
Abstract
Agricultural intensification has significantly impacted habitat structures in agricultural landscapes and is one of the main drivers of biodiversity decline, especially in farmland birds. Birds are considered to reflect well the trends in other biodiversity elements and are therefore often used as indicator species. We studied common pheasant (Phasianus colchicus) brood habitat use in a small-grain-dominated farmland in southern Finland. The broods significantly preferred field margins compared to their availability. The importance of field margins was underlined, as 68% of pheasant brood observations in grain fields were within a 25-m-wide zone from the field edge, despite the availability being only 40% of the field area. Our results support the idea that field margins and their proximity act as possible biodiversity reservoirs even in intensive farming systems. Increasing the amount of field margins can be an effective management method when aiming to improve success of common pheasant broods while simultaneously benefitting farmland biodiversity. Identifying key habitats and landscape features that allow the co-existence of biodiversity and effective food production is crucial when aiming to halt the ongoing biodiversity collapse.
Zusammenfassung
Auf die Ränder kommt es an: Die Bedeutung von Ackerrändern als Brutraum für Vögel in einer landwirtschaftlich intensiv genutzten Landschaft.
Die Intensivierung der Landwirtschaft hat sich erheblich auf die Strukturierung der Lebensräume von Agrarlandschaften ausgewirkt und ist eine der Hauptursachen für den Rückgang der biologischen Vielfalt, insbesondere bei den Vögeln der Agrarlandschaft. Man geht davon aus, dass Vögel die Entwicklungen anderer Aspekte der biologischen Vielfalt gut widerspiegeln und deshalb oft als Indikatorarten betrachtet werden können. Wir untersuchten die Nutzung des Bruthabitats des Jagdfasans (Phasianus colchicus) in einem von Getreide dominierten Ackerland in Südfinnland. Die Elterntiere zogen Feldraine in Hinblick auf deren Verfügbarkeit deutlich vor. Die Bedeutung der Feldränder wurde dadurch betont, dass 68% der Fasane in Getreidefeldern innerhalb einer 25 m breiten Zone vom Feldrand aus beobachtet wurden, obwohl deren Verfügbarkeit nur 40% der Feldfläche ausmachte. Unsere Ergebnisse unterstützen die Annahme, dass selbst in landwirtschaftlich intensiv genutzten Strukturen Feldränder und deren Nahbereiche als mögliche Reservoirs für die biologische Vielfalt dienen. Die Ausweitung von Feldrainen kann eine wirksame Art der Bewirtschaftung sein, um den Bruterfolg von Fasanen zu erhöhen und gleichzeitig die Artenvielfalt in der Agrarlandschaft zu fördern. Die Erfassung der wichtigsten Lebensräume und derjenigen Landschaftsmerkmale, die gleichzeitig die biologische Vielfalt und auch eine effektive Nahrungsmittelproduktion ermöglichen, ist von entscheidender Bedeutung, wenn der andauernde Rückgang der biologischen Vielfalt aufgehalten werden soll.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Agricultural intensification has significantly impacted the habitat structure of agricultural landscapes worldwide and is one of the main drivers of biodiversity decline (Altieri 1999; Chamberlain et al. 2000; Benton et al. 2003; Evans 2004; Ponce et al. 2014). Intensification has caused a loss of heterogeneity, an increase in field patch size and in the use of pesticides and fertilizers, a shift in the timing of agricultural activities, and other changes in land use that have impacted trophic cascades within communities inhabiting the agricultural landscape (Stoate et al. 2001, 2009; Hietala-Koivu 2002). This has set many European farmland bird populations in severe, and ongoing, declines (BirdLife International 2015; IUCN 2021). While the seriousness of this threat to biodiversity is evident (Donald et al. 2006; Butler et al. 2010), there is an ever-increasing demand for enhancing the efficiency of agricultural enterprises (Tilman et al. 2011; Gabriel et al. 2013; Searchinger et al. 2018; Clough et al. 2020). While the most productive areas are experiencing land use intensification, less productive areas concurrently face the abandonment of agricultural practices, which can be equally detrimental to agricultural biodiversity and farmland birds (Kleijn et al. 2011; Uchida and Ushimaru 2014; González del Portillo et al. 2021; Silva-Monteiro et al. 2021). Combining actions to halt ongoing biodiversity loss and managing a sustainable and productive agriculture that provides food security and economic viability is a complex task. However, at the current rate of urbanization, fragmentation and climate change, biodiversity conservation needs to extend beyond nature conservation areas in order to be successful. This requires agricultural landscapes capable of supporting wildlife (Bennett et al. 2006; Frei et al. 2018). Identifying key habitats and landscape features that support biodiversity and allow its co-existence with effective farming is one solution toward more sustainable agriculture (Fischer et al. 2017, Sirami et al. 2019, Clough et al. 2020, Šalek 2021).
Birds are considered to reflect well the trends in other biodiversity elements (Gregory et al. 2005). The EU farmland bird indicator uses multiple species to assess the biodiversity status of agricultural landscapes in Europe (EFBI). The Gray partridge (Perdix perdix) is one such species due to its tight habitat demands within the agricultural landscape (Potts 1986; European Council 2001; Gregory et al. 2005). During the last 50 years, gray partridges (hereafter partridge) have suffered a population loss of over 90% in Europe (EBCC European Bird Census Council, 2017). Evidence shows that the large-scale collapse of native partridge populations across Europe has been followed by a decrease in non-native common pheasant (Phasianus colchicus) populations. The large-scale introductions of common pheasants (hereafter pheasant) into the agricultural landscape, mainly for hunting purposes, have somewhat masked this population decline. Established wild pheasant populations have nonetheless declined over recent decades (Powell 2015; Robertson et al. 2017). Pheasant declines in the USA have been linked to changes in agriculture practices (Coates et al. 2017).
The pheasant and partridge are the main non-migratory gallinaceous bird species inhabiting the agricultural landscape of Northern Europe. These species require agricultural areas that offer food and shelter from predators, such as vegetation cover, and an adequate distance to forests that inhabit both avian and mammalian predators, nesting habitat in the form of perennial grasses, and insect-rich habitats such as segetal flora for brood rearing (Hill 1985; Potts 1986; Jorgensen et al. 2014; Ronnenberg et al. 2016). Several studies in the USA have shown that pheasants benefit from heterogeneous vegetation that provides varying successional stages (Taylor et al. 2018).
Reasons behind the partridge decline have been identified with extensive studies across Europe. The indirect effects of herbicides and direct effects of insecticides have led to a loss of arthropods on agricultural land, causing a lack of food for the chicks (Potts 1986; Potts and Aebischer 1995). Nesting habitats have been lost with intensified land use (Aebischer and Ewald 2004), and predation has increased, leading to higher mortality, particularly of nesting females (Panek and Bresinski 2002; Newton 2004; Langgemach and Bellebaum 2005; Ewald et al. 2010; Smith et al. 2010; Roos et al. 2018).
The factors affecting pheasant breeding success are in many ways similar to those influencing the partridge. Both species require a high-quality arthropod diet for the first 3 weeks of their lives, along with guidance and protection provided by an adult female (Hill 1985; Potts 1986; Smith et al. 2015). The home range size of a pheasant brood increases in monoculture habitats, thus leading to increased movement of the chicks, which subsequently increases their mortality (Warner 1979; Hill 1985). This implies that a pheasant hen is adept at judging the insect richness and quality of habitats, and that chick survival is a good indicator of habitat quality when considering the other factors that work against brood survival, e.g., predation and poor weather conditions (Brittas et al. 1992; Riley et al. 1998; Musil and Connelly 2009). Thereby, in the absence of natural partridge populations, pheasant broods could also be used as an indicator for farmland insect biodiversity and habitat quality. Recognizing the most valuable farmland areas makes it easier to target habitat management measures and to emphasize these landscape features while upholding a high food production capacity of the farming enterprise. Nielson et al. (2008) considered the pheasant a good indicator of agricultural landscapes and of the successional habitat created by a conservation reserve program (CRP) in the USA.
