
Abstract
Purpose
We hypothesised that during a rest-to-exercise transient in hypoxia (H), compared to normoxia (N), (i) the initial baroreflex sensitivity (BRS) decrease would be slower and (ii) the fast heart rate (HR) and cardiac output (CO) response would have smaller amplitude (A1) due to lower vagal activity in H than N.
Methods
Ten participants performed three rest-to-50 W exercise transients on a cycle-ergometer in N (ambient air) and three in H (inspired fraction of O2 = 0.11). R-to-R interval (RRi, by electrocardiography) and blood pressure profile (by photo-plethysmography) were recorded non-invasively. Analysis of the latter provided mean arterial pressure (MAP) and stroke volume (SV). CO = HR·SV. BRS was calculated by modified sequence method.
Results
Upon exercise onset in N, MAP fell to a minimum (MAPmin) then recovered. BRS decreased immediately from 14.7 ± 3.6 at rest to 7.0 ± 3.0 ms mmHg−1 at 50 W (p < 0.01). The first BRS sequence detected at 50 W was 8.9 ± 4.8 ms mmHg−1 (p < 0.05 vs. rest). In H, MAP showed several oscillations until reaching a new steady state. BRS decreased rapidly from 10.6 ± 2.8 at rest to 2.9 ± 1.5 ms mmHg−1 at 50 W (p < 0.01), as the first BRS sequence at 50 W was 5.8 ± 2.6 ms mmHg−1 (p < 0.01 vs. rest). CO-A1 was 2.96 ± 1.51 and 2.31 ± 0.94 l min−1 in N and H, respectively (p = 0.06). HR-A1 was 7.7 ± 4.6 and 7.1 ± 5.9 min−1 in N and H, respectively (p = 0.81).
Conclusion
The immediate BRS decrease in H, coupled with similar rapid HR and CO responses, is compatible with a withdrawal of residual vagal activity in H associated with increased sympathetic drive.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
In acute hypoxia, the control of heart rate (HR) and blood pressure is challenged by several mechanisms. To sustain the oxygen delivery, the decrease in blood oxygen concentration must be accompanied by an increase in cardiac output (CO), both at rest and during dynamic exercise (Stenberg et al. 1966; Ferretti et al. 1990; Adami et al. 2014). This CO increase is mainly sustained by an increase of HR with preserved stroke volume (SV) (Hartley et al. 1973; Siebenmann and Lundby 2015). Data suggest that these changes are the result of a different equilibrium in the autonomic nervous system, implying higher sympathetic and lower parasympathetic activity (Moore et al. 1986; Saito et al. 1988; Rowell et al. 1989; Sagawa et al. 1997; Boushel et al. 2001; Weisbrod et al. 2001; Halliwill and Minson 2002; Hopkins et al. 2003; Buchheit et al. 2004; Ferretti et al. 2005; Lador et al. 2008; Siebenmann and Lundby 2015).
When analysing the rest-to-exercise transient in normoxic condition, the first rapid HR response can be suppressed by vagal blockade with atropine (Fagraeus and Linnarsson 1976) suggesting a prompt withdrawal of the vagal activity (Fagraeus and Linnarsson 1976; Lador et al. 2006; Fontolliet et al. 2021). Following this concept, it has been hypothesised that conditions of reduced vagal activity at rest, as acute hypoxia, might be characterised by a reduced amplitude of the initial HR and CO response to exercise onset (phase I) (Lador et al. 2006). To the best of our knowledge, this hypothesis has been tested experimentally only once (Lador et al. 2008), although with a small number of observations (5 subjects). They reported that in acute hypoxia (inspired fraction of O2, FIO2, = 0.11) phase I of the cardiovascular response to exercise onset was characterised by smaller HR and CO amplitudes.
Hypoxia affects also arterial baroreflexes. Changes in slope of a baroreflex curve around the operating point (OP) (Kent et al. 1972) under different physiological conditions have been ascribed to modulation by the autonomic nervous system (Ogoh et al. 2005). The acute exposure to simulated high altitude (~ 4300 m) modifies the cardiac response of the carotid arterial baroreflex by resetting the entire response curve towards higher HR values and by decreasing the maximum gain of the reflex (Sagawa et al. 1997, neck pressure-suction technique). Coherently, application of the sequence method in acute hypoxia revealed a decrease of the spontaneous baroreflex sensitivity (BRS) around the OP (Klemenc and Golja 2011; Fisher et al. 2022).
Bringard et al. (2017), who applied the sequence method during the exercise transient, demonstrated an immediate reduction of the BRS at exercise start, which they ascribed to sudden vagal withdrawal, in line with the interpretation of the rapid cardiovascular response to exercise (Lador et al. 2006). Thus, if the vagal withdrawal conjecture holds, we would expect that, in acute normobaric hypoxia, a rest-to-light exercise transient would be characterised by: i) a smaller decrease of BRS and ii) a smaller amplitude of the phase I of HR and CO (corroborating the preliminary results of Lador et al 2008), than in normoxic conditions. The aim of the present study was to test these hypotheses by analysing the dynamics of arterial baroreflexes (Bringard et al. 2017; Taboni et al. 2021a, b, 2022) and the HR an CO kinetics (Lador et al. 2006; Fontolliet et al. 2021) during a rest-to-50 W exercise transient in normoxia and acute normobaric hypoxia.
Materials and methods
Subjects
Ten healthy subjects were enlisted (9 males and 1 female). Age, height, and body mass were 32 ± 6 years, 176 ± 9 cm, and 71 ± 13 kg, respectively. All subjects were moderately active. Their maximal aerobic capacity measured on the cycle ergometer (Lode Corival, Lode B.V., Groningen, The Netherlands) was 276 ± 67 W, 3.9 ± 1.0 W kg−1. They were already familiar with the laboratory protocols since they had participated in previous experiments. None reported history of cardiovascular, pulmonary, or neurological diseases or was taking medications at the time of the study. The subjects were asked to refrain from drinking coffee or smoking for 24 h before the experiments. All subjects gave their informed consent after having received a detailed description of the methods and experimental procedures of the study. Every subject was aware of the right of withdrawing from the study at any time without jeopardy. This study was performed in line with the principles of the Declaration of Helsinki. Approval was granted by the Commission Cantonale d’Éthique de la Recherche, Canton de Genève, CH (Date 11th July 2018–No. 2018–00913).
