
Abstract
Main conclusion
Biphenyl and dibenzofuran phytoalexins are differentially distributed among species of the rosaceous subtribe Malinae, which includes apple and pear, and exhibit varying inhibitory activity against phytopathogenic microorganisms.
Abstract
Biphenyls and dibenzofurans are specialized metabolites, which are formed in species of the rosaceous subtribe Malinae upon elicitation by biotic and abiotic inducers. The subtribe Malinae (previously Pyrinae) comprises approximately 1000 species, which include economically important fruit trees such as apple and pear. The present review summarizes the current status of knowledge of biphenyls and dibenzofurans in the Malinae, mainly focusing on their role as phytoalexins. To date, 46 biphenyls and 41 dibenzofurans have been detected in 44 Malinae species. Structurally, 54 simple molecules, 23 glycosidic compounds and 10 miscellaneous structures were identified. Functionally, 21 biphenyls and 21 dibenzofurans were demonstrated to be phytoalexins. Furthermore, their distribution in species of the Malinae, inhibitory activities against phytopathogens, and structure–activity relationships were studied. The most widely distributed phytoalexins of the Malinae are the three biphenyls aucuparin (3), 2ʹ-methoxyaucuparin (7), and 4ʹ-methoxyaucuparin (9) and the three dibenzofurans α-cotonefuran (47), γ-cotonefuran (49), and eriobofuran (53). The formation of biphenyl and dibenzofuran phytoalexins appears to be an essential defense weapon of the Malinae against various stresses. Manipulating phytoalexin formation may enhance the disease resistance in economically important fruit trees. However, this approach requires an extensive understanding of how the compounds are formed. Although the biosynthesis of biphenyls was partially elucidated, formation of dibenzofurans remains largely unclear. Thus, further efforts have to be made to gain deeper insight into the distribution, function, and metabolism of biphenyls and dibenzofurans in the Malinae.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
The subtribe Malinae (previously Pyrinae) of the subfamily Amygdaloideae within the Rosaceae consists of about 30 genera and approximately 1000 species and includes economically important fruit trees, such as apple (Malus domestica), pear (Pyrus communis), and quince (Cydonia oblonga) (Sun et al. 2018; Ulaszewski et al. 2021). The major phytoalexins of the Malinae are biphenyls and dibenzofurans (Watanabe et al. 1982; Kemp et al. 1983; Kokubun and Harborne 1995). Phytoalexins are de novo formed specialized metabolites exhibiting antimicrobial activity. They accumulate in plants after exposure to biotic and abiotic stresses as a component of a complex system of disease resistance (Jeandet 2015; Bizuneh 2021). Biphenyls and dibenzofurans are biogenically related phenolic compounds. Biphenyls consist of two benzene rings connected by a single C–C bond, whereas dibenzofurans have two benzene rings connected by a furan ring (Fig. 1). In the Malinae, the core structures of both classes of compounds are usually decorated by hydroxy, methoxy, sugar (glucose and/or rhamnose), and other moieties. The last review of biphenyl and dibenzofuran phytoalexins of the Malinae was published over ten years ago. It reported 10 biphenyl and 17 dibenzofuran phytoalexins, which were isolated from 29 species belonging to 14 Malinae genera (Chizzali and Beerhues 2012).



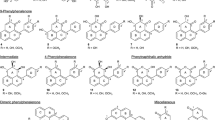
Biphenyls (1–46) and dibenzofurans (47–87) isolated from species of the rosaceous subtribe Malinae
Here, a survey was conducted by going through large electronic databases, such as PubMed, Google Scholar, and SciFinder etc. Based on these literature searches, 46 biphenyls and 41 dibenzofurans were detected in 44 plant species, which belong to 14 genera of the subtribe Malinae. Of the recorded compounds, 21 biphenyls and 21 dibenzofurans were formed in response to biotic and/or abiotic inducers. Thus, they were considered as phytoalexins. We summarize the chemical diversity of the reported biphenyls and dibenzofurans and their distribution among the Malinae species. Progress in elucidating their biosynthetic pathway is also presented. In the context of phytoalexin induction conditions, apple replant disease (ARD) is introduced as an in vivo model for eliciting the formation of biphenyl and dibenzofuran phytoalexins in apple roots. Finally, new information is added to the current knowledge of the antimicrobial activity of the phytoalexins, particularly regarding their inhibitory effect on phytopathogens.
Chemical diversity and distribution of biphenyls and dibenzofurans in the subtribe Malinae
As presented in Fig. 1, the chemical structures of biphenyls and dibenzofurans in the Malinae are diverse. They involve simple molecules with hydroxy and/or methoxy substitutions at the aromatic rings of the basic biphenyl and dibenzofuran skeletons (1–13, 17–20, 27, 28, 33–36, 44, 47–55, 57–63, 66–70, 72–76, 83, 84, 86, 87; in total 54 structures), O-glycosides with glucose and/or rhamnose moieties (14–16, 21–26, 32, 40–43, 45, 46, 56, 77–82; in total 23 compounds), and miscellaneous structures with aliphatic, aromatic, and/or heterocyclic substituents at the aromatic rings of the basic skeletons (29–31, 37–39, 64, 65, 71, 85; in total 10 structures). These biphenyl and dibenzofuran compounds were detected in 44 species of 14 Malinae genera, which were distributed among three clades in a phylogenetic tree (Fig. 2; Tables 1, 2) (Sun et al. 2018). A detailed analysis indicated that most species in clade III (7 out of 9) contain only biphenyls. In contrast, approximately half of the species in clade II (13 out of 25) and clade I (6 out of 10) form only dibenzofurans. In addition, the types of phytoalexin structures differed between the three clades. In species of clades I, II, and III, simple molecules constituted 63.3%, 87.5%, and 85.4%, respectively. Glycosidic biphenyls and dibenzofurans made up 36.7% in species of clade I, whereas their portions in clades II and III were only 5.2% and 9.8%, respectively. Furthermore, the clade I species did not have any miscellaneous structures which, however, constituted 7.3% and 4.9% in clade II and III species, respectively. The genes for biphenyl and dibenzofuran biosynthetic enzymes are thus differentially regulated in the Malinae species.

Phylogenetic distribution of biphenyl and dibenzofuran phytoalexins in the Malinae. The phylogenetic tree of the Malinae genera is modified after Sun et al. (2018). Values beside the branches are the bootstrap percentages from maximum parsimony analysis, maximum likelihood analysis, and Bayesian posterior probabilities. “-” indicates a branch collapse in the maximum parsimony and maximum likelihood trees. BP, biphenyl; DB, dibenzofuran; N, number of phytoalexins; R, ratio of the number of phytoalexins in a class to the total number of phytoalexins; Shaded in orange, plants producing biphenyls only; Green, plants producing dibenzofurans only; Grey, plants producing both biphenyls and dibenzofurans; Red, values indicating clear differences among the three clades
The co-occurrence of the two classes of compounds was reported for 13 species of nine Malinae genera, which were distributed among the three clades in the phylogenetic tree: two genera (Malus and Chaenomeles) in clade III, five genera (Photinia, Eriobotrya, Rhaphiolepis, Sorbus, and Pyrus) in clade II, and two genera (Pyracantha and Crataegus) in clade I (Fig. 2; Table 3). In these nine genera, co-occurrence of biphenyls and dibenzofurans was not found in all species. For example, Sorbus aucuparia, S. domestica, S. lanata, and S. pohaushanensis contained both classes of phytoalexins, whereas S. commixta and S. chamaemespilus had only dibenzofurans and S. decora had only biphenyls. On the other hand, two genera in clade III (Aronia and Osteomeles) contained only biphenyls, whereas one genus in clade II (Cotoneaster) and two genera in clade I (Cydonia and Mespilus) contained only dibenzofurans.
Likewise, the substitution patterns of the biphenyl and dibenzofuran basic skeletons varied among the three clades of the Malinae genera (Table 3). In the clade III species, none of the 35 biphenyls were substituted by methoxy (-OCH3) at C-4, whereas 10.9% and 17.6% of the biphenyls in clade II and I, respectively, had a methoxy moiety at C-4. In addition, 23.5%, 17.6%, 11.8%, and 11.8% of the biphenyls in species of clade I carried substitutions at positions C-2, C-6, C-5ʹ, and C-6ʹ, respectively. On the other side, no substitutions at these positions were observed for biphenyls of clade II and III species, except for 2.2% of clade II biphenyls, which had a substitution at C-5ʹ. For dibenzofurans, 33.3% of clade III species lacked a substitution at C-3 (equal to C-4 in biphenyls), whereas the C-3 position of all dibenzofurans was substituted in clades I and II. Furthermore, the C-8 position (equal to C-5ʹ in biphenyls) was free of substitution in clade III species only. Regiospecific substitutions on the basic skeletons are accomplished by modifying enzymes, such as hydroxylases, methyltransferases, and glycosyltransferases. The diversity within the individual enzyme classes is commonly due to gene duplication events. Obviously, the Malinae species differ in array and activity of genes for modifying enzymes.
