
Abstract
Fire offers both opportunities and risks for wildlife. Its impact will depend on the fire’s scale, how it alters key resources and how animals move. Understanding how wildlife respond to fire is crucial as climate change is predicted to increase wildfire risk and will likely result in more frequent prescribed fire to reduce wildfire risk. Invasive predators and inappropriate fire regimes in south-eastern Australia threaten the long-nosed potoroo (Potorous tridactylus), a vulnerable marsupial often residing in areas frequently exposed to fire. The cumulative impacts of fire and predation may increase the threat to P. tridactylus after fire, as predators can be more effective in the immediate post-fire environment and P. tridactylus is often dependent on thick ground cover. We present a before-after control-impact experiment describing the influence of prescribed fire on P. tridactylus. We fitted GPS collars to 52 individuals at nine independent sites to test if exposure to prescribed burning reduced their survival or altered their movement behavior. Prescribed fire reduced P. tridactylus survival, yet range size and diffusion (movement) rate remained largely unaffected. With limited fire exposure, P. tridactylus tended to continue using burnt areas whereas activity became restricted to unburnt areas when larger proportions of their home range burnt. Site fidelity was very high - individuals rarely moved their home ranges after fire, regardless of fire exposure. Our results suggest recently burnt areas may be particularly dangerous for P. tridactylus: areas that can be attractive yet confer lower fitness outcomes. P. tridactylus may benefit from smaller fire scars, retention of structurally complex vegetation, and integrating invasive predator control with prescribed burning.
Similar content being viewed by others

Avoid common mistakes on your manuscript.
Introduction
Animals often alter their movement behavior to survive disturbance events (Sergio et al. 2018; Nimmo et al. 2019; Doherty et al. 2021. An animal’s movements reflect its environment, internal state, and perceptual and movement capacity (Nathan et al. 2008). Understanding the drivers of animal movement is important: movement behaviors determine key aspects of animal fitness, including access to food and mates, and exposure to predators and competitors (Nathan et al. 2008; Allen and Singh 2016; Nimmo et al. 2019). Movement behaviors are flexible, varying through space and time from daily foraging to seasonal migration (Rettie and Messier 2000; Nathan et al. 2008). This flexibility is critical to individual survival and population persistence in disturbed environments (Winkler et al. 2014; Nimmo et al. 2019).
Fire affects fauna globally (Yibarbuk et al. 2001; Bond and Keeley 2005) by consuming plant matter and causing species turnover in the plant community (Phillips and Waldrop 2008; Higgins et al. 2007; Haslem et al. 2016). Consequently, fire offers both opportunities and risks for fauna (Nimmo et al. 2019). For example, vegetation growth and a more open environment after fire can benefit large herbivores (e.g. kangaroos, antelope, deer) by offering food and potentially lowering predation risk from ambush predators (Meers and Adams 2003; Archibald and Bond 2004; Cherry et al. 2018). In contrast, species that need thick vegetation for shelter may be disadvantaged by fire as the post-fire habitat may be less suitable or riskier to move through (Fordyce et al. 2016; Nimmo et al. 2019; Doherty et al. 2022). Prescribed burning is often used to reduce wildfire risk however climate change is predicted to increase the frequency, scale, and severity of wildfire in many systems (Morgan et al. 2020; Jones et al. 2022). This may result in more frequent application of prescribed fire in many areas to manage this risk and therefore, studies describing the survival and movement behavior of fauna after such fire are sorely needed (Nimmo et al. 2019; Doherty et al. 2022).
Animal movements can be characterized in many ways. Examples include home range size (an animal’s space use requirements, Burt 1943); movement rate (how frequently they move around their range to acquire resources, Novellie 1978); site fidelity (an animal’s association with a specific area through time, Switzer 1993); and resource selection (how frequently an animal uses a resource relative to its availability, Boyce and McDonald 1999). Using multiple approaches to describe movement behavior may help overcome the limitations of any one metric and better elucidate ecologically important patterns. For example, two prey species with similar home range sizes and spatial overlap with a predator can have contrasting predator encounter rates if they vary in their movement rate and resource selection (Suraci et al. 2022).
