
Abstract
Inflammation and glutamate (GLU) are widely thought to participate in the pathogenesis of depression, and current evidence suggests that the development of depression is associated with the activation of the kynurenine pathway (KP). However, the exact mechanism of KP among the inflammation, GLU and depression remain poorly understood. In this study, we examined the involvement of KP, inflammation and GLU in depressive phenotype induced by chronic unpredictable mild stress (CUMS) in C57B/6 J mice. Our results showed that CUMS caused depressive like-behavior in the sucrose preference test, tail suspension test and forced swimming test. From a molecular perspective, CUMS upregulated the peripheral and central inflammatory response and activated indoleamine 2,3-dioxygenase (IDO), the rate-limiting enzyme of KP, which converts tryptophan (TRP) into kynurenine (KYN). KYN is a precursor for QA in microglia, which could activate the N-methyl-D-aspartate receptor (NMDAR), increasing the GLU release, mirrored by increased IDO activity, quinolinic acid and GLU levels in the hippocampus, prefrontal cortex and serum. However, intervention with IDO inhibitor 1-methyl-DL-tryptophan (50 mg/kg/s.c.) and 1-methyl-L-tryptophan (15 mg/kg/i.p.) reversed the depressive-like behaviors and adjusted central and peripheral KP’s metabolisms levels as well as GLU content, but the inflammation levels were not completely affected. These results provide certain evidence that KP may be a vital pathway mediated by IDO linking inflammation and glutamate, contributing to depression.
Similar content being viewed by others

Introduction
Depression is a recurrent mental disorder with a high prevalence in current times [1,2,3]. As of January 2020, the depressive disorder has developed into a disease that impacts over 264 million people of all ages worldwide [4]. In brief, depression is characterized by low mood, cognitive deficits, anhedonia, and even a high suicidal tendency [5, 6]. Over the years, numerous causes of depression have been identified, involving hypothalamic–pituitary–adrenal axis disorders, monoamine, neurotrophic factors, oxidative stress, cytokines, oxidative stress, neurotransmitter receptors, inflammation, and so on [7, 8]. Lately, the kynurenine pathway has been reported to play a key role. At present, the pharmacological treatment of major depression is often suboptimal and associated with substantial side effects [9,10,11,12]. Therefore, it is essential to identify the exact mechanisms of depression.
An increasing body of evidence suggests that inflammation is tightly related to stress and depression [13,14,15], depression and inflammation fuel one another [16]. Patients with depression have increased pro-inflammatory cytokines in the blood, such as interleukin-6 (IL-6), interleukin-1β (IL-1β), tumor necrosis factor-α (TNF-α) and other acute-phase proteins and C-reactive protein (CRP) [17, 18]. Moreover, accumulating evidence suggests that chronic stress can lead to exaggerated or prolonged inflammatory responses, resulting in a series of sickness behaviors (such as pain and disturbed sleep) and depressive symptoms (lack of pleasure) and act as mediating pathways prompting further severe inflammation and depression.
In addition, the levels of brain major excitatory and inhibitory neurotransmitters are related to the variation in brain connectivity in major depression disorder (MDD) [19], and glutamate (GLU), has an essential role in the central nervous system as the predominant excitatory neurotransmitter [20]. In neuroimaging and post-mortem studies, the glutamate levels of most depressed patients’ were elevated in the plasma, cerebrospinal fluid and brain [21, 22]. Therefore, dysregulation of glutamatergic neurotransmission is related to depressive-like behaviors [23, 24]. Moreover, our previous studies confirmed that increased glutamate concentration in the hippocampus (HIP) of depressed rats leads to neurotoxicity and apoptosis of neurons and astrocytes [25], and chronic stress leads to structural and morphological alternations such as dendritic spine loss, dendritic atrophy, and volume loss in the HIP and prefrontal cortex (PFC) [26, 27]
What's more, the kynurenine pathway (KP) plays an essential role in tryptophan degradation mediated by indoleamine 2,3-dioxygenase (IDO), an initial and rate-limiting enzyme. Given that the terminal metabolites of KP all act on N-methyl-D-aspartate (NMDA) receptors, the dysregulated activity of quinolinic acid (QA) and kynurenic acid (KYNA) on NMDA receptors and the hyperactivity of the glutamate system may be the critical point of KP’s involvement in depression. When activated, KP can induce the metabolism of TRP to shift towards kynurenine (KYN) and eventually degrades to QA and KYNA, impacting GLU neurotransmission at NMDA glutamate receptors, the former as an agonist at the glutamate binding site of the NMDA receptor, leading to excess GLU release which is increasingly implicated in neurodegenerative disorders, while the latter blocks the glycine-site of NMDA, which is a standard for the identification of glutamate-releasing synapses [28,29,30], endowing with neuroprotective agents [31,32,33,34].
KP is easily activated during inflammation, which is among the main pathological characteristics of depression and alters glutamate metabolism. Interestingly, IDO has also emerged as a potential target for depression treatment. However, the mechanism of KP linking glutamate and inflammatory factors to depression remains obscure and requires further investigation. Therefore, we are focusing on the questions of ‘how’ (pathways) and ‘for whom’ (predispositions) these links exist.
Materials and methods
Animals
Male C57BL/6 J mice (8–10 weeks; 20–25 g) with the approval number 20210315–15 were housed in the Laboratory Animal Center with access to food and water provided ad libitum. Standard laboratory conditions (12 h light: 12 h dark cycle, lights on at 08:00 pm, T = 21 ± 1 °C) were maintained in animal housing facilities. At the end of the tests, serum was collected after overnight fasting via eyeball blood while mice were anesthetized with Isoflurane. Data collected were subjected to statistical analysis with at least 5 mice per group.