The most widely distributed biphenyls are aucuparin (3), 2'-methoxyaucuparin (7), and 4'-methoxyaucuparin (9) (Table 1). Aucuparin (3) and 2'-methoxyaucuparin (7) were isolated for the first time from the healthy heartwood of S. aucuparia. Thereafter, aucuparin (3) was detected in fourteen species belonging to seven genera (Aronia, Chaenomeles, Eriobotrya, Malus, Photinia, Pyrus, Sorbus). Its role as phytoalexin was disclosed in ten species. 2'-Methoxyaucuparin (7) was isolated from fourteen species belonging to six genera (Aronia, Chaenomeles, Crataegus, Malus, Photinia and Sorbus). Its function as phytoalexin was observed in eight species. Furthermore, 4'-methoxyaucuparin (9) was detected as phytoalexin in eight species belonging to six genera (Aronia, Chaenomeles, Malus, Photinia, Rhaphiolepis, Sorbus). Interestingly, the biphenyls 5, 8, 13–22, 24–27, and 30–46 were exclusively detected in individual species (Table 1).
Regarding dibenzofurans, α-cotonefuran (47), γ-cotonefuran (49), and eriobofuran (53) are the most widely distributed compounds (Table 2). γ-Cotonfuran (49) was detected as phytoalexin in eleven species belonging to four genera (Cotoneaster, Crataegus, Pyrus, Sorbus). α-Cotonefuran (47) was found as phytoalexin in eight species belonging to three genera (Cotoneaster, Crataegus, Mespilus). Moreover, eriobofuran (53) was isolated from eight species belonging to six genera (Eriobotrya, Malus, Photinia, Pyracantha, Pyrus, Sorbus). However, its role as phytoalexin was not confirmed in Photinia lucida. Interestingly, the dibenzofurans 56, 58, 60, 62–79, 81, and 83–87 were exclusively isolated from individual species.
The function of biphenyls and dibenzofurans as phytoalexins was reported for a total of 31 out of 44 species in the subtribe Malinae (Tables 1 and 2). Eight species form only biphenyls (Aronia arbutifolia, Chaenomeles cathayensis, Chaenomeles japonica, Malus sieversii, Malus sylvestris, Photinia glabra, Pyrus pyrifolia, Rhaphiolepis umbellata), sixteen species contain only dibenzofurans (Cotoneaster acutifolius, Cotoneaster divaricatus, Cotoneaster henryanus, Cotoneaster horizontalis, Cotoneaster lacteal, Cotoneaster splendens, Cotoneaster veitchii, Crataegus monogyna, C. oblonga, Mespilus germanica, Photinia davidiana, Pyracantha. coccinea, Pyrus nivalis, Pyrus pyraster, Pyrus ussuriensis, S. chamaemespilus), and seven species have both biphenyls and dibenzofurans (Eriobotrya japonica, M. domestica, P. communis, S. aucuparia, S. pohaushanensis), including two species (Chaenomeles sinensis, S. domestica), in which the role of the biphenyls as phytoalexins remains open. In 13 out of 44 species in the subtribe Malinae (Tables 1 and 2), neither biphenyls nor dibenzofurans were examined for their role as phytoalexins. Four species produce only biphenyls (Chaenomeles speciosa, Crataegus dahurica, Osteomeles schwerinae, S. decora), three species have only dibenzofurans (Crataegus pontica, Crataegus pycnoloba, S. commixta), and six species form both classes of compounds (Crataegus pinnatifida, Photinia lucida, Pyracantha fortuneana, Pyracantha koidzumii, Rhaphiolepis indica, S. lanata).
It is noteworthy that 21 biphenyls (1–20 and 23) and 21 dibenzofurans (47–67) were reported to be produced as phytoalexins in response to stress factors (Tables 1 and 2). In contrast, the phytoalexin function of 25 biphenyls (21, 22, 24–46) and 20 dibenzofurans (68–87) has not yet been explored. Curiously, 2,8-dihydroxy-3,4,7-trimethoxydibenzofuran (52) was produced as phytoalexin in five species (C. oblonga, P. communis, P. nivalis, P. pyraster, P. ussuriensis) but constitutively formed in C. potinica.
Taken together, biphenyls and dibenzofurans are not only diverse in their chemical structures but also variable in their abundance among the species of the subtribe Malinae. Some species accumulate a wide array of biphenyls and dibenzofurans, whereas others form only a few compounds. Furthermore, different parts of the producing plants contain the phytoalexins. Some compounds were isolated from the roots of a limited number of species, whereas the majority of phytoalexins were found in the aerial parts. Most biphenyls were detected in sapwood, leaf, and fruit, whereas most dibenzofurans were restricted to the sapwood. Functionally, nearly half of the reported biphenyls (21 out of 46) and dibenzofurans (21 out of 41) were demonstrated to act as phytoalexins, indicating that these two classes of compounds are the inducible defense substances of the Malinae, although residual compounds still need to be explored for their phytoalexin role. The clade-related distribution of biphenyls and dibenzofurans and the substitution patterns of their basic skeletons are interesting issues for chemotaxonomy and evolution of the Malinae. However, we must bear in mind that the number of species explored for the formation of biphenyls and dibenzofurans (44 species) is still small compared to the total number of species present in this subtribe (1000 species). With an increasing number of species investigated, more structures of biphenyls and dibenzofurans will be identified, which in turn will allow clear-cut chemotaxonomic conclusions.
Biosynthesis of biphenyls and dibenzofurans
The biosynthetic pathway of biphenyl and dibenzofuran phytoalexins is only fragmentarily known. A completely elucidated route at both the biochemical and molecular levels is the biosynthesis of the most common biphenyl, aucuparin (3) in cell cultures of S. aucuparia (Fig. 3). Biphenyl synthase (BIS), a type III polyketide synthase, catalyzes the first committed step of the pathway to form 3,5-dihydroxybiphenyl (10) (Liu et al. 2004, 2007). O-Methyltransferase 1 (OMT1) then catalyzes methylation of 3,5-dihydroxybiphenyl (10) to form 3-hydroxy-5-methoxybiphenyl (1), which in turn is hydroxylated at the C-4 position by a cytochrome P450 enzyme, biphenyl 4-hydroxylase (B4H), to yield noraucuparin (2) (Khalil et al. 2013, 2015; Sircar et al. 2015). The hydroxy group at C-3 of noraucuparin (2) is subsequently methylated by O-methyltransferase 2 (OMT2) to form aucuparin (3). Notably, the counterparts of these aucuparin biosynthetic genes present in M. domestica were also cloned and functionally characterized (Chizzali et al. 2012b; Sircar et al. 2015; Sarkate et al. 2019).

Biosynthesis of biphenyls and dibenzofurans. Solid arrows with blue enzyme names indicate reactions established at the gene level; solid arrows with black enzyme names indicate reactions demonstrated at the biochemical level; broken arrows with red enzyme names indicate proposed reactions. B4H biphenyl 4-hydroxylase; BD benzaldehyde dehydrogenase; BIS biphenyl synthase; BS benzaldehyde synthase; BZL benzoate-CoA ligase; CHD cinnamoyl-CoA hydratase/dehydrogenase; CHL cinnamoyl-CoA hydratase/lyase; CNL cinnamoyl-CoA ligase; CYP_H cytochrome P450 hydroxylase; CYP_C cytochrome P450 cyclase; KAT ketoacylthiolase; OMT O-methyltransferase; PAL phenylalanine ammonia lyase (Abd El-Mawla and Beerhues 2002; Liu et al. 2004, 2007; Hüttner et al. 2010; Chizzali et al. 2012a, b; Khalil et al. 2013, 2015; Sircar et al. 2015; Saini et al. 2017, 2019, 2020; Sarkate et al. 2019; Teotia et al. 2019; Busnena et al. 2021; Song et al. 2021)
Based on the structures of the isolated biphenyls, 3,5-dihydroxybiphenyl (10) can undergo single or multiple hydroxylation and methylation steps at the carbons 2, 4, 2ʹ, 3ʹ, and 4ʹ. However, detailed biochemical and molecular studies are still needed to elucidate these individual steps. In S. pohaushanensis, the presence of 3,4,5-trimethoxybiphenyl (4) (Song et al. 2021) points to another O-methyltransferase which methylates the 4-hydroxy group of aucuparin (3), however, this step is also open at the enzyme and gene levels.
