
Abstract
Psychedelics are powerful psychoactive substances that alter perception and mood processes. Their effectiveness in the treatment of psychiatric diseases was known before their prohibition. An increasing number of recent studies, due to the indisputable resurgence of serotonergic hallucinogens, have shown their efficacy in alleviating depression, anxiety, substance abuse therapies, and existential distress treatment in patients facing life-threatening illness. Psychedelics are generally considered to be physiologically safe with low toxicity and low addictive potential. However, their agonism at serotonergic receptors should be considered in the context of possible serotonin-related cardiotoxicity (5-HT2A/2B and 5-HT4 receptors), influence on platelet aggregation (5-HT2A receptor), and their proarrhythmic potential. The use of psychedelics has also been associated with significant sympathomimetic effects in both experimental and clinical studies. Therefore, the present review aims to provide a critical discussion of the cardiovascular safety of psilocybin, d-lysergic acid diethylamide (LSD), N,N-dimethyltryptamine, ayahuasca, and mescaline, based on the results of experimental research and clinical trials in humans. Experimental studies provide inconsistent information on the potential cardiovascular effects and toxicity of psychedelics. Data from clinical trials point to the relative cardiovascular safety of psychedelic-assisted therapies in the population of “healthy” volunteers. However, there is insufficient evidence from therapies carried out with microdoses of psychedelics, and there is still a lack of data on the safety of psychedelics in the population of patients with cardiovascular disease. Therefore, the exact determination of the cardiovascular safety of psychedelic therapies (especially long-term therapies) requires further research.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Psychedelics are a class of psychoactive substances, whose “acute” effects resulting primarily from serotonergic 5-HT2A receptors agonism cover a wide spectrum of behavioral, psychological, and physiological effects [1,2,3].
A range of symptoms, particularly those affecting the state of consciousness, caused by plants or plant-derived substances and fungi containing psychedelic substances, have led to their use for millennia in religious practices, tribal rituals, and also for the treatment of fever and rheumatic pain [4,5,6]. The modern era of psychedelic research did not begin with the synthesis of a new substance in this group, d-lysergic acid diethylamide (LSD) in 1938, but with the accidental discovery of its effects by Albert Hoffmann on 19 April, 1943, (known as Bike Day) [7]. This opened the door to extensive research into the use of the LSD molecule (then produced under the trade name Delysid) in the treatment of mental illnesses, such as: mood disorders, anxiety, and acute stress reaction [1, 7, 8]. Unfortunately, it was also a time of tumultuous social and political change (for example, the ongoing war in Vietnam and the emergence of the “counterculture” movement). Anti-war sentiments, rising crime rates and the growing wave of drug addiction, psychedelic abuse, the sale of powerful hallucinogens (e.g., 2,5-dimethoxy-4-methylamphetamine, DOM, known as STP) as LSD on the black market and increasing numbers of hospital admissions related to the experience of “bad trips” in the United States of America and many other Western countries led to the stigmatization, vilification, and finally the criminalization of psychedelics in 1970 with the Controlled Substances Act (Schedule I). Following the restrictions introduced in the United States, strict drug control laws were introduced in other Western countries. These historical and political factors have severely limited scientific research into the clinical potential of psychedelic substances.
Recently, there has been renewed interest in psychedelics in the context of neuropharmacotherapy for mental illness. This is related to the growing global burden of mood and anxiety disorders and the emergence of promising clinical research results on psychedelic substances in the pharmacotherapy of depressive and anxiety disorders, in particular, post-traumatic stress disorder (PTSD), substance use disorders, and chronic pain [9,10,11,12,13]. Classical psychedelics have been shown to catalyze relatively long-lasting improvements in mental health after a small number of doses, especially when combined with psychotherapy [12]. Following the rapidly developing field of clinical research in psychedelic medicine, there have also been important changes at the regulatory level, with the decriminalization of natural psychedelics in Western countries (certain states in the USA, Canada, Australia, Switzerland, and the Netherlands) and the publication of guidelines for conducting research involving the use of psychedelics (U.S. Food and Drug Administration Guidance issued 20 June 1987; FDA-2023-D-1987) [14].
About the renaissance of psychedelics, apart from the clinical benefits, an analysis of the adverse effects and possible risks associated with their use in a medical context should be essential. In particular, we should focus on the pharmacological and molecular mechanisms and assess the cardiovascular safety associated with psychedelic therapy in randomized clinical trials. In the context of applied medical therapies, psychedelics are generally considered to be physiologically safe. They have low toxicity and low addictive potential [14, 15]. However, their agonism at serotonergic receptors should be investigated because of possible serotonin-related cardiotoxicity, influence on platelet aggregation, and proarrhythmic potential. The use of psychedelics has also been associated with significant sympathomimetic effects in both experimental and clinical studies [1, 16].
The present review aimed to undertake a critical discussion of the cardiovascular safety of classical psychedelics in pharmacotherapy based on pharmacological mechanisms as well as experimental and clinical research results. First, the cardiovascular effects of classical psychedelics in experimental studies were analyzed by searching the PubMed database. Then, documentation of clinical trials, observational studies of classical psychedelics was collected using the PubMed database. The searches were performed between July 1 and August 15, 2023, using the search terms psilocybin, mescaline, LSD, d-lysergic acid, ayahuasca, dimethyltryptamine, DMT and the clinical trials filter. Of these articles, only primary reports of classic psychedelics administered by a health professional alone were included. The presence of information on the observation and reporting of the presence or absence of cardiovascular effects after psychedelic treatment was analyzed in all retrieved articles and their supplementary materials (if available). The apparent adverse effects of this group of substances in terms of toxicity associated with non-medical use are, therefore, not addressed in this review.
The mechanism of action of psychedelics
Classical psychedelics can be classified as tryptamines (e.g., psilocybin, its metabolite psilocin, N,N-dimethyltryptamine – DMT), phenethylamines (e.g., mescaline and its derivatives) and lysergamides (LSD) [9]. Experimental studies in the 1960s suggested that LSD activates the serotonergic system in the brain [1, 13, 17, 18]. The discovery of 5-HT receptor populations and the subsequent synthesis of selective 5-HT2 receptor antagonists enabled further development of knowledge on the mechanisms of the action of psychedelics. In particular, in a rat drug discrimination model study, Glennon et al. [19] were the first to show that blockade of 5-HT2 receptors with ketanserin and pirenperone abolished the effects produced by psychedelics. In support of this study, the binding of various hallucinogens to different populations of serotoninergic receptors using radioligand techniques indicated a common affinity only for 5-HT2, specifically 5-HT2A receptors [19,20,21].
Further animal and human studies have shown that different efficacy at 5-HT2A receptors is a key pharmacological mechanism responsible for the psychedelic effects [22,23,24] and that different psychedelic substances also have affinities for other receptors: serotonergic, dopaminergic, glutamatergic, adrenergic, and histaminergic [25,26,27,28]. Psychedelic stimulation of 5-HT2A receptors disrupts the cortical rhythm and large-scale brain networks through an increase in neocortical neuron excitation and subsequent augmentation of extracellular glutamate release in the prefrontal cortex [21, 29, 30]. In addition, psychedelics can affect neurotransmission by inhibiting transporters for the monoamine serotonin transporter (SERT) and vesicular monoamine transporter (VMAT2), trace amine-associated receptors (TAARs), and also monoamine oxidase (MAO) [31, 32]. Numerous experimental and human studies have confirmed the hypothesis of psychedelics as so-called ‘psychoplastogens’ as they were shown to enhance neurogenesis, neuroplasticity, and synaptic plasticity by increasing the expression of markers of neuroplastic processes (c-fos Arc) and brain-derived neurotrophic factor (BDNF) [33,34,35,36]. Recent animal studies suggest that the neuroplastic effects of psilocybin and LSD are associated with potent agonism (1000 times greater than classical antidepressants) at the tropomyosin-related kinase receptor B (TrkB) receptor for BDNF. This results in the enhancement of neurotrophic, plasticity and antidepressant-like behavior. It also promotes endogenous BDNF signaling [36].
Mechanisms of the cardiovascular effects of psychedelics
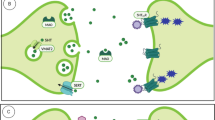
The effects of psychedelics on the cardiovascular system should be considered in terms of their effects on the 1) serotonergic system in the heart and the vasculature and/or their 2) sympathomimetic effects (Fig. 1).

Cardiovascular effects of classical psychedelics according to experimental and clinical data (black text) and possible effects that need experimental and clinical investigation (blue text). 5-HT serotonin; MAO monoamine oxidase; PAH pulmonary arterial hypertension; SERT serotonin transporter; TAARs trace amine-associated receptors; VMAT vesicular monoamine transporter
Cardiovascular effects of serotonin
Early experiments at the beginning of the twentieth century showed that blood platelets, as well as a substance isolated from enterochromatophilic cells (at that time called enteramine, later serotonin), act on smooth muscle by causing it to contract [37,38,39]. This substance was later identified as 5-hydroxytryptamine [39]. In adults, serotonin is mainly stored in dense platelet granules from which it is released during activation and aggregation phenomena, causing the vasoconstrictive effects described above. This circulating pool of 5-HT originates from the loading of platelets with serotonin synthesized in the intestinal wall. Serotonergic receptors are present in both the vascular endothelium (5-HT1B, 5-HT2B, and 5-HT7) and vascular smooth muscle (5-HT1B, 5-HT2A, and 5-HT7). Serotonin can produce both pressor and depressor effects. The diversity of vascular responses to 5-HT depends on the concentration, receptor site, and signaling mechanisms activated [40, 41]. In both humans and animals, 5-HT causes direct vasoconstriction predominantly via the 5-HT2A receptor and, to a lesser extent, via the 5-HT1B and 5-HT1D receptors [41, 42]. Serotonin also plays an important role in regulating coronary smooth muscle tone. Animal models and in vitro studies on the human coronary endothelium have shown that serotonin increases nitric oxide production, coronary vasodilation, and coronary flow via the 5-HT1B and 5-HT2B receptors [43,44,45]. Several observations provided evidence for increased vascular reactivity, i.e., a vasoconstrictor response to serotonin in damaged vessels (e.g., arterial hypertension). Namely, in an animal model of spontaneous hypertension (SHR), a six-fold and five-fold higher contractile sensitivity in the aorta and mesenteric artery, respectively [46], and an excessive central pressor response after exposure to 5-HT [47] have been demonstrated when compared to a normotensive control. The hypersensitivity to serotonin in experimental models of arterial hypertension may be related to enhanced expression of 5-HT2B serotonergic receptors in the smooth muscle of the vasculature [41, 48].
