
Abstract
Governments are under increasing pressure to reduce greenhouse gas emissions, and large-scale wind farms are being developed in marine environments worldwide. However, top predators are strongly affected by environmental change and anthropogenic activities. Common guillemots (Uria aalge, hereafter guillemots), as one of the world’s most numerous seabird species, are prone to interference with offshore wind farms (OWFs). This study assessed the cumulative impacts of all operating OWFs on guillemots in the German North Sea. These estimates were applied to quantify the possible conflicts between guillemot occurrence and current German government plans to implement large-scale OWFs. If OWFs were implemented according to the current maritime spatial plan for the German Exclusive Economic zone, they would cover 13% of the German North Sea. Guillemot numbers peak during autumn, with German North Sea offshore waters hosting approximately 90,000 individuals. Guillemot density in autumn was significantly reduced within a radius of 19.5 km around operating OWFs. Applying this disturbance distance to current installation plans, about 70% of the German North Sea would be affected, and an estimated 68% of guillemots in the German North Sea would experience habitat loss. This highlights the possible threat to guillemots in the southern North Sea if the current German government plans are implemented. The current estimates are highly relevant to decisions regarding marine spatial planning and management recommendations. Such evaluations are essential for developing sustainable scenarios including reducing the human CO2 footprint, whilst also conserving biodiversity.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Rapid progress in climate change (IPCC 2018, 2021) has increased the commitment of governments to reduce greenhouse gas emissions (UNFCCC 2015a, b; European Commission 2019, 2021). In addition, the current international political situation has highlighted the need for western countries to become independent of foreign mineral oil and gas supplies (The Federal German Government 2023). Renewable energy resources are thus of increasing importance to allow for a change in energy supply on a global scale. Large-scale offshore wind farms (OWFs) are thus being planned and constructed worldwide (4COffshoreWind 2023; WindEurope 2022). To cover the expected electricity demands, the EU strategy on offshore renewable energy targets an installed capacity of 300 GW of offshore wind by 2050 (European Commission 2020). A report commissioned by the wind energy industry expects 85% of the OWF capacity to be developed in northern Europe (i.e., 380 GW), with 212 GW potentially implemented in the North Sea (WindEurope 2019). In the recently signed Esbjerg Declaration, Denmark, Belgium, The Netherlands, and Germany agreed to develop the North Sea as a European Green Power Plant, with the objective of delivering > 50% of the energy needed for the EU to achieve climate neutrality. Installed offshore wind capacities of ≥ 65 GW by 2030 and at least 150 GW by 2050 are needed to reach this target (The Federal German Government 2022). Under increasing pressure to address the climate crisis, the German target for OWFs in its Exclusive Economic Zone (EEZ) was also increased from 15 GW by 2030 (BSH 2017) to ≥ 30 GW by 2030, ≥ 40 GW by 2035, and ≥ 70 GW by 2045 (The Federal German Government 2021; Deutscher Bundestag 2022).
However, offshore waters provide essential habitats for top predators, such as seabirds and marine mammals (Schreiber and Burger 2001; Wilson and Mittermeier 2014), resulting in potential conflicts with the planned large-scale development of offshore renewable energy. Top predators are already strongly affected by environmental changes, e.g., through climate change and changes at lower trophic levels (Croxall et al. 1999; Descamps et al. 2017; Dias et al. 2019), as well as by anthropogenic activities (Frederiksen et al. 2004; Dias et al. 2019). Top predators are thus important indicators of the state of our ecosystems (Parsons et al. 2008; Sergio et al. 2006; Velarde et al. 2019), and play a central role in the health of the marine environment (Duffy 2003; Estes et al. 2011; Wilmers et al. 2012). On a global scale, climate change is currently one of the greatest threats to seabirds (Dias et al. 2019); however, the expansion of renewable energy production has strong impacts on seabirds and adds to existing threats (Dias et al. 2019; Garthe et al. 2023). The implementation of OWFs fundamentally changes the ecological characteristics of the construction areas (e.g., de Mesel et al. 2015; Vandendriessche et al. 2015; Daewel et al. 2022), leading to substantial decreases in habitat availability for various species (Dierschke et al. 2016; Garthe et al. 2023). Reactions to OWFs vary between avoidance and attraction (Perrow 2019; Dierschke et al. 2016), depending on the species, area, and season, potentially leading to increased energy expenditure (Masden et al. 2010) and/or mortality of individuals (Drewitt and Langston 2006).
Anthropogenic activities in the marine habitat have increased greatly, both on a global scale (Halpern et al. 2015, 2019) and in the North Sea (Emeis et al. 2015). The North Sea is already among the most intensively utilised sea areas worldwide, with impacts from activities such as fishing, transport, oil and gas exploitation, and gravel extraction (Halpern et al. 2008, 2019; Emeis et al. 2015). Many top predators, including seabirds, marine mammals, and fish, are affected by the various human activities in the southern North Sea, e.g., through reduced availability of foraging habitats (Garthe et al. 2023; Peschko et al. 2020a), noise emissions during OWF construction (Dähne et al. 2013; Hastie et al. 2015; Russel et al. 2016), shipping activities (Jones et al. 2017; Fliessbach et al. 2019), or habitat changes after OWF installation (de Mesel et al. 2015; Vandendriessche et al. 2015).
Common guillemots (Uria aalge, hereafter guillemot) are distributed widely in the northern Hemisphere (BirdLife International 2021) and is among the most abundant seabird species in the world, and in the North Sea (Skov et al. 1995). Its high abundance means that guillemots are a key seabird species in the southern North Sea. However, the species has previously been shown to have strong reactions to OWFs (see overview by Dierschke et al. 2016; Vanermen et al. 2015; Peschko et al. 2020a, b; Mercker et al. 2021a). Guillemots are listed as being of least concern in the IUCN Red List for the EU member states and for European waters (regional level) (BirdLife International 2021). However, the number of individuals overwintering in German waters decreased by > 75%, representing the strongest decline of all wintering waterbird species in Germany over the past 12 years (Gerlach et al. 2019). The offshore waters of the German North Sea are highly important for guillemots breeding in Europe (Camphuysen and Leopold 1994; Harris et al. 2015; Buckingham et al. 2023). Their abundance and widespread occurrence in the North Sea, their strong but not complete avoidance of OWFs, as well as the existing broad and detailed knowledge of their biology and ecology (e.g., Dunn et al 2020) mean that guillemots are an ideal model species for studying the effects of OWFs on the marine ecosystem.
Using a comprehensive long-term dataset of the distribution and abundance of guillemots, this study aimed to quantify the cumulative effects of all OWFs currently operating in the German North Sea on guillemot occurrence during two sensitive seasons, including autumn, when the highest number of guillemots occurs in the study area. The study also aimed to quantify the conflict between guillemot occurrence in German offshore areas and the German government’s current plans to implement large-scale OWFs, and to provide data to inform relevant national and international management plans and decisions regarding the consequences of large-scale construction plans on an important indicator species for the marine ecosystem. Finally, the study aimed to provide an example of the application of knowledge about species distribution, abundance, and species-specific OWF avoidance to provide essential information for future planning of offshore renewable energy installations.
Methods
Study area
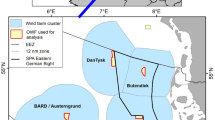
The study focused on the German EEZ in the North Sea (Fig. 1), where 22 wind farms with a power of approximately 7 GW were operating as of December 2022 (Fig. 1, BSH 2022a). The German government plans to construct offshore wind farms in the German EEZ with capacities of ≥ 30 GW by 2030, ≥ 40 GW by 2035, and ≥ 70 GW by 2045 (The Federal German Government 2021; Deutscher Bundestag 2022). The maritime spatial plan for the German EEZ defined priority areas for wind energy to ensure the implementation of this increased expansion target of the federal government for 2030 (BSH 2021b, priority areas, Fig. 1), with additional areas reserved to secure sites for further expansion (BSH 2021b, reservation areas, Fig. 1). Plans for OWF implementation in the areas defined by the maritime spatial plan are currently under development but have not been finalised. The current preliminary draft site-development plan indicates that an area greater than the combined priority and reserved areas included in the maritime spatial plan is needed to implement 70 GW of offshore wind power in the German EEZ (BSH 2023b). This study refers to the priority and reserved areas defined by the maritime spatial plan (BSH 2021b), as shown in Fig. 1, but does not focus on the OWF capacity or the year of the planned implementation, which is subject to change.

