
Abstract
Background
Bacillus spp. seem to be an alternative to antimicrobial growth promoters for improving animals’ health and performance. However, there is little information on the effect of Bacillus spp. in combination with different dietary crude protein (CP) levels on the ileal digestibility and microbiota composition. Therefore, the objective of this study was to determine the effect of Bacillus spp. supplementation to low- (LP) and high-protein diets (HP) on ileal CP and amino acid (AA) digestibility and intestinal microbiota composition.
Methods
Eight ileally cannulated pigs with an initial body weight of 28.5 kg were randomly allocated to a row-column design with 8 pigs and 3 periods of 16 d each. The assay diets were based on wheat-barley-soybean meal with two protein levels: LP (14% CP, as-fed) and HP diet (18% CP, as-fed). The LP and HP diets were supplemented with or without Bacillus spp. at a level of 0.04% (as-fed). The apparent ileal digestibility (AID) and standardized ileal digestibility (SID) of CP and AA was determined. Bacterial community composition from ileal digesta was analyzed by Illumina amplicon sequencing and quantitative real-time PCR. Data were analyzed as a 2 × 2 factorial design using the GLIMMIX procedures of SAS.
Results
The supplementation with Bacillus spp. did not affect both AID and SID of CP and AA in growing pigs. Moreover, there was no difference in AID of CP and AA between HP and LP diets, but SID of cystine, glutamic acid, glycine, and proline was lower (P < 0.05) in pigs fed the HP diets. The HP diets increased abundance of Bifidobacterium spp. and Lactobacillus spp., (P < 0.05) and by amplicon sequencing the latter was identified as predominant genus in microbiota from HP with Bacillus spp., whereas dietary supplementation of Bacillus spp. increased (P < 0.05) abundance of Roseburia spp..
Conclusions
The HP diet increased abundance of Lactobacillus spp. and Bifidobacterium spp.. The supplementation of Bacillus spp. resulted in a higher abundance of healthy gut associated bacteria without affecting ileal CP and AA digestibility, whereas LP diet may reduce the flow of undigested protein to the large intestine of pigs.
Similar content being viewed by others


Background
Due to the ban of antimicrobial growth promoters in livestock feeding by the European Union in 2006 [1], probiotics are considered as an alternative for improving animals’ health and performance [2, 3]. Within this regard, Bacillus spp. have the ability to sporulate, thereby making them stable during thermal treatment of feed, and resistant to enzymatic digestion along the gastrointestinal tract (GIT) [4]. Thus, Bacillus spp. such as Bacillus subtilis (B. subtilis) and Bacillus licheniformis (B. licheniformis) are frequently supplemented to pig diets [4–6] as these two species have been listed to be added as non-toxigenic, biological supplements to livestock diets [7], and additionally, they are widely used for the large-scale industrial production of proteins including extracellular enzymes [8]. Positive effects of dietary supplementation of B. subtilis and B. licheniformis on pigs’ growth performance have been reported before [9, 10].
Activity of probiotics is influenced by diet composition [11] and variations in dietary protein supply, thereby possibly affecting microbial composition in the gut [12, 13]. Accordingly, reducing the dietary crude protein (CP) level has been reported to markedly reduce the production of potentially harmful microbial metabolites such as ammonia and amines due to the lower availability of undigested protein for microbial fermentation [14]. Thus, excessive nitrogen (N) excretion by pigs is mitigated, resulting in a decrease of environmental pollutants [15, 16]. In contrast, increasing the dietary CP intake may stimulate the proliferation of almost all bacteria groups over the entire GIT including beneficial bacteria, such as Bifidobacterium spp., and potentially pathogenic bacteria, such as Bacteroides groups [17]. Furthermore, there is increasing evidence that interactions of supplemental probiotics with dietary CP level affect the intestinal microbiome at the ileal level [18].
According to the results of previous studies [19, 20] Bacillus spp. enhanced the development and activities of digestive enzymes in the GIT, which was associated with a numerical increase in apparent ileal digestibility (AID) and standardized ileal digestibility (SID) of some amino acids (AA) in weaning pigs [21]. However, studies with growing pigs in which Bacillus spp. were supplemented to diets varying in CP content are still lacking. Therefore, the objective of the present study was to test the hypothesis, if B. subtilis and B. licheniformis supplementation to low- and high-protein diets will affect ileal CP and AA digestibility and intestinal microbiota composition in growing pigs.
Methods
The research protocol was reviewed and approved by the German Ethical Commission for Animal Welfare, and care of the animals throughout this experiment was in accordance with guidelines issued by the Council Directive [22].
Animals, housing, and surgical procedures
Eight pigs were obtained from the University of Hohenheim Research Station. The average initial and final body weight (BW) of the experimental animals were 28.5 ± 0.8 and 64.3 ± 1.5 kg, respectively. The pigs were housed individually in stainless steel metabolic crates (0.8 m × 1.5 m). Each metabolic crate was equipped with an infrared heating lamp and a low pressure drinking nipple which allowed free access to water. The research unit was equipped with an automated temperature control system kept at 20 °C. Until the beginning of the experiment, the pigs were fed a commercial starter diet at a daily level of 4% (as-fed)/kg of average BW (Porcigold® SMA 134, Raiffeisen Kraftfutterwerke Süd GmbH, Würzburg, Germany; 17.5% CP and 13.4 MJ metabolizable energy (ME)/kg, as-fed). After arrival at the research unit, the pigs were surgically fitted with a simple T-cannula at the distal ileum as described by Li et al. [23]. The pigs were allowed a recovery period of at least 7 d. During this period, the feed allowance was gradually increased, starting from 50 g/d the day after surgery until 1000 g/d (as-fed) were consumed.
Experimental design, diets, and procedures
The experiment was arranged as a row-column design with 8 pigs and 3 experimental periods of 16 d each. Pigs were fed assay diets twice daily at 0700 and 1900 h at a level of 4% (as-fed)/kg of their average BW corresponding to 3 times their energy requirement for maintenance (i.e. 0.44 MJ ME/kg BW0.75) [24]. Pigs’ BW was determined at the beginning of each experimental period.
The assay diets were based on wheat, barley, and soybean meal with 2 protein levels resulting in a low-protein (14% CP, as-fed; LP) and a high-protein diet (18% CP, as-fed; HP). The LP diet was accomplished by blending the HP diet with 25% of native cornstarch. The contents of oil, minerals, vitamins, and titanium dioxide were the same for all diets. The Bacillus spp. product is comprised of a mixture of spray-dried spores of B. licheniformis and B. subtilis. The LP and HP diets were supplemented with (+) or without (-) Bacillus spp. at a level of 0.04% (as-fed). All assay diets were formulated (Table 1) to meet or exceed the dietary threshold levels for CP and AA according to Fan et al. [25] and NRC [26] nutrient recommendations for pigs from 25 to 50 kg BW. Vitamins and minerals were supplemented to all diets to meet or exceed NRC [26] standard, and all diets contained titanium dioxide at a level of 0.4% (as-fed basis) as an indigestible marker.
The assay diets were in a mash form mixed with water (1/1, w/v). During each of the 3 experimental periods, the pigs were allowed to adapt to their assay diets for 14 d before ileal digesta was collected for a total of 24 h from 0700 to 1900 h on d 15 and from 1900 on d 16 to 0700 h on d 17. Digesta collection procedure was adapted from Li et al. [23] using soft plastic bags attached to the barrel of the cannula by elastic bands. The bags were changed at least every 20 min. To minimize further bacterial fermentation 4 mL of 2.5 mol/L formic acid were added to the sampling bags and then immediately frozen at -18 °C. The individual digesta samples of each pig were pooled for each sampling period, freeze-dried, and ground to 0.5 mm prior to analyses. For analyses of intestinal microbiota composition, ileal digesta and feces samples were taken prior to the first experimental period (starter period) and on d 15 once for each experimental period. Ileal digesta and feces samples for microbial community analysis were immediately put on ice before being stored in a freezer at -80 °C for subsequent treatment and analyses.