Experimental design in chronic unpredictable mild stress (CUMS) procedure
After 1 week of adaptive feeding, mice were assigned to four groups, including the Control + PBS group, CUMS + PBS group, CUMS + DL-1MT group and CUMS + L-1MT group. Unstressed mice were group-housed in standard laboratory cages while the other groups were housed in individual cages, the latter were subjected to unpredictable mild stress protocol for 3 weeks. Two different stressors and sequences were applied each day. The protocol consisted of fourteen stressors as shown in Fig. 1B. After 3 weeks of the CUMS exposure, the behavioral approaches were performed to whether the model was successfully established. The body weights of the mice were measured weekly before and after the CUMS. The timeline of the experimental protocol is depicted in Fig. 1A.

Experimental design. A Experimental timeline for the animals in the study. B The detailed protocol in the process of establishing a mice model of depression. The animals were habituated for 7 days and subjected to chronic unpredictable mild stress or administered with 1-MT for 21 days. Inhibitors were intraperitoneally or subcutaneously injected, and the behaviors were tested. Subsequently, brain tissue and serum were collected
Drugs
-
1.
Control + PBS and CUMS + PBS group received phosphate buffer solution (PBS, pH 7.4, without Ca2+ and Mg,2+) daily.
-
2.
The IDO inhibitor, 1-methyl-D, L-tryptophan (DL-1-MT, Sigma-Aldrich, USA) was subcutaneously injected with a 5 ml/kg volume to deliver a dose of 50 mg/kg. The injections were administered twice daily with a 12-h interval between two administrations, and the effect is equivalent to the studies using 5 mg/day pellets [32]. The final pH was adjusted to 9.0 using 0.1 M NaOH.
-
3.
The IDO inhibitor, 1-methyl-L-tryptophan (L-1-MT, Sigma-Aldrich, USA) was intraperitoneally injected for 21 d at 15 mg/kg/d. The drug was dissolved in 0.1 M sodium hydroxide and the pH was adjusted to 9.0 using hydrochloric acid in a volume ratio of 1:1 before administration [35].
Behavioral analyses
Body weight measurement
Each mouse’s body weight was evaluated with an electronic balance at 9:00 am every Thursday.
Sucrose preference test (SPT)
The SPT was divided into training (as a baseline measurement) and testing periods. In brief, single-housed animals were trained with 1% sucrose solution for 24 h. Then after 24 h of food and water deprivation, all mice were provided with a bottle of pure water and another bottle of 1% sucrose solution simultaneously. Two hours later, the volumes of the remaining pure water and sucrose solution were recorded. It is defined as follows: sucrose preference percentage (%) = sucrose solution consumption (g)/(sucrose solution consumption [g] + water consumption [g]) × 100%.
Open-field test (OFT)
During the open filed test, mice were transported to the test room at least 2 h before the experiment for habituation. The room was kept quiet and the mice were placed in the center and allowed to move freely. The mice's behaviors were recorded for five minutes using a video camera mounted above the maze, which was analyzed using ANYmaze software (Stoelting). The time spent in the central area and total distance traveled were recorded automatically. After each session, the arena walls and floors were thoroughly wiped down with 75% ethanol to eliminate odor cues.
Tail-suspension test (TST)
During the tail-suspension test, mice were isolated and suspended by the tail taped on a stand at the edge of the tabletop 35 cm above [36]. The 6 min test was recorded and time spent immobile was measured during the last 4 min of the test.
Forced swimming test (FST)
During the forced swimming test, mice were placed in a transparent cylinder (30 cm height × 16 cm diameter, 14 cm of water depth, 24 °C) and they could neither touch the bottom nor climb out the top. Mice were forced to swim for 6 min. Consistent with TST, the animals were adapted for the first 2 min and the time spent immobile was recorded over the next 4 min.
Quantitative real-time PCR analysis
Twenty-four hours after the final behavioral test, mice were sacrificed by rapid decapitation. Total RNA was extracted from brain tissue using TRIzol reagent (Invitrogen, USA), according to the manufacturer's instructions, cDNA Synthesis was performed the Prime Script First Strand Kit (Takara Biotechnology). The cDNA was amplified by PCR using standard methods. The following specific primers were used (see Table 1).
Enzyme-linked immunosorbent assay (ELISA)
Twenty-four hours after the last behavioral test, mice were sacrificed by decapitation. Brain tissue and serum were dissected. After naturally clotting for more than 20 min, the blood was centrifuged for 20 min (3000 rpm) and the supernatant was collected in the following experiment. Weights were taken of prefrontal cortex and hippocampal tissue, which was placed in a PBS solution (1 mL/10 mg) and homogenized rapidly. The samples of tissue and serum were centrifuged (5000 × g for 5 min), then the supernatant was collected and quantified. On the microplate reader, absorbance was quickly determined at 450 nm. And according to the standard curve, the levels of inflammatory cytokines IL-1β(JL18442), IL-6(JL20268) and TNF-α(JL10484), and KP enzyme IDO1(JL47052), IDO2(JL33530) concentrations were calculated by commercial ELISA kits (Jianglai, Shanghai, China) according to the manufacturer's protocol.
High-performance liquid chromatography analysis (HPLC)
To understand the impact of stress on KP and its metabolic enzymes, the brain tissue and serum concentrations of tryptophan (TRP) and its IDO-catalyzed metabolite KYN、KYNA、QA were measured by HPLC and individual metabolite peaks detected were collected as HPLC fractions. The analysis was performed on an HPLC-1100 system (Agilent Technologies, USA) equipped with a quaternary pump and a UV detector for TRP and KYN, a fluorescence detection was used for measure KYNA and QA. HPLC analysis of the samples was performed using an Agilent C18 column (5 μm particle size, L × I.D. 25 cm × 4.6 mm) preceded by a C18 guard column (Agilent Technologies, USA). For TRP and KYN, the mobile phase was 15 mM acetic acid-sodium acetate buffer (pH 5.3) containing 6% acetonitrile by volume [37]. For KYNA and QA, Water and methanol with acetonitrile formate (0.1%) and formic acid (0.1%v/v) were used as mobile phase solvents [38].