A large number of experimental studies and clinical observations have linked serotonergic hyperactivity to the development of pulmonary arterial hypertension (PAH). This is due to its vasoconstrictive and mitogenic effects on the vascular smooth muscle cells [49]. In addition, patients with PAH have been found to have high plasma serotonin concentrations [50]. Furthermore, both in humans and in experimental models of pulmonary hypertension, increased expression of the serotonergic receptors 5-HT2B and, to a lesser extent, 5-HT2A has been observed in pulmonary arteries [51,52,53]. There is also strong evidence that hypoxia alters protein expression and proliferative processes through mechanisms involving serotonin, its receptors, and its transporter. This leads to the physiological pulmonary responses to hypoxia, namely vasoconstriction and remodeling of the pulmonary vasculature [54].
Serotonin potentiates platelet aggregation via the 5-HT2A receptors. It promotes thrombosis and increases the risk of thrombotic events [55]. Serotonin 5-HT2A receptors are expressed on the membrane of human platelets, and their blockade by small drug-like molecules suppresses platelet aggregation induced by the extracellular release of 5-HT and P-selectin from platelets [56]. Further studies have provided evidence that 5-HT2A antagonists were effective in the inhibition of collagen-induced platelet aggregation and in offering protection against experimental thrombosis in animal models [57]. More recently, experimental studies have shown that sarpogrelate, a selective 5-HT2A receptor antagonist, prevented restenosis and thrombosis after stent implantation [58]. In addition, in patients with coronary artery disease, the expression of the 5-HT2A receptor was increased [59]. Interestingly, a very recent clinical study revealed that acute treatment with psychotropic drugs with 5-HT2A antagonist properties was associated with significantly lower mortality in elderly patients with severe COVID-19 infection. The mechanism of the protective effect of 5-HT2A receptor antagonists in COVID-19 is unknown but may involve potential immunomodulatory or antiplatelet effects [60].
Apart from the vasculature, serotonin receptors were found in the myocardium as well as in the valve apparatus of the heart. While two main populations of serotonin receptors, 5-HT2 (2A and 2B) and 5-HT4, have been described in the cardiac muscle (in both the atria and the ventricles), the expression of mainly 5-HT2B receptors has been described in cardiac valve tissue [40]. The serotonergic 5-HT2B receptors are expressed in heart valves from various species, including dogs, rats, pigs, monkeys, and humans [40, 61]. The messenger RNAs of the 5-HT1B and 5-HT1D receptor subtypes have been isolated from the human cardiac valve interstitial cells in only one study [62]. Serotonin increases contractility (positive inotropic effect), and reduces diastolic time (positive lusitropic effect) of the atria and ventricles of some mammals, including humans [40, 63, 64]. In addition, serotonin has been shown to increase heart rate (positive chronotropic effect) by activation of the sinus node cells in isolated preparations from humans and isolated samples from pig atria [40, 65, 66]. These effects were mediated by the 5-HT4 receptor and associated with an increase in cAMP (cyclic adenosine monophosphate) content and the subsequent elevation of the phosphorylation state of Ca2+ regulatory proteins [67]. Under normal conditions, stimulation of the 5-HT4 serotonin receptor did not produce significant effects due to the degradation of cAMP by phosphodiesterases. However, in the presence of a phosphodiesterase inhibitor, an effect of serotonin through 5-HT4 receptors has been demonstrated, leading to increased contractility, relaxation, and sometimes arrhythmias. These effects were reminiscent of those produced by catecholamines acting on beta-adrenergic receptors [68].
In the physiological state, the expression level of serotonin 5-HT4 receptors is low, but increases with ventricular dysfunction. Increased expression of the 5-HT4 receptor in the failing heart reflects a cardiac fetal reprogramming process [69, 70]. Cardiac expression of 5-HT2A and 5-HT2B receptors increases in hypertensive left ventricular hypertrophy and heart failure in humans and animals [70,71,72]. In mice, infusion of 5-HT2A antagonists reduced cardiac hypertrophy in a model of cardiac hypertrophy induced by transverse aortic constriction [72]. Therefore, the use of 5-HT2A/2B and 5-HT4 receptor stimulants in heart failure or myocardial hypertrophy should be considered given the possibility of the potentiation of serotonergic effects.
Serotonin has been implicated in pathological valve remodeling. The cardiac changes seen in the carcinoid syndrome, in which about 50 percent of the cases develop valvular lesions, are a clinical model of the effect of excessive serotonergic activity on the development of valvular lesions [61]. As mentioned above, the main serotonergic receptor on heart valve cells is the 5-HT2B receptor. The 5-HT2B receptor initiates several intracellular signaling cascades through both Gq proteins and β-arrestin. The 5-HT2B receptor also activates mitogenic pathways through Src kinase and extracellular regulated kinases (ERK), and enhances the activity of transforming growth factor β (TGF-β) [73]. In myofibroblasts and smooth muscle cells, the biological result of 5-HT2B receptor activation is mitosis, secretion of inflammatory cytokines, and extracellular matrix components [74, 75]. Huang et al. observed a proliferative response to 5-HT2B agonists in 5-HT2B-expressing HEK-293 cells [76]. Recently, in an animal model of transgenic mice, the overexpression of the Gq-coupled 5-HT2B receptor led to an increase in extracellular matrix, hypertrophic remodeling of the heart, and ultimately to reduced ventricular function [77]. In addition, blockade of the 5-HT2B receptor by cyproheptadine prevented pergolide-induced valvulopathy in rats [78].
Cardiovascular effects of psychedelics in experimental studies
As agonists of serotonergic receptors, psychedelics have vasoconstrictive and, therefore, pressor effects. Dyer and Gant [79] reported that psilocin, psilocybin, mescaline, and LSD produced contractions of the isolated umbilical vasculature of humans and sheep, with LSD being the most potent. In another study, mescaline and LSD were found to induce cerebral vasospasm [80]. Apparently, the pressor effect after exposure to classical psychedelics depends on the type of substance used (the strongest effect in this group was shown for LSD) and, most importantly, on the dose. High doses of psychedelics has been responsible for dangerous reactions associated with severe vasospasm in different clinical case studies of drug intoxication in a non-medical context [1, 16, 81, 82]. This effect is exacerbated by the non-serotonergic effects of psychedelics, such as interaction with monoaminergic targets (adrenergic, dopaminergic, histaminergic, MAO, monoamine transport) [25,26,27,28, 31, 32].
As mentioned above, serotonergic hyperactivity is associated with an increased risk of the development of pulmonary arterial hypertension. However, information on the effects of psychoactive substances on pulmonary arterial pressure or in animal models of pulmonary hypertension is lacking in the available literature. Given the role of serotonin in the above pathophysiological mechanisms of PAH, it seems reasonable to investigate the effects of psychedelics on pulmonary pressure in chronic use and to assess the safety of their use in patients with pulmonary hypertension and/or chronic hypoxia.
Serotonin increases platelet aggregation via 5-HT2A receptors. This raises the question of whether psychedelics increase the risk of thromboembolic complications. Hallucinogens such as LSD, DMT, and mescaline have been found to induce the human thrombocyte shape-changing response [83]. However, serious thromboembolic events associated with psychedelic use have only been described in situations of reactive drug abuse [82, 84, 85].
Experimental studies on the effects of psychedelics on heart function provided inconsistent information. In endothelin-1-induced hypertrophy and tumor necrosis factor-α (TNF-α)-induced cell injury in H9C2 cardiomyocytes, water extracts of Panaeolus cyanescens and Psilocybe cubensis did not aggravate the pathological hypertrophy induced by endothelin-1 and also protected against the TNF-α-induced injury and cell death. Specifically, psilocybin-containing mushroom extracts have been shown to reduce the size of cardiomyoblasts, increase mitochondrial activity, and reduce TNF-α and oxygen-free radical levels [86]. Recently, Gergs et al. [87] demonstrated the inotropic and chronotropic effects of LSD via the serotonin 5-HT4 and histamine H2 receptors. In this study, LSD in a dose-dependent manner increased contractility and beating rates in spontaneously beating right atrial preparations and in spontaneously beating Langendorff-perfused hearts from transgenic mice overexpressing the human 5-HT4 receptor, the H2-histamine receptor, and also in the human right atrial muscle. These effects were antagonized by H2-histamine and 5-HT4 antagonists. In another study, psilocin administration to Wistar rats by intraperitoneal injection for 12 weeks produced ECG abnormalities in the form of tachycardia, myocardial ischaemia, abnormal intraventricular conduction and mitochondrial degeneration, and myocardial purine profile changes [88]. Moreover, psilocybin intoxication induced systolic dysfunction and takotsubo cardiomyopathy, possibly due to its toxic effect on the cardiomyocytes or strong sympathomimetic effect and interaction with not only serotonergic but also adrenergic and dopaminergic receptors [89, 90]. Consistent with these studies, in an animal model and the H9C2 cell lines, the use of the phenethylamines, 2-((2-(4-Iodo-2,5-dimethoxyphenyl)ethylamino)methyl)phenol (25I-NBOH) and 2-(((2-(4-chloro-2,5-dimethoxyphenyl)ethyl)amino)methyl)phenol (25C-NBOH), dramatically reduced the viability of the H9C2 cardiomyocytes and downregulated p21 CDC42/RAC1-activated kinase 1 (PAK1). In addition, 25I-NBOH demonstrated a proarrhythmic potential as it was shown to inhibit the potassium channels in the human ether-a-go-go-related gene (hERG) assay and to significantly prolong the QT intervals and RR intervals in the rat ECG measurement [91]. The risk of the proarrhythmic effects of psilocin was also investigated in another experimental study. In tsA201 cells, psilocin reduced the hERG currents in a concentration-dependent manner. However, in this observation, a 500-fold higher concentration of psilocin than the concentration actually achieved in human plasma in clinical studies was required to produce a relevant inhibition of the hERG channel [92]. Thus, the proarrhythmic effect of psilocybin, resulting in a significant prolongation of the QT interval, appears to occur at high doses or to depend on mechanisms other than inhibiting the hERG current.
