
Abstract
Human disturbance has been shown to provoke physiological and behavioral responses in birds, so nature-based tourism might reduce bird abundance and diversity. The negative consequences of human disturbance might be expected to be maximized during eventual massive events in highly protected areas such as national parks. In this study, the consequences for soundscapes of human presence and disturbance of thousands of visitors during an ornithological fair (massive event) on the bird community of the Monfragüe National Park (Spain) were analyzed. We found that the number and diversity of bird vocalizations did not decrease during the massive event. In contrast, the presence of people in the Monfragüe National Park was associated with an increase in the number and diversity of vocalizations. The effect of human presence on the number of calls and songs differed: the number of calls mainly increased during the massive event when people were present, while the number of songs increased when people were present, particularly during the measurement campaign without the massive event. The human shield hypothesis, along with other behavioral and environmental factors, might potentially explain the results obtained.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Humans may be perceived by animals as predators (Beale and Monaghan 2004; Frid and Dill 2002; Meisingset et al. 2022). Human presence can have direct impacts on wildlife by altering the behavior, physiology, and reproduction of animals (French et al. 2017; Knapp et al. 2013; Ohashi et al. 2013; Sibbald et al. 2011; Stillman and Goss-Custard 2002; van der Mescht et al. 2021; Williams et al. 2006). These effects can be observed in prey species but also in common predators (Lovell et al. 2022), which, in turn, can determine the presence and activity of prey species.
Impacts of human presence have been studied in birds. Human disturbance can alter heart rates and stress hormone levels or increase vigilance or flight (Beale and Monaghan 2004; Blumstein 2014; Caro 2005; Formenti et al. 2015; Fernández-Juricic and Tellería 2000; Møller 2008, 2010, 2012; Price 2008; Stankowich and Blumstein 2005; Weimerskirch et al. 2002). The consequences of human disturbance on birds’ physiology and behavior might drive to a decrease in survival or reproduction and the abandonment of the area (Anderson and Keith 1980; Arlettaz et al. 2015; Bötsch et al. 2017; Tablado and Jenni 2017; Tarjuelo et al. 2015). Accordingly, human disturbance has been shown to reduce bird density and species richness (Arlettaz et al. 2015; Bötsch et al. 2017, 2018; Kangas et al. 2010; Pfister et al. 1992; Steven and Castley 2013; van der Zande et al. 1984).
Nature-based tourism involves human disturbance and, therefore, has the potential to cause considerable effects on bird communities (Bötsch et al. 2018; Kangas et al. 2010; Kerbiriou et al. 2009; Rösner et al. 2014; Steven and Castley 2013). On the other hand, nature-based tourism can involve the well-being of local people and favors conservation (Das and Hussain 2016; Mayer 2014; Robalino and Villalobos 2015; Stronza et al. 2019). Research is necessary to ensure the sustainability of this human activity, mainly in protected areas.
National parks are widely known, highly protected areas around the world (Dudley 2008). In these areas, human activities which might be expected to affect nature conservation should be considered and monitored. Impacts of tourism on wildlife conservation in national parks have been found to be positive (conservation support of local people, funds for conservation, increase the number of wild animals) or negative (introduce invasive species, increase conflicts between human and wild animals; see review in Rhama 2020). In these protected areas, the relevance of human disturbance on bird communities can be maximized during eventual touristic massive events. Because of their special protection regimes, this type of event in national parks is expected to be rare. When massive human events are conducted in national parks, the impacts on wildlife deserve research.
Monfragüe National Park is a Spanish protected area where tourism benefits local communities (e.g., Sánchez et al. 2020). However, human presence might have negative consequences on animal species such as birds. Additionally, a massive human event occurs annually in Monfragüe National Park that might have a deep impact on wildlife: the International Ornithological Tourism Fair (FIO according to its Spanish name). During this event, there is a significant increase in the number of people throughout the park. In addition, the FIO fair is held in winter, when thermoregulatory energy demands increase (Stillman and Goss-Custard 2002; Wiersma and Piersma 1994), and therefore the impact on species such as birds is likely to be severe. In view of this, the aim of this study was to analyze the consequences of human presence and the massive disturbance of visitors during the FIO fair on the bird community in Monfragüe National Park.
Birds might avoid the areas in Monfragüe National Park where human presence increases. Additionally, the abandonment of the area by birds might be expected during the FIO fair due to the massive presence of people. The impact of human disturbance on bird communities can be evaluated by monitoring bird numbers and distribution through transect or point count surveys conducted by human observers. The drawbacks of these survey methods, such as observer biases or limited detectability of certain species, have prompted the application of alternative techniques, including acoustic methods (Abrahams and Geary 2020; Zwart et al. 2014). Therefore, the soundscape was the variable we analyzed to infer the impact on bird community.
Sound has been used for decades to survey birds and obtain population census information (Ralph and Scott 1981; Robbins and van Welzen 1969) and is nowadays recorded to monitor biodiversity trends (Sueur et al. 2019). However, in addition to the study of animal sound (bioacoustics), there is growing interest in the relationship of natural and anthropogenic sounds to the state of ecological systems with different levels of protection and human presence (ecoacoustics; Sueur and Farina 2015; Xie et al. 2020). Therefore, ecoacoustics broadens the scope of acoustic research to include bioacoustics and soundscape ecology. There are two aspects of sound included in ecoacoustics that are related to ecosystem management: response indicator (estimating the diversity of vocal organisms) and stress indicator (examining the effects of human activity; Farina and Gage 2017). Both aspects are related to the objective of the present study.