Statistical analysis
Statistical analyses were performed using GraphPad Prism 8 (GraphPad Software, San Diego, CA, USA). Moreover, our sample sizes were similar to those reported in previous publications. The data were expressed as the mean ± SEM and compared by a two-sided Student’s-t-test or one-way analysis of variance (ANOVA) followed by Bonferroni tests. Body weight and changes were analyzed via a repeated measures two-factor ANOVA. A p-value < 0.05 was statistically significant.
Results
CUMS induced depressive-like behavior
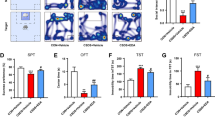
Mice underwent 3 weeks of CUMS exposure, then were weighed and tested for sickness or depression-like behavior before being euthanized to collect brain samples and serum. The CUMS-induced sickness was measured on the first day of week one by assessing body weight loss. (Fig. 2A) and one day after the final stress session by assessing the sucrose solution consumption (Fig. 2B). To investigate CUMS-induced depressive-like behaviors, all mice underwent behavioral tests including OFT, TST, and FST (Fig. 2C–F). As expected, there was a significant difference in body weight change after CUMS (p < 0.0001). Similarly, the sucrose consumption of CUMS-exposed mice was significantly reduced (p < 0.0001), which reflected anhedonia (Fig. 2B). After CUMS exposure, mice spent less time on the center zone than controls during the OFT test (p < 0.05) (Fig. 2C), however, the distance traveled in the open field was comparable among the groups, which indicated that mice's motor function was not impaired (p > 0.05) (Fig. 2D). The mice exposed to CUMS exhibited increased time spent immobile in TST (p < 0.01) (Fig. 2D) and FST (p < 0.0001) (Fig. 2E) compared to the control mice. These results indicate that mice exposed to CUMS displayed significant depressive-like behaviors,

C57BL/6 J mice exhibited alterations in depression-like behaviors. A Results of the body weight of mice after a 3-week exposure to CUMS (mean ± SEM, n = 6- 8). B Results of sucrose preference of mice after a 3-week exposure to CUMS. C Results of central time in OFT test of mice after a 3-week exposure to CUMS. D Results of total distance in OFT test of mice after a 3-week exposure to CUMS. E Results of tail suspension test of mice after a 3-week exposure to CUMS. F Results of forced swimming test of mice after a 3-week exposure to CUMS. Data are presented as mean ± SEM. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001 vs Control + PBS, #p < 0.05, ##p < 0.01, ###p < 0.001, ####p < 0.0001 vs CUMS + PBS
CUMS induces central and peripheral cytokine expression
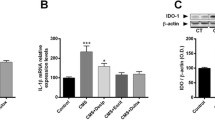
To validate that our model was successful and verify the relationship between central and peripheral cytokines and depression, we measured the mRNA expression and protein level of pro-inflammatory cytokines, including IL-6, IL-1β and TNF-α. As shown in Fig. 3A–C, CUMS increased the expression of IL-6 (p < 0.05), IL-1β (p < 0.05) and TNF-α (p < 0.001) mRNA compared with the Control + PBS group. Meanwhile, our data showed that stress stimuli dramatically increased the levels of IL-6 (p < 0.01), IL-1β (p < 0.0001) and TNF-α (p < 0.01) (Fig. 3D–I) in the brain and serum.

CUMS mice activate cytokines. A–I Results of the IL-6, IL-1β and TNF-α of mice after a 3-week exposure to CUMS through qPCR and ELISA. Data are presented as mean ± SEM, n = 5–8 per group. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001 vs Control + PBS, #p < 0.05, ##p < 0.01, ###p < 0.001, ####p < 0.0001 vs CUMS + PBS
CUMS increased the release of GLU in mouse serum and brain
GLU, is a major excitatory neurotransmitter, that plays a vital role in the central nervous system [20]. However, the changes in GLU levels in depressed mice serum and brain remain poorly understood. We observed central and peripheral GLU levels by HPLC and found that GLU was a significantly elevated in the serum of CUMS mice model (Fig. 5J) (p < 0.01). In contrast, GLU was elevated in brain tissues in the model group but did not differ from control mice (p > 0.05) (Fig. 5D).
CUMS activated IDO and mediated KP and its metabolism
Given that IDO is the first rate-limiting and inflammatory-inducing enzyme in the KP [39], we explored the molecular mechanisms underlying stress-induced depression. Accordingly, we measured two IDO isoforms, IDO1and IDO2, KP and its metabolites in the serum and brain tissue of mice harvested at the end of behavioral experiments by q-PCR, ELISA and HPLC. We found that brain (Fig. 4A, B, D, E) and serum (Fig. 4C, F) expression of IDO1 and IDO2 were significantly increased in CUMS induced-mice (p < 0.01), indicating that CUMS-exposure elevated IDO expression in mice.

CUMS mice activate IDO and cytokines. A–F Results of the IDO1 and IDO2 of mice after a 3-week exposure to CUMS through qPCR and ELISA. Data are presented as mean ± SEM, n = 5–8 per group. *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001 vs Control + PBS, #p < 0.05, ##p < 0.01, ###p < 0.001, ####p < 0.0001 vs CUMS + PBS
In addition to IDO, other downstream enzymes in the KP—including TRP, KYN, KYNA and QA, were analyzed in the CUMS group. As expected, KYN/TRP was an indicator of IDO activity [40], we found the ratio of KYN to TRP was increased in the brain and serum (Fig. 5A, E) (p < 0.01), suggesting it has a significant value as a surrogate marker of IDO activation [32, 41]. Moreover, the level of KYN was increased (p < 0.05) (Fig. 5B) while QA exhibited no significant change (p > 0.05) (Fig. 5C) during HPLC. However, the KP exhibited significant changes in the peripheral region, the level of TRP and KYNA was decreased (p < 0.001) (Fig. 5F, G) while QA was elevated (p < 0.0001) (Fig. 5I). Of note, there was a difference observed for KYNA/QA ratio (Fig. 5H) (p < 0.001), a measure of NMDA agonist/antagonist balance. These findings established that IDO could be activated by stress and then mediated the change in KP. To better verify these results, we also performed the correlation analysis between the level of QA and GLU, QA and cytokines respectively (see Additional file 1: Fig. S1, Table S1).