Our aim is to obtain information that will help in planning future drivers that enhance the biodiversity value of farmland, irrespective of the intensity of the implemented agricultural practices. To achieve this goal, we use the habitat selection process of pheasant broods as an indicator of habitat quality. Habitat selection is also examined, to elucidate the scale at which biodiversity drivers must be implemented in potential future management actions. We predict that 1) the broods use agricultural fields with vegetation cover and 2) the hens show some preference when choosing feeding areas for their broods.
Materials and methods
Study area
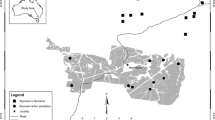
The study was conducted during 1995–1998 in Siuntio, southern Finland (N 60º 11,176′ E 24º 11,843′, ETRS89). This part of Finland is characterized by boreal forest, mainly conifers, mixed with some deciduous trees, interspersed with open agricultural landscapes (see Pic 1 in Electronic Supplementary Material). All fields in the area are subsurface drained and separated from one another by open ditches that also provide field margin habitat with herbaceous vegetation and occasional willow stands (Salix spp.). The median field patch size was 1.9 ha (mean 5.8, range 0.15–41.0).
At the time of the study, the agricultural practices in our area were mainly regular farming methods used in southern Finland (see also Vepsäläinen 2007): grain field spraying, one treatment with an herbicide (e.g., MCPA) spread on winter cereals in May and on spring cereals in June, and one treatment with a growth regulator (e.g., CCC) in June, also containing a fungicide if needed. Winter wheat, rye, and barley were the most sprayed grains. Because the study farm had dairy cattle, slurry was spread on the fields that were plowed in the fall. During summer (June–August), cattle grazed on the pastures.
We delineated the study area as a circle, to study the habitat use of brood-rearing pheasant hens that had previously been released from pens to the study area. The central point of the circle was fixed halfway between the two furthest pens that the pheasants were released from. All three release pens were in a row close together, within 250 m of each other. The pens were placed beside a tall spruce row, within a park, providing the birds with protection against the weather. The circle radius was 1800 m (see Pic 1 in Electronic Supplementary Material), which was based on the furthest tracking fix of a radio-tagged hen during the study period from the central point. This gave us an area of 1020 ha.
The study area was first divided into five land use categories: (1) cultivated fields, fallows, meadows, gardens and parks, (2) forests, (3) rivers and creeks, (4) roads, (5) yard areas, houses, and other non-natural areas. All areas were counted using QGIS 2.18 Development Team (2019). Map data were obtained from the National Land Survey of Finland (2019) and digitized manually to correspond to the land use at the time of the study. Categories (1) (397 ha) and (3) (26.9 ha) and a 1-m broad field margin area around each field patch (67 km equaling 6.7 ha) were included as suitable areas for pheasants, altogether 430.6 ha. All habitat analyses were performed per year considering the yearly changes in cultivated areas.
A field margin is defined as the area left between the main crop and the field boundary. In our study area, the field boundary usually consists of a drainage ditch with perennial vegetation, mainly grasses, on its banks. These areas are managed so that only a few Salix bushes or single trees are growing along the drainage areas. Another field patch, or a road, houses, gardens, forest, or other non-crop areas may be located on the other side of the drainage ditch. The ditches are often deep and covered in dense vegetation, making them unusable for foraging by pheasant broods. Some of our fields are bordered by creeks and riverbanks bearing year-round grassy vegetation and occasional trees. At the time of our study, no obligatory EU regulations were in place for field margins. Despite the lack of regulation and the farmers striving to sow the crops as close to the field boundary as possible, an area bearing less vegetation was always left between the crop and the boundary itself. We estimated this margin area to average a width of one meter in our study area. For further analyses of the grain fields, we also calculated zones at 5-m intervals within the cultivated fields, in the main crop, resulting in five zones between 0 and 25 m starting from the field margin (see Pic 2 in Electronic Supplementary Material). We calculated the field surface area within each zone by creating buffers with the buffer tool provided by the QGIS 2.18 Development Team (2019).
Fieldwork
During 1995–1998, we introduced 31 wild translocated pheasant hens and 32 hand-reared pheasant hens to the study area. All hens were fitted with VHF radio collars enabling location via a portable antenna. Information of the study pheasants, capture and rearing techniques, radio equipment used, and release methods, along with survival and breeding success, are described in detail in Kallioniemi et al. (2015).
We located the nesting sites and observed the broods from hatching to approximately 4 weeks of age (29–31 days), at least three times per week. Chick occurrence was verified whenever possible, without disturbing the brood or hen excessively. To avoid any timing-based bias, we collected data at varying times of the day. To avoid autocorrelation, i.e., correlation between subsequent observations of the same brood, we observed the broods at random times, with at least two hours between each observation. Still, obtaining true independence is difficult in radio-tracking studies, as shown by Rooney et al. (1998), and this must be considered when interpreting our results. Tracking was conducted between 5.30 am and 12 pm. During our study period, the time between sundown and sunrise is ca. five hours (11 pm–4 am), and there is no period of actual darkness during that time.
During the study years, we were able to collect sufficient data (Aebischer et al. 1993) from 15 broods for habitat analysis (Table 1). Of these 15 hens, 12 were wild and three were hand reared. As we only had three hand-reared hens, we combined their data with the wild hens. Altogether, we obtained 458 brood observations in eleven habitat categories.
Home range
We used minimum convex polygon (MCP) (Mohr 1947) home range estimates to describe the outer limits of each brood’s movements. Movements were recorded after the brood left the nest. We measured each brood’s distance from the nest for the first two days. We also measured the distance from the nests to the home range centroids. Nest locations were not included in the home range analysis. The home ranges for 10-day and 31-day observations were counted with MCP100, MCP95, and MCP50 using ArcGIS 9.2. (ESRI 2006).
The minimum number of tracking fixes per bird required to accurately determine the home range size is difficult to assess. For example, the home range of a pheasant brood grows as the chicks grow and eventually begin flying (Warner 1979). Therefore, obtaining asymptotic data that would determine the point where the home range ceases to grow is not possible (Harris et al. 1990). As a general guideline, Dowell et al. (1993) suggest that approximately 30 locations should be adequate for assessing the home range and proportional habitat utilization of most pheasant species. As our study object is a hen with chicks, its movements are more restricted than those of a regular adult pheasant. To obtain samples that show uniform monitoring intensity, we only included broods with observations from every study week. Hence, we assumed that 17 tracking fixes, i.e., the least number of observations, divided evenly over the study period, was sufficient for describing the home range of a pheasant brood over the first 31 days after hatching.
Habitat analysis
We used a pheasant hen with chicks as a sample unit to achieve an appropriate sampling level (Kenward 1992). The habitat description was performed on a 10 × 10 m square, so we tried to locate the brood very precisely. We collected information on the habitat category and the distances to the forest and the field margin in meters. Habitats were classified into eleven categories: (1) grain fields, (2) grasslands mowed twice during the summer, (3) pea fields, (4) rape fields, (5) other cultivated fields (potato, linseed, mustard, onions), (6) pastures, (7) field margins, (8) gardens (including parks), (9) fallows (including meadows and set-aside), (10) forest, and (11) roads.
The use of compositional analysis solves the unit-sum constraint typical of compositional data for assessing habitat selection (Aebischer et al. 1993; Dowell et al. 1993). This method assumes that each animal provides an independent measure of habitat use within the population, and that the composition of various habitats and their usage by various animals are equally accurate. Unequal sampling of individual animals does not affect the overall analysis provided that sampling intensity is adequate for deriving accurate estimates of mean habitat use or, at the very least, the estimates from individual animals are equally accurate or stable (see Smith and Racey 2005).
