
Abstract
Animal welfare concerns are becoming a central issue in wildlife management and conservation. Thus, we investigated stress response of wild ungulates to potentially traumatic situations (shooting injuries, vehicle collisions, entanglement, injuries or diseases) and hunting methods (stalking, battues and hunts with dogs) by means of serum cortisol concentrations from blood collected from killed animals. Cortisol levels in roe deer ranged below and in wild boar above levels for moose, red deer and fallow deer (hence, pooled as a group “deer”). Apart from species, cortisol concentration in trauma situations was mainly explained by trauma type and presence of disturbance after the trauma event. Effect of trauma type differed significantly for “deer”, with animals caught in fences and suffering vehicle collisions experiencing higher cortisol levels than animals injured by shooting. Differences between hunting methods were observed in the cervids (“deer” and roe deer), with stalking leading to lower cortisol levels than hunts with dogs (both groups) and battues (roe deer). Events both before and after the shot, such as duration of pursuit prior to shooting, location of injury, trauma length and presence of disturbance after the shot were relevant for cortisol levels in hunted cervids. Our results indicate that search teams tracking and euthanising wounded animals should behave in a calm way to minimise disturbance. Still, it is important to acknowledge that many situations described in the literature, i.e. reindeer handling, roe deer captures and red deer yarding, seem even more stressful, beside vehicle collisions, than most hunting methods.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Animal welfare concerns are becoming increasingly important in most contemporary societies (Fraser 2001, Keeling 2005, Lawrence 2008) and apply to many, if not all aspects of wildlife management and conservation (Paquet and Darimont 2010, McMahon et al. 2012). Over the last decades, they also have challenged the justification of old established capture, trapping and hunting methods (Proulx and Barrett 1989, the United Kingdom‘s Hunting Act 2004 c. 37, Littin and Mellor 2005, King et al. 2005). Thus, traditional activities involving wildlife whose legitimacy cannot be scientifically proven are expected to become increasingly subject of opinion-based conflicts.
Solid and rational scientific studies on the measurable and quantifiable effects of different problematic situations involving wildlife are needed (Weilnböck 2013). Studies measuring the impact of human-directed activities on ungulates on a physiological base are, so far, mostly limited to livestock (e.g. Mitchell et al. 1988, Dalin et al. 1992, Bristow and Holmes 2007) or captive animals (e.g. Abeyesinghe et al. 1997, Carragher et al. 1997), to specific situations (e.g. DeNicola and Swihart 1997) as live captures in traps (Bergvall et al. 2017) or chemical immobilisation (e.g. Sandegren et al. 1987, Arnemo 1995, Mentaberre et al. 2010). Still, few studies exist on physiological effects of hunting (e.g. Bateson and Bradshaw 1997, Cockram et al. 2011) and similar stressors or traumata (e.g. Hattingh 1988) are restricted to red deer (Cervus elaphus) (Bateson and Bradshaw 1997, Cockram et al. 2011) and antelopes (Hattingh 1988).
The term stress is suggested to be restricted to conditions which are uncontrollable and unpredictable and as a consequence is potentially life threatening and thus has an obvious link to animal welfare (Koolhaas et al. 2011). Cortisol is generally referred to as a classical “stress hormone” (Morton et al. 1995) and has been implemented as the standard stress indicator in animal welfare research (Mormède et al. 2007). Measurably increased cortisol concentrations can be observed from about 3–5 min after the onset of the stressor (Sheriff et al. 2011) and peak levels are reached within 15–30 min (De Kloet et al. 2005). A cortisol-mediated negative feedback loop causes the system to return to baseline activity again after 60–90 min (Sheriff et al. 2011, De Kloet et al. 2005).
Blood has been the primary choice of substance for obtaining glucocorticoid levels in many vertebrates and indicates an immediate snapshot of the focal animal state (Sheriff et al. 2011). Whereas a trauma causes a substantial increase in cortisol concentration, ordinary stress and physical activity induce only moderate augmentation (Sheriff et al. 2011).
In this study, we investigate effects of different hunting methods (stalking, battues and hunting with dogs) and other potentially traumatic situations (e.g. wounding during hunts, vehicle collisions and entanglement) on wild ungulates in Sweden (moose Alces alces, roe deer Capreolus capreolus, red deer, fallow deer Dama dama and wild boar Sus scrofa). This is accomplished by examining serum cortisol concentrations in blood samples collected from killed or euthanised animals with a known history during the last hours in life. We investigate the following questions: (1) Do the study species differ in terms of cortisol levels? Based on species biology and phylogeny, differences between wild boar and the cervid species are expected (Price et al. 2005). (2) Do the cortisol responses vary between stressors – which trauma situation causes the highest, which one the lowest response? We hypothesise that the longer elapsed time between trauma and death and the more severe and harmful the injuries, the higher cortisol response and presumably greater suffering. Disturbance before and after a trauma situation may additionally elevate stress levels (e.g. Rehbinder et al. 1982, Carragher et al. 1997, Creel et al. 2002). (3) Do additional factors such as experience (i.e. age), gender (different adaptations between males and females), ambient temperature (Mormède et al. 2007), physiological status (i.e. season) and time until blood sampling affect cortisol levels? Seasonal and diurnal variations in ungulate cortisol levels have been described with contrasting results (see e.g. Bubenik and Brown 1989, Ingram et al. 1999, Huber et al. 2003 for differing results). However, we hypothesise that ambient temperature does not affect cortisol levels in our study; firstly, because only one value was obtained for each individual (preventing analysis of changes over time) and secondly, because traumata are expected to have an overriding and more important impact on hormonal balance than seasonal changes (Rehbinder 1990, Sheriff et al. 2011). The effect of age and gender seems unclear (see e.g. Huber et al. 2003, Rostal et al. 2012, Küker et al. 2015 for differing results) and no considerable changes in cortisol concentration have been observed up to 60 min postmortem (Wesson et al. 1979, Holst 1996), thus we predict that sampling time does not affect the measured cortisol levels. (4) Do cortisol levels vary between hunting methods? Severe disturbance and more intruding hunting methods (such as drive hunts) are hypothesised to result in higher cortisol concentrations than calmer approaches (e.g. stalking) (Bradshaw and Bateson 2000, Cederlund and Kjellander 1991).