Experimental procedure
The subjects came to the laboratory on one occasion, at least 2 h after a light meal. In the laboratory, ambient temperature was set to 24 ± 1 °C and barometric pressure was 731 ± 8 mmHg. After instrumentation and wearing cycling shoes, the subject took place on an electromagnetically braked cycle ergometer (Lode Corival, Lode B.V., Groningen, The Netherlands). The experimental protocol was carried out in normoxia (N) and in hypoxia (H), which were administered in random order and were separated by 30 min to allow for rest and hydration. Both in N and H, participants wore an oro-nasal mask (7450 V2 Mask™, Hans Rudolph, Inc., Shawnee, KS, USA) connected to an ultrasonic flowmeter (Spiroson®, ECO MEDICS AG, Duernten, Switzerland). In H, the experiments were carried out while the subject was breathing a hypoxic gas mixture (FIO2 = 0.11), which was delivered by means of a low resistance, two-way non-rebreathing T-shape valve (Hans Rudolph, Inc., Shawnee, KS, USA). The inhalation port was connected to a 200 l Douglas bag, used as pressure buffer system, and filled with a gas mixture containing 11% oxygen in nitrogen coming from a high-pressure tank. After connection, 10 min of quiet breathing were allowed to attain alveolar gas equilibration before performing the procedure.
The experimental protocol was as follows. After subjects’ instrumentation and equipment calibration, the subjects spent at least 10 min in quiet rest. Data were continuously recorded during the last 5 min of this period. At the 4th minute of resting recording, a 20 µl capillary blood sample was taken from the right earlobe for blood lactate concentration ([La]) measurement. At the same time, a 35–55 µl arterialised capillary blood sample was taken from the left earlobe for measurement of pH, of oxygen and carbon dioxide partial pressures (pO2 and pCO2, respectively), and of bicarbonate concentration ([HCO3−]). Afterwards, the subject performed three square-wave rest-to-exercise transients at the constant power of 50 W. The first exercise bout lasted 10 min to ensure at least 5 min of steady state condition; blood samples for [La], pH, pO2, pCO2, and [HCO3−] were taken at the 9th minute. After the first exercise bout, the subject rested on the cycle-ergometer of 6 min and then performed two additional 50 W exercise bouts lasting 5 min, separated by 6 min of rest. Both in N and H, every rest-to-exercise transient started without previous flywheel acceleration; this usually implies that the mechanical power necessary for the flywheel acceleration may compensate for the delayed activation of the magnetic brake of the cycle ergometer (Hibi et al. 1996).
Measurements
Continuous non-invasive arterial blood pressure profiles were recorded at the medium finger (Portapres, Finapres® Medical Systems, Enschede, The Netherlands) and peripheral blood oxygen saturation (SpO2) was continuously monitored at the index finger of the left arm (Nellcor N-595, Medtronic, Minneapolis, MN, USA). The left arm was positioned on a support at the heart level. Beat-by-beat HR was recorded by electrocardiography (ECG100C module, BIOPAC® Systems Inc., Goleta, CA, USA). All signals were collected and sampled at 400 Hz (MP150 system with AcqKnowledge acquisition and analysis software, BIOPAC® Systems Inc., Goleta, CA, USA) and stored on a personal computer for subsequent analysis. [La] was measured by an enzymatic-amperometric method (Biosen C-Line Glucose and Lactate analyser, EKF Diagnostics, Cardiff, UK) on 20 μl capillary blood samples. Arterialised blood pH, pO2, pCO2, and [HCO3−] were measured (ABL800 FLEX, Radiometer, Brønshøj, Denmark) on 35–55 μl capillary blood samples.
Data treatment
Arterial blood pressure profiles were analysed to obtain beat-by-beat values of systolic (SAP), diastolic (DAP), and mean (MAP) arterial pressure using the Beatscope® software (Finapres® Medical Systems, Enschede, The Netherlands). The same software provided a beat-by-beat calculation of SV by the Modelflow method (Wesseling et al. 1993). Beat-by-beat CO was calculated as the SV times the corresponding HR and total peripheral resistances (TPR) as the ratio between MAP and CO. Data in steady state conditions were computed on the last 5 min of the first resting period and of the 10 min exercise bout.
In steady state condition, at rest and at exercise, the BRS was calculated with the sequence method (Bertinieri et al. 1988), using MAP and R-to-R interval (RRi) as independent and dependent variable, respectively (Taboni et al. 2018). A phase shift of one beat between MAP and RRi was introduced (Steptoe and Vogele 1990), then, sequences of 3 or more consecutive beats characterised by consensual increase or decrease in MAP and RRi were identified. Within each sequence, the relationship between RRi and MAP was analysed by linear regression to compute the slope and the coefficient of determination (R2). When R2 > 0.85, the slope was retained (Iellamo et al. 1997). In steady state conditions, the mean slope of the RRi versus MAP relationship was considered representative of the BRS for each subject and the mean RRi and MAP value was considered as the corresponding OP.