Psychedelic drugs may increase the risk of heart valve disease by binding to the 5-HT2B receptors. This risk should be considered both in terms of prolonged exposure to substance used and the affinity of the psychedelic for the 5-HT2B receptor itself. Surprisingly, neither experimental nor clinical data have comprehensively assessed this risk. The only experimental studies that have included a histological evaluation of the heart have been conducted with ayahuasca – a hallucinogenic beverage containing the beta-carbolines (harmine, harmaline, and tetrahydroharmine) and N,N-dimethyltryptamine [93, 94]. In the first study, no abnormalities were evident in a histological examination of the cardiac tissues of Wistar rats carried out 14 days after the administration of ayahuasca at a dose 50 times higher than in religious ceremonies (15.1 mg/kg DMT) [93]. A second animal study, in which ayahuasca was administered daily for 28 days in doses that exceeded those typically used in religious ceremonies, also showed that there were no histopathological changes in the heart [94]. The results of these experimental studies must be treated with some caution because ayahuasca is a mixture of DMT and beta-carbolines, which have different effects on the cardiovascular system. Indeed, in cardiovascular research, harmine has been shown to reduce systemic arterial blood pressure and peripheral vascular resistance through the inhibition of L-type calcium voltage-dependent channels. Finally, a recent study provided several lines of evidence for the anti-hypertrophic effects of harmine. Namely, in an animal model of spontaneous hypertension (SHR), harmine reduced myocardial hypertrophy. In addition, in vitro observations (of human embryonic stem cell-derived cardiomyocytes) showed that by inhibiting NF-κB phosphorylation and reducing inflammation, harmine inhibited the phenotypes of norepinephrine-induced hypertrophy and also downregulated the expression of hypertrophy-related genes [95]. There are no experimental studies on the risk of valve injury associated with the prolonged use of psilocybin and LSD. This contrasts with the other hallucinogen 3,4-methylenedioxymethamphetamine (MDMA, ‘Ecstasy’), for which prolonged mitogenic effects and induction of valvular interstitial fibroplasia through activation of the 5-HT2B receptors have been demonstrated [96]. In the absence of sufficient in vivo data, we must rely on in vitro data to assess the risk of valvular heart disease associated with medical psychedelic therapies (particularly microdosing with LSD and psilocybin). Psilocybin is a prodrug as it is almost completely metabolized to its active metabolite psilocin during absorption. Holze et al. [97] showed that the geometric mean plasma Cmax of 15 mg psilocybin was 13 ng/ml psilocin (63.5 nM). Considering psilocybin/psilocin dose proportional exposure, a typical microdoses of 1 and 2 mg of psilocin would have a Cmax of 4.2 and 8.4 nM, respectively. For LSD, pharmacokinetic studies following a 100 µg dose have reported a geometric mean Cmax of 0.279 ng/ml (0.86 nM) [98]. The concentrations of the two major metabolites of LSD [2-oxo-3-hydroxy-LSD and N-demethyl-LSD] used in in vitro studies at the 100 µg dose were very low and showed very low affinity for the 5HT-2B receptor. Therefore, in theory, the toxic effects of LSD metabolites at the LSD doses normally used for microdosing seem unlikely [99]. Pharmacokinetic data (Table 1) indicate that psilocybin/psilocin has a high potency for the 5-HT2B receptor, whereas in the case of LSD, the affinity for the 5-HT2B receptor is similar to its affinity for the 5-HT2A receptor. This suggests that psilocybin may have a relatively higher risk of valvular heart disease than LSD in microdosing therapies [100].
At this point it should also be noted that the development of experimental models (mainly in rodents) of psychedelics-associated valvulopathies and their translation to human studies is difficult, possibly because of interspecies differences in response to valvulotoxic agents or sensitivity to induction of valvular defects. Heart rate and local hemodynamic pressures during the cardiac cycle vary considerably depending on the body weight of the species. An example of this is the development of an animal model of heart valve damage caused by the drugs fenfluramine and phentermine (Fen-Phen). Despite the demonstration of serotonin receptor-mediated degenerative changes induced by Fen-Phen administration in vitro in cultured valve interstitial and endothelial cells and in humans, replication of these changes in an in vivo model has had limited success. In one study, Bratter et al. administered a continuous subcutaneous infusion of Fen-Phen to pregnant rats at a dose approximately 10 times the clinical dose for humans. As a result, large lesions of the mitral valve were observed in 25 percent of the pups [109].
The effects of psychedelics on the cardiovascular system such as the pressor and tachycardic response are also related to their sympathomimetic mechanism [110]. As mentioned earlier, psychedelics have been shown to interact with other than serotonergic monoaminergic targets, i.e., adrenergic receptors (α2A and α1A), dopaminergic receptors (D1-3), histaminergic receptors, TAARs1, monoamine transporters, and MAO inhibition.
Observations suggesting effects of psychedelics (psilocybin and LSD) on the TrkB/BDNF pathway may also be relevant to potential cardiovascular effects of psychedelics [36]. Many tissues and non-neuronal cells can synthesize BDNF. These include myocardium, endothelium and platelets [111]. In the adult heart, an impairment of the BDNF-TrkB pathway has been linked to reduced cardiac function, myocyte death, increased cardiac inflammation and oxidative stress, leading to the development of HF. [112, 113]. Through the promotion of TrkB/BDNF signaling, LSD and psilocybin may also have possible effects on cardiomyocytes and vascular endothelial function. Research into the mechanism of action of these drugs would be of interest.
Evidence-based cardiovascular risk related to psychedelic therapy
Assessing the cardiovascular safety of psychedelics in pharmacotherapy is challenging because the efficacy and potential adverse effects of these substances depend on complex factors such as the type of substance used, its dose, and, arguably, the duration of therapy. A patient’s comorbidities and drug–drug interactions must also be taken into account. There is also a need for a differentiated assessment of the potential risk of psychedelic use in relation to the potential benefits that patients may derive from the therapy. In randomized controlled trials (RCTs), observational data studies, and systematic reviews, psychedelics have been shown to produce short-term and clinically non-significant sympathomimetic effects, including increased heart rate and blood pressure. This effect was found to be dose dependent for all the classical psychedelics.
However, the remote effects of psychedelics have not been investigated in any of these studies. An interesting study on the possible long-term effects of psychedelics has recently been published. Namely, Simonnson et al. [114] showed that people who reported at least one-lifetime use of classical psychedelics (for non-medical purposes) had significantly lower odds of hypertension. It is worth noting that when the associations between hypertension in the past year and lifetime use of the main classes of classical psychedelics were analyzed, only the association with lifetime use of tryptamines was significant.
Psilocybin
An analysis of the clinical data on psilocybin suggests that it is relatively safe at the doses that have been used (Table 2). However, some published studies, especially those in which psilocybin was administered over a longer period (2–3 weeks in the microdose regimen), do not mention cardiovascular monitoring and cardiovascular effects [115,116,117,118,119]. The most commonly observed adverse effects following psilocybin were increased heart rate and blood pressure. These changes were transient, dose dependent and there was no need for medical intervention. A more serious adverse effect after psilocybin administration was a prolongation of the QTc interval [120]. In many studies, ECGs were performed only at enrolment, and electrocardiographic screening was not performed, so the magnitude of the effect of psilocybin on QT interval prolongation may be greater. Given the risk of serious life-threatening arrhythmias, electrocardiographic monitoring should be a conditio sine qua non not only in studies with psilocybin but also in studies with other psychedelics. Also, in view of the risk of valvulopathy associated with the pharmacodynamics of psilocybin (strong agonism at the 5-HT2B receptor), its chronic use in the ‘microdose’ form requires echocardiographic monitoring.
Mescaline
Data from clinical trials on the therapeutic use of mescaline are scarce. A recent study compared the immediate acute subjective, cognitive, and cardiovascular effects of 2,5-dimethoxy-4-bromophenethylamine (2C-B), a mescaline-derived hallucinogenic phenethylamine, with psilocybin in a group of 11 women [134]. The participants were given 20 mg of 2C-B, 15 mg of psilocybin, or a placebo on three separate occasions. Both compounds induced pressor effects (systolic and diastolic blood pressure). Systolic hypertension (greater than 140 mmHg) was observed in 5 cases with 2C-B and 8 cases with psilocybin. No significant differences in the heart rate of patients were observed. Recently, Ley et al. [139] compared the acute effects of mescaline (300 mg and 500 mg), LSD (100 µg), and psilocybin (20 mg) in 32 healthy subjects. Systolic blood pressure and heart rate were similarly increased by the high dose of mescaline (500 mg), LSD, and psilocybin. Notably, the increase in heart rate in response to the low dose of 300 mg mescaline exceeded the increase in heart rate in response to the high dose of 500 mg mescaline. When heart rate and blood pressure increases were combined using the rate-pressure product, the overall cardiovascular stimulation was comparable for all three substances.
N,N-dimethyltryptamine (DMT) or ayahuasca
Clinical trials using N,N-dimethyltryptamine have included DMT alone and ayahuasca, which contains DMT. In studies where cardiovascular effects were reported, administration of DMT or ayahuasca had transient, non-clinically significant sympathomimetic effects (tachycardia or an increase in blood pressure) (Table 3). When analyzing the results of these studies, it is important to remember that ayahuasca contains many other potentiating ingredients in addition to DMT. These include MAO inhibitors such as harmaline, which may affect depression (and the results of the studies) independently of DMT.
d-lysergic acid diethylamide (LSD)
Like psilocybin, LSD has shown no significant cardiovascular adverse effects in studies primarily involving healthy volunteers (Table 4). A pressor response and an increase in heart rate were the most common dose-dependent cardiovascular effects observed in these clinical trials. LSD increased blood pressure and heart rate in a dose-dependent manner. It is worth noting, however, that the cardiovascular response to LSD tends to differ. Holze et al. showed that LSD at doses of 100 and 200 µg accelerated the heart rate more than psilocybin (at doses of 15 mg and 30 mg) and that psilocybin increased blood pressure more than LSD [97]. Similar changes in the cardiovascular system after identical doses of LSD have been reported by Dodler [146]. Whereas Gasser [147] reported that LSD at a dose of 200 µg had no effect on blood pressure or heart rate. In other studies, much lower doses of LSD (below 26 µg) induced a pressor response and tachycardia [148, 149]. In another case, no changes in cardiovascular haemodynamic parameters were observed after administration [150], even in a healthy elderly population [151]. Perhaps these differences are related to the heterogeneous patterns of LSD use (Table 4) or to the study population itself (healthy volunteers with no history of reactive use of psychedelics versus those who reported a history of psychedelic use). Another explanation could be the different pharmacokinetics following the administration of LSD, as a large variability in blood concentrations of LSD has been found after microdosing [151].