Location of the study area in the southern North Sea (inserted map) and in the German Bight, indicating locations of the planned OWF areas in the German EEZ based on the maritime spatial plan (dark grey areas = priority areas for OWF implementation by 2030, shaded areas = reserved for OWF implementation by 2035/2040 (BSH 2021a, b); planned OWF areas are visualised according to BSH (2021b)). Red lines = areas with OWFs operating or under construction, dashed red lines = areas with approved OWFs
Datasets used for analysis of effects of operating OWFs
Data on guillemot occurrence were obtained from aerial and ship-based surveys between 2003 and 2020. The records originate from several seabird monitoring and research projects conducted by the University of Kiel (e.g., the German Marine Biodiversity Monitoring for seabirds on behalf of the Federal Agency for Nature Conservation, Borkenhagen et al. 2019), from monitoring during the construction and operation of offshore wind farms in the German EEZ and from environmental impact studies. The data were collected using ship-based surveys, observer-based aerial surveys, or digital aerial surveys. Details of the field methods are given in Supplementary file 1. The datasets of the different sources were reviewed, harmonised and stored in a combined database (for details of the database, data sources, field methods, and sea states selected for analysis, see Garthe et al. 2015, 2023; Peschko et al. 2020a). Because the effects of OWFs on guillemots during spring and the breeding season are well known (Peschko et al. 2020a, b), the current study was restricted to periods for which there is currently no information on the effects of OWFs on guillemots: autumn (the time between the breeding season and winter; 16. July–30. September, species-specific classification slightly modified from Garthe et al. 2007) and winter (01. October–29. February, Garthe et al. 2007). Autumn is also the season with peak guillemot numbers in the German North Sea. The final data set comprised a total of 26,307 observations of guillemots and 90,264 km2 of survey effort. Following the method recently presented by Garthe et al. (2023), the data were assigned to five wind farm clusters based on the minimum distance to the next OWF (after construction) (see also Supplementary file 2 and below). Further information on the data-collection method for this database can be found in Supplementary file 1.
Quantification of wind farm avoidance by guillemots
A ‘before–after control impact’ (BACI) analysis approach was used to estimate the relative change in guillemot density in the area influenced by OWFs and the response range to the OWFs. An impacted area and a control area were defined (areas or rings depending on the nearest distance to the OWF) and relative comparisons of spatial and temporal differences before and after OWF construction were subsequently used to extract the unbiased OWF impact (Smith 2002; Schwarz 2014; Mendel et al. 2019). We quantified the potential effects of currently operating wind farms using a similar approach to Garthe et al. (2023). We describe the main aspects of the modelling approach below. For further details, please see Garthe et al. (2023).
BACI model covariates
BACI analysis was carried out using generalised additive models, as described in several studies investigating the effects of OWFs on seabirds (e.g., Peschko et al. 2020a; Garthe et al. 2023). To evaluate potential OWF effects on the distribution and abundance of guillemots, the binary variable period was introduced and defined for each wind farm cluster separately. The before period encompassed the period before any construction work in the wind farm cluster (see Supplementary file 2), and the after period started after completion of any construction work at the turbines, for each OWF separately. This approach was chosen because operation started independently for each wind farm in a cluster and an increasing number of OWFs started operating successively during the study period. To minimize the effect of possible long-term changes in guillemot abundance and distribution, the maximum duration of the before period was restricted to 6 years (see Supplementary file 2), while the after period included all data available until 2020. The length of the after period and the time between the before and the after periods ranged from < 1 year to several years (data used per cluster and OWF is shown in Supplementary file 2), and changes in overall distribution could thus have occurred in the meantime. However, the BACI approach is relatively robust against such changes, by comparing densities outside vs. inside the OWF between the before and after periods. Hence, the outcome depends on relative changes in OWF-related distributions, and is not influenced by changes in absolute abundance between different periods. The continuous distance to the nearest OWF was described by the variable dist_owf, which measured the distance to an OWF that did not exist in the before period, to evaluate changes in response to this variable in the before vs. after periods. All data for ≤ 35 km from any wind farm were used (see Supplementary file 2), while larger distances were not expected to affect the birds (Mendel et al. 2019; Peschko et al. 2020a; Garthe et al. 2023).
The change in density between ‘inside affected area’ vs. ‘outside affected area’ was analysed by defining the binary variable B_dist_owf (based on dist_owf, Garthe et al. 2023), as the OWF area plus various radii of R km around the OWFs as 'inside OWF', and all distances further from the OWF as 'outside OWF'. Different radii (R = 1, 5, or 10 km) were applied.
The spatial range of disturbance was investigated by defining an impact area in the shape of a 'belt' around the OWFs, with an inner distance radius x and outer distance radius y to the nearest OWF (see “Habitat loss due to operating OWFs”, Supplementary file 2 and Garthe et al. 2023). The area outside the belt was defined as the control area (from outer radius y up to a distance of 35 km), and the area within the belt was not included in the regression. A width of 3 km was chosen for the belt to allow for good spatial resolution of the measured effects, while maintaining a sufficiently large database for each belt. This analysis was increased in a stepwise manner from x = 0 km (comprising the area occupied by the OWFs) to x = 30 km, to determine the distance up to which the BACI effect within the belt was still significant, corresponding to an estimate of the disturbance distance (Garthe et al. 2023).
The spatial distribution patterns in the before vs. after periods were investigated using the above regression methods but excluding dist_owf-related variables from the predictors, to allow for a maximally objective prediction of patterns related to OWF sites. In particular, the dependency on the variables depth, dist_coast, and the 2D spline depending on spatial coordinates were independently estimated for each period and no information about OWF locations was used (Garthe et al. 2023). For distribution maps, the predict-function of the fitted regression model was used by prescribing detection-related covariate values (sea state, method) to provide optimal detectability (approximating the real number without detectability-dependent loss). Notably however, these patterns do not represent temporally homogeneous situations, given that time frames for the before periods differ between OWF clusters and for the after periods between OWFs.
Details of the environmental covariates included in the models, BACI regression model structure, model-validation strategy, and calculation of the distribution maps can be found in Garthe et al. (2023).
All statistical analyses, validation procedures, and visualizations were carried out using R software (R Core Team 2023), with the following packages: ggplot2 (Wickham 2009) for visualizations and plots, MASS (Venables and Ripley 2002), pscl (Zeileis and Kleiber 2008), and mgcv (Wood 2006) for regression analyses, and Distance (Thomas et al. 2010) for distance-sampling-related procedures.
Dataset used for analysis of bird patterns in relation to OWFs scenarios
This dataset included Seabirds at Sea data collected during observer-based ship and aerial surveys in the German North Sea from 1990 to 2016, as part of several seabird monitoring and research projects conducted by the University of Kiel (e.g., the German Marine Biodiversity Monitoring on behalf of the Federal Agency for Nature Conservation). Population estimates (Gerlach et al. 2019) and distribution maps for guillemots in the German North Sea in autumn were based on predictions of guillemot abundance generated by an integrative statistical approach (Mercker et al. 2021b). Details of the field methods and methods used to determine abundance estimates and patterns are provided in Supplementary file 1.
Estimation of potential habitat loss due to OWF scenarios
This analysis focused on the distribution and abundance of guillemots in autumn, when guillemot numbers in the German North Sea are highest. The avoidance reaction quantified for autumn (see “Quantification of wind farm avoidance by guillemots”) was applied to their modelled distribution in the German North Sea (see “Dataset used for analysis of bird patterns in relation to OWFs scenarios”), as detailed below.