Material and methods
Sample collection and laboratory analysis
Data from dead animals were collected in several hunting districts in southern Sweden over a 14-year period, from 1998 until 2012, by one of the co-authors (BR) together with 10 engaged and interested hunters specialised in tracking and euthanising wounded animals. Blood samples were retrieved as soon as possible after the killing of the animal. In most cases, wounded animals were sampled closer to death than animals culled during regular hunts. For the latter group, it sometimes took 1–2 h after death until the blood sample was taken, wherefore the elapsed time between death and blood sampling was recorded for each individual. In order to obtain comparative values, we consider measurements from “undisturbed animals” (defined as animals not being followed by dogs or humans and shot during normal hunting practice, i.e. animals that died within 5 min without apparent suffering) as reference values for “unharmed” animals as opposed to traumatised ones. A similar approach had been taken by Cockram et al. (2011).
Whole blood was collected in 13 ml 100 × 16 mm polypropylene tubes (Sarstedt, Nümbrecht, Germany). When possible, liquid blood was gained directly after the animal’s death by cutting the jugular vein or by inserting a knife into the chest cavity and cutting large blood vessels in front of the heart. In case sampling occurred during exsanguination, blood samples were usually collected when Vena cava caudalis had been cut off in front of the liver and the blood flowed back into the abdominal cavity. Alternatively, free-flowing or partially coagulated blood was obtained from the heart cavities or from the chest cavity after opening the diaphragm. Blood samples contaminated by rumen, stomach or intestinal contents were discarded. Filled blood tubes were sent to the veterinary clinic at Kolmården Zoo (Kolmården, Sweden). Sera were separated by centrifugation and stored in 2 ml plastic tubes at − 23 °C. Frozen samples were sent to the Department of Anatomy, Physiology and Biochemistry at the Swedish University of Agricultural Sciences (SLU, Uppsala, Sweden) in 2007 and 2011 and analysed for serum cortisol concentrations using a radioimmunoassay (RIA) with Coat-A-Count® (Diagnostic Products Corporation, Los Angeles, California, USA). Forty single samples obtained in autumn 2011 and 2012 were analysed at the Department of Clinical Chemistry, Linköping University Hospital (Linköping, Sweden) using a direct chemiluminescent immunoassay analyser on an ADVIA Centaur (Siemens AG, Munich, Germany). The intra-assay % CV was < 10 and the inter-assay % CV was 6.9 at the 78 nmol/L level with the Coat-A-Count® system. The intra-assay % CV was < 3.6 at the 107 nmol/L level and the inter-assay % CV was 7.2 at the 267 nmol/L level with the ADVIA Centaur system.
Additional information by direct observations about each animal was recorded on prepared forms. General information included date, ambient temperature, sex, estimated age category and presence or absence of disturbance before the trauma and between trauma and death. Disturbance was hereby defined as contact with dogs or people; i.e. preceding trauma, pursuit or unaware scaring of animals, and after trauma, close contact in connection to death. For animals killed during regular hunting, further parameters included hunting method, duration of pursuit before shooting, number of shots and hits, injured organs and time of death (defined as in Cockram et al. 2011 as either the time when the animal collapsed to the ground and showed no further signs of life or, if the animal was found still alive after shooting, as the time when the hunter euthanised the animal). Note that type of hunting dog varies according to target game species—with short-legged chasing dogs used for deer, and long-legged baying elkhounds for moose and wild boar—but consistency is assured as the same dog category is used within one species group. For animals that were found injured and subsequently euthanised, type of traumatic accident, start time and duration of the search for the wounded animal, time of death (defined as dispatch time) and distance from killing site to accident site in meters were registered. Elapsed time from trauma to death and from death to sampling was calculated based on time of death, shooting and sampling recorded for each trauma event. All included variables are summarised in Table 1 .
In total, cortisol concentrations were analysed from blood samples of 406 different animals (Supplement, Table S1). Of these, 238 samples contained complete information about hunting method and were grouped in three different categories based on the most common hunting techniques (Supplement, Table S2).
The invasiveness of the blood-sampling techniques is commonly assumed to bias measured hormone levels, making it difficult or impossible to receive proper baseline values (Sheriff et al. 2011). This limitation did however not apply to our study, with the analysed samples being taken postmortem. The definition of reference cases as internal “baseline values” replaced otherwise necessary reference ranges and allowed for comparison with trauma events.
Statistical analysis
We investigated two separate aspects of cortisol levels by first modelling the effects of trauma situations in one analysis and the effects of different hunting strategies on wildlife in a second analysis, by using two different sets of data. We applied linear models with cortisol concentration (in nmol/L) as the response variable. Initial data exploration (graphically and by Shapiro-Wilk tests) led to the cortisol data being log-transformed in order to obtain normality of errors and in line with Ferre et al. (1998). The variable “time to sampling” was log(x + 1)-transformed to reduce the degree of its right-skewed data distribution. “Type of trauma” [four modalities: shooting injury, vehicle collision, reference, “other”] was included as a fixed variable for the general trauma analysis and correspondingly replaced by “hunting method” [three modalities: stalking, battue, hunt with dog] for the hunting techniques analysis. All included continuous explanatory variables were standardised due to their differences in magnitude.
