
Abstract
The tibial organ of Chloropidae and Milichiidae flies is an obscure feature with taxonomic significance. This study provides the first in-depth investigation into the ultrastructure of the hind leg of 11 genera from all Chloropidae subfamilies using a multimodal microscopy approach. The modified dermal tissue associated with the tibial organ indicates glandular function, as evidenced by the presence of secretory vesicles containing non-proteinaceous elements, potentially indicating lipidic secretion. The overall similarity of the tibial organ between Chloropidae and Milichiidae indicates a shared homology. However, the evolutionary history of this structure is still contentious due to limitations in the phylogenetic relationships of both lineages. Moreover, our findings enable future comparative investigations of other Diptera leg organs that possess secretory ability, which could be homologous across schizophoran families, but not necessarily the organs themselves.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Various insect lineages possess special secretory organs with particularly relevant roles in their biology (Betz 2010; Billen 2015). In some Chloropidae and Milichiidae flies, the hind tibia features an organ located at the inner posterior margin, covering at least one-third of the tibial length. This organ is present in both males and females and can be externally identified in some genera as an oval or rectangular pilose patch, often with a more prominent furrow. The size, shape, and color of the tibial organ are valuable features for distinguishing genera and species of the families Chloropidae and Milichiidae (Andersson 1977; Brake 2000).
The term “tibial organ” is commonly used to describe various modifications of the tibia across a wide range of fly families, as well as in Euglossini bees (Landim et al. 1965). In some Mycetophilidae, there are modified areas of the mid tibia in different genera (see Kurina et al. 2019; Kjærandsen et al. 2023), but there are conspicuous modifications in the setae of the hind tibia of Acomopterella Zaitzev with unknown function (Kallweit 2013). In Bibionidae, Sciaridae, Mycetophilidae, Keroplatidae and Lygistorrhinidae, sometimes the anteroapical depressed area of the fore tibia is referred to as a tibial organ (e.g., Fitzgerald 2004; Blagoderov et al. 2009; Hoffeins and Jaschhof 2010; Mohrig et al. 2017). In the Dolichopodidae genus Dolichopus Latreille, a tibial organ is restricted to males, consisting of a slit with a row of setae on the ventro-distal surface of the hind tibia, assumed to be related to the release of sex pheromones (Negrobov et al. 2018). Despite data from the external morphology, there are no studies featuring the internal morphology of these aforementioned leg organs of flies.
Among the schizophoran families of flies, Sepsidae feature a significant tibial modification known as the osmeterium, which is an exocrine gland on the edge of the hind tibia of males (see e.g., Pont and Meier 2002; Sǔlc 1928). Oxygenated hydrocarbon and putative isoprenoid secretion have been documented (Araujo et al. 2014). It is worth noting that despite the external similarity, the presence of the osmeteria solely in male sepsids is different from the tibial organ found in Chloropidae.
The term “tibial organ” referred in this paper was first described by Andersson (1977). Andersson (1977) coined this term for a “sensory area” recognized by other authors and reported the tibial organ in 50 of the 200 genera of Chloropidae. Subsequently, Brake (2000) documented a rather similar structure in Milichiidae. Notably, Chloropidae and Milichiidae are considered sister families (Buck 2006; Wiegmann et al. 2011), which raises the question of whether there is a strict evolutionary homology between the two structures.
The tibial organ of chloropids and milichiids is present in both sexes and so far has not been related to courtship or mating. While the majority of chloropids—a hyperdiverse taxon with over 3000 described species—exhibit the tibial organ, there are exceptions such as Chlorops Meigen and Ectecephala Macquart in which the organ is absent (Riccardi and Amorim 2020). Milichiids, on the other hand, include 18 genera and about 350 described species (Brake 2023) and the tibial organ has been detected only in Madiza Fallen (except M. Africana Brake and Papp) and in a few species of Milichiella Giglio-Tos (Brake 2000, 2023). The tibial organ has been described solely based on external morphology, while its ultrastructure and function remained unexplored until this study.
In this paper, we conduct a comparative morphology study of the hind tibia of 11 genera of Chloropidae through a multimodal microscopy approach. Our results provide the first in-depth description of the tibial organ’s ultrastructure, which indicate a glandular function. Furthermore, this study addresses the constraints and evolutive implications related to the homology of the tibial organ within Schizophora.
