
Key message
The Arabidopsis KASH protein SINE3 is involved in male and female gametophyte development, likely affecting the first post-meiotic mitosis in both cases, and is required for full seed set.
Abstract
Linker of nucleoskeleton and cytoskeleton (LINC) complexes are protein complexes spanning the inner and outer membranes of the nuclear envelope (NE) and are key players in nuclear movement and positioning. Through their roles in nuclear movement and cytoskeletal reorganization, plant LINC complexes affect processes as diverse as pollen tube rupture and stomatal development and function. KASH proteins are the outer nuclear membrane component of the LINC complex, with conserved C-termini but divergent N-terminal cytoplasmic domains. Of the known Arabidopsis KASH proteins, SUN-INTERACTING NUCLEAR ENVELOPE PROTEIN 3 (SINE3) has not been functionally characterized. Here, we show that SINE3 is expressed at all stages of male and female gametophyte development. It is located at the NE in male and female gametophytes. Loss of SINE3 results in a female-derived seed set defect, with sine3 mutant ovules arresting at stage FG1. Pollen viability is also significantly reduced, with microspores arresting prior to pollen mitosis I. In addition, sine3 mutants have a minor male meiosis defect, with some tetrads containing more than four spores. Together, these results demonstrate that the KASH protein SINE3 plays a crucial role in male and female gametophyte development, likely affecting the first post-meiotic nuclear division in both cases.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Sexual reproduction requires the production of haploid gametes that fuse to form a diploid zygote. Reproduction in flowering plants has evolved to use pollen tubes to deliver sperm cells to female gametes (Johnson et al. 2019). Shortly after pollination, the pollen grain is hydrated. The pollen tube penetrates the pistil and grows through the transmitting tract towards the female gametophyte, or embryo sac (Palanivelu and Tsukamoto 2011). Upon arrival at the ovule, the pollen tube is guided towards the micropylar opening where it contacts one of the two synergid cells of the embryo sac (Dresselhaus et al. 2016). The pollen tube ultimately ruptures and releases the sperm cells for fertilization (Dresselhaus et al. 2016).
Plants have a two-staged life cycle in which a haploid gametophyte generation alternates with a diploid sporophyte generation. Flowering plants have male and female multi-cellular haploid gametophytes. Haploid cells in the gametophyte undergo two to three rounds of post-meiotic mitosis, to form gametes and the accessory cells required for reproduction (Drews and Yadegari 2002). During male gametogenesis, the diploid pollen mother cell (PMC) undergoes meiosis and produces a tetrad of haploid microspores encased in callose (Twell et al. 1998; Twell 2011; Oh et al. 2011). Once released from the tetrad, the microspores grow in size and the nucleus migrates to the germ cell pole (Twell et al. 1998; Twell 2011; Oh et al. 2011). The polarized microspore undergoes an asymmetric mitotic division, termed pollen mitosis I (PMI), which produces a generative cell within the vegetative cell (Twell et al. 1998; Twell 2011; Oh et al. 2011). The generative cell then undergoes a second mitosis (PMII) to form two identical sperm cells.
During female gametogenesis, the megaspore mother cell (MMC) undergoes meiosis, resulting in four megaspores. Subsequently, three megaspores degenerate, leaving one functional megaspore (FM) (Erbasol Serbes et al. 2019). The FM then undergoes three rounds of mitosis without cytokinesis (Erbasol Serbes et al. 2019). After subsequent cellularization and polar nuclei fusion, the FM becomes a seven-celled female gametophyte, containing an egg cell, two synergid cells, a diploid central cell, and three antipodal cells, which undergo cell death before fertilization (Erbasol Serbes et al. 2019).
Similar to animals, flowering plants undergo open meiosis and mitosis. This involves breakdown of the nuclear envelope (NE), allowing for the connection of the kinetochores to the spindle fibers and for chromosome separation. The NE then reforms upon completion of nuclear division (Pradillo et al. 2019). The proteins of nucleoplasm and cytoplasm that interact with the NE are crucial for the successful completion of mitosis and meiosis (Pradillo et al. 2019). The linker of nucleoskeleton and cytoskeleton (LINC) complex is an important player in various protein interactions at the NE. LINC complexes are embedded in the NE and are composed of outer NE Klarsicht/ANC-1/Syne Homology (KASH) proteins and inner NE Sad1/UNC-84 (SUN) proteins that interact in the NE lumen. The terminal four amino acids of KASH proteins interact with the C-terminal SUN domain of the SUN proteins to form a bridge between the nucleoplasm and the cytoplasm (Starr and Fridolfsson 2010; Zhou et al. 2014).
Animals and plants have homologous SUN proteins (Graumann et al. 2010; Oda and Fukuda 2011). The outer NE KASH proteins appear to have evolved separately in plants, because they have nothing other in common with animal KASH proteins than being tail-anchored proteins with a highly conserved, immediately C-terminal short amino acid sequence (Xu et al. 2007; Zhou et al. 2012, 2014). Several plant KASH proteins have been functionally investigated in Arabidopsis, and are involved in a variety of tissues and processes (Meier et al. 2017). The plant KASH proteins WIP1-WIP3, along with their outer nuclear envelope interaction partners WIT1 and WIT2, are involved in male fertility and the movement of the pollen nucleus (Zhao et al. 2008; Zhou and Meier 2014; Zhou et al. 2015b; Moser et al. 2020) and shape and movement of the root hair nucleus (Zhou et al. 2012, 2015a; Tamura et al. 2013). The plant KASH proteins SINE1 and SINE2 are involved in stomatal development and stomatal dynamics (Gumber et al. 2019; Biel et al. 2020a; Biel et al. 2020b; Biel et al. 2022; Biel and Moser et al. 2024). In the model legume Medicago truncatula, plant KASH proteins are involved in initiation of nodulation (Newman-Griffis et al. 2019). All identified Arabidopsis KASH proteins bind to the INM-localized SUN proteins SUN1 and SUN2 (Zhou et al. 2012, 2014). The underlying assumption is that all these roles are, in analogy to the function of animal KASH proteins, related to movement or positioning of the nucleus or chromatin organization in different cellular situations.
SUN-BINDING NUCLEAR ENVELOPE PROTEIN 3 (SINE3) was identified by its plant KASH C-terminus and shown to be associated with the Arabidopsis nuclear envelope in a SUN-dependent manner, but has not been functionally investigated (Zhou et al. 2014). Here, we show that Arabidopsis SINE3 plays a role in gametophyte development. SINE3 is located at the NE in developing male and female gametophytes. Loss of SINE3 results in reduction in seed set and silique length, that is driven by defects in female gametophyte development, as sine3 mutant ovules arrest prior to the first post-meiotic mitosis. Pollen viability is significantly reduced in sine3 mutants as well, as a result of microspores arresting prior to the first post-meiotic mitosis during male gametophyte development. Together, these data indicate that SINE3 is involved in both male and female gametogenesis in Arabidopsis, and likely has a role in the first post-meiotic nuclear division.
Materials and methods
Plant material and growth
Arabidopsis thaliana (Columbia-0 ecotype) was germinated on Murashige and Skoog (MS) medium plates (Caisson Laboratories) containing 1% sucrose under constant light. Plants at the two-leaf stage were transplanted to soil and grown at an average temperature of 22–23 °C under a 16-h light/8-h dark regime. sine3-1 (SALK_032814C), sine3-2 (SALK_029812), and sine3-3 (SAIL_248_C12) were obtained from the Arabidopsis Biological Resource Center (Alonso et al. 2003; Sessions et al. 2002). The primers used to genotype the T-DNA insertion mutants are listed in Supplemental Table 1.
