
Abstract
This paper formulates an infected predator-prey model with Beddington-DeAngelis functional response from a classical deterministic framework to a stochastic differential equation (SDE). First, we provide a global analysis including the global positive solution, stochastically ultimate boundedness, the persistence in mean, and extinction of the SDE system by using the technique of a series of inequalities. Second, by using Itô’s formula and Lyapunov methods, we investigate the asymptotic behaviors around the equilibrium points of its deterministic system. The solution of the stochastic model has a unique stationary distribution, it also has the characteristics of ergodicity. Finally, we present a series of numerical simulations of these cases with respect to different noise disturbance coefficients to illustrate the performance of the theoretical results. The results show that if the intensity of the disturbance is sufficiently large, the persistence of the SDE model can be destroyed.
Similar content being viewed by others
1 Introduction
Mathematical inequalities play a large role in mathematics analysis and its application. Recently, the inequality technique was applied to impulsive differential systems [1, 2] and stochastic differential systems [3–5], thus some new results were obtained.
Predation can have far-reaching effects on biological communities. Thus many scientists have studied the interaction between predator and prey [6–10]. Interaction between predator and prey is hard to avoid being influenced by some factors. One of the most common factors is the disease. Therefore, there are many scholars who have studied the infected predator-prey systems [11–17]. For instance, Hadeler and Freedman [16] considered a predator-prey system with parasitic infection. They proved the epidemic threshold theorem for where there is coexistence of the predator with the uninfected prey. Han and Ma [15] analyzed four modifications of a predator-prey model to include an SIS or SIR parasitic infection. They obtained the thresholds and global stability results of the four systems.
Species may be subject to uncertain environmental disturbances, such as fluctuations of birth rate and death rate, food, habitat and water, etc. These phenomena can be described by stochastic processes. Recently, the stochastic predator-prey systems have received much attention from scholars [18–21]. Zhang and Jiang [18] studied a stochastic three species eco-epidemiological system. They analyzed the stochastic stability and asymptotic behaviors around the equilibrium points of its deterministic model. Liu and Wang [19] considered a two-species non-autonomous predator-prey model with white noise. They obtained the sufficient criteria for extinction, non-persistence in the mean, and weak persistence in the mean.
The functional response of predator is a very important factor of predator-prey system, which reflects the average consumption rate of predator to prey. Therefore, many scholars prefer to study the predator-prey system with functional response [22–25]. For instance, Wang and Wei [22] explored a predator-prey system with strong Allee effect and an Ivlev-type functional response. Liu and Beretta [23] studied a predator-prey model with a Beddington-DeAngelis functional response. Some biologists have argued that in many instances, especially when predators have to hunt for food and, therefore, have to share or compete for food, the functional response in a prey-predator model should be predator-dependent. This view has been supported by some practical observations [26, 27]. Skalski and Gilliam [26] collected observation data from 19 predator-prey communities, they found that three predator-dependent functional responses (Crowley-Martin [28], Hassell-Varley [29] and Beddington-DeAngelis [30, 31]) were in agreement with the observation data, and in many instances, the Beddington-DeAngelis type looked better than the other two.
The Beddington-DeAngelis type functional response of per capita feeding rate can be expressed as follows:
where a (units: time−1) represents the effects of capture rate on the feeding rate, p (units: prey−1) denotes the effects of handling time on the feeding rate, q (units: predator−1) represents the magnitude of interference among predators. Compared with the Holling II functional response, the Beddington-DeAngelis type functional response has an additional term qy in the denominator. In other words, this type of functional response is affected by both predator and prey. Therefore, the effect of mutual interference on the dynamics of population is worth studying.
To the best of our knowledge, the research on global asymptotic behaviors of a stochastic infected predator-prey system with Beddington-DeAngelis has not gone very far yet. Therefore, according to a deterministic predator-prey model, this paper investigates the stationary distribution and ergodic property of a stochastic infected predator-prey with Beddington-DeAngelis and explores the influence of white noise on the persistence in mean and extinction of the predator-prey-disease system.
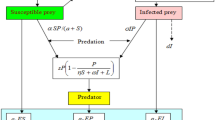
First of all, a deterministic predator-prey system is described in [32] by
where \(X(t)\) is the population density of prey at time t, \(S(t)\) and \(I(t)\), respectively, stand for the densities of susceptible predator and infected predator at time t, b is the intrinsic growth rate of \(X(t)\), c is the natural mortality rate of \(S(t)\), d is the diseased death rate of \(I(t)\). \(a_{11}, a_{22}, a_{33}\), respectively, stand for the density coefficients of \(X(t), S(t)\) and \(I(t)\). \(a_{12}\) is the captured rate of \(X(t)\), \(\frac{a_{21}}{a_{12}}\) is the conversion rate from \(X(t)\) to \(S(t)\), β represents the infection rate from \(S(t)\) to \(I(t)\), \(p,q>0\) are constant coefficients.
Second, the world is full of uncertainty and random phenomena, so species in the ecosystem may be subject to different forms of random interference. In this paper, we assume that the disturbance in the environment affects not only the rate of predation but also the infection rate of the disease, so that
where \(B_{1}(t)\) and \(B_{2}(t)\) are standard Brownian motions, \(\sigma _{12}^{2},\sigma_{21}^{2}\), and \(\sigma^{2}\) are the intensities of the Brownian motions.