As with psilocybin, many papers do not report changes in cardiovascular parameters during the use of LSD [156], or the authors simply monitored baseline BP and heart rate without assessing them after LSD administration [157].
The safety of the cardiovascular system during psychedelic therapy—what more do we need to know? What else should be done?
The physiological safety of single doses of psychedelics is now relatively well established. However, the potential risks of prolonged low-dose administration of psychedelics, particularly in relation to the cardiovascular system, require further research. Despite the growing evidence for the potential benefits of classical psychedelics in treating psychiatric disorders and the relative cardiovascular safety of psychedelic-assisted therapies, it should be remembered that, based on previous experience, prolonged pharmacological induction of serotonergic 5-HT2B receptor activation can lead to serious cardiac complications. Several drugs with relatively high 5-HT2B receptor binding affinity (Ki < 500 nM) have been unequivocally associated with valvular heart diseases, including fenfluramine and dexfenfluramine, methylergonovine, methysergide, ergotamine, pergolide, and cabergoline. Approximately 25 percent of patients developed new onset valvulopathy, including some cases of valvular thickening leading to death or requiring heart surgery [158, 159]. For this reason, it is necessary to include echocardiographic monitoring during and after prolonged use of psychedelics.
Clinical trials with psychedelics in the form of microdosing (mainly LSD and psilocybin) have not provided convincing evidence, and the potential risks may outweigh the benefits of the treatment. More convincing evidence and rigorous placebo-controlled clinical trials using representative populations are needed to establish medical indications for the chronic use of psychedelics. The largest “self-blinding” clinical trial to date, with 191 participants who received an average of 13 µg of LSD or 0.2 g of psilocybin mushrooms microdosed every 3–4 days for 4 weeks, showed equal improvement in both the placebo and psilocybin groups. This suggests that the beneficial effects of psychedelics could be explained by the placebo response [160]. In a recently published randomized clinical trial with groups assigned to placebo, 13 µg LSD, or 26 µg LSD, low doses of LSD were shown to be safe but produced negligible changes in mood or cognition in healthy volunteers [150]. It is also important to develop consistent, similar regimens for the use of microdosing therapies and to define the maximum duration of therapy.
Finally, existing clinical trials using psychedelics in a medical context have excluded patients with cardiovascular disease. Given the lack of evidence in this area, the relative safety of classical psychedelics does not apply to cardiovascular patients.
Another important issue that should be the subject of specific experimental and clinical studies is the assessment of interactions between psychedelics and their metabolites and other therapies. Given that psychedelics affect serotonergic neurotransmission, there is a potential for drug-drug interactions with medications that also modulate the serotonin system such as selective serotonin and serotonin-norepinephrine reuptake inhibitors, tricyclic antidepressants, monoamine oxidase inhibitors, atypical antipsychotics, and others. Some of these interactions may increase the risk of serotonin toxicity and even the development of symptoms of life-threatening serotonergic syndrome [161, 162]. Nevertheless, some clinical cases have shown that the chronic administration of the serotonin transporter inhibitors fluoxetine and sertraline reduced the effects of LSD and psilocybin in recreational drug users [163, 164]. This is supported by the results of a clinical study investigating whether escitalopram pretreatment for 2 weeks enhances the acute response to 25 mg of psilocybin in a group of 23 healthy volunteers. In this study, escitalopram did not have a relevant effect on the positive mood effects of psilocybin, but significantly reduced bad drug effects, anxiety, adverse cardiovascular effects and other adverse effects of psilocybin compared with placebo. In addition, the pharmacokinetics of psilocin were not altered by escitalopram [165]. Liver enzyme polymorphisms and the co-medication of patients undergoing psychedelic-assisted psychotherapy may potentially influence the pharmacology of psychedelics, and the chronic use of psychedelics may influence the activity of other drugs. Luethi et al. provided evidence for LSD metabolism in human liver microsomes (i.e., CYP2D6, 1A2, 3A4, and 2E1). Therefore, there is a possibility of drug–drug interaction with cardiovascular drugs such as statins (atorvastatin and lovastatin), warfarin, or ticagrelor [99].
Summary
The resurgence of scientific interest in psychedelic medicine has led to new insights into an ancient class of pharmacological substances used by humans for ritual, therapeutic, and cultural reasons. As this field of research evolves, there may be a need for an update of knowledge about psychedelic drugs. For patients who do not benefit from currently available treatments, continued medical research and scientific exploration of psychedelic drugs may offer new ways to treat mental illness and addiction. However, there are still many unknowns that need to be addressed to determine the safety and minimize the risk of psychedelic pharmacotherapy. Cardiovascular safety is of particular concern because of potential serotonin-related cardiotoxicity. The exact determination of the cardiovascular safety of psychedelic therapies (especially long-term therapies) requires further research.
Data availability
Data sharing is not applicable to this article as no datasets were generated or analyzed during the current study.
References
Nichols DE. Psychedelics. Pharmacol Rev. 2016;68(2):264–355. https://doi.org/10.1124/pr.115.011478.
Osmond H. A review of the clinical effects of psychotomimetic agents. Ann N Y Acad Sci. 1957;66:418-e434. https://doi.org/10.1111/j.1749-6632.1957.tb40738.x.
Rucker JJH, Iliff J, Nutt DJ. Psychiatry and the psychedelic drugs. Past, present and future. Neuropharmacology. 2018;142:200–18. https://doi.org/10.1016/j.neuropharm.2017.12.040.
Politi M, Tresca G, Menghini L, Ferrante C. Beyond the psychoactive effects of ayahuasca: cultural and pharmacological relevance of its emetic and purging properties. Planta Med. 2022;88:1275–86. https://doi.org/10.1055/a-1675-3840.
Schultes RE, Hofmann A. Plants of the Gods: Origins of Hallucinogenic Use. New York: McGraw-Hill; 1979.
Bruhn JG, De Smet PA, El-Seedi HR, Beck O. Mescaline use for 5700 years. Lancet. 2002;359:1866. https://doi.org/10.1016/S0140-6736(02)08701-9.
Hofmann A. How LSD originated. J Psychedelic Drugs. 1979;11:53–60.
Freedman DX. LSD and psychiatry: a personal trip. In: Kales A, Pierce CM, Greenblatt M, editors. The mosaic of contemporary psychiatry in perspective. New York: Springer-Verlag; 1992. p. 182–90.
Lowe H, Toyang N, Steele B, Grant J, Ali A, Gordon L, et al. W. Psychedelics: alternative and potential therapeutic options for treating mood and anxiety disorders. Molecules. 2022;27(8):2520. https://doi.org/10.3390/molecules27082520.
Belouin SJ, Henningfield JE. Psychedelics: Where we are now, why we got here, what we must do. Neuropharmacology. 2018;142:7–19. https://doi.org/10.1016/j.neuropharm.2018.02.018.
Nutt D, Carhart-Harris R. The current status of psychedelics in psychiatry. JAMA Psychiat. 2021;78(2):121–2. https://doi.org/10.1001/jamapsychiatry.2020.2171.
McClure-Begley TD, Roth BL. The promises and perils of psychedelic pharmacology for psychiatry. Nat Rev Drug Discov. 2022;21(6):463–73. https://doi.org/10.1038/s41573-022-00421-7.
Inserra A, De Gregorio D, Gobbi G. Psychedelics in psychiatry: neuroplastic, immunomodulatory, and neurotransmitter mechanisms. Pharmacol Rev. 2021;73(1):202–77. https://doi.org/10.1124/pharmrev.120.000056.
U.S. Food and Drug Administration. 2023. https://www.fda.gov/regulatory-information/search-fda-guidance-documents/psychedelic-drugs-considerations-clinical-investigations
Nutt DJ, King LA, Phillips LD. Independent Scientific Committee on Drugs. Drug harms in the UK: a multicriteria decision analysis. Lancet. 2010;376(9752):1558–65. https://doi.org/10.1016/S0140-6736(10)61462-6.
Schlag AK, Aday J, Salam I, Neill JC, Nutt DJ. Adverse effects of psychedelics: from anecdotes and misinformation to systematic science. J Psychopharmacol. 2022;36(3):258–72. https://doi.org/10.1177/02698811211069100.
Andén NE, Corrodi H, Fuxe K, Hökfelt T. Evidence for a central 5-hydroxytryptamine receptor stimulation by lysergic acid diethylamide. Br J Pharmacol. 1968;34(1):1–7. https://doi.org/10.1111/j.1476-5381.1968.tb07943.x.
Freedman DX. Effects of LSD-25 on brain serotonin. J Pharmacol Exp Ther. 1961;134:160–6.
Glennon RA, Young R, Rosecrans JA. Antagonism of the effects of the hallucinogen DOM and the purported 5-HT agonist quipazine by 5-HT2 antagonists. Eur J Pharmacol. 1983;91(2–3):189–96. https://doi.org/10.1016/0014-2999(83)90464-8.
Glennon RA, Titeler M, Young R. Structure-activity relationships and mechanism of action of hallucinogenic agents based on drug discrimination and radioligand binding studies. Psychopharmacol Bull. 1986;22:953–8.
Glennon RA. Do classical hallucinogens act as 5-HT2 agonists or antagonists? Neuropsychopharmacology. 1990;3:509–17.
González-Maeso J, Weisstaub NV, Zhou M, Chan P, Ivic L, Ang R, et al. Hallucinogens recruit specific cortical 5-HT(2A) receptor-mediated signaling pathways to affect behavior. Neuron. 2007;53(3):439–52. https://doi.org/10.1016/j.neuron.2007.01.008.
Schmid Y, Enzler F, Gasser P, Grouzmann E, Preller KH, Vollenweider FX, et al. Acute effects of lysergic acid diethylamide in healthy subjects. Biol Psychiatry. 2015;78(8):544–53. https://doi.org/10.1016/j.biopsych.2014.11.015.
Vollenweider FX, Vollenweider-Scherpenhuyzen MF, Bäbler A, Vogel H, Hell D. Psilocybin induces schizophrenia-like psychosis in humans via a serotonin-2 agonist action. NeuroReport. 1998;9(17):3897–902. https://doi.org/10.1097/00001756-199812010-00024.
Aghajanian GK, Marek GJ. Serotonin and hallucinogens. Neuropsychopharmacology. 1999;21(2 Suppl):16S-23S. https://doi.org/10.1016/S0893-133X(98)00135-3.
