
Abstract
Stopover periods between flights are essential for migrating birds, and the time birds spend at stopover sites as well as the refuelling rate is determined by intrinsic and extrinsic factors, such as competition and environmental conditions. While most studies on stopover ecology have been conducted along the Nearctic-Neotropical and the Palaearctic-African flyways, little is known about species migrating along the East Asian flyway. To address this, we compared stopover duration and body mass change of 13 closely related Emberiza bunting species during autumn migration in the Russian Far East. We found significant differences in stopover duration between the species but no differences in body mass change. Overall, stopover duration decreased during the season, suggesting that late-arriving individuals leave earlier due to external factors like unfavourable climatic conditions or food availability or as a result of their endogenous spatiotemporal migration program. We also found that stopover duration correlates with the fat score at arrival and the geographic position of breeding grounds. Juveniles stayed longer at the stopover site than adults, suggesting that the latter might be able to migrate more efficiently. No sex-specific differences in stopover duration were found. We found that body mass change was positively correlated with stopover duration and arrival date. We argue that buntings modulate their stopover duration depending on the energy required to continue migration. Our results suggest the applicability of optimal migration theory for the East Asian flyway.
Zusammenfassung
Unterschiede in Rastdauer und Veränderungen der Körpermasse zwischen Emberiza Ammern während des Herbstzuges im russischen Fernen Osten
Zwischenstopps sind für Vögel auf ihren Zugwegen unverzichtbar. Die Aufenthaltsdauer und die Auffüllung der Körperreserven an diesen Rastplätzen werden sowohl von intrinsischen als auch von extrinsischen Faktoren bestimmt, wie zum Beispiel durch Konkurrenz und Umweltbedingungen. Die meisten Studien zur Rastökologie wurden entlang der nearktisch-neotropischen und paläarktisch-afrikanischen Vogelzugwege durchgeführt, wohingegen nur wenig zu den Vogelarten bekannt ist, welche sich entlang des ostasiatischen Zugweges bewegen. Hier vergleichen wir die Rastdauer und Veränderungen der Körpermasse von bis zu 13 nahe verwandter Emberiza Ammer-Arten während des Herbstzuges im russischen Fernen Osten. Wir fanden signifikante Unterschiede in der Rastdauer, aber keine Unterschiede in der Körpermassezunahme. Generell nahm die Rastdauer während der Herbstsaison ab, möglicherweise, weil spät ankommende Individuen den Rastplatz aufgrund ungünstiger Wetterbedingungen oder Nahrungsverfügbarkeit eher verlassen müssen, oder aufgrund ihres endogenen Zugprogramms. Wir konnten außerdem zeigen, dass die Rastdauer mit dem Fettscore und der geographischen Position des Brutgebietes korreliert. Jungvögel blieben zudem länger am Rastplatz als Altvögel, was ein Hinweis auf effizientere Zugstrategien Letzterer sein könnte. Wir konnten keine geschlechtsspezifischen Unterschiede im Rastverhalten finden. Die Veränderung der Körpermasse war positiv mit Rastdauer und Ankunftsdatum korreliert. Wir schließen daraus, dass die Ammern ihre Rastdauer an die Anforderungen des weiteren Zugverlaufs anpassen. Unsere Ergebnisse bestätigen die Anwendbarkeit der Theorie des optimalen Vogelzugs auch für den ostasiatischen Zugweg.
Similar content being viewed by others


Avoid common mistakes on your manuscript.
Introduction
Migration is a critical phase during the annual life cycle of birds, proven to be 15 times more lethal than stationary periods (Sillett and Holmes 2002). Migratory birds are able to fly thousands of kilometres without interruption (Wikelski et al. 2003), but such long-distance movements are energetically costly (Mcwilliams et al. 2004; Wikelski et al. 2003). Birds have to store a high quantity of fat before departure (Jenni-Eiermann and Jenni 2003; McGuire et al. 2012; Mcwilliams et al. 2004), and during migration, accumulated fatty acids are sent to active muscles, resulting in enhanced energy loss (Mcwilliams et al. 2004). Accordingly, birds optimize long-distance flights to refuel in either strategic or random areas known as “stopovers” (Alerstam 2001; Dunning 2007; Klaassen 1996).
Stopovers have been found to be two times more costly than the flight itself (Hedenström and Alerstam, 1998; McGuire et al. 2012; Schmaljohann et al. 2012; Wikelski et al. 2003). One reason for this high cost is that when birds stop, they must thermoregulate (McGuire et al. 2012; Wikelski et al. 2003). Furthermore, stopover periods are usually much longer than flights, occupying 90% of the migration time (Hedenström and Alerstam 1998; Polak and Szewczyk 2007; Schaub et al. 2001).