Taking into account the effects of random interference gives
The rest of this paper is organized as follows. In the next section, we consider the existence of a global positive solution and the stochastically ultimate boundedness of model (2). In Section 3, we study the global asymptotic behaviors of model (2) around the equilibrium points of its deterministic system. In addition, we explore the stationary distribution and ergodic property of model (2). In Section 4, we obtain the conditions for the persistence in mean and extinction of model (2). In the last section, we summarize our main results and give some numerical simulations.
Throughout this paper, let \((\Omega,\mathcal{F},\{\mathcal{F}\}_{t\geq 0},\mathcal{P})\) be a complete probability space with a filtration \(\{\mathcal{F}_{t}\} _{t\geq0}\) satisfying the usual conditions (i.e. it is increasing and right continuous while \(\mathcal{F}_{0}\) contains all \(\mathcal{P}\)-null sets). The function \(B_{i}(t)\) (\(i=1,2\)) is a Brownian motion defined on the complete probability space Ω. For an integrable function \(X(t)\) on \([0,+\infty)\), we define \(\langle X(t)\rangle=\frac{1}{t}\int ^{t}_{0}X(s)\,ds, \langle X(t)\rangle_{*}=\liminf_{t\rightarrow+\infty}\langle X(t)\rangle, \langle X(t)\rangle^{*}=\limsup_{t\rightarrow+\infty }\langle X(t)\rangle\).
2 Global positive solution and stochastically ultimate boundedness
2.1 Global positive solution
Due to the physical meaning, variables \(S(t)\), \(I(t)\), and \(Y(t)\) in model (2) should remain nonnegative for \(t\geq0\). We next prove that this is actually the case and, furthermore, the positive solution is unique.
Lemma 2.1
For any initial value \((X(0),S(0),I(0))\in R_{+}^{3}\), model (2) has a local unique positive solution \((X(t),S(t),I(t))\) on \(t\in[0,\tau _{e})\), where \(\tau_{e}\) is the explosion time.
Theorem 2.1
For any initial value \((X(0),S(0),I(0))\in R_{+}^{3}\), model (2) has a unique positive solution \((X(t),S(t),I(t))\in R_{+}^{3}\) on \(t\geq0\) with probability 1.
Proof
By Lemma 2.1, we only need to prove that \(\tau_{e}=\infty\) a.s. To this end, let \(k_{0}>0\) be a sufficiently large constant such that \(X(0),S(0)\) and \(I(0)\) all lie in \([\frac{1}{k_{0}},k_{0}]\). For each \(k\geq k_{0}\) (\(k\in N_{+}\)), define the stopping time
As is easy to see, \(\tau_{k}\) is a monotonically increasing function when \(k\rightarrow\infty\). Let \(\tau_{\infty}= {\lim_{k\rightarrow\infty}}\tau_{k}\), thus \(\tau_{\infty}\leq\tau_{e}\) a.s. Now we need to prove \(\tau_{\infty}=\infty\) a.s., otherwise, there exist two constants \(T>0\) and \(\epsilon\in(0,1)\) such that \(P\{\tau _{\infty}\leq T\}>\epsilon\). Thus, there is an integer \(k_{1}\geq k_{0}\) such that
Define a \(C^{3}\)-function \(V:R^{3}_{+}\rightarrow R_{+}\),
The non-negativity of the function \(V(X,S,I)\) can be seen by \(u-1-\ln u\geq0,u>0\).
Applying Itô’s formula to the stochastic differential system (2) yields
where
Since
where \(C_{0}\) is a positive constant.
Then we have
and
Therefore, we have
where \(K_{0}\) is a positive constant.
So we have
Integrating (5) from 0 to \(\tau_{k}\wedge T\) and taking expectation on both sides, we have
Let \(\Omega_{k}=\{\tau_{k}\leq T\}\), from inequality (3) we can see that \(P(\Omega_{k})\geq\epsilon\). Note that, for every \(\omega\in \Omega_{k}\), there exists at least one of \(X(\tau_{k},\omega),S(\tau _{k},\omega),I(\tau_{k},\omega)\) that equals either k or \(\frac{1}{k}\). As a result, we have
Applying equation (6) and equation (7), we get
where \(1\Omega_{k}\) is the indicator function of \(\Omega_{k}\).
When \(k\rightarrow\infty\), we have
This is a contradiction. So \(\tau_{\infty}=\infty\). □
2.2 Stochastically ultimate boundedness
Theorem 2.1 shows that \(R_{+}^{3}\) is the positive invariant set of model (2). Now we prove the stochastically ultimate boundedness of model (2).
Definition 2.1
Let \((X(t),S(t),I(t))\) be the solution of model (2) with initial value \((X(0), S(0),I(0))\in R^{3}_{+}\). If, for any \(\varepsilon\in(0,1)\), there exists a \(\chi(=\chi(\omega ))>0\) such that the solution of model (2) satisfies
then model (2) has stochastically ultimate boundedness.
Lemma 2.2
The following elementary inequality will be used frequently in the sequel.