Compositional analysis also addresses the definition of available habitat (Aebischer et al. 1993). An animal’s use of the available habitat is an outcome of choices made at different levels affected by several factors. The habitat use of animals is an outcome of various ecological and behavioral variables and the mere area of a habitat class may not in reality fully match the amount of habitat deemed available by the animal itself (Wiens 1973, Johnson 1980). Because habitat selection may occur on several levels, habitat analyses should be carried out in stages as we did in our analyses (Aebischer et al. 1993).
For analyzing the data, all grasslands and pastures were combined under “Grass” and gardens and fallows under “Fallow”. To avoid possible misclassification problematics discussed by Bingham et al. (2007), we combined two categories of cultivated areas that are more seldomly frequented by pheasant broods, i.e., “Rape” and “Pea”, into the category “Other”. Habitat class 11, “Roads”, was excluded with zero observations, and class 10, “Forest”, was removed with only two observations, thus leaving us with six habitat classes. Despite this, our habitat use data still contained a high proportion of zeros. To avoid the statistical problems associated with many use proportions being zero, we replaced them with 0.01%, as recommended by Aebischer et al. (1993). We used randomization-based p values, as the data were not normally distributed.
As suggested by Aebischer et al. (1993), we compared habitat utilization with the availability of each habitat class at two levels. First, we examined home range selection within the entire study area by comparing the proportion of each habitat in the minimum convex polygon (MCP100) with that available in the study area. Secondly, we examined habitat use within the home ranges by comparing the proportion of tracking fixes in each habitat class with the availability of habitat classes within the home range. We used compositional analysis to test whether habitat use by the broods differs from random habitat use. All analyses were conducted in R 4.0.5 (R Core Team, 2021), and we used the R package adehabitatHS for the compositional analysis (Calenge 2006).
Distance to margin in grain fields and trends in habitat use
We separately examined the grain field observations to ascertain whether the broods preferred some certain part of the field. To search for an edge effect, we compared how the observations were divided across the 5-m buffer zones (0–25 m) and in the rest of the field area with the proportion of the areas. Finally, we compared habitats selected during weeks 1, 2, 3, and 4, to see whether they showed a phenology trend in habitat use during our observation time of approximately 4 weeks.
Results
Nests and early movements
Pheasant nests (n = 15) were mainly located on fallows, forest edges, pastures, and in gardens. None of these areas are managed annually in our study area. The average vegetation height at the nest sites was 78.9 cm (SD 25.7, min 40 cm, max 130 cm), measured after a verified hatching.
The average hatching day was the 3rd of July, 42 days after release (SD 7, min 34, max 53 days). From this, we can estimate that a successful incubation began an average of 17 days after the release. As stated earlier, the average clutch-laying time is 15 days, meaning that the birds found a successful nesting site only a few days after being released.
After hatching, the broods moved to their to-be home ranges. The average distance moved during the first day was 95 m (n = 13; SD = 75, range 8–201 m), and after the second day they had already moved an average of 185 m away from the nest (n = 12; SD 135, min 41 m, max 550 m).
The median distance from the nests to the MCP50 centroids for the 31-day home ranges was 187 m (SD 150, min 62 m, max 528 m).
Home ranges
Home range sizes were measured for 10-day and 31-day observations. In the 10-day analysis, we left out two broods with too few observations. The mean number of observations was 12.8 (SD 2.35, min 10, max 18). The median home range size for 10 days (MCP95) was 1.3 hectares (mean 1.4, SD 0.9, min 0.2 ha, max 3 ha) and 4.8 ha for 31 days (mean 6.1, SD 5.1, min 1.72 ha, max 21.7 ha) (MCP95). The median MCP50 area for 31 days was 0.7 hectares (mean 1.2, SD 1.4, min 0.4 ha, max 5.8 ha).
Habitat use
Most observations were made in the grain fields (43.6%), which was also the most common habitat type in the area (52.5%). The “Fallow” category had the second most observations (18%), which correlates with an average proportion of 17.9% in the study area. Field margins were the most used area in relation to availability, with 15.8% of observations recorded from an area that had a cover of only 1.6%. Even if the field margins were counted as being relatively broader, e.g., as 5-m-wide buffers, their occurrence was only 8%. Field margin was also the only habitat present on all brood home ranges. “Grain” and “Fallow”, the other two most used habitat categories, were absent from two home ranges. An overall comparison between tracking fixes and habitats available in the study area indicates that field margins seem to be the most preferred area for broods (Fig. 1).

Brood observations divided into the following categories: “Grain”, “Grass” (grasses, pastures), “Other” (pea-, rape-, linseed-, potato fields), “Margin” (field margins), “Fallow” (gardens, meadows, fallows), and their proportions of the total observations (n = 456) compared to the proportions of available habitat (430.6 ha)
We investigated habitat use more closely using compositional analysis, where each tracking fix is directly linked to the habitat availability of a brood’s home range and home ranges are compared with the availabilities of the corresponding year.
Habitat analysis level 1—study area versus home range—showed that the habitat distribution within the pheasant brood home ranges differed significantly from the occurrence of habitat types in the study area (Wilk’s lambda = 0.174, p < 0.001). A matrix (Table 2.) of the mean and standard error log-ratio differences for all possible pairs of habitat types indicates which habitat types were relatively most occupied and which were least occupied. The habitats in the home ranges compared with the study area rank Margin > Fallow > Grain > Grass > Other.
Habitat analysis on level 2—home range versus tracking fixes. Comparing habitat utilization (tracking fixes) with the habitat composition of the home ranges differed significantly (Wilk’s lambda = 0.108, p = 0.0154).
The ranking matrix (Table 3) indicates that the margin was by far the most used habitat, with a significant difference to grain fields, i.e., the second most used habitat. Habitats used by the broods in their home ranges rank Margin > > > Grain > Grass > Fallow > Other.
Compared with availability in the home ranges, “Margin” was used significantly more often than the other habitats. “Grain” was significantly more preferred than “Other”, but otherwise no significant differences in usage were observed between “Grain”, “Fallow”, “Grass”, and “Other”.
Distance to field margin—tracking fixes on the grain fields
The median (mean, ± SD, range) distance from the brood location to the field margin for grain fields was 15 (22.4, ± 20.4, 0.5–120) m (n = 199), and 15 (22.9, ± SD 22.6, 0.5–150) m for the rest of the habitat groups (“Fallow”, “Grass”, and “Other”, n = 185).
We compared the observations of the most used habitat “Grain” with its availability at various distances from the field edges (Fig. 2). This showed a significant preference for the edge zone, as 68% of all grain field observations occurred within a 25-m-wide zone from the field edge, despite the proportion being 40% (G = 65.14, df = 1, p < 0.0001). Also, 54% of pheasant brood observations on grain fields were less than 15 m from the field edge, despite the proportion being 25% (G = 74.86, df = 1, p < 0.0001). This strongly indicates a preference for margins over the field centers. The median (mean, ± SD, range) distance from the tracking fixes to the “Forest” for grain field observations was 100 (120.9, ± 82.1, 2–500) meters, (n = 199) and 50 m for all other observations (68.2, ± 64.3, 0.5–500) (n = 257).

Observation distances of grain fields from the field margin. Cumulative observations show that 68% of the observations are within a 25-m zone from the field margin, but this only represents 40% of the field area available. Only 32% of the observations are over 25 m from the field margin, although this area represents 60% of the field area available
Trends in habitat use vs. brood age
To elucidate the changes occurring in habitat use as a brood ages, we compared the weekly changes occurring during the study period. We made 407 observations over 4 weeks, with 107 observations during the 1st week, and 104, 101, and 95 observations for the 2nd, 3rd, and 4th week, respectively (Fig. 3.). A slight change in the use of “Margin” and “Fallow” areas was detectable.