Cloning
The SINE3 promoter was amplified from whole seedling genomic DNA (~ 2.2 kb; primers used are listed in Supplemental Table 2). Restriction sites for enzymes SacI and SpeI were added to the 5’ and 3’ ends and the amplified fragment was digested with the appropriate restriction enzymes. The SINE3 promoter fragment (approximately 2200 basepairs upstream of the SINE3 start codon) was isolated and purified with the QIAquik PCR Purification kit (Qiagen). The isolated fragment was subsequently ligated into a pH7WGF2 Gateway vector to obtain SINE3pro@pH7WGF2 (Takagi et al. 2011). PCR-based cloning was used previously to generate β-glucuronidase (GUS) and SINE3 coding regions, which were cloned into pENTR/D-TOPO vectors (Zhou et al. 2014). By LR reaction, the GUS, SINE3, and SINE3ΔPLPT sequences were moved from the pENTR/D-TOPO to SINE3pro@pH7WGF2 to obtain the SINE3pro::GFP-GUS, SINE3pro::GFP-SINE3, and SINE3pro::GFP-SINE3ΔPLPT constructs, respectively.
Generation of transgenic plants expressing GFP-tagged constructs
Binary vectors were transformed into Agrobacterium tumefaciens strain ABI by triparental mating (Wise et al. 2006). The Agrobacterium-mediated floral dip method was used to transform either Col-0 ecotype (WT) or sine3-1 (Clough and Bent 1998). Transgenic plants were isolated on MS plates supplemented with 30 µg/mL hygromycin, and the positive transformants (T1 plants) were confirmed by using confocal microscopy to detect GFP fluorescence. Each T1 transgenic plant was a result of an independent insertional event of the T-DNA of interest within the Arabidopsis genome. Progeny from each T1 plant were grown on hygromycin selection and floral tissue was imaged to confirm presence of GFP fluorescence (T2 plants). Hemizygous or homozygous status was determined based on the amount of GFP-positive haploid pollen (50% or 100%). Plants with 100% GFP-positive pollen grains were considered homozygous. Progeny of homozygous T2 plants were again grown on hygromycin selection and pollen grains were imaged to confirm presence of fluorescence (T3 plants). In case of SINE3pro::GFP-SINE3, no T2 plant with 100% fluorescent pollen could be identified. Eighteen SINE3pro::GFP-SINE3 in sine3-1 individual transgenic lines were isolated (T1 plants). Of those, 9 SINE3pro::GFP-SINE3 in sine3-1 lines were taken to the T3 plant generation. T3 progeny from hemizygous T2 plants were grown on hygromycin selection, pollen grains were imaged to confirm presence of fluorescence. In both the T2 and T3 generations, all the plants were hemizygous. Hemizygous plants were used for the GFP imaging experiment shown in Fig. 4.
Identifying T-DNA insertional mutants
Putative insertional lines were identified using T-DNA Express, an Arabidopsis gene mapping tool created by the Salk Institute Genomic Analysis Laboratory (http://signal.salk.edu/cgi-bin/tdnaexpress). Lines where the T-DNA insertion was predicted to be in an exon or intron were selected and acquired from the Arabidopsis Biological Resource Center (ABRC). Primers used for genotyping were generated from the T-DNA Primer Design tool also created by the Salk Institute Genomic Analysis Laboratory (http://signal.salk.edu/tdnaprimers.2.html) (Supplemental Table 1). The left borders of T-DNA insertion sites were confirmed by sequencing.
Sequencing to determine the T-DNA insertion site
The DNA fragment between the left border of the T-DNA insert and the 3’ end of the SINE3 gene was cloned using the left border forward primer (BP) and the corresponding sine3-1 or sine3-2 reverse primer (RP) (Supplemental Fig. 1a). The DNA fragment was sequenced and a sequence alignment was generated to compare to the SINE3 genomic DNA sequence (Supplemental Fig. 1b).

Analysis of SINE3 mutant alleles. A Protein domain organization of SINE3. (Green) transmembrane domain helix; (gray) unknown; (numbers) amino acids. The terminal four amino acids are shown in single-letter code. B Predicted tertiary structure of SINE3. The per-residue confidence score between 0 and 100 produced by Alphafold is shown in colors; dark blue > 90, very high; blue > 70, confident; yellow > 50, low; orange < 50, very low. C Gene diagram of the SINE3 locus and insertion sites of T-DNA insertional mutants used in this study. The T-DNA insertions in sine3-1, sine3-2, and sine3-3 are depicted as triangles, and the arrows indicate the orientation of the T-DNA insert within the chromosome. Exons, red bars; introns, lines; 5’ and 3’ untranslated regions, gray bars. Numbers indicate SALK or SAIL T-DNA insertional mutant collection code. D RT-PCR determination of the expression level of full length SINE3 in sine3 mutants. Primers amplifying the coding region were used for RT-PCR from floral tissue and are listed in Supplemental Table 1
RT-PCR analysis
Flowers from Arabidopsis plants were ground in liquid nitrogen, and total RNA was extracted using RNeasy Plant Mini kit (QIAGEN). First-strand cDNA was synthesized using SuperScript III First-Strand Synthesis System (Life Technologies) and oligodT as a primer. Primers used for PCR are listed in Supplemental Table 2.
Pollen grain staining
Pollen viability was determined using a dual fluorescent stain containing propidium iodide and fluorescein diacetate (Hamilton et al. 2015). To visualize nuclei and determine male gametophyte developmental stages, fresh anthers were squashed in 3 µg/mL DAPI in pollen isolation buffer (PIB; 100 mM NaPO4, pH 7.5, 1 mM EDTA, and 0.1% [v/v] Triton X-100) and imaged (Backues et al. 2010).
Floral Staging
Flowers were selected for use in ovule experiments at approximate stages of floral development, as previously described (Alvarez-Buylla et al. 2010).
Alphafold tertiary structure analysis
The three-dimensional (3-D) structure of the AtSINE3 protein was predicted using the Alphafold Protein Structure Database (https://alphafold.ebi.ac.uk/) (Evans et al. 2021; Jumper et al. 2021; Varadi et al. 2021).
Ovule development
For the analysis of embryo sac development in wild type and sine3 mutants, ovules were fixed and cleared as previously described (Min et al. 2019) with a few modifications. Briefly, pistils of floral stage 6 to stage 12 were harvested. For fixation, dissected ovules were incubated in 4% glutaraldehyde (in 12.5 mM K3PO4, pH 6.9) for 4 h. The tissues were dehydrated through an ethanol series (10, 20, 40, 60, 80, 95, and 100% (v/v) in ddH2O) with 10 min per step and left in 100% ethanol overnight. The dehydrated tissues were subsequently cleared in 2:1 (v/v) benzyl benzoate: benzyl alcohol for 5 h, and then observed with a Nikon C2plus confocal laser microscope. Samples were excited with 561 nm wavelength and emission was detected at 566–640 nm.
Imaging GFP localization in ovules
For imaging GFP-SINE3 subcellular localization in ovules, ovules were fixed and cleared as previously described (Tofanelli et al. 2019). Briefly, pistils were excised from flowers at different developmental stages and fixed in 4% paraformaldehyde in 1X PBS solution for 2 h at room temperature. The fixed pistils were washed twice in 1X PBS for 1 min. Next, the pistils were transferred to 1 mL of ClearSee solution (10% [w/v] xylitol, 15% [w/v] sodium deoxycholate, 25% [w/v] urea) and cleared overnight at room temperature. The cleared pistils were mounted in immersion oil and imaged with a Nikon C2plus confocal laser microscope.
