
Abstract
Many potential neurotherapeutic agents fail in the later stages during development due to insufficient blood-brain barrier (BBB) permeability or neurotoxic effects. To address this, we developed an in vitro model incorporating the neurovascular unit (NVU) — astrocytes, pericytes, neurons, and brain microvessel endothelial cells — designed to simulate the in vivo BBB and improve early drug screening. This model uses a direct contact triculture system enhanced by integrating SH-SY5Y neuron-like cells, enabling the study of permeability-linked neuronal responses. Our results show that this expanded NVU model, employing a Transwell® system, enhances the BBB’s restrictive properties and neuronal viability, potentially due to improved cell-cell signaling. Additionally, the model demonstrated increased efflux transporter expression, providing a more physiologically relevant assessment of neuroactivity in relation to BBB permeability. This innovative NVU model offers a predictive and robust tool for evaluating neurotherapeutic agents, facilitating the prioritization of candidates in large compound libraries and potentially reducing attrition rates in drug development. It represents a significant advancement in the methodology for early-stage neurotherapeutic screening, aligning in vitro findings more closely with in vivo responses.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Currently, the clinical translation of neurotherapeutics lags the rapid increase in neurological disorders and cases being seen worldwide. Thus, it is imperative that new methods are developed to help facilitate the pharmaceutical development of these agents addressing these needs. It has been theorized that a majority of to the difficulties associated with translation of these agents arise from the highly restrictive nature of the in vivo blood brain barrier (BBB). Preclinical in vitro screens do not provide the physiologically accurate BBB needed for proper lead candidate selection and optimization. Compound design and selection has traditionally focused on selected physiochemical properties (e.g., MW < 400, high lipophilicity, and poor solubility) that have been conventionally considered as necessary and sufficient prerequisites for the ability to traverse the BBB [1, 2]. In addition, a significant focus has also been placed on designing lower affinity and capacity for efflux transporters like P-glycoprotein (P-gp) as a route to increase parenchymal exposure. Often overlooked is fact that many of these properties also favor biotransformation, although the role of metabolism in the BBB or the potential for compounds possessing these physicochemical properties contributing to neurotoxic effects after biotransformation have not been widely investigated [3, 4]. We have hypothesized that by developing an in vitro BBB model where neuroactivity (defined as either a beneficial or toxic effect to neurons here) could also be assessed leading to a better evaluation of risks in earlier stages of drug development. This would aid in Go/No Go decision making while simultaneously directing the drug development process in the direction of appropriate in vivo BBB permeability.
Early development of the Bovine Brain Microvessel endothelial cells (BMECs) culture model by Borchardt and colleagues led to the development of in vitro BBB screening models. Approaches to developing physiologically relevant cell-based models of the BBB have been the focus since then. These models place an emphasis on the properties of the BMECs and have been the industrial standard to attempt to mitigate the attritions rates of neurotherapeutic. However, these efforts typically lead to in vitro models that are somewhat predictive, but lack in vivo physiological relevancy in the configuration of cells in the model. Moreover, the models are predicated on the fact that they are more readily amenable to higher throughput screening demands associated with large compound libraries [5]. Additionally, these models emphasize BBB permeability and do not incorporate the complete repertoire of cell-cell signaling, neuroactivity, assess associated toxicity, or induction of neuronal function, into the in vitro screening.
When developing an in vitro model for BBB permeability or neuroactivity screening the structure and the multiple cell types of the in vivo neurovascular unit (NVU) should be considered. We postulate that the conventional in vitro BBB screening methodologies place too much emphasis on the BMECs, where the synergistic interaction between supporting cells (astrocytes, pericytes, and neurons) that lead to the in vivo phenotype and neuroactivity are not included. In fact, most neuroactivity screens are performed separately in neuron only models, despite the recognition that cell-cell signaling between all four-cell types governs in vivo response. Thus, BBB permeability screening model predictability will be limited unless permeability and neuroactivity can be integrated into an enriched in vitro model system representing the in vivo structure of the NVU [6,7,8,9,10,11].
Specifically, the in vivo NVU is in part comprised of BMECs that express restrictive tight junctions, ubiquitous efflux transporters, and highly specific drug transporters, which together contribute the physical barrier and selective transport of solutes into and out of the brain parenchyma [12]. Surrounding the endothelium are pericytes and astrocytes, which fully envelop the BMECs and synergistically improve the BBB phenotype through the secretion of soluble factors that modulate BMEC protein and transporter expression, function, and even regulate capillary blood flow [9, 13,14,15,16,17,18]. These three cell types, in conjunction with the non-cellular basal lamina and neurons, make up the NVU and should be considered in BBB permeability models and neuroactivity screening tools. For example, it has been established that neurons in contact with nearby astrocytes lead to neuronal-glial interactions and signaling, which is propagated to the rest of the NVU to meet the metabolic demands of the brain parenchyma [19,20,21]. A number of vasoactive molecules are released by both astrocytes and neurons in response to synaptic glutamate to influence cerebral blood flow via vasoconstriction or dilation by pericytes and smooth muscle cells [17, 22, 23]. Cerebral blood flow is also regionally modulated based on neural activity, and there is evidence that the pericytes act to regulate vasoconstriction [22].
Given the cellular signaling between the neurons and the other NVU cell types, in vitro screening models that would be comprised of all four would best provide a predictive and physiologically relevant cell-based permeability linked neuroactivity assessment of potential drug candidates. Additionally, a synergistic model would also serve as an effective screen for many therapeutic agents theorized to possess potential neurotoxic off target effects driven by brain parenchymal exposure based on several scenarios including direct neuronal effects, significant accumulation, or disruption of the BBB [24]. Traditionally, in vitro neuroactivity screening is performed on cultures of neuronal cells independent from BBB permeability and assessed based on neuronal health and neurite outgrowth or retraction in response to incubation with chemical entities [24,25,26]. Although these assays allow for high throughput screening of large compound libraries and assessment of direct neuronal effects, they do not mimic the state of neuronal function found in vivo as these models neglect the permeation barrier of the BBB or the potential implications of therapeutic metabolites. Therefore, including the BBB in neuroactivity screening tools enhances the utility of the model [24, 27]. Work has been done to assess neuroactivity of compounds linked to BBB permeability and determine relative toxicity of a compound based on associated effects on BMECs as well as neurons, with neuroactivity measured based drug accumulated in the receiver chamber of a permeation assay [28, 29]. Although these works measure relative neuroactivity effect of a compound as it relates to BBB permeability, we believe that it is possible to further increase the in vivo relevancy of the models.
The study herein describes the development and early optimization of an in vitro NVU permeability-linked neuroactivity screening model. The model is predicated on utilizing a design of experiments optimized novel, direct contact BBB triculture for permeability assessment across human BMECs, pericytes, and astrocytes layered atop one another on a permeable filter support [30]. The resulting flux of the compound then leads to exposure in the basal chamber, where fap are seeded and time dependent response can be evaluated (Fig. 1). The SH-SY5Y cell line was utilized here due to their use in neurotoxicity studies, however it should be noted that this is proof of concept and the limitations of the SH-SY5Y cells are taken into account [29, 31, 32].