Materials and methods
Specimens
Voucher specimens representing all sampled species were deposited in the Museu de Zoologia da Universidade de São Paulo, Brazil (MZUSP) and Museum für Naturkunde Berlin, Germany (MfN). Historical specimens were used for the scanning electron microscopy. For the remaining techniques, fresh specimens were sampled with a sweep net, removed alive from the net using an aspirator, then transferred to glutaraldehyde. All specimens were identified by PRR using an unpublished identification key for Neotropical Chloropidae (Riccardi comm. pers.) and the key for Palearctic Chloropidae (Ismay and Nartshuk 2000). Detailed material information is presented in Table 1.
Scanning electron microscopy (SEM)
The legs were stored in ethanol 99.5% for at least 24 h, then immersed twice in a solution of hexamethyldisilazane (HMDS) Sigma-Aldrich® for 30 min each. Samples were left to dry overnight in a fume hood. The samples were mounted on aluminium stubs and gold coated for 200 s on a sputtering device. Scanning electron micrographs (SEM) were obtained using JEOL JCM-6000plus high vacuum equipment.
Light microscopy (LM)
Tibial organs were dissected in saline solution (0.1 M NaCl, 20 mM KH2PO4, and 20mMNa2HP4), under a stereoscope microscope. Samples were fixed in 3% glutaraldehyde solution for 48 h followed by cleaning in 0.1 M sodium phosphate buffer (PBS, pH 7.2) for at least 24 h. Washed samples were dehydrated in an ascending series of ethanol (70%, 80%, 90%, 95%, and 99.5%), soaked in Leica® historesin (Leica Biosystems Nussloch GmbH, Heidelberg, Germany), and sectioned (4 µm thick) with a glass knife on an automatic microtome. The sections were stained with hematoxylin (15 min) and eosin (1 min) (H&E), mounted on slides with Eukitt (Fluka) mounting medium, and photographed under a microscope coupled to a Moticam 580 digital camera (Motic, British Columbia, Canada).
Histochemistry
To detect neutral carbohydrates, sections were submitted to Periodic Acid–Shiff (PAS) reaction. The sections were incubated with a 0.4 M aqueous solution of periodic acid for 30 min, washed with distilled water and incubated in Shiff reagent for 60 min in the dark. The slides were washed with tap water for 15 min, mounted, and photographed as above.
To determine the total protein content, sections were submitted to the bromophenol blue reaction. The slides were incubated with a mercury solution of bromophenol for 135 min and washed with 0.5% acetic acid for 10 min. Afterward, the slides were washed in tap water for 5 min, mounted, and photographed.
Fluorescence microscopy
Sections were incubated for 1 h with WGA (Wheat Germ Agglutinin), FITC-conjugated (Fluorescein Isothiocyanate, Sigma-Aldrich, #3050, St. Louis, USA; 1:400) to detect glycoconjugates and polysaccharides containing β-1-4 N-acetylglucosamine residues in the tibial organ. Subsequently, sections were washed three times with PBS, and cell nuclei were stained by incubating the samples in DAPI (Diamidino-2-fenilindole, Polysciences, Inc., Warrington, USA; 1:500) for 30 min. After a triple wash, the slides were mounted in 30% sucrose solution, analyzed, and photographed using a fluorescence microscope connected to an Olympus DP71 digital camera (Olympus Optical Corp., Tokyo, Japan).
Transmission electron microscopy (TEM)
Fixed samples (see above) were post-fixed in 1% aqueous osmium tetroxide at 4 °C for 2 h then rinsed in PBS. The samples were dehydrated through an ascending series of acetone (30%, 50%, 70%, 90%, 95%, and 100% acetone, with 10–20 min in each), followed by a 24 h infiltration procedure using one part of Araldite® epoxy resin and one part of acetone. Samples were then embedded in Araldite® epoxy resin at 60 °C for 72 h. Longitudinal sections of 0.5 μm were stained using toluidine Blue for LM and digital images were obtained under a Leica M205C microscope coupled to the software Las X and assembled in Helicon Focus 6.
Ultrathin Sections (70 nm thick) were obtained using a Leica RM2235 ultramicrotome. The sections of the tibial organ were collected on copper grids (300 mesh) and stained with uranyl acetate with lead citrate. Images of TM were taken with the microscope JEOL Jem100CXII.