Regarding the quantification of biodiversity from sound recordings, two methodologies are commonly used. Listening to sound recordings for species identification is one of them. Previous studies show a high correlation between bird species identified by listening to recordings and in field observations (Haselmayer and Quinn 2000; Joo 2009). Nowadays there are also web applications based on artificial neural network for bird recognition (BirdNET). Bioacoustic indices are also used to measure biodiversity (Sueur et al. 2014; Zhao et al. 2019). These indices obtained from environmental recordings depend on the characteristics of the sounds in the recording, i.e., the diversity, complexity, degree of uniformity, or spectral composition of the sounds (Farina and Gage 2017). Moreover, the number and richness of bird vocalizations have been used to determine the presence and abundance of bird species, mainly in forest areas (Chen and Maher 2006; Pérez-González et al. 2021). Therefore, we hypothesize that human presence and the FIO fair (massive event) reduce bioacoustic indices of biodiversity in Monfragüe National Park. We compared diversity measurements related to bird vocalizations in situations with low and high human disturbance.
Despite bird abundance and biodiversity can be quantified using sound recordings, it is important to consider the behavioral aspects of the bird soundscape. Birds utilize vocalizations (songs and calls) as communicative signals to transmit information (Catchpole and Slater 2008). Songs and calls differ in terms of complexity, length, and the context of vocalization. Songs are more complex and longer than calls. Both types of vocalizations also serve different functions and are motivated by distinct stimuli. While songs are typically associated with territorial defense, courtship, and mating, calls are generally linked to alarm functions and social cohesion (Catchpole and Slater 2008). Therefore, we predict that human presence might have different effects on each type of vocalization. In response to perceiving human presence as a risk, birds might choose to fly away (Frid and Dill 2002), resulting in a decrease in the frequency of songs. Alternatively, if birds choose to remain in the presence of humans, the frequency of defensive behaviors and alarm calls might increase (Knight and Temple 1986; Leavesley and Magrath 2005). In order to gain further insights into the effect of human presence on bird behavior, we also investigated whether human presence differentially affects the frequency of bird songs and calls.
Material and methods
Monfragüe National Park and International Ornithological Tourism Fair
The Monfragüe National Park is located in the Spanish region of Extremadura (39° 49′ 12″ N, 5° 58′ 12″ W). Monfragüe has 18,396 ha and it is considered one of the biggest and best preserved areas of Mediterranean forest in Europe. Within the national park, there are patches of highly conserved forest and areas with different levels of human intervention such as dehesas. The Mediterranean forest is composed of cork and holm oak trees (Quercus suber, Q. ilex) and several scrub species such as strawberry tree (Arbutus unedo), laurustinus (Viburnum tinus), myrtle (Mirtus communis), false olive (Phillyrea angustifolia), heaths (Erica spp.), or rockroses (Cystus spp.). The dehesas are open grassland areas with scattered cork oaks and holm oaks. They were created by transforming the Mediterranean forest to favor livestock. In the park, there are two main rivers (Tajo and Tiétar) surrounded by quarzitic mountain ranges with large rocks and cliffs. These habitats are important natural refuges for the nesting of raptors, vultures, or storks. Most of these species can be easily observed from conditioned lookouts dispersed throughout the park. In addition to the flagship species that can be easily observed, the existence of an abundant and diverse bird community makes Monfragüe National Park an important area for bird conservation and the development of ornithological tourism in Europe. The estimated number of visitors was around 67,000 per year (Junta de Extremadura 2019; data 2016–2019). Therefore, around 183 visitors per day can be estimated.
Within Monfragüe National Park, managers offer three routes for visitors: green, red, and yellow (Figs. 1 and S1). The three routes start in the only village within the national park, Villarreal de San Carlos (Fig. S1). The green route (7.9 km) is a trail that connects the riparian forest of the Malvecino stream with the Gimio hill. The red route (16 km) is a trail throughout the most conserved Mediterranean forest in the park. The yellow route (8.9 km) is a road throughout the Tajo and Tiétar rivers. Along the routes, there are several observation points and lookouts where people congregate to observe birds and to enjoy the landscape. The starting point for all routes is Villarreal de San Carlos (Fig. S1).

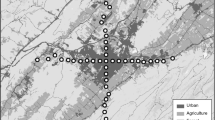
Sampling points in Monfragüe National Park (Google Earth). The figure shows the sampling points in the red route (A), in the yellow route (B), and in the green route (C). Routes are drawn with colored lines. The color of the points represents the mean of the total number of vocalizations for both measurement campaigns (see also Fig. S1). The UTM coordinates of some of the sampling points are as follows: Point 1: 29 S, 753751 m E, 4413075 m N; Point 2: 29 S, 755892 m E, 4415116 m N; Point 3: 29S, 753269 m E, 4415410 m N
The consideration of Monfragüe as one of the most important areas to ornithological tourism in Europe is also due to the hold of the FIO fair. The last FIO fair that was carried out previously to the Spanish confinement for the COVID-19 pandemic (February 2020) received 16,900 visitors. Around 480 professional encounters took place and more than 55 companies related to ornithology attended the event. More than 100 activities and conferences for visitors were conducted. During the FIO fair, the number of vehicles on the roads of Monfragüe increases significantly. Professional encounters and activities took place in Villarreal de San Carlos. However, most people use the attendance at the FIO fair to visit other areas, lookouts, or routes in the national park.