Mastinu A, Anyanwu M, Carone M, Abate G, Bonini SA, Peron G, et al. The bright side of psychedelics: latest advances and challenges in neuropharmacology. Int J Mol Sci. 2023;24(2):1329. https://doi.org/10.3390/ijms24021329.
Ray TS. Psychedelics and the human receptorome. PLoS ONE. 2010;5(2): e9019. https://doi.org/10.1371/journal.pone.0009019.
Marona-Lewicka D, Chemel BR, Nichols DE. Dopamine D4 receptor involvement in the discriminative stimulus effects in rats of LSD, but not the phenethylamine hallucinogen DOI. Psychopharmacology. 2009;203(2):265–77. https://doi.org/10.1007/s00213-008-1238-0.
Carhart-Harris RL, Leech R, Hellyer PJ, Shanahan M, Feilding A, Tagliazucchi E, et al. The entropic brain: a theory of conscious states informed by neuroimaging research with psychedelic drugs. Front Hum Neurosci. 2014;8:20. https://doi.org/10.3389/fnhum.2014.00020.
Vollenweider FX, Preller KH. Psychedelic drugs: neurobiology and potential for treatment of psychiatric disorders. Nat Rev Neurosci. 2020;21(11):611–24. https://doi.org/10.1038/s41583-020-0367-2.
Blough BE, Landavazo A, Decker AM, Partilla JS, Baumann MH, Rothman RB. Interaction of psychoactive tryptamines with biogenic amine transporters and serotonin receptor subtypes. Psychopharmacology. 2014;231(21):4135–44. https://doi.org/10.1007/s00213-014-3557-7.
Cozzi NV, Gopalakrishnan A, Anderson LL, Feih JT, Shulgin AT, Daley PF, et al. Dimethyltryptamine and other hallucinogenic tryptamines exhibit substrate behavior at the serotonin uptake transporter and the vesicle monoamine transporter. J Neural Transm (Vienna). 2009;116(12):1591–9. https://doi.org/10.1007/s00702-009-0308-8.
Calder AE, Hasler G. Towards an understanding of psychedelic-induced neuroplasticity. Neuropsychopharmacology. 2023;48(1):104–12. https://doi.org/10.1038/s41386-022-01389-z.
Davoudian PA, Shao LX, Kwan AC. Shared and distinct brain regions targeted for immediate early gene expression by ketamine and psilocybin. ACS Chem Neurosci. 2023;14(3):468–80. https://doi.org/10.1021/acschemneuro.2c00637.
Rijsketic DR, Casey AB, Barbosa DAN, Zhang X, Hietamies TM, Ramirez-Ovalle G, et al. UNRAVELing the synergistic effects of psilocybin and environment on brain-wide immediate early gene expression in mice. Neuropsychopharmacology. 2023. https://doi.org/10.1038/s41386-023-01613-4.
Moliner R, Girych M, Brunello CA, Kovaleva V, Biojone C, Enkavi G, et al. Psychedelics promote plasticity by directly binding to BDNF receptor TrkB. Nat Neurosci. 2023;26(6):1032–41. https://doi.org/10.1038/s41593-023-01316-5.
Janeway TC, Richardson HB, Park EA. Experiments on the vasoconstrictor action of blood serum. Arch Intern Med (Chic). 1918;XXI(5):565–603. https://doi.org/10.1001/archinte.1918.00090100002001.
Vialli M, Erspamer V. Cellule enterocromaffini e cellule basigranuloseacidofile nei Vertebrati. Z Zellforschg Mikrosk Anatomie. 1933;19:743–73.
Rapport MM, Green AA, Page IH. Serum vasoconstrictor, serotonin; isolation and characterization. J Biol Chem. 1948;176(3):1243–51.
Kaumann AJ, Levy FO. 5-hydroxytryptamine receptors in the human cardiovascular system. Pharmacol Ther. 2006;111(3):674–706. https://doi.org/10.1016/j.pharmthera.2005.12.004.
Watts SW, Morrison SF, Davis RP, Barman SM. Serotonin and blood pressure regulation. Pharmacol Rev. 2012;64(2):359–88. https://doi.org/10.1124/pr.111.004697.
Balasubramaniam G, Lee HS, Mah SC. Differences in the chronic hypotensive mechanism of action of ketanserin in spontaneously hypertensive and Wistar-Kyoto rats. J Hypertens. 1994;12(1):7–14.
Ishida T, Kawashima S, Hirata K, Yokoyama M. Nitric oxide is produced via 5-HT1B and 5-HT2B receptor activation in human coronary artery endothelial cells. Kobe J Med Sci. 1998;44(2):51–63.
Takano S, Hoshino Y, Li L, Matsuoka I, Ono T, Kimura J. Dual roles of 5-hydroxytryptamine in ischemia-reperfusion injury in isolated rat hearts. J Cardiovasc Pharmacol Ther. 2004;9(1):43–50. https://doi.org/10.1177/107424840400900i107.
Chang Chien CC, Hsin LW, Su MJ. Activation of serotonin 5-HT7 receptor induces coronary flow increase in isolated rat heart. Eur J Pharmacol. 2015;748:68–75. https://doi.org/10.1016/j.ejphar.2014.08.027.
Huzoor-Akbar, Chen NY, Fossen DV, Wallace D. Increased vascular contractile sensitivity to serotonin in spontaneously hypertensive rats is linked with increased turnover of phosphoinositide. Life Sci. 1989;45(7):577–83. https://doi.org/10.1016/0024-3205(89)90042-8.
Tsukamoto K, Kurihara T, Nakayama N, Isogai O, Ito S, Komatsu K, et al. Pressor responses to serotonin injected into the nucleus tractus solitarius of Sprague-Dawley rats and spontaneously hypertensive rats. Clin Exp Hypertens. 2000;22(1):63–73. https://doi.org/10.1081/ceh-100100062.
Russell A, Banes A, Berlin H, Fink GD, Watts SW. 5-Hydroxytryptamine(2B) receptor function is enhanced in the N(omega)-nitro-L-arginine hypertensive rat. J Pharmacol Exp Ther. 2002;303(1):179–87. https://doi.org/10.1124/jpet.102.037390.
Neumann J, Hofmann B, Dhein S, Gergs U. Cardiac roles of serotonin (5-HT) and 5-HT-receptors in health and disease. Int J Mol Sci. 2023;24(5):4765. https://doi.org/10.3390/ijms24054765.
Hervé P, Launay JM, Scrobohaci ML, Brenot F, Simonneau G, Petitpretz P, et al. Increased plasma serotonin in primary pulmonary hypertension. Am J Med. 1995;99(3):249–54. https://doi.org/10.1016/s0002-9343(99)80156-9.
Dumitrascu R, Kulcke C, Königshoff M, Kouri F, Yang X, Morrell N, et al. Terguride ameliorates monocrotaline-induced pulmonary hypertension in rats. Eur Respir J. 2011;37(5):1104–18. https://doi.org/10.1183/09031936.00126010.
Königshoff M, Dumitrascu R, Udalov S, Amarie OV, Reiter R, Grimminger F, et al. Increased expression of 5-hydroxytryptamine2A/B receptors in idiopathic pulmonary fibrosis: a rationale for therapeutic intervention. Thorax. 2010;65(11):949–55. https://doi.org/10.1136/thx.2009.134353.
Launay JM, Hervé P, Peoc’h K, Tournois C, Callebert J, Nebigil CG, et al. Function of the serotonin 5-hydroxytryptamine 2B receptor in pulmonary hypertension. Nat Med. 2002;8(10):1129–35. https://doi.org/10.1038/nm764.
Esteve JM, Launay JM, Kellermann O, Maroteaux L. Functions of serotonin in hypoxic pulmonary vascular remodeling. Cell Biochem Biophys. 2007;47(1):33–44. https://doi.org/10.1385/cbb:47:1:33.
Holmsen H. Platelet metabolism and activation. Semin Hematol. 1985;22(3):219–40.
Nakamura K, Kariyazono H, Moriyama Y, Toyohira H, Kubo H, Yotsumoto G. Effects of sarpogrelate hydrochloride on platelet aggregation, and its relation to the release of serotonin and P-selectin. Blood Coagul Fibrinolysis. 1999;10(8):513–9. https://doi.org/10.1097/00001721-199912000-00009.
Fujio M, Kuroita T, Sakai Y, Nakagawa H, Matsumoto Y. N-[1-(2-Phenylethyl)pyrrolidin-3-yl]-1-adamantanecarboxamides as novel 5-HT2 receptor antagonists. Bioorg Med Chem Lett. 2000;10(21):2457–61. https://doi.org/10.1016/s0960-894x(00)00492-3.
Kodama A, Komori K, Hattori K, Yamanouchi D, Kajikuri J, Itoh T. Sarpogrelate hydrochloride reduced intimal hyperplasia in experimental rabbit vein graft. J Vasc Surg. 2009;49(5):1272–81. https://doi.org/10.1016/j.jvs.2008.11.071.
Yamada S, Akita H, Kanazawa K, Ishida T, Hirata K, Ito K, Kawashima S, Yokoyama M. T102C polymorphism of the serotonin (5-HT) 2A receptor gene in patients with non-fatal acute myocardial infarction. Atherosclerosis. 2000;150(1):143–8. https://doi.org/10.1016/s0021-9150(99)00356-1.
Zimering MB, Razzaki T, Tsang T, Shin JJ. Inverse association between serotonin 2A receptor antagonist medication use and mortality in severe COVID-19 infection. Endocrinol Diabetes Metab J. 2020;4(4):1–5.
Ayme-Dietrich E, Lawson R, Da-Silva S, Mazzucotelli JP, Monassier L. Serotonin contribution to cardiac valve degeneration: new insights for novel therapies? Pharmacol Res. 2019;140:33–42. https://doi.org/10.1016/j.phrs.2018.09.009.
Roy A, Brand NJ, Yacoub MH. Expression of 5-hydroxytryptamine receptor subtype messenger RNA in interstitial cells from human heart valves. J Heart Valve Dis. 2000;9(2):256–60 (discussion 260-1).
Kaumann AJ. Blockade of human atrial 5-HT4 receptors by GR 113808. Br J Pharmacol. 1993;110(3):1172–4. https://doi.org/10.1111/j.1476-5381.1993.tb13937.x.
Chai W, Chan KY, de Vries R, van den Bogeardt AJ, de Maeyer JH, Schuurkes JA, et al. Inotropic effects of prokinetic agents with 5-HT(4) receptor agonist actions on human isolated myocardial trabeculae. Life Sci. 2012;90(13–14):538–44. https://doi.org/10.1016/j.lfs.2012.01.009.