To estimate the effect of the OWF installation on guillemot abundance, the geographical extents of three different OWF scenarios in the German EEZ were visualised, based on the maritime spatial plan (Fig. 1, BSH 2021b): (a) present situation = currently operating OWFs (approximately 7 GW); (b) priority areas = priority areas for OWF implementation up to 2030 (approximately 30 GW, BSH 2023a) including the present situation; and (c) priority areas + reserved areas = priority + reserved areas for implementation of offshore wind power up to 2035/2040 (roughly 50 GW, BSH 2021a, b, 2022b). These three scenarios were applied to illustrate the changing magnitudes of effects depending on different OWF development scenarios. The area that would be covered by OWFs was calculated for each scenario (see “Possible habitat loss due to OWF scenarios”). The area affected by the OWFs (‘affected area’) was estimated based on the spatial extent of significant avoidance in autumn (see “Habitat loss due to operating OWFs” and “Possible habitat loss due to OWF scenarios”). The number of guillemots present in the affected area during species-specific autumn (16 July–30 September; Garthe et al. 2007) was extracted, based on their modelled distribution, and this value was set in relation to the total population size of guillemots in the German EEZ and the whole German North Sea, respectively, to estimate the proportion affected (see “Possible habitat loss due to OWF scenarios”). The value of the reduction in guillemot density inside the affected area (obtained from the above BACI analysis) was then applied to assess the number of guillemots actually encountering habitat loss as a consequence of the different OWF installation scenarios.
Results
Distribution patterns before and after OWF installation
High densities of guillemots were found in autumn, in both the before and after periods, in the submerged portion of the glacial Elbe River valley in the south-east of the study region, as well as in the western and north-western parts of the study region (Fig. 2). In winter, guillemots were concentrated in the south before OWF construction, and additionally concentrated in the north-west after OWF construction (Fig. 3). Some areas of future OWF clusters did not include areas with high densities in the before period for either season, but some areas showed medium-to-high densities before construction and very low densities after OWF construction (e.g., autumn & winter: BARD-Cluster, see also Supplementary file 2). After OWF construction, guillemot densities were very low in autumn. Densities after construction were also low in winter in most clusters, except in the southernmost OWF cluster, where medium-to-high guillemot densities were present.

Predicted density of guillemots in all wind farm clusters in autumn (individuals per km2) before (left) and after construction of OWFs (right). Future wind farms are indicated in grey (left) and operating wind farms are indicated in black (right). Time periods (before vs. after) for the different OWF clusters differ such that depicted patterns represent an artificial mosaic of spatially varying time frames

Predicted density of guillemots in all wind farm clusters in winter (individuals per km2) before (left) and after construction of OWFs (right). Future wind farms are indicated in grey (left) and operating wind farms are indicated in black (right). Time periods (before vs. after) for the different OWF clusters differ such that depicted patterns represent an artificial mosaic of spatially varying time frames
Habitat loss due to operating OWFs
Guillemot abundance declined strongly and significantly after, compared with before construction of OWFs, with a reduction of 91% within the OWFs + 1 km radius (95% confidence interval (CI) = 84%–94% reduction) and 76% within the OWFs + 10 km radius (95% CI = 71%–81%) in autumn, and by 67% within the OWFs + 1 km radius (95% CI = 53%–77%) and 50% within the OWFs + 10 km radius (95% CI = 41%–57%) in winter (Table 1; here statistically significant refers to p-values of p < 0.05, which is equivalent to 95% CIs not intersecting with the value zero).
Guillemot density in autumn was significantly affected up to a distance of 18–21 km (mean = 19.5 km) from the border of the OWFs (Fig. 4, mean and 95% CI were below the expected abundance levels based on the ratio during the before period, i.e., blue line in Fig. 4), and guillemot density inside this affected area (OWF + 19.5 km radius) was reduced by 79% (95% CI = 74%–83%; Table 1). In winter, the density was significantly affected up to a distance of 15–18 km (mean = 16.5 km) from the border of the OWFs (Fig. 5), and guillemot density inside this area (OWF + 16.5 km radius) was reduced by 51% (95% CI = 42%–58%; Table 1).

Relative guillemot abundance after OWF construction in autumn in % of expected abundance without disturbance (100%, blue line). Values shown for stepwise analysis of 3-km-wide belts around the OWFs (x-axis). Mean values = black dots; 95% CIs = length of bars

Relative guillemot abundance after OWF construction in winter in % of expected abundance without disturbance (100%, blue line). Values shown for stepwise analysis of 3-km-wide belts around the OWFs (x-axis). Mean values = black dots; 95% CIs = length of bars
Possible habitat loss due to OWF scenarios
If offshore wind power was realised within all the currently planned areas (priority + reserved areas + operating OWFs), OWFs would cover 13% of the German North Sea (German North Sea = German EEZ and territorial sea areas combined) and 18% of the German EEZ (Fig. 6, Table 2). The planned OWF areas show substantial overlap with abundance hotspots for guillemots in the German North Sea in autumn (Fig. 6), when the offshore waters host approximately 90,000 guillemots (Table 3).

Modelled distribution of guillemots in the German North Sea (German EEZ and territorial sea areas combined) in autumn. Priority areas for OWF implementation up to 2030 are shown in grey (including OWFs already in use or under construction), shaded areas are reserved for OWF implementation up to 2035/2040 (BSH 2021a, b); solid red lines = operating OWFs, dashed red lines = approved OWFs
Applying the response radius of 19.5 km to the OWF areas indicates that 70% of the German North Sea and 82% of the German EEZ would be affected by the planned wind farms (Table 2), including a major part of the core areas used by guillemots in autumn in the German North Sea (Fig. 7). In the present scenario, approximately 26,000 (i.e., 28%) guillemots occurring in the German North Sea use the affected area (Table 3), of which approximately 20,000 individuals experience habitat loss, because the density is reduced by 79% up to a radius of 19.5 km around the OWFs. Thus, in the present scenario, approximately 22% of the German guillemot population in the North Sea in autumn experience habitat loss (Table 3). In the scenario including both priority and reserved areas, approximately 77,000 (i.e., 86%) guillemots in the German North Sea use the potentially affected area (Table 3), of which 61,000 individuals would experience habitat loss due to a reduction in density of 79% up to a radius of 19.5 km around the OWFs. Thus, about 68% of the German guillemot population in the North Sea in autumn would experience habitat loss (Table 3).

Modelled distribution of guillemots in the German North Sea during autumn. Priority areas for OWFs up to 2030 are shown in grey (including OWFs already in use or under construction), shaded areas reserved for OWF implementation up to 2035/2040 (BSH 2021a, b); solid red lines = operating OWFs, dashed red lines = approved OWFs. The response radius of 19.5 km around the OWFs is shown in white
Discussion
Based on extensive data regarding the occurrence of guillemots in the German North Sea, this study examined the impact of currently operating OWFs on guillemot abundance in the German North Sea, as well as the possible impact of German plans for further large-scale OWFs in a guillemot hotspot in the southern North Sea in autumn. Large numbers of guillemots are present in the area where OWFs are to be built, making this part of the North Sea an important area for the European guillemot population in autumn (c.f. BirdLife International 2015).
The current results suggest that operating OWFs have a much larger impact on guillemot densities in the German North Sea in autumn and winter than previously reported for other seasons or for the entire yearly cycle (Leopold et al. 2013; Vanermen et al. 2015; Vallejo et al. 2017; Peschko et al. 2020a). Especially in autumn, areas formerly used by guillemots in medium to high numbers were no longer used or were used by fewer guillemots after OWF construction. Although birds tend to be more flexible in their choice of habitat outside the breeding season, the seasons investigated in this study are considered to be especially sensitive for guillemots, encompassing the moulting period in autumn and winter (July to October, Mendel et al. 2008), as well as the period with cold water temperatures and increased energy demand and mortality in winter (Gaston and Jones 1998; Sonntag 2001; Fort et al. 2009, detailed discussion see below).