Data collection
Sampling points (N = 18) were located in the three routes of Monfragüe National Park (6 in the green route, 8 in the red route, and 4 in the yellow route; Fig. 1). Locations with sonorous and landscaped interest, as well as signposted lookouts, were included as sampling points. The minimum distance between the two sampling points was 90.5 m. At these sampling points, a portable Zoom H6 recorder with Roland Binaural microphones was used to record bird sounds for 5 min. Bird sounds were recorded in two measurement campaigns: 15 days before the International Ornithological Tourism Fair (February 13th, 2020) and during the fair (February 28th, 2020). Therefore, data were collected in two conditions: “no event” (15 days before the FIO fair) and “event” (during the FIO fair). Both measurement campaigns began at the same time during the morning and the surveys along the sampling points were conducted in the same order (red route from the west to the east, green route from the south to the north, and yellow route from the west to the east). Both measurement campaigns finished at a similar time during the afternoon. We recorded the order of data collection at the sampling points within each route (time variable) to account for the potential effect of the time of day on our results. Weather conditions on the days of data collection were similar (temperature ranged between 14 °C and 15 °C, sun and clouds, wind speed ≤ 7 km/h). For each sampling point, we registered the presence of people in the area during data collection. Due to the importance of water on habitat features and the presence of wildlife in Mediterranean ecosystems (Olea et al. 2005), we registered whether there were water resources less than 50 me apart from each sampling point. This distance was arbitrary, but variations of the value did not change the obtained results (not shown).
The free software Sonic Visualiser (https://www.sonicvisualiser.org/) and Audacity (https://www.audacityteam.org) were used to perform the analyses and management of recordings. All the vocalizations (songs and calls, see below) in the recordings were analyzed in order to identify the bird species that emitted the sound. Species identification was conducted mainly aurally by the authors’ experience, although the process was supported by the visual inspection of spectrograms. We used the BirdNET web page (https://birdnet.cornell.edu/) to verify identifications and to manage doubts. A small proportion of calls could not be identified as belonging to a specific species (see "Results" section). We used Birds of the World from The Cornell Lab of Ornithology (https://birdsoftheworld.org) as reference for scientific names.
In those sampling points with high sound levels proceeding from other environmental sound sources (motors of vehicles, or people talking loudly), sounds were filtered out by their frequencies to clearly identify bird vocalizations.
Different types of vocalizations were identified in passerine birds. In these species, vocalizations were categorized as calls or sounds. Songs were considered complex longer vocalizations for which the spectrogram shows different segments with different frequencies. We referred calls as shorter and simpler vocalizations for which the spectrogram shows a continuous segment or spot.
From each recording, the number of recorded vocalizations produced by each bird species was counted. Calls and short songs (less than 1 s) were counted as discrete vocalizations. Longer songs were registered as a number of vocalizations equal to the number of seconds that the song lasted (see Pérez-González et al. 2021). Unidentified vocalizations were included as vocalizations of an additional species. The number of vocalizations was used to obtain three diversity measures for each recording: species richness (number of species that emitted at least one vocalization), and the Shannon and Simpson indices. These three measurements were used to assess the impact of the massive event on the bioacoustic diversity in the National Park. Additionally, calls and songs were analyzed separately to obtain a more comprehensive understanding of the potential effect of human presence on bird behavior.
Statistical analyses
We firstly assessed the effect of human presence on the general soundscape of the Monfragüe National Park. The effects of the massive event and the presence of people on acoustic diversity measures were assessed by conducting three models, each with a different response variable: species richness (number of bird species), and the Shannon and Simpson indices which take into account both the number of species and the number of vocalizations per species. Note that these diversity measures estimate the diversity of vocalizations rather than the biodiversity of species. Therefore, for each sampling point, a single value was obtained for each of the three diversity measures. In all three models, the measurement campaign (no event vs. event), the presence of people (sampling point with and without people), and the interaction of both were included as a fixed factor, and time within route and sampling point within route were incorporated as nested random effects. Water resource (sampling point with or without water resources) and route were initially included as fixed factors. However, the models were simplified by conducting a background stepwise procedure in which the model with the lowest AIC value was selected. Water resource and route did not appear in the simplified models. For the species richness, we conducted a generalized linear mixed effect model (GLMMs) fitted by maximum likelihood using the Poisson distribution. For the Shannon and Simpson indices, linear mixed effect models (LMMs) fitted by maximum likelihood were performed.
On the other hand, we categorized bird vocalizations into calls and songs to explore further insights into the potential factors influencing the relationship between human presence and the number of vocalizations. For that, the effects of the massive event and the presence of people on the number of calls and songs were assessed by conducting two GLMMs for zero-inflated count data fitted by maximum likelihood using the Poisson distribution. The model was repeated separately for calls and songs. In both models, the number of vocalizations (calls or songs) produced by each bird species was included as the response variable. The fixed factors included in the models were people (sampling point with or without people), measurement campaign (no event vs. event), and the interaction of both. The bird species was considered a random factor, and time within the route and sampling point within the route were incorporated as nested random effects. Additionally, the water resource variable was incorporated into the model as a fixed factor. As the interactions between people and measure campaign were significant in both models (with calls and songs as response variables; see "Results" section), we repeated the models by changing the reference for the measure campaign factor.
Biodiversity measures were obtained with a vegan package (Oksanen et al. 2020) in R software (R Core Team 2019). LMMs were performed with the nlme package (Pinheiro et al. 2020) in R software. GLMMs for the total number of vocalizations and the species richness were conducted with lme4 package (Bates et al. 2015) in R software. GLMMs for zero-inflated count data with a single zero-inflation parameter applying to all observations were performed with the glmmTMB package (Brooks et al. 2017) in R software. Predict effect plots conducted with the effects package (Fox 2003) in R were used to observe the effect of the factors’ interaction on zero-inflated count data.
Results
All recordings registered 4375 bird vocalizations. The mean number of recorded vocalizations per minute was 24.306. We identified 33 bird species and 7.406% of the vocalizations were not identified. Table 1 shows the number of vocalizations for each bird species.