The time spent at the stopover site is depended on the bird’s required fuel rate, which in turn depends on their energy expenditure during the flight (Lindstrom and Alerstam 1992). Thus, fuel deposition rate and stopover duration are associated and are key determinants for successful migration (Schaub and Jenni 2001). The stopover duration is ruled by internal factors, such as the innate knowledge of the rhythm of flight and stop periods, fuel stores and fuel deposition rate, body mass and moult (Alerstam and Lindström 1990; Deppe et al. 2015; Goymann et al. 2010; Kaiser 1999; Schaub and Jenni 2001; Schmaljohann and Eikenaar 2017; Weber and Houston 1997). Yet, stopover duration also depends on external factors such as weather (wind conditions aloft or temperatures), food availability, competition and predation risk (Bozó et al. 2018; Fransson and Weber 1997; Kaiser 1999). All in all, migratory birds are expected to opt for a time-minimizing strategy, with short stopovers in productive areas to refuel fast and quickly continue their migration (Lindstrom and Alerstam 1992). Alternatively, birds can adopt an energy-minimizing strategy, with lower fuel loads and more frequent stopovers to reduce the costs of transport or the overall costs of migration (Hedenström and Alerstam 1997).
While most theories on stopover ecology were developed based on data from the Nearctic-Neotropical and the Palaearctic-African flyways (Goymann et al. 2010; Moore and Kerlinger 1987; Safriel and Lavee 1988; Van Loon et al. 2017), little is known about species migrating along the East Asian flyway (Yong et al. 2021), except for a few single-species studies (Wang et al. 2006; Bozó et al. 2020).
Here we address this knowledge gap by comparing stopover duration and body mass changes of a set of closely related Emberiza bunting species at a stopover site in the Russian Far East. Up to 14 Emberiza species use the same stopover site during migration, and they overlap both in habitat use and phenology (Heim et al. 2018). Furthermore, there are no pronounced differences in diet between the species (Byers et al. 1995). We, therefore, expect interspecific competition (Moore and Yong 1991), resulting in species-specific differences in stopover duration. We assume a longer stay for birds that arrive sooner than for individuals that reach the stopover site later in the fall (Polak and Szewczyk 2007). Further, we test whether stopover duration is correlated to species-specific distances to breeding and non-breeding grounds. We expect the northernmost breeding species as well as the southernmost wintering species to stay longer to gain more fuel (Conklin et al. 2010). In addition, we test whether birds’ age or sex affect stopover duration. While we expect no differences between females and males (Ellegren 1991), juveniles are expected to stay longer than adults (Rguibi-Idrissi et al. 2003). To examine body mass, we test whether the body mass of recaptured birds increases during a stopover. We assume a positive correlation between body mass change and stopover duration (Polak and Szewczyk 2007). We also assume a longer stopover duration for birds with a lower body mass at arrival (Arizaga et al. 2008). As such, we examine whether individuals that stay longer had arrived with lower fat reserves (Arizaga et al. 2008).
Materials and methods
Fieldwork and data selection
Fieldwork was conducted at Muraviovka Park (49°5508, 27 N, 127°4019, 93E) in the Russian Far East as part of the Amur Bird Project (Heim and Smirenski 2013). We captured birds on a daily basis from sunrise to sunset with standardized mist netting (up to 20 nets, total length 216 m) during autumn migration periods (August to November) from 2011 to 2017 (for details, see Heim et al. 2018). Each bird was ringed, measured and weighed, and we estimated fat scores (based on Eck et al. 2011). Birds caught twice in the same season were considered as “recaptures” of birds during stopover; birds not recaptured are referred to as “transients”. We calculated relative body mass by dividing body mass by wing length. Body mass change was calculated as the percentage of the body mass at “departure” (last capture) from the body mass at “arrival” (first capture) for each individual.
Statistical analysis
We estimated the minimum stopover duration as the number of days between first and last captures within a season (Yosef and Chernetsov 2005) using the lubridate package (Grolemund and Wickham 2011). This method is known to underestimate true stopover duration because the true arrival and departure dates are unknown (Schaub and Jenni 2001). However, it permits a comparable individual estimation of the time spent at the stopover (Yosef and Chernetsov 2005). Additionally, we have also calculated the species-specific minimum stopover duration including all transients. We defined transients to have a stopover duration of 0 days, as they might continue migration on the same day.
