
Abstract
In this study, the seedlings of two wheat cultivars were used: drought-resistant Chinese Spring (CS) and drought-susceptible (SQ1). Seedlings were subjected to osmotic stress in order to assess the differences in response to drought stress between resistant and susceptible genotype. The aim of the experiment was to evaluate the changes in physiological and biochemical characteristics and to establish the optimum osmotic stress level in which differences in drought resistance between the genotypes could be revealed. Plants were subjected to osmotic stress by supplementing the root medium with three concentrations of PEG 6000. Seedlings were grown for 21 days in control conditions and then the plants were subjected to osmotic stress for 7 days by supplementing the root medium with three concentrations of PEG 6000 (D1, D2, D3) applied in two steps: during the first 3 days of treatment −0.50, −0.75 and −1.00 and next −0.75, −1.25 and −1.5 MPa, respectively. Measurements of gas exchange parameters, chlorophyll content, height of seedlings, length of root, leaf and root water content, leaf osmotic potential, lipid peroxidation, and contents of soluble carbohydrates and proline were taken. The results highlighted statistically significant differences in most traits for treatment D2 and emphasized that these conditions were optimum for expressing differences in the responses to osmotic stress between SQ1 and CS wheat genotypes. The level of osmotic stress defined in this study as most suitable for differentiating drought resistance of wheat genotypes will be used in further research for genetic characterization of this trait in wheat through QTL analysis of mapping population of doubled haploid lines derived from CS and SQ1.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
It is known that water is an important environmental factor and a major limitation for plant growth, development and yield. Plants usually experience a fluctuating water supply during their life cycle due to continuously changing climatic factors (Blum 1998; Bohnert et al. 1995; Chaves et al. 2002; Passioura et al. 1993; Tan et al. 2006). Variability of resistance to drought within plants belonging to the same species has not been completely explained. The responses of plants to water stress depend on plant species, plant age, phase of growth and development, level and duration of drought and physical parameters. Differences in resistance to drought stress are known to exist amongst genotypes of plant species, e.g. in maize (Martiniello and Lorenzoni 1985; Lorens et al. 1987), wheat (Winter et al. 1988), and triticale (Grzesiak et al. 2003). Plants develop different mechanisms (morphological, physiological and biochemical) which inhibit or remove the harmful effects of stresses.
Water deficit caused by drought and osmotic stress effects changes in morphology, water status, gas exchange and chlorophyll content which are connected with the onset of protective mechanisms in the plant (Blum and Ebercon 1981; Mansfield and Davies 1981; Jackson et al. 1996). Application of polyethylene glycol (PEG) in a hydroponic solution causes osmotic stress, which results in changes in the water status of the tissues and a decrease of plant growth and biomass production (Berkowitz et al. 1983; Kicheva et al. 1994; P’erez-Alfocea and Larcher 1995; Grzesiak et al. 2003). Usually drought-resistant genotypes accumulate more biomass in leaves than susceptible ones (Kerepesi and Galiba 2000). Leaf water content and gas exchange are physiological processes very susceptible to drought stress. Under moderate drought, a decrease in photosynthesis is generally considered to be the result of reduced availability of CO2 due to stomatal closure (Mansfield and Davies 1981). However, when drought is prolonged, a decrease in photosynthesis is caused by “non-stomatal” mechanisms. The changes in photosynthetic activity are connected with membrane damage in mesophyll cells, decrease in chlorophyll content, and disturbance in synthesis and transport of assimilates (Cornic and Massacci 1996). Limitations of photosynthesis by stomatal as well as non-stomatal mechanisms depend not only on the duration and intensity of drought stress but also on plant species, stage of plant development, and leaf age (Berkowitz et al. 1983; Kicheva et al. 1994; Passioura et al. 1993).
A decrease in net photosynthetic rate under water stress is also related to disturbances in biochemical processes of a non-stomatal nature, caused by oxidation of chloroplast lipids and changes in the structure of pigments and proteins. Drought stress causes an increase in the content of reactive oxygen species (ROS) (Dhanda et al. 2004; Li et al. 2004; Miller et al. 2010; Rauf et al. 2007; Shao et al. 2005a). In response to drought-induced oxidative stress, plants increase the activity of anti-oxidative peroxidases or glutathione reductase (Gill and Tuteja 2010; Miller et al. 2010; Neill et al. 2002). In photosystem 2 (PS2), the occurrence of ROS is caused by damage to thylakoid membranes, when electrons from water are transferred to oxygen. Chloroplasts are very susceptible to oxidative stress, mainly due to high oxygen concentrations inside these organelles, which is a result of irradiation generating singlet oxygen (Baker 1993; Cornic and Briantais 1991). When water is not available to the leaves in sufficient quantity, higher water use efficiency (WUE) appears to be an alternative strategy to improve crop performance (Araus et al. 2002). WUE may be modified, not only through a decrease in stomata conductance but also through an increase in photosynthetic capacity.