We met people observing wildlife in 4 lookouts used as sampling points (2 in the red route and 2 in the yellow route). In all these lookouts, there were more than 5 people in both measurement campaigns. However, the number of observed people was very different between both measurement campaigns. We counted 5 and 8 people in the lookouts of the red route and 5 and 6 people in the lookouts of the yellow route during the measurement campaign of “no event.” However, we counted 37 and 41 people in the lookouts of the red route, and 26 and 22 people in the lookouts of the yellow route during the FIO fair (see Table S1 in Supplementary Material for additional information that illustrates human disturbance in both measurement campaigns). For all three measures (species richness, and the Shannon and Simpson indices), the values of acoustic biodiversity were higher during the massive event compared to the measurement campaign with no event (Table 2, Fig. 2). The measurement campaign (“no event” vs. “event”) was the only factor that reached statistical significance for these measures of acoustic biodiversity.

Acoustic biodiversity measures for both measurement campaigns (“no event” vs. “event”) and the people presence (sampling point without and with people). A–C graphs show results for each diversity measure (see Table 2)
After categorizing vocalizations into calls and songs, the number of vocalizations varied depending on the measurement campaign and the presence of people in the sampling point (Table 3, Fig. 3). Furthermore, the interaction between both fixed factors had a significant effect in both models, but this effect differed for calls and songs (Table 3, Fig. 3). The number of calls was higher during the massive event, particularly when people were present at the sampling point (Table 3A and right panel in Fig. 3A). When “no event” was employed as the reference for the Event factor, the effect of People did not reach statistical significance (estimate = 0.596, standard error = 0.433, z = 1.377, p = 0.168; left panel in Fig. 3A). The number of songs was also higher during the massive event, regardless of the presence of people. However, we also recorded a high number of songs during the measurement campaign of “no event” when people were present (Table 3B and left panel in Fig. 3B). When “event” was used as reference for the Event factor, the effect of People did not reach significance (estimate = 0.601, standard error = 0.604, z = 0.995, p = 0.320; right panel in Fig. 3B). The water resource factor was also positively related to both the number of calls and the number of songs (Table 3).

Effect of the interaction between measurement campaign (event vs. no event) and the presence of people on the number of calls (A) and songs (B). The figure shows predicted values and 95% confidence limits
Discussion
Bird soundscape was analyzed in two very different situations regarding human disturbance. During the measurement campaign of “no event,” there were few people in the national park. The highest number of people we met on this day was 20 (see Table S1). However, during the FIO fair, there were more than a thousand people in the national park. Most of them were in Villarreal de San Carlos, but there were people throughout the park. The number of people in the lookouts was not very different between measurement campaigns due to their limited space, which prevented the gathering of a large number of people. However, human presence and human activities differed between both measurement campaigns.
Human disturbance was expected to reduce the presence of birds in Monfragüe National Park, at least in popular lookouts or during events with high people influx (Bötsch et al. 2017, 2018; Coppes et al. 2017; Rösner et al. 2014). However, we did not find a negative relationship between the presence of people and the bioacoustic indices of biodiversity. Contrarily, the diversity of bird vocalizations increased during the International Ornithological Tourism Fair. Furthermore, the number of calls and songs was positively related to the human presence. These results can have important implications on bird conservation in protected areas where touristic activities are implemented.
Non-assessed variables might induce a spurious relationship between human presence and bioacoustic indices. For instance, high habitat quality in lookouts might be responsible for the positive association between human presence and bioacoustic indices of biodiversity. Accordingly, we found a positive effect of the existence of water resources on the number of calls and songs. Food availability or the presence of refuges might also impact the spatial distribution of bird vocalizations. However, differences in habitat quality do not explain why the frequency of bird vocalizations was higher during the massive event. The significant associations observed in both the spatial (People term in Table 3) and temporal (Event term in Tables 2 and 3) dimensions of human presence in our study support the notion that the relationship between human presence and the number and diversity of vocalizations might not be spurious.
On the other hand, the massive event occurred 2 weeks later than the measurement campaign of “no event.” This temporal delay makes the FIO fair closer to the reproductive period of Iberian birds and more frequent vocalizations related to mating might be expected. Songs are mainly associated with mating behaviors, while calls serve various functions (contact, alarm, mating; Catchpole and Slater 2008), so a prediction of this possible explanation would be that the increase in the number of vocalizations during the massive event is primarily driven by a higher number of songs. Accordingly, the intensity of the measurement campaign effect was greater for songs compared to calls (Table 3 and Fig. 3). However, the higher number of songs in the presence of people, particularly during the measurement campaign of “no event,” suggests that the temporal delay was not the sole variable associated with the increase in the number of vocalizations.
The decrease in bird presence and vocalizations, as a means to avoid detection, was expected since birds can perceive humans as potential predators (Beale and Monaghan 2004; Frid and Dill 2002). The absence of a negative effect of human presence on the number and diversity of bird vocalizations may be attributed to a habituation process taking place in the bird community of Monfragüe National Park (see e.g. Villanueva et al. 2012). The intense reiterative benign interactions with humans can induce bird habituation (Blumstein 2014; Fowler 1999; Webb and Blumstein 2005). This type of interaction is frequent in cities where birds show high levels of habituation to humans (Blumstein 2014; Møller 2012; Vincze et al. 2016).
Habituation can explain why human presence does not seem to have a negative impact on bird community. However, why do the number and diversity of vocalizations increase in the presence of people? Firstly, we assume that the increase in the number of vocalizations should not be due to an increase in bird abundance or biodiversity. Contrarily, it might be attributed to behavioral changes resulting from factors associated with human presence. The following possible explanations are based on this assumption.
As a possible explanation, human activities might produce environmental background noise and, as a consequence, birds might emit higher sound levels favoring sound transmission and the detection from higher distances (Brumm 2004; Brumm and Slabbekoorn 2005). Alternatively, humans might increase resource availability by food provisioning (consciously or unconsciously) and, hence, increase interactions between birds or change bird distribution and abundance (Geffroy et al. 2015; Knight 2009). Moreover, the higher number of vocalizations might also be explained by the increase in the emitting of alarm calls in the presence of people (Catchpole and Slater 2008; Frid and Dill 2002). According to these possible explanations, we found a higher frequency of calls mainly during the massive event in sampling points with the presence of people, when high human presence might motivate birds to communicate diverse information (contact, alarm, mating).