BRS during the exercise transients was computed with the same approach as previously proposed for exercise onset (Bringard et al. 2017), breath holding onset (Taboni et al. 2021b), fast whole body tilting (Taboni et al. 2021a), and light-to-moderate exercise transient (Taboni et al. 2022). A phase shift of one beat for HR was introduced and the same criteria used at steady state were applied to retain sequences. The mean value of the RRi versus MAP relationship over the three repetitions of exercise transient was considered representative of the mean BRS for each subject. In N during the exercise transient, MAP showed an abrupt fall, until a minimum MAP value was recorded, then a recovery. Consequently, the baroreflex sequences retrieved during the first seconds of exercise in N have been grouped in two categories: (i) before the attainment of minimum MAP (“pre MAPmin” in Fig. 3), and (ii) the first sequence after the attainment of minimum MAP (“post MAPmin” in Fig. 3). In H during the exercise transient, MAP did not show any abrupt fall, which prevened identification of a minimum MAP. Thus, the baroreflex sequences retrieved during the first seconds of exercise in N have been grouped as follows: (i) the very first sequence identified after the exercise start (“first slope” in Fig. 3), and (ii) the second sequence identified after the exercise start ("second slope” in Fig. 3).
The dynamics of the CO and HR changes over time (f(t)) during the two exercise transients was analysed using a bi exponential model (Barstow and Molé 1987; Lador et al. 2006):
where b is the baseline value, A is the response amplitude, d is the time delay, and τ is the time constant. The subscripts 1 and 2 refer to the initial (phase I) and the primary (phase II) components, respectively. H(t−d) is the Heaviside function, when t < d it equals 0 and the last term of the right-hand branch of Eq. 1 cancels out. When the amplitude of one phase resulted equal to 0 l min−1 for CO or 0 bpm for HR, the corresponding time constant was not considered for the statistical analysis.
Statistical analysis
Data are presented as mean ± standard deviation. Two-way ANOVA for repeated measures was used to investigate differences in the four steady state conditions in order to isolate the effect of exercise and of hypoxia separately. One way ANOVA for repeated measures was used to investigate differences between MAP and BRS measured at different time points during exercise transients. Tukey’s multiple comparisons test was used to isolate differences when necessary. Student’s T test for repeated measures was used to compare Eq. 1 parameters obtained in N and H. Differences were considered significant when p < 0.05. The statistical software Prism (version 8, GraphPad®, La Jolla, CA, USA) was used. Data fitting with Eq. 1 was performed after superimposition of the three exercise transients for each subject in order to avoid timeline distortion due to averaging (Francescato et al. 2014a, b; Bringard et al. 2014); MATLAB (version 9.5.0.944444 with Curve Fitting Toolbox, The MathWorks, Inc., Natick, MA, USA) was used with this aim. Figure 1, all Panels and Fig. 2, Panels C-D report average data from all rest-to-exercise transients and from all subjects (n = 30) interpolated at 0.1 s.
Results
Mean steady state data are reported in Table 1. All cardiovascular data changed from rest to exercise except DAP (both in N and H). Most data differed between H and N except for resting and exercising MAP, DAP, and [HCO3−], and resting SV, CO, and [La].
The time course of the main investigated parameters is shown in Fig. 1. In H, with respect to N, the exercise transient showed a greater increase of CO and HR, a slightly lower SV, MAP and TPR, and a progressive decrease of SpO2.

Time course of the cardiac output (CO), heart rate (HR), stroke volume (SV), mean arterial pressure (MAP), total peripheral resistances (TPR), and peripheral blood oxygen saturation (SpO2) during the rest to 50 W exercise transient in normoxia (red line) and in hypoxia (blue line). Mean values from all subjects (n = 10). The time scale refers to the time elapsed from the exercise onset
During the rest-to-exercise transient in N, MAP showed a sudden decrease at the very beginning of exercise (Fig. 1, Panel D, red line). Minimum MAP was 73 ± 11 mmHg (p = 0.0002 and p < 0.0001 vs. rest and 50 W steady state, respectively) and appeared after 8.1 ± 1.8 s from exercise onset. Such a pattern was not observed during the same transient in H (Fig. 1, Panel D, blue line). This implied that the pattern of the MAP-RRi relationship differed between conditions, as shown by Fig. 2.

Contour plots of the relationship between R-to-R interval (RRi) and mean arterial pressure (MAP) from 10 s before to 60 s after exercise onset (black arrowhead). Panels A and B: beat-by-beat value from a representative subject with a time shift of 1 beat applied between MAP and RRi. Panels C and D: mean values obtained from all rest-to-exercise transients (n = 30) with a time shift of 1 beat applied between MAP and RRi. In all panels, dots and squares represent, respectively, rest and 50 W steady states
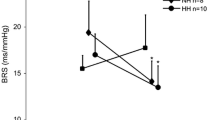
The BRS measured during the exercise transients are reported in Fig. 3. In N, the first slope was always computed on sequences that occurred before minimum MAP was reached, and thus was characterised by consensual decrease in MAP and RRi. The BRS values before and after minimum MAP were lower than in resting steady state (p ≤ 0.0148), but similar to those during exercise steady state (p ≥ 0.6796). In H, the first BRS slope was taken regardless of its direction and 22 out of 29 sequences were characterised by consensual decrease in MAP and RRi. Moreover, these first sequences appeared within 3.6 ± 3.1 s after the exercise onset. As in N, they were lower than in resting steady state (p = 0.0001).

Tukey representation of the baroreflex sensitivity (BRS) measured at different time points in normoxia and in hypoxia. *: significantly different vs. rest steady state (**: p < 0.01; ***: p < 0.001; ****: p < 0.0001); #: significantly different vs. exercise steady state (p < 0.05). MAPmin: minimum mean arterial pressure
Parameters obtained from the analysis of the CO and HR kinetics are reported in Table 2. Both the CO and HR baselines were higher in H than in N. Regarding phase I, CO A1, CO τ1, and HR A1 were similar in H and N whereas HR τ1 was higher in H than in N. Regarding phase II, both CO and HR A2 and τ2 were higher in H than in N.