Aniline blue staining
Aniline blue staining was conducted as previously described (Mori et al. 2006; Wu et al. 2010). Briefly, Arabidopsis pistils from one- or two-day-old flowers after flowering were fixed in a 3:1 ethanol/acetic acid solution for at least 2 h at room temperature. The fixed pistils were then washed in distilled water three times for 5 min each. The pistils were softened in 8 M NaOH overnight at room temperature. Carefully, the softened pistils were washed in distilled water three times for 1 h each and then stained with aniline blue solution (0.1% aniline blue in 0.1 M K3PO4 buffer, pH 11) for 3 to 5 h in the absence of light. After incubation, stained pistils were carefully mounted and imaged under Nikon C1 confocal laser microscope.
β-glucuronidase staining
A. thaliana seedlings and floral tissue were fixed in 90% acetone on ice for 30 min. Samples were washed in reaction buffer (50 mM sodium phosphate buffer, pH 7.2, 0.1% [vol/vol] Triton X-100, 2 mM K3Fe(CN)6, 2 mM K4Fe(CN)6) and incubated in the GUS staining solution (50 mM sodium phosphate buffer, pH 7.2, 0.1% [v/v] Triton X-100, 2 mM K3Fe(CN)6, 2 mM K4Fe(CN)6, and 2 mM X-Gluc [GoldBio]) for 48–72 h at 37 °C. Staining solution was then removed, and the samples were washed with 70% ethanol until the tissue was cleared. Samples were imaged under a Nikon SMZ1270 stereo microscope.
Quartet analysis
The qrt1-4 mutant was obtained from the Arabidopsis Biological Research Center (SALK_024104, Francis et al. 2006). Homozygous sine3-1 was crossed with qrt1-4. The resulting F1 seeds were grown and allowed to self-fertilize. F2 seeds were screen for homozygosity for both the sine3-1 and qrt1-4 alleles via PCR genotyping (See Supplemental Table 1 for primers). In the F3 generation, sine3-1 qrt1-4 plants were used for Alexander Staining to determine viability in each quartet (Supplemental Fig. 2, Table 3).
Results
SINE3 is a plant KASH protein of unknown function
SINE3 was identified previously as a putative Arabidopsis Klarsicht/ANC-1/Syne Homology (KASH) protein (Zhou et al. 2014). SINE3 is a 193 amino acid protein that contains a transmembrane domain (TMD) and KASH tail at its C-terminus (Fig. 1A). The cytoplasmic domain at the N-terminus (amino acids 1–158) is predicted to be highly disordered and has no known domains (Fig. 1B). SINE3 is a plant-specific protein that is not deeply conserved, with homologues only found in the Brassicaceae (Poulet et al. 2017).
To identify biological roles of SINE3, we chose three T-DNA insertion mutant alleles, sine3-1 (SALK_032804C), sine3-2 (SALK_029812), and sine3-3 (SAIL_248_C12). Through sequencing, the T-DNA insertion sites were confirmed to be within the third exon for both sine3-1 and sine3-2 (Fig. 1C). Although sine3-1 and sine3-2 are independent T-DNA insertion lines, the insertion is at the same exact position, between nucleotides 690 and 691 (Supplemental Fig. 1). The parental line for sine3-2 contained an additional heterozygous T-DNA insertion in the promoter for the gene locus At4g29780, which was removed through segregation, indicating that the two original SALK lines had arisen independently. The T-DNA insertion site for sine3-3 (determined by the Salk Institute Genomic Analysis Laboratory (Alonso et al. 2003)) was at nucleotide 144 within the first intron (Fig. 1C). RT-PCR analysis revealed that no residual full-length SINE3 transcripts were detected in sine3-1, sine3-2, and sine3-3 (Fig. 1D).
Loss of SINE3 leads to a female-derived seed set defect
When growing homozygote sine3 mutant plants, a reduction in silique length was noted (Fig. 2A). Quantification showed that both seed number and silique length were significantly reduced. In WT, the average seed number per silique was 63, and the average silique length was 16.3 mm (Fig. 2B, C). In sine3-1 and sine3-2, seeds per silique were reduced to about 25, with siliques measuring on average 11.7 mm (Fig. 2B, C). In contrast, seeds per silique and silique length in sine3-3 were reduced to 52 seeds and 14.1 mm, respectively, suggesting that sine3-3 is a weaker allele (Fig. 2B, C). To determine if the seed set defect was derived from the male or female parent, we performed reciprocal crosses between WT and homozygous sine3-1 and sine3-2 mutants. When WT stigmas were pollinated with WT, sine3-1, or sine3-2 pollen, the number of seeds per silique was between 48 and 50 (Fig. 2D). In contrast, when either sine3-1 or sine3-2 stigmas were pollinated with WT, sine3-1, or sine3-2 pollen, the number of seeds per silique was significantly reduced to approximately 20 (Fig. 2D). This suggests that the reduction in seeds in the two sine3 mutant alleles was driven by defects related to the female parent.

Loss of SINE3 leads to a seed set defect. A Representative images of mature siliques in WT and indicated sine3 mutants. Scale bar = 2 mm. B-C The average number of seeds per silique (B) and silique length (C) in WT and sine3 mutants. Data are mean values ± SE (N ≥ 50 siliques per background). Asterisks denote statistical significance to WT, as determined by Student’s t test, (p < 0.001). D Number of seeds per silique after reciprocal crosses between WT and sine3-1 or between WT and sine3-2. Data are mean values ± SE (N ≥ 20 siliques per cross). Asterisks denote statistical significance to WT X WT, as determined by Student’s t test, (p < 0.001)
SINE3 is expressed in Arabidopsis seedling roots and in reproductive tissues.
To examine SINE3 expression, 2.2. kb of SINE3 promoter sequence were fused with an in-frame fusion of GFP and β-glucuronidase (GUS) and transformed into WT Arabidopsis plants (SINE3pro::GFP-GUS). In 10 day old SINE3pro::GFP-GUS WT transgenic seedlings, GUS activity was detected throughout the root, the hypocotyl, the shoot apical meristem and faintly in the cotyledon vasculature (Fig. 3A). The strongest signal was detected in and around the root meristems (insert in Fig. 3A). In the inflorescence, a GUS signal was detected in the anthers throughout floral development and in open flowers (Fig. 3B). A GUS signal was observed in unfertilized ovules, specifically in the embryo sac of the ovule, and in pollen grains (Fig. 3C–E). A GUS signal was also detected in siliques, specifically in fertilized ovules and in seeds (Fig. 3F and G). The GUS signal observed is consistent with SINE3 expression data in the eFP Browser and Arabidopsis Heat Tree Viewer expression databases (Winter et al. 2007; Boavida et al. 2011; Borges et al. 2008; Honys and Twell 2004; Qin et al. 2009; Schmid et al. 2005). In particular, the SINE3 promoter-driven GUS signal in reproductive development is consistent with publicly available transcriptomic data from these databases.