The changes in KP and its metabolites under the stress. A–D The level of KYN/TRP (A), KYN (B), QA (C) and GLU (D) in the brain were measured via HPLC. E–I The results of KYN/TRP (E), TRP (F), QA (G), KYNA/QA (H), KYNA (I) and GLU (J) in the serum. Data are expressed as the mean ± SEM. ** p < 0.01, ****p < 0.0001 vs. Control group; #p < 0.05, ##p < 0.01, ###p < 0.001, ####p < 0.0001 vs model
1-MT specifically inhibits IDO and depressive-like behavior
To directly target IDO for in vivo experiments, mice were injected with DL-1-MT (50 mg/kg) or L-1-MT (15 mg/kg) for 3 weeks. First, we verified the inhibitory effect on IDO1 and IDO2 by q-PCR, as shown in Fig. 4A and D, respectively, 1-MT specifically inhibited the mRNA expression and protein level of IDO1 (p < 0.001) and IDO2 (p < 0.01); and through HPLC, we found that L-1-MT attenuated the increased ratio of kynurenine to tryptophan in the brain (Fig. 5A) (p < 0.05) and peripheral blood (Fig. 5E) (p < 0.001), however, DL-1-MT only affected the peripheral ratio value (Fig. 5E) (p < 0.01), and decreased the level of TRP in the serum (Fig. 5F) (p < 0.05). What’s more, the ratio of KYNA/QA as well as the concentration of QA (Fig. 5H, I) could only be reversed by two inhibitors, suggesting the KP was activated strongly in the peripheral blood and propelled kynurenine transport from the blood to the brain, which agreement with the Robert Danzer’s results [42].
Depressive-like behaviors were also measured post-1-MT injection. In contrast with mice treated with PBS, the 1-MT intervention did not affect the CUMS-induced reduction in body weight (Fig. 2A) (p > 0.05). However, injection with DL-1-MT (p < 0.0001) or L-1-MT (p < 0.001) significantly reversed the decrease in sucrose preference percentage induced by CUMS (Fig. 2B), increased the time spent in the center area during the open field test (Fig. 2D) (p < 0.01), and reduced the time spent during TST (p < 0.01) and FST (p < 0.001) compared with the CUMS group (Fig. 2E, F). Overall, these findings indicated that IDO inhibitors could ameliorate depressive-like behavior (Additional file 2, 3, 4).
1-MT blocks peripheral CUMS-induced cytokine and GLU content
Then, to determine whether DL-1-MT or L-1-MT yielded an anti-inflammatory effect, IL-6, IL-1β and TNF-α mRNA expression (Fig. 3A–C) and protein levels (Fig. 3D–I) were measured in the brains and serum in mice exposed to CUMS. Our results found that 1-MT did not modify the levels and mRNA expression of cytokines; however, as shown in Fig. 3GH, L-1-MT inhibited the protein level of IL-6 (p < 0.01) and IL-1β (p < 0.05), suggesting L-1-MT had a definite anti-inflammatory effect.
We also assessed the function of inhibitors on GLU and found that DL-1-MT or L-1-MT significantly decreased GLU levels in the serum (Fig. 5J) (p < 0.001) than in brain tissues (Fig. 5D) (p > 0.05), showing the inhibitors had a more significant effect on peripheral GLU levels.
Discussion
Two evolving theories about the development of mood disorders involve hyperactivation of inflammatory cytokines and altered glutamate metabolism [43,44,45]. Stress can cause depression, perhaps partly because it propels inflammation and then alters glutamate metabolism [46,47,48], and KYN metabolism has been identified as a critical neurochemical pathway linking inflammation and depression [49], which also regulates the release of glutamine. Therefore, in our study, we established a CUMS mice model and, observed alleviation of depression via changes in the behavioral test, glutamate and proinflammatory factor levels; we explored whether associations between inflammation and glutamine in depression were associated with the kynurenine pathway.
It is well-established that inflammation is linked to depression. An increasing body of evidence from recently published, human studies has associated increased peripheral and central cytokines production with the development of mood disorders [50, 51]. This increased activation of inflammatory, in turn, contributes to the glutamatergic system’s pathological activation, which leads to excitotoxicity and decreased neurotrophic support in the CNS. In fact, the dysfunction of the immune, monoaminergic, and glutamatergic systems is implicated in the pathology of depression. KP is a potential intersection of these systems, where immune responses and serotonergic neurotransmission are catabolized by TRP to KYN, ultimately altering downstream synaptic glutamate neurotransmission [52]. Enhanced pro-inflammatory cytokine levels may over-activate the KP, leading to tryptophan depletion and reduced serotonin levels, which can subsequently precipitate depressive symptoms, however, only a small fraction of the TRP pool is converted to serotonin, the vast majority (~ 95%) is metabolized via KP [53].
Apart from depression and inflammation, altered glutamate metabolism also implicates mood disorders [54,55,56]. Dysregulation of glutamate release is one of the major triggers of depression and is involved in the mechanism of antidepressant effects [25, 43, 57] which is the most abundant amino acid in the blood and cerebrospinal fluid and is the precursor of major central nervous system stimulants [20]. QA was an excitotoxic kynurenine pathway metabolite of tryptophan, enhancing glutamate release and inhibiting glutamate uptake. In our research, increased levels of pro-inflammatory cytokines and GLU in peripheral and central regions made us focus attention on the relationship of inflammation and glutamate release to explore the mechanisms underlying depression-like behavior induced by CUMS. So we did a correlation analysis to better illustrate the relationship between QA and glutamate and inflammatory factors, and found QA had a strong positive correlation with GLU, while hardly related with IL-6, IL-1β and TNF-α in the brain. Meanwhile, we observed that the QA had a strong linear correlation with GLU, and a moderate positive correlation with IL-6 and IL-1β, a weak correlation with TNF-α in the serum, suggesting QA may act as a bridge between the peripheral cytokines and glutamate.