Chemical analyses
Official standard methods [27] were used to determine contents of proximate nutrients, neutral detergent fiber (NDF), acid detergent fiber (ADF), acid detergent lignin (ADL), and microbial numbers of B. subtilis and B. licheniformis in assay diets. The assay diets and digesta samples were analyzed for DM (method 3.1). In addition, assay diets were analyzed for ash (method 8.1); ether extract (EE; method 5.1.1 using petroleum ether), NDF assayed with a heat-stable amylase and expressed inclusive of residual ash (method 6.5.1), ADF expressed inclusive of residual ash (method 6.5.2), and ADL determined by solubilization of cellulose with sulphuric acid (method 6.5.3). Moreover, microbial numbers of B. subtilis and B. licheniformis in assay diets were determined by method 28.2.2 [27]. Nitrogen contents in assay diets and ileal digesta samples were analyzed using a gas combustion method according to official method 990.03 of the AOAC International [28] (FP-2000, Leco Corp., St Joseph, MI, US). Ethylenediaminetetraacetic acid was used as a reference standard before and after all N analyses. Crude protein contents were calculated by multiplying the content of N with 6.25. Amino acid contents in assay diets and ileal digesta samples were determined by using ion-exchange chromatography with postcolumn derivatization with ninhydrin [29]. Tryptophan was determined by HPLC with fluorescence detection (extinction 280 nm, emission 356 nm), after alkaline hydrolysis with barium hydroxide octahydrate for 20 h at 110 °C according to the procedure as outlined by Commission Directive [30]. The titanium dioxide content in the assay diets and ileal digesta samples was performed according to the procedure described by Brandt and Allam [31].
DNA extraction of ileal digesta and feces samples
Genomic DNA was extracted from 250 mg ileal digesta and feces using Fast DNA Spin Kit for Soil (MP Biomedicals GmbH, Heidelberg, Germany). Extraction procedure was performed with slight modifications to manufacturer’s instructions as described by Burbach et al. [32].
Amplicon sequencing analysis
Illumina amplicon sequencing libraries of the V1-2 region of the 16S rRNA gene was performed similar to procedures described previously [33]. Library preparation, however, was modified as follows: the V1-2 region was amplified with a 27 F-modified forward primer (AGRGTTHGATYMTGGCTCAG) in a 20 μL reaction. 1 μL of this first PCR was used as template in a second PCR using multiplexing and indexing primers as described previously [33]. Amplicons were verified by agarose gel electrophoresis and normalized using SequalPrepTM Normalization Plate Kit (Invitrogen, Thermo Fisher Scientific, Waltham, USA). Libraries were pooled by index, purified with MinElute PCR Purification Kit (Qiagen, Hilden, Germany), quantified with Qubit® 2.0 Fluorometer (Invitrogen) and sequenced on Illumina MiSeq platform using 250 bp paired end sequencing chemistry. All analyzed samples comprised around 2.8 million reads, with an average of 43,646 reads per sample. Reads were quality filtered, assembled and aligned using Mothur pipeline [34]. UCHIME was used to find possible chimeras and reads were clustered at 97% identity into 2601 operative taxonomic units (OTU). The closest representative was manually identified with seqmatch from RDP [35]. Sequences classified as Chloroplast/Cyanobacteria were removed from OTU dataset as it was assumed that they represent undigested plant material. Sequences were submitted to European Nucleotide Archive under the accession number PRJEB14413 (http://www.ebi.ac.uk/ena/data/view/PRJEB14413).
Quantitative real time PCR
Quantitative real-time PCR (qPCR) was used to analyze the following bacteria groups in the ileal digesta samples: Total bacteria, Lactobacillus spp., Bifidobacterium spp., Roseburia spp., Enterobacteriaceae, Bacteroides-Prevotella-Porphyromonas group, Clostridium Cluster IV, and Bacillus spp.. All used primers were selected from literature and are listed in Table 2. Optimization of primer conditions was done in order to determine optimal annealing temperatures and primer concentrations by running a standard PCR with diverse primer concentrations (200 nmol/L, 400 nmol/L, 600 nmol/L) and a temperature gradient from 55.0 °C to 65.0 °C. According to melt curves on standard PCR and the agarose gel electrophoresis results, optimal primer concentration and annealing temperature was set for each primer.
Standard curves for each primer were designed using serial dilutions of the purified and quantified PCR products generated by standard PCR and genomic DNA from pig feces. The PCR products were checked by agarose gel electrophoresis (2% agarose) to ensure correct primer specific products. Quantity of purified PCR amplification products was determined using Qubit® 2.0 Fluorometer (Invitrogen).
Quantification was carried out using the CFX Connect™ Real-Time System (Bio-Rad Laboratories GmbH, Munich, Germany), associated with the Bio Rad CFX Manager™ Software 3.1 (Bio-Rad Laboratories GmbH, Munich, Germany). All samples were determined in duplicate and all standards were pipetted in triplicate on each plate. The order of samples and standards on the plates was randomized. The reaction mixture for each bacterial group consisted of 10 μL of KAPA SYBR FAST (PEQLAB Biotechnologie GmbH, Erlangen, Germany), 1 μL template DNA (ileal digesta samples and standards), the optimized primer concentrations of forward and reverse primers (Table 2), and was filled up to a total volume of 20 μL with PCR grade water (Carl Roth GmbH, Karlsruhe, Germany). Amplification conditions were: activation of polymerase at 95.0 °C for 3 min, followed by 40 cycles consisting of denaturation at 95.0 °C for 5 s, primer annealing for 20 s (at optimized temperatures, Table 2), and extension at 72.0 °C for 1 s. Subsequently, a final elongation step at 72.0 °C for 1 min followed. The melt curve was obtained by stepwise (0.5 °C) increase of temperature from 55 °C to 95 °C. Results were reported as log10 16S rRNA gene copies/g digesta.
Calculations
The AID of CP and AA in the assay diets was calculated according to the following equation:
where AIDD = AID of CP or AA in the assay diet (%), ID = marker content in the assay diet (g/kg DM), AI = CP or AA content in ileal digesta (g/kg DM), AD = CP or AA content in the assay diet (g/kg DM), and II = marker content in ileal digesta (g/kg DM).
According to Stein et al. [36] and Jansman et al. [37], the basal ileal endogenous loss of CP and AA (IAAend) is considered to be constant among groups of pigs, and therefore, mean values for IAAend [37] can be used for transformation of AID into their SID values.
The SID of CP and AA in assay diets was estimated according to the following equation:
where SIDD = SID of CP or AA in the assay diet (%).
Statistical analyses
Homogeneity of variances and normal distribution of the data were confirmed using the UNIVARIATE procedure of SAS (SAS Inst., Inc., Cary, NC). Data were analyzed as a 2 × 2 factorial using the GLIMMIX procedures of SAS. The model included the protein level, probiotic supplementation, and the interactive effects of protein level and probiotic supplementation as the fixed effects, and pig and period as the random effects. In case of interaction, the significant differences between treatments based on a t-test were set at α = 0.05 using the algorithm for letter-based representation of all pair-wise comparisons according to Piepho [38]. For microbiota analyses, bacterial 16S rRNA gene copy numbers in pre-treatment period was considered as covariate. Least squares means and standard error of the means are presented, and a probability level of P < 0.05 was considered to be statistically significant, whereas a P < 0.10 was considered to constitute a tendency.