SINE3 promoter-driven GUS expression in various cell types. A-G Expression pattern of SINE3 revealed by SINE3pro::GFP-GUS transformed WT plants. GUS staining shows the overall expression in an Arabidopsis seedling [the red inset is a magnified image showing lateral root tip expression] (A), inflorescence (B), pistil (C), ovule [black dashed line outlines whole ovule] (D), pollen grain (E), seed (F), silique (G). Scale bars = 2 mm (A-B); 0.2 mm (C); 20µm (D-E); 0.25 mm (F); 1 mm (G)
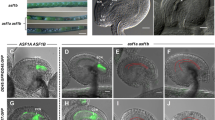
Next, hemizygous SINE3pro::GFP-SINE3 in sine3-1 transgenic plants (see Methods) were utilized to examine SINE3 expression and subcellular localization during gametophyte development. We first analyzed SINE3pro::GFP-SINE3 in sine3-1 during pollen development (Fig. 4A). GFP was not detected at the tetrad stage but was present at the NE in unpolarized and polarized microspores, bicellular pollen, and tricellular pollen. In bicellular pollen, the GFP-SINE3 signal was stronger at the vegetative NE than at the generative NE. In mature pollen grains, the GFP-SINE3 signal was only detectable at the vegetative NE (Fig. 4A).

SINE3 is expressed throughout male and female gametophyte development. A Representative microscopic images of the expression and localization of SINE3pro::GFP-SINE3 in sine3-1 in developing pollen grains at the tetrad, unpolarized microspore, polarized microspore, bicellular, and tricellular pollen stages. Tricellular pollen grain counterstained with DAPI to show the position of the vegetative nucleus (VN) and sperm cells (SCs). Scale bar = 10 μm. B Representative microscopic images of the expression and localization of SINE3pro::GFP-SINE3ΔPLPT in sine3-1 in the unpolarized microspore, bicellular, and tricellular pollen stages. Tricellular pollen grain counterstained with DAPI to show the position of the vegetative nucleus (VN) and sperm cells (SCs). Scale bar = 10 μm. C Images of developing ovules at stages FG1 to FG7. FM, functional megaspore; ACN, antipodal cell nucleus; SCN, synergid cell nucleus; ECN, egg cell nucleus; CCN, central cell nucleus. All images were captured using confocal microscopy. Scale bar = 20 μm
In the sporophyte, we have shown that the association of GFP-SINE3 with the nuclear envelope depends on the most C-terminal four amino acids (PLPT), which are required for binding SUN proteins (Zhou et al. 2014). To assess if the association with the gametophytic NE followed the same requirements, we also expressed the truncated SINE3pro::GFP-SINE3ΔPLPT in sine3-1. As shown in Fig. 4b, this led to diffuse GFP fluorescence in unpolarized microspores and at the bicellular and tricellular stage, indicating that the mechanism of SINE3 association with the nuclear envelope is the same in the sporophyte and gametophyte.
When GFP-SINE3 expression and localization was determined during female gametophyte development, GFP-SINE3 was detected at the NEs in ovules at all developmental stages (Female Gametophyte 1 (FG1) through FG7; Fig. 4C). The GFP-SINE3 fluorescent signal was detected on nuclear envelopes across all developmental stages. Together, these data show that the SINE3 promoter is specifically active in both male and female gametophytes, that SINE3 is expressed during all stages of gametophyte development, and that the protein is associated with the nuclear envelope at all stages, dependent on the last four amino acids. In pollen, nuclear envelope localization of SINE3 is limited to the vegetative cell.
sine3 mutant ovules arrest at the FG1 stage of female gametophyte development
Because the seed set defect in sine3 mutants was driven by a defect from the female, we observed WT, sine3-1, and sine3-2 ovules at various stages of female gametophyte development using whole-mount tissue clearing (Min et al. 2019). In WT, female gametogenesis proceeded normally from female gametophyte stage 1 (FG1) to FG7 (Fig. 5A–F). WT ovules at FG1 contained one functional megaspore (FM) which then underwent three rounds of mitosis without cytokinesis resulting in eight nuclei (Fig. 5A–D; FG2-FG5). Following the subsequent nuclear fusion of two polar nuclei and the degeneration of the three antipodal cells (Fig. 5E; FG6), WT ovules reached maturity (Fig. 5F; FG7). In the sine3-1 and sine3-2 mutants, female gametogenesis proceeded normally in approximately half of the ovules, however, the other half appeared to arrest at FG1 (Fig. 5G–L for sine3-1 and Fig. 5M–R for sine3-2). When analyzing the female gametophytes at the mature stage in WT, sine3-1, and sine3-2, 95.4% of WT gametophytes reached the FG7 stage (Fig. 5F; Table 1). By contrast, only 47% of the sine3-1 ovules and 47.8% of the sine3-2 ovules reached the FG7 stage, while 51.4% of sine3-1 and 51% of sine3-2 ovules were arrested at FG1 (Fig. 5L, R; Table 1). Taken together, these results indicate that SINE3 plays an important role at the onset of the nuclear divisions during female gametophyte development and that the mutant phenotype has an about 50% penetrance.

Loss of SINE3 leads to defects in female gametophyte development. Ovules at different developmental stages in WT (A-F), sine3-1 (G-L), and sine3-2 (M-R): FG1 (A, G, M), FG2/3 (B, H, N), FG4 (C, I, O), FG5 (D, J, P), FG6 (E, K, Q), FG7 (E, L, R). DM, degenerated megaspore; FM, functional megaspore; ECN, egg cell nucleus; PN, polar nucleus; ACN, antipodal cell nucleus; CCN, central cell nucleus; SCN, synergid cell nucleus; V, vacuole. Bright white spots indicate nucleoli. Scale bar = 20 µm
Loss of SINE3 leads to a pollen viability defect
Approximately half of the genes identified as functioning in gametogenesis are required for both female and male gametophyte development (Pagnussat et al. 2005). Thus, we next determined if the loss of SINE3 also affected the male gametophyte. Using a dual fluorescent stain containing propidium iodide and fluorescein diacetate (Hamilton et al. 2015), we observed a reduction in pollen viability in sine3-1 and sine3-2 mutants. Many of the nonviable pollen grains in sine3-1 and sine3-2 mutants were shriveled or collapsed (Fig. 6A, red arrows; white arrows mark non-viable normal-size pollen grains). 53.9% of sine3-1 pollen grains (n = 1264) and 50.7% of sine3-2 pollen grains (n = 1269) were viable, compared to 92% of WT pollen grains (n = 1310) (Fig. 6B).

Loss of SINE3 perturbs pollen development and leads to defects in viability. A-B Representative images (A) and quantification of viability (B) of WT and sine3 mutant pollen after hydration. Pollen was incubated for 30 min in pollen germination media containing fluorescein diacetate (FDA) and propidium iodide (PI), dyes that stain viable and non-viable pollen, respectively. White arrows mark non-viable normal-size pollen grains and red arrows mark non-viable shriveled or collapsed pollen grains. N > 1260 pollen grains per background. Scale bar = 25 μm. (C) Aniline blue-stained pollen tubes in self-pollinated WT and sine3-1 pistils. The white dotted lines indicate the pollen tube growth front in the pistils. Scale bar = 100 µm. (D) Developing spores and pollen grains stained with DAPI from WT (D-H) and sine3-1 (I-R) plants at tetrad (TET; D, I, N), unpolarized microspore (UMS; E, J, O), polarized microspore (PMS; F, K, P), bicellular (BC; G, L Q), and tricellular (TC; H, M, R) pollen stages. Developing sine3-1 pollen grains were split into 2 groups: pollen exhibiting WT-like development (I-M) and pollen exhibiting abnormal development (N-R). Images in each column correspond to the stages indicated above, with the exception of P and Q, which were present at the UMS, PMS, and BC stages
We then outcrossed the homozygous sine3-1 mutant to a quartet1 (qrt1-4) mutant (Francis et al. 2006). In qrt1-4 plants, the four products of a single meiosis remain attached to each other throughout pollen development. Alexander staining showed that the qrt1-4 plant produced four attached yet otherwise normal mature pollen grains (Supplemental Fig. 2). However, between zero and four shriveled and nonviable pollen grains were detected in tetrads from the F3 sine3-1 qrt1-4 homozygous double mutant plants at an about equal distribution (Supplemental Fig. 2 and Supplemental Table 3), suggesting that the male defect occurs with about 50% penetrance and occurs randomly after meiosis. These data also show that the gametophyte development phenotype is linked to the sine3-1 allele, which was followed in this cross by PCR detection of the T-DNA insertion to the F3 generation.