-
(1)
\(x^{r}\leq1+r(x-1), x\geq0, 1\geq r\geq0\),
-
(2)
\(n^{(1-p/2)\wedge0}|x|^{p}\leq\sum_{i=1}^{n}x_{i}^{p}\leq n^{(1-p/2)\vee0}|x|^{p}\),
where \(R_{+}^{n}:=\{x\in R^{n}:x_{i}>0,1\leq i\leq n\}, n\in R_{+}, p>0\).
Theorem 2.2
Let \((X(t),S(t),I(t))\) be the solution of model (2) with initial value \((X(0),S(0), I(0))\in R^{3}_{+}\), then \((X(t),S(t),I(t))\) is stochastically ultimate boundedness.
Proof
Define
Applying Itô’s formula to stochastic differential system (2) yields
where
Applying the Hölder inequality \(ab\leq\frac{a^{p}}{p}+\frac {b^{q}}{q}, \frac{1}{p}+\frac{1}{q}=1\) (\(p,q>1\)), we have
Therefore,
where \(H_{0}>0\) is a positive constant.
Thus
Applying Itô’s formula to \(e^{t}V(X,S,I)\) yields
So we have
and
Applying the second inequality of Lemma 2.2 and letting \(n=3, p=\frac{1}{2}\), we have
Thus, we obtain
Therefore, for any \(\varepsilon>0\), set \(\chi=\frac{H^{2}}{\varepsilon ^{2}}\), applying the Chebyshev inequality, we have
that is,
□
3 Asymptotic behaviors
System (1) has three equilibrium points [32]: (i) when \(R_{0}=\frac{a_{21}b}{c(a_{11}+pb)}<1\), system (1) has an equilibrium point \(E_{1}(K,0,0)\); (ii) when \(R_{0}=\frac {a_{21}b}{c(a_{11}+pb)}>1\) and \(R_{1}=\frac{a_{21}b}{ (c+\frac{da_{22}}{\beta} ) (a_{11}+pb+\frac{qda_{11}}{\beta} )}<1\), system (1) has another disease free equilibrium point \(E_{2}(\overline{X},\overline {S},0)\); (iii) when \(R_{1}=\frac{a_{21}b}{ (c+\frac{da_{22}}{\beta } ) (a_{11}+pb+\frac{qda_{11}}{\beta} )}>1\), system (1) has a positive equilibrium point \(E_{3}(X^{*},S^{*},I^{*})\). For its stochastic system (2), however, these equilibrium points do not exist.
In this section, we study the asymptotic behaviors of model (2) around the three equilibrium points \(E_{1}(K,0,0)\), \(E_{2}(\overline {X},\overline{S},0)\), and \(E_{3}(X^{*},S^{*},I^{*})\) of its deterministic model (1), respectively.
3.1 Asymptotic behaviors around the equilibrium point \(E_{1}\) of system (1)
When \(R_{0}<1\), system (1) has an equilibrium point \(E_{1}(K,0,0)=(\frac{b}{a_{11}},0,0)\), but it is not the equilibrium point of system (2). In this subsection, we study the asymptotic behaviors of system (2) around \(E_{1}(K,0,0)\).
Theorem 3.1
Let \((X(t),S(t),I(t))\) be the solution of model (2) with initial value \((X(0),S(0), I(0))\in R^{3}_{+}\). If \(R_{0}<1\) and \(K=\frac {b}{a_{11}}\leq\frac{c}{a_{21}}\), then
where \(W_{1}=\min \{a_{11},\frac{a_{12}a_{22}}{a_{21}},\frac {a_{12}a_{33}}{a_{21}} \}\).
Proof
Note that \((K,0,0)\) is the equilibrium point of system (1), where \(K=\frac{b}{a_{11}}\).
Define
Applying Itô’s formula to stochastic differential system (2) yields
where
Integrating equation (8) from 0 to t, we obtain
where
is a real-valued continuous local martingale.
Thus
Applying the strong law of large numbers, we obtain \({\lim_{t\rightarrow+\infty}}\frac{M_{1}(t)}{t}=0\).
Dividing equation (9) by t and taking the limit superior, we have
thus
□
Corollary 3.1
From Theorem 3.1, when \(\sigma_{12}=0\), we have
thus when \(R_{0}<1\) and \(K=\frac{b}{a_{11}}\leq\frac{c}{a_{21}}\) hold, the equilibrium point \(E_{1}(K,0,0)\) of system (1) is globally asymptotically stable.
Remark 3.1
From Theorem 3.1, if the interference intensity is sufficiently small, the solution of model (2) will fluctuate around the equilibrium point \(E_{1}(K,0,0)\). Moreover, the fluctuation intensity is related with the disturbance intensity: the fluctuation intensity is positively correlated with the value of \(\sigma_{12}\).
3.2 Asymptotic behaviors around the equilibrium point \(E_{2}\) of system (1)
When \(R_{0}>1\) and \(R_{1}<1\), system (1) has an equilibrium point \(E_{2}(\overline{X},\overline{S},0)\), but it is not the equilibrium point of system (2). In this subsection, we study the asymptotic behaviors of system (2) around \(E_{2}(\overline{X},\overline{S},0)\).