We examined which factors predict stopover duration using generalized mixed-effects models (GLMM) with a Poisson error distribution in the lme4 (Bates et al. 2014) and MuMIn packages (Barton and Barton 2015). We fitted stopover duration as a dependent variable and built 15 different models (for details see Supplement 1) adding the explanatory variables one by one: species, day of arrival, relative body mass, fat score at arrival, age and sex. Due to limited sample size, sex and age were not fitted into the same model. We also checked which of the random factors (year and the interactions between species and age, sex, day of arrival, relative body mass and fat score at arrival) explained a significant proportion of the variance (> 0.5). We used the Akaike Information Criterion (AIC) to select the best model (models with ΔAIC < 2 were considered equally well) and the significant explanatory variables.
We ran linear mixed-effect models (LMERs) (Bates et al. 2014) with a Poisson error distribution including migration distance, northernmost breeding latitude and southernmost non-breeding latitude as explanatory variables to check influences on stopover duration (response variable) with species and year as random factor applying backward stepwise model selection in the MASS package (Ripley et al. 2013) to obtain the best model out of the initial LMERs (Supplement 1). Data on species-specific migration distances as well as breeding and non-breeding latitudes were retrieved from a previous study at this site (Heim et al. 2018).
We built linear mixed-effect models (LMERs) with a Poisson error distribution to see if body mass change was linked to species (all species combined), stopover duration and day of arrival as explanatory variables for all species with n > 15 recaptures. We obtained the best model using again the backward elimination method (Supplement 1). Furthermore, we added the time difference between the first and last capture as co-variate (trecapture − tfirst capture, Schaub and Jenni 2000). We also added year as a random factor.
All analyses were conducted using the program R version 4.0.2 (R Core Team 2019).
Results
We captured 7502 buntings of 13 species, including 453 recaptures (Table 1). A minimum number of > 15 recaptures were available for six species (Black-faced Bunting Emberiza spodocephala, Elegant Bunting E. elegans, Little Bunting E. pusilla, Pallas’s Reed Bunting E. pallasi, Rustic Bunting E. rustica and Yellow-browed Bunting E. chrysophris).
Stopover duration
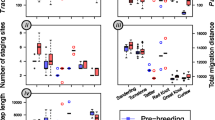
We found significant differences in stopover duration and the percentage of transients among the 13 studied species (χ2 = 288.89, df = 12, p < 0.001, Table 1). The majority (92.2%) of all ringed buntings were transients. Mean stopover duration (including transients) ranged from 0 days for the Chestnut-eared Bunting to 3.7 days for the Elegant Bunting (Table 1). The strongest differences were found between the Black-faced (1.0 days) and Elegant Buntings (3.7 days) and the remaining species (~ 0 days). When considering only recaptured individuals (representing 7.8% of all captured birds), the mean stopover duration ranged from 4.7 days (Pallas’s Reed Bunting) to 13.4 days (Elegant Bunting). Mean stopover duration (without transients) differed significantly between those six species (χ2 = 25.201, df = 5, p < 0.001, Fig. 1).

Minimum stopover duration (in days) of six bunting species at a stopover site in the Russian Far East. Only individuals which were recaptured at least once were included. Photos by Arend Heim & Wieland Heim (Amur Bird Project)
Drivers of stopover duration
We found that the model containing species, day of arrival, age, relative body mass and fat score at arrival was the best model to explain stopover duration (AIC = 2761.1, Table 2, Supplement 2). We found significant differences in stopover duration between species (Fig. 1). Furthermore, we found strong evidence that earlier-arriving individuals stay longer than later-arriving birds (Fig. 2). We also found evidence for a significant difference in the stopover duration of adults and juveniles (Fig. 3), with a mean stopover duration in adult buntings of 6.8 days and 8.9 days in juveniles. We found no evidence for differences in stopover duration between females and males), with a mean stopover duration of 5.8 days in females and 6.2 days in males. There was no evidence that relative body mass was linked to stopover duration. Individuals arriving at the stopover site with a lower fat score stayed significantly longer than birds with higher fat scores at arrival. The interaction of species and day of arrival as well as species and relative body mass explained a significant proportion of the variance, whereas other random factors did not.

Minimum stopover duration by day of arrival (= day of the first capture) of six bunting species with at least 15 recaptured individuals. Each dot represents one individual, lines are regression lines and shaded areas depict confidence intervals. Days are given as julian days (210 = 29 July, 310 = 6 November)

Stopover duration (in days) at the stopover site for both adult (n = 100) and juvenile (n = 353) buntings based on data of six bunting species with at least 15 recaptured individuals. Note that only two adults were recaptured for Little and Yellow-browed Bunting
Migration distance, breeding and non-breeding latitudes effects
We found that the northernmost breeding latitude was negatively correlated with stopover duration (AIC = 3952.4, Fig. 4, Supplement 2). The best model also contained year as a random factor, suggesting significant variation between years (Table 2). We found no evidence for a link between southernmost wintering latitude or migration distance with stopover duration.