The following discussion emphasizes the results for autumn, when guillemot numbers in the German North Sea are at their highest; however, the effect in winter is also strong and needs consideration, especially in areas with high numbers of wintering guillemots.
Current plans for future OWFs would affect 70% of the German North Sea for guillemots, leaving limited undisturbed space and alternative foraging areas in German waters. The only breeding colony of guillemots in German waters is very small (mean of 2,685 breeding pairs on Helgoland between 2000–2019; Dierschke et al. 2011, 2020), and most of the 77,000 individuals that use the affected area of the German North Sea in autumn originate from other colonies, e.g., along the North Sea coast (Camphuysen 2002; Harris et al. 2015; Dunn et al. 2020; Buckingham et al. 2023). The current extent of the planned anthropogenic activities would therefore affect guillemots from different parts of the North Sea.
Applying estimates of guillemot OWF avoidance to data on their abundance and distribution in the German North Sea revealed that 68% of guillemots occurring in this area in autumn are likely to experience habitat loss if offshore wind power is implemented in all the currently designated areas.
Substantial habitat loss can have several consequences for guillemots. The area planned to be covered by OWFs not only serves as a foraging area for guillemots in autumn, but is also important as an area where they moult and rear their chicks in autumn. This has implications for their physical condition and behaviour, given that guillemots are flightless while moulting, which restricts their ability to move (Harris and Wanless 1990; Dunn et al. 2020). A high proportion of adult guillemots have to forage for themselves and their offspring (Harris et al. 1991; Gaston and Jones 1998; Camphuysen 2002; Burke et al. 2015), making them especially dependent on productive foraging areas (Dunn et al. 2020) and vulnerable to changes in habitat conditions. Displacement by large-scale OWFs may prevent access to profitable foraging areas in their usual habitat in autumn. The negative effect in autumn could be further exacerbated, given the high energy demand and auk mortality in winter (Sonntag 2001; Fort et al. 2009; Burke and Montevecchi 2018); if guillemots already experience suboptimal foraging conditions in autumn and start the winter season in poor body condition, their winter mortality is likely to increase. In addition, carry-over effects of poor body condition of guillemot chicks (possibly related to high summer temperatures in 2018) were recently found to be the main cause of a mass mortality event in the southern North Sea in winter 2019 (Leopold et al. 2019). Furthermore, an increased number of individuals sharing a substantially reduced area of foraging habitat will lead to higher intraspecific competition.
It can be speculated that the reef effect (Lindeboom et al. 2011) might increase the abundance of possible prey species in the OWF areas, making them more valuable foraging areas for guillemots in the future. However, most studies have found that guillemots avoided OWFs (Vanermen et al. 2015; Welcker and Nehls 2016; Peschko et al. 2020a, b), despite the increased abundance of some prey species in OWFs in general (Van Deurs et al. 2012; Reubens et al. 2013; Stenberg et al. 2015). Even in the breeding season when guillemots are strongly restricted in their foraging range, GPS tracking showed that individuals strictly avoided OWFs located close to their colony on Helgoland (Peschko et al. 2020b). In contrast, no effect of OWFs on guillemots was detected in the Solway Firth (UK, Vallejo et al. 2017). Estimates for the winter season analysed in the current study revealed that guillemots avoided some OWF areas in the south-western part of the German North Sea less than they avoided other areas further north and east; however, the overall effect for all clusters combined still revealed a strong negative reaction. We assume that the current findings represent the reaction of guillemots in the German North Sea well. When interpreting the current findings however, it is important to consider that the reactions of guillemots towards OWFs in areas of future wind farm developments might differ from the presented effects. Further studies are needed to determine if habituation to OWFs might occur over time, and how guillemots will be affected by future OWF developments, especially as larger areas become occupied by OWFs.
When interpreting the current findings, it is necessary to note that the study only considered direct (e.g., visual) negative effects of OWFs on bird densities, while additional dynamic, large-scale rearrangements could also take place, e.g., displacement of birds into distant regions not covered by the present approach. Thus, future studies should also consider possible dynamic large-scale changes in guillemot distributions due to the presence of future OWFs, to generate a more comprehensive picture of the possible future effects on the guillemot population.
Guillemots are also sensitive to ship traffic (Mendel 2012; Fliessbach et al. 2019), which strongly increases in and around OWFs because of maintenance activities at the turbines and transport to and from the OWFs (Burger et al. 2019; Mendel et al. 2019). The negative effects of OWFs thus extend to areas not directly affected by the turbines. In addition to OWF-related ship traffic, an intensively used ship-traffic lane crossing the southern North Sea from north-east to south-west (Alessandrini et al. 2017; BSH 2020) occupies a large part of the remaining area between the planned OWFs. General ship traffic is expected to increase (OSPAR 2010) and will exacerbate this situation. Furthermore, other anthropogenic activities, e.g., fisheries, will also become concentrated in the space between the OWFs and other available areas, increasing disturbance in these areas and further reducing the space available for guillemots. Alternative foraging areas with similar conditions that might compensate for this lost habitat and are located within an appropriate range will be restricted in German waters and in neighbouring countries which also use their offshore areas intensively and also plan to implement even larger scale OWFs (4COffshoreWind 2023).
Considering the various negative impacts, guillemots occurring in this area could be affected at a population level. If wind farms in Germany are realised as currently planned, 2.5%–3% of the European guillemot population would be affected by habitat loss (based on the European population estimate of 2,350,000–3,060,000 mature individuals; BirdLife International 2015). Moreover, the potential effects presented here do not include plans by other countries neighbouring the German EEZ, which also intend to cover large parts of their offshore areas with OWFs to meet the European goal to implement 300 GW of offshore wind power by 2050 (European Commission 2020; 4COffshoreWind 2023).
This study used a key top predator species in the southern North Sea as an example to assess the possible effects of large scale OWF implementation on the marine ecosystem. The strong disturbance responses exhibited by guillemots, combined with the large-scale implementation of OWFs, need to be considered when evaluating the conservation status of guillemots in the EU, especially in light of strong regional population declines (BirdLife International 2015; Gerlach et al. 2019). It is currently unknown how habitat loss will ultimately impact individual guillemots and whether it will affect the species at the population level. However, there are strong indications that displacement from preferred foraging habitats decreases the body condition, survival, and reproductive success of seabirds (Masden et al. 2010; Langton et al. 2014; Laursen et al. 2016), suggesting that large-scale OWFs have the potential to affect species at a population level (Busch and Garthe 2016).
Climate change is progressing rapidly and all possible efforts are needed to reduce human greenhouse gas emissions (Steffen et al. 2015a; IPCC 2018, 2021). However, the world is simultaneously facing one of the largest biodiversity losses in its history (Butchard et al. 2010; Hill et al. 2018; EEA 2019). Biodiversity is critically important for the stability of ecosystems (Hooper et al. 2012; Hautier et al. 2015; Steffen et al. 2015b) and stopping biodiversity loss is as fundamental for human societies as stopping climate change (Rockström et al. 2009; Cardinale et al. 2012; Steffen et al. 2015b). A sustainable scenario to reduce the human CO2 footprint is therefore needed (Fuso Nerini et al. 2019) that can balance these two major challenges (Steffen et al. 2015b): i.e., it is necessary to address climate change and simultaneously preserve biodiversity. This could be achieved by gradually increasing the area reserved for OWF implementation while allowing for the thorough monitoring of their effects on the marine environment, and including the lessons learned during the planning process. Planning also needs to consider research on and implementation of mitigation of OWF effects on wildlife, e.g., by adapting OWF locations, size and design, turbine height and spacing, as well as reducing OWF-associated ship traffic, light emissions, and noise emitted during construction.