Theorem 3.2
Let \((X(t),S(t),I(t))\) be the solution of model (2) with initial value \((X(0),S(0), I(0))\in R^{3}_{+}\). If \(R_{0}>1, R_{1}<1\) and \(a_{11}q>a_{22}p\), then we have
where
and
Proof
Noting that \((\overline{X},\overline{S},0)\) is the equilibrium point of system (1), thus
Define
Applying Itô’s formula to stochastic differential system (2) yields
where
Similarly,
where
Also, we have
Hence
where
Since \(\beta\overline{S}< d\), thus
Integrating both sides of equation (10) from 0 to t yields
where
and
are real-valued continuous local martingales.
Thus
and
Applying the strong law of large numbers, we have \(\lim_{t\rightarrow+\infty}\frac{M_{i}(t)}{t}=0 \) (\(i=2,3\)).
Dividing equation (11) by t and taking the limit superior, we have
Thus
□
Corollary 3.2
From Theorem 3.2, when \(\sigma_{12}=\sigma_{21}=\sigma=0\), we have
thus when \(a_{11}q>a_{22}p\), \(R_{0}>1\) and \(R_{1}<1\) hold, the equilibrium point \(E_{2}(\overline{X},\overline{S},0)\) of system (1) is globally asymptotically stable.
Remark 3.2
From Theorem 3.2, if the interference intensity is sufficiently small, the solution of model (2) will fluctuates around the equilibrium point \(E_{2}(\overline{X},\overline{S},0)\). Moreover, the fluctuation intensity is related with the disturbance intensity: the fluctuation intensity is positively correlated with the value of \(\sigma_{12},\sigma_{21}\) and σ.
3.3 Asymptotic behaviors around the positive equilibrium point \(E_{3}\) of system (1)
When \(R_{1}>1\), system (1) has a positive equilibrium point \(E_{3}(X^{*},S^{*},I^{*})\), but it is not the equilibrium point of model (2). Now, we explore the asymptotic behaviors of system (2) around \(E_{3}(X^{*},S^{*},I^{*})\).
\(X(t)\) is a temporally homogeneous Markov process in \(E_{l}\), which is given by the stochastic differential equation
where \(E_{l}\subset R^{l}\) represents a l-dimensional Euclidean space.
The diffusion matrix of \(X(t)\) is given by
Assumption 3.1
[33]
Assume that there is a bounded domain \(U\subset E_{l}\) with regular boundary, satisfying the following conditions:
-
(1)
In the domain U and some of its neighbors, the minimum eigenvalue of the diffusion matrix \(A(x)\) is nonzero.
-
(2)
When \(x\in E_{l}\backslash U\), the mean time τ at which a path starting from x to the set U is limited, and \(\sup_{x\in H} E_{x}\tau<\infty\) for every compact subset \(H \subset E_{l}\).
Lemma 3.1
[33]
When Assumption 3.1 holds, the Markov process \(X(t)\) has a stationary distribution \(\mu(\cdot)\) with density in \(E_{l}\). Let \(f(x)\) be a function integrable with respect to the measure μ, where \(x\in E_{l}\), then, for any Borel set \(B\subset E_{l}\), we have
and
Theorem 3.3
Let \((X(t),S(t),I(t))\) be the solution of model (2) with initial value \((X(0),S(0), I(0))\in R^{3}_{+}\). If \(a_{11}q>a_{12}p\) and \(R_{1}>1\) hold, then
where
and
Proof
Noting that \((X^{*},S^{*},I^{*})\) is the equilibrium point of system (1), thus
Define
Applying Itô’s formula to the stochastic differential system (2) yields
where
Similarly,
where
Also, we have
where
Then we have
where
It is easy to see that, for any
the ellipsoid
lies entirely in \(R_{+}^{3}\). Let U to be any neighborhood of the ellipsoid with \(\bar{U}\subseteq E_{3}=R_{+}^{3}\), thus for any \(x\in U\backslash E_{l}\), we have \(LV\leq-\overline{M}\) (M̅ is a positive constant). Therefore, condition (2) in Assumption 3.1 is satisfied. Moreover, there exists a \(G=\min\{\sigma _{1}^{2}x_{1}^{2},\sigma_{2}^{2}x_{2}^{2},\sigma_{3}^{2}x_{3}^{2},(x_{1},x_{2},x_{3})\in\overline {U}\}>0\) such that
for all \(x\in\bar{U}, \xi\in R^{3}\), which means condition (1) in Assumption 3.1 is satisfied. Therefore, the stochastic model (2) has a unique stationary distribution \(\mu(\cdot)\), it also has the ergodic property.
Integrating equation (12) from 0 to t on both sides yields
where
and
are real-valued continuous local martingales.
Thus
and
Applying the strong law of large numbers, we have \(\lim_{t\rightarrow+\infty}\frac{M_{i}(t)}{t}=0\) (\(i=4,5\)).
Dividing equation (13) by t and taking the limit superior, we have
thus
□
Corollary 3.3
From Theorem 3.3, when \(\sigma_{12}=\sigma_{21}=\sigma=0\), we have
Thus when \(a_{11}q>a_{22}p\) and \(R_{1}>1\) hold, the positive equilibrium point \(E_{3}(X^{*},S^{*},I^{*})\) of system (1) is globally asymptotically stable.