Illumina amplicon sequencing data were analyzed using statistic software PRIMER (v.6.1.16, PRIMER-E; Plymouth Marine Laboratory, Plymouth, UK) [39]. Samples were standardized by total and resemblance matrix was calculated using Bray-Curtis coefficient. Overall community structures were explored by nonmetric multidimensional scaling (MDS). One way analysis of similarity (ANOSIM) and permutational multivariate analysis of variance (PERMANOVA) were used to evaluate similarity between different dietary groups, different protein levels and probiotic treatments, and a probability level of P ≤ 0.05 was considered to be significant different. The ANOSIM R values range from -1 to 1; the farer from zero the more distinct and the closer to zero the more similar are the compared groups. Variables contributing to observed differences were identified by similarity percentages routine. The bacterial families contributing to overall 70% of dissimilarities among treatment groups were considered to be the most important and their abundance data were graphically plotted according to a color key from zero to maximal abundance. Shannon index was used to measure diversity in bacterial communities from different sample groups, taking into account the number of OTUs and the proportion of each OTU. A Mantel-type test (RELATE) on Bray-Curtis matrices was used to quantify the correlation between results from bacterial community analysis. To enable comparison between amplicon sequencing and qPCR approaches, RELATE routine was run on untransformed datasets, restricted to bacteria groups targeted by qPCR primers and the generated Spearman Rho was considered to be significant if P ≤ 0.05.
Results
All pigs remained healthy throughout the experiment and readily consumed their daily feed allowances. The analyzed CP and AA contents of the assay diets and microbial numbers of B. subtilis and B. licheniformis in assay diets are presented in Table 3. As expected, CP and AA contents in LP were approximately 76.5 and 76.6% that of HP, respectively. The contents of ash, EE, NDF, ADF, and ADL in the HP diets were also greater than in the LP diets. The Bacillus spores determined in the experimental diets amounted to 1.54 × 109 CFU/kg feed for HP + and LP + diets, whereas HP - and LP - diets contained 0.02 × 109 and 0.04 × 109 CFU/kg feed, respectively.
The AID and SID of CP and AA in the assay diets are shown in Tables 4 and 5, respectively. The supplementation with Bacillus spp. did not affect both AID and SID of CP and AA. Furthermore, there was no difference in AID of CP and AA between HP and LP diets, but SID of cystine, glutamic acid, glycine, and proline was lower (P < 0.05) in the HP diets than in the LP diets. Moreover, SID of CP, alanine, aspartic acid, and serine also tended to be lower (P < 0.10) in the HP diets. However, no interactions between CP level and Bacillus spp. supplementation could be observed for AID and SID of CP and AA in the present study.
The overall structure in bacterial communities from ileal digesta was evaluated by 16S rRNA gene amplicon sequencing. Analysis of similarity revealed significant differences in microbiota composition due to different dietary treatments (P = 0.05), but a statistic R value close to zero (R = 0.176) suggests a weak separation of the different treatment groups (Fig. 1a).

Microbiota composition in ileal digesta samples from pigs fed starter diet and assay diets. a Multidimensional scaling plot based on Bray Curtis similarity matrix of 16S rDNA sequence data from ileal digesta. b Abundance plot of most important bacterial families in overall microbiota structure of ileal digesta. Phyla: Firmicutes (Fi), Bacteroidetes (Ba), Proteobacteria (Pr)
When compared with the starter period, bacterial communities were different (P < 0.01) between dietary treatments. Within assay diets, however, there were no effects (Table 6).
Taxonomical composition of ileal digesta samples demonstrated some variation among dietary treatments. At phylum level, the bacterial communities were dominated by Firmicutes and Bacteroidetes. Within the assay diets from periods 1 to 3, the relative abundance of Firmicutes was higher than Bacteroidetes when compared to the starter period. The reduction of Bacteroidetes was mainly due to lower abundance of Prevotellaceae, with an average abundance of 27% in the starter diet compared to 5% in the HP diets, 4% in LP - and 11% in LP +. Nine bacterial families contributed to the overall dissimilarities among microbiota structure in ileal digesta samples of different dietary treatments (Fig. 1b). Ileal microbiota from dietary treatments without probiotic supplementation consisted mainly of Peptostreptococcaceae, Clostridiaceae 1, Streptococcaceae, Lactobacillaceae and Erysipelotrichaceae with even proportions, except for Peptostreptococcaceae and Streptococcaceae being the predominant family in the HP and LP treatment, respectively. Streptococcus alactolyticus accounted for 15% of total microbiota in samples of LP - treatment. Compared to this, ileal digesta samples from LP + were enhanced in Clostridiaceae 1, Erysipelotrichaceae and Prevotellaceae. In HP +, the bacterial composition was dominated by Lactobacillaceae, with an average abundance of 40%. Here, an uncultured Lactobacillus from porcine intestine (relative abundance of 21.5%) and Lactobacillus amylovorus (14.2%) were the predominant species.
Lactobacillus spp. and other bacteria groups of interest were quantified in ileal digesta by qPCR (Table 7). Mantel test showed a significant correlation between the two approaches, sequencing and qPCR (Rho = 0.852, P < 0.01), thus confirming that both methodological approaches resulted in comparable results. The HP diets increased abundance of Lactobacillus spp. and Bifidobacterium spp. (P < 0.05). No effects of CP content on ileal gene copy numbers of total bacteria, Roseburia spp., Enterobacteriaceae, Bacteroides-Prevotella-Porphyromonas, Clostridium cluster IV and Bacillus spp. were found. Likewise, no significant effect of supplementation of Bacillus spp. was observed for ileal gene copy numbers of total bacteria, Lactobacillus spp., Bifidobacterium spp., Enterobacteriaceae, Clostridium cluster IV and Bacillus spp.. However, dietary supplementation of Bacillus spp. increased (P < 0.05) abundance of Roseburia spp., while it tended (P < 0.10) to promote Bacillus spp. and total bacteria. Furthermore, there was an interaction (P < 0.05) of protein level and Bacillus spp. supplementation for ileal gene copy numbers of Bacteroides-Prevotella-Porphyromonas. The LP + resulted in higher (P < 0.05) abundance of Bacteroides-Prevotella-Porphyromonas than the LP -, but did not differ from the HP diets.
The analysis of fecal microbiota by 16S rRNA gene amplicon sequencing showed no statistical effect on overall community structure. However, feces microbiota from each assay diet revealed to be significant different to that from the starter period (P < 0.01; Fig. 2a). At family level, Prevotellaceae exhibited the strongest impact on these dissimilarities (Fig. 2b), with Prevotella being the predominant genus. The average abundances of Prevotella showed slight variations for treatment groups with different protein levels; starter (15%), LP diets (13%), and HP diets (19%).

Microbiota composition in fecal samples from pigs fed starter diet and assay diets. a Multidimensional scaling plot based on Bray Curtis similarity matrix of 16S rDNA sequence data from fecal samples. b Abundance plot of most important bacterial families in overall microbiota structure of feces. Phyla: Firmicutes (Fi), Bacteroidetes (Ba), Spirochaetes (Sp)
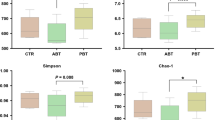
Comparing sequencing results from porcine ileal digesta and feces revealed distinct differences in bacterial communities structure (R = 0.924, P < 0.01) (Fig. 3a). Samples from ileal digesta showed a lower diversity compared to feces (Shannon index in average 2.9 vs. 4.7) (Fig. 3b and c). Mainly Streptococcus alactolyticus contributed to the dissimilarity with an average abundance of 9.7% in ileal digesta compared to 2.0% in feces. At family level differences were mainly due to Lactobacillaceae and Ruminococcaceae. The abundance of Lactobacillaceae was higher in ileal digesta (16%) than in feces (2%), and contrary the abundance of Ruminococcaceae was higher in feces (23%) than in ileal digesta (0.5%).