Conclusion
The current findings clearly demonstrate the effect of currently operating OWFs, as well as the potential future threat to guillemots in the southern North Sea if the German government’s plans to implement large-scale OWFs are realised. The combination of species-specific OWF avoidance values with data on seabird distribution and abundance provides essential information on the magnitude of possible effects of large-scale OWF implementation on seabird abundance. Similar approaches are important to allow the development of sustainable planning scenarios for renewable energy developments in offshore areas, while preserving marine biodiversity and ecosystem functioning. Similar estimates should thus be made for other seabird species and regions. Against the background of the recently signed Esbjerg and Ostend Declarations (The Federal German Government 2022, 2023), estimates such as those provided here are urgently needed to support planning and management decisions. Such evaluations are essential to allow the development of a sustainable scenario for reducing the human CO2 footprint, whilst balancing the demands of climate change and biodiversity.
Data availability
Data are stored at the German Federal Maritime and Hydrographic Agency (BSH, Hamburg, Germany, https://www.geoseaportal.de), and German Federal Agency for Nature Conservation (BfN, Bonn, Germany, https://geodienste.bfn.de), and available on request.
References
Alessandrini A, Guizzardi D, Janssens-Maenhout G, Pisoni E, Trombetti M, Vespe M (2017) Estimation of shipping emissions using vessel Long Range Identification and Tracking data. J Maps 13:946–954. https://doi.org/10.1080/17445647.2017.1411842
BirdLife International (2015) Uria aalge. The IUCN Red List of Threatened Species 2015: e.T22694841A60108623. Downloaded on 27 May 2020
BirdLife International (2021) Uria aalge. The IUCN Red List of Threatened Species 2021: e.T22694841A166288433. https://doi.org/10.2305/IUCN.UK.2021-3.RLTS.T22694841A166288433.en. Accessed 20 June 2023
Borkenhagen K, Markones N, Garthe S (2019) Das Wirbeltiermonitoring in den deutschen Meeresgebieten und die Rolle digitaler Erfassungen im Monitoringkonzept. Jahrbuch Für Naturschutz Und Landschaftspflege 62:56–60
BSH (2017) Bundesfachplan Offshore für die deutsche ausschließliche Wirtschaftszone der Nordsee 2016/2017 und Umweltbericht https://www.offshore-stiftung.de/sites/offshorelink.de/files/documents/BFO_Nordsee_2016_2017.pdf. Accessed 20 June 2023
BSH (2020) https://www.bsh.de/DE/THEMEN/Offshore/Meeresraumplanung/meeresraumplanung_node.html. Accessed 20 June 2023
BSH (2021a) Anlage zur Verordnung über die Raumordnung in der deutschen ausschließlichen Wirtschaftszone in der Nordsee und in der Ostsee vom 19. August 2021. Anlagenband zum Bundesgesetzblatt Teil I Nr. 58 vom 26. August 2021. https://www.bsh.de/DE/THEMEN/Offshore/Meeresraumplanung/Raumordnungsplan_2021/_Anlagen/Downloads/Raumordnungsplan_2021.pdf;jsessionid=31A7766D314FB5DC8E1F8C7F8F502B87.live21323?__blob=publicationFile&v=10. Accessed 20 June 2023
BSH (2021b) Raumordnungsplan AWZ. https://www.geoseaportal.de/atomfeeds/Raumordnungsplan_AWZ_de.xml#download=1
BSH (2022a) https://www.bsh.de/DE/THEMEN/Offshore/Nutzungskarten/_Anlagen/Downloads/Nordsee-OffshoreWindparks.pdf;jsessionid=19635421247C3E99383ED6CB34BDC6C3.live11311?__blob=publicationFile&v=15. Accessed 20 June 2023
BSH (2022b) Zweiter Entwurf Flächenentwicklungsplan. https://www.bsh.de/DE/THEMEN/Offshore/Meeresfachplanung/Flaechenentwicklungsplan/_Anlagen/Downloads/FEP_2022_3/Zweiter_Entwurf_Flaechenentwicklungsplan.html;jsessionid=F0301A5B7C49502297628C8B9F950ED3.live21321?nn=1653366. Accessed 20 June 2023
BSH (2023a) Flächenentwicklungsplan 2023 für die deutsche Nordsee und Ostsee. https://www.bsh.de/DE/THEMEN/Offshore/Meeresfachplanung/Flaechenentwicklungsplan/_Anlagen/Downloads/FEP_2023_1/Flaechenentwicklungsplan_2023.html;jsessionid=F0301A5B7C49502297628C8B9F950ED3.live21321?nn=1653366. Accessed 20 June 2023
BSH (2023b) Laufende Fortschreibung Flächenentwicklungsplan. https://www.bsh.de/DE/THEMEN/Offshore/Meeresfachplanung/Laufende_Fortschreibung_Flaechenentwicklungsplan/lfd_forts_flaechenentwicklungsplan_node.html
Buckingham L, Daunt F, Bogdanova MI, Furness RW, Bennett S, Duckworth J, Dunn RE, Wanless S, Harris MP, Jardine DC, Newell MA, Ward RM, Weston ED, Green JA (2023) Energetic synchrony throughout the non-breeding season in common guillemots from four colonies. J Avian Biol e03018. https://doi.org/10.1111/jav.03018
Burger C, Schubert A, Heinänen S, Dorsch M, Kleinschmidt B, Zydelis R, Morkunas J, Quillfeld P, Nehls G (2019) A novel approach for assessing effects of ship traffic on distributions and movements of seabirds. J Environ Manage 251:109511
Burke CM, Montevecchi WA (2018) Taking the bite out of winter: common murres (Uria aalge) push their dive limits to surmount energy constraints. Front Mar Sci 5:63. https://doi.org/10.3389/fmars.2018.00063
Burke CM, Montevecchi WA, Regular PM (2015) Seasonal variation in parental care drives sex-specific foraging by a monomorphic seabird. PLoS One 10(11):e0141190. https://doi.org/10.1371/journal.pone.0141190
Busch M, Garthe S (2016) Approaching population thresholds in presence of uncertainty: assessing displacement of seabirds from offshore wind farms. Environ Impact Assess Rev 56:31–42
Butchard SHM, Walpole M, Collen B, van Strien A, Scharlemann JPW, Almond REA, Baillie JEM, Bomhard B, Brown C, Bruno J, Carpenter KE, Carr GM, Chanson J, Chenery AM, Csirke J, Davidson NC, Dentener F, Foster M, Galli A, Galloway JN, Genovesi P, Gregory RD, Hockings M, Kapos V, Lamarque J-F, Leverington F, Loh J, McGeoch MA, McRae L, Minasyan A, Hernández Morcillo M, Oldfield TEE, Pauly D, Quader S, Revenga C, Sauer JR, Skolnik B, Spear D, Stanwell-Smith D, Stuart SN, Symes A, Tierney M, Tyrrell TD, Vié J-C, Watson R (2010) Global biodiversity: indicators of recent declines. Science 328(5982):1164–1168
Camphuysen CJ (2002) Post-fledging dispersal of Common Guillemots Uria aalge guarding chicks in the North Sea: the effect of predator presence and prey availability at sea. Ardea 90(1):103–119
Camphuysen CJ, Leopold MF (1994) Atlas of seabirds in the southern North Sea. (IBN research report; No. 94/6), (NIOZ – rapport; No. 1994–8). NIOZ [etc.]. https://edepot.wur.nl/384797
Cardinale B, Duffy JE, Gonzales A, Hooper DU, Perrings C, Venail P, Narwani A, Mace GM, Tilman D, Wardle DA, Kinzig AP, Daily GC, Loreau M, Grace JB, Larigauderie A, Srivastava DS, Naeem S (2012) Biodiversity loss and its impact on humanity. Nature 486:59. https://doi.org/10.1038/nature11148
4COffshoreWind (2023) https://map.4coffshore.com/offshorewind/. Accessed 20 June 2023
Croxall JP, Prince PA, Reid K (1999) Diet, provisioning and productivity responses of marine predators to differences in availability of Antarctic krill. Mar Ecol Prog Ser 177:115–131