Bunting species with a higher northernmost breeding latitude were found to have a shorter stopover duration. Shown are six species with at least 15 recaptured individuals
Body mass change
The final model showed that body mass change was linked to stopover duration, day of arrival and species (AIC = 3313.9, Fig. 5, Table 2, Supplement 2). Those individuals that stayed longer and arrived earlier showed stronger positive body mass changes. However, this pattern was significant in only four out of six species (Fig. 5). We also found that the time difference between the first and last capture affected the body mass change (Supplement 1).

Effect of stopover duration on the body mass change between the first and the last capture of individual buntings. Each dot represents one individual. Regression lines and confidence intervals (shaded areas) are given for species with a significant correlation
Discussion
We found significant differences in the stopover duration of 13 bunting species in the Russian Far East (Table 1). However, most birds were trapped only once, and were therefore considered transients. When comparing the six species most commonly recaptured, the mean stopover duration was close to or greater than two times higher for Black-faced, Elegant, Little and Yellow-browed buntings than for Pallas’s Reed and Rustic Buntings (Fig. 1). These differences were consistent between years, confirming rather fixed migration schedules of buntings at this stopover site (Heim et al. 2018).
Drivers of stopover duration
Differences in stopover duration have been linked to feeding habits: Species using a scarce or unpredictable food resource might require longer stopovers than species with a more flexible diet (Bairlein 1998; Bibby and Green 1981). The similar stopover durations of different warbler species in the Russian Far East have also been explained with similarities in diet (Bozó et al. 2020). However, the studied buntings at our study site have a similar bill morphology and are expected to have a similar diet outside the breeding season (Heim et al. 2018), but they still differ greatly in stopover duration. This might be explained by interspecific competition between the species, as overlap in feeding niches enhances interspecific competition (Moore and Yong 1991). Such competition can impede a time-minimizing strategy for the least competitive species and could explain the longer stays for four of the six bunting species most commonly recaptured.
Further, the differences in stopover duration might also be explained with species-specific arrival dates. Individuals arriving in August stayed around two weeks (mean: 18.8 days), while those arriving in September and October left the site after 4 or 5 days (Table 1). The first Elegant Buntings, the species with the longest mean stopover duration, tend to arrive at our study site already in August, whereas Rustic Buntings, the species with the shortest stopover duration, do not arrive before September (Fig. 2). To prevent competition, there might be a strict partitioning at the temporal scale between closely related species with similar foraging habits during migration and at stopover sites (Byers et al. 2013; Schoener 1974); staggered departures from the breeding grounds might reduce competition over resources. Such differences in timing have been reported previously for migrating buntings at our stopover site (Heim et al. 2018; Smirenski and Smirenski 2010). On the other hand, the decreasing stopover duration during the autumn season might simply reflect the availability of food (Eikenaar and Bairlein 2014). Seeds, as preferred by the studied buntings, might be more readily available later in autumn and might allow for faster refuelling and, therefore, shorter stopover durations.
First-year birds stayed at the stopover site significantly longer than adults (Fig. 3). This result is consistent with other studies on songbirds (Ellegren 1991; Newton 2010; Polak and Szewczyk 2007; Rguibi-Idrissi et al. 2003). Adults are more experienced and, therefore, are able to find suitable stopovers and accumulate fat faster than juveniles new to the migration challenge (Polak and Szewczyk 2007).
We found no significant effect of sex on stopover duration (Fig. 3). Earlier studies suggested that males might be more dominant, which could allow them to refuel faster, while females would stay longer to avoid competition (Newton 2010). In the absence of significant differences, intersexual competition might be low, and both sexes of the studied buntings might be able to follow a time-minimizing strategy.
Contrary to our expectations and most studies (e.g., Arizaga et al. 2008; Cherry 1982), we found that relative body mass at arrival did not predict stopover duration. However, fat score at arrival was included in the best model (Table 2)—buntings arriving with lower fat scores tend to stay longer at the study site than birds arriving with higher fat scores. Similar patterns were found in other songbirds (Cherry 1982; Moore and Kerlinger 1987). Fat accumulation is thought to be responsible for most of the body mass increase in birds during a stopover (Connell et al. 1960). As mentioned above, species breeding in high latitudes might refuel prior to arrival at our study site, which could explain why, e.g., Pallas’s Reed and Rustic Buntings arrive later at our study site and with higher fat scores, leading to a shorter stopover duration.