Comparison of microbiota from ileal digesta and feces. a MDS plot based on Bray-Curtis similarity matrix of all samples from ileal digesta and feces. b Shannon diversity calculated on operative taxonomic units data from ileal digesta samples (c) and from fecal samples
Discussion
According to previous studies, B. subtilis and B. licheniformis produce extracellular enzymes including proteases and α-amylase [19, 20], which may enhance nutrient digestibility resulting in improved feed conversion in finisher pigs [40]. In addition, B. subtilis exceeds B. licheniformis in production of glycosyl hydrolases [4], which assist in the hydrolysis of glycosidic bonds in complex sugars. However concerning antibiotic resistance, which is considered to be an important key requirement for probiotics, a higher concentration of antibiotics is tolerated by B. licheniformis than by B. subtilis [4]. Recently, probiotic characteristics were described for spores of B. subtilis, although interactions with porcine epithelial cells are not understood so far [4]. For example, the supplementation of B. subtilis to a soybean meal diet showed slight improvements in AID and SID of some AA in weaning pigs as described by Kim et al. [21]. However, in the present study, there was no improvement in AID and SID of CP and AA in growing pigs fed diets supplemented with B. subtilis and B. licheniformis. Similarly, previous studies [6, 41] failed to demonstrate that the inclusion of B. subtilis and B. licheniformis in diets would affect apparent total tract digestibility of CP in growing-finishing pigs. The lack of probiotic treatment effects may be due to low quantity of the supplemented bacterial species in porcine intestine, as in treatments with probiotic supplementation the Bacillus spp. numbers were not significantly higher compared to numbers in treatments without probiotic supplementation. The gene copy numbers of Bacillus spp. in the treatments without probiotic supplementation correspond to results of a study by Dowd et al. [42] on Bacillus spp. in the ileum of piglets using 16S rRNA gene sequencing. In addition to the qPCR results, further Bacillus species (B. pumilus and B. cereus) were identified by amplicon sequencing. Operative taxonomic units corresponding to Bacillus genus appeared in very low abundance (<1%), and were present in samples with and without Bacillus spp. supplementation. These results are in accordance with previous studies demonstrating the ability of germinated Bacillus spores to proliferate in mammal GIT, even if only at a low rate [5], and therefore might not be persistent [43].
Positive effects of diets supplemented with B. subtilis and B. licheniformis on feed conversion in pigs have been reported before [40, 44], however, the underlying mechanisms of Bacillus’ probiotic activity are little understood, and may be attributed to competitive adhesion and immunomodulation by Bacillus spores or to enzymes and other substances produced by the germinated, vegetative cells of Bacillus [5]. Notably, probiotic supplements may be more effective under stress such as practical field conditions [45, 46]. This might be one reason for the missing effect of Bacillus spp. supplementation on digestibility values in the present study, as pigs were individually housed and kept in a clean environment under optimal temperature and minimal stress conditions. Furthermore, the age of pigs may be associated with probiotic efficacy [47]. The use of probiotics tended to be more effective in early age of pigs rather than the growing period [48, 49]. In the present study, grower pigs (13- and 20-week old at the initial and final BW, respectively) fed diets supplemented with Bacillus spp. did not show any differences in ileal digestibility of CP and AA. It has been suggested that increasing age may be a contributing factor in building up the complexity of the microbial community [50] with growing pigs being more resistant to intestinal disorders than young pigs [51].
Dietary content of CP has been reported to be associated with AID due to the variation in endogenous CP and AA losses in ileal digesta [52]. Previous research [53] suggests that AID shows segmented quadratic with plateau relationships as the level of CP and AA in the diet increased from 4 to 24% (as-fed). Alternatively, SID has been widely accepted to overcome this problem by correcting AID values for basal endogenous losses of CP and AA [54]. In general, SID values are higher in comparison to their corresponding AID values as the basal endogenous losses of CP and AA are subtracted from ileal CP and AA outflow [36]. In the present study, SID of some AA was lower in HP diets than in LP diets. Apparently, higher fiber contents in HP diets, associated with enhanced secretion of endogenous AA [55, 56], may have contributed to higher rate of digesta passage in the digestive tract of pigs [57], thereby, decreasing SID values. This is confirmed by the results of a recent study [58], where SID of CP and most AA decreased linearly with increasing dietary CP from 6.8 to 21.4% (as-fed) due to the greater NDF and ADF contents.
The higher numbers of Lactobacillus spp. and Bifidobacterium spp. in ileal digesta of HP treatments are in agreement with a recent study by Rist et al. [17], where piglets fed high dietary CP levels showed an increased growth and proliferation of lactic-acid bacteria in ileal digesta. As content of soybean meal in the present study was greater in HP than in LP diets, enhanced availability of fermentable carbohydrates in the small intestine can be suggested, thereby stimulating ileal growth of lactobacilli and bifidobacteria [17]. Furthermore, HP diets could increase the availability of free AA in the small intestine [17], contributing much more preformed AA of dietary and endogenous origin to bacterial growth in the upper part of the digestive tract than microbial de novo synthesis of AA [13]. Furthermore, analysis of overall microbiota composition in ileal digesta by amplicon sequencing supported an increasing effect on Lactobacillus proportion upon feeding of HP + diets. The presence of Lactobacillus spp. and Bifidobacterium spp. in the GIT has been reported to be beneficial for the host animal [17] due to their ability for bacteriocin production [59]. Moreover, proliferation of pathogenic bacteria may be inhibited through the production of short-chain fatty acids (SCFA) and lactic acid, being associated with a lower pH, causing a hostile environment for some acid-sensitive bacteria strains [60]. The presented sequencing results for Lactobacillus spp. are supported by qPCR results, which revealed a higher number of Lactobacillus gene copies in HP diets. The identified Lactobacillus spp. were dominated by an uncultured bacterium, previously isolated from porcine intestine [61], and the species L. amylovorus. L. amylovorus is a synonym expression for Lactobacillus sobrius, which is characterized by amylolytic activity, and being previously identified with high prevalence in porcine intestine [62–65]. Application of an oral probiotic mixture including a L. amylovorus strain has been shown to promote growth performance of pigs [66]. In general, the enhancement of potential beneficial Lactobacillus spp. is considered to promote gut health. However, the above described supporting effect of HP + diet on abundance of Lactobacillus caused a reduced community diversity compared to microbiota from ileal digesta of other dietary treatments. A high diversity in intestinal microbiota might be preferable to cope effectively with potential challenging conditions [67].
Regardless of dietary protein level, the supplementation of B. subtilis and B. licheniformis had a stimulating effect on targeted quantity of Roseburia spp., known as an important butyrate producer [68]. Butyrate represents the most preferential energy source of colonocytes [69], resulting in the stimulation of epithelial cell proliferation and mucus secretion [70]. Therefore, the supplementation of B. subtilis and B. licheniformis may contribute to an improved gut health of pigs.