The conversion of biphenyls to dibenzofurans has not yet been confirmed experimentally. This intramolecular cyclization requires biphenyl 2ʹ-hydroxylation. The resulting 2ʹ-hydroxylated intermediate does not originate from salicoyl-CoA as a BIS starter substrate (Liu et al. 2010; Chizzali and Beerhues 2012). Upon salicoyl-CoA usage, all the known BIS enzymes release 4-hydroxycoumarin after a single condensation with malonyl-CoA rather than 2ʹ,3,5-trihydroxybiphenyl after three decarboxylative condensations. In S. aucuparia, the only isolated 2ʹ-hydroxylated biphenyl was 2ʹ-hydroxyaucuparin (6) (Hüttner et al. 2010) and no dibenzofuran lacking substitution at C-3 (corresponding to C-4 in biphenyls) was detected. These facts argue against the transition of 3,5-dihydroxybiphenyl (10) and 3-hydroxy-5-methoxybiphenyl (1) to the corresponding dibenzofurans. Since noreriobofuran (54) was detected in S. aucuparia cell cultures, noraucuparin (2) was believed to be the metabolic branch point, from which biphenyls derive on one side and dibenzofurans on the other side (Khalil et al. 2015). However, 2ʹ,3-dihydroxy-5-methoxybiphenyl (11) and 2ʹ-hydroxy-noraucuparin (12) have recently been detected in roots and medium of in vitro cultured and yeast-extract-treated plants of the apple rootstock M26 (Busnena et al. 2021). The profile of biphenyls in M. domestica indicates that biphenyl 2ʹ-hydroxylation can take place before hydroxylation at C-4. Interestingly, intramolecular cyclization of 2ʹ,3-dihydroxy-5-methoxybiphenyl (11) forms 2-hydroxy-4-methoxydibenzofuran (66), which was found for the first time in apple (Weiß et al. 2017a). In fact, 2-hydroxy-4-methoxydibenzofuran (66) is the predominant dibenzofuran in nearly all apple root samples studied (Reim et al. 2020; Rohr et al. 2020; Balbín-Suárez et al. 2021; Busnena et al. 2021). Thus, 3-Hydroxy-5-methoxybiphenyl (1) seems to be the target of 2ʹ-hydroxylation and the resulting product, 2ʹ,3-dihydroxy-5-methoxybiphenyl (11), seems to be the branch point of the biphenyl and the dibenzofuran biosynthetic routes in apple (Fig. 3). The cyclization of 2ʹ,3-dihydroxy-5-methoxybiphenyl (11) to form 2-hydroxy-4-methoxydibenzofuran (66) is proposed to be a major biosynthetic step in apple which, however, is absent from S. aucuparia. Another main phytoalexin in apple is the eriobofuran isomer 2 (Busnena et al. 2021), which is supposed to be synthesized through hydroxylation of 2-hydroxy-4-methoxydibenzofuran (66).
The formation of O-glycosides of biphenyl and dibenzofuran compounds is likely to be catalyzed by glycosyltransferase (GTF) enzymes. Although no public report about biphenyl and dibenzofuran GTFs was published, a phytoalexin GTF from S. pohaushanensis was described in two patents (Huang et al. 2017a, b). The biosynthesis of sorbusin A (64) and sorbusin B (65), which are sulfur and nitrogen-containing dibenzofurans, was proposed to be a condensation of cysteine with the oxidated dibenzofurans, followed by a series of reactions like cyclization, decarboxylation, re-arrangement, and methylation (Fig. 4) (Gao et al. 2021). However, this hypothesis remains to be proven experimentally. Chaenomin A (29), chaenomin B (30), berbekorin A (31), and sorbalanin (85) share a common component (4-hydroxyphenylglycerol), which is probably derived from the phenylpropanoid metabolism and attached to the biphenyl and dibenzofuran skeletons by either the 4-hydroxy group or the middle hydroxy group of glycerol. Biphenyls 38 and 39 lack the typical substitution pattern of 3,5-dihydroxybiphenyl. Instead, we propose that they arise from the C–C coupling of two phenylpropanoid units, which undergo esterification with either hexanol or heptanol and hydroxylation.

Proposed biosynthetic pathway of sulfur and nitrogen-containing dibenzofurans (Gao et al. 2021)
The biosynthesis of benzoyl-CoA, the starter substrate for BIS besides malonyl-CoA as extender, is a branch pathway of phenylpropanoid metabolism (Fig. 3). Cinnamic acid is formed from the aromatic amino acid L-phenylalanine by phenylalanine ammonium lyase (PAL). In the Malinae species, cinnamic acid is converted to benzoic acid by two pathways. The first pathway is CoA-independent and non-β-oxidative, which involves benzaldehyde synthase (BS) and benzaldehyde dehydrogenase (BD). This pathway was demonstrated at the biochemical level in P. pyrifolia (Saini et al. 2017, 2019). Alternatively, cinnamic acid is converted to cinnamoyl-CoA by cinnamoyl-CoA ligase (CNL), which is further converted to benzaldehyde through the CoA-dependent and non-β-oxidative pathway. Although cinnamoyl-CoA formation was demonstrated in apple (Teotia et al. 2019), the conversion of cinnamoyl-CoA to benzaldehyde by cinnamoyl-CoA hydratase/lyase (CHL) was only detected in Hypericum androsaemum (Abd El-Mawla and Beerhues 2002). Benzoyl-CoA ligase (BZL) activity has recently been detected at the biochemical level in cell cultures of P. pyrifolia (Saini et al. 2020).
The above findings indicate that a number of reactions related to biphenyl and dibenzofuran biosynthesis were elucidated. The enzymes that catalyze these reactions and the genes that encode the responsible enzymes were investigated. However, progress was mainly achieved with biphenyl biosynthesis, whereas dibenzofuran formation remains largely obscure. This is particularly true for the central biosynthetic steps of dibenzofuran biosynthesis, i.e. 2ʹ-hydroxylation and intramolecular cyclization to yield the tricyclic scaffold. The efforts have to be made to fully clarify the formation of the metabolic grid of biphenyl and dibenzofuran compounds.
Induction of phytoalexin formation by biotic and abiotic stimuli
Generally, phytoalexins formation is induced by biotic and abiotic factors. Abiotic inducers are not derived from biotic sources. They are stress conditions like extremely high or low temperatures, plant wounding, or exposure to UV light and heavy metals (Moesta and Grisebach 1980; Kokubun and Harborne 1994; Pedras et al. 2007). Biotic inducers refer to either intact microorganisms or extracts and molecules derived thereof (Darvill and Albersheim 1984; Khan et al. 2021). Only three studies reported phytoalexin formation in Malinae species upon elicitation by abiotic inducers, mainly heavy metals (Table 4). Mercuric chloride induced the formation of 2'-methoxyaucuparin (7) and 4'-methoxyaucuparin (9) in the leaves of P. glabra. It also stimulated the formation of rhaphiolepsin (8) and 4'-methoxyaucuparin (9) in leaves of Rhaphiolepis umbellata. Aucuparin (3) was induced in the leaves of S. aucuparia upon exposure to copper sulfate solution. Heavy metals as inducers are of ecological relevance because plants can be exposed to various heavy metals as pollutants in air, water, and soil.
In contrast to the few reports of abiotic effects, many research papers documented the formation of phytoalexins in Malinae species upon induction by biotic inducers, i.e. phytopathogens. Included are also yeast extract, harpin protein crude extract (Hpa1), and chitosan but these inducers lack ecological relevance for phytoalexins induction in plants. Nonetheless, they were used to study the phytoalexins profile of some species, mainly under cell culture conditions, aiming at understanding the structural diversity and biosynthetic route of the induced phytoalexins. In fact, fungal attack is considered a major inducer of biphenyl and dibenzofuran phytoalexins in the Malinae. Ten phytopathogenic fungal species were reported to initiate the formation of phytoalexins in this subtribe (Table 4). Nectria galligena and Phomopsis perniciosa (canker-causative agents in apple and pear) stimulated the production of the phytoalexins 3, 5–7, 9,47, 49–53, 55, 57, 59, 60 in the sapwood of 20 plant species. Notably, they induced the formation of either biphenyls (in seven plant species) or dibenzofurans (in 13 plant species). In addition, Nectria cinnabarina (coral spot and canker-causative agent) induced the formation of the biphenyls 3, 5–7, and 9 in the sapwood of S. aucuparia and the dibenzofurans 47- 51, 53, 57, and 61–63 in the sapwood of seven other plant species. Interestingly, Venturia inaequalis (scab-causative agent) was the only fungal species to induce the formation of both biphenyls and dibenzofurans in cell cultures of apple (2, 3, 53) and S. aucuparia (2, 3, 6, 53, 54).