When self-pollinated Arabidopsis pistils from WT and sine3-1 were fixed and stained with aniline blue to observe pollen tube growth within the transmitting tract, abundant sine3-1 pollen tubes germinated and grew successfully through the stigma and the pollen tube transmitting tract, and neared the ovules throughout the pistil, similar to WT (Fig. 6C). This indicates that the viable sine3-1 pollen grew normally and that pollen tube growth and guidance were normal in sine3-1 pistils.
Together, these data suggest that loss of SINE3 also perturbs male gametophyte development. Like the female gametophyte defect, the male defect occurs with about 50% penetrance and occurs randomly after meiosis. This defect had no effect on the male reciprocal cross (see Fig. 2D), likely because viable pollen was present in abundance.
sine3 mutant microspores arrest prior to pollen mitosis I
Based on the significant decrease in pollen viability and the collapsed pollen phenotype observed at the mature pollen stage, we wanted to determine when the initial defect appears during pollen development. We thus examined DAPI-stained microspores dissected from staged anthers of WT and sine3-1 mutant plants (Fig. 6D-R; Table 2). A typical progression of pollen development was observed in WT plants (Fig. 6D-H; Table 2). Meiosis resulted in a tetrad of 4 equally sized microspores (Fig. 6D). Upon callose degradation, the microspores were released from the tetrad (Fig. 6E). The microspore became polarized with the migration of the nucleus to the germ cell pole (Fig. 6F). The polarized microspore underwent an asymmetric mitotic division (pollen mitosis I, PMI) to form the germ cell nucleus and vegetative nucleus (Fig. 6G). A second symmetric mitotic division (pollen mitosis II, PMII) of the generative cell nucleus generated two sperm cells (Fig. 6H).
In sine3-1 plants, aberrant phenotypes were first evident at the tetrad stage (Fig. 6I and N). WT had 100% normal tetrads composed of four equally sized microspores (Fig. 6D). In sine3-1, the majority of tetrads (91.5%) were also normal (Fig. 6I; Table 2) However 8.5% of sine3-1 tetrads contained 5 or 6 spores that were not equally sized (Fig. 6N; Table 2). At later developmental stages, a population of abnormally small microspores was observed in sine3-1 (Fig. 6O, P, Table 2), in addition to normal microspores (Fig. 6J; Table 2). Nuclear migration towards the future germ cell pole occurred in sine3-1 mutant microspores but appeared slightly delayed (Fig. 6K, Q; Table 2). Only 56% of microspores underwent PMI to form bicellular pollen (Fig. 6L; Table 2), with a large proportion of microspores remaining polarized (Fig. 6Q; Table 2). The WT-like sine3-1 bicellular pollen underwent PMII to form tricellular pollen containing a vegetative nucleus and 2 sperm cells (Fig. 6M; Table 2). The proportion of aberrantly small microspores, and unpolarized and polarized microspores observed at the bicellular pollen stage accounts roughly for the proportion of collapsed pollen grains observed at the mature pollen stage (44%) (Fig. 6R; Table 2).
Together, these data show that SINE3 is also required for male gametophyte development and that the mutation leads to a variety of defects. Most prominent is the inability of mutant pollen to progress from the polarized microspore to the bicellular pollen stage, in other words to complete pollen mitosis I. Like in the female, this phenotype has about 50% penetrance, with remaining microspores developing to fully mature, viable pollen grains capable of pollen tube growth, female signal perception, and fertilization.
Discussion
In this study, we have shown that the plant KASH protein SINE3 plays an important role in plant gametophyte development. Both male and female gametophyte development are defective in sine3 mutants. Approximately half of the male gametophytes of sine3 were arrested at the polarized microspore stage prior to pollen mitosis I. Less than 50% of sine3 female gametophytes completed the three rounds of mitosis to form mature female gametes. The remaining ovules were arrested at the FG1 developmental stage, prior to the first post-meiotic mitosis. SINE3 was expressed in male and female gametophytes and located at the nuclear envelope (NE) at all developmental stages. Together, these data suggest that SINE3 is important for the initiation of the first post-meiotic nuclear/cell division in both female and male gametophytes. The incomplete penetrance of the mutant phenotype observed here has been reported in other reproductive mutants, indicating that this is not an unusual phenomenon among Arabidopsis reproductive mutants (Chen and McCormick 1996; Howden et al. 1998; Park et al. 1998).
Previous studies have dissected the mechanisms of male and female gametophyte development through mutant identification and characterization. In gemini pollen 1 (gem1) mutant microspores, nuclear migration towards the future germ cell pole was impaired, thus resulting in similar daughter cells and failed germ cell differentiation (Park et al. 1998). gem1 was later identified as an allele of MOR1, which is a member of the microtubule-associated protein (MAP) 215 family (Whittington et al. 2001; Twell et al. 2002; Oh et al. 2010b). In microspores of sidecar pollen (scp) mutants, nuclear migration occurs normally, but a proportion of pollen contains an extra vegetative cell, attributed to a delay in nuclear division and an altered spindle orientation (Chen and McCormick 1996; Oh et al. 2010c, 2011). Unlike gem1 and scp mutants, defective sine3 mutant microspores did not undergo division and the nucleus remained polarized.
A plethora of other female gametophyte mutants that affect cell cycle progression have been identified and many have a similar phenotype to sine3 mutants (Elliott et al. 1996; Baker et al., 1997; Christensen et al. 1998; Springer et al. 2000; Acosta-Garcia and Vielle-Calzada 2004; Huanca-Mamani et al. 2005; Colombo et al. 2008; Latrasse et al. 2008; Liu et al. 2008; Gallois et al. 2009; Backues et al. 2010; Sankaranarayanan et al. 2020; Qin et al. 2022). These mutants affect fundamental cellular processes, such as gene expression and regulation (Elliott et al. 1996; Baker et al., 1997; Huanca-Mamani et al. 2005; Colombo et al. 2008; Latrasse et al. 2008), DNA replication (Springer et al. 2000), and protein degradation (Liu et al. 2008; Gallois et al. 2009).
A mutation with similar characteristics to sine3-1 and sine3-2 is mos7-5, a mutant allele of the nucleoporin MOS7 (Modifier of Snc1, 7), the Arabidopsis homolog of Nup88 (Park et al. 2014). Nuclear migration in mos7-5 mutant microspores occurs normally but arrests at pollen mitosis I, similar to sine3 mutants. Loss of MOS7 also affected female gametophyte development, similar to sine3 mutants, with developing female gametophytes arrested at FG1 (Park et al. 2014). MOS7 was shown to be required for spindle assembly during pollen mitosis I and is localized at the spindle during mitosis (Park et al. 2014).