Assay diets did not significantly impact overall microbiota, but influence was demonstrated for bacterial copy numbers of Bacteroides-Prevotella-Porphyromonas. In the present study, the dietary CP level and the supplementation of B. subtilis and B. licheniformis showed an interaction, as supplementation of B. subtilis and B. licheniformis increased Bacteroides-Prevotella-Porphyromonas in the LP diets when compared to HP diets. The Bacteroides-Prevotella-Porphyromonas group includes phylogenetic related species from Bacteroidetes phylum that commonly inhabit GIT. Sequencing results confirmed an increased abundance of Prevotella in ileal digesta from LP + treatment when compared to the other assay diets. This finding is in agreement with other studies, which showed an enhancing effect of low protein diets on gene copy numbers of Bacteroides-Prevotella-Porphyromonas group in ileal digesta [17], and a significant increase in the abundance of Prevotella genus in cecum [71] when compared to samples of treatments with a higher protein level [17, 71]. Prevotella dominate the porcine fecal metagenome [72], play an important role in intestinal carbohydrate fermentation [73] and also show proteolytic activity [74]. Sequencing results also revealed members of Prevotella as main discriminators of community structure from ileal microbiota of starter and experimental periods. The observed decrease over experimental time is in agreement with a longitudinal study of Kim et al. [75]. Thus, the observations on relative proportion of Prevotella represent the general impact of diet and age on porcine intestinal microbiota. Contrary to ileal digesta, where abundance of Prevotella was highest in LP +, the fecal proportion of Prevotella was higher in HP than LP treatment and slightly increased over experimental time. This variation along sampling sites is in agreement with a previous study, investigating as well ileal digesta and fecal samples from pigs [17], where abundance patterns of Prevotella species in the GIT of pigs were different between ileal digesta and fecal samples.
The results of this study demonstrate an overall lower bacterial diversity for ileal digesta compared with fecal samples. Metagenome studies on porcine microbiota collected from different intestine sites revealed different contributions of bacterial species and activities along the GIT [76, 77]. The fecal collection is an easy accessible sampling site with samples showing high similarity to microbiota composition from proximal intestine. However, microbiota composition from fecal samples is not identical representatives to those from ileal digesta. Therefore, collecting samples of different sites of the GIT, where close interactions between the microbiome and the digestive processes occur, will improve understanding of probable functional changes and the effects of dietary treatments such as the addition of probiotics.
Undigested dietary components passing into the large intestine are subjected to fermentation by the intestinal microbiota [17]. As a result, fermentation products such as SCFA are rapidly absorbed across the gut wall, contributing up to 30% of growing pigs’ maintenance requirement for energy [78]. On the other hand, increasing protein fermentation may result in the formation of detrimental fermentation products such as ammonia and amines in the colon [79]. A lower dietary protein level may reduce ammonia production, as observed by Htoo et al. [14] in cecal samples of pigs, while supplementation of diets with B. subtilis and B. licheniformis showed similar results in slurry samples from pigs due to a lowering effect on the pH [6]. Therefore, LP diets supplemented with B. subtilis and B. licheniformis might be used to reduce the production of harmful microbial metabolites in the large intestine of pigs.
Conclusions
Supplementation with Bacillus spp. did not affect both AID and SID of CP and AA in growing pigs. The higher SID of some AA in the LP diets when compared to HP diets hints towards the possibility of reducing N excretion through diet manipulation. Regarding microbiota, the assay diets had no significant effect on overall community structure, neither in ileal digesta nor feces. Nevertheless, dietary protein content and Bacillus spp. supplementation may enhance various community members in ileal digesta. Within this regard, feeding of the HP diet resulted in a higher abundance of Lactobacillus spp. and Bifidobacterium spp., whereas LP diet may support bacteria important for carbohydrate degradation such as Prevotella. Furthermore, relative proportion of Prevotella was altered during pig’s age. The supplementation of Bacillus spp. promoted gene copy numbers of Roseburia spp., which may be beneficial due to ascribed health promoting properties of this butyrate producer, and this phenomenon may be more effective under stress condition. The LP diet supplemented with B. subtilis and B. licheniformis may be used as an alternative feeding strategy to support gut health in pigs.
Abbreviations
- AA:
-
Amino acid
- ADF:
-
Acid detergent fiber
- ADL:
-
Acid detergent lignin
- AID:
-
Apparent ileal digestibility
- ANOSIM:
-
Analysis of similarity
- BW:
-
Body weight
- CFU:
-
Colony forming units
- CP:
-
Crude protein
- DM:
-
Dry matter
- GIT:
-
Gastrointestinal tract
- HP:
-
High-protein diet
- LP:
-
Low-protein diet
- ME:
-
Metabolizable energy
- N:
-
Nitrogen
- NDF:
-
Neutral detergent fiber
- OTU:
-
Operativ taxonomic unit
- PERMANOVA:
-
Permutational multivariate analysis of variance
- qPCR:
-
Quantitative real-time PCR
- RELATE:
-
Mantel type test
- SCFA:
-
Short chain fatty acids
- SID:
-
Standardized ileal digestibility
References
Barton MD. Impact of antibiotic use in the swine industry. Curr Opin Microbiol. 2014;19:9–15. doi:10.1016/j.mib.2014.05.017 .
Chen YJ, Min BJ, Cho JH, Kwon OS, Son KS, Kim HJ, et al. Effects of dietary Bacillus-based probiotic on growth performance, nutrients digestibility, blood characteristics and fecal noxious gas content in finishing pigs. Asian Australas J Anim Sci. 2006;19:587–92. doi:10.5713/ajas.2006.587 .
Meng QW, Yan L, Ao X, Zhou TX, Wang JP, Lee JH, et al. Influence of probiotics in different energy and nutrient density diets on growth performance, nutrient digestibility, meat quality, and blood characteristics in growing-finishing pigs. J Anim Sci. 2010;88:3320–6. doi:10.2527/jas.2009-2308 .
Larsen N, Thorsen L, Kpikpi EN, Stuer-Lauridsen B, Cantor MD, Nielsen B, et al. Characterization of Bacillus spp. strains for use as probiotic additives in pig feed. Appl Microbiol Biotechnol. 2014;98:1105–18. doi:10.1007/s00253-013-5343-6 .
Leser TD, Knarreborg A, Worm J. Germination and outgrowth of Bacillus subtilis and Bacillus licheniformis spores in the gastrointestinal tract of pigs. J Appl Microbiol. 2008;104:1025–33. doi:10.1111/j.1365-2672.2007.03633.x .
Wang Y, Cho JH, Chen YJ, Yoo JS, Huang Y, Kim HJ, et al. The effect of probiotic BioPlus 2B® on growth performance, dry matter and nitrogen digestibility and slurry noxious gas emission in growing pigs. Livest Sci. 2009;120:35–42. doi:10.1016/j.livsci.2008.04.018 .
EFSA (European Food Safety Authority). Scientific Opinion on the maintenance of the list of QPS biological agents intentionally added to food and feed (2010 update). EFSA J. 2010;8:1944. doi:10.2903/j.efsa.2010.1944 .
Degering C, Eggert T, Puls M, Bongaerts J, Evers S, Maurer KH, et al. Optimization of protease secretion in Bacillus subtilis and Bacillus licheniformis by screening of homologous and heterologous signal peptides. Appl Environ Microbiol. 2010;76:6370–6. doi:10.1128/AEM.01146-10 .
Upadhaya SD, Kim SC, Valientes RA, Kim IH. The effect of Bacillus-based feed additive on growth performance, nutrient digestibility, fecal gas emission, and pen cleanup characteristics of growing-finishing pigs. Asian Australas J Anim Sci. 2015;28:999–1005. doi:10.5713/ajas.15.0066 .
Jørgensen JN, Laguna JS, Millán C, Casabuena O, Gracia MI. Effects of a Bacillus-based probiotic and dietary energy content on the performance and nutrient digestibility of wean to finish pigs. Anim Feed Sci Technol. 2016;221:54–61. doi:10.1016/j.anifeedsci.2016.08.008 .