Daewel U, Akhtar N, Christiansen N, Schrum C (2022) Offshore wind farms are projected to impact primary production and bottom water deoxygenation in the North Sea. Commun Earth Environ 3:292. https://doi.org/10.1038/s43247-022-00625-0
Dähne M, Gilles A, Lucke K, Peschko V, Adler S, Krügel K, Sundermeyer J, Siebert U (2013) Effects of pile-driving on harbor porpoises (Phocoena phocoena) at the first offshore wind farm in Germany. Environ Res Lett 8:025002
De Mesel I, Kerckhof F, Norro A, Rumes B, Degraer S (2015) Succession and seasonal dynamics of the epifauna community on offshore wind farm foundations and their role as stepping stones for non-indigenous species. Hydrobiologia 756:37–50
Descamps S, Anker-Nilssen T, Barrett RT et al (2017) Circumpolar dynamics of a marine top-predator track ocean warming rates. Glob Change Biol 23:3770–3780
Deutscher Bundestag (2022) Drucksache 20/1634. Gesetzentwurf der Bundesregierung. Entwurf eines Zweiten Gesetzes zur Änderung des Windenergie-auf-See-Gesetzes und anderer Vorschriften; 02.05.2022. https://www.google.com/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&cad=rja&uact=8&ved=2ahUKEwi4-ceCs6X4AhUXQ_EDHVenCdgQFnoECAkQAQ&url=https%3A%2F%2Fdserver.bundestag.de%2Fbtd%2F20%2F016%2F2001634.pdf&usg=AOvVaw2NGTq7VxBky7Wokkcemhzk. Accessed 20 June 2023
Dias MP, Martin R, Pearmain EJ, Burfield IJ, Small C, Phillips RA, Yates O, Lascelles B, Borboroglu PG and Croxall JP (2019) Threats to seabirds: a global assessment. Biol Conserv 237:525–537
Dierschke J, Dierschke V, Hüppop O, Jachmann KF (2011) Die Vogelwelt der Insel Helgoland. OAG Helgoland, Helgoland
Dierschke V, Furness RW, Garthe S (2016) Seabirds and offshore wind farms in European waters: avoidance and attraction. Biol Cons 202:59–68
Dierschke J, Dierschke V, Stühmer F (2020) Ornithologischer Jahresbericht 2019 für Helgoland. Ornithol Jber Helgoland 30:1–97
Drewitt AL, Langston RH (2006) Assessing the impacts of wind farms on birds. Ibis 148(s1):29–42
Duffy JE (2003) Biodiversity loss, trophic skew and ecosystem functioning. Ecol Lett 6:680–687
Dunn R, Wanless S, Daunt F, Harris MP, Green JA (2020) A year in the life of a North Atlantic seabird: behavioural and energetic adjustments during the annual cycle. Sci Rep 10:5993. https://doi.org/10.1038/s41598-020-62842-x
EEA (2019) The European environment — state and outlook 2020. Knowledge for transition to a sustainable Europe. European Environment Agency, Copenhagen
Emeis K-C, van Beusekom J, Callies U, Ebinghaus R, Kannen A, Kraus G, Kröncke I, Lenhart H, Lorkowski I, Matthias V, Möllmann C, Pätsch J, Scharfe M, Thomas H, Weisse R, Zorita E (2015) The North Sea – a shelf sea in the Anthropocene. J Mar Syst 141:18–33
Estes JA, Terborgh J, Brashares JS, Power ME, Berger J, Bond WJ, Carpenter SR, Essington TE, Holt RD, Jackson JBC, Marquis RJ, Oksanen L, Oksanen T, Paine RT, Pikitch EK, Ripple WJ, Sandin SA, Scheffer M, Schoener TW, Shurin JB, Sinclair ARE, Soulé ME, Virtanen R, Wardle DA (2011) Trophic downgrading of planet earth. Science 333:301–306
European Commission (2019) Communication from the Commission to the European Parliament, the European Council, the Council, the European Economic and Social Committee and the Committee of the Regions. The European Green Deal. Brussels, COM (2019) 640 final
European Commission (2020) Communication from the Commission to the European Parliament, the Council, the European Economic and Social Committee and the Committee of the Regions. An EU Strategy to harness the potential of offshore renewable energy for a climate neutral future. Brussels, COM(2020) 741 final
European Commission (2021) REGULATION (EU) 2021/1119 OF THE EUROPEAN PARLIAMENT AND OF THE COUNCIL of 30 June 2021 establishing the framework for achieving climate neutrality and amending Regulations (EC) No 401/2009 and (EU) 2018/1999 (‘European Climate Law’). Official Journal of the European Union. L 243/1
Fliessbach KL, Borkenhagen K, Guse N, Markones N, Schwemmer P, Garthe S (2019) A ship traffic disturbance vulnerability index for northwest European seabirds as a tool for marine spatial planning. Front Mar Sci 6:192
Fort J, Porter WP, Grémillet D (2009) Thermodynamic modelling predicts energetic bottleneck for seabirds wintering in the northwest Atlantic. J Exp Biol 212:2483–2490
Frederiksen M, Wanless S, Harris MP, Rothery P, Wilson LJ (2004) The role of industrial fisheries and oceanographic change in the decline of North Sea black-legged kittiwakes. J Appl Ecol 41:1129–1139
Fuso Nerini F, Sovacool B, Hughes N, Cozzi L, Cosgrave E, Howells M, Tavoni M, Tomei J, Zerriffi H, Milligan B (2019) Connecting climate action with other Sustainable Development Goals. Nat Sustain 2:674–680. https://doi.org/10.1038/s41893-019-0334-y
Garthe S, Schwemmer H, Markones N, Müller S, Schwemmer P (2015) Verbreitung, Jahresdynamik und Bestandsentwicklung der Seetaucheer Gavia spec. in der Deutschen Bucht (Nordsee). Vogelwarte 53:121–138
Garthe S, Sonntag N, Schwemmer P, Dierschke V (2007) Estimation of seabird numbers in the German North Sea throughout the annual cycle and their biogeographic importance. Vogelwelt 128:163–178
Garthe S, Schwemmer H, Peschko V, Markones N, Müller S, Schwemmer P, Mercker M (2023) Large-scale effects of offshore wind farms on seabirds of high conservation concern. Sci Rep 13:4779. https://doi.org/10.1038/s41598-023-31601-z
Gaston AJ, Jones IL (1998) Bird families of the world: The Auks. Oxford University Press, Oxford
Gerlach B, Dröschmeister R, Langgemach T, Borkenhagen K, Busch M, Hauswirth M et al (2019) Vögel in Deutschland: Übersichten zur Bestandssituation. Eigenverlag des Dachverbandes Deutscher Avifaunisten e. V, Münster
Halpern BS, Walbridge S, Selkoe KA, Kappel CV, Micheli F, D’Agrosa C, Bruno JF, Casey KS, Ebert C, Fox HE, Fujita R, Heinemann D, Lenihan HS, Madi EMP, Perry MT, Selig ER, Spalding M, Steneck R, Watson R (2008) A global map of human impact on marine ecosystems. Science 319:948–952
Halpern BS, Frazier M, Potapenko J, Case KS, Koenig K, Longo C, Lowndes JS, Rockwood C, Selig ER, Selkoe KA, Walbridge S (2015) Spatial and temporal changes in cumulative human impacts on the world’s ocean. Nat Commun 6:7615
Halpern BS, Frazier M, Afflerbach J, Lowndes JS, Micheli F, O’Hara C, Scarborough C, Selkoe KA (2019) Recent pace of change in human impact on the world’s ocean. Sci Rep 9:11609
Harris MP, Wanless S (1990) Moult and autumn colony attendance of auks. Brit Birds 83:55–66
Harris MP, Webb A, Tasker ML (1991) Growth of young Guillemots Uria aalge after leaving the colony. Seabird 13:40–44
Harris MP, Wanless S, Ballesteros M, Moe B, Daunt F, Erikstad KE (2015) Geolocators reveal an unsuspected moulting area for Isle of May Common Guillemots Uria aalge. Bird Study 62(2):267–270. https://doi.org/10.1080/00063657.2015.1006164