It is well-recognized that the relationship between inflammation and depression [32] is mediated by IDO activation which is an enzyme involved in kynurenine synthesis from tryptophan [58]. Importantly, the ratio of KYN/TRP is a proxy for IDO enzymatic activity [59]. Two isomers of IDO, IDO1 and IDO2, have been reported to participate in TRP conversion into KYN and other downstream metabolites [60, 61]. Nonetheless, the roles of IDO1 and IDO2 in this mechanism remain obscure [60], especially IDO2. Since DL-1-MT, a mixture of two isomers including L-1-MT and D-1-MT, could inhibit both IDO1 and IDO2. And L-1-MT is a more potent inhibitor of IDO1. We wanted to test which isomer could exert its observed effect on depression via this route. And we found that IDO1 and IDO2 activity were elevated both peripherally and centrally during the stress and changed the depressive-like behaviors. However, L-1-MT, a competitive inhibitor of IDO1, significantly lower the ratio of KYN/TRP, reversed the level of IL-6 and IL-1β in serum, possessing anti-inflammatory effects, which was consistent with the literature [62, 63]. It has been established that IDO2 plays a cytokine suppressor role while IDO1 is a pro-inflammatory one [63,64,65], and L-1-MT can block IDO1. In our study, L-1-MT suppressed the levels of IL-6 and IL-1β. Besides, DL-1-MT, a racemic compound inhibitor of IDO1 and IDO2 [62, 66, 67], yielded no significant effect on the proinflammatory cytokines.
In addition to IDO, KP and its metabolites have been reported to link inflammation and depression through effects on brain glutamate receptors. Increased glutamate in inflammatory conditions results from the effect of inflammatory cytokines on IDO [68, 69]. The KP is an alternative but distinct pathway to the generation of glutamate receptor ligands. The products of tryptophan metabolism via the kynurenine pathway include both quinolinic acid and kynurenic acid. As an agonist of NMDA receptors, QA leads to increased potential neurotoxic metabolites and releases large amounts of glutamate [70], and KYNA, a potentially neuroprotective compound [71], that aggravates depression at low levels [72, 73]. In addition to affecting excitatory neurotransmission by acting directly on glutamate receptors, QA and KYNA indirectly modulate glutamate function. For example, QA-induced stimulation of NMDA receptors in the rat cerebral cortex results in a substantial increase in glutamate release.
Previous studies have associated KP metabolites with the severity of depression including feelings of hopelessness and lack of motivation [49, 74]. Our data support these findings and suggest that the combined activation of inflammation and KP in the periphery may play an important role in depression, inflammation and GLU. Moreover, our data illustrated the specificity of IDO inhibitors, DL-1-MT and L-1-MT, which could reverse the depressive behaviors and may partly contribute to the release of inflammatory cytokines and GLU. In conclusion, this study describes a putative pathway that drives depression involving inflammatory mediators, GLU and KP metabolites. At last, there are also some limitations to this study. Firstly, in the behavioral experiment we did not include a placebo group as the objective was not to assess once more the effects of 1-MT but just to confirm that 1-MT did block the IDO-induced depression-like behaviors. Secondly, we only used male mice to verify our results and did not study female mice to examine the effect of KP on sex differences, which need further research. Thirdly, we ignore the tryptophan 2,3-dioxygenase (TDO), an enzyme with a similar function to IDO, which is capable of inducing stress-induced depression in the rat and driving the metabolism of tryptophan through the KP [38]. Despite these limitations, the conclusion still can be made that the kynurenine pathway involved the underlying etiopathology of CUMS between inflammation and glutamate.
Conclusion
A summary of our findings on the mechanisms of the kynurenine pathway linking inflammation and GLU in depression is provided in Fig. 6. These results corroborate that KP mechanisms may be used to prevent and treat CUMS-induced depression-like symptoms.

The directional associations of serum and brain kynurenine pathway (KP) among CUMS, inflammation, glutamate. The broken arrows represent indirect effects and the filled arrows represent direct effects, whereas truncated lines represent an inhibitory action or effect. CUMS = chronic mild unpredictable stress; KP = kynurenine pathway; 1-MT = 1-methyl-DL/L-tryptophan
Data availability statement
The data used and/or analyzed during the current study are available from the corresponding author on reasonable request.
References
Wigner P, Synowiec E, Jóźwiak P, et al. The changes of expression and methylation of genes involved in oxidative stress in course of chronic mild stress and antidepressant therapy with agomelatine. Genes (Basel). 2020;11(6):644.
Qiu X, Wu G, Wang L, et al. Lactobacillus delbrueckii alleviates depression-like behavior through inhibiting toll-like receptor 4 (TLR4) signaling in mice. Ann Transl Med. 2021;9(5):366.
Wang X, Cui S, Wu MS, et al. Victim sensitivity and its neural correlates among patients with major depressive disorder. Front Psychiatry. 2020;11:622.
Martínez-Castaño R, Pichel JC, Losada DE. A big data platform for real time analysis of signs of depression in social media. Int J Environ Res Public Health. 2020;17(13):4752.
Kessler RC, Bromet EJ. The epidemiology of depression across cultures. Annu Rev Public Health. 2013;34:119–38.
Swinkels JA. Revised Dutch GP guideline “Depression”: diagnosis not as simple as it appears. Ned Tijdschr Geneeskd. 2012;156(38):A5113.