Osmotic adjustment in plants subjected to water deficit may occur through an accumulation of low molecular weight organic solutes. The compatible osmolytes found in higher plants are soluble carbohydrates and proline. The accumulation of soluble carbohydrates in plants has been widely reported as a response to salinity (Ashraf and Harris 2004) and drought (Zhang et al. 2009) in addition to a significant decrease in the net CO2 assimilation rate. Proline, which is widely found in higher plants, accumulates in stressed plants in larger amounts than other amino acids (Ghaderi and Siosemardeh 2011). Proline accumulation is one of the common characteristics in many monocotyledons under water deficit. Proline regulates the accumulation of useable nitrogen, is osmotically active and contributes to membrane stability (Bandurska 2000; Bandurska et al. 2008; DaCosta and Huang 2006; Javadi et al. 2008). It may also act as a signaling regulatory molecule able to activate multiple responses that are components of the adaptation process (Maggio et al. 2002).
The wheat genotype Chinese Spring (CS) used in this study has been shown to be relatively drought-resistant (Galiba et al. 1989), and to maintain relatively high yields under drought field conditions compared to those of the breeding line SQ1 (Dodig and Quarrie, data unpublished). CS and SQ1 have been used to make a mapping population of doubled haploid lines for quantitative trait locus (QTL) analysis of stress responses (Quarrie et al. 2005). As a prerequisite for future work on the genetic control of these physiological and biochemical responses to osmotic stress, the experiment described here was performed under PEG-induced osmotic stress on the two parents. The objective of this experiment was to determine the optimum concentration of PEG to expose the differences in the physiological and biochemical traits described above between CS and SQ1 grown under osmotic stress.
Materials and methods
Plant material
Seedlings of drought-resistant (Chinese Spring, CS) and drought-susceptible (SQ1) hexaploid wheat (Triticum aestivum L.) were examined in the experiment. The genotype SQ1 was selected at the Plant Breeding Institute, UK, from the seventh cross generation (F7) between two wheat genotypes: Highbury × TW269/9/3/4. CS and SQ1 differ significantly in their physiological, morphological and developmental traits. In comparison to CS, SQ1 is shorter with a smaller leaf surface area, and fewer spikes which have awns (Quarrie et al. 1991).
Experimental design
Grains were disinfected in 96 % ethanol for 1.5 min followed by 15 min in 15 % Domestos, before being washed 4 times in sterile water. Afterwards, grains were germinated on wet filter paper for 3 days. Germinating seedlings were put into plastic pots containing water with a half-strength Hoagland solution and maintained in a hydroponics culture in a phytotronic greenhouse for 21 days. The hydroponic solution was aerated by air pumps. Every day, the hydroponic medium was supplemented with a fresh medium and every week, it was completely exchanged with a fresh medium. After 21 days of growth in control (C) treatment (0.0 MPa), seedlings were exposed to three levels of osmotic stress for the following 7 days, namely mild (D1), moderate (D2) and severe (D3). For these treatments, osmotic stress was applied with PEG 6000 dissolved in half-strength Hoagland medium in two steps: first, from day 1 to 3, the osmotic potentials in treatments D1, D2 and D3 were −0.50, −0.75 and −1.00 MPa, respectively, and second from day 4 to 7: −0.75, −1.25 and −1.5 MPa, respectively. The seedlings of each genotype were grown until the fourth leaf was fully expanded.
Measurements and analysis
Gas exchange parameters (Pn, E, g s) were measured using a CIRAS 2 analyzer (PP System), chlorophyll content was measured with a SPAD CL 01 meter (Hansatech) and leaf osmotic potential with a psychrometer HR 33T (WESCOR). Measurements of gas exchange and chlorophyll content were taken on first, third, fourth and seventh day after applying osmotic stress in eight replications (8 leaf seedlings grown in 4 hydroponic containers). On the last day of osmotic stress, plant height, length of roots, fresh weight (FW), fresh weight at full turgor (TW) and dry weight (DW) of stem and roots were measured in order to determine relative water content {RWC = (FW − DW/TW − DW) × 100} according to Slatyer (1967). Seedling samples were also collected on that day for the determination of leaf osmotic potential, level of lipid peroxidation (MDA), contents of proline and soluble carbohydrates.
Osmotic potential
The measurements were taken using the mode of “dew point” by microvoltmeter (model HR-33T with C-52 sample chambers, Wescor Inc., Logan, Utah, USA). Leaf discs (∅ = 5 mm) were collected for analysis from the middle part of leaves and were placed in an Eppendorf tube, frozen in liquid nitrogen and stored at −70 °C. During the measurement, leaf samples were thawed at room temperature and the sap from leaf discs was extracted with a syringe and quickly transferred to a leaf chamber. The time needed for the saturation of leaf chambers was set to 40 min. The measurements for each genotype were taken in the dew point mode at room temperature in five replicates.
Lipid peroxidation
Measurements of lipid peroxidation (the concentration of malondialdehyde, MDA) were taken according to Dhindsa et al. (1981). 1 g of fresh leaves was ground in 5 ml of 0.5 % trichloroacetic acid (TCA) and centrifuged (1,000g) for 15 min. The mixture containing 1 ml of the supernatant and 4 ml of 0.5 % thiobarbituric acid (TBA) in 20 % TCA was heated at 100 °C for 30 min and then cooled to room temperature. The specific absorbance (at 532 nm) of the extract (relative to the background absorbance at 600 nm) was detected. The concentration of MDA was expressed in μM g−1 FW (fresh weight of leaves), using a molar extinction coefficient equal to 155 × 105 mM−1 cm−1.