The long-nosed potoroo (Potorous tridactylus) is a medium-sized marsupial (~ 1 kg) that has declined following European human invasion of Australia (Burbidge and McKenzie 1989; Johnston 2008; Woinarski et al. 2015) and often resides in areas that undergo prescribed fire. It typically uses dense vegetation for shelter and more open environments for foraging (Bennett 1993; Norton et al. 2011). P. tridactylus contributes to ecosystem function by turning over soil in search of food, driving many ecosystem processes (Valentine et al. 2017; Nest et al. 2023). P. tridactylus may be particularly vulnerable to predation shortly after fire as invasive predators such as foxes (Vulpes vulpes) and feral cats (Felis catus) can sometimes select strongly for recently burnt environments (McGregor et al. 2016; Hradsky et al. 2017). Occurrence of P. tridactylus often declines immediately after fire and can take several years to recover (Claridge and Barry 2008; Arthur et al. 2012) Understanding how P. tridactylus responds to the immediate effects of fire can aid conservation efforts by identifying post-fire refuges and determining if invasive predator control is required.
We present a replicated before-after control-impact study to test if prescribed burning reduces P. tridactylus survival or alters their movement behavior, and describe how this impact varies with increasing burn exposure and sex of individual potoroos. We expected (1) P. tridactylus would be less likely to survive and more likely to alter their movement behavior as more of their pre-fire home range burnt; (2) P. tridactylus would avoid recently burnt areas due to the simplification of vegetation structure (Bennet 1993; Claridge et al. 2007; Norton et al. 2010); and (3) P. tridactylus would shift their home ranges away from recently burnt areas as seen in another similar-sized macropod elsewhere (Povh et al. 2023).
Methods
Site description
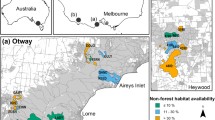
Our study was conducted in the western Heathy Woodlands of the Great Otway National Park (38.54 S, 143.47 E), Victoria, Australia. This locality is 130–250 m above sea level and has a mild, temperate climate: maximum daily temperatures average 26.1 OC in summer and 11.6 OC in winter; annual rainfall averages 538 mm with more rainfall in winter and spring (June to November) than the warmer months of summer and autumn (December to May) (Bureau of Meterology 2023). The overstorey is dominated by Eucalyptus baxteri and E. obliqua, with a mid-story dominated by Banksia marginata, Epacris impressa, Xanthorrhoea australis and Leptospermum continentale, with Melaleuca squarrosa becoming the dominant mid-story species in drainage lines. The understory predominantly comprises a diverse array of herbs, graminoids, and lichens. Camera trap detections suggest P. tridactylus tend to occur homogeneously throughout the structurally complex stands of the Austral Grass-tree (Xanthorrhoea australis) that occur along ridges within our study area (Le Pla, pers. obs., Le Pla et al. 2023; Figure S1).
Prescribed burning
Prescribed burning is often undertaken by land managers within our study region with the aim of reducing wildfire risk to local communities (Gazzard et al. 2020). Fires in our study region typically consume between 70 and 100% of available vegetation as the composition and structural complexity of the environment promotes high severity fires (Gazzard et al. 2020; Figure S1). Local fire practitioners conducted a series of eight prescribed burns during the cooler and wetter months (April – September) of 2020–2023. Each burn was within a designated burn “block” – a defined area (often bounded by roads or watercourses) that was designated for burning by local authorities as part of the study region’s bushfire management strategy (Gazzard et al. 2020). These areas tended to be bounded by roads or watercourses. Within each block, the total area burnt (7.37–383.67 hectares) and number of fire scars (1–19) varied substantially. One block remained unburnt, resulting in a total of nine different blocks where we followed P. tridactylus movements before, during and after fire.
