
Abstract
In this paper, we propose a discrete pollination mutualism in a plant-pollinator system with the Beddington-DeAngelis functional response and feedback controls. By applying the comparison theorem of a difference equation and constructing some suitable Lyapunov functions, sufficient conditions are obtained for the permanence and the extinction of the system. Moreover, under some suitable conditions, we show that the solution of the system is globally attractive. The paper ends by some numerical simulations and a brief discussion.
Similar content being viewed by others


1 Introduction
In theoretical ecology, there are several famous functional responses in the ecosystem, which we refer to as Holling type-I, type-II, type-III, type-IV, Monod-Haldane type, and Hassel-Verley type functional response etc. Some authors studied the ecosystem with different types of functional responses. Beddington [1] and DeAngelis et al. [2] first proposed a predator dependent functional response (known as the B-D functional response). After that, a lot of scholars did work on the ecosystems with the Beddington-DeAngelis functional response. Chen and You [3] studied the permanence, extinction, and periodic solution of the periodic predator-prey system with a Beddington-DeAngelis functional response and stage structure for prey. Xiao [4] analyzed the existence and uniqueness of the positive equilibrium and its global asymptotic stability by using the qualitative methods of ordinary differential equation. In [5], the author focused on the uniform persistence, local stability, and global stability for a Beddington-DeAngelis type stage structures predator-prey model. Furthermore, Chen et al. [6] with the help of a fluctuation lemma obtained a set of new conditions on the global asymptotic stability of the boundary solution.
In the ecological system, we know that it is more appropriate to use a discrete dynamic model to describe these systems when the populations have a short life or non-overlapping generations. Recently, Wu [7, 8] studied the permanence and global stability of a discrete competition feedback-control system with a Beddington-DeAngelis functional response, moreover, she generalized it to n species,
Li et al. [9] proposed a discrete predator-prey systems with a Beddington-DeAngelis functional response and feedback controls as follows:
By applying the comparison theorem of a difference equation, they obtained sufficient conditions to the permanence of the system.
Recently, Wang et al. [10] studied the interactions between pollinators, nectar robbers, defensive plants and non-defensive plants. Among them, the plant-pollinator system is described by a cooperative model with the Beddington-DeAngelis functional response,
where x and y denote population densities of non-defensive plants and pollinators, respectively. \(r_{1}\) represents the intrinsic growth rate of the plants and \(r_{1}/d_{1}\) is their carrying capacity in the absence of visitors. \(r_{2}\) denotes the pollinators’ per capita mortality rate. Since the Beddington-DeAngelis functional response represents a positive effect of the pollinators on the plants, \(\alpha_{12}\) can be regarded as the plants’ efficiency in translating the plant-pollinator interactions into fitness. Similarly, \(\alpha _{21}\) represents the corresponding value for the pollinators. The authors analyzed the global stability of the positive equilibrium of the system.
On the other hand, as was pointed out by Huo and Li [11], ecosystems in the real world are continuously disrupted by unpredictable forces which can result in changes in the biological parameters. For having a more accurate description of such a system, scholars introduced the feedback control into the ecosystems. Moreover, discrete time models, governed by difference equations, are more appropriate than the continuous ones. The above phenomena motivated us to propose and study the discrete non-autonomous pollination mutualism in a plant-pollinator system with a Beddington-DeAngelis functional response and feedback controls as follows:
where \(x_{i}(n)\) (\(i=1,2\)) is the density of \(x_{i}\) species at the nth generation and \(u_{i}(n)\) (\(i=1,2\)) is the feedback-control variable. The coefficients \(r_{i}(n)\), \(d_{1}(n)\), \(a(n)\), \(b(n)\), \(\alpha _{ij}(n)\), \(e_{i}(n)\), \(\eta_{i}(n)\), \(q_{i}(n)\) (\(i,j=1,2\), \(i\neq j\)) are all bounded nonnegative sequences.
Throughout this paper, we use the following notations for any bounded sequence \(\{f(n)\}\):
and assume that \(0<\eta^{l}_{i}\leq\eta^{u}_{i}<1\) (\(i=1,2\)).