Like MOS7, SINE3 is a NE-associated protein, currently of unknown function in reproductive development. SINE3 is a plant KASH protein, the ONM component of plant LINC complexes (Zhou et al 2014). LINC complexes, which facilitate nuclear movement and nuclear positioning, span the nuclear envelope, with KASH proteins frequently interfacing directly and indirectly with the cytoskeleton (reviewed in Meier et al. 2017). At this point, it is not known at which step of the male and female gametophyte cell cycles the SINE3 gene product acts. However, based on its nuclear envelope localization and its dual role in both gametophytes, one attractive hypothesis is that the protein is directly involved in a process required for the initiation of the first post-meiotic mitosis. In land plants, that do not have centrosomes, the nuclear envelope is involved in forming the microtubule organizing centers (MTOCs) for the outgrowth of the spindle apparatus at the onset of prophase (Zhang and Dawe 2011; Masoud et al. 2013). Further work is required to investigate this step specifically, e.g. by imaging fluorescently labeled microtubules in sine3 mutants during pollen mitosis I (Oh et al. 2010a).
Sine3 mutants also displayed a minor male meiosis defect, with approximately 8.5% of tetrads containing more than four spores (Table 2). In animals, LINC complexes have been shown to be involved in meiosis, in particular the KASH protein KASH5, which associates with Dynein (Morimoto et al. 2012; Horn et al. 2013; Agrawal et al. 2022). In plants, a KASH protein that functions during meiosis has not yet been identified. However, there is a precedent for a LINC-complex role in plant meiosis because in Oryza sativa and Arabidopsis, double mutants of SUN1 and SUN2 have severe meiotic defects, such as delayed meiotic progression, an absence of full synapsis, unresolved interlock-like structures, and a reduction in the mean cell chiasma frequency (Zhang et al. 2020; Varas et al. 2015). The meiotic defect reported here suggests that SINE3 too plays a minor role during meiosis.
Unlike many other gametophyte mutants, the function of SINE3 appears to be specific to the gametophyte. SINE3 is expressed in sporophytic tissues, such as the root and shoot apical meristems, but the homozygous sine3 mutants did not exhibit obvious growth defects. One hypothesis is that the presence of other proteins, which are expressed in the sporophyte but not the gametophyte, are acting redundantly to SINE3. Alternatively, the currently unknown molecular role of SINE3 might point towards a step required for the onset of gametophytic, but not sporophytic mitosis. For example, pollen mitosis I is an asymmetric division that produces a cell within a cell and occurs in the absence of a preprophase band (Terasaka and Niitsu 1995; Oh et al. 2010b, 2010c). Female gametophyte mitosis occurs in the absence of cytokinesis, producing a coenocyte. Thus, important regulatory or mechanistic steps might be unique to gametophyte mitosis. Investigating the molecular roles of SINE3 might shed light on such a mechanism.
A limitation of this study was the inability to recover any sine3-1 transgenic lines that were homozygous for SINE3pro::GFP-SINE3 or SINE3pro::GFP-SINE3ΔPLPT. Eighteen SINE3pro::GFP-SINE3 in sine3-1 and 13 SINE3pro::GFP-SINE3ΔPLPT in sine3-1 individual transgenic lines were isolated. Of those lines, 9 SINE3pro::GFP-SINE3 in sine3-1 and 3 SINE3pro::GFP-SINE3ΔPLPT in sine3-1 lines were further characterized. In both the T2 and T3 generations, all plants were hemizygous for the GFP-fusion construct. The inability to isolate a homozygous line seems to suggest that two copies of either SINE3pro::GFP-SINE3 or SINE3pro::GFP-SINE3ΔPLPT are detrimental. Possibly a dose-dependent dominant-negative or neomorphic effect of the fusion proteins is at play. Despite this limitation the two independently isolated mutant alleles sine3-1 and sine3-2 as well as the weaker allele sine3-3 (5’ intron insertion) all recapitulate the same phenotype. Sine3-1 was outcrossed from two, sine3-2 from one additional T-DNA insertion present in the original germplasm. Sine3-1 was additionally crossed with the qrt1-4 mutant. This strongly suggests that the disruption of the SINE3 open reading frame is the cause of the mutant phenotypes of the three lines.
Taken together, we provide evidence that the plant KASH protein SINE3 is involved in male and female gametophyte development, as related to male meiosis and male and female mitosis. Further examination of sine3 mutant defects in meiosis and mitosis will shed light on the molecular function of SINE3 and its potential interactors.
Author contribution statement
IM, NRG, and MM designed the experiments. MM performed and analyzed the experiments. MM, NRG, and IM wrote and edited the manuscript and IM provided oversight and funding for the study.
Data availability
All original data files will be made available upon request.
References
Acosta-Garcia G, Vielle-Calzada JP (2004) A classical arabinogalactan protein is essential for the initiation of female gametogenesis in Arabidopsis. Plant Cell 16:2614–2628
Agrawal R, Gillies JP, Zang JL, Zhang J, Garrott SR, Shibuya H, Nandakumar J, DeSantis ME (2022) The KASH5 protein involved in meiotic chromosomal movements is a novel dynein activating adaptor. eLife 11:e78201
Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen H, Shinn P, Stevenson DK, Zimmerman J, Barajas P, Cheuk R et al (2003) Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301:653–657
Alvarez-Buylla ER, Benitez M, Corvera-Poire A, Chaos Cador A, de Folter S, Gamboa de Buen A, Garay-Arroyo A, Garcia-Ponce B, Jaimes-Miranda F, Perez-Ruiz RV, Pineyro-Nelson A, Sanchez-Corrales YE (2010) Flower development. Arabidopsis Book 2010(8):e0127
Backues SK, Korasick DA, Heese A, Bednarek SY (2010) The Arabidopsis dynamin-related protein 2 family is essential for gametophyte development. Plant Cell 22:3218–3231
Biel A, Moser M, Meier I (2020a) A role for plant KASH proteins in regulating stomatal dynamics. Plant Physiol 182:1100–1113
Biel A, Moser M, Meier I (2020b) Arabidopsis KASH proteins SINE1 and SINE2 are involved in microtubule reorganization during ABA-induced stomatal closure. Front Plant Sci 11:575573
Biel A, Moser M, Groves NR, Meier I (2022) Distinct roles for KASH proteins SINE1 and SINE2 in guard cell actin reorganization, calcium oscillations, and vacuolar remodeling. Front Plant Sci 13:784342
Boavida LC, Borges F, Becker JD, Feijo JA (2011) Whole genome analysis of gene expression reveals coordinated activation of signaling and metabolic pathways during pollen-pistil interactions in Arabidopsis. Plant Physiol 155:2066–2080
Borges F, Gomes G, Moreno N, McCormick S, Feijo JA, Becker JD (2008) Comparative transcriptomics of Arabidopsis sperm cells. Plant Physiol 148:1168–1181