Cross sectional depiction of the neurovascular unit (NVU) with the endothelium (BMECs) lining the capillary, pericytes embedded within the basal lamina, astrocytes having nearly full coverage of the BMECs and surrounding pericytes, and neurons in close contact with the astrocytes (left). The direct contact triculture model on the apical surface of a Transwell® filter support with neurons in the basolateral chamber mimicking the in vivo NVU. Astrocytes are seeded first on the filter, followed by pericytes, then BMECs to generate a fully apical, direct contact triculture model and neurons seeded in the basolateral chamber to generate the in vitro NVU model for BBB permeability and neuroactivity screening (right)
To determine feasibility, BBB permeability linked neuroactivity was investigated utilizing marker compounds that were selected based upon reports indicating their effects on neuronal health and neurite outgrowth and the response to drug accumulation in the receiver chamber was determined. The model developed here encompasses the in vivo reality of an intended neurotherapeutic agent and its associated neuronal effects resulting in a physiologically relevant screening tool that may potentially be utilized to assess large libraries for hit and lead candidate selection.
Results
NVU optimization
A full factorial design was used to determine the optimal conditions using three levels of neuron density and two times of inclusion. Upon analysis, the seeding density and time of inclusion of the SH-SY5Y neurons with the DOE optimized direct contact triculture were determined to be the two most important factors that influence the barrier restrictiveness of the NVU model [30]. Neuronal influence on BBB properties was assessed based on the change in paracellular permeability marker compound, 4 kD FITC-dextran. Results revealed that, regardless of cell density, introducing neurons to the basal chamber at day 7 post HBEC-5i plating had a lesser impact on barrier properties in contrast to introduction earlier in culture at day 3. Introducing neurons on day 3 at 25,000 and 50,000 cells/cm2 resulted in the largest decreases in BBB permeation, 11% and 16% decreases respectively (Table 1). A neuron density of 25,000 cells/cm2 introduced at day 3 post endothelial cell plating were the conditions chosen for all further studies to facilitate neuroactivity measurements. The optimized conditions were repeated to confirm the decrease in the permeation rate for 4 kD FITC-dextran with the addition of neurons in the basolateral chamber. The Papp observed for the triculture alone was 5.83 ± 0.30 × 10−6 cm/sec compared to 5.34 ± 0.15 × 10−6 cm/sec with neurons in the basolateral chamber resulting in an apparent, but not statistically significant decrease of 9% in permeability (p = 0.065) (Fig. 2).

Apparent permeability (Papp) of 4 kD FITC-dextran across the direct contact triculture alone and the optimized NVU model with neurons in the basolateral chamber. Optimized conditions for the NVU model consist of 25,000 cells/cm2 SH-SY5Y neurons introduced to the apical direct contact triculture 3 days post endothelial cell plating. Error bars represent one standard deviation (n = 3)
Neuronal viability and NVU marker compounds
Viability and outgrowth of neurons throughout the length of coculture in the presence of the direct contact triculture was assessed. SH-SY5Y neuronal cells were cultured alone in a well plate for 8 days or for 24 hours alone and then combined with the apical triculture for an additional 7 days. Viability and outgrowth of neurons cultured alone was normalized to the control values of 100 ± 7% and 9% respectively. In the presence of the triculture neuron viability increased by 32 ± 3% and outgrowth increased by 28 ± 1% in comparison to the neurons alone (Fig. 3). This was determined based on quantifying relative fluorescence intensity of the neuron viability and neurite outgrowth stain using the Molecular Probes® Neurite Outgrowth Staining Kit.

Viability and outgrowth of SH-SY5Y neurons in culture with and without the presence of the direct contact triculture as measured by the Neurite Outgrowth Staining Kit. Viability and outgrowth of SH-SY5Y cells cultured alone were normalized to 100% for comparison. Statistical significance was determined using Student’s t-test between the two groups. Significance is labeled by (viability, outgrowth) where **p < 0.01. Error bars represent one standard deviation (n = 5)
Four paracellular permeability markers of increasing hydrodynamic radii were used to assess the tightness of the NVU model. The apparent permeability of [14C]-sucrose (4.6 Å) was 13.61 ± 1.94 × 10−6 cm/sec, followed by 4 kD FITC-dextran (14 Å) at 4.85 ± 0.20 × 10−6 cm/sec, 10 kD FITC-dextran (23 Å) at 3.64 ± 0.20 × 10−6 cm/sec, and 40 kD FITC-dextran (45 Å) at 1.92 ± 0.05 × 10−6 cm/sec (Fig. 4).

Apparent permeability of paracellular marker compounds [14C]-sucrose and FITC-dextrans of 4 kD, 10 kD, and 40 kD across the optimized NVU model. Error bars represent one standard deviation (n = 3)
The function of efflux transporter P-gp is also a key validation characteristic of any in vitro BBB model. P-gp function was assessed in the NVU model using P-gp substrate R123 in the presence and absence of P-gp inhibitor elacridar (Fig. 5). In the NVU model, the Papp of R123 alone was 12.12 ± 0.57 × 10−6 cm/sec versus 13.56 ± 0.50 × 10−6 cm/sec in the presence of elacridar (p < 0.05). In comparison, the Papp of R123 alone and in the absence of elacridar across the BBB direct contact triculture alone was 18.52 ± 0.58 × 10−6 cm/sec and 21.14 ± 0.46 × 10−6 cm/sec respectively (p < 0.01). The triculture alone shows a greater increase in the Papp of R123 in the presence of inhibitor (14% increase) compared to the NVU model (12% increase), however the overall permeability of R123 with and without inhibitor is decreased in the NVU model compared to the triculture alone (p < 0.001).

Apparent permeability of P-gp substrate rhodamine 123 (R123) in the presence and absence of elacridar, a P-gp inhibitor. Papp was measured across the triculture without neurons (dark grey) and the NVU model (light grey). One way ANOVA with a Tukey-Kramer post-hoc test was used to determine significance between R123 with and without elacridar for the triculture with neurons and without where *p < 0.05, **p < 0.01, and ***p < 0.001. Error bars represent one standard deviation (n = 3)
The apparent permeability of BBB high and low permeating compounds across the NVU model, whose structures are shown in Fig. 6, was measured for several compounds to evaluate the NVU assay’s ability for ranking potential therapeutic agents based on permeation rates (Fig. 7) [33]. Caffeine (Papp = 30.70 ± 1.18 × 10−6 cm/sec), carbamazepine (Papp = 25.37 ± 2.80 × 10−6 cm/sec), melatonin (Papp = 18.29 ± 0.50 × 10−6 cm/sec), and R123 in the presence of elacridar (Papp = 13.56 ± 0.50 × 10−6 cm/sec) are positive BBB permeants. R123 alone (Papp = 12.12 ± 0.57 × 10−6 cm/sec), clozapine (Papp = 11.44 ± 0.78 × 10−6 cm/sec), digoxin (Papp = 8.78 ± 0.37 × 10−6 cm/sec), prazosin (Papp = 3.90 ± 0.35 × 10−6 cm/sec), and cyclosporine A (Papp = 2.61 ± 0.37 × 10−6 cm/sec) are BBB negative permeants. Lapatinib was also tested for permeability but was undetectable in the receiver chamber after 3 hours (data not shown).

Compounds structure of caffeine, carbamazepine, melatonin, R123, elacridar, clozapine, digoxin, prazosin, and cyclosporin A

Apparent permeability of BBB positive (grey) and negative (patterned) permeants across the optimized NVU model. Assays were performed in triplicate and error bars represent one standard deviation (n = 3-6)
The BBB permeants that were tested across both the optimized triculture without neurons and the NVU model were plotted for comparison [30]. Of the markers screened, significant decreases in permeation rates across the NVU model were observed for R123 with elacridar (p < 0.001), R123 alone (p < 0.001), and prazosin (p < 0.01) while a significant increase in permeation was seen for clozapine (p < 0.01) all in comparison to the optimized triculture alone (Fig. 8).