Habitat use shifted during the study weeks, as “Margin” became more favorable, while “Fallow” lost preference
Discussion
Our results show that field margin, the area where the main crop abuts the field boundary, e.g., a ditch, road, or non-crop vegetation, is an important and preferred habitat for pheasant broods throughout the brood stage. The majority of our brood observations were made in grain fields, which were the most common habitat type in the area. However, taking habitat availability into account, the most preferred habitat was field margins. The mere 1-m-wide strip of field margin may seem like too small a landscape feature for animals to prefer, yet the total length of this strip spread along our study area, equaling 15 km/100 ha, describes the significance of such features, especially when considering the interchange between the semi-natural habitat, the margin, and the adjacent crop (Bianchi et al. 2006). The amount of field margin is also in relation to field patch size, which then again reflects the heterogeneity in the landscape composition and configuration that is often considered to enhance biodiversity (Benton et al. 2003; Fahrig et al. 2011, 2015, Clough et al. 2020, but see Hiron et al. 2015).
The importance of margins in our study may arise from at least two, mutually non-exclusive effects. Firstly, the preference for field margins may be due to a less disturbed habitat leading to higher arthropod availability. Secondly, pheasant broods may prefer margins due to this habitat providing more sparse vegetation, leading to openness, more weed sprouts, and a more suitable microclimate. Segetal vegetation is needed for arthropod habitat (Potts 1986), but concurrently it must be remembered that bare ground offers movement corridors for young pheasants, where they can capture prey and avoid predators (Doxon and Carroll 2010). An indication toward a preference for sparser vegetation could be detected in the habitat use shift during our study; fallow use declined and margin use increased as the season progressed. An alternative explanation could be that the pheasant hens dared to spend more time in the open margin habitats when their chicks were larger and more agile. Increased use of strip vegetation by older broods was also shown by Meyers et al. (1988). The concept of offering the chicks habitat containing both cover and open patches has proven effective when restoring partridge habitats in the North Sea Partridge Project (Brewin et al. 2020).
Field margins, along with other semi-natural habitats, have been described as biodiversity reservoirs in intensified agricultural landscapes, and their biodiversity value has been shown by many studies (Bianchi et al. 2006; Vickery et al. 2009; Holland et al. 2017; Martin et al. 2019; Šálek et al. 2021). The concept of biodiversity-rich margins has been developed further with the idea of arable margins placed between field boundaries and the main crops (Meek et al. 2002; Vickery et al. 2009). Herzon and O’Hara (2007) stated that out of all non-crop elements, the extent of ditches and small rivers was the strongest positive predictor of bird communities, and the only one with an exclusively positive effect on individual species. Ekroos et al. (2019) also pointed out that edge density, which is not supported by the Agri-Environmental Scheme (AES) greening measures, had the highest impact on farmland bird assemblages. The value of field margins, along with other non-crop habitat elements adjacent to fields, extends beyond conservation values. Even wildlife welfare may benefit from areas offering refuge to animals displaced by in-field operations (Mathews 2010).
The role of field margins as the most preferred habitat was supported by our other finding, which showed that most grain field observations, i.e., the most numerous observations, were made in the proximity of the margins. This suggests that the combination of margin and grain field offers an ideal habitat for pheasant broods that need high-quality arthropods and shelter, also indicating high biodiversity. This margin effect has been noted by earlier studies: both gray and red-legged partridge (Alectoris rufa) broods avoided areas more than 50 m from the field edge and preferred areas within 25 m of the edge (Green 1984). Best et al. (1990) found that the length of linear field margin relative to the field area had a major effect on cornfield use by birds, as larger fields were used proportionately less. Holland et al. (1999) showed that many arthropod species occur within 60 m of the field edge. Furthermore, Green (1984) found that arthropods and weeds used as food by partridges occurred at higher densities 5 m from the field boundary compared with 50 m. The importance of field boundaries to many carabid and staphylinid beetles was also noted by Andersen (1997). This field edge effect could be strengthened by decreasing pesticide use, as seen with unsprayed headlands that enhance arthropod diversity and numbers (Chiverton and Sotherton 1991; Helenius et al. 1995; Meek et al. 2002). Schindler et al. (2020) showed the importance of a mosaic of croplands and grasslands as a habitat for pheasants in the USA.
The lack of proper nesting places has been indicated as a threshold for pheasant success in certain areas (Robertson 1996). However, pheasant hens in our study area found successful nesting places quickly after release, indicated by the fact that the hens in our study were able to hatch broods within an average of 42 days from release. Our assumption that the hens would choose an inconspicuous nesting place, and therefore prefer set-asides and other areas with covering vegetation already present (dead grasses from last year, bushes, or other perennial crops that offer nesting cover), proved correct. Our study pheasant hen nests were located in semi-natural areas, as expected. Hill and Robertson (1988) showed that re-nesting later in the season was more common on cultivated lands than first nests were, probably because of how the vegetation developed. The vegetation was just beginning to regain its growth during the bird release.
After hatching, the broods left the nests and, within approximately 2 days, reached the area where they then remained. Some broods even crossed tarmac roads and dense fallows to reach cultivated fields. It is possible that during the incubation period, hens search for areas that offer suitable invertebrate supply and other preferred habitat qualities for the broods, and then guide their chicks there as quickly as possible. Such behavior has been shown in ducks (Casazza et al. 2020). The hen leaves the nest for only short periods during the egg-laying and incubation periods, to keep the eggs from chilling. However, even throughout this period, the hen will avoid feeding in close proximity to the nest, avoiding an area under 50 m from the nest. This has been considered a way to prevent nest predation (Hill and Robertson 1988), and perhaps the transition from the nest to the home range also serves this purpose.
Previous studies have shown rather large home ranges for 10-day-old broods spanning from ca. 5 to 11 ha (Hanson and Progulske 1973; Warner 1979; Hill 1985). Even though our value of 1.4 ha (0.2–3.0 ha) can only be used as descriptive because of the small number of observations, it seems that the broods in our study quickly found high-quality home ranges. Larger home ranges have been connected to higher chick mortality and they also correlate with fewer arthropods (Warner 1979; Hill 1985). Warner (1979) showed that a brood home range increases as the chicks grow. This could also be seen in our results. Riley et al. (1998) observed broods for 28 days in the USA on two row crop production areas for 5 years and calculated home range averages of 66 and 76 ha (range 15–179 ha). Using MCP for home range analysis is occasionally criticized for its deficiencies and robustness (Laver and Kelly 2008), but our sampling methods and data size, along with our basic knowledge of the research object and its habitat use allowed us to assume that the MCP estimate will provide accurate enough information of home range sizes for pheasant broods and allowed us to compare the results with earlier studies. One explanation for the smaller home ranges in our study may be that, according to FAO (2020), pesticide use is not as intensive in Finland as in the USA (five times more per ha) or in Europe (UK/France) (ten times more per ha), likely leading to less disturbed arthropod communities (Stoate et al. 2009). The average small field patch size in our study area increases the quantity of margin area, which increases diversity per se (Fahrig et al. 2015). Brood movements are much broader and home ranges larger on large monoculture blocks than in broods feeding within a more diverse complex of farming (Warner 1984; Riley et al. 1998). Indeed, Hill (1985) showed that habitats selected by broods largely reflected the levels of arthropod food that they contained.