Finally, the reiterative benign interactions with humans might induce a phenomenon previously detected in urban environments: the human shield (Geffroy et al. 2015; Møller 2012; Valcarcel and Fernández-Juricic 2009). The human shield hypothesis explains the increase in the abundance of certain bird species in cities because of a decrease in native predators. Therefore, the human presence may create a safe situation that prevents predation to birds. In Monfragüe National Park, the recorded vocalizations were mostly emitted by bird species that can be predated by raptors such as sparrowhawk (Accipiter nisus), peregrine falcon (Falco peregrinus), kites (Milvus spp.), goshawk (Accipiter gentilis), or booted eagle (Hieraaetus pennatus) (García et al. 1998; García-Dios 2006; Hernández 2018; Mañosa 1994; Rizolli et al. 2005). The presence of people in Monfragüe National Park might act as a human shield that decreases the risk of being attacked by a predator. The decrease in the predation risk can favor the emission of bird vocalizations to transmit information (Catchpole and Slater 2008). But also, the abundance and diversity of the bird community might increase in those areas with human presence that act as attractive safe zones (Valcarcel and Fernández-Juricic 2009). According to the human shield hypothesis, the frequency of songs was maximized when people were present during both measurement campaigns. Despite the human shield hypothesis can explain the obtained results, we cannot rule out other explanations and probably the outputs were due to the combination of several factors and processes.
In summary, we found that the number and diversity of vocalizations increased in the presence of people. Therefore, we might conclude that the human impact was positive on the bird community of Monfragüe National Park. However, according to our sampling procedure, we were unable to provide evidence of the negative impacts of human presence on the bird community. These impacts might be related to increased energetic demand and food provisioning and reduced reaction to predators or negative effects on predator populations (Carrascal et al. 2012; Geffroy et al. 2015; Orams 2002). Future studies about the effect of human presence on the entire bird community, including raptors or rates of food provisioning, can help to design strategies to make compatible nature-based tourism and conservation in highly visited national parks.
Data availability
The data have been included as Electronic Supplementary Material.
References
Abrahams C, Geary M (2020) Combining bioacoustics and occupancy modelling for improved monitoring of rare breeding bird populations. Ecol Indic 112:106131. https://doi.org/10.1016/j.ecolind.2020.106131
Anderson DW, Keith JO (1980) The human influence on seabird nesting success: conservation implications. Biol Conserv 18:65–80. https://doi.org/10.1016/0006-3207(80)90067-1
Arlettaz R, Nusslé S, Baltic M, Vogel P, Palme R, Jenni-Eiermann S, Patthey P, Genoud M (2015) Disturbance of wildlife by outdoor winter recreation: allostatic stress response and altered activity-energy budgets. Ecol Appl 25:1197–1212. https://doi.org/10.1890/14-1141.1
Bates D, Mächler M, Bolker B, Walker S (2015) Fitting linear mixed-effects models using lme4. J Stat Softw 67:1–48
Beale CM, Monaghan P (2004) Human disturbance: people as predation-free predators? J Appl Ecol 41:335–343. https://doi.org/10.1111/j.0021-8901.2004.00900.x
Blumstein DT (2014) Attention, habituation, and antipredator behaviour: implications for urban birds. In: Gil D, Brumm H (eds) Avian urban ecology. Oxford University Press, UK, Oxford, pp 41–53. https://doi.org/10.1093/acprof:osobl/9780199661572.003.0004
Bötsch Y, Tablado Z, Jenni L (2017) Experimental evidence of human recreational disturbance effects on bird-territory establishment. Proc Biol Sci 284:20170846. https://doi.org/10.1098/rspb.2017.0846
Bötsch Y, Tablado Z, Scherl D, Kéry M, Graf RF, Jenni L (2018) Effect of recreational trails on forest birds: human presence matters. Front Ecol Evol 6:175. https://doi.org/10.3389/fevo.2018.00175
Brooks ME, Kristensen K, van Benthem KJ, Magnusson A, Berg CW, Nielsen A, Skaug HJ, Maechler M, Bolker BM (2017) glmmTMB balances speed and flexibility among packages for zero-inflated generalized linear mixed modeling. R J 9:378–400. https://journal.r-project.org/archive/2017/RJ-2017-066/index.html
Brumm H (2004) The impact of environmental noise on song amplitude in a territorial bird. J Anim Ecol 73:434–440. https://doi.org/10.1111/j.0021-8790.2004.00814.x
Brumm H, Slabbekoorn H (2005) Acoustic communication in noise. Adv Stud Behav 35:151–209. https://doi.org/10.1016/S0065-3454(05)35004-2
Catchpole CK, Slater PJB (2008) Bird song: biological themes and variations, 2nd edn. Cambridge Univ. Press, Cambridge
Caro T (2005) Antipredator defenses in birds and mammals. University of Chicago Press, Chicago
Carrascal LM, Villén-Pérez S, Seoane J (2012) Thermal, food and vegetation effects on winter bird species richness of Mediterranean oakwoods. Ecol Res 27:293–302. https://doi.org/10.1007/s11284-011-0900-x
Chen Z, Maher RC (2006) Semi-automatic classification of bird vocalizations using spectral peak tracks. J Acoust Soc Am 120:2974–2984. https://doi.org/10.1121/1.2345831