Chen Y-CS, McCormick S (1996) sidecar pollen, an Arabidopsis thaliana male gametophytic mutant with aberrant cell divisions during pollen development. Development 122:3243–3253
Christensen CA, Subramanian S, Drews GN (1998) Identification of gametophytic mutations affecting female gametophyte development in Arabidopsis. Dev Biol 202:136–151
Clough SJ, Bent AF (1998) Floral dip: a simplified method for agrobacterium-mediated transformation of Arabidopsis thaliana. Plant J 16:735–743
Colombo M, Masiero S, Vanzulli S, Lardelli P, Kater MM, Colombo L (2008) AGL23, a type I MADS-box gene that controls female gametophyte and embryo development in Arabidopsis. Plant J 54:1037–1048
Dresselhaus T, Sprunck S, Wessel GM (2016) Fertilization mechanisms in flowering plants. Curr Biol 26:R125–R139
Drews GN, Yadegari R (2002) Development and function of the angiosperm female gametophyte. Annu Rev Genet 36:99–124
Elliott RC, Betzner AS, Huttner E, Oakes MP, Tucker WQ, Gerentes D, Perez P, Smyth DR (1996) AINTEGUMENTA, an APETALA2-like gene of Arabidopsis with pleiotropic roles in ovule development and floral organ growth. Plant Cell 8:155–168
Erbasol Serbes I, Palovaara J, Groß-Hardt R (2019) Development and function of the flowering plant female gametophyte. Curr Top Dev Biol 131:401–434
Evans R, Pritzel A, Green T, Figurnov M, Ronneberger O, Tunyasuvunakool K, Bates R, Žídek A, Potapenko A et al (2021) Highly accurate protein structure prediction with AlphaFold. Nature 596:583–589
Francis KE, Lam SY, Copenhaver GP (2006) Separation of Arabidopsis pollen tetrads is regulated by QUARTET1, a pectin methyltransferase gene. Plant Physiol 142:1004–1013
Gallois J-L, Guyon-Debast A, Lécureuil A, Vezon D, Carpentier V, Bonhomme S, Guerche P (2009) The Arabidopsis proteasome RPT5 subunits are essential for gametophyte development and show accession-dependent redundancy. Plant Cell 21:442–459
Graumann K, Runions J, Evans DE (2010) Characterization of SUN-domain proteins at the higher plant nuclear envelope. Plant J 61:134–144
Gumber HK, McKenna JF, Tolmie AF, Jalovec AM, Kartick AC, Graumann K, Bass HW (2019) MLKS2 is an ARM domain and F-actin-associated KASH protein that functions in stomatal complex development and meiotic chromosome segregation. Nucleus 10:144–166
Hamilton ES, Jensen GS, Maksaev G, Katims A, Sherp AM, Haswell ES (2015) Mechanosensitive channel MSL8 regulates osmotic forces during pollen hydration and germination. Science 350:438–441
Honys D, Twell D (2004) Transciptome analysis of haploid male gametophyte development in Arabidopsis. Genome Biol 5:R85
Horn HF, Kim DI, Wright GD, Wong ESM, Stewart CL, Burke B, Roux KJ (2013) A mammalian KASH domain protein coupling meiotic chromosomes to the cytoskeleton. J Cell Biol 202:1023–1039
Howden R, Park SK, Moore JM, Orme J, Grossniklaus U, Twell D (1998) Selection of T-DNA-tagged male and female gametophytic mutants by segregation distortion in Arabidopsis. Genetics 149:621–631
Huanca-Mamani W, Garcia-Aguilar M, León-Martínez G, Grossniklaus U, Vielle-Calzada J-P (2005) CHR11, a chromatin-remodeling factor essential for nuclear proliferation during female gametogenesis in Arabidopsis thaliana. Proc Natl Acad Sci USA 102:17231–17236
Johnson MA, Harper JF, Palanivelu R (2019) A fruitful journey: pollen tube navigation from germination to fertilization. Annu Rev Plant Biol 70:809–837
Jumper J, Evans R, Pritzel A, Green T, Figurnov M, Ronneberger O, Tunyasuvunakool K, Bates R, Zidek A, Potapenko A, Bridgland A, Meyer C, Kohl SAA, Ballard AJ, Cowie A, Romera-Paredes B, Nikolov S, Jain R, Adler J, Back T, Petersen S, Reiman D, Clancy E, Zielinski M, Steinegger M, Pacholska M, Berghammer T, Bodenstein S, Silver D, Vinyals O, Senior AW, Kavukcuoglu K, Kohli P, Hassabis D (2021) Highly accurate protein structure prediction with AlphaFold. Nature 596:583–589
Latrasse D, Benhamed M, Henry Y, Domenichini S, Kim W, Zhou DX, Delarue M (2008) The MYST histone acetyltransferases are essential for gametophyte development in Arabidopsis. BMC Plant Biol 8:121
Liu J, Zhang Y, Qin G, Tsuge T, Sakaguchi N, Luo G, Sun K, Shi D, Aki S, Zheng N, Aoyama T, Oka A, Yang W, Umeda M, Xie Q, Gu H, Qu L-J (2008) Targeted degradation of the cyclin-dependent kinase inhibitor ICK4/KRP6 by RING-type E3 ligases is essential for mitotic cell cycle progression during Arabidopsis gametogenesis. Plant Cell 20:1538–1554
Masoud K, Herzog E, Chabouté M-E, Schmit A-C (2013) Microtubule nucleation and establishment of the mitotic spindle in vascular plant cells. Plant J 75:245–257
Meier I, Richards EJ, Evans DE (2017) Cell biology of the plant nucleus. Annu Rev Plant Biol 68:139–172
Min Y, Frost JM, Choi Y (2019) Nuclear chaperone ASF1 is required for gametogenesis in Arabidopsis thaliana. Sci Rep 9:13959
Mori T, Kuroiwa H, Higashiyama T, Kuroiwa T (2006) Generative cell specific 1 is essential for angiosperm fertilization. Nature Cell Biol 8:64–71
Morimoto A, Shibuya H, Zhu X, Kim J, Ishiguro K-I, Han M, Watanabe Y (2012) A conserved KASH domain protein associates with telomeres, SUN1, and dynactin, during mammalian meiosis. J Cell Biol 198(2):165–72
Moser M, Kirkpatrick A, Groves NR, Meier I (2020) LINC complex-mediated positioning of the vegetative nucleus is involved in calcium and ROS signaling in Arabidopsis pollen tubes. Nucleus 11:149–163
Moser M, Groves NR, Meier I (2024) Plant KASH proteins SINE1 and SINE2 have synergistic and antagonistic interactions with actin-branching and actin-bundling factors. J Exp Bot 75:73–87
Newman-Griffis AH, del Cerro P, Charpentier M, Meier I (2019) Medicago LINC complexes function in nuclear morphology, nuclear movement, and root nodule Symbiosis. Plant Physiol 179:491–506
Oda Y, Fukuda H (2011) Dynamics of Arabidopsis SUN proteins during mitosis and their involvement in nuclear shaping. Plant J 66:629–641
Oh SA, Allen T, Twell D (2010a) A ticket for the live show: microtubules in male gametophyte development. Plant Signal Behav 5:614–617
Oh S, Das Pal MD, Park S, Johnson A, Twell D (2010b) The tobacco MAP215/Dis1-family protein TMBP200 is required for the functional organization of microtubule arrays during male germline establishment. J Exp Bot 61:969–981
Oh S, Park K, Twell D, Park S (2010c) The SIDECAR POLLEN gene encodes a microspore-specific LOB/AS2 domain protein required for the correct timing and orientation of asymmetric cell division. Plant J 64:839–850
Oh SA, Twell D, Park SK (2011) SIDECAR POLLEN suggests a plant specific regulatory network underlying asymmetric microspore division in Arabidopsis. Plant Signal Behav 6:416–419
Pagnussat GC, Yu H-J, Ngo QA, Rajani S, Mayalagu S, Johnson CS, Capron A, Xie L-F, Ye D, Sundaresan V (2005) Genetic and molecular identification of genes required for female gametophyte development and function in Arabidopsis. Development 132:603–614
Palanivelu R, Tsukamoto T (2011) Pathfinding in angiosperm reproduction: pollen tube guidance by pistils ensures successful double fertilization. Wiley Interdiscip Rev Dev Biol 1:96–113