To investigate question 1, we checked for differences in cortisol levels between species using non-parametric Kruskal-Wallis tests and post-hoc pairwise Mann-Whitney tests with Bonferroni correction to adjust p values for multiple comparisons (Holm 1979) due to variance violating assumptions of equality (assessed graphically and by Bartlett’s test). In order to decrease the amount of levels contributing to the analysis, species were pooled when cortisol concentrations did not differ significantly from each other. In the same way, we tested for differences between trauma types and hunting methods for each species group. A species was included in the following modelling only if any of the following prerequisite criteria were met: (1) differences between reference cases and trauma, or (2) differences between hunting methods.
We applied the same modelling approach to test which stressors (question 2) and additional factors (question 3) that explained the cortisol response in trauma situations and to test how the cortisol response was affected by different hunting methods (question 4). Our approach began with defining a baseline (null) model against which to compare the more complex models, including only species and type of trauma or hunting method. In order to examine the effects of each single variable on cortisol concentration and subsequently limit the number of potential predictors, we performed univariate tests with each potential variable in the baseline model (Zuur et al. 2010). We then created a global model with the potential predictors that were significant in the univariate tests. Using correlation matrices (cut-off value Pearsons’ p < 0.7) and variance inflation factors (VIFs, cut-off value VIF < 2.5), we checked and corrected for collinearity between all continuous explanatory variables. We also examined if including meaningful interactions and polynomial terms improved the models. We then analysed each explanatory variable’s significance (p value) and contribution (assessed by hierarchical partitioning, see Allen (2016)) and consequently excluded non-relevant variables. Having obtained the final combination of variables for each model group, we tried to improve model quality. We checked each model for heteroscedasticity by plotting Q-Q plots and model residuals vs. fitted values. Finally, we performed hierarchical partitioning with each final model in order to report the percentage of explained variation for each variable. We then conducted 10-fold cross validation and leave-one-out cross validation (LOOCV) to determine each model’s predictive accuracy as described in Allen (2016).
In case hierarchical partitioning indicated ungulate species to be a major factor in the analyses, we complemented our approach with single species (species group) models in order to detect potential species-specific differences.
We conducted all statistical analyses using R (version 3.3.1, R Core Team 2016) combined with the user interface RStudio (version 1.0.136, RStudio 2016). Unless specified otherwise, statistical procedures were performed using R‘s internal package stats (R Core Team 2016). For creating plots, we used the package ggplot2 (Wickham and Chang 2016). Variance inflation factors were calculated using the package car (Fox et al. 2016). Hierarchical partitioning was conducted with the hier.part package (Walsh and Mac Nally 2015). Cross validation was implemented with the DAAG package (Maindonald and Braun 2015). Model effects were visualised using the package effects (Fox et al. 2017).
Results
Effects of different trauma situations
There was a clear and significant difference between the different species groups’ cortisol levels (Kruskal-Wallis test, H = 166.25, df = 4, p < 2.2 × 10−16). Roe deer and wild boar differed from each other and from the group of moose, red deer and fallow deer, whereas the latter three did not differ significantly from each other (Table 2 ). Hence, they were pooled, leading to three species groups; wild boar, roe deer and the group “deer” (moose, red deer and fallow deer).
Each species group had significantly lower cortisol levels for reference cases than for trauma situations, justifying the use of the former as reference values (mean concentrations ± SE [nmol/L], reference vs trauma: “deer” 29.89 ± 5.07 vs 104.26 ± 5.98, roe deer 13.42 ± 2.32 vs 64.43 ± 6.17, wild boar 249.58 ± 36.80 vs 775.24 ± 64.66; Mann-Whitney test; “deer”: U = 1086.5, p = 8.651 × 10−6; roe deer: U = 134.5, p = 6.776 × 10−8; wild boar: U = 30, p = 3.262 × 10−3).
Trauma types (excluding reference cases) differed significantly for “deer”, with shooting injuries resulting in lower cortisol levels than vehicle collisions and “other” situations (pairwise Mann-Whitney tests with Bonferroni correction with threshold p value = 0.017; shooting injury vs vehicle collision U = 923.5, p = 1.754 × 10−4; shooting injury vs “other” U = 350, p = 4.786 × 10−4; vehicle collision vs “other” U = 52, p = 0.050). “Other” included injured (n = 4), diseased (n = 2) and entangled (in fences or nets, n = 4) moose and fallow deer. In roe deer, trauma categories did not differ significantly, but vehicle collisions were connected to slightly higher cortisol levels than shooting injuries (shooting injury vs vehicle collision U = 99, p = 0.018). Wild boar showed no difference between shooting injuries and “other” situations.
The global model used as a starting point contained the potential explanatory variables “type of trauma”, “species”, “trauma length”, “pre-stress”, “post-stress” and “time to sampling”. No collinearity between continuous variables was found and no heteroscedasticity was apparent in model checking. Cortisol concentration was best explained by ungulate species (57.13% of total variance excluding interactions explained by that variable, estimated by hierarchical partitioning), type of trauma (13.63%), presence of pre-stress (1.46%) and post-stress (27.79%) disturbance and the interaction between pre- and post-stress (contribution of interactions is not supported by the applied hierarchical partitioning approach). The selected top model (Table 3 , Fig. 1 ) explained 64.1% of the variation in cortisol concentration (adjusted r2) as compared to 54.4% of the baseline model (containing only species and type of trauma) and yielded an overall mean square error (MSE) of 0.729 (10-fold) and 0.731 (LOOCV) in cross validation, respectively.