Soluble carbohydrates
Sugar content was analysed spectrophotometrically according to Dubois et al. (1951) with modifications. About 5 mg of lyophilized and homogenized samples were extracted in 1.5 ml of 96 % ethanol for 15 min. Then the samples were centrifuged at 21,000g for 15 min and 40 μl of supernatant was transferred to 10 ml test tubes containing 400 μl of deionised water. After that, 400 μl of 5 % phenol and 2 ml of concentrated sulphuric acid were added. Samples were incubated for 20 min and transferred to 96-well plates. The absorbance was measured at 490 nm. The level of carbohydrates was expressed in μg g−1 DW (dry weight of leaves).
Proline
Proline content was measured spectrophotometrically according to Ting and Rouseff (1979) with modifications. About 5 mg of lyophilized and homogenized samples were extracted in 0.5 ml of 3 % 5-sulphosalicylic acid for 15 min. Then, the samples were centrifuged at 21,000g for 15 min. The clear supernatant (200 μl) was transferred to polypropylene screw cap vials and 200 μl of concentrated formic acid and 400 μl of 3 % ninhydrin reagent in 2-methoxyethanol were added. Samples were heated for 0.5 h at 100 °C in a water-bath, and then transferred to 96-well plates. The absorbance was measured at 514 nm on a micro-plate reader. The level of proline was expressed in μg g−1 DW (dry weight of leaves).
Statistical analysis
The experiment was performed according to a completely randomized design. Standard errors of means were calculated for all gas exchange parameters. Data were analysed using ANOVA and Duncan’s multiple range test at p < 0.05 with the statistical package STATISTICA 9.0 (Stat-Soft, Inc., USA).
Results
Gas exchange parameters and chlorophyll content
Changes in gas exchange parameters (photosynthesis rate Pn, transpiration rate E, stomatal conductance g s), water use efficiency WUE and chlorophyll content SPAD in CS and SQ1 genotypes were observed during days 1–7 of the experiment and are compared as a percentage of the control (Figs. 1, 2, 3, 4, 5). On the first and third day of stress, differences between the drought-resistant (CS) and drought-susceptible genotype (SQ1) for all treatments (D1, D2 and D3) were not high. Comparable decreases in the values of Pn and E rate were observed for CS and SQ1 after the next 4 and 7 days (Figs. 1, 2). Values of g s differed depending on the genotype, the day of the treatment and also the interaction between them (Table 1; Fig. 3). On the fourth and seventh day of stress, g s decreased more for CS than SQ1 for all treatments. On the third day, the WUE for CS was higher than for SQ1 for all treatments (Fig. 4). On the fourth day of stress, the opposite changes were observed; namely the increase of WUE was greater for SQ1 than for CS. On the last (7th) day of stress, WUE decreased more in SQ1 leaves than CS leaves for all treatments (Fig. 4). Chlorophyll content (SPAD) for SQ1 was slightly higher than for CS on the third and fourth day of stress and maintained this level up to the seventh day of stress (Fig. 5).

Changes in photosynthesis (Pn) in CS and SQ1 wheat seedlings after 1, 3, 4 and 7 days of growth in three levels of osmotic stress (D1, D2 and D3). Data are presented as percentage of control (C). Error bars indicate SE (n = 16)

Changes in transpiration rate (E) in CS and SQ1 wheat seedlings after 1, 3, 4 and 7 days of growth in three levels of osmotic stress (D1, D2 and D3). Data are presented as percentage of control (C). Error bars indicate SE (n = 16)

Changes in stomatal conductance (g s) in CS and SQ1 wheat seedlings after 1, 3, 4 and 7 days of growth in three levels of osmotic stress (D1, D2 and D3). Data are presented as percentage of control (C). Error bars indicate SE (n = 16)

Changes in water use efficiency (WUE) in CS and SQ1 wheat seedlings after 1, 3, 4 and 7 days of growth in three levels of osmotic stress (D1, D2 and D3). Data are presented as percentage of control (C). Error bars indicate SE (n = 16)

Changes in chlorophyll content (SPAD) in CS and SQ1 wheat seedlings after 1, 3, 4 and 7 days of growth in three levels of osmotic stress (D1, D2 and D3). Data are presented as percentage of control (C). Error bars indicate SE (n = 16)
ANOVA analysis of gas exchange parameters and chlorophyll content (Table 1) showed significant differences between CS and SQ1 for all parameters, both under control and three concentrations of PEG 6000. Variation amongst treatment duration (day 1–7) was significant for Pn, E, g s, and SPAD, but for WUE only under severe osmotic stress (D3). Interactions between genotype and treatment duration were highly significant under both control and all stress treatments for g s, but among other gas exchange parameters only for Pn and WUE under the conditions of moderate stress treatment D2.