Migration distance, breeding and non-breeding latitudes effects
Another explanation for the differences in stopover duration might stem from differences in migration distance and the geographic origin of the species (Monti et al. 2018). During autumn migration, buntings breeding further south and/or migrating longer distances are known to arrive earlier at our study site than species breeding further north and/or migrating shorter distances (Heim et al. 2018). This could affect the stopover duration as well since longer-distance migrants might have to refuel more extensively (Lindstrom and Alerstam 1992). However, we found no evidence for a significant correlation between migration distance and stopover duration. This can be explained by the early occurrence of a number of individuals in species that migrate otherwise relatively late in the season, such as the Elegant Bunting. The migration distance of this species is relatively short, but those individuals that arrive early will stay for extended periods at our study site. While we found no effect of migration distance, we did observe that stopover duration decreased with the northernmost latitude of breeding grounds (Fig. 4). This pattern has been observed in other songbirds (Smetzer and King 2018), but it may reverse if northern populations have to cross significant ecological barriers (Conklin et al. 2010; Van Loon et al. 2017). Buntings breeding in higher latitudes might have access to more favourable Siberian stopovers and might refuel before reaching our study site. Alternatively, they might stopover more frequently and for shorter amounts of time than species breeding further south. The higher proportion of transients in species breeding further north, such as Little, Pallas’s Reed and Rustic Buntings (Table 1), supports this assumption. Given that “year” was included as a random factor in the best model (Table 2), interannual differences depending e.g. on local weather or food availability might be expected in this pattern.
Body mass change
One of the ultimate drivers of stopover duration is the refuelling rate, which can be affected by competition, geographic origin, or age, as discussed above. We, therefore, investigated changes in body mass of the recaptured buntings and found a mean increase of 0.14 g, similar to other studies on songbirds (e.g. Arizaga et al. 2008). However, only one of six species showed a significant increase in body mass, the Little Bunting; overall, differences between species were not significant. This suggests low fuel deposition rates at our study site, which could be the result of competition (Moore and Yong 1991). Nevertheless, we found a positive correlation of body mass change with stopover duration (Fig. 5), confirming a higher mass gain in birds staying longer (Arizaga et al. 2008; Safriel and Lavee 1988). Furthermore, we found that birds arriving earlier would also show higher mass gains. This is most likely linked to the fact that earlier arriving birds are more likely to stay longer (see Fig. 2). As the random factor “year” explained a significant proportion of the variance (Table 2), interannual differences in refuelling rate can be expected. This might be linked to differences in stopover habitat quality, which could be linked to drastic differences in vegetation at our study site between years with and without fires (Heim et al. 2019). Unsurprisingly, we also found that the time difference between first and last capture events was explaining body mass change, which can be explained with the body mass gain during the course of the day (Delingat et al. 2009).
Conclusions
We found that the autumn stopover durations of bunting species differ at our study site in the Russian Far East and are determined by geographic origin, arrival date, age, and fat score at arrival, whereas body mass change is linked to stopover duration and arrival date. The observed patterns are similar to observations made on songbirds migrating along other flyways, suggesting the applicability of the optimal migration theory for the East Asian flyway. Our study site provides a unique opportunity to compare the migratory behaviour of a large number of closely related species occurring sympatrically during a stopover.
The populations of many of the studied bunting species have declined in the past decades (Choi et al. 2020; Edenius et al. 2017, Kamp et al. 2015), and some of the species are now globally threatened. A better understanding of their migration ecology and protection throughout the annual cycle is urgently required.