Pino R, Cerbai E, Calamai G, Alajmo F, Borgioli A, Braconi L, et al. Effect of 5-HT4 receptor stimulation on the pacemaker current I(f) in human isolated atrial myocytes. Cardiovasc Res. 1998;40(3):516–22. https://doi.org/10.1016/s0008-6363(98)00198-9.
Kaumann AJ. Piglet sinoatrial 5-HT receptors resemble human atrial 5-HT4-like receptors. Naunyn Schmiedebergs Arch Pharmacol. 1990;342(5):619–22. https://doi.org/10.1007/BF00169055.
Gergs U, Neumann J, Simm A, Silber RE, Remmers FO, Läer S. Phosphorylation of phospholamban and troponin I through 5-HT4 receptors in the isolated human atrium. Naunyn Schmiedebergs Arch Pharmacol. 2009;379(4):349–59. https://doi.org/10.1007/s00210-008-0371-y.
Levy FO, Qvigstad E, Krobert KA, Skomedal T, Osnes JB. Effects of serotonin in failing cardiac ventricle: signalling mechanisms and potential therapeutic implications. Neuropharmacology. 2008;55(6):1066–71. https://doi.org/10.1016/j.neuropharm.2008.07.010.
Qvigstad E, Brattelid T, Sjaastad I, Andressen KW, Krobert KA, Birkeland JA, et al. Appearance of a ventricular 5-HT4 receptor-mediated inotropic response to serotonin in heart failure. Cardiovasc Res. 2005;65(4):869–78. https://doi.org/10.1016/j.cardiores.2004.11.017.
Ayme-Dietrich E, Aubertin-Kirch G, Maroteaux L, Monassier L. Cardiovascular remodeling and the peripheral serotonergic system. Arch Cardiovasc Dis. 2017;110(1):51–9. https://doi.org/10.1016/j.acvd.2016.08.002.
Jaffré F, Bonnin P, Callebert J, Debbabi H, Setola V, Doly S, et al. Serotonin and angiotensin receptors in cardiac fibroblasts coregulate adrenergic-dependent cardiac hypertrophy. Circ Res. 2009;104(1):113–23. https://doi.org/10.1161/CIRCRESAHA.108.180976.
Lairez O, Cognet T, Schaak S, Calise D, Guilbeau-Frugier C, Parini A, et al. Role of serotonin 5-HT2A receptors in the development of cardiac hypertrophy in response to aortic constriction in mice. J Neural Transm (Vienna). 2013;120(6):927–35. https://doi.org/10.1007/s00702-013-1011-3.
Wang Q, Zhou Y, Huang J, Huang N. Structure, function, and pharmaceutical ligands of 5-hydroxytryptamine 2B receptor. Pharmaceuticals. 2021;14(2):76. https://doi.org/10.3390/ph14020076.
Nebigil CG, Choi DS, Dierich A, Hickel P, Le Meur M, Messaddeq N, et al. Serotonin 2B receptor is required for heart development. Proc Natl Acad Sci USA. 2000;97(17):9508–13. https://doi.org/10.1073/pnas.97.17.9508.
Rajamannan NM, Caplice N, Anthikad F, Sebo TJ, Orszulak TA, Edwards WD, et al. Cell proliferation in carcinoid valve disease: a mechanism for serotonin effects. J Heart Valve Dis. 2001;10(6):827–31.
Huang XP, Setola V, Yadav PN, Allen JA, Rogan SC, Hanson BJ, et al. Parallel functional activity profiling reveals valvulopathogens are potent 5-hydroxytryptamine(2B) receptor agonists: implications for drug safety assessment. Mol Pharmacol. 2009;76(4):710–22. https://doi.org/10.1124/mol.109.058057.
Nebigil CG, Jaffré F, Messaddeq N, Hickel P, Monassier L, Launay JM, et al. Overexpression of the serotonin 5-HT2B receptor in heart leads to abnormal mitochondrial function and cardiac hypertrophy. Circulation. 2003;107(25):3223–9. https://doi.org/10.1161/01.CIR.0000074224.57016.01.
Droogmans S, Roosens B, Cosyns B, Degaillier C, Hernot S, Weytjens C, et al. Cyproheptadine prevents pergolide-induced valvulopathy in rats: an echocardiographic and histopathological study. Am J Physiol Heart Circ Physiol. 2009;296(6):H1940–8. https://doi.org/10.1152/ajpheart.01177.2008.
Dyer DC, Gant DW. Vasoconstriction produced by hallucinogens on isolated human and sheep umbilical vasculature. J Pharmacol Exp Ther. 1973;184(2):366–75.
Altura BM, Altura BT. Pharmacologic inhibition of cerebral vasospasm in ischemia, hallucinogen ingestion, and hypomagnesemia: barbiturates, calcium antagonists, and magnesium. Am J Emerg Med. 1983;1(2):180–90. https://doi.org/10.1016/0735-6757(83)90087-6.
Raval MV, Gaba RC, Brown K, Sato KT, Eskandari MK. Percutaneous transluminal angioplasty in the treatment of extensive LSD-induced lower extremity vasospasm refractory to pharmacologic therapy. J Vasc Interv Radiol. 2008;19(8):1227–30. https://doi.org/10.1016/j.jvir.2008.05.008.
Lieberman AN, Bloom W, Kishore PS, Lin JP. Carotid artery occlusion following ingestion of LSD. Stroke. 1974;5(2):213–5. https://doi.org/10.1161/01.str.5.2.213.
McClue SJ, Brazell C, Stahl SM. Hallucinogenic drugs are partial agonists of the human platelet shape change response: a physiological model of the 5-HT2 receptor. Biol Psychiatry. 1989;26(3):297–302. https://doi.org/10.1016/0006-3223(89)90042-5.
Wadowski PP, Giurgea GA, Schlager O, Luf A, Gremmel T, Hobl EL, et al. Acute limb ischemia after intake of the phenylethylamine derivate NBOMe. Int J Environ Res Public Health. 2019;16(24):5071. https://doi.org/10.3390/ijerph16245071.
Borowiak KS, Ciechanowski K, Waloszczyk P. Psilocybin mushroom (Psilocybe semilanceata) intoxication with myocardial infarction. J Toxicol Clin Toxicol. 1998;36(1–2):47–9. https://doi.org/10.3109/15563659809162584.
Nkadimeng SM, Steinmann CML, Eloff JN. Effects and safety of Psilocybe cubensis and Panaeolus cyanescens magic mushroom extracts on endothelin-1-induced hypertrophy and cell injury in cardiomyocytes. Sci Rep. 2020;10(1):22314. https://doi.org/10.1038/s41598-020-79328-5.
Gergs U, Jacob H, Braekow P, Hofmann B, Pockes S, Humphrys LJ, et al. Lysergic acid diethylamide stimulates cardiac human H2 histamine and cardiac human 5-HT4-serotonin receptors. Naunyn Schmiedebergs Arch Pharmacol. 2023. https://doi.org/10.1007/s00210-023-02591-6.
Borowiak K, Machoy-Mokrzyńska A, Majdanik S, Waloszczyk P, Piasecka M, Janus T, et al. Psilocin multiple intake resulted and in cardiotoxic effects. Acta Toxicol. 2006;14(1/2):23–30.
Kotts WJ, Gamble DT, Dawson DK, Connor D. Psilocybin-induced takotsubo cardiomyopathy. BMJ Case Rep. 2022;15(5): e245863. https://doi.org/10.1136/bcr-2021-245863.
Nef HM, Möllmann H, Hilpert P, Krause N, Troidl C, Weber M, et al. Apical regional wall motion abnormalities reminiscent to Tako-Tsubo cardiomyopathy following consumption of psychoactive fungi. Int J Cardiol. 2009;134(1):e39-41. https://doi.org/10.1016/j.ijcard.2007.12.064.
Yoon KS, Cha HJ, Choi SO, Lee JM. 2-((2-(4-Iodo-2,5-dimethoxyphenyl)ethylamino)methyl)phenol (25I-NBOH) and 2-(((2-(4-chloro-2,5-dimethoxyphenyl)ethyl)amino)methyl)phenol (25C-NBOH) induce adverse effects on the cardiovascular system. Toxicol Lett. 2022;355:160–9. https://doi.org/10.1016/j.toxlet.2021.11.016.
Hackl B, Todt H, Kubista H, Hilber K, Koenig X. Psilocybin therapy of psychiatric disorders is not hampered by hERG potassium channel-mediated cardiotoxicity. Int J Neuropsychopharmacol. 2022;25(4):280–2. https://doi.org/10.1093/ijnp/pyab085.
Pic-Taylor A, da Motta LG, de Morais JA, Junior WM, Santos Ade F, Campos LA, et al. Behavioural and neurotoxic effects of ayahuasca infusion (Banisteriopsis caapi and Psychotria viridis) in female Wistar rat. Behav Processes. 2015;118:102–10. https://doi.org/10.1016/j.beproc.2015.05.004.
Colaço CS, Alves SS, Nolli LM, Pinheiro WO, de Oliveira DGR, Santos BWL, et al. Toxicity of ayahuasca after 28 days daily exposure and effects on monoamines and brain-derived neurotrophic factor (BDNF) in brain of Wistar rats. Metab Brain Dis. 2020;35(5):739–51. https://doi.org/10.1007/s11011-020-00547-w.
Huang J, Liu Y, Chen JX, Lu XY, Zhu WJ, Qin L, et al. Harmine is an effective therapeutic small molecule for the treatment of cardiac hypertrophy. Acta Pharmacol Sin. 2022;43(1):50–63. https://doi.org/10.1038/s41401-021-00639-y.
Setola V, Hufeisen SJ, Grande-Allen KJ, Vesely I, Glennon RA, Blough B, et al. 3,4-methylenedioxymethamphetamine (MDMA, “Ecstasy”) induces fenfluramine-like proliferative actions on human cardiac valvular interstitial cells in vitro. Mol Pharmacol. 2003;63(6):1223–9. https://doi.org/10.1124/mol.63.6.1223.
Holze F, Ley L, Müller F, Becker AM, Straumann I, Vizeli P, et al. Direct comparison of the acute effects of lysergic acid diethylamide and psilocybin in a double-blind placebo-controlled study in healthy subjects. Neuropsychopharmacology. 2022;47(6):1180–7. https://doi.org/10.1038/s41386-022-01297-2.