According to the biological background of system (1.4), we only consider the solution of the system (1.4) with the following initial conditions:
It is easy to prove that the solution of the system (1.4) which satisfies the initial condition is positive.
We mention here that, as far as the system (1.4) is concerned, whether the system is to persist is a question that we need to solve. Furthermore whether the feedback-control variables have influence on the extinction and the stability of the system or not is an interesting problem. The aim of this paper is to solve the above questions.
The remainder of the paper is organized as follows: in Section 2, we introduce some useful lemmas and obtain the sufficient conditions to guarantee the permanence and the extinction of system (1.4). In Section 3, a set of sufficient conditions which ensure the stability of the system are obtained. In Section 4, we give some examples to illustrate our results, and we end this paper by a brief discussion.
2 Permanence
In this section, we establish a permanence result for system (1.4). First, let us state several lemmas which will be useful in proving the main results.
Now let us consider the first order difference equation
where A and B are positive constants.
Lemma 2.1
[12]
Assume that \(|A|<1\), for any initial value \(y(0)\), there exists a unique solution \(y(n)\) of (2.1) which can be expressed as follows:
where \(y^{\ast}=B/(1-A)\). Thus, for any solution \(y(n)\) of system (2.1),
Lemma 2.2
[12]
Let \(n\in N^{+}_{n_{0}}=\{n_{0}, n_{0}+1,\ldots,n_{0}+l,\ldots\}\), \(r\geq0\). For any fixed n, \(g(n,r)\) is a nondecreasing function with respect to r, and for \(n\geq n_{0}\), the following inequalities hold:
If \(y(n_{0})\leq u(n_{0})\), then \(y(n)\leq u(n)\) for all \(n\geq n_{0}\).
Now let us consider the following single species discrete model:
where \(a(n)\) and \(b(n)\) are strictly positive sequences of real numbers defined for \(n\in N=\{0,1,2,\ldots\}\) and \(0< a^{l}\leq a^{u}\), \(0< b^{l}\leq b^{u}\). We have the following lemma.
Lemma 2.3
Any solution of system (2.5) with initial condition \(N(0)>0\) satisfies
where
Lemma 2.4
[13]
Let \(x(n)\) and \(b(n)\) be nonnegative sequences defined on N and \(c\geq0\) is a constant. If
then
Lemma 2.5
[14]
Assume that \(A>0\) and \(y(0)>0\), and further suppose that
Then, for any integer \(k\leq n\),
Proposition 2.6
Assume that
holds, then
where
Proof
Let \((x_{1}(n), x_{2}(n),u_{1}(n),u_{2}(n))\) be any positive solution of system (1.4), it follows from the first equation of system (1.4) that
By applying Lemmas 2.2 and 2.3, we have
Let \(x_{2}(n)=\exp\{v(n)\}\), then
where
Condition (2.10) shows that Lemma 2.4 could be applied to (2.12), it immediately follows that
This is
For any small enough positive constant ε, it follows from (2.11) and (2.13) that there exists a large enough \(N_{0}\) such that
From the third and fourth equations of the system (1.4) and (2.14), we can obtain
By applying Lemmas 2.1 and 2.2, it immediately follows that
Setting \(\varepsilon\rightarrow0\) in the above inequalities leads to
This completes the proof of Proposition 2.6. □
Theorem 2.7
In addition to (2.10), assume further that
then the system (1.4) is permanent.
Proof
By applying Proposition 2.6, it is easy to see that, to end the proof of Theorem 2.7, it is enough to show that under the conditions of Theorem 2.7,
From Proposition 2.6, we know that for the above ε, there exists a \(N_{1}>N_{0}\) such that
From the first equation of system (1.4) and (2.16), we have
for all \(n>N_{1}\), where \(D_{\varepsilon}=-d^{u}_{1} M_{1\varepsilon }-e^{u}_{1}W_{1\varepsilon}\). So for \(n\geq k\) we have
From the third equation of system (1.4), we have
where \(A=1-\eta^{l}_{1}\), \(B=q^{u}_{1}x_{1}(n)\). Then Lemma 2.5 implies that, for any \(n\geq k\),
Note that
We can choose \(N_{2}=\max\{N_{1}, \frac{\ln P_{1}}{\ln{A}} \}+1\), where \(P_{1}= \frac{r^{u}_{1}}{e^{u}_{1}W_{1\varepsilon}}\). As \(n>N_{2}\), we have \(r^{l}_{1}-e^{u}_{1}A^{N_{2}}W_{1\varepsilon}>0\), then we get
where \(G_{\varepsilon}=q^{u}_{1} \sum^{N_{2}-1}_{i=0}A^{i}\exp\{ -D_{\varepsilon}(i+1)\}\).