References
Alerstam T (2001) Detours in bird migration. J Theor Biol 209:319–331. https://doi.org/10.1006/jtbi.2001.2266
Alerstam T, Lindström Å (1990) Optimal bird migration: the relative importance of time, energy, and safety. In: Gwinner E (ed) Bird migration. Springer, Berlin, Heidelberg, pp 331–351
Arizaga J, Barba E, Belda EJ (2008) Fuel management and stopover duration of Blackcaps Sylvia atricapilla stopping over in northern Spain during autumn migration period. Bird Study 55:124–134
Bairlein F (1998) The effect of diet composition on migratory fuelling in Garden Warblers Sylvia borin. J Avian Biol 29:546–551
Barton K, Barton MK (2015) Package ‘MuMIn.’ Version 1, 18
Bates D, Mächler M, Bolker B, Walker S (2014) Fitting linear mixed-effects models using lme4. ArXiv e-prints arXiv:1406. https://doi.org/10.18637/jss.v067.i01
Bibby CJ, Green RE (1981) Autumn migration strategies of reed and sedge warblers. Ornis Scand 12:1–12. https://doi.org/10.2307/3675898
Bozó L, Csörgő T, Heim W (2018) Weather conditions affect spring and autumn migration of Siberian leaf warblers. Avian Res 9:33. https://doi.org/10.1186/s40657-018-0126-5
Bozó L, Csörgő T, Heim W (2020) Stopover duration and body mass change of two Siberian songbird species at a refuelling site in the Russian Far East. Ornithol Sci 19:159–166
Byers C, Olsson U, Curson J (1995) Buntings and sparrows: a guide to the buntings and North American sparrows. A&C Black, London
Byers C, Olsson U, Curson J (2013) Buntings and sparrows. Bloomsbury Publishing Plc, UK
Cherry JD (1982) Fat deposition and length of stopover of migrant white-crowned sparrows. Auk 99:725–732
Choi CY, Nam HY, Kim HK, Park SY, Park JG (2020) Changes in Emberiza bunting communities and populations spanning 100 years in Korea. PloS one 15(5):e0233121
Conklin JR, Battley PF, Potter MA, Fox JW (2010) Breeding latitude drives individual schedules in a trans-hemispheric migrant bird. Nat Commun 1:1–6. https://doi.org/10.1038/ncomms1072
Connell CE, Odum EP, Kale H (1960) Fat-free weights of birds. Auk 77:1–9
Delingat J, Dierschke V, Schmaljohann H, Bairlein F (2009) Diurnal patterns of body mass change during stopover in a migrating songbird, the northern wheatear Oenanthe oenanthe. J Avian Biol 40:625–634
Deppe JL, Ward MP, Bolus RT, Diehl RH, Celis-Murillo A, Zenzal TJ, Moore FR, Benson TJ, Smolinsky JA, Schofield LN, Enstrom DA, Paxton EH, Bohrer G, Beveroth TA, Raim A, Obringer RL, Delaney D, Cochran WW (2015) Fat, weather, and date affect migratory songbirds’ departure decisions, routes, and time it takes to cross the Gulf of Mexico. Proc Natl Acad Sci 112:E6331–E6338. https://doi.org/10.1073/pnas.1503381112
Dunning JB Jr (2007) CRC handbook of avian body masses. CRC Press, Boca Raton
Eck S, Töpfer T, Fiebig J, Heynen I, Fiedler W, Nicolai B, van den Elzen R, Winkler R, Woog F (2011) Measuring birds. Deutsche Ornithologen-Gesellschaft
Edenius L, Choi CY, Heim W, Jaakkonen T, De Jong A, Ozaki K, Roberge JM (2017) The next common and widespread bunting to go? Global population decline in the Rustic Bunting Emberiza rustica. Bird Conserv Int 27:35–44. https://doi.org/10.1017/s0959270916000046
Eikenaar C, Bairlein F (2014) Food availability and fuel loss predict Zugunruhe. J Ornithol 155:65–70
Ellegren H (1991) Stopover ecology of autumn migrating Bluethroats Luscinia s. svecica in relation to age and sex. Ornis Scand 22:340–348
Francis CM, Cooke F (1986) Differential timing of spring migration in wood warblers (Parulinae). Auk 103:548–556
Fransson T, Weber TP (1997) Migratory fuelling in blackcaps (Sylvia atricapilla) under perceived risk of predation. Behav Ecol Sociobiol 41:75–80
Goymann W, Spina F, Ferri A, Fusani L (2010) Body fat influences departure from stopover sites in migratory birds: evidence from whole-island telemetry. Biol Lett 6(478):481. https://doi.org/10.1098/rsbl.2009.1028
Grolemund G, Wickham H (2011) Dates and times made easy with lubridate. J Stat Softw 40:1–25
Harrison XA, Donaldson L, Correa-Cano ME, Evans J, Fisher DN, Goodwin CE, Robinson BS, Hodgson DJ, Inger R (2018) A brief introduction to mixed effects modelling and multi-model inference in ecology. PeerJ 6:e4794
Hedenström A, Alerstam T (1997) Optimum fuel loads in migratory birds: distinguishing between time and energy minimization. J Theor Biol 189:227–234