Apparent permeability of high and low permeating marker compounds across the optimized direct contact triculture (dark grey) and the optimized NVU model (light grey). Student’s t-test was used to determine statistical significance between Papp of markers across the two models where *p < 0.05, **p < 0.01, and ***p < 0.001. Error bars represent one standard deviation (n = 3–6)
Permeability linked neuroactivity
Relative neuroactivity was measured in the NVU model following 3-hour BBB permeability at an initial apical concentration of 50 µM in the NVU model where drug accumulated in the receiver chamber containing SH-SY5Y neuronal cells over the course of permeation across the apical direct contact triculture (Fig. 9). This study was performed at n = 4 and neuroactivity (neuronal viability and outgrowth) data is reported as a percent of control NVU SH-SY5Y cells with vehicle (0.50% DMSO) alone, and flux reported for each compound to represent the amount accumulated in the neuronal chamber. In comparison to the control (neuron viability was 100 ± 8%, where neurite outgrowth was 100 ± 26%) caffeine accumulation resulted in a significant increase in viability with a non-significant increase in outgrowth (159 ± 34%, p < 0.05; 122 ± 26%; flux = 269 ± 14 pg/(cm2 · sec)). Carbamazepine (96 ± 18%; 96 ± 9%; flux = 310 ± 9 pg/(cm2 · sec)) demonstrated negligible changes in viability and outgrowth while clozapine (143 ± 20%; 111 ± 12%; flux = 372 ± 19 pg/(cm2 · sec)) and prazosin (137 ± 19%; 106 ± 22%; flux = 166 ± 14 pg/(cm2 · sec)) demonstrated insignificant increases in both viability and outgrowth in comparison to the control. Permeability linked neuroactivity of the SH-SY5Y cells in response to digoxin accumulation resulted in significant increases in both viability and outgrowth when compared to the control neuronal cells (157 ± 18%, p < 0.05; 147 ± 15%, p < 0.05; flux = 450 ± 19 pg/(cm2 · sec)). Lastly, cyclosporin A accumulation resulted in the largest increase in neuronal viability and insignificant changes in neurite outgrowth (365 ± 30%, p < 0.001; 106 ± 15%; flux = 127 ± 27 pg/(cm2 · sec)).

Neuroactivity of marker compounds after 3 hour permeability measured by neuronal viability (solid) and outgrowth (checkered) of SH-SY5Y cells represented as percentage of control NVU neuronal cells with vehicle (0.50% DMSO) alone and relative flux of each compound (red circle). Initial concentrations of 50 µM were placed in the apical chamber of the NVU model, SH-SY5Y neurons were stained for viability and outgrowth at the end of 3 hours of marker permeation. Viability stain was measured at ex: 483 nm and em: 525 nm and outgrowth stain was measured at ex: 535 nm and em: 590 nm. Markers are ordered based on the highest to lowest permeation rate across the NVU model. Statistical significance was determined using a one-way ANOVA followed by a Tukey-Kramer post-hoc test where *p < 0.05, **p < 0.01, and ***p < 0.001. Error bars represent one standard deviation (n = 4)
Representative images of neuronal viability and outgrowth staining are presented for the control, digoxin, and cyclosporin A samples (Fig. 10). In comparison to the control (panel a), punctate neurite projections are observable in the fluorescent (red) and bright field images of digoxin SH-SY5Y neurons (panel b), while a qualitative intensified green fluorescence and diffuse outgrowth is observed for cyclosporin A neuronal cells (panel c). These qualitative observations correlate with the quantifiable data obtained from the relative fluorescence results (Fig. 9).

Qualitative fluorescence (top) and bright field (bottom) images of SH-SY5Y neuronal cells after 3 hours of a control vehicle (0.50% DMSO), b digoxin, and c cyclosporin A accumulation after permeation across the apical triculture in the optimized NVU model. Images were obtained using the BioTek Cytation 3 with 20x objective and Green Fluorescent Protein and Texas Red filters for neuronal viability and outgrowth respectively. Scale bars represent 100 µm
BBB triculture viability
Viability of the apical triculture cells was inferred using an MTT assay to determine if observed neuronal effects or flux were due to changes in the integrity of the triculture cells. Inferred viability of the triculture cells is normalized to the triculture cells of the control NVU and reported as percent of the control (Fig. 11). The viability of the triculture following 3 hours of caffeine (104 ± 25%), carbamazepine (103 ± 5%), clozapine (99 ± 4%), digoxin (94 ± 5%), prazosin (82 ± 6%), and cyclosporin A (103 ± 2%) permeation resulted in no significant changes in comparison to the triculture of the control NVU (100 ± 2%, p > 0.05).

Viability of the apical triculture cells after 3-hour permeation linked neuroactivity measurement inferred using an MTT assay. Data is normalized to the triculture cells of the control NVU samples containing vehicle (0.50% DMSO) alone. One way ANOVA determined no significant differences between the triculture samples with markers and the control (p > 0.05). Error bars represent one standard deviation (n = 4)
P-Glycoprotein immunofluorescence imaging
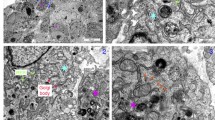
P-glycoprotein expression was evaluated in three in vitro models: monoculture of BMECs, BBB triculture model, and the NVU model. Semiquantitative immunofluorescence imaging, using the same antibody titer solution was conducted after the full culture period indicated enhanced cell densities in both the BBB triculture and NVU models compared to the monoculture. Notably, the NVU model appeared to exhibit the highest expression of P-gp when compared with the monoculture and the BBB (Fig. 12). The P-gp expression results are consistent with the rhodamine and elacridar functional data presented in Fig. 8, suggesting P-gp expression is increased in the NVU model.

The results of immunofluorescence staining of the HBEC-5i monoculture, the BBB triculture and NVU models. The NVU model exhibits the most pronounced expression of P-gp in NVU model in contrasted with the monoculture and the BBB triculture model
Discussion
Using a previously optimized in vitro model of the BBB that encompasses the 3 major cell types of the NVU [30], we have further improved upon the utility of the model by including neurons as the fourth cellular component for use in both permeability and associated neuroactivity evaluation of a potential drug candidate. Although we encourage direct, physiologically relevant contact between the different cell types of the NVU, it is important to consider that for the utility of the NVU model it is necessary to separate the neurons for subsequent neuroactivity screening. However, as evidenced by our results, there is apparent cell-cell signaling between the neurons and the apical triculture based on the decrease in permeation of paracellular markers, an index of tightness, across the BMECs and an increase in both neuronal viability and outgrowth when in the presence of the triculture as compared to being cultured separately. In addition, the presence of neurons may lead to an increase in P-gp expression, which is also a significant marker of barrier properties in vivo.