Effects of the parameters type of trauma, species and the interaction pre-stress*post-stress disturbance included in the selected all-species model predicting cortisol response in different trauma situations
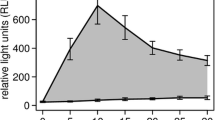
As ungulate species was the most decisive factor in the analysis, we also performed species (group) specific models. For these analyses, two extreme outliers were removed for roe deer (cortisol values > 200 nmol/L). Relevant variables included in the global starting models varied between species groups (Table 4 ) and also cortisol concentration was explained by different factors per species group (Table 5 ). For “deer”, the quadratic term of trauma length was significant, indicating that cortisol concentration increased with trauma length following an approximated logarithmic trajectory (Fig. 2 ). Relative importance of variables based on hierarchical partitioning varied between model groups (Table 6 ). Model parameters and visualised model effects for each selected model can be found in the Supplement (Table S3−S5, Fig. S1−S3).

Relationship between trauma length and cortisol concentration for “deer” up to 24 h of trauma. The curve is fitted using a LOESS function and each data point corresponds to one sample. The shaded area shows the standard error. Cortisol levels increase with trauma length up to around 200 min where the effect evens out, following an approximated logarithmic trajectory
Effects of different hunting techniques
Differences between the hunting methods were found for the “deer” group (Kruskal-Wallis test, H = 7.43, df = 2, p = 0.024) and for roe deer (Kruskal-Wallis test, H = 18.02, df = 2, p = 1.220 × 10−4). For both groups, cortisol levels for stalking differed from those for hunts with dogs and for roe deer also from those for battues (Table 7 ). Neither “deer” nor roe deer showed significant differences between battues and hunts with dogs. Due to wild boar not fulfilling the prerequisites for the analysis (i.e. difference between hunting methods), modelling was restricted to “deer” and roe deer.
The global model used as a starting point consisted of the potential explanatory variables “hunting method”, “species”, “trauma length”, “post-stress”, “injured organs” and “pursuit”. No collinearity between continuous variables or heteroscedasticity was detected. The originally fixed variable “species” became insignificant in the model (p > 0.3) and was therefore removed during model selection. The variables predicting cortisol concentration best were hunting method (36.66% of total variance explained), injured organs (20.48%), length of trauma (15.22%), post-stress disturbance (15.09%) and duration of pursuit (12.55%). The quadratic term of trauma length was significant as in the trauma model for “deer”. The chosen top model (Table 8 , Fig. 3 ) explained 27.6% of the variation in cortisol concentration (adjusted r2) as compared to 12.7% of the baseline model (containing only species and hunting method) and yielded an overall mean square error (MSE) of 0.88 (10-fold) and 0.89 (LOOCV) in cross validation, respectively.

Effects of the parameters hunting method, post-stress disturbance, injured organs, trauma length and duration of pursuit included in the selected model predicting cortisol response with different hunting techniques in cervids
Discussion
In this study, and in relation to the initially asked questions, we show the following: (1) cortisol levels in roe deer were lower and in wild boar higher compared to moose, red deer and fallow deer (the later three pooled as “deer”); (2a) cortisol concentration in trauma situations was mainly affected by species, type of trauma and disturbance after trauma; (2b) predictors for cortisol concentration differed slightly between species groups, with type of trauma and post-stress disturbance being important for all three groups and trauma length relevant only for “deer”; (3) additional factors such as experience, gender and physiological status only played minor roles in influencing cortisol levels in trauma situations; and finally, (4) the cervids (“deer” and roe deer) exhibited differences between hunting methods, with presence of disturbance after the shot, location of injury, trauma length and duration of pursuit being additional relevant parameters influencing cortisol concentration. These general results will now be discussed in depth.
Effects of different trauma situations
Cortisol concentrations in wild boar ranged about 10 times above the levels in the other species, confirming clear physiological differences between the phylogenic cervid and suid groups (Price et al. 2005). Similar observations have been made by Morton et al. (1995) and no clear species differences were found between red and fallow deer by Bubenik and Bartos (1993), strengthening the generality of our result. The reason for the lower levels we found in roe deer is unclear.
As hypothesised, animals in this study that were undisturbed, shot and killed instantly, had lower cortisol levels than animals killed after trauma. Trauma cortisol levels in this study were comparable to capture and transport situations (Mentaberre et al. 2010, Montané et al. 2002) and agreed well with maximum levels determined by Bubenik and Reyes-Toledo (1994), indicating a marked stress responses of particularly roe deer to most trauma situations in our study. Still, there are several other situations described with elevated and much higher cortisol levels than in our study and all relate to human disturbance (see e.g. Rehbinder et al. 1982, Hattingh 1988, Morton et al. 1995, DeNicola and Swihart 1997), as for example stressful immobilisation (Rostal et al. 2012 in moose, see also Lundstein 2013), velvet antler removal in red deer (Cook and Schaefer 2002) and physically restrained individuals (Küker et al. 2015). Little information exists on cortisol levels in wild boar, but our reference and trauma levels lay in the range of a study on anti-parasite-treated wild boar (López-Olvera et al. 2006). Morton et al. (1995) found warthogs (Phacochoerus africanus), the only suids in their comparative study, to have the highest capture stress cortisol levels.