Mosenthin R, Hambrecht E, Sauer WC. Utilisation of different fibres in piglet feeds. In: Garnsworthy PC, Wiseman J, editors. Recent advances in animal nutrition. Nottingham: Nottingham University Press; 1999. p. 227–56.
Wellock IJ, Fortomaris PD, Houdijk JGM, Kyriazakis I. The effect of dietary protein supply on the performance and risk of post-weaning enteric disorders in newly weaned pigs. Anim Sci. 2006;82:327–35. doi:10.1079/ASC200643 .
Libao-Mercado AJO, Zhu CL, Cant JP, Lapierre H, Thibault JN, Sève B, et al. Dietary and endogenous amino acids are the main contributors to microbial protein in the upper gut of normally nourished pigs. J Nutr. 2009;139:1088–94. doi:10.3945/jn.108.103267 .
Htoo JK, Araiza BA, Sauer WC, Rademacher M, Zhang Y, Cervantes M, et al. Effect of dietary protein content on ileal amino acid digestibility, growth performance, and formation of microbial metabolites in ileal and cecal digesta of early-weaned pigs. J Anim Sci. 2007;85:3303–12. doi:10.2527/jas.2007-0105 .
Ferket PR, van Heugten E, van Kempen TATG, Angel R. Nutritional strategies to reduce environmental emissions from nonruminants. J Anim Sci. 2002;80(E-Suppl 2):E168–82. doi:10.2527/animalsci2002.80E-Suppl_2E168x .
Lee JH, Kim JH, Kim JD, Kim SW, Han IK. Effects of low crude protein diets supplemented with synthetic amino acids on performance, nutrient utilization and carcass characteristics in finishing pigs reared using a phase feeding regimen. Asian Australas J Anim Sci. 2001;14:655–67. doi:10.5713/ajas.2001.655 .
Rist VTS, Weiss E, Sauer N, Mosenthin R, Eklund M. Effect of dietary protein supply originating from soybean meal or casein on the intestinal microbiota of piglets. Anaerobe. 2014;25:72–9. doi:10.1016/j.anaerobe.2013.10.003 .
Bhandari SK, Opapeju FO, Krause DO, Nyachoti CM. Dietary protein level and probiotic supplementation effects on piglet response to Escherichia coli K88 challenge: Performance and gut microbial population. Livest Sci. 2010;133:185–8. doi:10.1016/j.livsci.2010.06.060 .
Priest FG. Extracellular enzyme synthesis in the genus Bacillus. Bacteriol Rev. 1977;41:711–53.
Carlisle GE, Falkinham III JO. Enzyme activities and antibiotic susceptibility of colonial variants of Bacillus subtilis and Bacillus licheniformis. Appl Environ Microbiol. 1989;55:3026–8.
Kim DH, Heo PS, Jang JC, Jin SS, Hong JS, Kim YY. Effect of different soybean meal type on ileal digestibility of amino acid in weaning pigs. J Anim Sci Technol. 2015;57:11. doi:10.1186/s40781-015-0041-9 .
Council Directive. Council Directive of 24 November 1986 on the approximation of laws, regulations and administrative provisions of the Member States regarding the protection of animals used for experimental and other scientific purposes (86/609/EEC). Off J Eur Union. 1986;L 358:1–28.
Li S, Sauer WC, Fan MZ. The effect of dietary crude protein level on ileal and fecal amino acid digestibility in early-weaned pigs. J Anim Physiol Anim Nutr. 1993;70:117–28. doi:10.1111/j.1439-0396.1993.tb00314.x .
NRC. Nutrient requirements of swine. 10th ed. Washington: National Academies Press; 1998.
Fan MZ, Sauer WC, Hardin RT, Lien KA. Determination of apparent ileal amino acid digestibility in pigs: effect of dietary amino acid level. J Anim Sci. 1994;72:2851–9. doi:10.2527/1994.72112851x .
NRC. Nutrient requirements of swine. 11th ed. Washington: National Academies Press; 2012.
Verband Deutscher Landwirtschaftlicher Untersuchungs- und Forschungsanstalten (VDLUFA). Handbuch der Landwirtschaftlichen Versuchs- und Untersuchungsmethodik (VDLUFA-Methodenbuch), Bd. III Die chemische Untersuchung von Futtermitteln mit 1. – 8. Ergänzungslieferung (1983 – 2012). (In German). 3rd ed. Darmstadt: VDLUFA-Verlag; 1976.
AOAC. Official methods of analysis. 17th ed. Gaithersberg: AOAC International; 2000.
Llames CR, Fontaine J. Determination of amino acids in feeds: collaborative study. J Assoc Off Anal Chem. 1994;77:1362–402.
Commission Directive. Commission Directive 2000/45/EC of 6 July 2000 on establishing community methods of analysis for the determination of vitamin A, vitamin E and tryptophan in feedingstuffs. Annex part C. Determination of tryptophan. Off J Eur Union. 2000;L 174:32–50.
Brandt M, Allam SM. Analytik von TiO2 im Darminhalt und Kot nach Kjeldahlaufschluß. Arch Anim Nutr. 1987;37:453–4.
Burbach K, Seifert J, Pieper DH, Camarinha-Silva A. Evaluation of DAN extraction kits and phylogenetic diversity of the porcine gastrointestinal tract based on Illumina sequencing of two hypervariable regions. Microbiologyopen. 2016;5:70–82. doi:10.1002/mbo3.312 .
Camarinha-Silva A, Jáuregui R, Chaves-Moreno D, Oxley APA, Schaumburg F, Becker K, et al. Comparing the anterior nare bacterial community of two discrete human populations using Illumina amplicon sequencing. Environ Microbiol. 2014;16:2939–52. doi:10.1111/1462-2920.12362 .
Schloss PD, Westcott SL, Ryabin T, Hall JR, Hartmann M, Hollister EB, et al. Introducing mothur: open-source, platform-independent, community-supported software for describing and comparing microbial communities. Appl Environ Microbiol. 2009;75:7537–41. doi:10.1128/AEM.01541-09 .
Wang Q, Garrity GM, Tiedje JM, Cole JR. Naïve Bayesian classifier for rapid assignment of rRNA sequences into the new bacterial taxonomy. Appl Environ Microbiol. 2007;73:5261–7. doi:10.1128/AEM.00062-07 .
Stein HH, Sève B, Fuller MF, Moughan PJ, de Lange CFM. Invited review: amino acid bioavailability and digestibility in pig feed ingredients: terminology and application. J Anim Sci. 2007;85:172–80. doi:10.2527/jas.2005-742 .
Jansman AJM, Smink W, van Leeuwen P, Rademacher M. Evaluation through literature data of the amount and amino acid composition of basal endogenous crude protein at the terminal ileum of pigs. Anim Feed Sci Technol. 2002;98:49–60. doi:10.1016/S0377-8401(02)00015-9 .
Piepho H-P. A SAS macro for generating letter displays of pairwise mean comparisons. CBCS. 2012;7:4–13.
Clarke KR, Warwick RM. Change in marine communities: an approach to statistical analysis and interpretation. Plymouth: PRIMER-E Ltd; 2001.
Davis ME, Parrott T, Brown DC, de Rodas BZ, Johnson ZB, Maxwell CV, et al. Effect of a Bacillus-based direct-fed microbial feed supplement on growth performance and pen cleaning characteristics of growing-finishing pigs. J Anim Sci. 2008;86:1459–67. doi:10.2527/jas.2007-0603 .