Seeding density of neurons in the basolateral chamber and the time of introduction of the apical BBB filter were chosen as the two key factors that would influence the characteristics of the model. Optimization was performed using a full factorial design to observe the impact of a low, medium, and high SH-SY5Y seeding density and two different times of BBB filter incorporation on the paracellular tightness of the model evaluated by permeation of a 4 kD FITC-dextran (Table 1). Results showed that allowing neurons and the apical triculture to coculture for a longer period of time, regardless of density, resulted in greater restrictiveness of the BMECs as compared to shorter coculturing times. This can be explained by the active cross-talk that occurs between the neurons and astrocytes in vivo [19,20,21]. The presence of neurons in companion wells has been shown to increase expression levels of occludin and claudin-5, essential tight junction proteins, in BMECs cultured on apical filter supports, which suggests that the presence of neurons in our NVU model may be increasing tight junction protein expression in the endothelium thus resulting in the reduced permeability of paracellular markers compared to the triculture alone.8,30 However, the decrease in permeability of the 4 kD dextran in the NVU model is not significant, therefore it is difficult to say with certainty what effects the presence of the neuronal cells has on tight junction expression in the absence of molecular based techniques (i.e. Western blot, mRNA expression). Secondly, lower densities of SH-SY5Y cells, 25,000 – 50,000 cells/cm2, resulted in greater tightening over the higher 75,000 cells/cm2 density used. This can be attributed to the possibility of overgrowth of the SH-SY5Y neurons at higher densities leading to possible decreases in viability and deleterious effects on the apical BBB cells. Moreover, reducing the confluency of the neurons in the basolateral chamber could facilitate the observation of neurite outgrowth as the potential for cellular overgrowth may be significantly reduced. In addition to the neurons influencing barrier properties, results of our study demonstrate that the presence of the apical triculture has synergistic effects on the SH-SY5Y cells. The increase in neuronal viability and outgrowth when cocultured with the apical triculture suggests that the other cell types of the NVU are essential for optimal neuronal health by way of cellular crosstalk. Note that the increase in viability and outgrowth observed for the neuronal cells in the NVU model may also be attributed to higher cell counts in these cultures compared to the neurons cultured alone. Both systems were seeded at the same initial density with the same neuronal cultures and grown for the same length of time before analysis, therefore an increase in neuron proliferation over the course of culture in the NVU model further supports the possibility of the different cell types of the NVU positively influencing each other via synergistic cell-cell signaling.
The NVU was evaluated using paracellular markers in a range of sizes ([14C]-sucrose < 4 kD FITC-dextran < 10 kD FITC-dextran < 40 kD FITC-dextran). The model performed as expected, showing a sequential reduction in permeability with increasing marker size suggesting that the model can prevent substantial paracellular permeation by a potential therapeutic compound—forcing the molecule to permeate through a transcellular route. The P-gp marker compound, R123 was used in the presence and absence of elacridar to determine the functional efflux of P-gp, which is a intrinsic characteristic of a BBB permeability screening tool. The NVU model showed a significant increase of R123 permeation when in the presence of P-gp inhibitor, which can be inferred to represent the relative function of P-gp in the in vitro model. The presence of neurons in the NVU model further increases P-gp function in comparison to the triculture alone reported previously. [30], as seen by the decrease in R123 permeation alone and in the presence of elacridar. The results appear to suggest that an increase in the P-gp expression occurs in the NVU when contrasted with the monoculture and the BBB triculture, which is apparent upon inspection of the immunofluorescence data illustrated in Fig. 11. P-gp expression is believed to be a key factor in the inherent restrictive barrier function of the BBB that acts to protect the brain parenchyma. Thus, the presence of neurons increases the physiological relevancy of the basolateral chamber, therefore leading to increased function of efflux transporters.
The NVU model’s utility for BBB permeability ranking was tested using several compounds known to be high or low BBB permeants as previously reported in vivo [34]. Caffeine is a psychoactive compound known for its ability to permeate and have an effect on the brain parenchyma and ranks as the most permeable compound to cross the NVU model [35]. Carbamazepine, an anticonvulsant shown to have limited P-gp interaction, is a positive BBB permeant showing adequate permeation across the NVU model [36]. The differentiation for positive and negative permeants was placed between the permeation of R123 in the presence and absence of elacridar as R123 alone should not positively permeate into the brain parenchyma. Although R123 is a P-gp substrate its BBB permeability in the NVU model is greater than that of other substrates tested, which suggests that P-gp substrates have different affinities and efflux capacity, and different efflux substrates can infer varying degrees of functionality of the transporter. Concentration dependent analysis could be done to determine which substrates might be best suited for the assessment of efflux transporter function in in vitro models. Clozapine is a psychoactive molecule which has been shown to be a possible P-gp substrate, however it is susceptible to metabolism with the metabolites being able to readily crossing the BBB [36,37,38,39,40]. Further analysis of the permeation samples should be done to determine the extent of permeation of the clozapine metabolites as the HPLC analytical technique used here may not be adequate for separation. Digoxin is a P-gp substrate that is actively effluxed from the BBB and its lack of permeation across the NVU model corresponds with expected outcomes from this molecule in BBB screening tools [41]. Prazosin is a substrate for the other predominantly expressed efflux transporter in the BBB, Breast Cancer Resistance Protein (BCRP)—the lack of permeation of this molecule across the NVU model suggests that there is functional expression of BCRP [42]. Lastly, the permeation of cyclosporin A (CsA) in the NVU was the lowest observed as it is a broad spectrum inhibitor for a number of efflux transporters [43]. P-glycoprotein expression suggests that the NVU model more effectively mimics the efflux transporter properties of the in vivo BBB, highlighting its potential superiority for neuropharmacological investigations.
Of the compounds that overlap between the libraries used, the observed rates of permeation for BBB positive compounds caffeine and carbamazepine are not significant between the two models [30]. These markers passively diffuse across the barrier via a transcellular pathway with some reported transporter contribution and limited efflux potential [35, 36, 44]. Of greater interest is the significant differences in permeation rates of effluxed or readily metabolized compounds in each in vitro model. As stated previously, R123 permeation with and without the inhibitor elacridar is significantly decreased in the NVU model, which may imply that the presence of the SH-SY5Y neuronal cells is modulating overall P-gp function to some degree. A significant difference is not observed for the P-gp substrate digoxin, however the permeation rate of BCRP substrate prazosin is significantly low in the NVU model compared to the optimized triculture alone. We postulate, based on the observed permeation rates in each model, that the presence of the neuronal cells in the basolateral chamber is facilitating synergistic signally between the neurons and the BBB triculture resulting in modulation of either efflux transporter function or overall expression. Additionally, the significantly higher permeation rate of metabolically active clozapine across the NVU model may imply a similar possibility for drug metabolizing enzymes. Ultimately, comparing the NVU model with the optimized BBB triculture alone reveals significant functional differences in observed permeation of compounds with transporter contributions.
Initial assessment of BBB permeability linked neuroactivity was performed after a 3-hour permeation period to determine how the permeation barrier of the apical triculture would impact neuroactivity of the SH-SY5Y cells in comparison to reported effects in direct neuronal experiments. Of the six markers chosen, caffeine and prazosin have limited reports on neuroactivity, cyclosporin A and digoxin are reported to inhibit neuroactivity, and clozapine and carbamazepine have been observed to induce neuroactivity, which are all based on direct neuronal incubation of these compounds [25, 28, 39, 45]. Compared to the control neuronal cells, caffeine accumulation resulted in a significant increase in neuroactivity but non-significant changes in outgrowth, which is expected based on its psychoactive and neuroprotective attributes [35]. Although carbamazepine and clozapine have been reported to induce neuroactivity as observed by increases in neurite outgrowth, the same was not seen in the NVU model [39, 45]. In the NVU model, digoxin was the only marker which showed significant neuroactivity in both neuronal viability and outgrowth by the observed increases in both. This is in opposition of what has been reported from direct neuron evaluation and its potential for causing systemic toxicity [25, 46]. Cyclosporin A 3-hour incubation in the NVU model resulted in the highest and most significant observed increase in neuronal viability relative to the other compounds tested. Cyclosporin A is an immunosuppressive compound which has been shown to increase the viability of neuronal precursor cells (NPCs), supporting the higher viability of the SH-SY5Y neurons in the NVU model [47]. Of interest here is the lack of correlation of neuroactivity to flux measurements or total accumulation in the neuronal chamber. Except digoxin being the highest accumulated compound with significant increases in viability and outgrowth, the neuronal response to cyclosporin A is the highest despite having the lowest flux of all compounds tested. In order to ensure that the neuroactivity responses were due to accumulation and not disruption of the triculture the relative viability of the triculture cells was assessed using via MTT assay, but revealed no significant changes in viability in comparison to the control suggesting that the barrier cells were not disrupted throughout the course of screening.