Live trapping, data collection and data pre-processing
P. tridactylus were trapped using wire cage traps and GPS collars were fitted to animals weighing more than 500 g (see Table S1 for details). Females with large pouch young (~ 7% of total captures) were not included in this study to minimize the risk of pouch young ejection (Le Pla et al. 2023). During 2020 deployments, GPS collars collected fixes every half hour during the night (1800 –0600 h) and at three-hourly intervals during the day (0600–1800 h). Initial data suggested P. tridactylus were more active during the day than expected and therefore, GPS collars in 2021–2023 deployments collected fixes at hourly intervals continuously. Estimated battery life for GPS collars was approximately six weeks. We timed collar deployments with the goal of capturing two weeks of pre-fire and four weeks of post-fire movement data.
We attached GPS collars to 52 animals between 2020 and 2023. Forty of these animals were exposed to prescribed fire. Beyond minor hair loss and light rubbing around the neck of some individuals, no observable negative effects of collar attachment, such as loss of body mass was observed (Le Pla et al. 2023). Prior to analysis, GPS data were pre-processed to remove outliers, account for location uncertainty, and remove fixes associated with resting behavior. More detail on these procedures can be found in the supplementary information.
Survival and movement behavior response variables
Survival
The survival status (alive or dead) of each individual at the completion of the study (up to 52 days after collar fitting).
Home range size
We fit continuous-time movement models and estimated 95% utilization distributions (hereafter “home range”) before and after fire for all individuals using an error-informed, optimally weighted autocorrelated kernel density estimator (wAKDE; Fleming et al. 2015; Fleming et al. 2018) within the ctmm package (version 1.2.1, Fleming and Calabrese 2023). Prior to estimating home range size, we ensured individuals were range residents by visually assessing empirical variograms and confirming semi-variance had plateaued (Calabrese et al. 2016). Because wAKDE corrects for differences in autocorrelation (Noonan et al. 2019), sampling regime (Fleming et al. 2018) and measurement error (Fleming et al. 2021), it allows for reliable comparisons across time periods, sites and individuals.
Diffusion rate
We estimated diffusion rate, the mean rate of each individual’s movements within their home range before and after fire, using the ctmm package (version 1.2.1, Fleming and Calabrese 2023). Diffusion rates are proportional to the area over which an animal can be expected to range on an average day (Gill et al. 2023) and serve as a robust measure of an animal’s movement rate when tracking data are temporally autocorrelated and the fix schedule is too coarse to estimate speed effectively.
Home range overlap
We compared the similarity of the pre- and postfire utilization distributions for each individual using the Bhattacharyya coefficient (BC; Bhattacharyya 1943). The BC satisfies many desirable criteria when comparing utilization distributions, including statistical validity, objectivity, asymptotic consistency, and quantifiable uncertainty (Winner et al. 2018). The BC is a function of the product of the two distributions being compared and produces values ranging from 0 to 1, wherein a BC of 0 suggests the two distributions have no similarity and a BC of 1 suggests the two distributions are identical (Winner et al. 2018).
Resource selection
We modelled resource selection by fitting integrated resource selection functions (iRSF) to each individual before and after fire using the rsf.fit function using the ctmm package. These functions are functionally equivalent to an inhomogeneous Poisson point-process model (Alston et al. 2023; Gill et al. 2023). Locations within each individual’s 95% wAKDE home-range estimate were used as the known locations, while the rsf.fit function generated randomly distributed non-detections. iRSF’s do not require a pre-defined number of pseudo-absences; rather one must specify a target standard error, here 0.01, and increasingly large samples of points (on the order of thousands to tens of thousands) are generated until this threshold is reached (Alston et al. 2023; Gill et al. 2023). This approach estimates the area available to each individual (i.e. the sampling window) alongside selection coefficients simultaneously, negating the need to arbitrarily define an availability domain.