Hedenström A, Alerstam T (1998) How fast can birds migrate? J Avian Biol 29:424–432. https://doi.org/10.2307/3677161
Heim W, Smirenski S (2013) The Amur bird project at Muraviovka Park in far East Russia. Bird Asia 19:31–33
Heim W, Eccard JA, Bairlein F (2018) Migration phenology determines niche use of East Asian buntings (Emberizidae) during stopover. Curr Zool 64:681–692
Heim RJ, Hölzel N, Heinken T, Kamp J, Thomas A, Darman GF, Smirenski SM, Heim W (2019) Post-burn and long-term fire effects on plants and birds in floodplain wetlands of the Russian Far East. Biodivers Conserv 28:1611–1628
Iijima K (1973) A comparative study on the wintering ecology of four species of Emberiza in the basin of Chikuma River. J Yamashina Inst Ornithol 7:179–201
Jenni-Eiermann S, Jenni L (2003) Interdependence of flight and stopover in migrating birds: possible effects of metabolic constraints during refuelling on flight metabolism. In: Berthold P, Gwinner E, Sonnenschein E (eds) Avian migration. Springer, Berlin, pp 293–306
Kaiser A (1999) Stopover strategies in birds: a review of methods for estimating stopover length. Bird Study 46:S299–S308
Kamp J, Oppel S, Ananin AA, Durnev YA, Gashev SN, Hölzel N, Mishchenko AL, Pessa J, Smirenski SM, Strelnikov EG, Timonen S, Wolanska K, Chan S (2015) Global population collapse in a superabundant migratory bird and illegal trapping in China. Conserv Biol 29(6):1684–1694
Ketterson ED, Nolan V Jr (1976) Geographic variation and its climatic correlates in the sex ratio of eastern-wintering dark-eyed juncos (Junco hyemalis hyemalis). Ecology 57:679–693
Klaassen M (1996) Metabolic constraints on long-distance migration in birds. J Exp Biol 199:57–64
Lindstrom A, Alerstam T (1992) Optimal fat loads in migrating birds: a test of the time-minimization hypothesis. Am Nat 140:477–491. https://doi.org/10.1086/285422
McGuire LP, Guglielmo CG, Mackenzie SA, Taylor PD (2012) Migratory stopover in the long-distance migrant silver-haired bat, Lasionycteris noctivagans. J Anim Ecol 81:377–385
Mcwilliams SR, Guglielmo C, Pierce B, Klaassen M (2004) Flying, fasting, and feeding in birds during migration: a nutritional and physiological ecology perspective. J Avian Biol 35:377–393. https://doi.org/10.1111/j.0908-8857.2004.03378.x
Menz M, Arlettaz R (2012) The precipitous decline of the ortolan bunting Emberiza hortulana: time to build on scientific evidence to inform conservation management. Oryx 46:122–129
Monti F, Grémillet D, Sforzi A, Dominici JM, Bagur RT, Navarro AM, Fusani L, Klaassen RHG, Alerstam T, Duriez O (2018) Migration distance affects stopover use but not travel speed: contrasting patterns between long- and short-distance migrating ospreys. J Avian Biol 49:e01839. https://doi.org/10.1111/jav.01839
Moore F, Kerlinger P (1987) Stopover and fat deposition by North American wood-warblers (Parulinae) following spring migration over the Gulf of Mexico. Oecologia 74:47–54. https://doi.org/10.1007/BF00377344
Moore FR, Yong W (1991) Evidence of food-based competition among passerine migrants during stopover. Behav Ecol Sociobiol 28:85–90
Newton I (2010) The migration ecology of birds. Elsevier Science, Amsterdam
Pacala S, Roughgarden J (1982) Resource partitioning and interspecific competition in two two-species insular Anolis lizard communities. Science 217:444–446
Polak M, Szewczyk P (2007) Relation between stopover length and time and body parameters of European Robin Erithacus rubecula (L., 1758) during autumn migration (Central Poland). Pol J Ecol 55:511–517
R Core Team (2019) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Rappole JH (2013) The avian migrant: the biology of bird migration. Columbia University Press, Columbia
Rguibi-Idrissi H, Julliard R, Bairlein F (2003) Variation in the stopover duration of Reed Warblers Acrocephalus scirpaceus in Morocco: effects of season, age and site. Ibis 145:650–656. https://doi.org/10.1046/j.1474-919X.2003.00208.x
Ricklefs RE, Cox GW (1977) Morphological similarity and ecological overlap among passerine birds on St. Kitts, British West Indies. Oikos 29:60–66. https://doi.org/10.2307/3543293
Ripley B, Venables B, Bates DM, Hornik K, Gebhardt A, Firth D, Ripley MB (2013) Package ‘mass.’ Cran R 538:113–120
Safriel U, Lavee D (1988) Weight changes of cross-desert migrants at an oasis—do energetic considerations alone determine the length of stopover? Oecologia 76:611–619. https://doi.org/10.1007/BF00397878