In this work the SH-SY5Y neuroblastoma cell line was used as the neuronal cell source despite the limitations associated with its use in neuroactivity models [31, 32]. The cells used in this work were in their undifferentiated stated based on reports on their sensitivity to neurotoxins and potential for use in neurotoxicity screening. Based on the results of this work we postulate that there may be modulation of the state of the SH-SY5Y cells in response to the synergistic signaling and that the state of the neuronal cells should be looked at in future investigations.
Taken together, we have demonstrated that the presence of a BBB model in neuroactivity screening is essential for adequately mimicking the path of a compound in vivo. In vivo the NVU responds to neuronal demands based on synergistic signaling from the neurons through direct contacts with astrocytes via neuronal secretion of a number of soluble factors [17, 22, 48]. This in turn modulates vascular diameter by pericyte action to protect the parenchyma [17]. We have demonstrated, in the optimized NVU model, that there is synergistic signaling occurring through all cell types by soluble factor secretion.
Conclusion
Presently, there are few therapeutic agents for many neurological disorders such as neurodegenerative and neurodevelopmental diseases and mental illness [49]. This may be due in large part to the high attrition rates in later stage clinical trials and the high costs associated with developing this class of compounds [50, 51]. High attrition rates are often attributed to a lack of successful delivery methods due to the restrictive BBB, a lack of efficacy when moving to in vivo studies and clinical trials, and neurotoxic side effects of drugs that permeate in excess into the brain parenchyma [52]. In an effort to provide a low-cost solution to the current need of the field, we have developed a physiologically relevant cell-based model of the NVU that incorporates BBB permeation and linked neuroactivity into a single screening tool. In addition, we have shown that incorporating all four cell types of the NVU leads to increased phenotypic expression of the BBB and cellular viability of the neuronal cells. The utility of the model serves to mimic the in vivo situation a therapeutic agent may encounter when attempting to cross the BBB into the brain parenchyma. By implementing this screening tool in pharmaceutical development of neurotherapeutic agents, as well as other classes of drugs, there is potential to decrease the resources needed for ranking hit and lead candidate compounds through the evaluation of BBB permeability linked neuroactivity using a single in vitro screening tool.
Methods
Materials
Human brain astrocytes and vascular pericytes, astrocyte medium, pericyte medium, and astrocyte and pericyte growth factors were all obtained from ScienCell Research Laboratories (Carlsbad CA, USA). HBEC-5i cells were purchased from ATCC (Manassas, VA, USA). SH-SY5Y neurons were graciously provided by Dr. Jean-Christophe Rochet (Purdue University, Department of Medicinal Chemistry and Molecular Pharmacology, West Lafayette, IN, USA). Transwell® filters of 12 mm 0.4 µm pore size, T75 culture flasks, Matrigel®, type I rat tail collagen, NuSerumTM, penicillin/streptomycin, and RPMI-1640 were purchased from Corning (Corning, NY, USA). Hank’s balanced salt solution (HBSS) and Dulbecco’s Modified Eagle Medium/Nutrient Mixture F-12 (DMEM/F-12) were obtained from Gibco (Carlsbad, CA, USA). Fetal bovine serum (FBS), hydrocortisone, rhodamine 123 (R123), elacridar, carbamazepine, colchicine, clozapine, caffeine, melatonin, digoxin, cyclosporin A, and prazosin hydrochloride were purchased from MilliporeSigma (St. Louis, MO, USA). Lapatinib was purchased from Attix Pharmaceuticals (Ontario, Canada). Radiolabeled [14C]-sucrose was obtained from Moravek Biochemicals Inc. (Brea, CA, USA). HEPES (2-[4-(2-hydroxyethyl)piperazin-1-yl]ethanesulfonic acid) and calcium chloride dehydrate were obtained from J.T. Baker (Phillipsburg, NJ, USA). Dexamethasone and MTT (3-(4,5-dimethylthiazolyl-2)-2,5-diphenyl tetrazolium bromide) were obtained from MP Biomedicals (Santa Ana, CA, USA). Endothelial cell growth supplement (ECGS) was purchased from Alfa Aesar (Haverhill, MA, USA). Fluorescein isothiocyanate (FITC) labeled 4 kD and 40 kD dextrans were purchased from Chondrex (Redmond, WA, USA). FITC labeled 10 kD dextran was purchased from TCI America (Portland, OR, USA). Poly-L-lysine (PLL) was purchased from Trevigen (Gaithersburg, MD, USA). Neurite Outgrowth Staining Kit was purchased from Molecular Probes (Eugene, OR, USA). P-Glycoprotein Monoclonal Antibody, Goat anti-Mouse IgG Secondary Antibody Alexa Fluor™ 488, DAPI (4’,6-Diamidino-2-Phenylindole, Dihydrochloride), Permount™ Mounting Medium, Microscope Slides, and Cover Glass were purchased from Invitrogen (Waltham, MA, USA). CoverGrip™ Coverslip Sealant was purchased from Biotium (Fremont, CA, USA). Blocking One Histo was purchased from Nacalai (Kyoto, Japan). 4% Paraformaldehyde (formaldehyde) aqueous solution was purchased from Electron Microscopy Sciences (Hatfield, PA, USA). Triton® X-100 Surfactant was purchased from Thermo Scientific Chemicals (Waltham, MA, USA).
Cell culture
Human Brain Endothelial Cells (HBEC-5i) were maintained in T-75 culture flasks pre-coated with Type I rat tail collagen with medium changes every 3 days and subculturing at 80–90% confluency—cells were utilized between passage 22 and 30. HBEC-5i culture medium was made up of DMEM/F-12 supplemented with 10% FBS, 15 mM HEPES, and 40 ug/mL ECGS. Human astrocytes and human brain vascular pericytes are maintained in T-75 culture flasks pre-coated with poly-L-lysine with medium changes every 3 days and were passaged at 80–90% confluency, where the cells were utilized between passages 4 and 10. Astrocyte culture medium was made up of Astrocyte Medium supplemented with 5% FBS, astrocyte growth supplement, and penicillin/streptomycin. Pericyte culture medium was comprised of Pericyte Medium supplemented with 5% FBS, pericyte growth supplement, and penicillin/streptomycin. Human neuroblastoma cell line, SH-SY5Y, was maintained in T-75 culture flasks with medium changes every 3 days and passaging at 80–90% confluency. SH-SY5Y cells were grown in RPMI 1640 with L-glutamine and 25 mM HEPES supplemented with 10% NuSerumTM and 1% penicillin/streptomycin. SH-SY5Y cells were used between passage 10 and 17 in all experiments.
Optimizing plating of neurons with direct contact triculture
Seeding density and time of introduction of the direct contact triculture with the SH-SY5Y neurons was optimized using a full factorial design of the two factors (Table 1). Plating methods of the BBB triculture is explained in the following section. Neurons were plated in a separate 12-well plate at 25,000, 50,000, or 75,000 cells/cm2 24 hours prior to placing the direct contact triculture atop the neurons. Neurons were cultured with the apical well of the triculture starting at 3 or 7 days post endothelial cell seeding and cultured until day 9 post based on the day of endothelium plating. Cultures were maintained at 37 °C and 5% CO2 with complete endothelial medium in the apical chamber and neuronal medium in the basolateral chamber with medium changes every other day. Optimized conditions were selected based on paracellular permeability of a 4 kD fluorescein labeled dextran.