Holze F, Vizeli P, Ley L, Müller F, Dolder P, Stocker M, et al. Acute dose-dependent effects of lysergic acid diethylamide in a double-blind placebo-controlled study in healthy subjects. Neuropsychopharmacology. 2021;46(3):537–44. https://doi.org/10.1038/s41386-020-00883-6.
Luethi D, Hoener MC, Krähenbühl S, Liechti ME, Duthaler U. Cytochrome P450 enzymes contribute to the metabolism of LSD to nor-LSD and 2-oxo-3-hydroxy-LSD: implications for clinical LSD use. Biochem Pharmacol. 2019;164:129–38. https://doi.org/10.1016/j.bcp.2019.04.013.
Tagen M, Mantuani D, van Heerden L, Holstein A, Klumpers LE, Knowles R. The risk of chronic psychedelic and MDMA microdosing for valvular heart disease. J Psychopharmacol. 2023;12:2698811231190865. https://doi.org/10.1177/02698811231190865.
Glatfelter GC, Pottie E, Partilla JS, Sherwood AM, Kaylo K, Pham DNK, et al. Structure-activity relationships for Psilocybin, baeocystin, aeruginascin, and related analogues to produce pharmacological effects in mice. ACS Pharmacol Transl Sci. 2022;5(11):1181–96. https://doi.org/10.1021/acsptsci.2c00177.
Halberstadt AL, Geyer MA. Multiple receptors contribute to the behavioral effects of indoleamine hallucinogens. Neuropharmacology. 2011;61(3):364–81. https://doi.org/10.1016/j.neuropharm.2011.01.017.
Klein AK, Chatha M, Laskowski LJ, Anderson EI, Brandt SD, Chapman SJ, et al. Investigation of the structure-activity relationships of psilocybin analogues. ACS Pharmacol Transl Sci. 2020;4(2):533–42. https://doi.org/10.1021/acsptsci.0c00176.
Rickli A, Moning OD, Hoener MC, Liechti ME. Receptor interaction profiles of novel psychoactive tryptamines compared with classic hallucinogens. Eur Neuropsychopharmacol. 2016;26(8):1327–37. https://doi.org/10.1016/j.euroneuro.2016.05.001.
Sard H, Kumaran G, Morency C, Roth BL, Toth BA, He P, et al. SAR of psilocybin analogs: discovery of a selective 5-HT 2C agonist. Bioorg Med Chem Lett. 2005;15(20):4555–9. https://doi.org/10.1016/j.bmcl.2005.06.104.
Nichols DE, Frescas S, Marona-Lewicka D, Kurrasch-Orbaugh DM. Lysergamides of isomeric 2,4-dimethylazetidines map the binding orientation of the diethylamide moiety in the potent hallucinogenic agent N, N-diethyllysergamide (LSD). J Med Chem. 2002;45(19):4344–9. https://doi.org/10.1021/jm020153s.
Porter RH, Benwell KR, Lamb H, Malcolm CS, Allen NH, Revell DF, et al. Functional characterization of agonists at recombinant human 5-HT2A, 5-HT2B and 5-HT2C receptors in CHO-K1 cells. Br J Pharmacol. 1999;128(1):13–20. https://doi.org/10.1038/sj.bjp.0702751.
Rickli A, Luethi D, Reinisch J, Buchy D, Hoener MC, Liechti ME. Receptor interaction profiles of novel N-2-methoxybenzyl (NBOMe) derivatives of 2,5-dimethoxy-substituted phenethylamines (2C drugs). Neuropharmacology. 2015;99:546–53. https://doi.org/10.1016/j.neuropharm.2015.08.034.
Bratter J, Gessner IH, Rowland NE. Effects of prenatal co-administration of phentermine and dexfenfluramine in rats. Eur J Pharmacol. 1999;369(3):R1-3. https://doi.org/10.1016/s0014-2999(99)00100-4.
Costa VM, Grando LGR, Milandri E, Nardi J, Teixeira P, Mladěnka P, et al. Natural sympathomimetic drugs: from pharmacology to toxicology. Biomolecules. 2022;12(12):1793. https://doi.org/10.3390/biom12121793.
Kermani P, Hempstead B. BDNF actions in the cardiovascular system: roles in development, adulthood and response to injury. Front Physiol. 2019;10:455. https://doi.org/10.3389/fphys.2019.00455.
Donovan MJ, Lin MI, Wiegn P, Ringstedt T, Kraemer R, Hahn R, et al. Brain derived neurotrophic factor is an endothelial cell survival factor required for intramyocardial vessel stabilization. Development. 2000;127(21):4531–40. https://doi.org/10.1242/dev.127.21.4531.
Li L, Guo H, Lai B, Liang C, Chen H, Chen Y, et al. Ablation of cardiomyocyte-derived BDNF during development causes myocardial degeneration and heart failure in the adult mouse heart. Front Cardiovasc Med. 2022;9: 967463. https://doi.org/10.3389/fcvm.2022.967463.
Simonsson O, Hendricks PS, Carhart-Harris R, Kettner H, Osika W. Association between lifetime classic psychedelic use and hypertension in the past year. Hypertension. 2021;77(5):1510–6. https://doi.org/10.1161/HYPERTENSIONAHA.120.16715.
Barnett L, Muthukumaraswamy SD, Carhart-Harris RL, Seth AK. Decreased directed functional connectivity in the psychedelic state. Neuroimage. 2020;209: 116462. https://doi.org/10.1016/j.neuroimage.2019.116462.
Cavanna F, Muller S, de la Fuente LA, Zamberlan F, Palmucci M, Janeckova L, et al. Microdosing with psilocybin mushrooms: a double-blind placebo-controlled study. Transl Psychiatry. 2022;12(1):307. https://doi.org/10.1038/s41398-022-02039-0.
Marschall J, Fejer G, Lempe P, Prochazkova L, Kuchar M, Hajkova K, et al. Psilocybin microdosing does not affect emotion-related symptoms and processing: a preregistered field and lab-based study. J Psychopharmacol. 2022;36(1):97–113. https://doi.org/10.1177/02698811211050556.
Mason NL, Kuypers KPC, Müller F, Reckweg J, Tse DHY, Toennes SW, et al. Me, myself, bye: regional alterations in glutamate and the experience of ego dissolution with psilocybin. Neuropsychopharmacology. 2020;45(12):2003–11. https://doi.org/10.1038/s41386-020-0718-8.
van Elk M, Fejer G, Lempe P, Prochazckova L, Kuchar M, Hajkova K, et al. Effects of psilocybin microdosing on awe and aesthetic experiences: a preregistered field and lab-based study. Psychopharmacology. 2022;239(6):1705–20. https://doi.org/10.1007/s00213-021-05857-0.
Goodwin GM, Aaronson ST, Alvarez O, Arden PC, Baker A, Bennett JC, et al. Single-dose psilocybin for a treatment-resistant episode of major depression. N Engl J Med. 2022;387(18):1637–48. https://doi.org/10.1056/NEJMoa2206443.
Bogenschutz MP, Forcehimes AA, Pommy JA, Wilcox CE, Barbosa PC, Strassman RJ. Psilocybin-assisted treatment for alcohol dependence: a proof-of-concept study. J Psychopharmacol. 2015;29(3):289–99. https://doi.org/10.1177/0269881114565144.
Brown RT, Nicholas CR, Cozzi NV, Gassman MC, Cooper KM, Muller D, et al. Pharmacokinetics of escalating doses of oral psilocybin in healthy adults. Clin Pharmacokinet. 2017;56(12):1543–54. https://doi.org/10.1007/s40262-017-0540-6.
Carhart-Harris RL, Bolstridge M, Rucker J, Day CM, Erritzoe D, Kaelen M, et al. Psilocybin with psychological support for treatment-resistant depression: an open-label feasibility study. Lancet Psychiatry. 2016;3(7):619–27. https://doi.org/10.1016/S2215-0366(16)30065-7.
Carhart-Harris R, Giribaldi B, Watts R, Baker-Jones M, Murphy-Beiner A, Murphy R, et al. Trial of psilocybin versus escitalopram for depression. N Engl J Med. 2021;384(15):1402–11. https://doi.org/10.1056/NEJMoa2032994.
Carbonaro TM, Johnson MW, Hurwitz E, Griffiths RR. Double-blind comparison of the two hallucinogens psilocybin and dextromethorphan: similarities and differences in subjective experiences. Psychopharmacology. 2018;235(2):521–34. https://doi.org/10.1007/s00213-017-4769-4.
Dahmane E, Hutson PR, Gobburu JVS. Exposure-response analysis to assess the concentration-QTc relationship of psilocybin/psilocin. Clin Pharmacol Drug Dev. 2021;10(1):78–85. https://doi.org/10.1002/cpdd.796.
Davis AK, Barrett FS, May DG, Cosimano MP, Sepeda ND, Johnson MW, et al. Effects of psilocybin-assisted therapy on major depressive disorder: a randomized clinical Trial. JAMA Psychiat. 2021;78(5):481–9. https://doi.org/10.1001/jamapsychiatry.2020.3285.
Goodwin GM, Aaronson ST, Alvarez O, Atli M, Bennett JC, Croal M, et al. Single-dose psilocybin for a treatment-resistant episode of major depression: Impact on patient-reported depression severity, anxiety, function, and quality of life. J Affect Disord. 2023;327:120–7. https://doi.org/10.1016/j.jad.2023.01.108.
Griffiths RR, Johnson MW, Carducci MA, Umbricht A, Richards WA, Richards BD, et al. Psilocybin produces substantial and sustained decreases in depression and anxiety in patients with life-threatening cancer: a randomized double-blind trial. J Psychopharmacol. 2016;30(12):1181–97. https://doi.org/10.1177/0269881116675513.
Grob CS, Danforth AL, Chopra GS, Hagerty M, McKay CR, Halberstadt AL, et al. Pilot study of psilocybin treatment for anxiety in patients with advanced-stage cancer. Arch Gen Psychiatry. 2011;68(1):71–8. https://doi.org/10.1001/archgenpsychiatry.2010.116.
Gukasyan N, Davis AK, Barrett FS, Cosimano MP, Sepeda ND, Johnson MW, et al. Efficacy and safety of psilocybin-assisted treatment for major depressive disorder: prospective 12-month follow-up. J Psychopharmacol. 2022;36(2):151–8. https://doi.org/10.1177/02698811211073759.