Spatial scales matter when investigating avian communities (Wiens 1989). We must scale out from individual habitat requirements to observe processes that drive biodiversity and are governed by landscape composition. These processes are affected by the levels of semi-natural habitats in the landscape, and, on the other hand, by landscape configuration, such as the amount of field margins. Both these factors affect ecological processes independently and interactively (Fahrig et al. 2011). Landscape composition and structure may have more of an impact on the success of a certain species than suitable habitats, such as field margins, have on a smaller scale (Bennett et al. 2006; Frei et al. 2018). For example, the taxonomic richness and diversity of invertebrates in field edges have been observed to positively relate to large-scale landscape complexity (Evans et al. 2016). Sasaki et al. (2020) noted the importance of open land in maintaining farmland biodiversity in a forest-dominated landscape. Jorgensen et al. (2014) found that “Forest” habitat in a landscape was a limiting factor for pheasants, probably because of predators. In our study, 54% of the study area was defined as “Forest”, which could also be detected in the short mean distance to forest areas (under 100 m). In Finland, trees serve as perches for predators, e.g., for birds of prey and nest-predating corvids, which may be important nest predators of ground-nesting birds (Andrén 1992; Krüger et al. 2018; Holopainen et al. 2020a). Forests also provide dens for meso-predators, such as the red fox (Vulpes vulpes), badger (Meles meles), and raccoon dog (Nyctereutes procyonoides). The red fox can even be a threat to adult hens, while the latter two mainly cause nesting failures due to predation and disturbance (Kallioniemi et al. 2015; Holopainen et al. 2020a, 2021). Recent studies have highlighted the role of invasive alien predators in elevating nest and adult predation and thus reducing the resilience of ground-nesting birds. The increasing numbers of invasive raccoon dogs pose a severe threat to ground-nesting birds in general in Northern Europe (Krüger et al. 2018; Nummi et al. 2019; Holopainen et al. 2020a, 2020b; Koshev et al. 2020; Jaatinen et al. 2022). To ensure successful management, it is therefore crucial to understand that even though a habitat as such is suitable for brood rearing, predation pressure can be too high for an area to uphold a successful pheasant population.
Conclusions and recommendations for agricultural policy
We found that pheasant broods preferred field margins over all other farmland habitats. On an area-for-area basis, field margins are effective at providing food for birds and enhancing biodiversity (Marshall and Moonen 2002; Vickery et al. 2002, 2009) and could easily be integrated into whole-field management practices. These keystone structures (Tews et al. 2004) adjacent to croplands could be a cost-efficient way to preserve biodiversity even within intensive farming practices, when integrated with low-intensity practices such as methods that aim at minimized pesticide application near margins (Vickery et al. 2002). Agricultural areas with small grain crop cultivation could have particularly great potential in providing more biodiversity. Land use intensification methods, e.g., subsurface drainage, which reduce margins and the open-field ditch-bank habitat (Helenius et al. 1995; Tarmi 2011), should be compensated for with, e.g., beetle banks, flower strips, and other biodiversity-enhancing methods (Brewin et al. 2020). The quantity of field margins, i.e., field edge density, is currently not addressed in national agri-environment schemes, while the enlargement of field parcels and replacement of open ditches with subsurface drainage is still an active agricultural policy target aiming to further modernize agriculture in Finland (Häggblom et al. 2020). Field margins can also offer a supplementary flowering season for pollinators (Memmott et al. 2010), which are of special concern in the new EU biodiversity strategy for 2030 (EU Publications Office, 2020).
Recent studies evaluating the value of AES measures in Finland recognized animal farms practicing ecological production (Santangeli et al. 2019), along with increasing the proportion of grasslands and fallows (Ekroos et al. 2019), as beneficial for farmland birds. The need to begin appreciating field margins as vital biodiversity elements in agricultural landscapes is underlined by Ekroos et al. (2019), who found that field edge densities had a much stronger effect on farmland bird diversity than the AES greening measures they studied. The analysis by Martin et al. (2019) connected edge density to higher functional biodiversity and to higher yield-enhancing ecosystem services in European landscapes. Reducing crop field sizes and thereby increasing the amount of margin and structural diversification instead of focusing on set-asides and organic farming is recommended as a solution to halting biodiversity loss in agricultural areas (Sirami et al. 2019; Clough et al. 2020; Šálek et al. 2021). Considering these results, changes in agricultural policy toward favoring the biodiversity-boosting effects of margins and their surroundings should be obvious.
References
Aebischer NJ, Ewald JA (2004) Managing the UK grey partridge Perdix perdix recovery: population change, reproduction, habitat and shooting. Ibis 146:181–191. https://doi.org/10.1111/j.1474-919X.2004.00345.x
Aebischer NJ, Robertson PA, Kenward RE (1993) Compositional analysis of habitat use from animal radio tracking data. Ecology 75:1313–1325
Altieri MA (1999) The ecological role of biodiversity in agroecosystems. Agric Ecosyst Environ 74:19–31. https://doi.org/10.1016/S0167-8809(99)00028-6
Andersen A (1997) Densities of overwintering carabids and staphylinids (Col. Carabidae and Staphylinidae) in cereal and grass fields and their boundaries. J Appl Entomol 121:77–80. https://doi.org/10.1111/j.1439-0418.1997.tb01374.x
Andrén H (1992) Corvid density and nest predation in relation to forest fragmentation: a landscape perspective. Ecology 73:794–804. https://doi.org/10.2307/1940158
Bennett AF, Radford JQ, Haslem A (2006) Properties of land mosaics: implications for nature conservation in agricultural environments. Biol Conserv 133:250–264. https://doi.org/10.1016/j.biocon.2006.06.008
Benton TG, Vickery JA, Wilson JD (2003) Farmland biodiversity: is habitat heterogeneity the key? Trends Ecol Evol 18:182–188. https://doi.org/10.1016/S0169-5347(03)00011-9
Best LB, Whitmore RC, Booth GM (1990) Use of cornfields by birds during the breeding season: the importance of edge habitat. Am Midl Nat 123:84–99. https://doi.org/10.2307/2425762
Bianchi FJJA, Booij CJH, Tscharntke T (2006) Sustainable pest regulation in agricultural landscapes: a review on landscape composition, biodiversity and natural pest control. Proc R Soc Biol Sci Ser B 273:1715–1727. https://doi.org/10.1098/rspb.2006.3530
Bingham RL, Brennan LA, Ballard BM (2007) Misclassified resource selection: compositional analysis and unused habitat. J Wildl Manage 71:1369–1374. https://doi.org/10.2193/2006-072
BirdLife International (2015) European list of birds, Office for Official Publications of the European Communities, Luxembourg, Available at http://www.birdlife.org/datazone/info/euroredlist. Accessed 3 May 2021
Brewin J, Buner F, Ewald J (2020) Farming with nature—promoting biodiversity across Europe through partridge conservation. The Game and Wildlife Conservation Trust, Fordingbridge, UK
Brittas R, Marcström V, Kenward RE, Karlbom M (1992) Survival and breeding success of reared and wild ring-necked pheasants in Sweden. J Wildl Manage 56:368–376. https://doi.org/10.2307/3808836
Butler SJ, Boccaccio L, Gregory RD, Vorisek P, Norris K (2010) Quantifying the impact of land-use change to European farmland bird populations. Agric Ecosyst Environ 137:348–357. https://doi.org/10.1016/j.agee.2010.03.005
Calenge C (2006) The package “adehabitat” for the R software: tool for the analysis of space and habitat use by animals. Ecol Model 197:1035. https://doi.org/10.1016/j.ecolmodel.2006.03.017
Casazza ML, McDuie F, Lorenz AA, Keiter D, Yee J, Overton CT, Peterson SH, Feldheim CL, Ackerman JT (2020) Good prospects: high-resolution telemetry data suggests novel brood site selection behaviour in waterfowl. Anim Behav 164:163–172. https://doi.org/10.1016/j.anbehav.2020.04.013
Chamberlain DE, Fuller RJ, Bunce JC, Duckworth JC, Shrubb M (2000) Changes in the abundance of farmland birds in relation to the timing of agricultural intensification in England and Wales. J Appl Ecol 37:771–788. https://doi.org/10.1046/j.1365-2664.2000.00548.x
Chiverton PA, Sotherton NW (1991) The effects on beneficial arthropods of the exclusion of herbicides from cereal crops. J Appl Ecol 28:1027–1039. https://doi.org/10.2307/2404223
Clough Y, Kirchweger S, Kantelhardt J (2020) Field sizes and the future of farmland biodiversity in European landscapes. Conserv Lett 13(6):e12752. https://doi.org/10.1111/conl.12752
Coates PS, Brussee BE, Howe KB, Fleskes JP, Dwight IA, Connelly DP, Meshriy MG, Gardner SC (2017) Long-term and widespread changes in agricultural practices influence ring-necked pheasant abundance in California. Ecol Evol 7:2546–2559. https://doi.org/10.1002/ece3.2675
Donald PF, Sanderson FJ, Burfield IJ, van Bommel FPJ (2006) Further evidence of continent-wide impacts of agricultural intensification on European farmland birds, 1990–2000. Agr Ecosyst Environ 116:189–196. https://doi.org/10.1016/j.agee.2006.02.007
Dowell SD, Aebischer NJ, Robertson PA (1993) Analysing habitat use from radio-tracking data. In: Jenkins D (ed) Pheasants in Asia 1992. World Pheasant Association, Reading, UK, pp 62–66
Doxon ED, Carroll JP (2010) Feeding ecology of ring-necked pheasant and northern bobwhite chicks in conservation reserve program fields. J Wildl Manage 74:249–256. https://doi.org/10.2193/2008-522
EBCC European Bird Census Council (2017) Trends of Common Birds in Europe, 2017 Update, available at http://ebcc.birdlife.cz/trends-of-common-birds-in-europe-2017-update/. Accessed 5 May 2021
EFBI (2021) European Farmland bird index, available at https://agridata.ec.europa.eu/Qlik_Downloads/InfoSheetEnvironmental/infoC35.html. Accessed 1 April 2021
European Council (2001) Presidency conclusions, Gothenburg Council, 15 and 16 June 2001. SN/200/1/01 REV1, 8 pp.