Plant height, root length, leaf and root relative water and MDA content and leaf osmotic potential
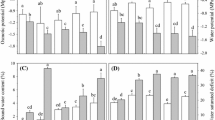
Plant height and root length, leaf and root relative water content (RWC), leaf osmotic potential and MDA content were determined only after 7 days of osmotic stress. Osmotic stress slightly decreased plant height only in the case of D3 treatment of SQ1 and CS genotypes in comparison to control (Fig. 6a). Root length of the CS seedlings grown in D1 treatment increased significantly in comparison to the control, and decreased in the case of D2 (about 40 %) and D3 (about 20 %) treatment (Fig. 6b). In treatments D2 and D3, significant differences between CS and SQ1 were also observed in leaf and root RWC (Fig. 7a, b). RWC of CS leaves decreased about 10 % after D2 and D3 treatment in comparison to the control and RWC of SQ1 leaves was on the same level. The highest decrease (double in comparison to the control) of leaf osmotic potential was noticed in CS under D2 treatment (Fig. 8a). MDA concentrations in leaves showed the greatest differences between CS and SQ1 under D1 osmotic stresses (Fig. 8b), although it should be noted that with the increase of stress level, a higher content of MDA was observed in CS than in SQ1.

Plant height (a) and root length (b) of CS and SQ1 wheat seedlings after 7 days of growth in three levels of osmotic stress (D1, D2 and D3). Values are mean ± SE (n = 3). Mean values followed by the same letter are not significantly different (p = 0.05)

Leaves (a) and roots (b) relative water content (RWC) of CS and SQ1 wheat seedlings after 7 days of growth in three level of osmotic stress (D1, D2 and D3). Values are mean ± SE (n = 3). Mean values followed by the same letter are not significantly different (p = 0.05)

Leaf osmotic potential (a) and content of malondialdehyde (MDA) (b) of CS and SQ1 wheat seedlings after 7 days of growth in three levels of osmotic stress (D1, D2 and D3). Values are mean ± SE (n = 3). Mean values followed by the same letter are not significantly different (p = 0.05)
Contents of proline and soluble carbohydrates
Osmotic stress-induced accumulation of proline in leaves (Fig. 9a), and proline concentration was comparable in response to osmotic stress both for CS and SQ1, although it was more visible in the case of D2 treatment. Under osmotic stress, carbohydrate concentrations tended to increase as the level of stress increased, though the increase was greater in CS than in SQ1 (Fig. 9b) and differences in carbohydrate concentrations between CS and SQ1 were significant only for D3.

Content of proline (a) and carbohydrates (b) in leaves of CS and SQ1 wheat seedlings after 7 days growth in three levels of osmotic stress (D1, D2 and D3). Values are mean ± SE (n = 3). Mean values followed by the same letter are not significantly different (p = 05)
The ANOVA interaction terms genotype × stress treatment and genotype × treatment duration (Tables 1, 2) were used to identify the most appropriate stress treatment for future research. Despite highly significant differences between genotypes and parameters investigated, all the interactions between genotypes and stress treatments (D1, D2, D3) were significant. Thus, stress treatment highlighted the optimum differential between CS and SQ1, though interactions between genotype and treatment duration were greatest for the medium stress treatment D2 (Table 2).
Discussion
We found that osmotic stress affected gas exchange parameters, chlorophyll content, seedlings morphology, water status, osmotic potential, lipid peroxidation, proline and soluble sugar contents in drought-resistant and drought-susceptible wheat genotype. Grzesiak et al. (2006) suggested that a major physiological reason for susceptibility differing between drought-susceptible and drought-resistant triticale and maize genotypes was the more efficient protection of tissue water status in resistant genotypes through greater decreases in stomatal conductance, and thus in the transpiration rate, compared with susceptible genotypes. Our results confirmed these observations and revealed higher decrease of gas exchange parameters on fourth and seventh day of PEG treatment for resistant CS in comparison to the more susceptible SQ1. At the same time, water use efficiency (WUE) was higher for this genotype. CS genotype showed a higher decrease in chlorophyll content (measured in SPAD units) than SQ1 in all three stress level treatments. There were also significant differences between treatment days, although the interactions between genotypes and drought treatment were not significant. The results are in agreement with Mafakheri et al. (2010) and Nyachiro et al. (2001) who described a significant decrease in total chlorophyll caused by water deficit in three chickpea cultivars and in six Triticum aestivum cultivars, respectively. A lower or unchanged chlorophyll level during drought stress has been reported in other species, depending on the duration and severity of the drought (Mafakheri et al. 2010). A decrease in total chlorophyll with drought stress implies a lowered capacity for light harvesting. Since the production of reactive oxygen species is mainly driven by excess energy absorption in the photosynthetic apparatus, this might be avoided by degrading the absorbing pigments (Mafakheri et al. 2010; Esfandiari et al. 2008). Lawlor and Cornic (2002) suggested that actual plant water status in leaves depends on the osmotic conditions of cells and transport of water from the shoots. During the inhibition of water transport from the root, osmotic regulation may actively influence water potential in assimilating tissues and limit the detrimental effects of water deficiency on photosynthesis. Adaptation to low water potential was observed in sunflower under mild water stress in comparison to non-acclimated plants subjected directly to severe drought, in which full inhibition of photosynthesis occurred (Cornic and Fresnau 2002; Medrano et al. 2002). According to the cited authors, osmotic regulation in leaf tissue is