Remark 3.3
From Theorem 3.3, if the interference intensity is sufficiently small, the solution of model (2) will fluctuates around the equilibrium point \(E_{3}(X^{*},S^{*},I^{*})\). Moreover, the fluctuation intensity is related with the disturbance intensity: the fluctuation intensity is positively correlated with the value of \(\sigma _{12},\sigma_{21}\) and σ.
Remark 3.4
If the conditions in Theorem 3.3 are hold, then the solution of model (2) has a unique stationary distribution, it also has the ergodic property.
4 Persistence in mean and extinction
When we consider a biological population system, persistence in mean and extinction are two very important properties. In this section, we investigate the persistence in mean and extinction of system (2).
Since there is no equilibrium point in system (2), we cannot determine the persistence of system (2) by proving the stability of the equilibrium point as a deterministic system.
Definition 4.1
[5]
The definition of persistence in mean and extinction are given as follows:
-
(1)
The species \(X(t)\) is said to be in extinction if \({\lim_{t\rightarrow+\infty}}X(t)=0\).
-
(2)
The species \(X(t)\) is said to be in persistence in mean if \({\lim_{t\rightarrow+\infty}}\langle X(t)\rangle_{*}>0\).
Lemma 4.1
[34]
Let \(X(t)\in C(\Omega\times[0,+\infty),R_{+})\).
(1) If there exist \(T>0,\lambda_{0}>0,\lambda,n_{i}\), when \(t\geq T\), we have
then
(2) If there exist \(T>0,\lambda_{0}>0,\lambda>0,n_{i}\), when \(t\geq T\), we have
then \(\langle X\rangle_{*}\geq\frac{\lambda}{\lambda_{0}}\) a.s.
4.1 Persistence in mean
Theorem 4.1
Let \((X(t),S(t),I(t))\) be the solution of model (2) with initial value \((X(0),S(0), I(0))\in R^{3}_{+}\). Model (2) has persistence in mean if conditions \(a_{11}q>a_{12}p, R_{1}>1\), and
hold, that is,
where
\(U_{3}\) and \(W_{3}\) are defined in Theorem 3.3.
Proof
Applying equation (14) in Theorem 3.3 we have
Applying the inequality \(2a^{2}-2ab\leq a^{2}+(a-b)^{2}\) to \(X(t)\), we have
Therefore
When \(\varrho< X^{*}\sqrt{\frac{W_{3}}{U_{0}}}\), we have
Similarly, when \(\varrho< S^{*}\sqrt{\frac{W_{3}}{U_{0}}}\), we have
When \(\varrho< I^{*}\sqrt{\frac{W_{3}}{U_{0}}}\), we have
□
Remark 4.1
From Theorem 4.1, when \(R_{1}>1,a_{11}q>a_{12}p\) and the intensity of random disturbance is sufficiently small, system (2) will persistence in mean. This shows that biological populations can resist a small environmental disturbance to maintain the original persistence.
4.2 Extinction
Extinction and persistence in mean are closely related, so we also concern ourselves with the situation of population extinction. In this subsection, we point out the conditions of predator extinction.
Theorem 4.2
Let \((X(t),S(t),I(t))\) be the solution of model (2) with initial value \((X(0),S(0), I(0))\in R^{3}_{+}\). If one of the following conditions holds:
-
(1)
\(\sigma_{21}>\max \{\frac{a_{21}}{\sqrt{2c}},\sqrt {a_{21}p} \}\),
-
(2)
\(R^{*}=\frac{a_{21}}{pc}-\frac{\sigma_{21}^{2}}{2p^{2}c}<1,\sigma _{21}\leq\sqrt{a_{21p}}\),
then
Proof
Applying Itô’s formula to the second equation of stochastic differential system (2) yields
Case I. When \(\sigma_{21}>\max \{\frac{a_{21}}{\sqrt{2c}},\sqrt {a_{21}p} \}\), inequality (16) takes its maximum value on the interval \([0,\frac{1}{p}]\) at \(\frac{a_{21}}{\sigma_{21}^{2}}\), so we have
Integrating (16) from 0 to t and dividing it by t, we get
Applying Lemma 4.1, we obtain
Case II. When \(R^{*}=\frac{a_{21}}{pc}-\frac{\sigma_{21}^{2}}{2p^{2}c}<1\) and \(\sigma_{21}\leq\sqrt{a_{21p}}\), inequality (16) takes its maximum value on the interval \([0,\frac{1}{p}]\) at \(\frac{1}{p}\), so we have
Integrating (16) from 0 to t and dividing it by t, we obtain
Applying Lemma 4.1, we obtain
Applying Itô’s formula to the third equation of stochastic differential system (2), one has
Since \({\lim_{t\rightarrow+\infty}}S(t)=0\), there is an arbitrarily small constant \(\varepsilon>0\) such that when \(t>T\), we have \(-\frac{\sigma^{2}S^{2}}{2}+\beta S<\varepsilon\), thus
Integrating equation (17) from 0 to t and dividing it by t yields
Applying Lemma 4.1 and the arbitrariness of ε, we obtain
Similarly,
Since \({\lim_{t\rightarrow+\infty}S(t)=0}\), there is an arbitrarily small constant \(\varepsilon>0\) such that when \(t>T\), we have \(\frac{S}{1+pX+qS}<\varepsilon\), thus
Integrating the above equation from 0 to t and dividing it by t, one has
Applying Lemma 4.1 and the arbitrariness of ε, we obtain
On the other hand,
Integrating equation (19) from 0 to t and dividing it by t, we have
Applying Lemma 4.1, we obtain
□
Remark 4.2
From Theorem 4.2, if the intensity of random disturbance is sufficiently large or \(R^{*}<1\) and \(\sigma_{12}\leq\sqrt{a_{21}p}\), the predator population will be extinct.