Species identity was by far the most important variable influencing cortisol response in this study, presumably due to the marked differences between wild boar and other ungulates. Post-stress disturbance was the second most important predictor and as expected, disturbance after the trauma increased cortisol levels. Also, pre-stress disturbance, such as pursuit by dogs preceding trauma, had an effect. The interaction between pre- and post-stress disturbance in the selected model, had larger effects on cortisol concentration than the two separate disturbance factors alone, confirming the hypothesis that stress factors may be cumulative (Rehbinder 1990). Type of trauma obviously had a strong influence on the cortisol response. Our findings suggest that vehicle collisions and entanglement, severe diseases and injuries involve more distress than hunting-related injuries. This could be explained by (i) the individual’s pre-trauma condition, with hunted game expected to be mainly in good shape and diseased animals often having already debilitated conditions before sampling, or (ii) stronger stress responses to the course of events itself, possibly related to higher degree of pain experienced in vehicle collisions and the usually close contact to humans following the accident. Length of stress situation, with previously injured or ill animals possibly remaining undetected for a longer period, can be neglected as a reason, as we controlled for trauma length in the analyses.
In the species-group specific models, total explained variance was higher for the general top model, probably due to larger sample sizes and the large amount of variance explained by species identity. Only in wild boar and roe deer did trauma type and post-stress disturbance remain after model selection, underlining the importance of these factors to cortisol response. For “deer”, trauma length was additionally relevant, as well as the interaction between trauma length and post-stress disturbance, further emphasising the importance of post-stress disturbance. The relationship between cortisol concentration and trauma length was characterised by a quadratic term, following an approximated and stabilising logarithmic curve. This suggests that length of trauma matters up to a certain threshold after which only minor changes in cortisol concentration occur. Such ceiling effects due to biological limitations are common in physiology (Veissier and Boissy 2007) and cortisol seems to quickly reach a ceiling (Bradshaw and Bateson 2000).
Effects of the different trauma types also varied between species groups. Cortisol response in “deer” was significantly higher for traffic accidents and entanglements (category “other”) than for shooting injuries, probably for the same reasons mentioned above about the general trauma model. Similar tendencies were apparent for roe deer, although not significant. Wild boar cortisol response did not differ between trauma situations, but was significantly higher under trauma conditions than in reference situations. The lack of difference was likely due to large inter-individual variation in each category or small sample sizes for this species. Our findings may also suggest a higher sensitivity of cervids to varying stressful situations compared to suids (Morton et al. 1995). The variables “ambient temperature”, “time to sampling”, “sex” and “age” were not included in any of the best models. This corroborates our hypothesis that these factors play only minor roles in affecting cortisol levels in trauma situations (Rehbinder 1990).
To conclude, vehicle collisions, diseases, injuries and entanglement (“other”) combined with post-stress disturbance evoke the greatest stress response of all analysed courses of events. Pre-stress disturbance increased cortisol response additionally, but only in combination with post-stress disturbance. Cortisol levels vary strongly between ungulate species and the cortisol response of “deer” was particularly sensitive to prolonged trauma durations. Hunting laws and animal ethics dictate that unnecessary suffering of animals has to be avoided by all means. Post-stress disturbance should hence not only be performed as soon as possible (to shorten suffering) but should also be reduced to a minimum of people involved. Earlier studies on stress in wildlife mainly investigate stress response in total without accounting for different impact factors such as pre- and post-stress disturbance (see e.g. Hattingh 1988). Exceptions are the studies by Rehbinder et al. (1982) and Rehbinder (1990) that found a relationship between extent of handling stress and different physiological parameters in semi-domestic reindeer (Rangifer tarandus) and identified usage of motor vehicles for herding as a crucial stress factor. They therefore also suggested shortening stressful handling procedures and transportation wherever possible.
Effects of different hunting techniques
Our found cortisol levels agree well with levels found in red deer killed after stalking in Great Britain (Bradshaw and Bateson 2000, Cockram et al. 2011). Hunting with hound packs in England leads to higher cortisol release (Bateson and Bradshaw 1997, Bradshaw and Bateson 2000) than the battues and dog hunts of our study, presumably due to the much more intense and disturbing situation involved in hunting with staghounds (Bradshaw and Bateson 2000). As cortisol levels for both cervid groups were similar for battues and hunts with dogs, we suggest that it is of minor importance whether cervids are disturbed by people or pursued by a limited number of dogs, with the disturbance itself being the major impact factor (Bradshaw and Bateson 2000).
As expected, hunting method was the most important variable predicting stress response of cervids to hunting. “Active” hunts with dogs or many people involved (battues) were related to higher cortisol responses than “calmer” stalking. Post-trauma circumstances such as wounding site, disturbance and time until dispatch (i.e. trauma length) also had high importance for cortisol levels in cervids. Contrary to our expectations damages in thorax, bleeding organs and injured abdominal organs caused lower cortisol levels in “deer” than injuries in CNS, skeleton or muscles, because CNS damage and fractures are regarded as less immediate painful than other wounds (Beecher 1946 on pain in wounded soldiers, Bradshaw and Bateson 2001, Gregory 2005). Obviously it is hard to generalise the subjective feeling of pain (Beecher 1946) and sample size is, owing to the preferred hit zone in hunting, skewed towards hits in thorax and bleeding organs (but see Short 1998 and Bateson and Bradshaw 2000).