responsible for such an impact of drought on photosynthesis by directly influencing stomatal regulation and adaptation of the photosynthetic apparatus. Stomatal (during mild drought) or non-stomatal (during severe drought) mechanisms are responsible for the decrease in photosynthesis under water deficit conditions. Grzesiak et al. (2006) compared the influence of osmotic stress simulated by mannitol solutions from −0.01 to −2.45 MPa to changes in leaf water potential and parameters of gas exchange in drought-resistant and drought-susceptible genotypes of triticale and maize. According to their findings, with the increase in the concentration of the mannitol solution, the impact of drought on gas exchange parameters and water potential was higher for susceptible genotypes than for the resistant ones. In comparison to resistant CS, there was a decrease in susceptible SQ1 of upper ground seedlings growth with the increase in concentration of PEG. The maximum value for plant height and root length was observed in the control (without PEG) and the minimum value was observed in SQ1 roots under D3 treatment. Other researchers, including Singh (2000), Rakesh et al. (1998), Khan et al. (2002) and Rauf et al. (2007) conducted similar studies and showed that growth of wheat seedlings was affected significantly by the change in water stress level. Osmotic stress induced by PEG 6000 caused a decrease in relative water content (RWC) in leaves and roots of drought-resistant CS under D2 and D3, compared to control and D1 treatment as well as susceptible SQ1. There were no significant differences in RWC of SQ1. These results are in agreement with Yagmur and Kaydan (2008) who also observed a decrease of RWC in hexaploid triticale when drought has been induced by PEG 6000. The greatest decrease (as much as twice the percentage of the control) of osmotic potential was observed in CS genotype in D2 treatment. Lower values of the osmotic potential lead to the conclusion that the resistant genotype has adapted to drought through effective osmoregulation (Hura et al. 2010). According to Hura et al. (2007), during osmotic stress induction by PEG, CS genotype displayed a capability for osmoregulation, which enabled it to maintain a relatively high volume of turgor protoplasts and high activity of the photosynthetic apparatus. Moreover, osmoregulation enables a high water content to be maintained in the leaf which, in turn, causes a decrease in the activity of the photosynthetic apparatus. It has been found that PEG treatment could induce alternative electron transport processes (e.g. Mehler reaction), thereby preventing the over-reduction of electron transport components of the photosynthetic apparatus. We observed that SQ1 genotype had lower lipid peroxidation measured as MDA content than CS under all D1, D2 and D3 treatments and displayed higher membrane integration under osmotic stress.
MDA is a common and important index for evaluating the redox status of wheat. Lower MDA content means higher antioxidative ability, reflecting higher drought resistance as suggested by Shao et al. (2005a) and Dhanda et al. (2004). In our experiment, it was shown that the genotypes tested clearly responded to water deficiency differently. This further implied that different wheat genotypes had discrete water stress thresholds (Shao et al. 2005b). Therefore, resistant CS has a different physiological adaptive mechanism to regulate its redox status from the more susceptible SQ1. Osmoregulation is usually considered to be the critical factor under water deficient conditions. Peroxidation of lipids can disturb the assembly of the membrane, causing changes in fluidity and permeability, alterations of ion transport and inhibition of metabolic processes (Catalá 2006). Peroxidation can also greatly alter the physicochemical properties of membrane lipid bilayers, resulting in severe cellular dysfunction and biosynthesis of MDA. A lower MDA content in CS shows its higher anti-oxidative capability, reflecting a higher resistance to drought in comparison to SQ1, where the content of MDA increased when osmotic stress increased. Plants respond to drought stress using different physiological and biochemistry strategies. Some osmolytes such as proline and soluble sugars are widespread throughout the plant kingdom. Proline and carbohydrates are the two most important organic solutes that are accumulated in higher plants under drought conditions (Changhai et al. 2010; P’erez-Alfocea and Larcher 1995; Sumera and Asghari 2010). It is well known that free proline level increases in response to drought (Chorfi and Taïbi 2011; Moayedi et al. 2011; Rampino et al. 2006). This was confirmed in our experiment, where the applied concentration of PEG caused a higher accumulation of proline under D2 and D3 than under D1 treatment and in the resistant CS than in the susceptible SQ1 genotype. The accumulation of proline serves as a depot for energy to regulate redox potentials (Hong-Boa et al. 2006; Saradhi and Saradhi 1991), and functions as a hydroxyl radical scavenger (Smirnoff and Cumbes 1989). Our results showed that concentration of soluble carbohydrates was higher after D2 and D3 treatments in comparison to the control. CS tended to accumulate more soluble carbohydrates than SQ1, indicating that there might be a genetic variation in the accumulation of these compounds. Carbohydrates seem to play a key role in the integration of plant growth and appear to be part of a wider mechanism for balancing carbon acquisition and allocation within and between organs (Farrar et al. 2000). It has been suggested that under water stress conditions, soluble sugars can function in two ways which are difficult to separate, namely osmotic agents and osmoprotectors (Bohnert et al. 1995; Ingram and Bartels 1996; Yong et al. 2006). As osmotic agents soluble sugars facilitate osmotic adjustment, as osmoprotectors they stabilize proteins and membranes, most likely substituting water in the formation of hydrogen bonds with polypeptide polar residues and phospholipid phosphate groups.