Considering the first equation of system (1.4), we have
where \(E_{1\varepsilon}=r^{l}_{1}-e^{u}_{1} A^{N_{2}}W_{1\varepsilon}\), \(E_{2\varepsilon}=e^{u}_{1}G_{\varepsilon}+d^{u}_{1}\).
By applying Lemmas 2.2 and 2.3, it immediately follows that
Setting \(\varepsilon\rightarrow0\) in (2.17) leads to
Then we assume that \(\varepsilon<\frac{1}{2}m_{1}\), from (2.18) we know that there exists a large enough \(N_{2}>N_{1}\) such that
From the second equation of system (1.4), (2.16), and (2.19), we have
for all \(n>N_{2}\).
By applying Lemmas 2.2 and 2.3, it immediately follows that
Setting \(\varepsilon\rightarrow0\), we have
Without loss of generality, we may assume that \(\varepsilon<(1/2)\min\{ m_{1},m_{2}\}\). It follows from (2.18) and (2.21) that there exists a large enough \(N_{3}>N_{2}\), such that
From the third and fourth equations of the system (1.4) and (2.22), we have
By applying Lemmas 2.1 and 2.2, it immediately follows that
Setting \(\varepsilon\rightarrow0\) in the above inequalities leads to
This completes the proof. □
Theorem 2.8
Assume that the inequality
holds. Let \((x_{1}(n), x_{2}(n),u_{1}(n),u_{2}(n))\) be any positive solution of system (1.4), then \(x_{2}(n)\rightarrow0\), \(u_{2}(n)\rightarrow0\) as \(n\rightarrow+\infty\).
Proof
Equation (2.23) is equivalent to the following inequality:
From (2.24), there exists a \(\delta>0\) such that
Let \((x_{1}(n), x_{2}(n),u_{1}(n),u_{2}(n))\) be any positive solution of system (1.4). For any \(q\in N\), according to the second equation of system (1.4), we obtain
Summating both sides of the above inequalities from 0 to \(n-1\), we obtain
then
From (2.27), \(x_{2}(n)\rightarrow0\) as \(n\rightarrow+\infty\).
Further, consider the fourth equation of system (1.4). Applying Lemma 2.3 in [15], we easily obtain \(u_{2}(n)\rightarrow0\) as \(n\rightarrow+\infty\). This completes the proof of Theorem 2.8. □
Theorem 2.9
Assume that (2.23) holds, and further assume that
then, for any two positive solutions \((x_{1}(n),x_{2}(n),u_{1}(n),u_{2}(n))\) and \((x^{\ast}_{1}(n),x^{\ast}_{2}(n), u^{\ast}_{1}(n),u^{\ast}_{2}(n)) \) of the system, we have
Proof
By conditions (2.28), there exist a positive constant ε and δ such that
From Theorems 2.7 and 2.8, for the above ε, there exists \(N_{4}>N_{3}\) such that
Using the mean value theorem, one has
where \(\theta(n)\) is between \(x_{1}(n)\) and \(x^{\ast}_{1}(n)\).
Now we define
From the first equation of the system, we have
From the third equation of the system, we have
Now we define a Lyapunov function as follows:
From (2.31) and (2.32), we have
Summating both sides of the above inequalities from \(N_{4}\) to n, we have
Hence
From Theorem 2.8, we have \(\sum^{+\infty }_{n=0}x_{2}(n)<+\infty\) and \(\sum^{+\infty}_{n=0}x^{\ast}_{2}(n)<+\infty\). We notice that \(V(N_{4})\) is bounded. So from the above inequalities, we have
Therefore
This means that
Consequently
This completes the proof of Theorem 2.9. □
3 Global attractivity
In this section, we will consider the stability of the system (1.4).