Our findings suggest largely similar factors affect stress response in hunting and trauma situations, although pre-stress factors appear to weigh higher in hunting situations and post-stress factors higher in trauma situations. This may be explained by the varying ratio of pre-and post-stress extent for the two settings: In hunting, the pre-stress phase (locating and possibly pursuing the animal) mostly lasts longer than the post-stress period (ideally only a few seconds from first shot to death). By contrast, trauma situations usually have short pre-stress periods (abrupt incident creating trauma) followed by longer post-stress phases until the animal eventually is euthanized. To minimise stress level, it should thus, whenever possible, be given preference to calm approaches (such as stalking) compared to more intensive and intruding methods (such as drive hunts with fast and long-legged dogs) (see also Bradshaw and Bateson 2000, Cockram et al. 2011). Shortening the period of pre-trauma disturbance and stress is also beneficial for the game‘s meat quality (see e.g. Rehbinder et al. 1982). Cortisol levels for the cervids in our study killed during battues and hunts with dogs ranged far below those determined for ungulates in other human-dominated situations (e.g. Rehbinder et al. 1982, Carragher et al. 1997, Küker et al. 2015). Furthermore, it has to be kept in mind that wild animals are regularly exposed to natural situations that may trigger cortisol release to yet unknown extent (e.g. predators passing by, unexpected sounds such as falling trees, inter- and intraspecific interactions) and that stress response as such is a vital biological phenomenon present in the daily life of every wild animal. Contrasting natural against anthropogenic stressors will be a challenging, but critical task for future research in order to allow a more sophisticated assessment of traumatic situations.
Conclusions and management implications
With an exceptional dataset we believe our results support that in traumatic situations (such as vehicle collisions and shooting injuries), disturbance between trauma and death influences cortisol concentrations in all analysed species the most. In moose, red deer and fallow deer, hunting and injuries caused by shooting in general seem less harmful than vehicle collisions.
While hunters should be aware that battues and hunts with fast dogs imply higher stress exposure for cervids than calm methods like stalking, cautions should also be taken by search teams when tracking wounded animals, as close encounters with people or dogs after trauma appear to trigger cortisol responses additionally, presumably reflecting the psychological stress response. In future research, we suggest to study the course of events early after trauma onset in detail, as trauma-mediated cortisol increase takes place during the first hour of trauma.
References
Abeyesinghe SM, Goddard PJ, Cockram MS (1997) The behavioural and physiological responses of farmed red deer (Cervus elaphus) penned adjacent to other species. Appl Anim Behav Sci 55(1):163–175. https://doi.org/10.1016/S0168-1591(97)00037-3
Allen A (2016) Linking movement ecology with the Management of Mobile Species. Managing moose in Sweden (paper VI), Faculty of Forest Sciences, Swedish University of Agricultural Sciences, Umeå
Arnemo JM (1995) Immobilization of free-ranging moose (Alces alces) with medetomidine-ketamine and remobilization with atipamezole. Rangifer 15(1):19–25. https://doi.org/10.7557/2.15.1.1153
Bateson P, Bradshaw EL (1997) Physiological effects of hunting red deer (Cervus elaphus). P Roy Soc Lond B Biol 264(1389):1707–1714. https://doi.org/10.1098/rspb.1997.0237
Bateson P, Bradshaw EL (2000) The effects of wound site and blood collection method on biochemical measures obtained from wild, free-ranging red deer (Cervus elaphus) shot by rifle. J Zool 252(3):285–292. https://doi.org/10.1111/j.1469-7998.2000.tb00623.x
Beecher HK (1946) Pain in men wounded in battle. Ann Surg 123(1):6–105
Bergvall AU, Jäderberg L, Kjellander P (2017) The use of box-traps for wild roe deer: behaviour, injuries and recaptures. Eur J Wildl Res 63:67
Bradshaw EL, Bateson P (2000) Welfare implications of culling red deer (Cervus elaphus). Anim Welf 9(1):3–24
Bradshaw EL, Bateson P (2001) Deer hunting and welfare—reply. Anim Welf 10(1):115–117
Bristow DJ, Holmes DS (2007) Cortisol levels and anxiety-related behaviors in cattle. Physiol Behav 90(4):626–628. https://doi.org/10.1016/j.physbeh.2006.11.015
Bubenik GA, Brown RD (1989) Seasonal levels of cortisol, triiodothyronine and thyroxine in male axis deer. Comp Biochem Physiol Phys A 92(4):449–503
Bubenik GA, Bartos L (1993) Cortisol levels in red deer (Cervus elaphus) and fallow deer (Dama dama) after an acute ACTH administration. Can J Zool 71(11):2258–2261. https://doi.org/10.1139/z93-317
Bubenik GA, Reyes-Toledo E (1994) Plasma levels of cortisol, testosterone and growth hormone in pudu (Pudu puda Molina) after ACTH administration. Comp Biochem Physiol 107(3):523–527. https://doi.org/10.1016/0300-9629(94)90035-3
Carragher JF, Ingram JR, Matthews LR (1997) Effects of yarding and handling procedures on stress responses of red deer stags (Cervus elaphus). Appl Anim Behav Sci 51(1):143–158. https://doi.org/10.1016/S0168-1591(96)01090-8