Discussion
Arterial baroreflex at steady state
At rest, BRS around the OP was lower in H than in N, in line with previous observations (Koller et al. 1988; Lucy et al. 2000; Buchheit et al. 2004; Siebenmann and Lundby 2015; Siebenmann et al. 2019). This may be a consequence of a lower vagal output to the heart. In fact, BRS is reduced by full vagal blockade with atropine (Fontolliet et al. 2018), and enhanced in conditions characterised by a strong vagal output to the heart, such as supine posture (O’Leary et al. 2003; Steinback et al. 2005), water immersion (Chouchou et al. 2020), and short-term microgravity exposure (Hirayanagi et al. 2004; Di Rienzo et al. 2008). Moreover, the decrease of vagal activity in H is often associated with an increased sympathetic activity (Robinson et al. 1966; Saito et al. 1988; Hopkins et al. 2003; Ferretti et al. 2005; Tan et al. 2013; Siebenmann et al. 2015, 2019).
The lower BRS in H was coupled with a displacement of the OP toward lower RRi without any changes in MAP (Table 1), indicating resetting of the baroreflex in H. It was previously suggested that a hypoxic hyperventilation may be responsible for a decrease in the arterial baroreflex gain (Melcher 1980; Mancia and Mark 1983). In agreement with this hypothesis, the maximum baroreflex gain, obtained with the neck pressure-suction technique, was found to be lower than at sea level at altitudes above 4000 m (Sagawa et al. 1997). Since a FIO2 of 0.11 corresponds to a simulated altitude around 5000 m, the observed BRS decrease in the present study may in fact reflect a reduction of the maximum gain of the open-loop arterial baroreflex relationship. However, the observed decrease of the BRS around the OP in H may also be due to an OP displacement along the baroreflex curve away from the point of maximum gain, i.e. the centring point, as occurs during exercise (Potts et al. 1993; Norton et al. 1999; Fadel et al. 2001; Ogoh et al. 2003, 2005; Raven et al. 2006). To the best of our knowledge, no open-loop based studies of the arterial baroreflex have analysed the relative OP position in acute hypoxia.
At exercise, the BRS measured around the OP was lower in H than in N due to a BRS decrease of similar extent from rest to exercise steady states (−7.4 ± 4.6 ms mmHg−1 in N and −8.0 ± 3.4 ms mmHg−1 in H, p = 0.6158). This observation is compatible with residual vagal activity in resting H. If this is so, BRS decrease during the exercise transient might be the result of an additional vagal withdrawal, not only in N but also in H. In this case, the reduction of resting vagal activity in H would be partial. We can speculate that it may be greater the stronger the level of acute hypoxia to which a subject is exposed.
Baroreflex dynamics during the exercise transient
In N, MAP and TPR showed a sudden fall at exercise onset (Fig. 1, Panel D and E), in line with previous findings (Bringard et al. 2017), possibly due to prompt muscle vasodilation (Rådegran and Saltin 1998). The observed MAP fall was coupled with a RRi decrease as in a baroreflex relationship, so that BRS could be measured, as previously proposed (Bringard et al. 2017; Taboni et al. 2021a, b). The measured BRS of this baroreflex sequence was significantly lower than that measured at rest, and similar to that at exercise (Fig. 3), thus witnessing a prompt decrease of the BRS around the OP, similarly to previous observations (Bringard et al. 2017). When passing from rest to exercise, the open-loop arterial baroreflex relationship is displaced without any changes in maximal gain but with a shift of OP away from the centring point (Potts et al. 1993; Norton et al. 1999; Fadel et al. 2001; Ogoh et al. 2003, 2005; Raven et al. 2006), so that the BRS decrease measured upon exercise onset may simply represent this OP shift. The BRS decrease around OP occurs rapidly, within the very first few seconds of exercise, therefore only a very fast mechanism may be implied. It has been hypothesised that this mechanism may be the sudden withdrawal of the vagal tone, since the activation of the sympathetic system would require a longer time (Warner and Cox 1962; Fagraeus and Linnarsson 1976; Lador et al. 2006, 2013; Fontolliet et al. 2021).
In H, contrary to N, MAP did not show a single transient nadir but rather multiple oscillatory adjustments (Fig. 1, Panel D and Fig. 2), whereas TPR showed a similar initial fall upon exercise onset in the two conditions despite a lower baseline value in H (Fig. 1, Panel D). This apparently different MAP control at exercise onset in H may be due to a greater baseline sympathetic drive than in N (Saito et al. 1988; Rowell et al. 1989; Halliwill and Minson 2002), which would sustain arterial blood pressure and counteract hypoxic vasodilation (Weisbrod et al. 2001; Halliwill and Minson 2002). In H, several oscillations were observed before the attainment of a new steady state MAP value (Fig. 1, Panel D and Fig. 2) and during these MAP oscillations, it was possible to identify several baroreflex sequences. The BRS around OP promptly decreased since the first sequence after exercise start (Fig. 3), similarly to N. This suggests that the mechanism leading to the BRS decrease may be the same in the two analysed conditions, so that vagal withdrawal can still partly determine it. This would reinforce the concept that some level of vagal activity is still present in H.
Concerning baroreflex resetting during the exercise transient, in N the results are in line with previous literature (Bringard et al. 2017; Fagoni et al. 2020). The attainment of minimum MAP may trigger the resetting process. In H instead, it is hard to identify clear patterns of resetting. Although the mean data in H suggest the attainment of a minimum MAP similar to that in N (Fig. 2, panel D), this is not easily identifiable in all subjects. The progressive decrease of SpO2 during the exercise transient in H suggests that other factors than baroreflex mechanisms or stronger sympathetic stimulation may participate in the HR response to exercise onset (Halliwill et al. 2003), which make the patterns followed by HR and MAP more complex than in N.