Besides fungi, only three pathogenic bacteria were reported to induce the formation of biphenyl and dibenzofuran phytoalexins in the Malinae (Table 4). Erwinia amylovora (fire blight disease-causing bacterium) stimulated the formation of the phytoalexins 2, 3, 6, 53, and 54 in apple, P. communis, and S. aucuparia. It also elicited the formation of 3-hydroxy-5-methoxybiphenyl (1) in both apple and pear, whereas 3,4,5-trimethoxybiphenyl (4) was only found in P. communis. Moreover, Pseudomonas syringae pv. tabaci (wildfire disease-causative agent) stimulated leaves of E. japonica to produce aucuparin (3), whereas Pseudomonas syringae pv. eriobotryae (loquat stem canker-causative agent) additionally induced eriobofuran (53) formation. Yeast extract elicited an extensive accumulation of biphenyl and dibenzofuran phytoalexins. It differentially induced the production of the phytoalexins 1–3, 6, 7, 9–16, 23, 53, 54, 56, and 64–67 in four species (M. domestica, S. aucuparia, S. pouhaushanensis, P. pyrifolia). In S. aucuparia cell cultures, Harpin protein crude extract (Hpa1) induced the formation of noraucuparin (2), while chitosan, a polysaccharide derived from chitin, induced the production of the phytoalexins 2, 3, 6, 53, and 54 (Table 4).
As described above, phytopathogens are the main inducers of phytoalexin biosynthesis in the Malinae. They stimulated the formation of biphenyls and dibenzofurans differentially, in terms of identity, number, and chemical class. The differences are due to the nature of the inducer, the induction mechanism, the response of the individual plant species, and the type of the plant system used, e.g. undifferentiated cell cultures vs. highly differentiated intact plants. The role of individual phytoalexins and especially their synergism in pathogen defense largely remains to be investigated.
Apple replant-diseased soil as an in vivo model for phytoalexins induction
Replant disease is a soil-borne disease in orchards and tree nurseries. It affects many economically important species belonging to the Rosaceae, such as apple, cherry, and peach. Apple replant disease (ARD) was reported to suppress the vegetative and generative performance of apple cultivars by up to 50% and to delay the bearing of fruits on trees by 2–3 years (Mai and Abawi 1978; Winkelmann et al. 2019; Cavael et al. 2020). Although the etiology of ARD is still not fully understood, biotic factors were proven to be the major causal agents. Indeed, there are many reports of an altered microbiome, i.e., changes in the fungal and bacterial communities in ARD-affected soil, which may exacerbate or ameliorate the effects of phytopathogens. There is increasing evidence that multiple soil-borne detrimental fungi (Rhizoctonia sp., Fusarium sp., Cylindrocarpon sp., and other Nectriaceae), oomycetes (Phytophthora sp. and Pythium sp.), and nematodes (Pratylenchus penetrans) form a disease complex of variable species (Kanfra et al. 2018; Manici et al. 2018; Radl et al. 2019; Balbín-Suárez et al. 2020).
The symptoms of ARD include general stunting and reduction in biomass. The root system becomes damaged, fragile, and dark-colored. The roots of diseased apple plants contain higher amounts of phenolics and antioxidants than healthy roots (Henfrey et al. 2015; Rohr et al. 2021). Specifically, the roots accumulate higher amounts of biphenyls and dibenzofurans (Kanfra et al. 2018; Reim et al. 2020; Rohr et al. 2020; Busnena et al. 2021). These phytoalexins were detected for the first time in roots of the apple rootstock M26 when cultivated in ARD-affected soil (Weiß et al. 2017a). M26, besides other apple rootstock genotypes like B63 and MAL0595, formed the biphenyls 1–3, 6, 11 and the dibenzofurans 53, 54, 66 (Reim et al. 2020; Rohr et al. 2020; Balbín-Suárez et al. 2021). Recently, we have even detected seven biphenyls (1–3, 6, 9–11) and twenty dibenzofurans (53, 54, 66) plus seventeen tentatively identified dibenzofurans in M26 rootstock upon growth in ARD-affected soil (Busnena et al. 2021). Notably, the phytoalexins were not only accumulated inside the roots but also exuded to the root surface and the surrounding soil, where they exert effects on the microbiome (Fig. 5).

Total biphenyl and dibenzofuran phytoalexins inside and outside roots of the apple rootstock M26 cultivated in either gamma-irradiated (γARD) or untreated ARD soils. Total is the sum of phytoalexin amounts detected in rhizoplane, rhizosphere, and bulk soil (adapted from Busnena et al. 2021)
Genes involved in phytoalexin biosynthesis, such as biphenyl synthase (BIS) and biphenyl-4-hydroxylase (B4H), were upregulated in ARD samples as early as three days after planting (Weiß et al. 2017b; Reim et al. 2020; Rohr et al. 2020; Balbín-Suárez et al. 2021). Consequently, the biphenyl and dibenzofuran phytoalexin amounts increased starting from day 3 and the highest amounts were observed in the root samples after 14 days of cultivation (Fig. 6). Furthermore, the genotype-specific BIS and B4H expression patterns were consistent with the phytoalexin contents measured in roots (Weiß et al. 2017b; Reim et al. 2020). The BIS gene appeared to be particularly useful as an early biomarker for ARD because its expression pattern well correlated with the phenotypic reaction of the Malus genotypes investigated (Rohr et al. 2020).

Kanfra et al. (2018) grew the apple rootstock M26 in a sterilized matrix of perlite and sand, which was inoculated with a suspension of nematodes isolated from replant–diseased soil. The roots developed a typical ARD morphology and the concentrations of noraucuparin (2), noreriobofuran (54), 2-hydroxy-4-methoxydibenzofuran (66), and another tentatively identified dibenzofurans significantly increased (Kanfra et al. 2018).
As pointed out, replant-diseased soil stimulates apple roots to form biphenyls and dibenzofurans. The compounds are also exuded to the root surface and the surrounding soil. These findings support their role as phytoalexins in shaping the soil microbiome, which in turn influences the development and severity of ARD. However, the detailed effects of phytoalexins on the rhizosphere and soil microbial communities need to be investigated. This is also true for the possible cytotoxic effects of the phytoalexins on the producing apple roots. It also remains to be explored which specific ARD-related microbes induce the formation of the phytoalexins. The extent to which the phytoalexins inhibit the ARD causative microbes must likewise be investigated. The answers to all these questions will help understand the correlation between phytoalexin formation and ARD development. This knowledge may finally help establish measures for alleviating the ARD syndrome.
Antimicrobial activity against phytopathogens and structure–activity relationships
Biphenyl and dibenzofuran phytoalexins differentially inhibit spore germination, germ tube development, and mycelial growth of various phytopathogenic fungi. Aucuparin (3) inhibited the spore germination rate of Colletotrichum lindemuthianum (black spot disease-causative agent) by 100% at 100 µg ml−1 (Watanabe et al. 1982). At the same concentration, it inhibited the spore germination rate of Pestalotia funereal (loquat gray leaf spot-causative agent) by 70% and reduced its germ tube length to 5 µm compared to the control (210 µm) after 24 h of inoculation. This activity was significantly reduced after 48 h, suggesting that aucuparin was inactivated by P. funereal, which may account for the pathogenic character of this fungus against loquat (Watanabe et al. 1982). Eriobofuran (53) was tested against eight plant pathogenic fungi and completely inhibited spore germination and germ tube growth of P. funereal at 43.2 µg ml−1 (Miyakodo et al. 1985). 2'-Methoxyaucuparin (7) and 4ʹ-methoxyaucuparin (9) reduced the spore germination rate of Pestalotia photiniae (leaf spot-causative agent) at 40 µg ml−1 by 64% and 71%, respectively (Widyastuti et al. 1992). Moreover, rhaphiolepsin (8) exhibited significant inhibitory activity against Pestalotia sp. because it was able to strongly reduce the spore germination rate and the germ tube length at 10 µg ml−1 (Watanabe et al. 1990; Widyastuti et al. 1991b). 4ʹ-Methoxyaucuparin (9) exhibited a significant effect on the hyphal growth of Pestalotia sp. at 40 µg ml−1 (Watanabe et al. 1990; Widyastuti et al. 1991a). The phytoalexins were more active against spore germination of Pestalotia sp. with increasing oxygenation (3 vs 7, 8, 9, and 53). Furthermore, phytoalexins with a furan ring, as in eriobofuran (53), were more active than the biphenyls tested (3, 7, 8, and 9), which may be related to the accessibility to the microbial target site.