Tian Q, Chen L, Luo B, et al. Hydrogen sulfide antagonizes chronic restraint stress-induced depressive-like behaviors via upregulation of adiponectin. Front Psychiatry. 2018;9:399.
Lang UE, Borgwardt S. Molecular mechanisms of depression: perspectives on new treatment strategies. Cell Physiol Biochem. 2013;31(6):761–77.
Srikumar BN, Paschapur M, Kalidindi N, et al. Characterization of the adrenocorticotrophic hormone - induced mouse model of resistance to antidepressant drug treatment. Pharmacol Biochem Behav. 2017;161:53–61.
Ceylan D, Erer S, Zarifoğlu M, et al. Evaluation of anxiety and depression scales and quality of LIFE in cervical dystonia patients on botulinum toxin therapy and their relatives. Neurol Sci. 2019;40(4):725–31.
Weber A, Heger S, Sinkgraven R, et al. Psychosocial aspects of patients with focal hyperhidrosis. Marked reduction of social phobia, anxiety and depression and increased quality of life after treatment with Botulinum toxin A. Br J Dermatol. 2005;152(2):342–5.
Ouyang X, Wang Z, Luo M, et al. Ketamine ameliorates depressive-like behaviors in mice through increasing glucose uptake regulated by the ERK/GLUT3 signaling pathway. Sci Rep. 2021;11(1):18181.
Beurel E, Toups M, Nemeroff CB. The bidirectional relationship of depression and inflammation: double trouble. Neuron. 2020;107(2):234–56.
Pinto-Sanchez MI, Hall GB, Ghajar K, et al. Probiotic Bifidobacterium longum NCC3001 reduces depression scores and alters brain activity: a pilot study in patients with Irritable Bowel Syndrome. Gastroenterology. 2017;153(2):448-59.e8.
Mazza MG, De Lorenzo R, Conte C, et al. Anxiety and depression in COVID-19 survivors: role of inflammatory and clinical predictors. Brain Behav Immun. 2020;89:594–600.
Kiecolt-Glaser JK, Derry HM, Fagundes CP. Inflammation: depression fans the flames and feasts on the heat. Am J Psychiatry. 2015;172(11):1075–91.
Koo J, Marangell LB, Nakamura M, et al. Depression and suicidality in psoriasis: review of the literature including the cytokine theory of depression. J Eur Acad Dermatol Venereol. 2017;31(12):1999–2009.
Goldsmith DR, Rapaport MH, Miller BJ. A meta-analysis of blood cytokine network alterations in psychiatric patients: comparisons between schizophrenia, bipolar disorder and depression. Mol Psychiatry. 2016;21(12):1696–709.
Duman RS, Sanacora G, Krystal JH. Altered connectivity in depression: GABA and glutamate neurotransmitter deficits and reversal by novel treatments. Neuron. 2019;102(1):75–90.
Chen YP, Wang C, Xu JP. Chronic unpredictable mild stress induced depression-like behaviours and glutamate-glutamine cycling dysfunctions in both blood and brain of mice. Pharm Biol. 2019;57(1):280–6.
Sanacora G, Zarate CA, Krystal JH, et al. Targeting the glutamatergic system to develop novel, improved therapeutics for mood disorders. Nat Rev Drug Discov. 2008;7(5):426–37.
Smaragdi A, Chavez S, Lobaugh NJ, et al. Differential levels of prefrontal cortex glutamate+glutamine in adults with antisocial personality disorder and bipolar disorder: a proton magnetic resonance spectroscopy study. Prog Neuropsychopharmacol Biol Psychiatry. 2019;93:250–5.
Kim HB, Yoo JY, Yoo SY, et al. Early-life stress induces EAAC1 expression reduction and attention-deficit and depressive behaviors in adolescent rats. Cell Death Discov. 2020;6:73.
Ferrero AJ, Cereseto M, Reinés A, et al. Chronic treatment with fluoxetine decreases seizure threshold in naïve but not in rats exposed to the learned helplessness paradigm: correlation with the hippocampal glutamate release. Prog Neuropsychopharmacol Biol Psychiatry. 2005;29(5):678–86.
Jiang S, Zhang QA, Guo Q, et al. The glutamatergic system and astrocytic impairment in rat hippocampus: a comparative study of underlying etiology and pathophysiology of depression. J Integr Neurosci. 2019;18(4):387–92.
Laird KT, Krause B, Funes C, et al. Psychobiological factors of resilience and depression in late life. Transl Psychiatry. 2019;9(1):88.
Alexopoulos GS. Mechanisms and treatment of late-life depression. Transl Psychiatry. 2019;9(1):188.
Stone TW. Endogenous neurotoxins from tryptophan. Toxicon. 2001;39(1):61–73.
Stone TW, Addae JI. The pharmacological manipulation of glutamate receptors and neuroprotection. Eur J Pharmacol. 2002;447(2–3):285–96.
Stone TW, Forrest CM, Mackay GM, et al. Tryptophan, adenosine, neurodegeneration and neuroprotection. Metab Brain Dis. 2007;22(3–4):337–52.
Corona AW, Huang Y, O’Connor JC, et al. Fractalkine receptor (CX3CR1) deficiency sensitizes mice to the behavioral changes induced by lipopolysaccharide. J Neuroinflammation. 2010;7:93.
O’connor JC, Lawson MA, André C, et al. Lipopolysaccharide-induced depressive-like behavior is mediated by indoleamine 2,3-dioxygenase activation in mice. Mol Psychiatry. 2009;14(5):511–22.
Zunszain PA, Anacker C, Cattaneo A, et al. Interleukin-1β: a new regulator of the kynurenine pathway affecting human hippocampal neurogenesis. Neuropsychopharmacology. 2012;37(4):939–49.
Kruse JL, Cho JH, Olmstead R, et al. Kynurenine metabolism and inflammation-induced depressed mood: a human experimental study. Psychoneuroendocrinology. 2019;109:104371.