Hasler F, Grimberg U, Benz MA, Huber T, Vollenweider FX. Acute psychological and physiological effects of psilocybin in healthy humans: a double-blind, placebo-controlled dose-effect study. Psychopharmacology. 2004;172(2):145–56. https://doi.org/10.1007/s00213-003-1640-6.
Johnson MW, Garcia-Romeu A, Cosimano MP, Griffiths RR. Pilot study of the 5-HT2AR agonist psilocybin in the treatment of tobacco addiction. J Psychopharmacol. 2014;28(11):983–92. https://doi.org/10.1177/0269881114548296.
Mallaroni P, Mason NL, Reckweg JT, Paci R, Ritscher S, Toennes SW, et al. Assessment of the acute effects of 2C-B vs. psilocybin on subjective experience, mood, and cognition. Clin Pharmacol Ther. 2023;114(2):423–33. https://doi.org/10.1002/cpt.2958.
Moreno FA, Wiegand CB, Taitano EK, Delgado PL. Safety, tolerability, and efficacy of psilocybin in 9 patients with obsessive-compulsive disorder. J Clin Psychiatry. 2006;67(11):1735–40. https://doi.org/10.4088/jcp.v67n1110.
Rucker JJ, Marwood L, Ajantaival RJ, Bird C, Eriksson H, Harrison J, et al. The effects of psilocybin on cognitive and emotional functions in healthy participants: results from a phase 1, randomised, placebo-controlled trial involving simultaneous psilocybin administration and preparation. J Psychopharmacol. 2022;36(1):114–25. https://doi.org/10.1177/02698811211064720.
Schindler EAD, Sewell RA, Gottschalk CH, Luddy C, Flynn LT, Lindsey H, et al. Exploratory controlled study of the migraine-suppressing effects of psilocybin. Neurotherapeutics. 2021;18(1):534–43. https://doi.org/10.1007/s13311-020-00962-y.
Schneier FR, Feusner J, Wheaton MG, Gomez GJ, Cornejo G, Naraindas AM, et al. Pilot study of single-dose psilocybin for serotonin reuptake inhibitor-resistant body dysmorphic disorder. J Psychiatr Res. 2023;161:364–70. https://doi.org/10.1016/j.jpsychires.2023.03.031.
Ley L, Holze F, Arikci D, Becker AM, Straumann I, Klaiber A, et al. Comparative acute effects of mescaline, lysergic acid diethylamide, and psilocybin in a randomized, double-blind, placebo-controlled cross-over study in healthy participants. Neuropsychopharmacology. 2023. https://doi.org/10.1038/s41386-023-01607-2.
Dos Santos RG, Grasa E, Valle M, Ballester MR, Bouso JC, Nomdedéu JF, et al. Pharmacology of ayahuasca administered in two repeated doses. Psychopharmacology. 2012;219(4):1039–53. https://doi.org/10.1007/s00213-011-2434-x.
D’Souza DC, Syed SA, Flynn LT, Safi-Aghdam H, Cozzi NV, Ranganathan M. Exploratory study of the dose-related safety, tolerability, and efficacy of dimethyltryptamine (DMT) in healthy volunteers and major depressive disorder. Neuropsychopharmacology. 2022;47(10):1854–62. https://doi.org/10.1038/s41386-022-01344-y.
Durante Í, Dos Santos RG, Bouso JC, Hallak JE. Risk assessment of ayahuasca use in a religious context: self-reported risk factors and adverse effects. Braz J Psychiatry. 2021;43(4):362–9. https://doi.org/10.1590/1516-4446-2020-0913.
Riba J, Valle M, Urbano G, Yritia M, Morte A, Barbanoj MJ. Human pharmacology of ayahuasca: subjective and cardiovascular effects, monoamine metabolite excretion, and pharmacokinetics. J Pharmacol Exp Ther. 2003;306(1):73–83. https://doi.org/10.1124/jpet.103.049882.
Strassman RJ, Qualls CR. Dose-response study of N, N-dimethyltryptamine in humans. I. Neuroendocrine, autonomic, and cardiovascular effects. Arch Gen Psychiatry. 1994;51(2):85–97. https://doi.org/10.1001/archpsyc.1994.03950020009001.
Vogt SB, Ley L, Erne L, Straumann I, Becker AM, Klaiber A, et al. Acute effects of intravenous DMT in a randomized placebo-controlled study in healthy participants. Transl Psychiatry. 2023;13(1):172. https://doi.org/10.1038/s41398-023-02477-4.
Dolder PC, Schmid Y, Steuer AE, Kraemer T, Rentsch KM, Hammann F, et al. Pharmacokinetics and pharmacodynamics of lysergic acid diethylamide in healthy subjects. Clin Pharmacokinet. 2017;56(10):1219–30. https://doi.org/10.1007/s40262-017-0513-9.
Gasser P, Holstein D, Michel Y, Doblin R, Yazar-Klosinski B, Passie T, et al. Safety and efficacy of lysergic acid diethylamide-assisted psychotherapy for anxiety associated with life-threatening diseases. J Nerv Ment Dis. 2014;202(7):513–20. https://doi.org/10.1097/NMD.0000000000000113.
Bershad AK, Schepers ST, Bremmer MP, Lee R, de Wit H. Acute subjective and behavioral effects of microdoses of lysergic acid diethylamide in healthy human volunteers. Biol Psychiatry. 2019;86(10):792–800. https://doi.org/10.1016/j.biopsych.2019.05.019.
Bershad AK, Preller KH, Lee R, Keedy S, Wren-Jarvis J, Bremmer MP, et al. Preliminary report on the effects of a low dose of LSD on resting-state amygdala functional connectivity. Biol Psychiatry Cogn Neurosci Neuroimaging. 2020;5(4):461–7. https://doi.org/10.1016/j.bpsc.2019.12.007.
de Wit H, Molla HM, Bershad A, Bremmer M, Lee R. Repeated low doses of LSD in healthy adults: a placebo-controlled, dose-response study. Addict Biol. 2022;27(2): e13143. https://doi.org/10.1111/adb.13143.
Family N, Maillet EL, Williams LTJ, Krediet E, Carhart-Harris RL, Williams TM, et al. Safety, tolerability, pharmacokinetics, and pharmacodynamics of low dose lysergic acid diethylamide (LSD) in healthy older volunteers. Psychopharmacology. 2020;237(3):841–53. https://doi.org/10.1007/s00213-019-05417-7.
Holze F, Caluori TV, Vizeli P, Liechti ME. Safety pharmacology of acute LSD administration in healthy subjects. Psychopharmacology. 2022;239(6):1893–905. https://doi.org/10.1007/s00213-021-05978-6.
Murray CH, Tare I, Perry CM, Malina M, Lee R, de Wit H. Low doses of LSD reduce broadband oscillatory power and modulate event-related potentials in healthy adults. Psychopharmacology. 2022;239(6):1735–47. https://doi.org/10.1007/s00213-021-05991-9.
Olbrich S, Preller KH, Vollenweider FX. LSD and ketanserin and their impact on the human autonomic nervous system. Psychophysiology. 2021;58(6): e13822. https://doi.org/10.1111/psyp.13822.
Ramaekers JG, Hutten N, Mason NL, Dolder P, Theunissen EL, Holze F, et al. A low dose of lysergic acid diethylamide decreases pain perception in healthy volunteers. J Psychopharmacol. 2021;35(4):398–405. https://doi.org/10.1177/0269881120940937.
Yanakieva S, Polychroni N, Family N, Williams LTJ, Luke DP, Terhune DB. The effects of microdose LSD on time perception: a randomised, double-blind, placebo-controlled trial. Psychopharmacology. 2019;236(4):1159–70. https://doi.org/10.1007/s00213-018-5119-x.
Bedford P, Hauke DJ, Wang Z, Roth V, Nagy-Huber M, Holze F, et al. The effect of lysergic acid diethylamide (LSD) on whole-brain functional and effective connectivity. Neuropsychopharmacology. 2023;48(8):1175–83. https://doi.org/10.1038/s41386-023-01574-8.
Connolly HM, Crary JL, McGoon MD, Hensrud DD, Edwards BS, Edwards WD, et al. Valvular heart disease associated with fenfluramine-phentermine. N Engl J Med. 1997;337(9):581–8. https://doi.org/10.1056/NEJM199708283370901.
Brenot F, Herve P, Petitpretz P, Parent F, Duroux P, Simonneau G. Primary pulmonary hypertension and fenfluramine use. Br Heart J. 1993;70(6):537–41. https://doi.org/10.1136/hrt.70.6.537.
Szigeti B, Kartner L, Blemings A, Rosas F, Feilding A, Nutt DJ, et al. Self-blinding citizen science to explore psychedelic microdosing. Elife. 2021;10: e62878. https://doi.org/10.7554/eLife.62878.
Sarparast A, Thomas K, Malcolm B, Stauffer CS. Drug-drug interactions between psychiatric medications and MDMA or psilocybin: a systematic review. Psychopharmacology. 2022;239(6):1945–76. https://doi.org/10.1007/s00213-022-06083-y.
Malcolm B, Thomas K. Serotonin toxicity of serotonergic psychedelics. Psychopharmacology. 2022;239(6):1881–91. https://doi.org/10.1007/s00213-021-05876-x.
Strassman RJ. Human hallucinogen interactions with drugs affecting serotonergic neurotransmission. Neuropsychopharmacology. 1992;7(3):241–3.
Bonson KR, Buckholtz JW, Murphy DL. Chronic administration of serotonergic antidepressants attenuates the subjective effects of LSD in humans. Neuropsychopharmacology. 1996;14(6):425–36. https://doi.org/10.1016/0893-133X(95)00145-4.
Becker AM, Holze F, Grandinetti T, Klaiber A, Toedtli VE, Kolaczynska KE, et al. Acute effects of psilocybin after escitalopram or placebo pretreatment in a randomized, double-blind, placebo-controlled, crossover study in healthy subjects. Clin Pharmacol Ther. 2022;111(4):886–95. https://doi.org/10.1002/cpt.2487.
Acknowledgements
I would like to express my gratitude to Mr. Marcin Kumosa for the preparation of the illustrations.
Funding
This review did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.
Author information
Authors and Affiliations
Contributions
AW: concept, design, literature review, preparing and revising the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The author declares no conflict of interest.
Additional information
Publisher’s Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Wsół, A. Cardiovascular safety of psychedelic medicine: current status and future directions. Pharmacol. Rep 75, 1362–1380 (2023). https://doi.org/10.1007/s43440-023-00539-4
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s43440-023-00539-4