5 Conclusions and numerical simulations
This paper investigates a stochastic infected predator-prey model with Beddington-DeAngelis functional response. The existence of a global positive solution of model (2) is first proved, then we show the stochastically ultimate boundedness of the solution. In addition, by using the Lyapunov method and Itô’s formula, we study the asymptotic properties and stationary distribution of the solution of model (2) around the equilibrium points of its deterministic. At last, we discuss the persistence in mean and extinction of model (2). The biological significance of the result shows that the external environment disturbance may have a certain effect on the stability of the biological system: the population’s ability to adapt to the environment is limited. If the disturbance in the environment is small enough, the stability of the population will not be destroyed; if large disturbances occur in the environment, it may lead to the extinction of species.
We next give some numerical simulations to support our results. We consider the following discrete equations:
where \(\Delta t=0.01\), \(\Delta W_{ik}\triangleq W(t_{k+1})-W(t_{k})\) obeys the Gaussian distribution \(N(0,\Delta t)\).
In Figure 1, we choose \(X(0)=2,S(0)=2,I(0)=2,b=1,c=0.4,d=0.2,a_{11}=0.8,a_{12}=0.55, a_{21}=0.2,a_{22}=0.1,a_{33}=0.1,\beta =0.2,p=1,q=1\), and step size \(\triangle t=0.001\).

Time sequence diagram and phase portrait of model ( 2 ). (a) The deterministic model; (b) the Brownian motion model with \(\sigma_{12}=\sigma_{21}=\sigma=0.5\); (c) phase portrait: the red ∘ is corresponding to the deterministic model, while the blue ∘ represents the stochastic model.
Under this condition,
The numerical simulation of Figure 1 is consistent with our conclusion in Theorem 3.1.
In Figure 2, we choose \(X(0)=2,S(0)=2,I(0)=2,b=0.6,c=0.2,d=0.3,a_{11}=0.8,a_{12}=0.6, a_{21}=0.8,a_{22}=0.1,a_{33}=0.4,\beta =0.2,p=1,q=1\), and step size \(\triangle t=0.001\).

Time sequence diagram and phase portrait of model ( 2 ). (a)-(c) are a Brownian motion model with \(\sigma_{12}=\sigma _{21}=\sigma=0.1,0.2,0.4\), respectively. (d)-(f) are phase portraits of (a)-(c), respectively. The red ∘ is corresponding to the deterministic model, while the blue ∘ represents the stochastic model.
Under this condition,
In Figure 2(a), we choose \(\sigma_{12}=\sigma_{21}=\sigma=0.1\), thus
In Figure 2(b), we choose \(\sigma_{12}=\sigma_{21}=\sigma=0.2\), thus
In Figure 2(c), we choose \(\sigma_{12}=\sigma_{21}=\sigma=0.4\), thus
Figure 2 shows that the solution of model (2) fluctuates around the equilibrium \(E_{2}(0.6,0.4,0)\). In addition, the fluctuation intensity is related with the disturbance intensity: with the increase of \(\sigma_{12},\sigma_{21},\sigma\), the fluctuation intensity is also increasing. These all meet the conditions of Theorem 3.2.
In Figure 3, we choose \(X(0)=2,S(0)=2,I(0)=2,b=1,c=0.1,d=0.1,a_{11}=0.5,a_{12}=0.3, a_{21}=1,a_{22}=0.1,a_{33}=0.2,\beta =0.5,p=1,q=1\), and step size \(\triangle t=0.001\).

Time sequence diagram and phase portrait of model ( 2 ). (a) The Brownian motion model with \(\sigma_{12}=\sigma _{21}=\sigma=0.03,0.06,0.1\), respectively. (d)-(f) are phase portraits of (a)-(c), respectively. The red ∘ is corresponding to the deterministic model, while the blue ∘ represents the stochastic model.
Under this condition,
In Figure 3(a), we choose \(\sigma_{12}=\sigma_{21}=\sigma=0.03\), thus
In Figure 3(b), we choose \(\sigma_{12}=\sigma_{21}=\sigma=0.06\), thus
In Figure 3(c), we choose \(\sigma_{12}=\sigma_{21}=\sigma=0.1\), thus
Figure 3 shows that the solution of model (2) fluctuates around \(E_{3}(1.9085,0.5231, 0.8071)\). In addition, the fluctuation intensity is related with the disturbance intensity: with the increase of \(\sigma_{12},\sigma_{21}\) and σ, the fluctuation intensity is also increasing. These all meet the conditions of Theorem 3.2.
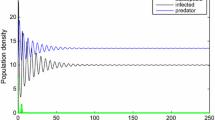
In Figure 4, we choose \(X(0)=2,S(0)=2,I(0)=2,b=1,c=0.1,d=0.1,a_{11}=0.5,a_{12}=0.3, a_{21}=1,a_{22}=0.1,a_{33}=0.2,\beta =0.5,p=1,q=1\), and step size \(\triangle t=0.001\). Figure 4 shows that the solution of model (2) fluctuates up and down in a small neighborhood. According to the density functions in Figure 4(b)-(d), we see that there is a stationary distribution. This is in line with our conclusions.