Conclusion and future prospects
The seedlings of wheat genotypes which were used in this experiment differed in their morphological, physiological and biochemical characteristics. In this experiment, the varying responses to osmotic stress of CS and SQ1 wheat genotypes were shown, as was their dependency on PEG concentration and osmotic stress duration. This variation has a genetic background and confirms the existence of a genetically determined resistance to water stress among different crop genotypes. The obtained results indicate that in the search for the right experimental design for the investigation of osmotic and drought stress resistance in wheat, a proper PEG concentration can be found to reveal the differences between resistant CS and susceptible SQ1 genotypes. This optimum concentration of PEG found in this study will be used in further research to evaluate the influence of PEG solution in hydroponics culture with chosen phytohormones and osmo-protectants, like abscisic acid, salicylic acid and polyamines on osmotic stress amelioration in CS and SQ1 genotypes and the double haploid mapping population derived from parents CS and SQ1 in order to identify QTLs involved in drought resistance.
Author contribution
IM, IC-M and ES designed the research; IM, IC-M, ES, MF, TH, MD, SG, MTG, AN and KD conducted the research; IM, IC-M, MTG and ES analysed the data; IM, FJ, and SAQ wrote the paper; IM had primary responsibility for the final content. All authors have read and approved the final manuscript.
References
Ashraf M, Harris PJC (2004) Potential biochemical indicators of salinity tolerance in plants. Plant Sci 166:3–16
Baker NR (1993) Light-use efficiency and photoinhibition of photosynthesis in plants under environmental stress. In: Smith JAC, Griffiths H (eds) Water deficits. Plant responses from cell to community. BIOS Scientific Publishers, Oxford, pp 221–235
Bandurska H (2000) Does proline accumulated in leaves of water deficit stressed barley plants confine cell membrane injury? I. Free proline accumulation and membrane injury index in drought and osmotically stressed plants. Acta Physiol Plant 22:409–415
Bandurska H, Górny AG, Zielezińska M (2008) Effects of water deficit on relative water content, proline accumulation and injury of cell membranes in leaves of old and modern cultivars of winter wheat. Zeszyty Problemowe Postępów Nauk Rolniczych 524:115–126
Berkowitz GA, Chen C, Gibbs M (1983) Stromal acidification mediates in vivo water stress inhibition of non stomatal controlled photosynthesis. Plant Physiol 72:1123–1126
Blum A (1998) Improving wheat grain filling under stress by stem reserve mobilization. Euphytica 100:77–83
Blum A, Ebercon A (1981) Cell membrane stability as a measure of drought and heat tolerance in wheat. Crop Sci 21:43–47
Bohnert HJ, Nelson DE, Jensen RG (1995) Adaptations to environmental stresses. Plant Cell 7:1099–1111
Catalá A (2006) An overview of lipid peroxidation with emphasis in outer segments of photoreceptors and the chemiluminescence assay. Int J Biochem Cell Biol 38(9):1482–1495
Changhai S, Baodi D, Yunzhou Q et al (2010) Physiological regulation of high transpiration efficiency in winter wheat under drought conditions. Plant Soil Eviron 56(7):340–347
Chaves MM, Pereira JS, Maroco J et al (2002) How plants cope with water stress in the field. Photosynthesis and growth. Ann Bot 89:907–916
Chorfi A, Taïbi K (2011) Biochemical screening for osmotic adjustment of wheat genotypes under drought stress. Tropicult 29(2):82–87
Cornic G, Briantais J-M (1991) Partitioning of photosynthetic electron flow between CO2 and O2 reduction in a C3 leaf (Phaseolus vulgaris L.) at different CO2 concentrations and during drought stress. Planta 183:178–184
Cornic G, Fresnau C (2002) Photosynthetic carbon reduction and carbon oxidation cycles are the main electron sinks for photosystem II activity during a mild drought. Ann Bot 89:887–894
Cornic C, Massacci A (1996) Leaf photosynthesis under drought stress. In: Baker NR (ed) Photosynthesis and environment. Kluwer Academic Publishers, The Netherlands, pp 347–366
DaCosta M, Huang BR (2006) Osmotic adjustment associated with variation in bentgrass tolerance to drought stress. J Am Soc Hortic Sci 131(3):338–344
Dhanda SS, Sethi GS, Behl RK (2004) Indices of drought tolerance in wheat genotypes at early stages of plant growth. J Agron Crop Sci 190(1):6–12
Dhindsa RS, Plumb-Dhindsa P, Thorpe TA (1981) Leaf senescence: correlation with increased levels of membrane permeability and lipid peroxidation, and decreased levels of superoxide dismutase and catalase. J Exp Bot 32:96–101
Dubois M, Gilles K, Hamilton JK et al (1951) A colorimetric method forth determination of sugars. Nature 168:167–168