The phytoalexins 3, 6, 7, 9, 47–51, 53, 55, 57, and 61–63 affected the spore germination rate of Alternaria alternata (leaf spots, rots, and blight-causative agent), Botrytis cinerea (gray mold-causative agent), and Fusarium culmorum (seedling blight, foot rot, ear blight, stalk rot, and common root rot-causative agent) with ED50 values in the range of 12–328 µg ml−1. The dibenzofurans showed higher antifungal activity than the tested biphenyls, which was hypothesized to be related to the greater potential to penetrate the biological membrane and interfere with fungal growth (Kokubun et al. 1995a, b, c, d; Harborne 1997). Malusfuran (56), a dibenzofuran detected only in M. domestica, inhibited spore germination of V. inaequalis (scab-causative agent) at 1 mM. Interestingly, its aglycone, 2,4-dimethoxy-3,9-dihydroxydibenzofuran, showed higher toxicity to V. inaequalis than malusfuran. It inhibited spore germination of V. inaequalis at 0.5 mM, suggesting that the aglycone may be the actual phytoalexin (Hrazdina et al. 1997). Noraucuparin (2) and eriobofuran (53) had significant inhibitory activities against Physalospora piricola (fungal pathogen of apple and pear trees) with MIC values of 6.25 and 3.13 µg ml−1, respectively (Li et al. 2019).
Noraucuparin (2), aucuparin (3), and eriobofuran (53) inhibited conidial germination of V. inaequalis at 5 µM by 28%, 46%, and 34%, respectively. Interestingly, a combination of the three compounds synergistically inhibited the conidial germination of V. inaequalis by 74% (Sarkate et al. 2018). This report is the only study to explore the synergistic effects of biphenyl and dibenzofuran phytoalexins upon combination. Biphenyls 2, 3, 4, 6, 17, 18, 19, and 20 inhibited the Alternaria tenuissi growth at 100 µg ml−1 by 48.5, 51, 54, 37, 50.3, 66.4, 58.8, and 33.3%, respectively (Song et al. 2021). In addition, 4ʹ-hydroxy-3,4,5-trimethoxybiphenyl (18) and 4ʹ,3,4,5-tetramethoxybiphenyl (19) showed inhibitory activities against four crop pathogens, i.e. Fusarium graminearum (stem rot-causative agent), Helminthosporium maydis (southern corn leaf blight-causative agent in maize), Sclerotinia sclerotiorum (vegetable sclerotinia-causative agent) and Exserohilum turcicum (northern corn leaf blight-causative agent in maize) at 100 µg ml−1, with F. graminearum being inhibited up to 63% by 4ʹ-hydroxy-3,4,5-trimethoxybiphenyl (18). 4ʹ,3,4,5-Tetramethoxybiphenyl (19) affected the growth of F. graminearum and E. turcicum with inhibition rates of 59.4 and 58.8%, respectively (Song et al. 2021). This study preliminarily concluded that biphenyl substitution at C-4ʹ increased the antifungal activity of 4ʹ-hydroxy-3,4,5-trimethoxybiphenyl (18) and 4ʹ,3,4,5-tetramethoxybiphenyl (19) as compared to 2ʹ-hydroxy-3,4,5-trimethoxybiphenyl (17) and 2ʹ-hydroxyaucuparin (6). In addition, a methoxy substitution usually showed more potent antifungal effect than a hydroxy substitution in the same position, as exemplified by 3,4,5-trimethoxybiphenyl (4) and 3,4,5-trihydroxybiphenyl (20). In another study, the phytoalexins 14, 15, 64, and 65 were tested against ten plant pathogenic fungi, sorbusin A (64) showing strong inhibitory activity against Alternaria longipes (brown spot-causative agent in tobacco) with a MIC value of 12.5 µg ml−1 (Gao et al. 2021).
It should be noted that only a single study reported the inhibitory activity of biphenyl and dibenzofuran phytoalexins on phytopathogenic bacteria. When a set of thirteen biphenyls and four dibenzofurans was tested against a panel of E. amylovora strains (fire blight disease-causing bacterium), noraucuparin (2), aucuparin (3), 3,5-dihydroxybiphenyl (10), 2,4-dihydroxydibenzofuran, eriobofuran (53), and noreriobofuran (54) exhibited antibacterial activity with MIC50 values equal to 43, 138, 17, 50, 195, 173 µg ml−1, respectively. Interestingly, a general tendency regarding structure–activity relationship was observed in this study. Biphenyls had somewhat stronger antibacterial activity than the structurally related dibenzofurans, e.g. 3,5-dihydroxybiphenyl vs. 2,4-dihydroxydibenzofuran, noraucuparin vs. 3,4-dihydroxy-2-methoxydibenzofuran, and aucuparin vs. eriobofuran (Chizzali et al. 2012a).
The above observations indicate that biphenyls and dibenzofurans possess antimicrobial activity against various plant pathogens. They appear to play a role in enhancing the resistance of Malinae species against pathogenic infections. However, all the above-mentioned antimicrobial studies lacked any information about the ecological concentrations of the phytoalexins. However, testing the compounds within their ecological concentrations is critical for understanding their role as part of the disease defense arsenal. Therefore, future works should consider the ecological concentrations of the phytoalexins to obtain reliable and informative data about their in vivo antimicrobial activity. Biphenyls and dibenzofurans need to be tested both individually and in combination against various phytopathogens to draw solid conclusions about their structure–activity relationships and possible synergistic effects. Studies of the antimicrobial activity of the phytoalexins may also be extended to human pathogens, thereby using the chance to possibly develop new human medicines. However, mechanistic studies are in demand to understand how the phytoalexins exert their antimicrobial activities.
Conclusions
Biphenyls and dibenzofurans act as phytoalexins in the subtribe Malinae of the family Rosaceae upon induction by biotic and abiotic stimuli. They are preferably accumulated in the sapwood and their chemical structures are diverse. In a phylogenetic tree of the Malinae, their distribution is clade-related, and their substitution pattern is helpful for chemotaxonomy. However, the number of Malinae species explored for phytoalexins formation is still relatively small and it needs to be expanded.
Biphenyl and dibenzofuran phytoalexins exhibit differential antimicrobial activity against plant pathogenic fungi and bacteria. However, their antimicrobial activities have not yet been correlated with their ecological concentrations in specific plant tissues, which is of great interest to support their role as an efficient component of the intricate disease resistance arsenal. The insights into the synergistic effects of biphenyls and dibenzofurans against phytopathogens, their mechanisms of action, and their structure−activity relationships are still limited. Thus, many more biphenyls and dibenzofurans need to be diligently tested for their antimicrobial properties under ecological conditions before reliable conclusions can be drawn.
Although crucial biosynthetic steps leading to the formation of biphenyls were disclosed, dibenzofuran biosynthesis is largely unclear. More efforts should be undertaken to completely elucidate the metabolism of both biphenyls and dibenzofurans. This knowledge will finally provide a solid basis for manipulating phytoalexin formation using molecular breeding strategies, thereby increasing the disease resistance potential of economically valuable fruit trees, such as apple.
Author contribution statement
BB and BL performed the literature search and drafted the manuscript. BL and LB critically revised the manuscript. All authors read and approved the manuscript.
Data availability
Data sharing is not applicable to this article as no datasets were generated or analyzed in the current study.