Theorem 3.1
In addition to the conditions of Theorem 2.7, assume that the following condition holds:
where \(m_{i}\), \(M_{i}\), \(i=1,2\), are defined as before and
then the solution \((x_{1}(n),x_{2}(n),u_{1}(n),u_{2}(n))\) of the system (1.4) is globally attractive.
Proof
Let \((x_{1}(n),x_{2}(n),u_{1}(n),u_{2}(n))\) and \((x^{\ast}_{1}(n),x^{\ast}_{2}(n),u^{\ast}_{1}(n),u^{\ast}_{2}(n))\) be any two positive solutions of the system (1.4), let
So from the first equation of the system, we have
In a similar way, we get
Also, one has
We have
where \(\xi_{i}(n)\) between \(\ln x_{i}(n)\) and \(\ln x^{\ast}_{i}(n)\).
As follows from the above equation, we have
By (3.1), we can choose a \(\varepsilon>0\) such that
In view of Theorem 2.7, there exists \(N_{4}>N_{3}\) such that
It follows from (3.6) that
Let \(\chi=\max\{\chi^{\varepsilon}_{1},\chi^{\varepsilon}_{2}, \chi^{\varepsilon}_{3},\chi^{\varepsilon}_{4}\}\), then \(0<\chi<1\). It follows from (3.7) that
for \(n>N_{4}\). Then we have
Thus
This completes the proof. □
4 Examples
In the section, we present some examples showing the feasibility of our main results.
Example 4.1
Consider the following system:
Through a simple computation, we have
So Proposition 2.6 holds, moreover,
Therefore the conditions of Theorem 2.7 hold. Then the system (4.1) has permanence.
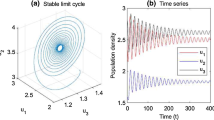
Numeric simulation (Figure 1) also supports this finding.

Dynamics behavior of the solution \(\pmb{{(x_{1}(n),x_{2}(n),u_{1}(n),u_{2}(n))}}\) to the system ( 4.1 ) with the initial conditions \(\pmb{(x_{1}(0),x_{2}(0),u_{1}(0),u_{2}(0))= (1.4,0.2,1.2,0.9), (3.2, 2.1,4.6,2.5), \mbox{and }(5.8,3.9,0.8,4.2)}\) , respectively.
Example 4.2
Consider the following system:
Through a simple computation, we have
Therefore the conditions of Theorems 2.8 and 2.9 hold. Then the species \(x_{2}\) is extinct and the species \(x_{1}\) has global stability.
Numeric simulation (Figure 2) also supports this finding.

Dynamics behavior of the solution \(\pmb{{(x_{1}(n),x_{2}(n),u_{1}(n),u_{2}(n))}}\) of the system ( 4.2 ) with the initial conditions \(\pmb{(x_{1}(0),x_{2}(0),u_{1}(0),u_{2}(0))= (0.05,0.07,0.01,0.04), (0.06, 0.045,0.032,0.021), \mbox{and } (0.04,0.028,0.042,0.008)}\) , respectively.
5 Conclusion
In this paper, we proposed a discrete non-autonomous plant-pollinator system with the Beddington-DeAngelis functional response and feedback controls. As we see it, plants can build a cooperative interaction with pollinators by providing a reward for the pollinators’ services. From Theorem 2.7, we discover that when \(\alpha_{21}(n)\) is large enough, then (2.10) must hold, that is, the system (1.4) has an upper bound. We know that \(\alpha_{21}(n)\) represents the pollinators’ efficiency in translating plant-pollinator interactions into fitness. In other words, when the efficiency of the pollinators is large enough, the system has an upper bound. What is more, when the coefficient \(e_{1}(n)\) is small enough, then the system has permanence. That is, when the interference of the plant is small, the system is to persist. From Theorem 2.8, it is obvious that when \(r_{2}(n)\) is large enough, \(x_{2}(n)\) will contribute to extinction. That is, if the mortality is large enough, then the population will go to extinction. From Theorem 2.9, we known that when \(e_{1}(n)\) is small enough, then the partial species is globally stable. That is, if the feedback control is small enough, the population may remain stable. Wang et al. [10] have shown that the system (1.3) is globally stable, and our work shows that the feedback controls have no influence on the attractivity of the system. The obtained results may be helpful to maintain the plant-pollinator cooperation and provide insight in the mechanisms by which pollination mutualism could persist and we have global attractivity, which may be helpful for understanding the complexity of these systems.