Time sequence diagram and density function of model ( 2 ) with \(\pmb{\sigma_{12}=\sigma_{21}=\sigma=0.01}\) . (a) Time sequence diagram; (b)-(d) the density functions of \(X(t)\), \(S(t)\), \(I(t)\), respectively.
In Figure 5, we choose \(X(0)=2,S(0)=2,I(0)=2,b=1,c=0.1,d=0.1,a_{11}=0.5,a_{12}=0.3, a_{21}=0.1,a_{22}=0.1,a_{33}=0.2,\beta =0.5,p=0.5,q=0.5\), and step size \(\triangle t=0.001\).

In this condition,
In Figure 5(b), we choose \(\sigma_{12}=\sigma_{21}=\sigma=0.006\). In this case,
which satisfies the conditions in Theorem 4.1. Figure 5(b) shows that \(X(t),S(t),I(t)\) have persistence in mean, this is in line with our conclusion in Theorem 4.1.
In Figure 5(c), we choose \(\sigma_{12}=\sigma=0.5\) and
which satisfies the conditions in Theorem 4.2. Figure 5(c) shows that \(S(t),I(t)\) are extinct and
this is in line with our conclusion in Theorem 4.2.
To sum up, we have the following conclusions:
-
I.
Asymptotic behaviors
-
(1)
When \(R_{0}<1\) and \(Ka_{21}< c\), the solution of model (2) is fluctuating around \(E_{1}\). Therefore, the intensity of the fluctuation is positively correlated with \(\sigma_{12}\).
-
(2)
When \(R_{0}>1,R_{1}<1\) and \(a_{11}q>a_{22}p\), the solution of model (2) is fluctuating around \(E_{2}\). Therefore, the intensity of the fluctuation is positively correlated with \(\sigma_{12},\sigma_{21}\) and σ.
-
(3)
When \(R_{1}>1\) and \(a_{11}q>a_{22}p\), the solution of model (2) is fluctuating around \(E_{3}\). Therefore, the intensity of the fluctuation is positively correlated with \(\sigma_{12},\sigma_{21}\) and σ. When the interference intensity is sufficient small, the solution of model (2) has a unique stationary distribution, it also has the ergodic property.
-
(1)
-
II.
Persistence in mean and extinction
-
(1)
When \(R_{1}>1,a_{11}q>a_{12}p\) and \(\varrho=\max\{\sigma_{12},\sigma _{21},\sigma\}<\min \{X^{*}\sqrt{\frac{W_{3}}{U_{0}}},S^{*}\sqrt{\frac {W_{3}}{U_{0}}},I^{*}\sqrt{\frac{W_{3}}{U_{0}}} \}\), the solution of model (2) can have persistence in mean.
-
(2)
When \(\sigma_{21}>\max \{\frac{a_{21}}{\sqrt{2c}},\sqrt {a_{21}p} \}\) or \(R^{*}<1\) and \(\sigma_{21}\leq\sqrt{a_{21p}}\), the predator of model (2) can be extinct.
-
(1)
References
Gao, SJ, Chen, LS, Nieto, JJ, Torres, A: Analysis of a delayed epidemic model with pulse vaccination and saturation incidence. Vaccine 24, 6037-6045 (2006)
Meng, XZ, Jiao, JJ, Chen, LS: The dynamics of an age structured predator-prey model with disturbing pulse and time delays. Nonlinear Anal., Real World Appl. 9, 547-561 (2008)
Tong, YL: Relationship between stochastic inequalities and some classical mathematical inequalities. J. Inequal. Appl. 1997(1), 364048 (1997)
Zu, L, Jiang, DQ, O’Regan, D: Conditions for persistence and ergodicity of a stochastic Lotka-Volterra predator-prey model with regime switching. Commun. Nonlinear Sci. Numer. Simul. 29, 1-11 (2015)
Meng, XZ, Zhao, SN, Feng, T, Zhang, TH: Dynamics of a novel nonlinear stochastic SIS epidemic model with double epidemic hypothesis. J. Math. Anal. Appl. 433, 227-242 (2016)
Meng, XZ, Liu, R, Liu, LD, Zhang, TH: Evolutionary analysis of a predator-prey community under natural and artificial selections. Appl. Math. Model. 39, 574-585 (2015)
Jiao, JJ, Chen, LS, Cai, SH, Wang, LM: Dynamics of a stage-structured predator-prey model with prey impulsively diffusing between two patches. Nonlinear Anal., Real World Appl. 11, 2748-2756 (2010)
Zhang, H, Georgescu, P, Chen, LS: An impulsive predator-prey system with Beddington-DeAngelis functional response and time delay. Int. J. Biomath. 1, 1-17 (2008)
Song, XY, Chen, LS: Harmless delays and global attractivity for nonautonomous predator-prey system with dispersion. Comput. Math. Appl. 39, 33-42 (2000)
Zhang, TQ, Ma, WB, Meng, XZ, Zhang, TH: Periodic solution of a prey-predator model with nonlinear state feedback control. Appl. Math. Comput. 2667, 95-107 (2015)
Chattopadhyay, J, Arino, O: A predator-prey model with disease in the prey. Nonlinear Anal., Theory Methods Appl. 36, 747-766 (1999)
Xiao, YN, Chen, LS: A ratio-dependent predator-prey model with disease in the prey. Appl. Math. Comput. 131, 397-414 (2002)
Haque, M, Jin, Z, Ezio, V: An ecoepidemiological predator-prey model with standard disease incidence. Math. Methods Appl. Sci. 32, 875-898 (2009)
