants regeneration from isolated microspores of hexaploid triticale (× Triticosecale Wittmack) cv. Bogo. Plant Cell Rep 22:885–893
Oleszczuk S, Rabiza-Swider J, Zimny J, Lukaszewski AJ (2011) Aneuploidy among androgenic progeny of hexaploid triticale (× Triticosecale Wittmack). Plant Cell Rep 30:575–586
Page RDM (1996) Tree view: an application to display phylogenetic trees on personal computers. Comput Appl Biosci 12:357–358
Pauk J, Manninen O, Mattila I, Salo Y, Puli S (1991) Androgenesis in hexaploid spring wheat F2 population and their parents using a multiple-step regeneration system. Plant Breed 107:18–27
Ponitka A, Slusarkiewicz-Jarzina A (2007) The effect of liquid and solid medium on production of winter triticale (× Triticosecale Wittm.) anther-derived embryos and plants. Cereal Res Commun 35:15–22
Raemakers CJJM, Jacobsen E, Visser RGF (1995) Secondary somatic embryogenesis and applications in plant-breeding. Euphytica 81:93–107
Rivard SR, Saba-El-Leil MK, Landry BS, Cappadocia M (1994) RFLP analyses and segregation of molecular markers in plants produced by in vitro anther culture, selfing, and reciprocal crosses of two lines of self-incompatible Solanum chacoense. Genome 37:775–783
Röder MS, Korzun V, Wendehake K, Plaschke J, Leroy P, Ganal MW (1998) A microsatellite map of wheat. Genetics 149:2007–2023
Rybczynski JJ, Simonson RL, Baenziger PS (1991) Evidence for microspore embryogenesis in wheat anther culture. In Vitro Cell Dev Biol 27P:168–174
Seguí-Simarro JM, Nuez F (2008) Pathways to doubled haploidy: chromosome doubling during androgenesis. Cytogenet Genome Res 120:358–369
Smykalova I, Horacek J, Kubosiova M, Smirous P Jr, Soukup A, Gasmanova N, Griga M (2012) Induction conditions for somatic and microspore-derived structures and detection of haploid status by isozyme analysis in anther culture of caraway (Carum carvi L.). In Vitro Cell Dev Biol 48:30–39
Tenhola-Roininen T, Immonen S, Tanhuanpaa P (2006) Rye doubled haploids as a research and breeding tool—a practical point of view. Plant Breed 125:584–590
Tyrka M, Perovic D, Wardynska A, Ordon F (2008) A new diagnostic SSR marker for selection of the Rym4/Rym5 locus in barley breeding. J Appl Genet 49:127–134
Veilleux RE, Shen LY, Paz MM (1995) Analysis of the genetic composition of anther-derived potato by randomly amplified polymorphic DNA and simple sequence repeats. Genome 38:1153–1162
Wan Y, Widholm JM (1992) Formation of multiple embryo-like structures from single microspores during maize anther culture. Plant Cell Rep 11:529–531
Warzecha R, Sowa S, Salak-Warzecha K, Oleszczuk S, Sliwinska E, Zimny J (2005) Doubled haploids in production of male sterility maintaining triticale (Triticosecale Wittmack) lines. Acta Physiol Plant 27:245–250
Würschum T, Tucker MR, Reif JC, Maurer HP (2012) Improved efficiency of doubled haploid generation in hexaploid triticale by in vitro chromosome doubling. BMC Plant Biol 12:109
Zhang L, La Zhang, Luo J, Chen W, Hao M, Liu B, Yan Z, Zhang B, Zhang H, Zheng Y, Liu D, Yen Y (2011) Synthesizing double haploid hexaploid wheat populations based on a spontaneous alloploidization process. J Genet Genomics 38:89–94
Zhuang JJ, Xu J (1983) Increasing differentiation frequencies in wheat pollen callus. In: Hu H, Vega MR (eds) Cell and tissue culture techniques for cereal crop improvement. Science Press, Beijing, p 431
Zwierzykowski Z, Zwierzykowska E, Slusarkiewicz-Jarzina A, Ponitka A (1999) Regeneration of anther-derived plants from pentaploid hybrids of Festuca arundinacea × Lolium multiflorum. Euphytica 105:191–195
Acknowledgments
The authors thank Prof. A. Lukaszewski for valuable suggestions and critical reading of the manuscript. We acknowledge the technical assistance from M. Dobrzyńska, A. Zimny, K. Makowska and A. Drąg.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
About this article
Cite this article
Oleszczuk, S., Tyrka, M. & Zimny, J. The origin of clones among androgenic regenerants of hexaploid triticale. Euphytica 198, 325–336 (2014). https://doi.org/10.1007/s10681-014-1109-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10681-014-1109-1