Coppes J, Ehrlacher J, Thiel D, Suchant R, Braunisch V (2017) Outdoor recreation causes effective habitat reduction in Capercaillie Tetrao urogallus: a major threat for geographically restricted populations. J Avian Biol 48:1–28. https://doi.org/10.1111/jav.01239
Das D, Hussain I (2016) Does ecotourism affect economic welfare? Evidence from Kaziranga National Park, India. J Ecotourism 15:241–260. https://doi.org/10.1080/14724049.2016.1192180
Dudley N (2008) Guidelines for applying protected area management categories. International Union for the Conservation of Nature (IUCN), Gland, Switzerland
Farina A, Gage SH (2017) Ecoacoustics: a new science. In: Farina A, Gage SH (eds) Ecoacoustics. https://doi.org/10.1002/9781119230724.ch1
Fernández-Juricic E, Telleria JL (2000) Effects of human disturbance on spatial and temporal feeding patterns of blackbird Turdus merula in urban parks in Madrid, Spain. Bird Study 47:13–21. https://doi.org/10.1080/00063650009461156
Formenti N, Vigano R, Bionda R, Ferrari N, Trogu T, Lanfranchi P, Palme R (2015) Increased hormonal stress reactions induced in an alpine black grouse (Tetrao tetrix) population by winter sports. J Ornithol 156:317–321. https://doi.org/10.1007/s10336-014-1103-3
Fowler GS (1999) Behavioral and hormonal responses of Magellanic penguins (Spheniscus magellanicus) to tourism and nest site visitation. Biol Conserv 90:143–149. https://doi.org/10.1016/S0006-3207(99)00026-9
Fox J (2003) Effect displays in R for generalised linear models. J Stat Softw 8:15:1–27. https://doi.org/10.18637/jss.v008.i15
French SS, Neuman-Lee LA, Terletzky PA, Kiriazis NM, Taylor EN, DeNardo DF (2017) Too much of a good thing? Human disturbance linked to ecotourism has a “dose-dependent” impact on innate immunity and oxidative stress in marine iguanas, Amblyrhynchus cristatus. Biol Conserv 210:37–47. https://doi.org/10.1016/j.biocon.2017.04.006
Frid A, Dill LM (2002) Human-caused disturbance stimuli as a form of predation risk. Conserv Ecol 6:11. https://www.ecologyandsociety.org/vol6/iss1/art11/
García JT, Viñuela J, Sunyer C (1998) Geographic variation of the winter diet of the Red Kite Milvus milvus in the Iberian Peninsula. Ibis 140:302–309. https://doi.org/10.1111/j.1474-919X.1998.tb04393.x
García-Dios IS (2006) Dieta del aguililla calzada en el sur de Ávila: importancia de los paseriformes. Ardeola 53:39–54. https://doi.org/10.1111/j.1474-919X.1998.tb04393.x
Geffroy B, Samia DSM, Bessa E, Blumstein DT (2015) How nature-based tourism might increase prey vulnerability to predators. Trends Ecol Evol 30:755–765. https://doi.org/10.1016/j.tree.2015.09.010
Haselmayer J, Quinn JS (2000) A comparison of point counts and sound recording as bird survey methods in Amazonian southeast Peru. Condor 102:887–893. https://doi.org/10.2307/1370317
Hernández A (2018) Diet of Eurasian Sparrowhawks in a northwest Iberian hedgerow habitat throughout the year. Ornithol Sci 17:95–101. https://doi.org/10.2326/osj.17.95
Joo W (2009) Environmental acoustics as an ecological variable to understand the dynamics of ecosystems, PhD dissertation. Michigan State University, East Lansing
Junta de Extremadura (2019) Visitantes del parque nacional de Monfragüe. Datos de 2007 a 2019.
Kangas K, Luoto M, Ihantola A, Tomppo E, Siikamäki P (2010) Recreation induced changes in boreal bird communities in protected areas. Ecol Appl 20:1775–1786. https://doi.org/10.1890/09-0399.1
Kerbiriou C, Le Viol E, Robert A, Porcher E, Gourmelon F, Julliard R (2009) Tourism in protected areas can threaten wild populations: from individual response to population viability of the chough Pyrrhocorax pyrrhocorax. J Appl Ecol 46:657–665. https://doi.org/10.1111/j.1365-2664.2009.01646.x
Knapp CR, Hines KH, Zachariah TT, Perez-Heydrich C, Iverson JB, Buckner SD, Halach SC, Lattin CR, Romero LM (2013) Physiological effects of tourism and associated food provisioning in an endangered iguana. Conserv Physiolo 1:cot032. https://doi.org/10.1093/conphys/cot032
Knight J (2009) Making wildlife viewable: habituation and attraction. Soc Anim 17:167–184. https://doi.org/10.1163/156853009x418091
Knight RL, Temple SA (1986) Why does intensity of avian nest defense increase during the nesting cycle? Auk 103:318–327. https://doi.org/10.1093/auk/103.2.318
Leavesley AJ, Magrath RD (2005) Communicating about danger: urgency alarm calling in a bird. Anim Behav 703:65–373. https://doi.org/10.1016/j.anbehav.2004.10.017
Lovell C, Li S, Turner J, Carbone C (2022) The effect of habitat and human disturbance on the spatiotemporal activity of two urban carnivores: the results of an intensive camera trap study. Ecol Evol 12:e8746. https://doi.org/10.1002/ece3.8746
Mañosa S (1994) Goshawk diet in a Mediterranean area of Northeastern Spain. J Raptor Res 28:84–92
Mayer M (2014) Can nature-based tourism benefits compensate for the costs of national parks? A study of the Bavarian Forest National Park, Germany. J Sustain Tour 22:561–583. https://doi.org/10.1080/09669582.2013.871020
Meisingset EL, Gusevik J, Skjørestad A, Brekkum Ø, Mysterud A, Rosell F (2022) Impacts of human disturbance on flight response and habitat use of red deer. Ecosphere 13:e4281. https://doi.org/10.1002/ecs2.4281