Park SK, Howden R, Twell D (1998) The Arabidopsis thaliana gametophytic mutant gemini pollen disrupts microspore polarity, division asymmetry and pollen cell fate. Development 125:3789–3799
Park GT, Frost JM, Park J-S, Kim TH, Lee JS, Oh SA, Twell D, Brooks JS, Fischer RL, Choi Y (2014) Nucleoporin MOS7/Nup88 is required for mitosis in gametogenesis and seed development in Arabidopsis. Proc Natl Acad Sci USA 111:18393–18398
Poulet A, Probst AV, Graumann K, Tatout C, Evans D (2017) Exploring the evolution of the proteins of the plant nuclear envelope. Nucleus 8:46–59
Pradillo M, Evans D, Graumann K (2019) The nuclear envelope in higher plant mitosis and meiosis. Nucleus 10:55–66
Qin Y, Leydon AR, Manziello A, Pandey R, Mount D, Denic S, Vasic B, Johnson MA, Palanivelu R (2009) Penetration of the stigma and style elicits a novel transcriptome in pollen tubes, pointing to genes crucial for growth in a pistil. PLoS Genet 5:e1000621
Qin Z, Wu Y, Sun T, Ma T, Xu M, Pang C, Li S, Li S (2022) Arabidopsis RAN GTPases are critical for mitosis during male and female gametogenesis. FEBS Lett 596:1892–1903
Sankaranarayanan S, Jamshed M, Delmas F, Yeung EC, Samuel MA (2020) Identification and characterization of a female gametophyte defect in sdk1-7 +/- abi3-6 +/ heterozygotes of Arabidopsis thaliana. Plant Signal Behav 15:1780038
Schmid M, Davison TS, Henz SR, Pape UJ, Demar M, Vingron M, Scholkopf B, Weigel D, Lohmann JU (2005) A gene expression map of Arabidopsis thaliana development. Nat Genet 37:501–506
Sessions A, Burke E, Presting G, Aux G, McElver J, Patton D, Dietrich B, Ho P, Bacwaden J, Ko C et al (2002) A high-throughput Arabidopsis reverse genetics system. Plant Cell 14:2985–2994
Springer PS, Holding DR, Groover A, Yordan C, Martienssen RA (2000) The essential Mcm7 protein PROLIFERA is localized to the nucleus of dividing cells during the G(1) phase and is required maternally for early Arabidopsis development. Development 127:1815–1822
Starr DA, Fridolfsson HN (2010) Interactions between nuclei and the cytoskeleton are mediated by SUN-KASH nuclear-envelope bridges. Annu Rev Cell Dev Biol 26:421–444
Takagi S, Islam MS, Iwabuchi K (2011) Chapter four-dynamic behavior of double-membrane-bounded organelles in plant cells. Int Rev Cell Mol Biol 286:181–222
Tamura K, Iwabuchi K, Fukao Y, Kondo M, Okamoto K, Ueda H, Nishimura M, Hara-Nishimura I (2013) Myosin XI-i links the nuclear membrane to the cytoskeleton to control nuclear movement and shape in Arabidopsis. Curr Biol 23:1776–1781
Terasaka O, Niitsu T (1995) The mitotic apparatus during unequal microspore division observed by a confocal laser scanning microscope. Protoplasma 189:187–193
Tofanelli R, Vijayan A, Scholz S, Schneitz K (2019) Protocol for rapid clearing and staining of fixed Arabidopsis ovules for improved imaging by confocal laser scanning microscopy. Plant Methods 15:120
Twell D (2011) Male gametogenesis and germline specification in flowering plants. Sex Plant Reprod 24:149–160
Twell D, Park SK, Lalanne E (1998) Asymmetric division and cell-fate determination in developing pollen. Trends Plant Sci 3:305–310
Twell D, Park SK, Hawkins TJ, Schubert D, Schmidt R, Smertenko A, Hussey PJ (2002) MOR1/GEM1 plays an essential role in the plant specific cytokinetic phragmoplast. Nat Cell Biol 4:711–714
Varadi M, Anyango S, Deshpande M, Nair S, Natassia C, Yordanova G, Yuan D, Stroe O, Wood G, Laydon A et al (2021) AlphaFold protein structure database: massively expanding the structural coverage of protein-sequence space with high-accuracy models. Nucleic Acids Res 50:D439–D444
Varas J, Graumann K, Osman K, Pradillo M, Evans DE, Santos JL, Armstrong SJ (2015) Absence of SUN 1 and SUN 2 proteins in Arabidopsis thaliana leads to a delay in meiotic progression and defects in synapsis and recombination. Plant J 81:329–346
Whittington AT, Vugrek O, Wei KJ, Hasenbein NG, Sugimoto K, Rashbrooke MC, Wasteneys GO (2001) MOR1 is essential for organizing cortical microtubules in plants. Nature 411:610–613
Winter D, Vinegar B, Nahal H, Ammar R, Wilson GV, Provart NJ (2007) An “Electronic Fluorescent Pictograph” browser for exploring and analyzing large-scale biological data sets. PLoS ONE 2:e718
Wise AA, Liu Z, Binns AN (2006) Three methods for the introduction of foreign DNA into agrobacterium. Methods Mol Biol 343:43–53
Wu Y, Yan J, Zhang R, Qu X, Ren S, Chen N, Huang S (2010) Arabidopsis FIMBRIN5, an actin bundling factor, is required for pollen germination and pollen tube growth. Plant Cell 22:3745–3763
Xu XM, Meulia T, Meier I (2007) Anchorage of plant RanGAP to the nuclear envelope involves novel nuclear-pore-associated proteins. Curr Biol 17:1157–1163
Zhang H, Dawe RK (2011) Mechanisms of plant spindle formation. Chromosom Res 19:335–344
Zhang M, Zhang X, Gao X (2020) ROS in the male-female interactions during pollination: function and regulation. Front Plant Sci 11:177
Zhao Q, Brkljacic J, Meier I (2008) Two distinct interacting classes of nuclear envelope-associated coiled-coil proteins are required for the tissue-specific nuclear envelope targeting of Arabidopsis RanGAP. Plant Cell 20:1639–1651
Zhou X, Meier I (2014) Efficient plant male fertility depends on vegetative nuclear movement mediated by two families of plant outer nuclear membrane proteins. Proc Natl Acad Sci USA 111:11900–11905
Zhou X, Graumann K, Evans DE, Meier I (2012) Novel plant SUN-KASH bridges are involved in RanGAP anchoring and nuclear shape determination. J Cell Biol 196:203–211
Zhou X, Graumann K, Wirthmueller L, Jones JDG, Meier I (2014) Identification of unique SUN-interacting nuclear envelope proteins with diverse functions in plants. J Cell Biol 205:677–692
Zhou X, Groves NR, Meier I (2015a) Plant nuclear shape is independently determined by the SUN-WIP-WIT2-myosin XI-i complex and CRWN1. Nucleus 6:144–153
Zhou X, Groves NR, Meier I (2015b) SUN anchors pollen WIP–WIT complexes at the vegetative nuclear envelope and is necessary for pollen tube targeting and fertility. J Exp Bot 66:7299–7307
Acknowledgements
We would like to thank all members of the Meier lab for fruitful discussions and reading of the manuscript and Dr. Emma Knee (ABRC, Columbus, Ohio) for help with tracing the ancestry of the sine3-1 and sine3-2 alleles.
Funding
This work was funded by a grant from the National Science Foundation (NSF-2023348) to IM.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare no conflicts of interest.
Additional information
Communicated by Tetsuya Higashiyama.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Moser, M., Groves, N.R. & Meier, I. The Arabidopsis KASH protein SINE3 is involved in male and female gametogenesis. Plant Reprod (2024). https://doi.org/10.1007/s00497-024-00508-8
Received:
Accepted:
Published:
DOI: https://doi.org/10.1007/s00497-024-00508-8