References
Abd El-Mawla AM, Beerhues L (2002) Benzoic acid biosynthesis in cell cultures of Hypericum androsaemum. Planta 214:727–733. https://doi.org/10.1007/s004250100657
Balbín-Suárez A, Lucas M, Vetterlein D, Sørensen SJ, Winkelmann T, Smalla K, Jacquiod S (2020) Exploring microbial determinants of apple replant disease (ARD): a microhabitat approach under split-root design. FEMS Microbiol Ecol. https://doi.org/10.1093/femsec/fiaa211
Balbín-Suárez A, Jacquiod S, Rohr AD, Liu B, Flachowsky H, Winkelmann T, Beerhues L, Nesme J, Sørensen SJ, Vetterlein D, Smalla K (2021) Root exposure to apple replant disease soil triggers local defense response and rhizoplane microbiome dysbiosis. FEMS Microbiol Ecol. https://doi.org/10.1093/femsec/fiab031
Bizuneh GK (2021) The chemical diversity and biological activities of phytoalexins. Adv Tradit Med 21:31–43. https://doi.org/10.1007/s13596-020-00442-w
Busnena BA, Beuerle T, Mahnkopp-Dirks F, Winkelmann T, Beerhues L, Liu B (2021) Formation and exudation of biphenyl and dibenzofuran phytoalexins by roots of the apple rootstock M26 grown in apple replant disease soil. Phytochemistry 192:112972. https://doi.org/10.1016/j.phytochem.2021.112972
Cavael U, Diehl K, Lentzsch P (2020) Assessment of growth suppression in apple production with replant soils. Ecol Indic 109:105846. https://doi.org/10.1016/j.ecolind.2019.105846
Chen J, Shaojiang S, Jun H, Suixu X (2006) A new biphenyl glucoside from the leaves of Crataegus pinnatifida. Shenyang Yaoke Daxue Xuebao 23:430–431
Chen JJ, Luo YT, Liao CH, Chen IS, Liaw CC (2009) A new dibenzofuran and further constituents from the stems of Pourthiaea lucida with inhibitory activity on superoxide generation by neutrophils. Chem Biodivers 6:774–778. https://doi.org/10.1002/cbdv.200800118
Chizzali C, Beerhues L (2012) Phytoalexins of the Pyrinae: Biphenyls and dibenzofurans. Beilstein J Org Chem 8:613–620. https://doi.org/10.3762/bjoc.8.68
Chizzali C, Khalil MN, Beuerle T, Schuehly W, Richter K, Flachowsky H, Peil A, Hanke MV, Liu B, Beerhues L (2012a) Formation of biphenyl and dibenzofuran phytoalexins in the transition zones of fire blight-infected stems of Malus domestica cv. ’Holsteiner Cox’ and Pyrus communis cv. ’Conference’. Phytochemistry 77:179–185. https://doi.org/10.1016/j.phytochem.2012.01.023
Chizzali C, Gaid MM, Belkheir AK, Hänsch R, Richter K, Flachowsky H, Peil A, Hanke MV, Liu B, Beerhues L (2012b) Differential expression of biphenyl synthase gene family members in fire-blight-infected apple ’Holsteiner Cox’. Plant Physiol 158:864–875. https://doi.org/10.1104/pp.111.190918
Chizzali C, Gaid MM, Belkheir AK, Beuerle T, Hänsch R, Richter K, Flachowsky H, Peil A, Hanke MV, Liu B, Beerhues L (2013) Phytoalexin formation in fire blight-infected apple. Trees Struct Funct 27:477–484. https://doi.org/10.1007/s00468-012-0808-2
Chizzali C, Swiddan AK, Abdelaziz S, Gaid M, Richter K, Fischer TC, Liu B, Beerhues L (2016) Expression of biphenyl synthase genes and formation of phytoalexin compounds in three fire blight-infected Pyrus communis cultivars. PLoS ONE 11:e0158713/1–e0158713/16. https://doi.org/10.1371/journal.pone.0158713
Choi SY, Jeong B, Jang HS, Lee J, Ko K, Kim H, Hong J, Bae YS, Yang H (2018) A new dibenzofuran from the barks of Sorbus commixta. Rec Nat Prod 12:179–183. https://doi.org/10.25135/rnp.20.17.06.112
Coyne S, Litomska A, Chizzali C, Khalil MN, Richter K, Beerhues L, Hertweck C (2014) Control of plant defense mechanisms and fire blight pathogenesis through the regulation of 6-thioguanine biosynthesis in Erwinia amylovora. ChemBioChem 15:373–376. https://doi.org/10.1002/cbic.201300684
Dai Y, Zhou GX, Kurihara H, Ye WC, Yao XS (2006) Biphenyl glycosides from the fruit of Pyracantha fortuneana. J Nat Prod 69:1022–1024. https://doi.org/10.1021/np0600853
Dai Y, Zhou GX, Kurihara H, Ye WC, Yao XS (2008) Fortuneanosides G-L, dibenzofuran glycosides from the fruit of Pyracantha fortuneana. Chem Pharm Bull (Tokyo) 56:439–442. https://doi.org/10.1248/cpb.56.439
Darvill AG, Albersheim P (1984) Phytoalexins and their elicitors-a defense against microbial infection in plants. Annu Rev Plant Physiol 35:243–275. https://doi.org/10.1146/annurev.pp.35.060184.001331
Deng Y, Xu Z, Yang B, Liu Y, Dai R, Yu Y (2006) Isolation and structure identification of chemical components from root of Crataegus pinnatifida. Beijing Ligong Daxue Xuebao 26:464–467
Gao Y, Yang J, Yang XL, Zhang L, Wang J, Li Q, Lin DM, Zhang M, Xia SN, Xu LL, Zhang Q, Hai P, Liu YH, Wang S, Guo LP (2021) Novel dibenzofuran and biphenyl phytoalexins from Sorbus pohuashanensis suspension cell and their antimicrobial activities. Fitoterapia 152:104914. https://doi.org/10.1016/j.fitote.2021.104914
Harborne JB (1997) Recent advances in chemical ecology. Nat Prod Rep 14:83. https://doi.org/10.1039/np9971400083
Henfrey JL, Baab G, Schmitz M (2015) Physiological stress responses in apple under replant conditions. Sci Hortic 194:111–117. https://doi.org/10.1016/j.scienta.2015.07.034
Hrazdina G, Borejsza-Wysocki W, Lester C (1997) Phytoalexin production in an apple cultivar resistant to Venturia inaequalis. Phytopathology 87:868–876. https://doi.org/10.1094/PHYTO.1997.87.8.868
Huang X, Li D, Li L, Guo D, Ren R, Song S (2012) Isolation and identification of chemical constituents from leaves of Crataegus pinnatifida Bge. Shenyang Yaoke Daxue Xuebao 29:340–347
Huang L, Zhou L, Guo L, Li J, Wang S, Jiang J, Liu Y (2017a) Phytoalexin glycosyltransferase from Sorbus aucuparia and application. Patent
Huang L, Zhou L, Guo L, Li J, Wang S, Jiang J, Liu Y (2017b) Sequence and application of Sorbus aucuparia phytoalexin glycosyl transferase. Patent
Huang W, He J, Nisar MF, Li H, Wan C (2018) Phytochemical and pharmacological properties of Chaenomeles speciosa: An edible medicinal chinese mugua. Evid Based Complement Alternat Med. https://doi.org/10.1155/2018/9591845
Hüttner C, Beuerle T, Scharnhop H, Ernst L, Beerhues L (2010) Differential effect of elicitors on biphenyl and dibenzofuran formation in Sorbus aucuparia cell cultures. J Agric Food Chem 58:11977–11984. https://doi.org/10.1021/jf1026857
Jeandet P (2015) Phytoalexins: current progress and future prospects. Molecules 20:2770–2774. https://doi.org/10.3390/molecules20022770
Jia HL, Zhou LY, Wang S, Liu YH, Mo G, Guo LP (2018) Jasmonate signal pathway involved in phytoalexin biosynthesis in Sorbus aucuparia cell elicited by Hpa1. China J Chin Materia Medica 43:2928–2934. https://doi.org/10.19540/j.cnki.cjcmm.20180510.013
Kanfra X, Liu B, Beerhues L, Sørensen SJ, Heuer H (2018) Free-living nematodes together with associated microbes play an essential role in apple replant disease. Front Plant Sci 9:1666. https://doi.org/10.3389/fpls.2018.01666
Kemp MS, Burden RS, Loeffler RS (1983) Isolation, structure determination, and total synthesis of the dibenzofurans α- and β-pyrufuran, new phytoalexins from the wood of Pyrus communis L. J Chem Soc Perkin Trans 1:2267–2272. https://doi.org/10.1039/p19830002267