Cardiovascular dynamics during the exercise transient
When analysing the rest-to-exercise transient, the phase I of the CO kinetics showed no significant differences in both amplitude and time constant in the two conditions (Table 2). This result goes against previous literature. In fact, when analysing a rest-to-50 W exercise transient in the same hypoxic conditions, lower A1 and lower τ1 were found in H than in N (Lador et al. 2008). This discrepancy may be due, at least in part, to the fact that in this study the number of participants was twice that of Lador et al (2008). Notwithstanding this, we note that, during full parasympathetic blockade, the CO kinetics at exercise onset shows a clear phase I, though with a smaller A1 than in control (Fontolliet et al. 2021). This was ascribed to sudden increase of SV by vagus-independent mechanisms active upon exercise onset, such as an increase of pre-load via muscle pump action (Chung et al. 1997; Sundblad et al. 2000, 2014; Naeije and Badagliacca 2017; Fagoni et al. 2020) and a reduction of after-load via prompt vasodilation at the level of the contracting muscles (Ferretti et al. 1995; DeLorey et al. 2003; Clifford 2007; Chin et al. 2010). Moreover, the phase I of the HR kinetics was characterised by similar amplitude and higher time constant in H than in N (Table 2). This is, at odds with the same hypoxic transient of Lador et al (2008), where HR A1 was lower and HR τ1 was equal compared to N. The phase I of HR responses to exercise onset is abolished under full parasympathetic blockade with atropine (Fontolliet et al. 2021). We speculate that, in H, the A1 of CO and HR were partially affected by vagal withdrawal, in line with the previous observation that at rest some degree of vagal activity may subsist. Notably, cardiovascular responses in N and H might have been influenced also by the respiratory apparatus used in the two conditions. In fact, in H a low resistance, two-way non-rebreathing T-shape valve was mounted at the mouth of each participant which increased the dead space by approximately 50%.
The phase II of both CO and HR was characterised by higher A2 and τ2 in H than in N, in line with previous findings (Lador et al. 2013). The current interpretation is that the incurring sympathetic stimulation may play a major role during this phase (Lador et al. 2006, 2013; Fontolliet et al. 2021), triggered by a more intense muscle-metabolic reflex (Houssiere et al. 2005) and chemoreflex (Jouett et al. 2015; Keir et al. 2019), given the progressive further SpO2 decrease after the exercise onset (Fig. 1, Panel F). Of course, a higher τ2 of CO should carry along a higher τ2 of oxygen uptake, which was not analysed in this study. Yet a hint suggesting that this may indeed be the case comes from the steady state lactate values in H, indicating early lactate accumulation in the exercise transient (Ferretti et al. 2022) leading to higher lactate steady state at exercise than at rest.
Conclusions
Upon exercise onset in acute normobaric hypoxia, the immediate fall of peripheral resistances at exercise start was not accompanied by a dramatic fall of mean arterial pressure. This last showed several oscillations until reaching a new steady state value. Moreover, the baroreflex sensitivity decreased immediately and was associated with the presence of a phase I of heart rate responses. These fast cardiovascular readjustments upon exercise onset are compatible with a withdrawal of residual vagal activity concomitant with increased sympathetic drive. After the first cardiovascular adjustments, the attainment of the new steady state was slower in hypoxia than in normoxia, as long as acute hypoxia was characterised by further peripheral deoxygenation.
Data availability
The datasets generated and analysed during the current study are available from the corresponding author on reasonable request.
Abbreviations
- [HCO3 −]:
-
Bicarbonate concentration
- [La]:
-
Blood lactate concentration
- A:
-
Amplitude of the bi-exponential model, subscripts 1 and 2 refers to phase I and phase II, respectively
- b :
-
Baseline value of the bi-exponential model
- BRS:
-
Spontaneous baroreflex sensitivity
- CO:
-
Cardiac output
- d :
-
Time delay of the bi-exponential model
- DAP:
-
Diastolic arterial pressure
- H:
-
Hypoxic condition
- HR:
-
Heart rate
- MAP:
-
Mean arterial pressure
- N:
-
Normoxic condition
- OP:
-
Operating point
- pCO2 :
-
Carbon dioxide partial pressures
- pO2 :
-
Oxygen partial pressures
- RRi:
-
R-to-R interval
- SAP:
-
Systolic arterial pressure
- SpO2 :
-
Peripheral blood oxygen saturation
- SV:
-
Stroke volume
- TPR:
-
Total peripheral resistances
- τ:
-
Time constant of the bi-exponential model, subscripts 1 and 2 refers to phase I and phase II, respectively
References
Adami A, Fagoni N, Ferretti G (2014) The Q-VO2 diagram: an analytical interpretation of oxygen transport in arterial blood during exercise in humans. Respir Physiol Neurobiol 193:55–61. https://doi.org/10.1016/j.resp.2014.01.007
Barstow TJ, Molé PA (1987) Simulation of pulmonary O2 uptake during exercise transients in humans. J Appl Physiol 63:2253–2261. https://doi.org/10.1152/jappl.1987.63.6.2253
Bertinieri G, Di Rienzo M, Cavallazzi A et al (1988) Evaluation of baroreceptor reflex by blood pressure monitoring in unanesthetized cats. Am J Physiol 254:H377–H383. https://doi.org/10.1152/ajpheart.1988.254.2.H377