Results
The tibial organ of males and females of the chloropid genera we examined in this study externally consist of a dense patch of hair-like microtrichia (Fig. 1), sometimes inserted at a small depression on the hind tibia. The same pattern is observed in the remaining chloropid genera. Despite a slight variation in the general shape and density of hairs, the external morphology of the tibial organ across the genera is quite similar. Sensilla and pores were not detected on the external surface of the tibial organ cuticle of eight genera of grass flies (Table 1; Fig. 1). Furthermore, the SEM micrographs show that the microtrichia associated with the tibial organ are not modified or could serve as stalked ducts (Fig. 1H2).

Scanning electron micrographs of the tibial organ of Chloropidae, latero-posterior view. A Apotropina sp. B Chaethippus carioca. C Medeventor sp. D Dasyopa sp. E Psilochlorops paganelliae. F Thaumatomyia sp. G Trigonomma sp. H1 Dicraeus sp. H2 Dicraeus sp., details of the microtrichiae; arrow pointing to hair apex. Scale bar 50 µm
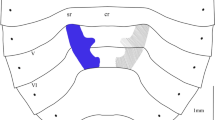
Histological sections of the mid and hind tibia of of Apotropina Hendel, Apallates Sabrosky (Fig. 2) and Olcella Enderlein (see Table 1) revealed a conspicuous glandular epithelium underneath the tibial organ, i.e., the external patch of microtrichia on the area of the tibia corresponding to the tibial organ. In contrast, the hind tibia of Ectecephala (tibial organ absent) and the mid tibia of Apotropina and Apallates have no modifications in the epithelial tissue underneath the inner posterior margin of the tibia. The epithelium is similar throughout the leg segment (as the lower margin of the hind tibia of Fig. 2A1). Furthermore, our results support the absence of reservoir.

Light Microscopy of the tibial organ of Chloropidae; histological longitudinal sections. A Apotropina sp. stained with toluidine blue; A1 hind tibia (scale bar 100 µm); A2 details of the tibial organ; A3 details of the internal tissue. B Apallates sp. stained with PAS (scale bar 25 µm); arrow pointing to a carbohydrate. C Apallates sp. stained with bromophenol (scale bar 25 µm). D Apallates sp. stained with H&E; D1 tissue. b Pilosity; cu cuticle; ms musculature; tb o tibial organ glandular tissue
The tibial organ’s epithelium exhibits PAS-positive staining (Fig. 2B). The cell nuclei have predominantly decondensed chromatin (Figs. 2D, 4D) and cytoplasm containing numerous round to oval vacuolar spaces (Fig. 2A2, 3), indicating glandular activity. Bromophenol-positive staining was not detected (Fig. 2C), as well as no WGA-FITC signal (Fig. 3). The secretory cells show mitochondria and Golgi apparatuses (Fig. 4E), with several vesicles (Fig. 4C) some of which containing lipid droplets, as evidenced by the TM preparation. Endoplasmic reticulum, neither of the smooth nor of the granular type were detected on the studied sections and the cellular membrane is shown in Fig. 4E; arrow). Therefore, according to Betz (2010) and Noirot and Quennedey (1974), these glandular cells may belong to class-1, as evidenced by Fig. 4A, B. This class indicates a low degree of modification in the glandular tissue and the absence of a reservoir.

Fluorescence Microscopy of the tibial organ of Apallates sp. (Chloropidae). A1 Hind leg stained with DAPI (scale bar 50 µm); A2 details of the tibial organ. B Hind leg stained with WGA-FITC (scale bar 50 µm). C Merged. fm Femur; tb tibia

Transmission electron micrographs of the tibial organ of Apotropina sp. (Chloropidae). A Details of the internal tissue and cuticle (scale bar 10 µm). B1 Details of the internal tissue (scale bar 5 µm); B2 cell between the cuticle and the glandular tissue. C Arrows indicating intracellular vesicles (scale bar 1 µm). D Nucleus (scale bar 1 µm). E Details of cellular membrane (scale bar 1 µm). F Arrow indicating nerve cell connected to the tibial organ internal tissue (scale bar 5 µm). cu Cuticle; g Golgi body; m mitochondria; nu nucleus
In addition, TEM preparations reveal a thin layer of cells between the cuticle and glandular tissue (Fig. 4A, B), not visible in other preparations. We observed a small nerve fiber associated to the proximal margin of the glandular tissue (Fig. 4F; arrow). Moreover, faint transversal markings in the adjacent cuticle may indicate a releasing duct (Fig. 4B2; arrow).