Ekroos J, Tiainen J, Seimola T, Herzon I (2019) Weak effects of farming practices corresponding to agricultural greening measures on farmland bird diversity in boreal landscapes. Landscape Ecol 34:389–402. https://doi.org/10.1007/s10980-019-00779-x
ESRI (2006) ArcGIS Desktop: Release 9.2. Environmental Systems Research Institute Inc, Redlands, CA, USA
EU Publications Office (2020) EU biodiversity strategy for 2030, available at https://eur-lex.europa.eu/legal-content/EN/LSU/?uri=CELEX:52020DC0380. Accessed 15 June 2021
Evans KL (2004) The potential for interactions between predation and habitat change to cause population declines of farmland birds. Ibis 46:1–13. https://doi.org/10.1111/j.1474-919X.2004.00231.x
Evans TR, Mahoney MJ, Cashatt ED, Noordijk J, de Snoo G, Musters CJ (2016) The impact of landscape complexity on invertebrate diversity in edges and fields in an agricultural area. Insects 7:7. https://doi.org/10.3390/insects7010007
Ewald JA, Aebischer NJ, Richardson SM, Grice PV, Cooke AI (2010) The effect of agri-environment schemes on grey partridges at the farm level in England. Agr Ecosyst Environ 138:55–63. https://doi.org/10.1016/j.agee.2010.03.018
Fahrig L, Baudry J, Brotons L, Burel FG, Crist TO, Fuller RJ, Sirami C, Siriwardena GM, Martin JL (2011) Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecol Lett 14:101–112. https://doi.org/10.1111/j.1461-0248.2010.01559.x
Fahrig L, Girard J, Duro D, Pasher J, Smith A, Javorek S, King D, Lindsay KF, Mitchell S, Tischendorf L (2015) Farmlands with smaller crop fields have higher within-field biodiversity. Agric Ecosyst Environ 200:219–234. https://doi.org/10.1016/j.agee.2014.11.018
FAO (2020) FAOSTAT, Agri-environmental Indicators/Pesticides, available at http://www.fao.org/faostat/en/#data/EP. Accessed 14 May 2021.
Fischer J, Abson DJ, Bergsten A, Collier NF, Dorresteijn I, Haspach J, Hylander K, Schultner J, Senbeta F (2017) Reframing the food-biodiversity challenge. Trends Ecol Evol 32:335–345. https://doi.org/10.1016/j.tree.2017.02.009
Frei B, Bennett EM, Kerr JT (2018) Cropland patchiness strongest agricultural predictor of bird diversity for multiple guilds in landscapes of Ontario, Canada. Reg Environ Change 18:2105–2115. https://doi.org/10.1007/s10113-018-1343-5
Gabriel D, Sait SM, Kunin WE, Benton TG (2013) Food production vs. biodiversity: comparing organic and conventional agriculture. J Appl Ecol 50:355–364. https://doi.org/10.1111/1365-2664.12035
González del Portillo D, Arroyo B, García Simón G, Morales MB (2021) Can current farmland landscapes feed declining steppe birds? Evaluating arthropod abundance for the endangered little bustard (Tetrax tetrax) in cereal farmland during the chick-rearing period: variations between habitats and localities. Ecol Evol 11:3219–3238
Green RE (1984) The feeding ecology and survival of partridge chicks (Alectoris rufa and Perdix perdix) on arable farmland in East Anglia. J Appl Ecol 21:817–830. https://doi.org/10.2307/2405049
Gregory RD, van Strien AJ, Vorisek P, Gmelig Meyling AW, Noble DG, Foppen RPB, Gibbons DW (2005) Developing indicators for European birds. Philos Trans R Soc B 360:269–288. https://doi.org/10.1098/rstb.2004.1602
Häggblom O, Härkönen L, Joensuu S, Keskisarja V, Äijö H (2020) Water management guidelines for agriculture and forestry. Publications of the Ministry of Agriculture and Forestry. In Finnish, summary in English, available at http://urn.fi/URN:ISBN:978-952-366-186-8
Hanson LE, Progulske DR (1973) Movements and cover preferences of pheasants in South Dakota. J Wildl Manage 37:454–461. https://doi.org/10.2307/3800308
Harris S, Creswell WJ, Forde PG, Trewhella WJ, Wollard T, Wray S (1990) Home range analysis using radio-tracking data; a review of problems and techniques particularly as applied to the study of mammals. Mammal Rev 20:97–123. https://doi.org/10.1111/j.1365-2907.1990.tb00106.x
Helenius J, Tuomola S, Nummi P (1995) Food availability for the grey partridge in relation to changes in the arable environment. In Finnish, summary in English. Suomen Riista 41:42–52
Herzon I, O’Hara RB (2007) Effects of landscape complexity on farmland birds in the Baltic States. Agric Ecosyst Environ 118:297–306. https://doi.org/10.1016/j.agee.2006.05.030
Hietala-Koivu R (2002) Landscape and modernizing agriculture: a case study of three areas in Finland in 1954–1998. Agr Ecosyst Environ 91:273–281. https://doi.org/10.1016/S0167-8809(01)00222-5
Hill DA (1985) The feeding ecology and survival of pheasant chicks on arable farmland. J Appl Ecol 22:645–654. https://doi.org/10.2307/2403218
Hill D, Robertson P (1988) The Pheasant: ecology, management and conservation, In BSP Professional Books
Holland M, Douma JC, Crowley L, James L, Kor L, Stevenson DRW, Smith BM (2017) Semi-natural habitats support biological control, pollination and soil conservation in Europe. A Review Agron Sustain Dev 37:31. https://doi.org/10.1007/s13593-017-0434-x
Holland JM, Perry JN, Winder L (1999) The within-field spatial and temporal distribution of arthropods in winter wheat. Bull Entomol Res 89:499–513. https://doi.org/10.1017/S0007485399000656
Holopainen S, Väänänen V-M, Fox A (2020a) Landscape and habitat affects frequency of native but not alien predation of artificial duck nests. Basic Appl Ecol 48:52–60. https://doi.org/10.1016/j.baae.2020.07.004
Holopainen S, Väänänen V-M, Fox A (2020b) Artificial nest experiment reveals inter-guild facilitation in duck nest predation. Global Ecol Conserv 24:e01305. https://doi.org/10.1016/j.gecco.2020.e01305
Holopainen S, Väänänen V-M, Vehkaoja M, Fox A (2021) Do alien predators pose a particular risk to duck nests in Northern Europe? Results from an artificial nest experiment. Biol Invasions. https://doi.org/10.1007/s10530-021-02608-2
IUCN (2021) The IUCN red list of threatened species. Version 2021-1, available at https://www.iucnredlist.org. Accessed 14 March 2021
Jaatinen K, Hermansson I, Mohring B, Steele B, Öst M (2022) Mitigating impacts of invasive alien predators on an endangered sea duck amidst high native predation pressure. Oecologia 198:543–552. https://doi.org/10.1007/s00442-021-05101-8
Jorgensen CF, Powell LA, Lusk JJ, Bishop AA, Fontaine JJ (2014) Assessing landscape constraints on species abundance: does the neighborhood limit species response to local habitat conservation programs? PLoS ONE 9:e99339. https://doi.org/10.1371/journal.pone.0099339
Kallioniemi H, Väänänen V-M, Nummi P, Virtanen J (2015) Bird quality, origin and predation level affect survival and reproduction of translocated common pheasants Phasianus colchicus. Wildl Biol 21:269–276. https://doi.org/10.2981/wlb.00052