Xiao, YN, Chen, LS: Modeling and analysis of a predator-prey model with disease in the prey. Math. Biosci. 171, 59-82 (2001)
Han, LT, Ma, ZE, Hethcote, HW: Four predator prey models with infectious diseases. Math. Comput. Model. 34, 849-858 (2001)
Hadeler, KP, Freedman, HI: Predator-prey populations with parasitic infection. J. Math. Biol. 27, 609-631 (1989)
Hethcote, HW, Wang, WD, Han, LT, Ma, ZE: A predator-prey model with infected prey. Theor. Popul. Biol. 66, 259-268 (2004)
Zhang, QM, Jiang, DQ, Liu, ZW, O’Regan, D: Asymptotic behavior of a three species eco-epidemiological model perturbed by white noise. J. Math. Anal. Appl. 433, 121-148 (2016)
Liu, M, Wang, K: Persistence, extinction and global asymptotical stability of a non-autonomous predator-prey model with random perturbation. Appl. Math. Model. 36, 5344-5353 (2012)
Das, K, Reddy, KS, Srinivas, MN, Gazi, NH: Chaotic dynamics of a three species prey-predator competition model with noise in ecology. Appl. Math. Comput. 231, 117-133 (2014)
Li, D, Cui, JA, Song, GH: Permanence and extinction for a single-species system with jump-diffusion. J. Math. Anal. Appl. 430, 438-464 (2015)
Wang, XC, Wei, JJ: Dynamics in a diffusive predator-prey system with strong Allee effect and Ivlev-type functional response. J. Math. Anal. Appl. 422, 1447-1462 (2015)
Liu, SQ, Beretta, E: Predator-prey model of Beddington-DeAngelis type with maturation and gestation delays. Nonlinear Anal., Real World Appl. 11, 4072-4091 (2010)
Jiang, J, Song, YL: Delay-induced Bogdanov-Takens bifurcation in a Leslie-Gower predator-prey model with nonmonotonic functional response. Commun. Nonlinear Sci. Numer. Simul. 19, 2454-2465 (2014)
Meng, XZ, Li, ZQ, Nieto, JJ: Dynamic analysis of Michaelis-Menten chemostat-type competition models with time delay and pulse in a polluted environment. J. Math. Chem. 47, 123-144 (2010)
Skalski, GT, Gilliam, JF: Functional responses with predator interference: viable alternatives to the Holling type II model. Ecology 82, 3083-3092 (2001)
Jost, C, Arditi, R: From pattern to process: identifying predator-prey models from time-series data. Popul. Ecol. 43, 229-243 (2001)
Crowley, PH, Martin, EK: Functional response and interference within and between year classes of a dragonfly population. J. North Am. Benthol. Soc. 8, 211-221 (1989)
Hassell, MP, Varley, GC: New inductive population model for insect parasites and its bearing on biological control. Nature 223, 1133-1137 (1969)
Beddington, JR: Mutual interference between parasites or predators and its effect on searching efficiency. J. Anim. Ecol. 44, 331-340 (1975)
DeAngelis, DL, Goldstein, RA, O’Neill, RV: A model for tropic interaction. Ecology 56, 881-892 (1975)
Li, S, Wang, XP: Analysis of stochastic predator-prey models with disease in the predator and Beddington-DeAngelis functional response. Adv. Differ. Equ. 2015, 224 (2015)
Hasminskij, RZ, Milstejn, GN, Nevelson, MB: Stochastic Stability of Differential Equations. Springer, Berlin (2012)
Liu, M, Wang, K: Survival analysis of a stochastic cooperation system in a polluted environment. J. Biol. Syst. 19, 183-204 (2011)
Acknowledgements
The authors would like to thank Dr. Tonghua Zhang, who helped them during the writing of this paper. This work was supported by National Natural Science Foundation of China (11371230, 11501331, 11561004), the SDUST Research Fund (2014TDJH102), Shandong Provincial Natural Science Foundation, China (ZR2015AQ001, BS2015SF002), Joint Innovative Center for Safe And Effective Mining Technology and Equipment of Coal Resources, the Open Foundation of the Key Laboratory of Jiangxi Province for Numerical Simulation and Emulation Techniques, Gannan Normal University, China, SDUST Innovation Fund for Graduate Students (No. SDKDYC170225).
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
All authors contributed equally to the writing of this paper. All authors read and approved the final manuscript.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Feng, T., Meng, X., Liu, L. et al. Application of inequalities technique to dynamics analysis of a stochastic eco-epidemiology model. J Inequal Appl 2016, 327 (2016). https://doi.org/10.1186/s13660-016-1265-z
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13660-016-1265-z