Deng Y, Zhou M, Wang J, et al. Involvement of the microbiota-gut-brain axis in chronic restraint stress: disturbances of the kynurenine metabolic pathway in both the gut and brain. Gut Microbes. 2021;13(1):1–16.
Vaugeois JM, Passera G, Zuccaro F, et al. Individual differences in response to imipramine in the mouse tail suspension test. Psychopharmacology. 1997;134(4):387–91.
Liang H, Chen M, Qi F, et al. The proatherosclerotic function of indoleamine 2, 3-dioxygenase 1 in the developmental stage of atherosclerosis. Signal Transduct Target Ther. 2019;4:23.
Gibney SM, Fagan EM, Waldron AM, et al. Inhibition of stress-induced hepatic tryptophan 2,3-dioxygenase exhibits antidepressant activity in an animal model of depressive behaviour. Int J Neuropsychopharmacol. 2014;17(6):917–28.
Noakes R. Frontal fibrosing alopecia. An example of disrupted aryl hydrocarbon receptor-mediated immunological homeostasis in the skin? Clin Cosmet Investig Dermatol. 2020;13:479–84.
Kiens O, Taalberg E, Ivanova V, et al. The effect of obstructive sleep apnea on peripheral blood amino acid and biogenic amine metabolome at multiple time points overnight. Sci Rep. 2021;11(1):10811.
Wang Y, Lawson MA, Dantzer R, et al. LPS-induced indoleamine 2,3-dioxygenase is regulated in an interferon-gamma-independent manner by a JNK signaling pathway in primary murine microglia. Brain Behav Immun. 2010;24(2):201–9.
Walker AK, Wing EE, Banks WA, et al. Leucine competes with kynurenine for blood-to-brain transport and prevents lipopolysaccharide-induced depression-like behavior in mice. Mol Psychiatry. 2019;24(10):1523–32.
Sanacora G, Treccani G, Popoli M. Towards a glutamate hypothesis of depression: an emerging frontier of neuropsychopharmacology for mood disorders. Neuropharmacology. 2012;62(1):63–77.
Sanacora G, Banasr M. From pathophysiology to novel antidepressant drugs: glial contributions to the pathology and treatment of mood disorders. Biol Psychiatry. 2013;73(12):1172–9.
Miller AH, Maletic V, Raison CL. Inflammation and its discontents: the role of cytokines in the pathophysiology of major depression. Biol Psychiatry. 2009;65(9):732–41.
Kiecolt-Glaser JK, Glaser R, Christian LM. Omega-3 fatty acids and stress-induced immune dysregulation: implications for wound healing. Mil Med. 2014;179(11 Suppl):129–33.
Wade AT, Davis CR, Dyer KA, et al. A Mediterranean diet with fresh, lean pork improves processing speed and mood: cognitive findings from the MedPork randomised controlled trial. Nutrients. 2019;11(7):1521.
Bozzatello P, Rocca P, Mantelli E, et al. Polyunsaturated fatty acids: what is their role in treatment of psychiatric disorders? Int J Mol Sci. 2019;20(21):5257.
Savitz J. The kynurenine pathway: a finger in every pie. Mol Psychiatry. 2020;25(1):131–47.
Hodes GE, Pfau ML, Leboeuf M, et al. Individual differences in the peripheral immune system promote resilience versus susceptibility to social stress. Proc Natl Acad Sci U S A. 2014;111(45):16136–41.
Maes M, Kubera M, Obuchowiczwa E, et al. Depression’s multiple comorbidities explained by (neuro)inflammatory and oxidative and nitrosative stress pathways. Neuro Endocrinol Lett. 2011;32(1):7–24.
Kadriu B, Farmer CA, Yuan P, et al. The kynurenine pathway and bipolar disorder: intersection of the monoaminergic and glutamatergic systems and immune response. Mol Psychiatry. 2021;26(8):4085–95.
Zádor F, Joca S, Nagy-grócz G, et al. Pro-inflammatory cytokines: potential links between the endocannabinoid system and the kynurenine pathway in depression. Int J Mol Sci. 2021;22(11):5903.
Kang JB, Lee SY, Park DJ, et al. Decrease of 14–3–3 proteins by glutamate exposure in the cerebral cortex of newborn rats. Lab Anim Res. 2020;36:1.
Delorenzo C, Sovago J, Gardus J, et al. Characterization of brain mGluR5 binding in a pilot study of late-life major depressive disorder using positron emission tomography and [11C]ABP688. Transl Psychiatry. 2015;5(12): e693.
Haroon E, Fleischer CC, Felger JC, et al. Conceptual convergence: increased inflammation is associated with increased basal ganglia glutamate in patients with major depression. Mol Psychiatry. 2016;21(10):1351–7.
Cui W, Ning Y, Hong W, et al. Crosstalk between inflammation and glutamate system in depression: signaling pathway and molecular biomarkers for ketamine’s antidepressant effect. Mol Neurobiol. 2019;56(5):3484–500.
Fang Y, Shi B, Liu X, et al. Xiaoyao pills attenuate inflammation and nerve injury induced by lipopolysaccharide in hippocampal neurons in vitro. Neural Plast. 2020;2020:1.
Bigenzahn S, Juergens B, Mahr B, et al. No augmentation of indoleamine 2,3-dioxygenase (IDO) activity through belatacept treatment in liver transplant recipients. Clin Exp Immunol. 2018;192(2):233–41.
Du L, Xing Z, Tao B, et al. Both IDO1 and TDO contribute to the malignancy of gliomas via the Kyn-AhR-AQP4 signaling pathway. Signal Transduct Target Ther. 2020;5(1):10.
Rago D, Rasmussen MA, Lee-Sarwar KA, et al. Fish-oil supplementation in pregnancy, child metabolomics and asthma risk. EBioMedicine. 2019;46:399–410.