Khalil MN, Beuerle T, Müller A, Ernst L, Bhavanam VB, Liu B, Beerhues L (2013) Biosynthesis of the biphenyl phytoalexin aucuparin in Sorbus aucuparia cell cultures treated with Venturia inaequalis. Phytochemistry 96:101–109. https://doi.org/10.1016/j.phytochem.2013.09.003
Khalil MN, Brandt W, Beuerle T, Reckwell D, Groeneveld J, Hänsch R, Gaid MM, Liu B, Beerhues L (2015) O-methyltransferases involved in biphenyl and dibenzofuran biosynthesis. Plant J 83:263–276. https://doi.org/10.1111/tpj.12885
Khan M, Al Azawi TNI, Pande A, Mun BG, Da Lee S, Hussain A, Lee BH, Yun BW (2021) The role of nitric oxide-induced ATILL6 in growth and disease resistance in Arabidopsis thaliana. Front Plant Sci 12:685156. https://doi.org/10.3389/fpls.2021.685156
Kim CS, Subedi L, Kwon OK, Kim SY, Yeo EJ, Choi SU, Lee KR (2016) Isolation of bioactive biphenyl compounds from the twigs of Chaenomeles sinensis. Bioorg Med Chem Lett 26:351–354. https://doi.org/10.1016/j.bmcl.2015.12.004
Kokubun T, Harborne JB (1994) A survey of phytoalexin induction in leaves of the Rosaceae by copper ions. Z Für Naturforschung Sect C 49:628–634. https://doi.org/10.1515/znc-1994-9-1014
Kokubun T, Harborne JB (1995) Phytoalexin induction in the sapwood of plants of the Maloideae (Rosaceae): Biphenyls or dibenzofurans. Phytochemistry 40:1649–1654. https://doi.org/10.1016/0031-9422(95)00443-B
Kokubun T, Harborne JB, Eagles J, Waterman PG (1995a) Antifungal biphenyl compounds are the phytoalexins of the sapwood of Sorbus aucuparia. Phytochemistry 40:57–59. https://doi.org/10.1016/0031-9422(95)00307-S
Kokubun T, Harborne JB, Eagles J, Waterman PG (1995b) Dibenzofuran phytoalexins from the sapwood of Cotoneaster acutifolius and five related species. Phytochemistry 38:57–60. https://doi.org/10.1016/0031-9422(94)00636-8
Kokubun T, Harborne JB, Eagles J, Waterman PG (1995c) Dibenzofuran phytoalexins from the sapwood tissue of Photinia, Pyracantha and Crataegus species. Phytochemistry 39:1033–1037. https://doi.org/10.1016/0031-9422(95)00128-T
Kokubun T, Harborne JB, Eagles J, Waterman PG (1995d) Four dibenzofuran phytoalexins from the sapwood of Mespilus germanica. Phytochemistry 39:1039–1042. https://doi.org/10.1016/0031-9422(95)00160-9
Lee IS, Jung SH, Lee YM, Choi SJ, Sun H, Kim JS (2015) Phenolic compounds from the leaves and twigs of Osteomeles schwerinae that inhibit rat lens aldose reductase and vessel dilation in zebrafish larvae. J Nat Prod 78:2249–2254. https://doi.org/10.1021/acs.jnatprod.5b00469
Li JX, Li HL, Zhou LY, Huang L, Yang J, Zhang WD, Guo LP (2019) Chemical constituents from Sorbus aucuparia cell suspension treated with yeast and their antifungal activity. Nat Prod Res Dev 31:2071–2076
Lin CH, Chang HS, Liao CH, Ou TH, Chen IS, Tsai IL (2010) Anti-inflammatory biphenyls and dibenzofurans from Rhaphiolepis indica. J Nat Prod 73:1628–1631. https://doi.org/10.1021/np100200s
Lin RD, Chen MC, Liu YL, Lin YT, Lu MK, Hsu FL, Lee MH (2015) New whitening constituents from taiwan-native Pyracantha koidzumii: structures and tyrosinase inhibitory analysis in human epidermalmelanocytes. Int J Mol Sci 16:28598–28613. https://doi.org/10.3390/ijms161226115
Liu B, Beuerle T, Klundt T, Beerhues L (2004) Biphenyl synthase from yeast-extract-treated cell cultures of Sorbus aucuparia. Planta 218:492–496. https://doi.org/10.1007/s00425-003-1144-y
Liu B, Raeth T, Beuerle T, Beerhues L (2007) Biphenyl synthase, a novel type III polyketide synthase. Planta 225:1495–1503. https://doi.org/10.1007/s00425-006-0435-5
Liu B, Raeth T, Beuerle T, Beerhues L (2010) A novel 4-hydroxycoumarin biosynthetic pathway. Plant Mol Biol 72:17–25. https://doi.org/10.1007/s11103-009-9548-0
Mai WF, Abawi GS (1978) Determining the cause and extent of apple, cherry, and pear replant diseases under controlled conditions. Phytopathology 68:1540–1544
Manici LM, Kelderer M, Caputo F, Saccà ML, Nicoletti F, Topp AR, Mazzola M (2018) Involvement of Dactylonectria and Ilyonectria spp. in tree decline affecting multi-generation apple orchards. Plant Soil 425:217–230. https://doi.org/10.1007/s11104-018-3571-3
Miyakodo M, Watanabe K, Ohno N, Nonaka F, Morita A (1985) Isolation and structural determination of eriobofuran, a new dibenzofuran phytoalexin from leaves of loquat, Eriobotrya japonica L. J Pestic Sci 10:101–106. https://doi.org/10.1584/jpestics.10.101
Moesta P, Grisebach H (1980) Effects of biotic and abiotic elicitors on phytoalexin metabolism in soybean. Nature 286:710–711. https://doi.org/10.1038/286710a0
Pedras MSC, Zheng Q, Sarma-Mamillapalle VK (2007) The phytoalexins from Brassicaceae: structure, biological activity synthesis and biosynthesis. Nat Prod Commun 2:1934578X0700200. https://doi.org/10.1177/1934578x0700200315
Qiu X, Lei C, Huang L, Li X, Hao H, Du Z, Wang H, Ye H, Beerhues L, Liu B (2012) Endogenous hydrogen peroxide is a key factor in the yeast extract-induced activation of biphenyl biosynthesis in cell cultures of Sorbus aucuparia. Planta 235:217–223. https://doi.org/10.1007/s00425-011-1545-2
Radl V, Winkler JB, Kublik S, Yang L, Winkelmann T, Vestergaard G, Schröder P, Schloter M (2019) Reduced microbial potential for the degradation of phenolic compounds in the rhizosphere of apple plantlets grown in soils affected by replant disease. Environ Microbiome 14:1–12. https://doi.org/10.1186/s40793-019-0351-5
Reim S, Rohr AD, Winkelmann T, Weiß S, Liu B, Beerhues L, Schmitz M, Hanke MV, Flachowsky H (2020) Genes involved in stress response and especially in phytoalexin biosynthesis are upregulated in four Malus genotypes in response to apple replant disease. Front Plant Sci 10:1724. https://doi.org/10.3389/fpls.2019.01724
Rohr AD, Schimmel J, Liu B, Beerhues L, Guggenberger G, Winkelmann T (2020) Identification and validation of early genetic biomarkers for apple replant disease. PLoS ONE 15:e0238876. https://doi.org/10.1371/journal.pone.0238876
Rohr AD, Staudt J, Cziborra K, Fritz A, Schmitz M, Winkelmann T (2021) Split-root approach reveals localized root responses towards apple replant disease (ARD) in terms of ARD biomarker gene expression and content of phenolic compounds. Sci Hortic 286:110117. https://doi.org/10.1016/j.scienta.2021.110117
Saini SS, Teotia D, Gaid M, Thakur A, Beerhues L, Sircar D (2017) Benzaldehyde dehydrogenase-driven phytoalexin biosynthesis in elicitor-treated Pyrus pyrifolia cell cultures. J Plant Pathol 215:154–162. https://doi.org/10.1016/j.jplph.2017.06.004
Saini SS, Teotia D, Gaid M, Sircar D (2019) A new enzymatic activity from elicitor-treated pear cell cultures converting trans-cinnamic acid to benzaldehyde. Physiol Plant 167:64–74. https://doi.org/10.1111/ppl.12871
Saini SS, Gaid M, Sircar D (2020) Benzoate-CoA ligase contributes to the biosynthesis of biphenyl phytoalexins in elicitor-treated pear cell cultures. Plant Cell Rep 39:207–215. https://doi.org/10.1007/s00299-019-02484-0
Sarkate A, Saini SS, Teotia D, Gaid M, Mir JI, Roy P, Agrawal PK, Sircar D (2018) Comparative metabolomics of scab-resistant and susceptible apple cell cultures in response to scab fungus elicitor treatment. Sci Rep 8:17844. https://doi.org/10.1038/s41598-018-36237-y