For providing a comprehensive micromorphological characterization of the tibial organ in chloropids, we used a variety of techniques. While the bromophenol, H&E, and DAPI/WGA-FITC staining methods only detected the nuclei of the glandular tissue (Figs. 2C, D, 3), the toluidine blue staining also highlighted the presence of numerous cellular vesicles. Although the exact nature of the vesicle content is still unknown, our results suggest the presence of lipidic secretion, as no strong evidence for protein or carbohydrate components was detected. Notably, the combined microscopy results did not provide strong evidence of a direct communication between the glandular tissue and the external surface of the tibial organ (as evidenced by the lack of WGA-FITC signal), with the exception of the microtrichia, which are positioned in close proximity to the glandular tissue, and faint transversal markings in the adjacent cuticle.
Discussion
The discovery of a previously unknown dermal gland associated with the hind leg in chloropid flies is a significant finding. Unlike our multimodal microscopy approach, preceding studies did not utilize high magnification and invasive techniques to characterize the tibial organ. The tissue morphology and the large number of vesicles suggest that the tibial organ produces lipid secretions, although the connection between the internal tissue and the external microtrichia remains elusive.
The overall internal anatomy of the tibial organ resembles the epidermal glands found in other arthropods, such as the ultimate legs of geophilomorph centipedes (Sombke and Müller 2021), wasp legs (Nijs and Billen 2015) and sepsids osmeterium (Sǔlc 1928). However, our results did not detect any stalked ducts on the microtrichia evidenced by the absence of perforations on the microtrichia apex (Fig. 1H2), nor the presence of reservoirs. Moreover, perforations (or canaliculi) on the cuticle, that would confirm an exocrine nature of the glands are not evident. Thus, the high density of microtrichia on the surface of the tibial organ makes external visualization of the cuticle challenging.
While ducts extending from the epidermis to the epicuticle are a universal feature in terrestrial hexapods, many glands of different kinds and with distinct roles have evolved through modifications of the cuticle. These gland secretions have different roles and singular evolutionary implications. Apart from the locomotory function of the hind leg of insects, the structural modification observed in in most chloropids must represent a novel function, yet to be discovered. Although the specific role of the glandular tissue and the microtrichia associated to the tibial organ remain elusive, our findings make it plausible to assume that the vesicle content may be released externally due to the position, morphology of the organ, and overall similarity of the exocrine glands of other arthropods. The presence of a nerve fiber suggests a neurosecretory control of the gland, although we still do not understand the mechanism of secretion release.
Faint longitudinal markings on the cuticle (Fig. 4B2; arrow) and substance accumulation at the base of the microtrichia (Fig. 1H2) indicate that the release mechanism of the tibial organ content may be hidden by the base of the microtrichia. Moreover, the absence of a direct connection between the microtrichia and content release indicates that the most probable function of this pilosity is to increase the friction surface (Gorb 1997). Most studies focus on the macrotrichia (innervated hairs inserted on sockets) modifications and function of insects, and there is little discussion in the literature about the microtrichia (minute cuticle projections lacking sockets) functions (Ghiradella 1998), particularly of flies. Therefore, further developments on the behavior, micromorphology and chemical content of the tibial organ vesicles are needed to unravel its function.
Other structures in the male abdomen of some chloropids, as the paired eversible vesicles of Thaumatomyia Zenker (Kotbra 2009), the sclerotized paddles of Chaetochlorops Malloch (Riccardi and Rodriguez-Sanches 2021), and the invaginations between the third and fourth tergites of Aragara Walker (Ismay 1990) seem to be related to intraspecific communication and reproduction. However, sex-related features are usually restricted to one gender, whereas both genders of chloropids and milichiids possess the tibial organ. Although the epithelial tissue of the osmeterium in sepsid flies (Sǔlc 1928) is similar to our findings, it is worth noting that the osmeterium demonstrates a distinct link to mating behavior (Araujo et al. 2014). This association is not yet observed for the tibial organ of chloropids.
The presence of tibial organ in most genera of Chloropidae strongly suggests that this feature is synapomorphy of the entire Chloropidae. However, a comprehensive phylogeny of this lineage would provide means to trace the evolution of this feature. Also, more information on the biology of the chloropid genera could indicate if the loss of the tibial organ could be related to shifts in life strategies.
Although the tibial organ in Madiza glabra Fallen (Brake 2023) is strikingly similar to the tibial organ in chloropids externally, the absence of this feature in most milichiids is intriguing. Therefore, future outputs in the micromorphological characterization of the tibial organ in Milichiidae and ancestral state reconstructions of Milichiidae + Chloropidae are required to address the origin and homology of the tibial organ within this lineage.