Metz R, Duhadaway JB, Kamasani U, et al. Novel tryptophan catabolic enzyme IDO2 is the preferred biochemical target of the antitumor indoleamine 2,3-dioxygenase inhibitory compound D-1-methyl-tryptophan. Cancer Res. 2007;67(15):7082–7.
LöB S, Königsrainer A, Zieker D, et al. IDO1 and IDO2 are expressed in human tumors: levo- but not dextro-1-methyl tryptophan inhibits tryptophan catabolism. Cancer Immunol Immunother. 2009;58(1):153–7.
Shon WJ, Lee YK, Shin JH, et al. Severity of DSS-induced colitis is reduced in Ido1-deficient mice with down-regulation of TLR-MyD88-NF-kB transcriptional networks. Sci Rep. 2015;5:1.
Yamamoto Y, Yamasuge W, Imai S, et al. Lipopolysaccharide shock reveals the immune function of indoleamine 2,3-dioxygenase 2 through the regulation of IL-6/stat3 signalling. Sci Rep. 2018;8(1):15917.
Moreno AC, Clara RO, Coimbra JB, et al. The expanding roles of 1-methyl-tryptophan (1-MT): in addition to inhibiting kynurenine production, 1-MT activates the synthesis of melatonin in skin cells. Febs J. 2013;280(19):4782–92.
Lob S, Konigsrainer A, Schafer R, et al. Levo- but not dextro-1-methyl tryptophan abrogates the IDO activity of human dendritic cells. Blood. 2008;111(4):2152–4.
Dantzer R, O’Connor JC, Freund GG, et al. From inflammation to sickness and depression: when the immune system subjugates the brain. Nat Rev Neurosci. 2008;9(1):46–56.
Rosenblat JD, Cha DS, Mansur RB, et al. Inflamed moods: a review of the interactions between inflammation and mood disorders. Prog Neuropsychopharmacol Biol Psychiatry. 2014;53:23–34.
Connick JH, Stone TW. Quinolinic acid effects on amino acid release from the rat cerebral cortex in vitro and in vivo. Br J Pharmacol. 1988;93(4):868–76.
Cho HJ, Savitz J, Dantzer R, et al. Sleep disturbance and kynurenine metabolism in depression. J Psychosom Res. 2017;99:1–7.
Haroon E, Welle JR, Woolwine BJ, et al. Associations among peripheral and central kynurenine pathway metabolites and inflammation in depression. Neuropsychopharmacology. 2020;45(6):998–1007.
Paul ER, Schwieler L, Erhardt S, et al. Peripheral and central kynurenine pathway abnormalities in major depression. Brain Behav Immun. 2022. https://doi.org/10.1016/j.bbi.2022.01.002.
Bay-richter C, Linderholm KR, Lim CK, et al. A role for inflammatory metabolites as modulators of the glutamate N-methyl-D-aspartate receptor in depression and suicidality. Brain Behav Immun. 2015;43:110–7.
Acknowledgements
We sincerely thank the 3rd Clinical College of Zhejiang Chinese Medical University, Hangzhou, Zhejiang, China for offering the experimental areas. We also thank all teachers and students there for directing empirical methods.
Funding
This research was supported by the National Natural Science Foundation of China (82004463), and Zhejiang Provincial Program for the Cultivation of High-level Innovative Health Talents (2020).
Author information
Authors and Affiliations
Contributions
XW and BC contributed equally to this work as co-first authors. QG and RM designed this study. XW and BC analyzed the data. XW, ZD, SJ wrote the manuscript. HX, MS, RH, SS, ZS and JL participated in the revision of the article, QG and RM validated the manuscript. All authors had read and approved the final manuscript.
Corresponding authors
Ethics declarations
Ethics approval and consent to participate
All experiments involving mice were carried out in accordance with the Institutional Animal Care and Use Committee (IACUC) approved protocol and Association for Assessment and Accreditation of Laboratory Animal Care approved Facility at the Animal Ethical and Welfare Committee of ZheJiang Chinese Medical University. The study was carried out in compliance with the ARRIVE guidelines.
Consent for publication
Not applicable.
Competing interests
The authors declare that they have no conflict of interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Additional file 1: Figure S1.
The correlation analysis between the level of QA and GLU, QA andAQ5 cytokines respectively. (A-E) The level of GLU(A), KYN/TRP(B), KYN(C) KYNA(D) and QA(E) in the brain were measured via HPLC. (F-I) The results of GLU(E), KYN/TRP(F), QA(G) and TRP(H) in the serum. Data are expressed as the mean ± SEM. ** p < 0.01, ****p < 0.0001 vs. control group; #p < 0.05, ##p < 0.01, ###p < 0.001, ####p < 0.0001 vs CUMS. The Pearson correlation coefficients were used to evaluate the correlation of continuous variables, which are scaled such that they range from –1 to + 1, where 0 indicates that there is no linear or monotonic association, and the relationship gets stronger and ultimately approaches a straight line, what’s more, datas with correlation coefficient < 0.25 were evaluated as ‘weak’correlation, ≥ 0.25 and < 0.5 as ‘moderate’ correlation, ≥ 0.5 and < 0.75 as ‘strong’ and ≥ 0.75 as very strong correlation(PMID: 30672319).
Additional file 2: Table S2.
The mean and SEM of metabolites for all the groups.
Additional file 3: Table S3.
Lowest level of quantification and the intra-assay percentage of coefficient of variation for the analytes measured by high performance liquid chromatography with tandem mass spectrometry.
Additional file 4.
The chromatogram of KYNA and QA.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated in a credit line to the data.
About this article

Cite this article
Wu, X., Chen, B., Di, Z. et al. Involvement of kynurenine pathway between inflammation and glutamate in the underlying etiopathology of CUMS-induced depression mouse model. BMC Neurosci 23, 62 (2022). https://doi.org/10.1186/s12868-022-00746-4
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s12868-022-00746-4