Cederlund G, Kjellander P (1991) Effects of chasing by hunting dogs on roe deer. Transactions of the 20th Congress of the International Union of Game Biologists. University of Agricultural Science, Gödöllö, Hungary, 21–26
Cockram MS, Shaw DJ, Milne E, Bryce R, McClean C, Daniels MJ (2011) Comparison of effects of different methods of culling red deer (Cervus elaphus) by shooting on behaviour and post mortem measurements of blood chemistry, muscle glycogen and carcase characteristics. Anim Welf 20(2):211–224
Cook NJ, Schaefer AL (2002) Stress responses of wapiti (Cervus elaphus canadensis) to removal of velvet antler. Can J Anim Sci 82(1):11–17. https://doi.org/10.4141/A01-031
Creel S, Fox JE, Hardy A, Sands J, Garrott B, Peterson RO (2002) Snowmobile activity and glucocorticoid stress responses in wolves and elk. Conserv Biol 16(3):809–814. https://doi.org/10.1046/j.1523-1739.2002.00554.x
Dalin AM, Magnusson U, Häggendal J, Nyberg L (1992) The effect of transport stress on plasma levels of catecholamines, cortisol, corticosteroid-binding globulin, blood cell count, and lymphocyte proliferation in pigs. Acta Vet Scand 34(1):59–68
De Kloet ER, Sibug RM, Helmerhorst FM, Schmidt M (2005) Stress, genes and the mechanism of programming the brain for later life. Neurosci Biobehav Rev 29(2):271–281. https://doi.org/10.1016/j.neubiorev.2004.10.008
DeNicola AJ, Swihart RK (1997) Capture-induced stress in white-tailed deer. Wildlife Soc Bull 25(2):500–503
Ferre I, Goddard PJ, Macdonald AJ, Littlewood CA, Duff EL (1998) Effect of method of blood sample collection on adrenal activity in farmed red deer and sheep following administration of ACTH. Anim Sci 67(1):157–164. https://doi.org/10.1017/S1357729800009899
Fraser D (2001) Farm animal production: changing agriculture in a changing culture. J Appl Anim Welf Sci 4(3):175–190. https://doi.org/10.1207/S15327604JAWS0403_02
Fox J, Weisberg S, Adler D, Bates D, Baud-Bovy G, Ellison S, Firth D, Friendly M, Gorjanc G, Graves S, Heiberger R, Laboissiere R, Monette G, Murdoch D, Nilsson H, Ogle D, Ripley B, Venables W, Winsemius D, Zeileis A (2016) Package ‘car’. Companion to Applied Regression
Fox J, Weisberg S, Friendly M, Hong J, Andersen R, Firth D, Taylor S (2017) Package ‘effects’. Effect displays for linear, generalized linear, and other models
Gregory NG (2005) Bowhunting deer. Anim Welf 14(2):111–116
Hattingh J (1988) Comparative quantitation of the physiological response to acute stress in impala and roan antelope. Comp Biochem Physiol 89(4):547–551. https://doi.org/10.1016/0300-9629(88)90831-6
Holm S (1979) A simple sequentially rejective multiple test procedure. Scand J Stat 6(2):65–70
Holst S (1996) Evaluation of stress in eland antelopes at the Kolmården Zoo. Master thesis, Swedish University of Agricultural Sciences, Report no. 23/1996, Sweden
Huber SR, Palme R, Arnold W (2003) Effects of season, sex, and sample collection on concentrations of fecal cortisol metabolites in red deer (Cervus elaphus). Gen Comp Endocrinol 130(1):48–54
Ingram JR, Crockford JN, Matthews LR (1999) Ultradian, circadian and seasonal rhythms in cortisol secretion and adrenal responsiveness to ACTH and yarding in unrestrained red deer (Cervus elaphus) stags. J Endocrinol 162(2):289–300. https://doi.org/10.1677/joe.0.1620289
Keeling LJ (2005) Healthy and happy: animal welfare as an integral part of sustainable agriculture. Ambio 34(4):316–319. https://doi.org/10.1579/0044-7447-34.4.316
King RJH, Bateson P, Eves HE, Wolf CM, Nelson F, Kerasote T, Yaich SC (2005) The ethics of hunting. Front Ecol Environ 3(7):392–397
Koolhaas JM, Bartolomucci A, Buwalda B, de Boer SF, Flugge G, Korte SM, Meerlo P, Murison R, Olivier B, Palanza P et al (2011) Stress revisited: a critical evaluation of the stress concept. Neurosci Biobehav Rev 35(5):1291–1301. https://doi.org/10.1016/j.neubiorev.2011.02.003
Küker S, Huber N, Evans A, Kjellander P, Bergvall UA, Jones KL, Arnemo JM (2015) Hematology, serum chemistry, and serum protein electrophoresis ranges for free-ranging roe deer (Capreolus capreolus) in Sweden. J Wildl Dis 51(1):269–273. https://doi.org/10.7589/2014-02-025
Lawrence AB (2008) Applied animal behaviour science: past, present and future prospects. Appl Anim Behav Sci 115:1):1–1)24
Littin KE, Mellor DJ (2005) Strategic animal welfare issues: ethical and animal welfare issues arising from the killing of wildlife for disease control and environmental reasons. Rev Sci Tech Oie 24(2):767–782. https://doi.org/10.20506/rst.24.2.1611
López-Olvera JR, Höfle U, Vicente J, Fernández-de-Mera IG, Gortázar C (2006) Effects of parasitic helminths and ivermectin treatment on clinical parameters in the European wild boar (Sus scrofa). Parasitol Res 98(6):582–587. https://doi.org/10.1007/s00436-005-0099-2
Lundstein LG (2013) Can cortisol be used to assess acute stress in moose? Bachelor Thesis, Högskolan I Hedmark, Campus Evenstad, Norway
Maindonald JH, Braun WJ (2015) Package ‘DAAG’. Data Analysis and Graphics Data and Functions
McMahon CR, Harcourt R, Bateson P, Hindell MA (2012) Animal welfare and decision making in wildlife research. Biol Conserv 153:254–256. https://doi.org/10.1016/j.biocon.2012.05.004