Kornegay ET, Risley CR. Nutrient digestibilities of a corn-soybean meal diet as influenced by Bacillus products fed to finishing swine. J Anim Sci. 1996;74:799–805. doi:10.2527/1996.744799x .
Dowd SE, Sun Y, Wolcott RD, Domingo A, Carroll JA. Bacterial tag-encoded FLX amplicon pyrosequencing (bTEFAP) for microbiome studies: bacterial diversity in the ileum of newly weaned Salmonella-infected pigs. Foodborne Pathog Dis. 2008;5:459–72. doi:10.1089/fpd.2008.0107 .
Tam NKM, Uyen NQ, Hong HA, Le Duc H, Hoa TT, Serra CR, et al. The intestinal life cycle of Bacillus subtilis and close relatives. J Bacteriol. 2006;188:2692–700. doi:10.1128/JB.188.7.2692-2700.2006 .
Alexopoulos C, Georgoulakis IE, Tzivara A, Kyriakis CS, Govaris A, Kyriakis SC. Field evaluation of the effect of a probiotic-containing Bacillus licheniformis and Bacillus subtilis spores on the health status, performance, and carcass quality of grower and finisher pigs. J Vet Med. 2004;A 51:306–12. doi:10.1111/j.1439-0442.2004.00637.x .
Shon KS, Hong JW, Kwon OS, Min BJ, Lee WB, Kim IH, et al. Effects of Lactobacillus reuteri-based direct-fed microbial supplementation for growing-finishing pigs. Asian Australas J Anim Sci. 2005;18:370–4. doi:10.5713/ajas.2005.370 .
Weiss E, Eklund M, Semaskaite A, Urbaityte R, Metzler-Zebeli B, Sauer N, et al. Combinations of feed additives affect ileal fibre digestibility and bacterial numbers in ileal digesta of piglets. Czech J Anim Sci. 2013;58:351–9.
Lessard M, Brisson GJ. Effect of a Lactobacillus fermentation product on growth, immune response and fecal enzyme activity in weaned pigs. Can J Anim Sci. 1987;67:509–16. doi:10.4141/cjas87-049 .
Pollman DS, Danielson DM, Peo Jr ER. Effects of microbial feed additives on performance of starter and growing-finishing pigs. J Anim Sci. 1980;51:557–81.
Link R, Kováč G. The effect of probiotic BioPlus 2B on feed efficiency and metabolic parameters in swine. Biologia. 2006;61:783–7. doi:10.2478/s11756-006-0158-x .
Baker AA, Davis E, Spencer JD, Moser R, Rehberger T. The effect of a Bacillus-based direct-fed microbial supplemented to sows on the gastrointestinal microbiota of their neonatal piglets. J Anim Sci. 2013;91:3390–9. doi:10.2527/jas2012-5821.
Nousiainen J, Setälä J. Lactic acid bacteria as animal probiotics. In: Salminen S, von Wright A, editors. Lactic acid bacteria. New York: Marcel Dekker; 1993. p. 315–56.
Eklund M, Mosenthin R, Piepho H-P, Rademacher M. Estimates of dietary threshold levels for crude protein and amino acids to obtain plateau values of apparent ileal crude protein and amino acid digestibilities in newly weaned pigs. Arch Anim Nutr. 2010;64:357–72. doi:10.1080/1745039X.2010.492139 .
Fan MZ, Sauer WC, McBurney MI. Estimation by regression analysis of endogenous amino acid levels in digesta collected from the distal ileum of pigs. J Anim Sci. 1995;73:2319–28. doi:10.2527/1995.7382319x .
Eklund M, Mosenthin R, Piepho H-P, Rademacher M. Effect of dietary crude protein level on basal ileal endogenous losses and standardized ileal digestibilities of crude protein and amino acids in newly weaned pigs. J Anim Physiol Anim Nutr. 2008;92:578–90. doi:10.1111/j.1439-0396.2007.00751.x.
Grala W, Verstegen MWA, Jansman AJM, Huisman J, van Leeusen P. Ileal apparent protein and amino acid digestibilities and endogenous nitrogen losses in pigs fed soybean and rapeseed products. J Anim Sci. 1998;76:557–68. doi:10.2527/1998.762557x.
Jondreville C, van den Broecke J, Gâtel F, Grosjean F, Van Cauwenberghe S, Sève B. Ileal amino acid digestibility and estimates of endogenous amino acid losses in pigs fed rapeseed meal, sunflower meal and soybean meal. Can J Anim Sci. 2000;80:495–506. doi:10.4141/A99-104 .
Calvert CC. Fiber utilization by swine. In: Miller ER, Ullrey DE, Lewis AJ, editors. Swine nutrition. Massachusetts: Butterworth-Heinemann; 1991. p. 285–96.
Zhai H, Adeola O. Apparent and standardized ileal digestibilities of amino acids for pigs fed corn- and soybean meal-based diets at varying crude protein levels. J Anim Sci. 2011;89:3626–33. doi:10.2527/jas.2010-3732 .
Vuotto C, Longo F, Donelli G. Probiotics to counteract biofilm-associated infections: promising and conflicting data. Int J Oral Sci. 2014;6:189–94. doi:10.1038/ijos.2014.52 .
Kleerebezem M, Vaughan EE. Probiotic and gut lactobacilli and bifidobacteria: molecular approaches to study diversity and activity. Annu Rev Microbiol. 2009;63:269–90. doi:10.1146/annurev.micro.091208.073341 .
Leser TD, Amenuvor JZ, Jensen TK, Lindecrona RH, Boye M, Møller K. Culture-independent analysis of gut bacteria: the pig gastrointestinal tract microbiota revisited. Appl Environ Microbiol. 2002;68:673–90. doi:10.1128/AEM.68.2.673-690.2002 .
Jakava-Viljanen M, Murros A, Palva A, Björkroth KJ. Lactobacillus sobrius Konstantinov et al. 2006 is a later synonym of Lactobacillus amylovorus Nakamura 1981. Int J Syst Evol Microbiol. 2008;58:910–3. doi:10.1099/ijs.0.65432-0 .
Nakamula LK. Lactobacillus amylovorus, a new starch-hydrolyzing species from cattle waste-corn fermentations. Int J Syst Evol Microbiol. 1981;31:56–63. doi:10.1099/00207713-31-1-56 .
Mann E, Schmitz-Esser S, Zebeli Q, Wagner M, Ritzmann M, Metzler-Zebeli BU. Mucosa-associated bacterial microbiome of the gastrointestinal tract of weaned pigs and dynamics linked to dietary calcium-phosphorus. PLoS One. 2014;9:e86950. doi:10.1371/journal.pone.0086950 .
Konstantinov SR, Awati AA, Williams BA, Miller BG, Jones P, Stokes CR, et al. Post-natal development of the porcine microbiota composition and activities. Environ Microbiol. 2006;8:1191–9. doi:10.1111/j.1462-2920.2006.01009.x .
Ross GR, Gusils C, Oliszewski R, de Holgado SC, González SN. Effects of probiotic administration in swine. J Biosci Bioeng. 2010;109:545–9. doi:10.1016/j.jbiosc.2009.11.007 .
Lozupone CA, Stombaugh JI, Gordon JI, Jansson JK, Knight R. Diversity, stability and resilience of the human gut microbiota. Nature. 2012;489:220–30. doi:10.1038/nature11550 .
Flint HJ, Scott KP, Louis P, Duncan SH. The role of the gut microbiota in nutrition and health. Nat Rev Gastroenterol Hepatol. 2012;9:577–89. doi:10.1038/nrgastro.2012.156 .