Plating direct contact triculture with neurons in the basolateral chamber
The seeding of the direct contact triculture was done following the design of experiments optimized procedure, as reported by our laboratory previously [30]. Briefly, 12 mm, 0.4 µm pore polyester Transwell® filters were pre-coated with 5 µg/cm2 poly-L-lysine. Astrocytes were seeded at a density of 20,000 cells/cm2 and allowed to grow for 48 hours prior to seeding pericytes atop the astrocyte cell layer at a density of 20,000 cells/cm2. After 48 hours of pericyte growth, Matrigel® at a density of 25 µL/cm2 in HBSS was added to the astrocyte-pericyte lawn and allowed to incubate at 37 °C for 45 minutes. Matrigel® was removed and HBEC-5i cells were seeded directly atop the astrocyte-pericyte ECM coated lawn at a density of 80,000 cells/cm2 and maintained. Cultures were maintained with complete endothelial medium both on the apical side of the filter and basolateral chamber, when the direct contact triculture model formed. SH-SY5Y cells were introduced to the direct contact triculture 3 days post endothelial cell plating. Neurons were seeded 24 hours before the BBB triculture introduction in a 12-well plate at a density of 25,000 cells/cm2. Filter supports containing the direct contact triculture were placed above the culture neurons in the basolateral chamber of the well plate. Cultures were maintained with complete endothelial medium on the apical side of the filter and complete neuronal medium in the basolateral chamber. Cultures were utilized for assessing permeability and neuroactivity screening at 9 days post endothelial cell seeding.
Permeability assays
Prior to commencing all assays, cells were washed (2x) with HBSS to remove residual medium and then left to incubate in HBSS for 30 minutes at 37 °C to equilibrate. Apparent permeability of paracellular markers 4 kD, 10 kD, and 40 kD FITC-dextran, and [14C]-sucrose was performed at 37 °C on a rocking platform with samples pulled at 15, 30, 45, 60, and 90 minutes, where all studies were conducted under sink conditions. Dextran solutions and [14C]-sucrose were prepared at initial concentrations of 250 μg/mL and 0.25 μCi respectively in HBSS containing 0.50% DMSO. Dextran solutions were analyzed using a BioTek Synergy 4 plate reader with excitation at 485 nm and emission at 530 nm while [14C]-sucrose was assessed via scintigraphy.
The effective permeability coefficients of selected markers (caffeine, carbamazepine, melatonin, clozapine, digoxin, cyclosporine A, lapatinib, and prazosin) was performed at initial concentrations of 25 or 50 μM in HBSS containing 0.50% DMSO from 10 mM concentrated stock solutions in DMSO for each compound. Media samples were removed at 30, 60, 90, 120, 150, and 180 minutes for determining permeation rates and the remaining neurons then washed for the evaluation of neuroactivity. All samples for these compounds were analyzed using high performance liquid chromatography (HPLC). Apparent permeability (Papp) and flux (J) were calculated using the following equation (Eq. 1):
where dM/dT is the amount of material that moves across the filter over time, Co is the initial concentration in the donor (apical) chamber, and A is the surface area of the filter support.
Apparent permeability of P-gp substrate R123 was measured in the presence and absence of elacridar, a P-gp inhibitor. Working solutions of R123 at 10 µM and elacridar at 2 µM were prepared in HBSS with 1% DMSO. Replicates in the presence of inhibitor were incubated with elacridar for 45 minutes prior to the start of R123 permeation, while samples without inhibitor were incubated in blank HBSS. Samples were collected at 30, 60, 90, and 120 minute intervals and assessed using a BioTek Synergy 4 plate reader at excitation of 485 nm and emission of 530 nm. Apparent permeability was measured using Eq. 1 above.
High-performance liquid chromatography
All compounds were analyzed using an Agilent 1100 reversed phase HPLC equipped with a variable wavelength detector (VWD). All methods were run isocratically using water and acetonitrile (ACN) through an Ascentis® C-18 15 × 4.6 mm, 5 µm column at 25 µL injections. Caffeine mobile phase consisted of 90:10, water:ACN, flow rate of 1.0 mL/min run at ambient temperature and analysis at 275 nm. Carbamazepine was run using 65:35, water:ACN with a flow rate of 1.5 mL/min with a column temperature of 40 °C and observed at 284 nm. Clozapine utilized a mobile phase of 45:55, water:ACN at 1.5 mL/min flow rate and 40 °C column temperature with analysis at 254 nm. Colchicine analysis was performed using a mobile phase of 75:25, water:ACN at a flow rate of 1.5 mL/min, 40 °C column temperature, and wavelength of 354 nm. Cyclosporin A was run using a mobile phase of 30:70, water:ACN at 1.5 mL/min flow rate, 40 °C column temperature, and measured at 214 nm. Digoxin utilized a mobile phase of 30:70, water:ACN at a flow rate of 1.1 mL/min, 40 °C column temperature, and observed at 218 nm. Lapatinib utilized a mobile phase of 40:60, water:ACN at a flow rate of 1.0 mL/min, a column temperature of 25 °C, and VWD detection at 232 nm. Melatonin was measured using a mobile phase of 75:25, water:ACN with a flow rate of 1.5 mL/min, 40 °C column temperature, and measured at 222 nm. Prazosin was analyzed with a mobile phase of 65:35, water:ACN at a flow rate of 1.5 mL/min, a 40 °C column, and wavelength of 254 nm.
Neuroactivity assessment
Neuroactivity was assessed after 3-hour BBB permeability of marker compounds caffeine, carbamazepine, clozapine, digoxin, prazosin, and cyclosporin A using the dual fluorescent dye Molecular Probes® Neurite Outgrowth Staining Kit which provides neuronal viability and degree of neurite outgrowth differences in comparison to a control. Following incubation, the test compound was removed from the neuronal cells and neurons were washed with fresh HBSS. A staining solution containing cell viability indicator and cell membrane stain was prepared according to manufacturer recommendations in fresh HBSS and added to neuron samples. After cells were incubated with the stain for 20 minutes at room temperature the stain was removed, cells were gently washed with fresh HBSS, and background suppression solution was added for analysis. Fluorescence quantification was measured using a BioTek Neo2 plate reader where the viability stain was measured at excitation and emission of 483 nm and 525 nm, whereas the cell membrane stain was measured at excitation and emission wavelength of 535 nm and 590 nm. All samples were compared to a control containing vehicle alone (0.50% DMSO in HBSS) and cell free controls were used to account for background fluorescence. Qualitative images were obtained using the BioTek Cytation 3 with the 20x objective for bright field and fluorescent pictures. Green Fluorescent Protein (GFP) and Texas Red filters were used to observe fluorescence in each sample.
Triculture cell viability assay
The viability of the direct contact triculture at the completion of the neuroactivity measurements was inferred from the mitochondrial oxidation of MTT dye. At the end of BBB permeability linked neuroactivity studies triculture plated filters were moved to a blank chamber well and the apical solution was removed from the filter support then washed with fresh HBSS. The triculture was incubated with 450 µL of fresh HBSS and 50 µL of 5 mg/mL MTT stock solution in HBSS for 4 hours at 37 °C, blank HBSS was kept in the basolateral chamber to ensure cells were not directly exposed to air. After incubation, MTT solution was removed and replaced with 300 µL of DMSO to lyse cells and solubilize the mitochondria-generated formazan salt. A sample volume of 50 µL was diluted an equal volume of fresh DMSO in a 96-well plate. Absorbance was measured at 560 nm using a BioTek Powerwave HT plate reader. Samples were compared to a control of triculture incubated with vehicle alone (0.50% DMSO in HBSS) over the course of permeability measurement.