Sarkate A, Saini SS, Gaid M, Teotia D, Mir JI, Agrawal PK, Beerhues L, Sircar D (2019) Molecular cloning and functional analysis of a biphenyl phytoalexin-specific O-methyltransferase from apple cell suspension cultures. Planta 249:677–691. https://doi.org/10.1007/s00425-018-3031-6
Sircar D, Gaid MM, Chizzali C, Reckwell D, Kaufholdt D, Beuerle T, Broggini GA, Flachowsky H, Liu B, Hänsch R, Beerhues L (2015) Biphenyl 4-hydroxylases involved in aucuparin biosynthesis in rowan and apple are cytochrome P450 736A proteins. Plant Physiol 168:428–442. https://doi.org/10.1104/pp.15.00074
Song SJ, Li LZ, Gao PY, Yuan YQ, Wang RP, Liu KC, Peng Y (2012) Isolation of antithrombotic phenolic compounds from the leaves of Crataegus pinnatifida. Planta Med 78:1967–1971. https://doi.org/10.1055/s-0032-1327877
Song C, Wang X, Yang J, Kuang Y, Wang Y, Yang S, Qin J, Guo L (2021) Antifungal biphenyl derivatives from Sorbus pohuashanensis leaves infected by Alternaria tenuissi and their effect against crop pathogens. Chem Biodivers 18:e2100079. https://doi.org/10.1002/cbdv.202100079
Suh WS, Park KJ, Kim DH, Subedi L, Kim SY, Choi SU, Lee KR (2017) A biphenyl derivative from the twigs of Chaenomeles speciosa. Bioorg Chem 72:156–160. https://doi.org/10.1016/j.bioorg.2017.04.003
Sun J, Shi S, Li J, Yu J, Wang L, Yang X, Guo L, Zhou S (2018) Phylogeny of Maleae (Rosaceae) based on multiple chloroplast regions: Implications to genera circumscription. Biomed Res Int 2018:7627191. https://doi.org/10.1155/2018/7627191
Teotia D, Gaid M, Saini SS, Verma A, Yennamalli RM, Khare SP, Ambatipudi K, Mir JI, Beuerle T, Hänsch R, Roy P, Agrawal PK, Beerhues L, Sircar D (2019) Cinnamate-CoA ligase is involved in biosynthesis of benzoate-derived biphenyl phytoalexin in Malus × domestica ‘Golden Delicious’ cell cultures. Plant J 100:1176–1192. https://doi.org/10.1111/tpj.14506
Termentzi A, Zervou M, Kokkalou E (2009) Isolation and structure elucidation of novel phenolic constituents from Sorbus domestica fruits. Food Chem 116:371–381. https://doi.org/10.1016/j.foodchem.2009.02.019
Uddin G, Latif A, Arfan M, Ali M, Hussain SH, Simpson TJ, Cox RJ, Choudhary MI (2013) Phytochemicals from the stem wood of Sorbus lanata (D. Don.) Schauer. Phytochem Lett 61:84–89. https://doi.org/10.1016/j.phytol.2012.10.002
Ulaszewski B, Jankowska-Wróblewska S, Świło K, Burczyk J (2021) Phylogeny of Maleae (Rosaceae) based on complete chloroplast genomes supports the distinction of Aria, Chamaemespilus and Torminalis as separate genera, different from Sorbus sp. Plants. https://doi.org/10.3390/plants10112534
Wang BB, Gao Y, Chen LY, Zhang CL, Zhang XQ, Zhang HL (2020) New constituents from the low polar fraction of the fruits of crataegus dahurica and their anti-inflammatory activity in RAW264.7 Cells. Chem Biodivers 17:e1900609. https://doi.org/10.1002/cbdv.201900609
Wang ZJ, Jin DN, Zhou Y, Sang XY, Zhu YY, He YJ, Xie TZ, Dai Z, Zhao YL, Luo XD (2021) Bioactivity ingredients of chaenomeles speciosa against microbes: characterization by LC-MS and activity evaluation. J Agric Food Chem 69:4686–4696. https://doi.org/10.1021/acs.jafc.1c00298
Watanabe K, Ishiguri Y, Nonaka F, Morita A (1982) Isolation and identification of aucuparin as a phytoalexin from Eriobotrya japonica L. Agric Biol Chem 46:567–568. https://doi.org/10.1080/00021369.1982.10865103
Watanabe K, Widyastuti SM, Nonaka F (1990) Two biphenyl compounds from Rhaphiolepsis umbellata as its phytoalexin. Agric Biol Chem 54:1861–1862. https://doi.org/10.1271/bbb1961.54.1861
Wei JJ, Wang WQ, Song WB, Xuan LJ (2020) Three new dibenzofurans from Cydonia oblonga Mill. Nat Prod Res 34:1146–1151. https://doi.org/10.1080/14786419.2018.1553877
Weiß S, Liu B, Reckwell D, Beerhues L, Winkelmann T (2017a) Impaired defense reactions in apple replant disease-affected roots of Malus domestica ’M26’. Tree Physiol 37:1672–1685. https://doi.org/10.1093/treephys/tpx108
Weiß S, Bartsch M, Winkelmann T (2017b) Transcriptomic analysis of molecular responses in Malus domestica ‘M26’ roots affected by apple replant disease. Plant Mol Biol 94:303–318. https://doi.org/10.1007/s11103-017-0608-6
Widyastuti SM, Nonaka F, Watanabe K, Maruyama E, Sako N (1991a) Accumulation and antifungal spectrum of 4ʹ-methoxyaucuparin as a new phytoalexin in Rhaphiolepsis umbellata Makino. Ann Phytopath Soc Japan 57:232–238
Widyastuti SM, Nonaka F, Watanabe K, Maruyama E, Sako N (1991b) Accumulation and antifungal spectrum of rhaphiolepsin as a second new phytoalexin in Rhaphiolepis umbellata Makino. Jpn J Phytopath 57:641–648. https://doi.org/10.3186/jjphytopath.57.641
Widyastuti SM, Nonaka F, Watanabe K, Sako N, Tanaka K (1992) Isolation and characterization of two aucuparin-related phytoalexins from Photinia glabra Maxim. Jpn J Phytopath 58:228–233. https://doi.org/10.3186/jjphytopath.58.228
Winkelmann T, Smalla K, Amelung W, Baab G, Grunewaldt-Stöcker G, Kanfra X, Meyhöfer R, Reim S, Schmitz M, Vetterlein D, Wrede A, Zühlke S, Grunewaldt J, Weiß S, Schloter M (2019) Apple replant disease: causes and mitigation strategies. Mol Biol 30:89–106. https://doi.org/10.21775/cimb.030.089
Yousuf S, Latif A, Arfan M, Choudhary MI (2010) 1,2,4-Trimethoxydibenzo[b, d]furan-3-ol. Acta Crystallogr Sect E Struct Rep. https://doi.org/10.1107/S1600536810044417
Zhao P, Guo R, Zhang YY, Zhang H, Yao GD, Lin B, Wang XB, Huang XX, Song SJ (2019) Phenylpropanoid and dibenzofuran derivatives from Crataegus pinnatifida with antiproliferative activities on hepatoma cells. Bioorg Chem 93:103354. https://doi.org/10.1016/j.bioorg.2019.103354
Zhou L, Yang J, Yang G, Kang C, Xiao W, Lv C, Wang S, Tang J, Guo L (2016) Biphenyl phytoalexin in Sorbus pohuashanensis suspension cell induced by yeast extract. Molecules 21:1180/1–1180/12. https://doi.org/10.3390/molecules21091180
Zhu Y, Fazio G, Mazzola M (2014) Elucidating the molecular responses of apple rootstock resistant to ARD pathogens: challenges and opportunities for development of genomics-assisted breeding tools. Hortic Res 1:14043/1–14043/8. https://doi.org/10.1038/hortres.2014.43
Acknowledgements
The work on biphenyl and dibenzofuran phytoalexins in the authors’ laboratory was supported by the German Research Foundation (DFG, BE 1174/19-1) and the Federal Ministry of Research and Education (BMBF) via the project BonaRes-ORDIAmur (031B0512Dand031B1070D). Belnaser A. Busnena thanks the German Academic Exchange Service (DAAD) for providing a doctoral scholarship and the Libyan Higher Education Ministry for general support.
Funding
Open Access funding enabled and organized by Projekt DEAL.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Additional information
Communicated by Gerhard Leubner.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Busnena, B.A., Beerhues, L. & Liu, B. Biphenyls and dibenzofurans of the rosaceous subtribe Malinae and their role as phytoalexins. Planta 258, 78 (2023). https://doi.org/10.1007/s00425-023-04228-7
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00425-023-04228-7