Roediger WEW. Utilization of nutrients by isolated epithelial cells of the rat colon. Gastroenterology. 1982;83:424–9.
Tsukahara T, Hashizume K, Koyama H, Ushida K. Stimulation of butyrate production through the metabolic interaction among lactic acid bacteria, Lactobacillus acidophilus, and lactic acid-utilizing bacteria, Megasphaera elsdenii, in porcine cecal digesta. Anim Sci J. 2006;77:454–61. doi:10.1111/j.1740-0929.2006.00372.x .
Zhou L, Fang L, Sun Y, Su Y, Zhu W. Effects of the dietary protein level on the microbial composition and metabolomic profile in the hindgut of the pig. Anaerobe. 2016;38:61–9. doi:10.1016/j.anaerobe.2015.12.009 .
Lamendella R, Domingo JWS, Ghosh S, Martinson J, Oerther DB. Comparative fecal metagenomics unveils unique functional capacity of the swine gut. BMC Microbiol. 2011;11:103. doi:10.1186/1471-2180-11-103 .
De Filippo C, Cavalieri D, Di Paola M, Ramazzotti M, Poullet JB, Massart S, et al. Impact of diet in shaping gut microbiota revealed by a comparative study in children from Europe and rural Africa. Proc Natl Acad Sci U S A. 2010;107:14691–6. doi:10.1073/pnas.1005963107 .
Purushe J, Fouts DE, Morrison M, White BA, Mackie RI, the North American Consortium for Rumen Bacteria, et al. Comparative genome analysis of Prevotella ruminicola and Prevotella bryantii: insights into their environmental niche. Microb Ecol. 2010;60:721–9. doi:10.1007/s00248-010-9692-8 .
Kim HB, Borewicz K, White BA, Singer RS, Sreevatsan S, Tu ZJ, et al. Longitudinal investigation of the age-related bacterial diversity in the feces of commercial pigs. Vet Microbiol. 2011;153:124–33. doi:10.1016/j.vetmic.2011.05.021 .
Looft T, Allen HK, Cantarel BL, Levine UY, Bayles DO, Alt DP, et al. Bacteria, phages and pigs: the effects of in-feed antibiotics on the microbiome at different gut locations. ISME J. 2014;8:1566–76. doi:10.1038/ismej.2014.12 .
Zhao W, Wang Y, Liu S, Huang J, Zhai Z, He C, et al. The dynamic distribution of porcine microbiota across different ages and gastrointestinal tract segments. PLoS One. 2015;10:e0117441. doi:10.1371/journal.pone.0117441 .
Varel VH, Yen JT. Microbial perspective on fiber utilization by swine. J Anim Sci. 1997;75:2715–22. doi:10.2527/1997.75102715x .
Macfarlane GT, Gibson GR, Beatty E, Cummings JH. Estimation of short-chain fatty acid production from protein by human intestinal bacteria based on branched-chain fatty acid measurements. FEMS Microbiol Lett. 1992;101:81–8. doi:10.1111/j.1574-6968.1992.tb05764.x .
Fuller Z, Louis P, Mihajlovski A, Rungapamestry V, Ratcliffe B, Duncan AJ. Influence of cabbage processing methods and prebiotic manipulation of colonic microflora on glucosinolate breakdown in man. Br J Nutr. 2007;98:364–72. doi:10.1017/S0007114507709091 .
Malinen E, Kassinen A, Rinttilä T, Palva A. Comparison of real-time PCR with SYBR Green I or 5′-nuclease assays and dot-blot hybridization with rDNA-targeted oligonucleotide probes in quantification of selected faecal bacteria. Microbiology. 2003;149:269–77. doi:10.1099/mic.0.25975-0 .
Rinttilä T, Kassinen A, Malinen E, Krogius L, Palva A. Development of an extensive set of 16S rDNA-targeted primers for quantification of pathogenic and indigenous bacteria in faecal samples by real-time PCR. J Appl Microbiol. 2004;97:1166–77. doi:10.1111/j.1365-2672.2004.02409.x .
Veiga P, Gallini CA, Beal C, Michaud M, Delaney ML, DuBois A, et al. Bifidobacterium animalis subsp. lactis fermented milk product reduces inflammation by altering a niche for colitogenic microbes. Proc Natl Acad Sci U S A. 2010;107:18132–7. doi:10.1073/pnas.1011737107 .
Bartosch S, Fite A, Macfarlane GT, McMurdo MET. Characterization of bacterial communities in feces from healthy elderly volunteers and hospitalized elderly patients by using real-time PCR and effects of antibiotic treatment on the fecal microbiota. Appl Environ Microbiol. 2004;70:3575–81. doi:10.1128/AEM.70.6.3575-3581.2004 .
Ramirez-Farias C, Slezak K, Fuller Z, Duncan A, Holtrop G, Louis P. Effect of inulin on the human gut microbiota: stimulation of Bifidobacterium adolescentis and Faecalibacterium prausnitzii. Br J Nutr. 2009;101:541–50. doi:10.1017/S0007114508019880 .
Lay C, Sutren M, Rochet V, Saunier K, Doré J, Rigottier-Gois L. Design and validation of 16S rRNA probes to enumerate members of the Clostridium leptum subgroup in human faecal microbiota. Environ Microbiol. 2005;7:933–46. doi:10.1111/j.1462-2920.2005.00763.x .
Fernández-No IC, Guarddon M, Böhme K, Cepeda A, Calo-Mata P, Barros-Velázquez J. Detection and quantification of spoilage and pathogenic Bacillus cereus, Bacillus subtilis and Bacillus licheniformis by real-time PCR. Food Microbiol. 2011;28:605–10. doi:10.1016/j.fm.2010.10.014.
Acknowledgements
The authors would like to thank S. Stabenow for taking care of pigs and M. Steffl and N. Nautscher for their excellent work with animal surgery. The support of M. Eklund in designing this study is acknowledged.
Funding
This research was financially supported by Chr. Hansen A/S (Hørsholm, Denmark) and the Foundation fiat panis (Ulm, Germany). The receipt of a scholarship for Chanwit Kaewtapee granted by Food Security Center (FSC), University of Hohenheim under the German Academic Exchange Service (DAAD) program exceed with funds of the Federal Ministry for Economic Cooperation and Development (BMZ) of Germany is gratefully acknowledged.
Availability of data and materials
All data generated or analyzed during this study are available from the corresponding author on reasonable request.
Authors’ contributions
CK, KB, PRK, SH, JS and RM conceived the study, performed the statistics and drafted the manuscript. KB, GT and TH participated in the animal care and digesta collection. KB, ACS, TH and SH conducted the DNA extraction and quantitative real-time PCR. MW analyzed contents of CP and AA in diets and ileal digesta. All authors read and approved the final manuscript.
Competing interests
The authors declare that they have no competing interests.
Consent for publication
Not applicable.
Ethics approval and consent to participate
The research protocol was approved by the German Ethical Commission for Animal Welfare.
Author information
Authors and Affiliations
Corresponding author
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made. The Creative Commons Public Domain Dedication waiver (http://creativecommons.org/publicdomain/zero/1.0/) applies to the data made available in this article, unless otherwise stated.
About this article

Cite this article
Kaewtapee, C., Burbach, K., Tomforde, G. et al. Effect of Bacillus subtilis and Bacillus licheniformis supplementation in diets with low- and high-protein content on ileal crude protein and amino acid digestibility and intestinal microbiota composition of growing pigs. J Animal Sci Biotechnol 8, 37 (2017). https://doi.org/10.1186/s40104-017-0168-2
Received:
Accepted:
Published:
DOI: https://doi.org/10.1186/s40104-017-0168-2