References
Beddington, JR: Mutual interference between parasites or predator and its effect on searching efficiency. J. Anim. Ecol. 44(1), 331-340 (1975)
DeAngelis, DL, Goldstein, RA, O’Neil, RV: A model for tropic interaction. Ecology 56(4), 881-892 (1975)
Chen, FD, You, MS: Permanence, extinction and periodic solution of the predator-prey system with Beddington-DeAngelis functional response and stage structure for prey. Nonlinear Anal., Real World Appl. 9(2), 207-221 (2008)
Xiao, HB: Positive equilibrium and its stability of the Beddington-DeAngelis’ type predator-prey dynamical system. Appl. Math. J. Chin. Univ. Ser. B 21(4), 429-436 (2006)
Khajanchi, S: Dynamic behavior of a Beddington-DeAngelis type stage structured predator-prey model. Appl. Math. Comput. 244, 344-360 (2014)
Chen, FD, Chen, YM, Shi, JL: Stability of the boundary solution of a nonautonomous predator-prey system with the Beddington-DeAngelis functional response. J. Math. Anal. Appl. 344(2), 1057-1067 (2008)
Wu, T: Permanence and global stability of a discrete competition feedback-control system with Beddington-DeAngelis functional response. J. Minjiang Univ. 31(2), 16-20 (2010) (in Chinese)
Wu, T: Permanence of a discrete n-species competition-predator system with Beddington-DeAngelis functional response. Pure Appl. Math. 27(4), 437-441 (2011) (in Chinese)
Li, XP, Yang, WS: Permanence of a discrete predator-prey systems with Beddington-DeAngelis functional response and feedback control. Discrete Dyn. Nat. Soc. 2008, Article ID 149267 (2008)
Wang, YS, Wu, H, Sun, S: Persistence of pollination mutualisms in plant-pollination-robber systems. Theor. Popul. Biol. 81, 243-250 (2012)
Huo, HF, Li, WT: Positive periodic solution of a class of delay differential system with feedback control. Appl. Math. Comput. 148(1), 35-46 (2004)
Chen, FD, Xie, XD: Study on the Dynamic Behaviors of Cooperation Population Modeling. Science Press, Beijing (2014) (in Chinese)
Takeuchi, Y: Global Dynamical Properties of Lotka-Volterra Systems. World Scientific, Singapore (1996)
Fan, YH, Wang, LL: Permanence for a discrete model with feedback control and delay. Discrete Dyn. Nat. Soc. 2008, Article ID 945109 (2008)
Chen, LJ, Chen, FD: Extinction in a discrete Lotka-Volterra competitive system with the effect of toxic substances and feedback controls. Int. J. Biomath. 8(1), 1550012 (2015)
Acknowledgements
The authors are grateful to the anonymous referees for their excellent suggestions, which greatly improved the presentation of the paper. The research was supported by the Natural Science Foundation of Fujian Province (2015J010121, 2015J01019).
Author information
Authors and Affiliations
Corresponding author
Additional information
Competing interests
The authors declare that they have no competing interests.
Authors’ contributions
All authors contributed to the writing of this paper. All authors read and approved the final manuscript.
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Han, R., Xie, X. & Chen, F. Permanence and global attractivity of a discrete pollination mutualism in plant-pollinator system with feedback controls. Adv Differ Equ 2016, 199 (2016). https://doi.org/10.1186/s13662-016-0889-0
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s13662-016-0889-0