Kleijn D, Rundlöf M, Scheper J, Smith HG, Tscharntke T (2011) Does conservation on farmland contribute to halting the biodiversity decline? Trends Ecol Evol 26:474-481. https://doi.org/10.1016/j.tree.2011.05.009
Kenward RE (1992) Quantity versus quality: programming for collection and analysis of radio tag data. In: Priede IG, Swift SM (eds) Wildlife telemetry: remote monitoring and tracking of animals. Ellis Horwood, Chichester, UK, pp 231–246
Koshev YS, Petrov MM, Nedyalkov NP, Raykov IA (2020) Invasive raccoon dog depredation on nests can have strong negative impact on the Dalmatian pelican’s breeding population in Bulgaria. Eur J Wildl Res 66:85. https://doi.org/10.1007/s10344-020-01423-9
Krüger H, Väänänen V-M, Holopainen S, Nummi P (2018) The new faces of nest predation on agricultural landscapes—a wildlife camera survey with artificial nests. Eur J Wildl Res 64:76. https://doi.org/10.1007/s10344-018-1233-7
Langgemach T, Bellebaum J (2005) Predation and the conservation of ground-breeding birds in Germany. Vogelwelt 126:259–298
Laver P, Kelly M (2008) A critical review of home range studies. J Wildl Manage 72:290–298. https://doi.org/10.2193/2005-589
Layerace (2022) Map of the world, available at https://www.freepik.com/vectors/map-world. Uploaded 14.4.2022
Marshall EJR, Moonen AC (2002) Field margins in northern Europe: their functions and interactions with agriculture. Agr Ecosyst Environ 89:5–21. https://doi.org/10.1016/S0167-8809(01)00315-2
Martin EA, Dainese M, Clough Y, Báldi A, Bommarco R, Gagic V et al (2019) The interplay of landscape composition and configuration new pathways to manage functional biodiversity and agroecosystem services across Europe. Ecol Lett 22:1083–1094. https://doi.org/10.1111/ele.13265
Mathews F (2010) Wild animal conservation and welfare in agricultural systems. Anim Welf 19:159–217
Meek B, Loxton D, Sparks T, Pywell R, Pickett H, Nowakowski M (2002) The effect of arable field margin composition on invertebrate biodiversity. Biol Cons 106:259–271. https://doi.org/10.1016/S0006-3207(01)00252-X
Memmott J, Carvell C, Pywell RF, Craze PG (2010) The potential impact of global warming on the efficacy of field margins sown for the conservation of bumblebees. Philos T R Soc B 365:2071–2079. https://doi.org/10.1098/rstb.2010.0015
Meyers SM, Crawford JA, Haensly TF, Castillo WJ (1988) Use of cover types and survival of ring-necked pheasant broods. Northw Sci 62:36–40
Mohr CO (1947) Table of equivalent populations of North American small mammals. Am Midl Nat 37:223–249. https://doi.org/10.2307/2421652
Musil DD, Connelly JW (2009) Survival and reproduction of pen-reared vs translocated wild pheasants Phasianus colchicus. Wildl Biol 15:80–88. https://doi.org/10.2981/07-049
National Land Survey of Finland (2019) Maastokarttarasteri, 1:50 000. CSC—Tieteen tietotekniikan keskus Oy, available at http://urn.fi/urn:nbn:fi:att:61bdc714-d16f-4dc4-8d5e-1d3080c80525
Newton I (2004) The recent declines of farmland bird populations in Britain: an appraisal of causal factors and conservation actions. Ibis 146:579–600. https://doi.org/10.1111/j.1474-919X.2004.00375.x
Nielson RM, McDonald LL, Sullivan JP, Burgess C, Johnson DS, Johnson DH, Bucholtz S, Hyberg S, Howlin S (2008) Estimating the response of ring-necked pheasants (Phasianus colchicus) to the conservation reserve program. Auk 125:434–444
Nummi P, Väänänen V-M, Pekkarinen A-J, Eronen V, Mikkola-Roos M, Nurmi J, Rautiainen A, Rusanen P (2019) Alien predation in wetlands—raccoon dog and waterbird breeding success. Baltic for 25:228–237. https://doi.org/10.46490/vol25iss2pp228
Panek M, Bresinski W (2002) Red fox Vulpes vulpes density and habitat use in a rural area of western Poland in the end of the 1990s, compared with the turn of the 1970s. Acta Theriol 47:433–442. https://doi.org/10.1007/BF03192468
Ponce C, Bravo C, Alonso JC (2014) Effects of agri-environmental schemes on farmland birds: do food availability measurements improve patterns obtained from simple habitat models? Ecol Evol 4:2834–2847. https://doi.org/10.1002/ece3.1125
Potts GR (1986) The Partridge: pesticides, predation and conservation. Collins, London
Potts GR, Aebischer NJ (1995) Population dynamics of the grey partridge Perdix perdix 1793–1993: monitoring, modelling and management. Ibis 137:29–37. https://doi.org/10.1111/j.1474-919X.1995.tb08454.x
Powell L (2015) Hitler’s effect on wildlife in Nebraska: world war II and farmed landscapes. Great Plains Quart 35:1–26. https://doi.org/10.1353/gpq.2015.0003
QGIS 2.18 Development Team (2019) QGIS Geographic Information System, Open Source Geospatial Foundation Project, available at http://qgis.osgeo.org
R Core Team (2021) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria
Riley TZ, Clark WR, Ewing DE, Vohs PA (1998) Survival of ring-necked pheasant chicks during brood rearing. J Wildl Manage 62:37–44. https://doi.org/10.2307/3802262
Robertson PA (1996) Does nesting cover limit abundance of ring-necked pheasants in North America? Wildlife Soc B 24:98–106
Robertson PA, Mill AC, Rushton SP, McKenzie AJ, Sage RB, Aebischer NJ (2017) Pheasant release in Great Britain: long-term and large-scale changes in the survival of a managed bird. Eur J Wildl Res 63:100. https://doi.org/10.1007/s10344-017-1157-7
Ronnenberg K, Strauß E, Siebert U (2016) Crop diversity loss as a primary cause of grey partridge and common pheasant decline in Lower Saxony. Germany BMC Ecol 16:39. https://doi.org/10.1186/s12898-016-0093-9
Rooney SM, Wolfe A, Hayden TJ (1998) Autocorrelated data in telemetry studies: time to independence and the problem of behavioural effects. Mammal Rev 28:89–98. https://doi.org/10.1046/j.1365-2907.1998.00028.x
Roos S, Smart J, Gibbons DW, Wilson JD (2018) A review of predation as a limiting factor for bird populations in mesopredator-rich landscapes: a case study of the UK. Biol Rev. https://doi.org/10.1111/brv.12426
Šálek M, Kalinová K, Daňková R, Grill S, Żmihorski M (2021) Reduced diversity of farmland birds in homogenized agricultural landscape: a cross-border comparison over the former Iron Curtain. Agric Ecosyst Environ 321:107628. https://doi.org/10.1016/j.agee.2021.107628
Santangeli A, Lehikoinen A, Lindholm T, Herzon I (2019) Organic animal farms increase farmland bird abundance in the Boreal region. PLoS ONE 14(5):e0216009. https://doi.org/10.1371/journal.pone.0216009
Sasaki K, Hotes S, Kadoya T, Yoshioka A, Wolters V (2020) Landscape associations of farmland bird diversity in Germany and Japan. Glob Ecol Conserv 21:e00891. https://doi.org/10.1016/j.gecco.2019.e00891