P-Glycoprotein immunofluorescence imaging
A neuron density of 25,000 cells/cm2 introduced at day 7 post endothelial cell plating were the conditions chosen for P-glycoprotein immunofluorescence imaging measurements. Monocultures of HBEC-5i cells, direct contact triculture BBB model cells were also seeded as control group. They were all seeded into 12-well Transwell® plates. Upon the completion of NVU model cells, the triculture model cells, and concurrent with the growth timeline of HBEC-5i monocultures, all sets of plates underwent fixation with a 4% paraformaldehyde aqueous solution for 15 minutes. Following fixation, cells were permeabilized with 0.1% Triton™ X-100 for 10 minutes and subsequently blocked with Blocking One Histo for 15 minutes at room temperature. Immunolabeling of efflux protein was performed using P-Glycoprotein Monoclonal Antibody at a concentration of 4 µg/mL in 5% Blocking One Histo for 3 hours at room temperature for each antibody. Subsequently, further incubation with Goat anti-Mouse IgG Secondary Antibody Alexa Fluor™ 488 at 0.5 µg/mL for 45 minutes. Nuclei were counterstained with DAPI at 5 µg/mL at room temperature for 15 minutes. The Transwell® membrane was then delicately excised with a scalpel, transferred onto microscope slides, and mounted with mounting medium. The assembly was covered with a glass coverslip and sealed using a Coverslip Sealant. Immunofluorescence images were captured using a Nikon A1Rsi Confocal microscope at 60X magnification.
References
Geldenhuys WJ, Mohammad AS, Adkins CE, Lockman PR. Molecular determinants of blood-brain barrier permeation. Ther Deliv. 2015;6:961–71. https://doi.org/10.4155/tde.15.32
Pajouhesh H, Lenz GR. Medicinal chemical properties of successful central nervous system drugs. NeuroRx. 2005;2:541–53. https://doi.org/10.1602/neurorx.2.4.541
Pardridge WM. The blood-brain barrier: bottleneck in brain drug development. NeuroRx. 2005;2:3–14.
Pardridge WM. Why is the global CNS pharmaceutical market so under-penetrated? Drug Discov Today. 2002;7:5–7. https://doi.org/10.1016/S1359-6446(01)02082-7
Bicker J, Alves G, Fortuna A, Falcão A. Blood–brain barrier models and their relevance for a successful development of CNS drug delivery systems: A review. Eur J Pharm Biopharm. 2014;87:409–32. https://doi.org/10.1016/j.ejpb.2014.03.012
Helms HC, Abbott NJ, Burek M, Cecchelli R, Couraud P-O, Deli MA, et al. In vitro models of the blood-brain barrier: An overview of commonly used brain endothelial cell culture models and guidelines for their use. J Cereb Blood Flow Metab. 2016;36:862–90. https://doi.org/10.1177/0271678X16630991
Helms HC, Waagepetersen HS, Nielsen CU, Brodin B. Paracellular tightness and Claudin-5 Expression is increased in the BCEC/Astrocyte blood–brain barrier model by increasing media buffer capacity during growth. AAPS J. 2010;12:759–70. https://doi.org/10.1208/s12248-010-9237-6
Kulczar C, Lubin KE, Lefebvre S, Miller DW, Knipp GT. Development of a direct contact astrocyte-human cerebral microvessel endothelial cells blood-brain barrier coculture model. J Pharm Pharmacol. 2017;69:1684–96. https://doi.org/10.1111/jphp.12803
Al Ahmad A, Taboada CB, Gassmann M, Ogunshola OO. Astrocytes and pericytes differentially modulate blood-brain barrier characteristics during development and hypoxic insult. J Cereb Blood Flow Metab. 2011;31:693–705. https://doi.org/10.1038/jcbfm.2010.148
Canfield SG, Stebbins MJ, Morales BS, Asai SW, Vatine GD, Svendsen CN, et al. An isogenic blood-brain barrier model comprising brain endothelial cells, astrocytes, and neurons derived from human induced pluripotent stem cells. J Neurochem. 2017;140:874–88. https://doi.org/10.1111/jnc.13923
Xue Q, Liu Y, Qi H, Ma Q, Xu L, Chen W, et al. A novel brain neurovascular unit model with neurons, astrocytes and microvascular endothelial cells of rat. Int J Biol Sci. 2013;9:174–89. https://doi.org/10.7150/ijbs.5115
Abbott NJ, Patabendige AAK, Dolman DEM, Yusof SR, Begley DJ. Structure and function of the blood-brain barrier. Neurobiol Dis. 2010;37:13–25. https://doi.org/10.1016/j.nbd.2009.07.030
Abbott NJ, Rönnbäck L, Hansson E. Astrocyte-endothelial interactions at the blood-brain barrier. Nat Rev Neurosci. 2006;7:41–53. https://doi.org/10.1038/nrn1824
Bouchaud C, Le Bert M, Dupouey P. Are close contacts between astrocytes and endothelial cells a prerequisite condition of a blood-brain barrier? The rat subfornical organ as an example. Biol Cell. 1989;67:159–65.
Daneman R, Zhou L, Kebede AA, Barres BA. Pericytes are required for blood-brain barrier integrity during embryogenesis. Nature. 2010;468:562–6. https://doi.org/10.1038/nature09513
Dohgu S, Takata F, Yamauchi A, Nakagawa S, Egawa T, Naito M, et al. Brain pericytes contribute to the induction and up-regulation of blood-brain barrier functions through transforming growth factor-beta production. Brain Res. 2005;1038:208–15. https://doi.org/10.1016/j.brainres.2005.01.027
Hamilton NB, Attwell D, Hall CN Pericyte-mediated regulation of capillary diameter: a component of neurovascular coupling in health and disease. Front Neuroenergetics 2010;2: https://doi.org/10.3389/fnene.2010.00005
Hall CN, Reynell C, Gesslein B, Hamilton NB, Mishra A, Sutherland BA, et al. Capillary pericytes regulate cerebral blood flow in health and disease. Nature. 2014;508:55–60. https://doi.org/10.1038/nature13165
Halassa MM, Fellin T, Takano H, Dong J-H, Haydon PG. Synaptic islands defined by the territory of a single astrocyte. J Neurosci. 2007;27:6473–7. https://doi.org/10.1523/JNEUROSCI.1419-07.2007
Bushong EA, Martone ME, Jones YZ, Ellisman MH. Protoplasmic astrocytes in CA1 stratum radiatum occupy separate anatomical domains. J Neurosci. 2002;22:183–92.