Boushel R, Calbet J-AL, Rådegran G et al (2001) Parasympathetic neural activity accounts for the lowering of exercise heart rate at high altitude. Circ 104:1785–1791. https://doi.org/10.1161/hc4001.097040
Bringard A, Adami A, Moia C, Ferretti G (2014) A new interpolation-free procedure for breath-by-breath analysis of oxygen uptake in exercise transients. Eur J Appl Physiol 114:1983–1994. https://doi.org/10.1007/s00421-014-2920-z
Bringard A, Adami A, Fagoni N et al (2017) Dynamics of the RR-interval versus blood pressure relationship at exercise onset in humans. Eur J Appl Physiol 117:619–630. https://doi.org/10.1007/s00421-017-3564-6
Buchheit M, Richard R, Doutreleau S et al (2004) Effect of acute hypoxia on heart rate variability at rest and during exercise. Int J Sports Med 25:264–269. https://doi.org/10.1055/s-2004-819938
Chin LMK, Heigenhauser GJF, Paterson DH, Kowalchuk JM (2010) Pulmonary O2 uptake and leg blood flow kinetics during moderate exercise are slowed by hyperventilation-induced hypocapnic alkalosis. J Appl Physiol 108:1641–1650. https://doi.org/10.1152/japplphysiol.01346.2009
Chouchou F, Pichot V, Costes F et al (2020) Autonomic cardiovascular adaptations to acute head-out water immersion, head-down tilt and supine position. Eur J Appl Physiol 120:337–347. https://doi.org/10.1007/s00421-019-04278-4
Chung DC, Niranjan SC, Clark JW et al (1997) A dynamic model of ventricular interaction and pericardial influence. Am J Physiol - Heart Circ Physiol 272:H2942–H2962. https://doi.org/10.1152/ajpheart.1997.272.6.h2942
Clifford PS (2007) Skeletal muscle vasodilatation at the onset of exercise. J Physiol 583:825–833. https://doi.org/10.1113/jphysiol.2007.135673
DeLorey DS, Kowalchuk JM, Paterson DH (2003) Relationship between pulmonary O2 uptake kinetics and muscle deoxygenation during moderate-intensity exercise. J Appl Physiol 95:113–120. https://doi.org/10.1152/japplphysiol.00956.2002
Di Rienzo M, Castiglioni P, Iellamo F et al (2008) Dynamic adaptation of cardiac baroreflex sensitivity to prolonged exposure to microgravity: data from a 16-day spaceflight. J Appl Physiol 105:1569–1575. https://doi.org/10.1152/japplphysiol.90625.2008
Fadel PJ, Ogoh S, Watenpaugh DE et al (2001) Carotid baroreflex regulation of sympathetic nerve activity during dynamic exercise in humans. Am J Physiol - Heart Circ Physiol 280:H1383–H1390. https://doi.org/10.1152/ajpheart.2001.280.3.H1383
Fagoni N, Bruseghini P, Adami A et al (2020) Effect of lower body negative pressure on phase i cardiovascular responses at exercise onset. Int J Sports Med 41:209–218. https://doi.org/10.1055/a-1028-7496
Fagraeus L, Linnarsson D (1976) Autonomic origin of heart rate fluctuations at the onset of muscular exercise. J Appl Physiol 40:679–682. https://doi.org/10.1152/jappl.1976.40.5.679
Ferretti G, Boutellier U, Pendergast D et al (1990) IV. Oxygen transport system before and after exposure to chronic hypoxia. Int J Sports Med 11:S15–S20. https://doi.org/10.1055/s-2007-1024848
Ferretti G, Binzoni T, Hulo N et al (1995) Kinetics of oxygen consumption during maximal exercise at different muscle temperatures. Respir Physiol 102:261–268. https://doi.org/10.1016/0034-5687(95)00071-2
Ferretti G, Licker MJ, Anchisi S et al (2005) The effects of β1-adrenergic blockade on cardiovascular oxygen flow in normoxic and hypoxic humans at exercise. Eur J Appl Physiol 95:250–259. https://doi.org/10.1007/s00421-005-1393-5
Ferretti G, Fagoni N, Taboni A et al (2022) A century of exercise physiology: key concepts on coupling respiratory oxygen flow to muscle energy demand during exercise. Eur J Appl Physiol 122:1317–1365. https://doi.org/10.1007/s00421-022-04901-x
Fisher JP, Roche J, Turner R et al (2022) Hypobaric hypoxia and cardiac baroreflex sensitivity in young women. Am J Physiol - Heart Circ Physiol 323:H1048–H1054. https://doi.org/10.1152/ajpheart.00452.2022
Fontolliet T, Pichot V, Bringard A et al (2018) Testing the vagal withdrawal hypothesis during light exercise under autonomic blockade: a heart rate variability study. J Appl Physiol 125:1804–1811. https://doi.org/10.1152/japplphysiol.00619.2018
Fontolliet T, Bringard A, Adami A et al (2021) Vagal blockade suppresses the phase I heart rate response but not the phase I cardiac output response at exercise onset in humans. Eur J Appl Physiol 121:3173–3187. https://doi.org/10.1007/s00421-021-04769-3
Francescato MP, Cettolo V, Bellio R (2014a) Confidence intervals for the parameters estimated from simulated O2 uptake kinetics: effects of different data treatments. Exp Physiol 99:187–195. https://doi.org/10.1113/expphysiol.2013.076208
Francescato MP, Cettolo V, Bellio R (2014b) Assembling more O2 uptake responses: is it possible to merely stack the repeated transitions? Respir Physiol Neurobiol 200:46–49. https://doi.org/10.1016/j.resp.2014.06.004
Halliwill JR, Minson CT (2002) Effect of hypoxia on arterial baroreflex control of heart rate and muscle sympathetic nerve activity in humans. J Appl Physiol 93:857–864. https://doi.org/10.1152/japplphysiol.01103.2001
Halliwill JR, Morgan BJ, Charkoudian N (2003) Peripheral chemoreflex and baroreflex interactions in cardiovascular regulation in humans. J Physiol 552:295–302. https://doi.org/10.1113/jphysiol.2003.050708