Schindler AR, Haukos DA, Hagen CA, Ross BE (2020) A multispecies approach to manage effects of land cover and weather on upland game birds. Ecol Evol 10:14330–14345. https://doi.org/10.1002/ece3.7034
Searchinger T, Waite R, Hanson C, Ranganathan J (2018) Creating a sustainable food future: a menu of solutions to feed nearly 10 billion people by 2050 (Synthesis Report). World Resources Institute
Silva-Monteiro M, Pehlak H, Fokker C, Kingma D, Kleijn D (2021) Habitats supporting wader communities in Europe and relations between agricultural land use and breeding densities: a review. Global Ecol Conserv 28:e01657. https://doi.org/10.1016/j.gecco.2021.e01657
Sirami C, Gross N, Baillod AB, Bertrand C, Carrie R, Hass A, Henckel L, Miguet P, Vuillot C, Alignier A, Girard J, Batary P, Clough Y, Violle C, Giralt D, Bota G, Badenhausser I, Lefebvre G, Gauffre B, Vialatte A, Calatayud F, Gil-Tena A, Tischendorf L, Mitchell S, Lindsay K, Georges R, Hilaire S, Recasens J, Sole-Senan XO, Robleno I, Bosch J, Barrientos JA, Ricarte A, Marcos-Garcia MA, Minano J, Mathevet R, Gibon A, Baudry J, Balent G, Poulin B, Burel F, Tscharntke T, Bretagnolle V, Siriwardena G, Ouin A, Brotons L, Martin JL, Fahrig L (2019) Increasing crop heterogeneity enhances multitrophic diversity across agricultural regions. PNAS 116(33):16442–16447. https://doi.org/10.1016/j.apgeog.2013.12.006
Smith RK, Pullin AS, Stewart GB, Sutherland WJ (2010) Effectiveness of predator removal for enhancing bird populations. Conserv Biol 24:820–829. https://doi.org/10.1111/j.1523-1739.2009.01421.x
Smith JA, Matthews TW, Holcomb ED, Negus LP, Davis CA, Brown MB, Powell LA, Taylor JS (2015) Invertebrate prey selection by ring-necked pheasant Phasianus colchicus broods in Nebraska. Am Midl Nat 173:318–325. https://doi.org/10.1674/amid-173-02-318-325.1
Smith PG, Racey PA (2005) Optimum effort to estimate habitat use when the individual animal is the sampling unit. Mammal Rev 35:295–301. https://doi.org/10.1111/j.1365-2907.2005.00074.x
Stoate C, Baldi A, Beja P, Boatman ND, Herzon I, van Doorn A et al (2009) Ecological impact of early 21st century agricultural change in Europe—a review. J Environ Manage 91:22–46. https://doi.org/10.1016/j.jenvman.2009.07.005
Stoate C, Boatman ND, Borralho RJ, Rio Carvalho C, de Snoo GR, Eden P (2001) Ecological impacts of arable intensification in Europe. J Environ Manage 63:337–365. https://doi.org/10.1006/jema.2001.0473
Tarmi S (2011) Plant communities of field margins: the effects of management and environmental factors on species composition and diversity, Academic Dissertation, University of Helsinki, available at http://urn.fi/URN:ISBN:978-952-10-4314-7
Taylor JS, Bogenschutz TD, Clark WR (2018) Pheasant responses to U.S. cropland conversion programs: a review and recommendations. Wildlife Soc B 42:184–194. https://doi.org/10.1002/wsb.882
Tews J, Brose U, Grimm V, Tielbörger K, Wichmann MC, Schwager M, Jeltsch F (2004) Animal species diversity driven by habitat heterogeneity/diversity: the importance of keystone structures. J Biogeogr 31:79–92. https://doi.org/10.1046/j.0305-0270.2003.00994.x
Tilman D, Balzer C, Hill J, Befort BL (2011) Global food demand and the sustainable intensification of agriculture. Proc Natl Acad Sci USA 108:20260–20264. https://doi.org/10.1073/pnas.1116437108
Uchida K, Ushimaru A (2014) Biodiversity declines due to abandonment and intensification of agricultural lands: Patterns and mechanisms. Ecol Mon 84:637–658. https://doi.org/10.1890/13-2170.1
Vepsäläinen V (2007) Farmland birds and habitat heterogeneity in intensively cultivated boreal agricultural landscapes. Academic dissertation. University of Helsinki. Available at http://hdl.handle.net/10138/22139
Vickery JA, Carter N, Fuller RJ (2002) The potential value of managed cereal field margins as foraging habitats for farmland birds in the UK. Agr Ecosyst Environ 89:41–52. https://doi.org/10.1016/S0167-8809(01)00317-6
Vickery JA, Feber RE, Fuller RJ (2009) Arable field margins managed for biodiversity conservation: a review of food resource provision for farmland birds. Agr Ecosyst Environ 133:1–13. https://doi.org/10.1016/j.agee.2009.05.012
Warner RE (1979) Use of cover by pheasant broods in east-central Illinois. J Wildl Manage 43:334–346. https://doi.org/10.2307/3800342
Warner RE (1984) Effects of changing agriculture on ring-necked pheasant brood movements in Illinois. J Wildl Manage 48:1014–1018. https://doi.org/10.2307/3801459
Wiens J (1989) The Ecology of Bird communities (Cambridge Studies in Ecology), Vol. 2 Processes and variations. Cambridge University Press, Cambridge. https://doi.org/10.1017/CBO9780511608568
Acknowledgements
This work was funded by the Niemi Foundation and the von Frenckell Foundation. Open access funding was provided by the University of Helsinki, including the Helsinki University Central Hospital. We thank Paula Haapanen, Teppo Jauhiainen, Teemu Lamberg, Keijo Taskinen, Seppo Tolvanen, and Juha-Pekka Väänänen, who assisted in the field work. We specially thank Stella Thompson for checking the language.
Funding
Open Access funding provided by University of Helsinki including Helsinki University Central Hospital. This work was supported by the Niemi Foundation and the von Frenckell Foundation. Open access funding was provided by University of Helsinki, including the Helsinki University Central Hospital.
Author information
Authors and Affiliations
Contributions
All authors contributed to the study conception and design. Material preparation, data collection and analysis were performed by HK, KJ, V-MV and JV. The first draft of the manuscript was written by HK and all authors commented on previous versions of the manuscript. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Conflict of interest
The authors have no conflicts of interest to declare that are relevant to the content of this article.
Data availability
We will make data available under request.
Animal welfare
This study involved wild and hand-reared animals that were handled according to Finnish animal welfare legislation including the Law on Animal welfare 4.4.1996/247.
Additional information
Communicated by T. Gottschalk.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Krüger, H., Jaatinen, K., Holopainen, S. et al. Margins matter: the importance of field margins as avian brood-rearing habitat in an intensive agricultural landscape. J Ornithol 164, 101–114 (2023). https://doi.org/10.1007/s10336-022-02014-y
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10336-022-02014-y