Mentaberre G, López-Olvera JR, Casas-Díaz E, Bach-Raich E, Marco I, Lavín S (2010) Use of haloperidol and azaperone for stress control in roe deer (Capreolus capreolus) captured by means of drive-nets. Res Vet Sci 88(3):531–535
Mitchell G, Hattingh J, Ganhao M (1988) Stress in cattle assessed after handling, after transport and after slaughter. Vet Rec 123(8):201–205
Montané J, Marco I, López-Olvera J, Manteca X, Lavín S (2002) Transport stress in roe deer (Capreolus capreolus): effects of a short-acting antipsychotic. Anim Welf 11(4):405–417
Mormède P, Andanson S, Aupérin B, Beerda B, Guémené D, Malmkvist J, Manteca X, Manteuffel G, Prunet P, van Reenen CG, Richard S, Veissier I (2007) Exploration of the hypothalamic–pituitary–adrenal function as a tool to evaluate animal welfare. Physiol Behav 92(3):317–339. https://doi.org/10.1016/j.physbeh.2006.12.003
Morton DJ, Anderson E, Foggin CM, Kock MD, Tiran EP (1995) Plasma cortisol as an indicator of stress due to capture and translocation in wildlife species. Vet Rec 136(3):60–63. https://doi.org/10.1136/vr.136.3.60
Paquet PC, Darimont CT (2010) Wildlife conservation and animal welfare: two sides of the same coin. Anim Welf 19(2):177–190
Price SA, Bininda-Emonds ORP, Gittleman JL (2005) A complete phylogeny of the whales, dolphins and even-toed hoofed mammals (Cetartiodactyla). Biol Rev 80(3):445–473. https://doi.org/10.1017/S1464793105006743
Proulx G, Barrett MW (1989) Animal welfare concerns and wildlife trapping: ethics, standards and commitments. T W Sec Wil 25:1–6
R Core Team (2016) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna. URL http://www.R-project.org
Rehbinder C, Edqvist LE, Lundström K, Villafañe F (1982) A field study of management stress in reindeer (Rangifer tarandus tarandus). Rangifer 2(2):2–21. https://doi.org/10.7557/2.2.2.404
Rehbinder C (1990) Management stress in reindeer. Rangifer 10(3):267–288. https://doi.org/10.7557/2.10.3.868
Rostal MK, Evans AL, Solberg EJ, Arnemo JM (2012) Hematology and serum chemistry reference ranges of free-ranging moose (Alces alces) in Norway. J Wildl Dis 48(3):548–559. https://doi.org/10.7589/0090-3558-48.3.548
RStudio (2016) Integrated development environment for R, Boston, Massachusetts. URL http://www.rstudio.org
Sandegren F, Pettersson L, Ahlqvist P, Röken BO (1987) Immobilization of moose in Sweden. Swedish Wildl Res Suppl 1:785–791
Sheriff MJ, Dantzer B, Delehanty B, Palme R, Boonstra R (2011) Measuring stress in wildlife: techniques for quantifying glucocorticoids. Oecologia 166(4):869–887
Short CE (1998) Fundamentals of pain perception in animals. Appl Anim Behav Sci 59(1):125–133. https://doi.org/10.1016/S0168-1591(98)00127-0
Veissier I, Boissy A (2007) Stress and welfare: two complementary concepts that are intrinsically related to the animal’s point of view. Physiol Behav 92(3):429–433. https://doi.org/10.1016/j.physbeh.2006.11.008
Walsh C, Mac Nally R (2015) Package ‘hier.part’. Hierarchical Partitioning
Weilnböck GEC (2013) Zur Stressbelastung des Rehwilds (Capreolus capreolus) beim Fang mit der Kastenfalle. Dissertation, Ludwig-Maximilians-Universität München
Wesson JA, Scanlon PF, Kirkpatrick RL, Mosby HS (1979) Influence of the time of sampling after death on blood measurements of the white-tailed deer. Can J Zool 57(4):777–780. https://doi.org/10.1139/z79-095
Wickham H, Chang W (2016) Package ‘ggplot2’. Create elegant data visualisations using the grammar of graphics
Zuur AF, Ieno EN, Elphick CS (2010) A protocol for data exploration to avoid common statistical problems. Methods Ecol Evol 1:3–14
Acknowledgements
We thank all the dedicated tracking hunters, primarily Torbjörn Johansson, without whom this study would not have been possible. The assistance of veterinary assistant Karin Magnusson, Kolmården Zoo, and many students with samples and lab work is greatly appreciated. Finally, we thank Luca Corlatti and two anonymous reviewers for helpful comments on a previous version of the manuscript.
Funding
This study was financed by a research grant from Karl-Erik Önnesjö Foundation, Norrköping, and the Swedish Association for Hunting and Wildlife Management (“Forskningstjugan”) to BR.
Author information
Authors and Affiliations
Corresponding author
Ethics declarations
Conflict of interest
The authors declare that they have no conflict of interest.
Ethical statement
As the described animal handling procedures were either conducted in the context of normal hunting actions or as a measure to euthanise accidentally injured animals, no ethical permit for animal experiments applies or has to be approved. All applicable international, and/or institutional guidelines for the care and use of animals were followed. This article does not contain any studies with human participants performed by any of the authors.
Electronic supplementary material
ESM 1
(DOCX 3729 kb).
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Gentsch, R.P., Kjellander, P. & Röken, B.O. Cortisol response of wild ungulates to trauma situations: hunting is not necessarily the worst stressor. Eur J Wildl Res 64, 11 (2018). https://doi.org/10.1007/s10344-018-1171-4
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10344-018-1171-4