Predictor variables
Burn exposure
To assess evidence for a generalizable impact of fire, we used a two-level categorical variable ‘burn’ wherein we categorized P. tridactylus as either: unburnt (0% of pre-fire range burnt and home range center > 100 m from a burnt area) and burnt (> 1% of pre-fire range burnt).
Proportion of home range burnt
Individuals exposed to fire experienced highly variable degrees of burn exposure, ranging from less than 5–100% of their pre-fire home range (Figure S2). To explore the potential for animals to display different movement behaviors as more of their pre-fire home range was burnt, we used a continuous numerical variable ‘prop_burned’ (defined as the proportion (%) of an individual’s pre-fire home range that burnt). We categorized habitat as either burnt or unburnt as fires tended to produce a fundamentally simplified structural environment (Figure S1).
Sex
Male P. tridactylus have larger home ranges relative to females in our study region (Le Pla et al. 2023). Therefore, we explored the potential for sex-based differences in survival and movement behavior post-fire using a two-level categorical variable ‘sex’: male or female.
Burnt area: For resource selection behaviors, we assessed support for the selection for or against burnt areas with a two-factor categorical variable ‘burn’: unburnt (the reference category) or burnt. The selection or avoidance of burnt areas was only assessed for animals that had exposure to burnt areas.
Analytical framework and rationale
We assessed the impact of prescribed fire on P. tridactylus survival and each movement behavior (home range size, diffusion rate, site fidelity and resource selection) within a two-stage modelling framework. In both stages, alternative model structures were compared within an information theoretic framework (Burnham and Anderson 2002). Models were compared using Akaike Information Criterion adjusted for small sample size values (AICC), wherein models within two AICC units of the top model were considered equally plausible, and models within five AICC units of the top model were considered to have moderate support (Burnham and Anderson 2002).
For each response variable, we first tested for a causal impact of fire by comparing a model including the predictor variable burn exposure (burnt, unburnt) to an intercept only (null) model. Support for a causal impact of fire in stage one was inferred by assessing the coefficient estimate and uncertainty of the ‘burn’ covariate, with statistical significance being inferred if the 95% confidence intervals of this estimate did not overlap 0. This is because information theoretic approaches to parameter selection apply a more liberal significance level of p < 0.157 (Sutherland et al. 2023).
We then explored the potential influence of greater fire exposure (‘prop_burned’) and sex (‘sex’) on each response variable. We fitted models including ‘prop_burned’ and ‘sex’ individually, alongside additive and interactive models, and an intercept only model. This resulted in a total of five candidate models for survival and each movement metric in the second stage of our modelling framework. In both stages we interpreted non-overlapping 85% confidence intervals around two predicted point estimates as evidence of their statistical difference. This is equivalent to observing that the 95% confidence interval of the difference between two estimates does not overlap 0 (MacGregor-Fors and Payton 2013).
Survival
We estimated the probability of an individual being alive at the end of collar deployments (approximately six weeks after fitting) by fitting a mixed effects binomial logistic regression model with a two-level categorical covariate (alive or dead) as the response variable and block as a random effect.
Home range size and diffusion rate
For each individual, the response variable was the percentage change in home range size and diffusion rate after fire, relative to the pre-fire period (for animals exposed to fire) or the first fortnight of tracking (for animals not exposed to fire). Home range and diffusion rate models were fitted using generalized linear mixed effect models with a Gaussian residual distribution and an identity link. Block was included as a random effect in all model fitting. Standardizing our response variable to percentage change in home range size allowed us to control for the highly variable pre-fire home range sizes and diffusion rates (Figure S2) and better elucidated fire-induced changes in these behaviors. Because the response variable (percentage change) explicitly accounts for time, support for either ‘burn’ (in stage one) or ‘prop_burned’ (in stage two) would be indicative of an effect of fire.