Esfandiari E, Shakiba MR, Mahboob SA et al (2008) The effect of water stress on the antioxidant content, protective enzyme activities, proline content and lipid peroxidation in wheat seedling Pakistan. J Biol Sci 11(15):1916–1922
Farrar J, Pollock C, Gallanger J (2000) Sucrose and the interaction of metabolism in vascular plants. Plant Sci 154:1–11
Galiba G, Simon-Sarkadi L, Salgo A et al (1989) Genotype dependent adaptation of wheat varieties to water stress in vitro. J Plant Physiol 134:730–735
Ghaderi N, Siosemardeh A (2011) Response to drought stress of two strawberry cultivars (cv. Kurdistan and Selva). Hortic Environ Biotechnol 52(1):6–12
Gill SS, Tuteja N (2010) Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiol Biochem 48:909–930
Grzesiak S, Grzesiak MT, Filek W et al (2003) Evaluation of physiological screening tests for breeding drought resistant triticale (×Triticosecale Wittmack). Acta Physiol Plant 25:29–37
Grzesiak MT, Grzesiak S, Skoczowski A (2006) Changes of leaf water potential and gas exchange parameters during and after drought in triticale and maize genotypes differing in drought tolerance. Photosynthetica 44:561–568
Hong-Boa S, Xiao-Yan C, Li-Ye C et al (2006) Investigation on the relationship of proline with wheat anti-drought under soil water deficits. Colloids Surf B Biointer 53:113–119
Hura T, Grzesiak S, Hura K, Thiemt E, Tokarz K, Wedzony M (2007) Physiological and biochemical tools useful in drought-tolerance detection in genotypes of winter triticale: accumulation of ferulic acid correlates with drought tolerance. Ann Bot 100:767–775
Hura T, Hura K, Grzesiak S, Banaszak Z (2010) Simulation of osmotic stress during the early stages of triticale development as a promising laboratory test for screening drought resistance. Cer Res Commun 38(3):327–334
Ingram J, Bartels D (1996) The molecular basis of dehydration tolerance in plants. Ann Rev Plant Physiol Plant Mol Biol 47:377–403
Jackson P, Rubertson M, Cupper M, Hammer G (1996) The role of physiological understanding in plant breeding from breeding perspective. Field Crops Res 49:11–37
Javadi T, Arzani K, Ebrahimzadeh H (2008) Study of proline, soluble sugar, and chlorophyll a and b changes in nine Asian and one European pear cultivar under drought stress. Acta Hortic 769:241–246
Jl Araus, Slafer GA, Reynolds MP, Royo C (2002) Plant breeding and drought in C3 cereals: what should we breed for? Ann Bot 89:925–940
Kerepesi I, Galiba G (2000) Osmotic and salt stress-induced alteration in soluble carbohydrate content in wheat seedlings. Crop Sci 40:482–487
Khan MQ, Anwar S, Khan MI (2002) Genetic variability for seedling Traits in wheat (Triticum aestivum L.) under moisture stress conditions. Asian J Plant Sci 1(5):588–590
Kicheva MI, Tsonev TD, Popova LP (1994) Stomatal and nonstomatal limitations to photosynthesis in two wheat cultivars subjected to water stress. Photosynthetica 30:107–116
Lawlor DW, Cornic G (2002) Photosynthetic carbon assimilation and associated metabolism in relation to water deficits in higher plans. Plant, Cell Environ 25:275–294
Li C-Z, Jiao J, Wang G-X (2004) The important roles of reactive oxygen species in the relationship between ethylene and polyamines in leaves of spring wheat seedlings under root osmotic stress. Plant Sci 166:303–315
Lorens GF, Bennett JM, Loggale LB (1987) Differences in drought resistance between two corn hybrids. II. Component analysis and growth rates. Agron J 79:808–813
Mafakheri A, Siosemardeh A, Bahramnejad B, Struik PC, Sohrabi E (2010) Effect of drought stress on yield, proline and chlorophyll contents in three chickpea cultivars. Aust J Crop Sci 4(8):580–585
Maggio A, Miyazaki S, Veronese P et al (2002) Does proline accumulation play an active role in stress induced growth reduction? Plant J 31:699–712
Mansfield TA, Davies WJ (1981) Stomata and stomatal mechanisms. In: Paleg LG, Aspinall D (eds) The physiology and biochemistry of drought resistance in plants. Academic Press, New York, pp 315–346
Martiniello P, Lorenzoni C (1985) Response of maize genotypes to drought tolerance tests. Maydica 30:361–370
Medrano H, Escalona JM, Bota J, Gulías J, Flexas J (2002) Regulation of photosynthesis of C3 plants in response to progressive drought: stomatal conductance as a reference parameter. Ann Bot 89:895–905
Miller G, Suzuki N, Ciftci-Yilmaz S, Mittler R (2010) Reactive oxygen species homeostasis and signalling during drought and salinity stresses. Plant Cell Environ 33:453–467
Moayedi AA, Nasrulhaq-Boyce A, Tavakoli H (2011) Application of physiological and biochemical indices for screening and assessment of drought tolerance in durum wheat genotypes. Aust J Crop Sci 5(8):1014–1018
Neill SJ, Desikan R, Clarke A, Hurst RD, Hancock JT (2002) Hydrogen peroxide and nitric oxide as signalling molecules in plants. J Exp Bot 53:1237–1247