Sander MM, Heim W, Schmaljohann H (2020) Seasonal and diurnal increases in energy stores in migratory warblers at an autumn stopover site along the Asian-Australasian flyway. J Ornithol 161:73–87. https://doi.org/10.1007/s10336-019-01701-7
Schaub M, Jenni L (2000) Fuel deposition of three passerine bird species along the migration route. Oecologia 122:306–317
Schaub M, Jenni L (2001) Stopover durations of three warbler species along their autumn migration route. Oecologia 128:217–227. https://doi.org/10.1007/s004420100654
Schaub M, Pradel R, Jenni L, Lebreton JD (2001) Migrating birds stop over longer than usually thought: an improved capture–recapture analysis. Ecology 82:852–859
Schmaljohann H, Eikenaar C (2017) How do energy stores and changes in these affect departure decisions by migratory birds? A critical view on stopover ecology studies and some future perspectives. J Comp Physiol A 203:411–429. https://doi.org/10.1007/s00359-017-1166-8
Schmaljohann H, Fox JW, Bairlein F (2012) Phenotypic response to environmental cues, orientation and migration costs in songbirds flying halfway around the world. Anim Behav 84:623–640
Schoener TW (1974) Resource partitioning in ecological communities. Science 185:27–39
Sillett TS, Holmes RT (2002) Variation in survivorship of a migratory songbird throughout its annual cycle. J Anim Ecol 71:296–308
Smetzer JR, King DI (2018) Prolonged stopover and consequences of migratory strategy on local-scale movements within a regional songbird staging area. Auk 135:547–560
Smirenski SM, Smirenski EM (2010) Migration timing of buntings in the south of the Middle Amur region. Russ Ornithol J 19:1452–1454 (in Russian)
Tamada K, Tomizawa M, Umeki M, Takada M (2014) Population trends of grassland birds in Hokkaido, focussing on the drastic decline of the Yellow-breasted Bunting. Ornithol Sci 13:29–40. https://doi.org/10.2326/osj.13.29
Van Loon A, Ray J, Savage A, Mejeur J, Moscar L, Pearson M, Pearman M, Hvenegaard G, Mickle N, Applegate K (2017) Migratory stopover timing is predicted by breeding latitude, not habitat quality, in a long-distance migratory songbird. J Ornithol 158:745–752
Wang Y, Chang J, Moore FR, Su L, Cui L, Yang X (2006) Stopover ecology of red-flanked bush robin (Tarsiger cyanurus) at Maoershan, Northeast China. Acta Ecol Sin 26:638–646
Weber TP, Houston AI (1997) Flight costs, flight range and the stopover ecology of migrating birds. J Anim Ecol 66:297–306
Wickham H (2016) ggplot2: Elegant Graphics for Data Analysis, Use R! Springer International Publishing, Switzerland
Wikelski M, Tarlow EM, Raim A, Diehl RH, Larkin RP, Visser GH (2003) Costs of migration in free-flying songbirds. Nature 423:704–704
Yong DL, Heim W, Chowdhury SU, Choi CY, Ktitorov P, Kulikova O, Kondratyev A, Round PD, Allen D, Trainor CR, Gibson L, Szabo JK (2021) The state of migratory landbirds in the East Asian Flyway: Distributions, threats, and conservation needs. Front Ecol Evol 9:613172. https://doi.org/10.3389/fevo.2021.613172
Yosef R, Chernetsov N (2005) Longer is fatter: body mass changes of migrant Reed Warblers (Acrocephalus scirpaceus) staging at Eilat, Israel. Ostrich 76:142–147
Acknowledgements
The authors want to thank all volunteers of the Amur Bird Project who supported the data collection, as well as Sergei M. Smirenski and the staff of Muraviovka Park. The fieldwork for this study was supported by grants of the German Ornithologist’s Society (Deutsche Ornithologen-Gesellschaft e.V.,), Förderkreis Allgemeine Naturkunde (Biologie) e.V., NABU Regionalverband Erzgebirge e.V., The Sound Approach, Mohamed bin Zayed Species Conservation Fund, Oriental Bird Club, ProRing e.V as well as private donors.
Funding
Open Access funding provided by University of Turku (UTU) including Turku University Central Hospital.
Author information
Authors and Affiliations
Corresponding author
Additional information
Communicated by N. Chernetsov.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Collet, L., Heim, W. Differences in stopover duration and body mass change among Emberiza buntings during autumn migration in the Russian Far East. J Ornithol 163, 779–789 (2022). https://doi.org/10.1007/s10336-022-01976-3
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-022-01976-3