Additionally, we estimated the effective sample size (an estimate of the information content of tracking data after accounting for autocorrelation; see Fleming and Calabrese 2016) of home range size and diffusion rate predictions within each time period (before and after fire). Larger effective sample sizes allow for more precise estimates of home range size and diffusion rate. Individuals with effective sampling sizes < 10 within either of the before or after fire periods were not included in this analysis as these home range and diffusion estimates had considerable uncertainty relative to other animals (see Noonan et al. 2019).
Home range overlap
We compared the similarity of home ranges from the pre-fire and post-fire periods for each individual using the BC (Bhattacharyya 1943; Winner et al. 2018). In our study context, BC values represent the relative change in the utilization distribution of GPS tracked animals between periods (before and after fire). As BC values are bounded between 0 and 1, we fitted binomial logistic regression models with a beta binomial residual distribution to test for an impact of fire and explore the influence of sex and burn exposure on site fidelity. We considered BC values between 1 and 0.75 to represent minor changes in home range location, values between 0.75 and 0.5 to represent moderate changes in home range location, and values < 0.5 to represent major changes in home range location.
Resource selection
We assessed evidence for attraction, indifference or avoidance of recently burnt areas by fitting a separate iRSF to each individual’s post-fire movement data. Individuals with no burn exposure were not included in this analysis as no contrast between burnt and unburnt areas was available. Therefore, only stage two analysis was conducted for this response variable. Population-level patterns in resource selection as a function of burn exposure level were obtained by averaging the individual coefficients and their associated uncertainty using the meta-regression models implemented in the R package metafor (ver. 3.8-1, Viechtbauer, 2010).
Model validation and diagnostics
All models were fit within the R programming environment (R Core Team 2023) and residual diagnostics in all fitted models was assessed using the performance package (v. 0.10.8, Lüdecke et al. 2021), with no issues detected.
Results
Fire reduced P. Tridactylus survival
Survival was low for animals exposed to fire (18/40 individuals, 45%) relative to those with no burn exposure (10/12 individuals, 83%). There was no evidence that the fires directly caused any mortality events (e.g. burnt GPS collars). The logistic regression model indicated that there was strong statistical evidence that prescribed burning reduced the probability of P. tridactylus being alive four weeks after fire (β = -3.01, 95% CI: -5.83 – -0.2, Table 1). Model estimates suggested animals in unburnt environments were almost three times as likely to be alive at study completion (91%, 85% CI: 64–98) relative to animals exposed to fire (34%, 85% CI: 16–58) (Fig. 1). Stage two analysis suggested that animals with greater proportions of their home range burnt were less likely to survive after fire (Fig. 1; Table 1). There was little evidence that survival differed between males and females - this covariate was an uninformative parameter (Arnold 2010).

Predictions from binomial regression showing the probability of P. tridactylus survival one month after fire as a function burn exposure (top) and the level of burn exposure (bottom). 84% confidence intervals are represented by error bars (top) and grey shading (bottom)
Fire had a varied impact on home range size
The mean pre-fire home range size of female and male P. tridactylus was 4.54 hectares (95%: CI: 3.54–5.54) and 14.17 hectares (95% CI: 10.40– 18.00) respectively. Home range size effective sample sizes were high in both time periods with a mean of 63 (range: 14–154) for the pre-fire period and 147 (range: 6–330) for the post-fire period (Table S2).
Whether or not an animal was exposed to fire was not a good predictor of how they changed the size of their home range (Table 2, Figure S4). That is, the best performing model in our stage one analysis was the ‘null’ model, and the coefficient estimate for ‘burn’ in the alternative model had confidence intervals that overlapped zero (β = 7.31, 95% CI: -30.44–15.82, Table 2). Instead, our stage two analysis supported an effect of both sex and proportion of home range burnt (Table 2; Fig. 2). The top performing model (‘prop_burned + sex’) indicated that males were more likely to increase their home range between periods (β = 16.99, 95% CI: 0.88–33.11), and animals with larger proportions of their home range burnt were more likely to decrease their home range size after fire (β = -7.95, 95% CI: -16.62 – -0.71). Point estimates predicted from this model suggested females were more likely to reduce their home range once more than 30% of their pre-fire home range was burnt, and males were more likely to increase their home range size when small proportions (< 30%) of their pre-fire home range burnt (Fig. 2).