Nyachiro JM, Briggs KG, Hoddinott J, Johnson-Flanagan AM (2001) Chlorophyll content, chlorophyll fluorescence and water deficit in spring wheat. Cereal Res Commun 29:135–142
P’erez-Alfocea F, Larcher F (1995) Sucrose and proline accumulated and sugar efflux in tomato leaf discs affected by NaCl and polyethylene glycol 6000 isoosmotic stresses. Plant Sci 107:9–15
Passioura JB, Condon AG, Richards RA (1993) Water deficits, the development of leaf area and crop productivity. In: Smith JAC, Griffiths H (eds) Water deficits. Plant responses from cell to community. BIOS Scientific Publishers, Oxford, pp 253–264
Quarrie SA, Steed A, Calestani C, Abugalieva A et al (1991) Implications of genetic differences in ABA accumulation for crop production. In: Davies WJ, Jones HG (eds) Abscisic acid physiology and biochemistry. BIOS Scientific Publishers, pp 227–243
Quarrie SA, Steed A, Calestani C et al (2005) A high density genetic map of hexaploid wheat (Triticum aestivum L.) from the cross Chinese Spring × SQ1 and its use to compare QTLs for grain yield across a range of environments. Theor Appl Genet 110:865–880
Rakesh V, Sethi SS, Vaishnavi R (1998) Relative drought tolerance of rye-introgressed bread wheat (Triticum aestivum L.) genotypes in osmoticum. Ann Biol Ludhiana 14(2):169–173
Rampino P, Pataleo S, Gerardi C et al (2006) Drought stress response in wheat: physiological and molecular analysis of resistant and sensitive genotypes. Plant Cell Environ 29:2143–2152
Rauf M, Munir M, Hassan M, Ahmad M, Afzal M (2007) Performance of wheat genotypes under osmotic stress at germination and early seedling growth stage. Afr J Biotechnol 6(8):971–975
Saradhi A, Saradhi PP (1991) Proline accumulation under heavy metal stress. J Plant Physiol 138:554–558
Shao HB, Liang ZS, Shao MA, Wang BC (2005a) Changes of anti-oxidative enzymes and membrane peroxidation for soil water deficits among 10 wheat genotypes at seedling stage. Colloids Surf B 42:107–113
Shao HB, Liang ZS, Shao MA, Wang BC (2005b) Impacts of PEG-6000 pretreatment for barley (Hordeum vulgare L.) seeds on the effect of their mature embryo in vitro culture and primary investigation on its physiological mechanism. Colloids Surf B Biointer 41(2–3):73–77
Singh KP (2000) Effect of osmotic water stress on germination of wheat. Ann Plant Physiol 14(1):98–100
Slatyer RO (1967) Plant–water relationships. Academic Press, London 366
Smirnoff N, Cumbes QJ (1989) Hydroxyl radical scavenging activity of compatible solute. Phytochemistry 28:1057–1060
Sumera I, Asghari B (2010) Effect of drought and abscisic acid application on the osmotic adjustment of four wheat cultivars. J Chem Soc Pakistan 32(1):13–19
Tan Y, Liang Z, Shao H et al (2006) Effect of water deficits on the activity of anti-oxidative enzymes and osmoregulation among three different genotypes of Radix Astragali at seeding stage. Colloid Surf B Biointer 49:60–65
Ting SV, Rouseff RL (1979) Proline content in Florida frozen concentrated orange juice and canned grapefruit juice. Proc Fla State Hortic Soc 92:143–145
Winter SR, Musick JT, Porter KB (1988) Evaluation of screening techniques for breeding drought resistant winter wheat. Crop Sci 28:512–516
Yagmur M, Kaydan D (2008) Alleviation of osmotic stress of water and salt in germination and seedling growth of triticale with seed priming treatments. Afr J Biotechnol 7:2156–2162
Yong T, Zongsuo L, Hongboc S, Feng D (2006) Effect of water deficits on the activity of anti-oxidative enzymes and osmoregulation among three different genotypes of Radix Astragali at seeding stage. Colloids Surf B 49:60–65
Zhang J, Dell B, Conocono E, Waters I, Setter T, Appels R (2009) Water deficits in wheat: fructan exohydrolase (1-FEH) mRNA expression and relationship to soluble carbohydrate concentrations in two varieties. New Phytol 81(4):843–850
Acknowledgments
This research was supported by the Ministry of Science and Higher Education in Poland, No. 480/N-COST-2009/0.
Open Access
This article is distributed under the terms of the Creative Commons Attribution License which permits any use, distribution, and reproduction in any medium, provided the original author(s) and the source are credited.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by K. Trebacz.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 2.0 International License (https://creativecommons.org/licenses/by/2.0), which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.
About this article
Cite this article
Marcińska, I., Czyczyło-Mysza, I., Skrzypek, E. et al. Impact of osmotic stress on physiological and biochemical characteristics in drought-susceptible and drought-resistant wheat genotypes. Acta Physiol Plant 35, 451–461 (2013). https://doi.org/10.1007/s11738-012-1088-6
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s11738-012-1088-6