Møller AP (2008) Flight distance of urban birds, predation, and selection for urban life. Behav Ecol Sociobiol 63:63–75. https://doi.org/10.1007/s00265-008-0636-y
Møller AP (2010) Interspecific variation in fear responses predicts urbanization in birds. Behav Ecol 21:365–371. https://doi.org/10.1093/beheco/arp199
Møller AP (2012) Urban areas as refuges from predators and flight distance of prey. Behav Ecol 23:1030–1035. https://doi.org/10.1093/beheco/ars067
Ohashi H, Saito M, Horie R, Tsunoda H, Noba H, Ishii H, Kuwabara T, Hiroshige Y, Koike S, Hoshino Y, Toda H, Kaji K (2013) Differences in the activity pattern of the wild boar Sus scrofa related to human disturbance. Eur J Wildlife Res 59:167–177. https://doi.org/10.1007/s10344-012-0661-z
Oksanen J, Blanchet FG, Friendly M, Kindt R, Legendre P, McGlinn D, Minchin PR, O’Hara RB, Simpson GL, Solymos P, Stevens MHH, Szoecs E, Wagner H (2020) Vegan: community ecology package, R package version 2.5-7. https://CRAN.R-project.org/package=vegan
Olea L, López-Bellido RJ, Poblaciones MJ (2005) Europe types of silvopastoral systems in the Mediterranean area: dehesa. In: Mosquera MR, Rigueiro A, McAdam J (eds) Silvopastoralism and sustainable land management. CABI, Wallingford, Oxfordshire, United Kingdom, pp 30–35. https://doi.org/10.1079/9781845930011.0030
Orams MB (2002) Feeding wildlife as a tourism attraction: a review of issues and impacts. Tourism Manag 23:281–293. https://doi.org/10.1016/S0261-5177(01)00080-2
Pérez-González J, Rey Gozalo G, Montes González D, Hidalgo de Trucios SJ, Barrigón Morillas JM (2021) Are quartzite scree slopes used by birds to promote sound transmission in the Mediterranean forest? Anim Biodiv Conserv 44:175–194. https://doi.org/10.32800/abc.2021.44.0175
Pinheiro J, Bates D, DebRoy S, Sarkar D (2020) nlme: linear and nonlinear mixed effects models. R package version 3.1-148. https://CRAN.R-project.org/package=nlme
Pfister C, Harrington BA, Lavine M (1992) The impact of human disturbance on shorebirds at a migration staging area. Biol Conserv 60:115–126. https://doi.org/10.1016/0006-3207(92)91162-L
Price M (2008) The impact of human disturbance on birds: a selective review. In: Lunney D, Munn A, Meikle W (eds) Too close for comfort: conflicts in human wildlife encounters. Royal Zoological Society of NSW, Sydney, pp 163–196. https://doi.org/10.7882/FS.2008.023
Ralph JC, Scott JM (1981) Estimating numbers of terrestrial birds. Studies in Avian Biology No. 6. Cooper Ornithological Society, Allen Press, Inc., Lawrence, Kansas
R Core Team (2019) R: a language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Rhama B (2020) The meta-analysis of ecotourism in national parks. Afr J Hosp Tour Leis 9:1–17
Rizolli F, Sergio F, Marchesi L, Pedrini P (2005) Density, productivity, diet and population status of the peregrine falcon Falco peregrinus in the Italian Alps. Bird Study 52:188–192. https://doi.org/10.1080/00063650509461390
Robalino J, Villalobos L (2015) Protected areas and economic welfare: an impact evaluation of national parks on local workers’ wages in Costa Rica. Environ Dev Econ 20:283–310. https://doi.org/10.1017/S1355770X14000461
Robbins CS, van Welzen WT (1969) The breeding bird survey, 1967 and 1968. Special scientific report–wildlife. US fish and wildlife service, Washington, DC
Rösner S, Mussard-Forster E, Lorenc T, Müller J (2014) Recreation shapes a “landscape of fear” for a threatened forest bird species in Central Europe. Landscape Ecol 29:55–66. https://doi.org/10.1007/s10980-013-9964-z
Sánchez JM, Rengifo JI, Sánchez M (2020) Protected areas as a center of attraction for visits from world heritage cities: Extremadura (Spain). Land 9:47. https://doi.org/10.3390/land9020047
Sueur J, Farina A (2015) Ecoacoustics: the ecological investigation and interpretation of environmental sound. Biosemiotics 8:493–502. https://doi.org/10.1007/s12304-015-9248-x
Sueur J, Krause B, Farina A (2019) Climate change is breaking earth beat. Trends Ecol Evol 34:971–973. https://doi.org/10.1016/j.tree.2019.07.014
Sibbald AM, Hooper RJ, McLeod JE, Gordon IJ (2011) Responses of red deer (Cervus elaphus) to regular disturbance by hill walkers. Eur J Wildlife Res 57:817–825. https://doi.org/10.1007/s10344-011-0493-2
Stankowich T, Blumstein DT (2005) Fear in animals: a meta-analysis and review of risk assessment. Proc Biol Sci 272:2627–2634. https://doi.org/10.1098/rspb.2005.3251
Steven R, Castley JG (2013) Tourism as a threat to critically endangered and endangered birds: global patterns and trends in conservation hotspots. Biodivers Conserv 22:1063–1082. https://doi.org/10.1007/s10531-013-0470-z
Stillman RA, Goss-Custard JD (2002) Seasonal changes in the response of oystercatchers Haematopus ostralegus to human disturbance. J Avian Biol 33:358–365. https://doi.org/10.1034/j.1600-048X.2002.02925.x
Stronza AL, Hunt CA, Fitzgerald LA (2019) Ecotourism for conservation? Annu Rev Env Resour 44:229–253. https://doi.org/10.1146/annurev-environ-101718-033046
Sueur J, Farina A, Gasc A, Pieretti N, Pavoine S (2014) Acoustic indices for biodiversity assessment and landscape investigation. Acta Acust United Ac 100:772–781. https://doi.org/10.3813/AAA.918757