Hastie GD, Russell DJF, McConnell B, Moss S, Thompson D, Janik VM (2015) Sound exposure in harbour seals during the installation of an offshore wind farm: predictions of auditory damage. J Appl Ecol 52:631–640
Hautier Y, Tilman D, Isbell F, Seabloom EW, Borer ET, Reich PB (2015) Anthropogenic environmental changes affect ecosystem stability via biodiversity. Science 348:6232
Hill SLL, Gonzalez R, Sanchez-Ortiz K, Caton E, Espinoza F, Newbold T, Tylianakis J, Scharlemann JPW, De Palma A, Purvis A (2018). Worldwide impacts of past and projected future land-use change on local species richness and the Biodiversity Intactness Index. https://doi.org/10.1101/311787
Hooper DU, Adair EC, Cardinale DJ, Byrnes JEK, Hungate BA, Matulich KL, Gonzalez A, Duffy JE, Gemfeldt L, O’Connor MI (2012) A global synthesis reveals biodiversity loss as a major driver of ecosystem change. Nature 486:105. https://doi.org/10.1038/nature11118
Intergovernmental Panel on Climate Change (IPCC) (2018) Summary for policymakers. In: Masson-Delmotte V, Zhai P, Pörtner H-O, Roberts D, Skea J, Shukla PR, Pirani A, Moufouma-Okia W, Péan C, Pidcock R, Connors S, Matthews JBR, Chen Y, Zhou X, Gomis MI, Lonnoy E, Maycock T, Tignor M, Waterfield T (eds) Global Warming of 1.5°C. An IPCC Special Report on the impacts of global warming of 1.5°C above pre-industrial levels and related global greenhouse gas emission pathways, in the context of strengthening the global response to the threat of climate change, sustainable development, and efforts to eradicate poverty. In Press
Climate Change 2021: The Physical Science Basis. Contribution of Working Group I to the Sixth Assessment Report of the intergovernmental panel on climate change. Masson-Delmotte, V, P Zhai, A Pirani, SL Connors, C Péan, S Berger, N Caud, Y Chen, L Goldfarb, MI Gomis, M Huang, K Leitzell, E Lonnoy, JBR Matthews, TK Maycock, T Waterfield, O Yelekçi, R Yu, B Zhou (eds). Cambridge University Press. In Press
Jones EL, Hastie GD, Smout S, Onoufriou J, Merchant ND, Brookes KL, Thompson D (2017) Seals and shipping: quantifying population risk and individual exposure to vessel noise. J Appl Ecol 54:1930–1940
Langton R, Davies IM, Scott BE (2014) A simulation model coupling the behaviour and energetics of a breeding central place forager to assess the impact of environmental changes. Ecol Model 273:31–43
Laursen K, Møller AP, Holm TE (2016) Dynamic group size and displacement as avoidance strategies by eiders in response to hunting. Wildl Biol 22(4):174–181. https://doi.org/10.2981/wlb.00197
Leopold MF, van Bemmelen RSA, Zuur AF (2013) Responses of local birds to the offshore wind farms PAWP and OWEZ off the Dutch mainland coast. Report C151/12. Imares, Wageningen
Leopold MF, Kik M, van Tulden P, van Franeker JA, Kühn S, Rijks J (2019) De Zoe en de zeekoet. Een onderzoek naar de doodsoorzaak en de herkomst van de zeekoeten die massaal strandden op de Nederlandse kust in januarifebruari 2019. Wageningen University & Research rapport C026/19
Lindeboom HJ, Kouwenhoven HJ, Bergman MJN, Bouma S, Brasseur S, Daan R, Fijn RC, de Haan D, Dirksen S, van Hal R, Hille Ris Lambers R, ter Hofstede R, Krijgsveld KL, Leopold M, Scheidat M (2011) Short-term ecological effects of an offshore wind farm in the Dutch coastal zone; a compilation. Environ Res Lett 60:035101
Masden EA, Haydon DT, Fox AD, Furness RW (2010) Barriers to movement: modelling energetic costs of avoiding marine wind farms amongst breeding seabirds. Mar Pollut Bull 60:1085–1091
Mendel V, Schwemmer P, Peschko V, Müller S, Schwemmer H, Mercker M, Garthe S (2019) Operational offshore wind farms and associated ship traffic cause profound changes in distribution patterns of loons (Gavia spp.). J Environ Manage 231:429–438
Mendel B, Sonntag N, Wahl J, Schwemmer P, Dries H, Guse N, Müller S, Garthe S (2008) Profiles of Seabirds and waterbirds of the German North and Baltic Seas. Distribution, ecology and sensitivities to human activities within the marine environment. Federal Agency for Nature Conservation. Bonn – Bad Godesberg
Mendel B (2012) Anthropogene Nutzungen und deren Auswirkungen auf Seevögel in der Deutschen Bucht: Entwicklung methodischer Ansätze und Bewertung der Effekte. Dissertation, University of Kiel
Mercker M, Dierschke V, Camphuysen K, Kreutle A, Markones M, Vanermen N, Garthe S (2021a) An indicator for assessing the status of marine-bird habitats affected by multiple human activities: a novel statistical approach. Ecol Ind 130:108036. https://doi.org/10.1016/j.ecolind.2021.108036
Mercker M, Markones N, Borkenhagen K, Schwemmer H, Wahl J, Garthe S (2021b) An integrated framework to estimate seabird population numbers and trends. J Wildlife Manage 1–21. https://doi.org/10.1002/jwmg.22026
OSPAR (2010) Quality Status Report 2010. OSPAR Commission, London
Parsons M, Mitchell I, Butler A, Ratcliffe N, Frederiksen M, Foster S, Reid JB (2008) Seabirds as indicators of the marine environment. ICES J Mar Sci 65:1520–1526
Perrow MR (ed) (2019) Wildlife and wind farms, conflicts and solutions, vol 3. Potential effects. Pelagic Publishing, Exeter/Offshore
Peschko V, Mendel B, Müller S, Markones M, Mercker M, Garthe S (2020a) Effects of offshore windfarms on seabird abundance: strong effects in spring and in the breeding season. Mar Environ Res 62:105157. https://doi.org/10.1016/j.marenvres.2020.105157
Peschko V, Mercker M, Garthe S (2020b) Telemetry reveals strong effects of offshore wind farms on behaviour and habitat use of common guillemots (Uria aalge) during the breeding season. Mar Biol 167:118
R Core Team (2023) R: A language and environment for statistical computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.Rproj ect.org/
Reubens JT, Braeckman U, Vanaverbeke J, Van Colen C, Degraer S, Vincx M (2013) Aggregation at windmill artificial reefs: CPUE of Atlantic cod (Gadus morhua) and pouting (Trisopterus luscus) at different habitats in the Belgian part of the North Sea. Fish Res 139:28–34
Rockström J, Steffen W, Noone K, Persson A, Chapin FS, Lambin EF, Lenton TM, Scheffer M, Folke C, Schellnhuber HJ, Nykvist B, de Wit CA, Hughes T, van der Leeuw S, Rodhe H, Sörlin S, Snyder PK, Costanza R, Svedin U, Falkenmark M, Karlberg L, Corell RW, Fabry VJ, Hansen J, Walker B, Liverman D, Richardson K, Crutzen P, Foley JA (2009) A safe operating space for humanity. Nature 461:472–475. https://doi.org/10.1038/461472a
Russell DJF, Hastie GD, Thompson D, Janik VM, Hammond PS, Scott-Hayward LAS, Matthiopoulos J et al (2016) Avoidance of wind farms by harbour seals is limited to pile driving activities. J Appl Ecol 53:1642–1652
Schreiber EA, Burger J (2001) Biology of marine birds. CRC Press, CRC Marine Biology Series, p 740