Hartley LH, Vogel J, a, Landowne M, (1973) Central, femoral, and brachial circulation during exercise in hypoxia. J Appl Physiol 34:87–90. https://doi.org/10.1152/jappl.1973.34.1.87
Hibi N, Fujinaga H, Ishii K (1996) Work and power outputs determined from pedalling and flywheel friction forces during brief maximal exertion on a cycle ergometer. Eur J Appl Physiol 74:435–442. https://doi.org/10.1007/BF02337724
Hirayanagi K, Iwase S, Kamiya A et al (2004) Functional changes in autonomic nervous system and baroreceptor reflex induced by 14 days of 6 degrees head-down bed rest. Eur J Appl Physiol 92:160–167. https://doi.org/10.1007/s00421-004-1067-8
Hopkins SR, Bogaard HJ, Niizeki K et al (2003) β-Adrenergic or parasympathetic inhibition, heart rate and cardiac output during normoxic and acute hypoxic exercise in humans. J Physiol 550:605–616. https://doi.org/10.1113/jphysiol.2003.040568
Houssiere A, Najem B, Ciarka A et al (2005) Chemoreflex and metaboreflex control during static hypoxic exercise. Am J Physiol - Heart Circ Physiol 288:H1724–H1729. https://doi.org/10.1152/ajpheart.01043.2004
Iellamo F, Legramante JM, Raimondi G, Peruzzi G (1997) Baroreflex control of sinus node during dynamic exercise in humans: effects of central command and muscle reflexes. Am J Physiol - Heart Circ Physiol 272:H1157–H1164. https://doi.org/10.1152/ajpheart.1997.272.3.H1157
Jouett NP, Watenpaugh DE, Dunlap ME, Smith ML (2015) Interactive effects of hypoxia, hypercapnia and lung volume on sympathetic nerve activity in humans. Exp Physiol 100:1018–1029. https://doi.org/10.1113/EP085092
Keir DA, Duffin J, Millar PJ, Floras JS (2019) Simultaneous assessment of central and peripheral chemoreflex regulation of muscle sympathetic nerve activity and ventilation in healthy young men. J Physiol 597:3281–3296. https://doi.org/10.1113/JP277691
Kent BB, Drane JW, Blumenstein B, Manning JW (1972) A mathematical model to assess changes in the baroreceptor reflex. Cardiol 57:295–310
Klemenc M, Golja P (2011) Baroreflex sensitivity in acute hypoxia and carbohydrate loading. Eur J Appl Physiol 111:2509–2515. https://doi.org/10.1007/s00421-011-1875-6
Koller EA, Drechsel S, Hess T et al (1988) Effects of atropine and propranolol on the respiratory, circulatory, and ECG responses to high altitude in man. Eur J Appl Physiol 57:163–172. https://doi.org/10.1007/BF00640657
Lador F, Azabji Kenfack M, Moia C et al (2006) Simultaneous determination of the kinetics of cardiac output, systemic O2 delivery, and lung O2 uptake at exercise onset in men. Am J Physiol-Regul, Integr Comp Physiol 290:R1071–R1079. https://doi.org/10.1152/ajpregu.00366.2005
Lador F, Tam E, Azabji Kenfack M et al (2008) Phase I dynamics of cardiac output, systemic O2 delivery, and lung O2 uptake at exercise onset in men in acute normobaric hypoxia. Am J Physiol-Regul, Integr Comp Physiol 295:R624–R632. https://doi.org/10.1152/ajpregu.00797.2007
Lador F, Tam E, Adami A et al (2013) Cardiac output, O2 delivery and V˙O2 kinetics during step exercise in acute normobaric hypoxia. Respir Physiol Neurobiol 186:206–213. https://doi.org/10.1016/J.RESP.2013.01.017
Lucy SD, Hughson RL, Kowalchuk JM et al (2000) Body position and cardiac dynamic and chronotropic responses to steady-state isocapnic hypoxaemia in humans. Exp Physiol 85:227–237. https://doi.org/10.1111/j.1469-445X.2000.01932.x
Mancia G, Mark AL (1983) Arterial baroreflexes in humans. comprehensive physiology. Wiley & Sons Inc, Hoboken, NJ, USA, pp 755–793
Melcher A (1980) Carotid baroreflex heart rate control during the active and the assisted breathing cycle in man. Acta Physiol Scand 108:165–171. https://doi.org/10.1111/j.1748-1716.1980.tb06514.x
Moore LG, Cymerman A, Huang SY et al (1986) Propranolol does not impair exercise oxygen uptake in normal men at high altitude. J Appl Physiol 61:1935–1941. https://doi.org/10.1152/jappl.1986.61.5.1935
Naeije R, Badagliacca R (2017) The overloaded right heart and ventricular interdependence. Cardiovasc Res 113:1474–1485. https://doi.org/10.1093/cvr/cvx160
Norton KH, Boushel R, Strange S et al (1999) Resetting of the carotid arterial baroreflex during dynamic exercise in humans. J Appl Physiol 87:332–338. https://doi.org/10.1152/jappl.1999.87.1.332
O’Leary DD, Kimmerly DS, Cechetto AD, Shoemaker JK (2003) Differential effect of head-up tilt on cardiovagal and sympathetic baroreflex sensitivity in humans. Exp Physiol 88:769–774. https://doi.org/10.1113/eph8802632
Ogoh S, Fadel PJ, Nissen P et al (2003) Baroreflex-mediated changes in cardiac output and vascular conductance in response to alterations in carotid sinus pressure during exercise in humans. J Physiol 550:317–324. https://doi.org/10.1113/jphysiol.2003.041517
Ogoh S, Fisher JP, Dawson EA et al (2005) Autonomic nervous system influence on arterial baroreflex control of heart rate during exercise in humans. J Physiol 566:599–611. https://doi.org/10.1113/jphysiol.2005.084541
Potts JT, Shi XR, Raven PB (1993) Carotid baroreflex responsiveness during dynamic exercise in humans. Am J Physiol 265:H1928–H1938
Rådegran G, Saltin B (1998) Muscle blood flow at onset of dynamic exercise in humans. Am J Physiol - Heart Circ Physiol 274:H314–H322. https://doi.org/10.1152/ajpheart.1998.274.1.h314
Raven PB, Fadel PJ, Ogoh S (2006) Arterial baroreflex resetting during exercise: a current perspective. Exp Physiol 91:37–49. https://doi.org/10.1113/expphysiol.2005.032250