Tablado Z, Jenni L (2017) Determinants of uncertainty in wildlife responses to human disturbance. Biol Rev 92:216–233. https://doi.org/10.1111/brv.12224
Tarjuelo R, Barja I, Morales MB, Traba J, Benítez-López A, Casas F, Arroyo B, Delgado MP, Mougeot F (2015) Effects of human activity on physiological and behavioral responses of an endangered steppe bird. Behav Ecol 26:828–838. https://doi.org/10.1093/beheco/arv016
Valcarcel A, Fernández-Juricic E (2009) Antipredator strategies of house finches: are urban habitats safe spots from predators even when humans are around? Behav Ecol Sociobiol 63:673–685. https://doi.org/10.1007/s00265-008-0701-6
van der Mescht AC, Pryke JS, Gaigher R, Samways MJ (2021) Ecological and acoustic responses of bush crickets to anthropogenic and natural ecotones. Biodivers Conserv 30:3859–3878. https://doi.org/10.1007/s10531-021-02278-5
van der Zande AN, Berkhuizen JC, van Latesteijn HC, ter Keurs WJ, Poppelaars AJ (1984) Impact of outdoor recreation on the density of a number of breeding bird species in woods adjacent to urban residential areas. Biol Conserv 30:1–39. https://doi.org/10.1016/0006-3207(84)90018-1
Villanueva C, Walker BG, Bertellotti M (2012) A matter of history: effects of tourism on physiology, behaviour and breeding parameters in Magellanic Penguins (Spheniscus magellanicus) at two colonies in Argentina. J Ornithol 153:219–228. https://doi.org/10.1007/s10336-011-0730-1
Vincze E, Papp S, Preiszner B, Seress G, Bókony V, Liker A (2016) Habituation to human disturbance is faster in urban than rural house sparrows. Behav Ecol 27:1304–1313. https://doi.org/10.1093/beheco/arw047
Webb NV, Blumstein DT (2005) Variation in human disturbance differentially affects predation risk assessment in Western Gulls. Condor 107:178–181. https://doi.org/10.1093/condor/107.1.178
Weimerskirch H, Schaffer SA, Mabiklle G, Martin J, Boutard O, Rouanet JL (2002) Heart rate and energy expenditure of incubating wandering albatrosses: basal levels, natural variation, and the effects of human disturbance. J Exp Biol 205:475–483. https://doi.org/10.1242/jeb.205.4.475
Wiersma P, Piersma T (1994) Effects of microhabitat, flocking, climate and migratory goal on energy expenditure in the annual cycle of red knots. Condor 96:257–279. https://doi.org/10.2307/1369313
Williams R, Lusseau D, Hammond PS (2006) Estimating relative energetic costs of human disturbance to killer whales (Orcinus orca). Biol Conserv 133:301–311. https://doi.org/10.1016/j.biocon.2006.06.010
Xie J, Hu K, Zhu M, Guo Y (2020) Data-driven analysis of global research trends in bioacoustics and ecoacoustics from 1991 to 2018. Ecol Inform 57:101068. https://doi.org/10.1016/j.ecoinf.2020.101068
Zhao Z, Xu Z-Y, Bellisario K, Zeng R-W, Li N, Zhou W-Y, Pijanowski BC (2019) How well do acoustic indices measure biodiversity? Computational experiments to determine effect of sound unit shape, vocalization intensity, and frequency of vocalization occurrence on performance of acoustic indices. Ecol Indic 107:105588. https://doi.org/10.1016/j.ecolind.2019.105588
Zwart MC, Baker A, McGowan PJ, Whittingham MJ (2014) The use of automated bioacoustic recorders to replace human wildlife surveys: an example using nightjars. PLoS One 9:e102770. https://doi.org/10.1371/journal.pone.0102770
Acknowledgements
The authors would like to thank Marcelo D’Amico and two anonymous reviewers for their comments on the manuscript. We also thank the Laboratorio de Acústica–Lambda (https://lambdauex.wordpress.com) for their technical and scientific support and to the managers of the Monfragüe National Park for the use of their facilities and the reception of the project, especially its Director Mr. Ángel Rodríguez Martín.
Funding
Open Access funding provided thanks to the CRUE-CSIC agreement with Springer Nature. This study was supported by the Diputación de Cáceres under grant AV–6 and was also the result of collaboration with the R&D&I project PID2020-114199RA-I00, funded by MCIN/AEI/10.13039/501100011033. G. Rey-Gozalo was supported by the Consejería de Economía, Ciencia y Agenda Digital of Junta de Extremadura (Spain) through grants for attracting and returning research talent to R&D&I centers belonging to the Extremadura Science, Technology and Innovation System (TA18019).
Author information
Authors and Affiliations
Contributions
Javier Pérez-González: methodology, validation, formal analysis, investigation, data curation, writing—original draft, writing—review and editing, visualization, supervision. Guillermo Rey-Gozalo: validation, formal analysis, resources, data curation, writing—review and editing, visualization, supervision, project administration, funding acquisition. Sebastián J. Hidalgo-de-Trucios: validation, investigation, writing—review and editing, visualization, supervision.
Corresponding author
Ethics declarations
Competing interest
The authors declare no competing interests.
Additional information
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Pérez-González, J., Rey-Gozalo, G. & Hidalgo-de-Trucios, S.J. Human presence is positively related to the number of bird calls and songs: Assessment in a national park. Eur J Wildl Res 70, 20 (2024). https://doi.org/10.1007/s10344-024-01772-9
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10344-024-01772-9