Schwarz CJ (2014) Analysis of BACI experiments (Chapter 12). In: Course notes for beginning and intermediate statistics. Available at: http://www.stat.sfu.ca/cschwarz/CourseNotes
Sergio F, Newton I, Marchesi L, Pedrini P (2006) Ecologically justified charisma: preservation of top predators delivers biodiversity conservation. J Appl Ecol 43:1049–1055
Skov H, Durinck J, Leopold MF, Tasker ML (1995) Important Bird Areas for seabirds in the North Sea, including the Channel and Kattegat. BirdLife international, Cambridge
Smith EP (2002) BACI design, ecological statistics. Wiley
Sonntag N (2001) Winterverbreitung der Trottellumme (Uria aalge) in der Nordsee und Koloniebesuche auf Helgoland: Einfluss von Wassertemperatur und Nahrungsverfügbarkeit. Diplomarbeit der Fakultät für Biologie der Eberhard-Karls-Universität Tübingen, 111 p
Steffen W, Broadgate W, Deutsch L, Gaffney O, Ludwig C (2015a) The trajectory of the Anthropocene: The Great Acceleration. Anthrop Rev 2(1):81–98. https://doi.org/10.1177/2053019614564785
Steffen W, Richardson K, Rockström J, Cornell SE, Fetzer I, Bennett EM, Biggs R, Carpenter SR, de Vries W, de Wit CA, Folke C, Gerten D, Heinke J, Mace GM, Persson LM, Ramanathan V, Reyers B, Sörlin S (2015b) Planetary boundaries: guiding human development on a changing planet. Science 347(6223):1259855. https://doi.org/10.1126/science.1259855
Stenberg C, Støttrup JG, van Deurs M, Berg CW, Dinesen GE, Mosegaard H, Grome TM, Leonhard SB (2015) Long-term effects of an offshore wind farm in the North Sea on fish communities. Mar Ecol Prog Ser 528:257–265
The Federal German Government (2021) Koalitionsvertrag 2021 – 2025 zwischen der Sozialdemokratischen Partei Deutschlands (SPD), BÜNDNIS 90/DIE GRÜNEN und den Freien Demokraten (FDP). https://www.bundesregierung.de/breg-de/service/gesetzesvorhaben/koalitionsvertrag-2021-1990800
The Federal German Government (2022) The Esbjerg Declaration on The North Sea as a Green Power Plant of Europe. Esbjerg, 18.05.2022. https://www.google.com/url?sa=t&rct=j&q=&esrc=s&source=web&cd=&cad=rja&uact=8&ved=2ahUKEwjd2bbYwqX4AhUKNOwKHcAHC84QFnoECAIQAQ&url=https%3A%2F%2Fwww.bundesregierung.de%2Fresource%2Fblob%2F974430%2F2040932%2Fb357fa6726099a0304ee97c3a64e411c%2F2022-18-05-erklaerung-nordsee-gipfel-data.pdf%3Fdownload%3D1&usg=AOvVaw3PoCV_9SWHIrLHSNZ8KTsU
The Federal German Government (2023) Ostend Declaration of Energy Ministers on the North Seas as Europe’s Green Power Plant Delivering Cross-Border Projects and Anchoring the Renewable Offshore Industry In Europe. https://www.bmwk.de/Redaktion/DE/Downloads/Energie/ostend-declaration-energy-ministers-north-seas-europes-green-power-plant.pdf?__blob=publicationFile&v=4
Thomas L et al (2010) Distance software: design and analysis of distance sampling surveys for estimating population size. J Appl Ecol 47:5–14
United Nations Framework Convention on Climate Change (UNFCCC) (2015a) Adoption of the Paris Agreement. Report No. FCCC/CP/2015/L.9/Rev.1. http://unfccc.int/resource/docs/2015/cop21/eng/l09r01.pdf
UNFCCC (2015b) INDCs as communicated by Parties. http://www4.unfccc.int/submissions/indc/Submission%20Pages/submissions.aspx
Vallejo GC, Grellier K, Nelson EJ, McGregor RM, Canning SJ, Caryl FM, McLean N (2017) Responses of two marine top predators to an offshore wind farm. Ecol Evol 7:8698–8708
Van Deurs M, Grome TM, Kaspersen M, Jensen H, Stenberg C, Sørensen TK, Støttrup J, Warnar T, Mosegaard H (2012) Short- and long-term effects of an offshore wind farm on three species of sandeel and their sand habitat. Mar Ecol Prog Ser 458:169–180
Vandendriessche S, Derweduwen J, Hostens K (2015) Equivocal effects of offshore wind farms in Belgium on soft substrate epibenthos and fish assemblages. Hydrobiologia 756(1):19–35
Vanermen N, Onkelinx T, Courtens W, Van dewalle M, Verstraete H, Stienen EWM, (2015) Seabird avoidance and attraction at an offshore wind farm in the Belgian part of the North Sea. Hydrobiologia 756:51–61
Velarde E, Anderson DW, Ezcurra E (2019) Seabird clues to ecosystem health. Science 365(6449):116–117. https://doi.org/10.1126/science.aaw9999
Venables WN, Ripley BD (eds) (2002) (2002) Modern applied statistics with S. Fourth Edition. Springer, New York
Welcker J, Nehls G (2016) Displacement of seabirds by an offshore wind farm in the North Sea. Mar Ecol Prog Ser 554:173–182
Wickham H (2009) ggplot2: elegant graphics for data analysis. Springer, New York
Wilmers CC, Estes JA, Edwards M, Laidre KL, Konar B (2012) Do trophic cascades affect the storage and flux of atmospheric carbon? An analysis of sea otters and kelp forests. Front Ecol Environ 10:409–415
Wilson DE, Mittermeier RA (2014) Handbook of the mammals of the world, vol 4. Sea mammals. Lynx Edicions, Barcelona
Wind energy in Europe. 2021 statistics and the outlook for 2022–2026. Windeurope, Brussels
WindEurope (2019) Our energy, our future. How offshore wind will help Europe go carbon-neutral. Report by WindEurope, Brussels
Wood SN (2006) Generalized additive models: an introduction with R. Chapman & Hall/CRC
Zeileis SJA & Kleiber C (2008) Regression models for count data in R. URL http://www.jstatsoft.org/v27/i08/. J Stat Softw 27:1–25
Acknowledgements
The following environmental consulting companies also recorded bird-distribution data: BioConsult SH GmbH, IBL Umweltplanung GmbH, and Institut für Angewandte Ökosystemforschung GmbH. S. Furness provided linguistic support. K. Fliessbach and S. Weiel provided valuable comments.
Funding
Open Access funding enabled and organized by Projekt DEAL. Major parts of the data collection and analyses were conducted within the research projects OWP-Seevögel (FKZ:3520860100), MONTRACK (project no.: Z 1.2-532 02/AWZ/2017/2), and TopMarine (Z 1.2 53202/AWZ/2017/7/CAU), funded by the German Federal Agency for Nature Conservation, as well as HELBIRD, funded by the German Federal Ministry for Economic Affairs and Energy, and CoastalFutures (grant number 03F0911C), funded by the German Federal Ministry of Education and Research.
Author information
Authors and Affiliations
Contributions
All authors jointly developed this study. VP initialized and led this project. MM, VP, HS analysed the data with support from the other authors. VP and MM wrote the paper. All co-authors contributed to the drafts and gave final approval for publication.
Corresponding author
Ethics declarations
Competing interests
The authors have no competing interests to declare.
Additional information
Communicated by David Hawksworth.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Peschko, V., Schwemmer, H., Mercker, M. et al. Cumulative effects of offshore wind farms on common guillemots (Uria aalge) in the southern North Sea - climate versus biodiversity?. Biodivers Conserv 33, 949–970 (2024). https://doi.org/10.1007/s10531-023-02759-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10531-023-02759-9