Robinson BF, Epstein SE, Beiser GD, Braunwald E (1966) Control of heart rate by the autonomic nervous system. Circ Res 19:400–411. https://doi.org/10.1161/01.RES.19.2.400
Rowell LB, Johnson DG, Chase PB et al (1989) Hypoxemia raises muscle sympathetic activity but not norepinephrine in resting humans. J Appl Physiol 66:1736–1743. https://doi.org/10.1152/jappl.1989.66.4.1736
Sagawa S, Torii R, Nagaya K et al (1997) Carotid baroreflex control of heart rate during acute exposure to simulated altitudes of 3,800 m and 4,300 m. Am J Physiol-Regul, Integr Comp Physiol 273:R1219–R1223. https://doi.org/10.1152/ajpregu.1997.273.4.R1219
Saito M, Mano T, Iwase S et al (1988) Responses in muscle sympathetic activity to acute hypoxia in humans. J Appl Physiol 65:1548–1552. https://doi.org/10.1152/jappl.1988.65.4.1548
Siebenmann C, Lundby C (2015) Regulation of cardiac output in hypoxia. Scand J Med Sci Sports 25:53–59. https://doi.org/10.1111/sms.12619
Siebenmann C, Rasmussen P, Sørensen H et al (2015) Hypoxia increases exercise heart rate despite combined inhibition of β-adrenergic and muscarinic receptors. Am J Physiol-Heart Circ Physiol 308:H1540–H1546. https://doi.org/10.1152/ajpheart.00861.2014
Siebenmann C, Ryrsø CK, Oberholzer L et al (2019) Hypoxia-induced vagal withdrawal is independent of the hypoxic ventilatory response in men. J Appl Physiol 126:124–131. https://doi.org/10.1152/japplphysiol.00701.2018
Steinback CD, O’Leary DD, Bakker J et al (2005) Carotid distensibility, baroreflex sensitivity, and orthostatic stress. J Appl Physiol 99:64–70. https://doi.org/10.1152/japplphysiol.01248.2004
Stenberg J, Ekblom B, Messin R (1966) Hemodynamic response to work at simulated altitude, 4,000 m. J Appl Physiol 21:1589–1594. https://doi.org/10.1152/jappl.1966.21.5.1589
Steptoe A, Vogele C (1990) Cardiac baroreflex function during postural change assessed using non-invasive spontaneous sequence analysis in young men. Cardiovasc Res 24:627–632. https://doi.org/10.1093/cvr/24.8.627
Sundblad P, Haruna Y, Tedner B, Linnarsson D (2000) Short-term cardiovascular responses to rapid whole-body tilting during exercise. Eur J Appl Physiol 81:259–270. https://doi.org/10.1007/s004210050041
Sundblad P, Spaak J, Kaijser L (2014) Time courses of central hemodynamics during rapid changes in posture. J Appl Physiol 116:1182–1188. https://doi.org/10.1152/japplphysiol.00690.2013
Taboni A, Fagoni N, Vinetti G, Ferretti G (2018) Baroreflex sensitivity: an algebraic dilemma. Acta Physiol. https://doi.org/10.1111/apha.12979
Taboni A, Fagoni N, Fontolliet T et al (2021a) A closed-loop approach to the study of the baroreflex dynamics during posture changes at rest and at exercise in humans. Am J Physiol-Regul Integr Comp Physiol 321:R960–R968. https://doi.org/10.1152/ajpregu.00167.2021
Taboni A, Vinetti G, Fontolliet T et al (2021b) Baroreflex responses during dry resting and exercise apnoeas in air and pure oxygen. Eur J Appl Physiol 121:539–547. https://doi.org/10.1007/s00421-020-04544-w
Taboni A, Fagoni N, Fontolliet T et al (2022) Dynamics of cardiovascular and baroreflex readjustments during a light-to-moderate exercise transient in humans. Eur J Appl Physiol. https://doi.org/10.1007/s00421-022-05011-4
Tan CO, Tzeng Y-C, Hamner JW et al (2013) Alterations in sympathetic neurovascular transduction during acute hypoxia in humans. Am J Physiol-Regul Integr Comp Physiol 304:R959–R965. https://doi.org/10.1152/ajpregu.00071.2013
Warner HR, Cox A (1962) A mathematical model of heart rate control by sympathetic and vagus efferent information. J Appl Physiol 17:349–355. https://doi.org/10.1152/jappl.1962.17.2.349
Weisbrod CJ, Minson CT, Joyner MJ, Halliwill JR (2001) Effects of regional phentolamine on hypoxic vasodilatation in healthy humans. J Physiol 537:613–621. https://doi.org/10.1111/j.1469-7793.2001.00613.x
Wesseling KH, Jansen JR, Settels JJ, Schreuder JJ (1993) Computation of aortic flow from pressure in humans using a nonlinear, three-element model. J Appl Physiol (bethesda, Md : 1985) 74:2566–2573. https://doi.org/10.1152/jappl.1993.74.5.2566
Funding
Open access funding provided by Università degli Studi di Brescia within the CRUI-CARE Agreement. This study was supported by Swiss National Science Foundation, Berne, CH (grant numbers 32003B_179448) to Guido Ferretti.
Author information
Authors and Affiliations
Contributions
GF conceived and designed research. AT, NF, and TF conducted experiments. AT and GV analysed data. AT wrote the manuscript. All authors read and approved the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical approval
Ethical approval was granted by the Commission Cantonale d’Éthique de la Recherche, Canton de Genève, CH (Date 11th July 2018–No. 2018–00913).
Consent to participate
All subjects gave their written informed consent after having received a detailed description of the methods and experimental procedures of the study.
Consent for publication
Informed consent was obtained from all participants included in the study.
Additional information
Communicated by Ellen Adele Dawson.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Taboni, A., Fagoni, N., Fontolliet, T. et al. Baroreflex dynamics during the rest to exercise transient in acute normobaric hypoxia in humans. Eur J Appl Physiol 124, 2765–2775 (2024). https://doi.org/10.1007/s00421-024-05485-4
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00421-024-05485-4