Banerjee S, Bhat MA. Neuron-Glial interactions in blood-brain barrier formation. Annu Rev Neurosci. 2007;30:235–58. https://doi.org/10.1146/annurev.neuro.30.051606.094345
McConnell HL, Kersch CN, Woltjer RL, Neuwelt EA. The translational significance of the neurovascular unit. J Biol Chem. 2017;292:762–70. https://doi.org/10.1074/jbc.R116.760215
Attwell D, Buchan AM, Charpak S, Lauritzen M, Macvicar BA, Newman EA. Glial and neuronal control of brain blood flow. Nature. 2010;468:232–43. https://doi.org/10.1038/nature09613
Bal-Price AK, Hogberg HT, Buzanska L, Coecke S. Relevance of in vitro neurotoxicity testing for regulatory requirements: Challenges to be considered. Neurotoxicol Teratol. 2010;32:36–41. https://doi.org/10.1016/j.ntt.2008.12.003
Sherman SP, Bang AG. High-throughput screen for compounds that modulate neurite growth of human induced pluripotent stem cell-derived neurons. Dis Model Mech 2018;11:. https://doi.org/10.1242/dmm.031906
Anderl JL, Redpath S, Ball AJ A Neuronal and Astrocyte Co-Culture Assay for High Content Analysis of Neurotoxicity. J Vis Exp JoVE 2009. https://doi.org/10.3791/1173
Radio NM, Mundy WR. Developmental neurotoxicity testing in vitro: Models for assessing chemical effects on neurite outgrowth. NeuroToxicol. 2008;29:361–76. https://doi.org/10.1016/j.neuro.2008.02.011
Hallier-Vanuxeem D, Prieto P, Culot M, Diallo H, Landry C, Tähti H, et al. New strategy for alerting central nervous system toxicity: Integration of blood–brain barrier toxicity and permeability in neurotoxicity assessment. Toxicol In Vitro. 2009;23:447–53. https://doi.org/10.1016/j.tiv.2008.12.011
Toimela T, Mäenpää H, Mannerström M, Tähti H. Development of an in vitro blood–brain barrier model—cytotoxicity of mercury and aluminum. Toxicol Appl Pharmacol. 2004;195:73–82. https://doi.org/10.1016/j.taap.2003.11.002
Lubin KE, Knipp GT. Design of experiment based optimization of an in vitro direct contact triculture blood brain barrier model for permeability screening. Pharm Pharmacol. Int J. 2021;9:143–58.
Kovalevich J, Langford D. Considerations for the use of SH-SY5Y Neuroblastoma cells in neurobiology. In: Amini S, White MK, editors. Neuronal Cell Cult., vol. 1078. Totowa, NJ: Humana Press; 2013. p. 9–21.
Shipley MM, Mangold CA, Szpara ML. Differentiation of the SH-SY5Y human Neuroblastoma cell line. J Vis Exp JoVE 2016. https://doi.org/10.3791/53193.
Wang Q, Rager JD, Weinstein K, Kardos PS, Dobson GL, Li J, et al. Evaluation of the MDR-MDCK cellline as a permeability screen for the blood-brain barrier. Int J Pharm. 2005;288:349–59. 2010.1016/
Savettieri G, Di Liegro I, Catania C, Licata L, Pitarresi GL, D’Agostino S, et al. Neurons and ECM regulate occludin localization in brain endothelial cells. Neuroreport. 2000;11:1081–4.
Chen X, Ghribi O, Geiger JD. Caffeine protects against disruptions of the blood-brain barrier in animal models of Alzheimer’s and Parkinson’s disease. J Alzheimers Dis JAD. 2010;20:S127–41. https://doi.org/10.3233/JAD-2010-1376
Maines LW, Antonetti DA, Wolpert EB, Smith CD. Evaluation of the role of P-glycoprotein in the uptake of paroxetine, clozapine, phenytoin and carbamazapine by bovine retinal endothelial cells. Neuropharmacology. 2005;49:610–7. https://doi.org/10.1016/j.neuropharm.2005.04.028
Masters A, Pandi-Perumal SR, Seixas A, Girardin J-L, McFarlane SI Melatonin, the hormone of darkness: from sleep promotion to Ebola treatment. Brain Disord Ther 2014;4:. https://doi.org/10.4172/2168-975X.1000151.
Liu Y, Zhang Z, Lv Q, Chen X, Deng W, Shi K, et al. Effects and mechanisms of melatonin on the proliferation and neural differentiation of PC12 cells. Biochem Biophys Res Commun. 2016;478:540–5. https://doi.org/10.1016/j.bbrc.2016.07.093
Lu X-H, Dwyer DS. Second-generation antipsychotic drugs, olanzapine, quetiapine, and clozapine enhance neurite outgrowth in PC12 cells via PI3K/AKT, ERK, and pertussis toxin-sensitive pathways. J Mol Neurosci MN. 2005;27:43–64.
Hellman K, Aadal Nielsen P, Ek F, Olsson R. An ex vivo model for evaluating blood–brain barrier permeability, efflux, and drug metabolism. ACS Chem Neurosci. 2016;7:668–80. https://doi.org/10.1021/acschemneuro.6b00024
Helms HC, Hersom M, Kuhlmann LB, Badolo L, Nielsen CU, Brodin B. An electrically tight in vitro blood–brain barrier model displays net brain-to-blood efflux of substrates for the ABC transporters, P-gp, Bcrp and Mrp-1. AAPS J. 2014;16:1046–55. https://doi.org/10.1208/s12248-014-9628-1
Mao Q, Unadkat JD. Role of the breast cancer resistance protein (BCRP/ABCG2) in drug transport—an update. AAPS J. 2014;17:65–82. https://doi.org/10.1208/s12248-014-9668-6
Qadir M, O’Loughlin KL, Fricke SM, Williamson NA, Greco WR, Minderman H, et al. Cyclosporin A is a broad-spectrum multidrug resistance modulator. Clin Cancer Res. 2005;11:2320–6. https://doi.org/10.1158/1078-0432.CCR-04-1725
Sun J, Xie L, Liu X. Transport of carbamazepine and drug interactions at blood-brain barrier. Acta Pharmacol Sin. 2006;27:249–53. https://doi.org/10.1111/j.1745-7254.2006.00246.x
Schultz L, Zurich M-G, Culot M, da Costa A, Landry C, Bellwon P, et al. Evaluation of drug-induced neurotoxicity based on metabolomics, proteomics and electrical activity measurements in complementary CNS in vitro models. Toxicol In Vitro. 2015;30:138–65. https://doi.org/10.1016/j.tiv.2015.05.016
Bauman JL, Didomenico RJ, Galanter WL. Mechanisms, manifestations, and management of digoxin toxicity in the modern era. Am J Cardiovasc Drugs Drugs Devices Interv. 2006;6:77–86. https://doi.org/10.2165/00129784-200606020-00002
Hunt J, Cheng A, Hoyles A, Jervis E, Morshead CM. Cyclosporin A has direct effects on adult neural precursor cells. J Neurosci. 2010;30:2888–96. https://doi.org/10.1523/JNEUROSCI.5991-09.2010
Iadecola C. The neurovascular unit coming of age: a journey through neurovascular coupling in health and disease. Neuron. 2017;96:17–42. https://doi.org/10.1016/j.neuron.2017.07.030
Miller G. Is pharma running out of brainy ideas. Science. 2010;329:502–4. https://doi.org/10.1126/science.329.5991.502
Kesselheim AS, Hwang TJ, Franklin JM. Two decades of new drug development for central nervous system disorders. Nat Rev Drug Discov. 2015;14:815–6. https://doi.org/10.1038/nrd4793
Choi DW, Armitage R, Brady LS, Coetzee T, Fisher W, Hyman S, et al. Medicines for the mind: policy-based ‘pull’ incentives for creating breakthrough CNS drugs. Neuron. 2014;84:554–63. https://doi.org/10.1016/j.neuron.2014.10.027
Gribkoff VK, Kaczmarek LK. The need for new approaches in CNS drug discovery: why drugs have failed, and what can be done to improve outcomes. Neuropharmacology. 2017;120:11–9. https://doi.org/10.1016/j.neuropharm.2016.03.021
Acknowledgements
This work was financially supported in part by Environmental Protection Agency OSAPE Star Grant #RD-84002701-0.
Author information
Authors and Affiliations
Corresponding author
Additional information
Publisher’s note Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Lubin, K.E., Xu, Z.(., Sluka, J.P. et al. Development of an in vitro model of the neurovascular unit for BBB permeability-linked neuroactivity screening. Med Chem Res 33, 1450–1463 (2024). https://doi.org/10.1007/s00044-024-03290-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00044-024-03290-4