Since there are barely any other cases of similar organs or conspicuous glands in the hind tibia of schizophoran families, the question of evolutionary homology across the clade is to be raised. If isolated secreting glands with similar function were to be found in other milichiids—although not as a tissue and visible externally by modifications of the cuticle shape—, this would suggest homology of biochemical and cell mechanisms in both lineages. Further developments in both families of isolated glands into morphological structures with conspicuous glandular activity would, then, correspond to parallel traits between chloropids and Madiza. In fact, if we had more information on isolated glandular cells and their secretions in the legs of other schizophorans, it would be possible to better understand the origin of these independent conspicuous organs and their functions in different families.
Data availability
The raw images generated in this study are available at Figshare (https://doi.org/10.6084/m9.figshare.24872301).
References
Andersson H (1977) Taxonomic and phylogenetic studies on Chloropidae (Diptera) with special reference to Old World genera. Entomologica Scandinavica 8:5–199
Araujo DP, Tuan MJM, Yew JY, Meier R (2014) Analysing small insect glands with UV-LDI MS: high-resolution spatial analysis reveals the chemical composition and use of the osmeterium secretion in Themira superba (Sepsidae: Diptera). J Evol Biol. https://doi.org/10.1111/jeb.12420
Betz O (2010) Adhesive exocrine glands in insects: morphology, ultrastructure, and adhesive secretion. In: Byern J, Grunwald I (eds) Biological adhesive systems. Springer, Vienna, pp 111–152. https://doi.org/10.1007/978-3-7091-0286-2_8
Billen J (2015) Insect exocrine glands. Arthropod Struct Dev 5:399–400
Blagoderov V, Hippa H, Ševčík J (2009) Asiorrhina, a new Oriental genus of Lygistorrhinidae (Diptera: Sciaroidea) and its phylogenetic position. Zootaxa. https://doi.org/10.5281/zenodo.191565
Brake I (2000) Phylogenetic systematics of the Milichiidae (Diptera, Schizophora). Entomologica Scandinavica 57:1–120
Brake I. Milichiidae online; 2023. Complete URL: http://milichiidae.info/. Accessed 26 May 2023
Buck M (2006) A new family and genus of acalypterate flies from the Neotropical region, with a phylogenetic analysis of Carnoidea family relationships (Diptera, Schizophora). Syst Entomol 31:377–404
Fitzgerald SJ (2004) Evolution and classification of bibionidae (Diptera: Bibionomorpha). Ph.D. Thesis, State University, Corvallis, OR, USA
Ghiradella H (1998) Hairs, bristles, and scales. In: Harrison FW, Locke M (eds) Microscopic anatomy of invertebrates, vol 11A. Wiley-Liss, pp 257–287
Gorb SN (1997) Ultrastructural architecture of the microtrichia of the insect cuticle. J Morphol 234(1):1–10
Hoffeins C, Jaschhof M (2010) First record of Antlemon Loew (Diptera: Keroplatidae) in Baltic amber. Studia Dipterological 17:143–149
Ismay JW (1990) A new Aragara Walker (Dipt., Chloropidae) from Malaysia. Entomologist’s Monthly Mag 126:217–221
Ismay JW, Nartshuk EP (2000) Family Chloropidae. In: Papp L, Darvas B (eds) Contributions to a manual of Palaearctic Diptera (with special reference to flies of economic importance). Appendix Volume. Science Herald Press, Budapest, pp 387–429
Kallweit U (2013) Review of the Palaearctic Acomopterella Zaitzev (Diptera, Sciaroidea, Mycetophilidae). ZooKeys 269:11–32. https://doi.org/10.3897/zookeys.269.4252
Kjærandsen J, Kerr PH, Lindemann JP, Kurina O (2023) When details matter: Integrative revision of Holarctic Coelophthinia Edwards (Diptera, Mycetophilidae), including mapping of its mitogenome, leads to the description of four new pseudocryptic species. Biodivers Data J. https://doi.org/10.3897/BDJ.11.e98741
Kotbra M (2009) Male flies with yellow balls – new observations on the eversible vesicles on the postabdomen of male Thaumatomyia notata (Diptera: Chloropidae). Eur J Entomol 106:57–62
Kurina O, Hippa H, Amorim DS (2019) Notes on Manota Williston (Diptera: Mycetophilidae) from Australia and Papua New Guinea, with description of two new species. Zootaxa. https://doi.org/10.11646/zootaxa.4555.3.7
Landim CC, Stort AC, Cruz MAC, Kitajima EW (1965) Órgāo Tibial Dos Machos De Euglossini. Estudo Ao Microscópio Óptico E Eletrônico. Rev Bras Biol 25(4):323–341
Mohrig W, Kauschke E, Heller K (2017) Austrosciara Schmitz and Mjöberg, 1924, a senior synonym of Ctenosciara Tuomikoski, 1960 (Diptera: Sciaridae) and the description of a new brachypterous species in the genus. Zootaxa. https://doi.org/10.11646/zootaxa.4344.2.10
Negrobov OP, Chursina MA, Selivanova OV (2018) Morphology and morphometry of the tibial organ of males of the genus Dolichopus Latreille, 1796 (Dolichopodidae, Diptera). Acta Biologica Sibirica 4:46–51
Nijs C, Billen J (2015) Exocrine glands in the legs of the social wasp Vespula vulgaris. Arthropod Struct Dev. https://doi.org/10.1016/j.asd.2015.08.011
Noirot C, Quennedey A (1974) Fine structure of insect epidermal glands. Annu Rev Entomol 19:61–80
Pont AC, Meier R (2002) The Sepsidae (Diptera) of Europe. Fauna Entomologica Scandinavica, vol 37, pp 1–221
Riccardi PR, Amorim DS (2020) Phylogenetic relationships and classification of the Chloropinae of the world (Diptera: Chloropidae). Zool J Linn Soc. https://doi.org/10.1093/zoolinnean/zlaa007
Riccardi PR, Rodríguez-Sánchez E (2021) New records and notes on the morphology and biology of Chaetochlorops Malloch (Diptera: Chloropidae). J Insect Biodivers. https://doi.org/10.12976/jib/2021.26.2.5
Sombke A, Müller CHG (2021) When SEM becomes a deceptive tool of analysis: the unexpected discovery of epidermal glands with stalked ducts on the ultimate legs of geophilomorph centipedes. Front Zool. https://doi.org/10.1186/s12983-021-00402-3
Sǔlc K (1928) Dosud neznámá osmeteria u Sepsid (Diptera). Biol Listy 14:428–439
Wiegmann BM, Trautwein MD, Winkler IS, Barr NB, Kim JW, Lambkin C, Bertone MA, Cassel BK, Bayless KM, Heimberg AM, Wheeler BM, Peterson KJ, Pape T, Sinclair BJ, Skevington JH, Blagoderov V, Caravas J, Kutty SN, Schmidt-Ott U, Kampmeier GE, Thompson FC, Grimaldi DA, Beckenbach AT, Courtney GW, Friedrich M, Meier R, Yeates DK (2011) Episodic radiations in the fly tree of life. Proc Natl Acad Sci. https://doi.org/10.1073/pnas.1012675108
Acknowledgements
PRR is grateful to the Laboratório Multiusuário de Microscopia Eletrônica, FMRP/University of Sao Paulo, for the part of the histological preparations and the transmission electron microscopy; to the Centro para Documentação da Biodiversidade, FFCLRP/University of Sao Paulo and to the Integrated Zoological Research Laboratory/Museum für Naturkunde Berlin, for the scanning electron microscopy. PRR has a CAPES-Humboldt grant #88881.512934/2020-01; DSA has a FAPESP grant #2021/08713-2; RSA has a CAPES-PNPD grant #001.
Funding
Open Access funding enabled and organized by Projekt DEAL. This article is funded by CAPES-Humboldt, 88881.512934/2020-01, CAPES-PNPD, 001, Fundação de Amparo à Pesquisa do Estado de São Paulo, 2021/08713-2.
Author information
Authors and Affiliations
Contributions
PRR: conceptualization, collecting and identifying the samples, light and electron microscopy, analyzing the data, writing the manuscript; DSA: discussing evolutionary aspects, writing the manuscript. RSA: light and fluorescence microscopy, analyzing the data. GFM: supervision, revising the manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Raile Riccardi, P., de Souza Amorim, D., dos Santos Araújo, R. et al. Comparative morphology and function of Chloropidae (Diptera) tibial organ. Zoomorphology 143, 99–106 (2024). https://doi.org/10.1007/s00435-023-00630-7
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s00435-023-00630-7