Predictions from a generalized linear model showing the percentage change (%) in P. tridactylus home range size as a function of sex (males = solid line, females = dotted line) and burn exposure level. Positive values suggest an increase in home range size relative to the pre-fire period, and negative values suggest the opposite. 84% confidence intervals are represented by grey shading. Raw data is overlayed for males (hollow triangles) and females (crosses
Fire had no impact on diffusion rate
The mean pre-fire diffusion rate of female and male P. tridactylus was 7.69 hectares per day (95%: CI: 6.61–8.77) and 20.16 hectares per day (95% CI: 15.3–25) respectively. Diffusion rate effective sample sizes were similarly high in both time periods with a mean of 113 (range: 11–473) for the pre-fire period and 261 (range: 33–986) for the post-fire period (Table S2). Diffusion rate scaled positively with home range size (Figure S3) and animals typically covered a larger area than their home range each day: the mean ratio of pre-fire home range size to diffusion rate was 1.92 (range: 0.48–4.01) for females and 1.68 (range: 0.48–2.88) for males.
Whether or not an animal was exposed to fire was not a good predictor of how they changed their diffusion rate (Table 3, Figure S5). That is, the best performing model in stage one was the ‘null’ model, and the coefficient estimate for ‘burn’ in the alternative model had confidence intervals that overlapped zero (β = 13.11, 95% CI: -9.91–36.13) (Table 3). Likewise, neither sex nor proportion of home range burnt substantially improved model fit compared to the null model in our stage two analysis (Table 3).
Fire had no impact on site fidelity
There was sufficient data to estimate the BC for 46 of the 52 individuals (Table S2). There was little evidence that animals moved their home ranges in a meaningful way post-fire (Figure S9). In both stages, all models failed to reach convergence, likely due to most BC values being close to numerical boundaries (i.e. near 1). Removing the random effect of ‘block’ did little to address model fitting issues. The mean BC value was 0.91 (95% CI: 0.88–0.95) for animals exposed to fire and 0.96 (95% CI: 0.94–0.98) for animals with no burn exposure. Visual inspection of the data suggested the largest deviations in home range location tended to occur in animals with high burn exposure, but only four individuals (9%) showed a moderate change in home range similarity (BC values between 0.75 and 0.5) and no animals changed their home range location by more than 50% (Figure S9). The few animals that moved their home range location in a meaningful way after fire typically shifted into the nearest adjacent unburnt habitat.
P. Tridactylus displayed contrasting selection for burnt areas
Of the 40 animals exposed to fire, we had sufficient data to estimate selection for recently burnt areas for 33 individuals. Of the remaining seven, resource selection could not be assessed for five individuals with high fire exposure as they had very little unburnt habitat within their post-fire home ranges and all succumbed to predation within the first few days post-fire, one collar failed to collect any fixes, and another slipped off the animal several days after collar attachment.
The best performing model included ‘prop_burned’, which had a coefficient estimate with confidence intervals only marginally overlapping zero (Table 4, β = -1.54, 95% CI: -3.16–0.07). This model suggested P. tridactylus were more likely to avoid recently burnt areas as more of their pre-fire home range was burnt. Animals with small proportions of their pre-fire range burnt tended to be attracted to recently burnt areas, albeit with some uncertainty (Fig. 3).

Integrated resource selection function (iRSF) coefficients of selection for burnt areas after fire for each individual. Estimates above 0 (dotted-line) suggest attraction to burnt areas and estimates below 0 suggest avoidance of burnt areas. 95% confidence intervals around each estimate describe uncertainty. Intervals that do not overlap 0 suggest statistical significance (in bold). The trend line is the estimated population level resource selection for burnt areas as a function of burn exposure as predicted by meta-regression models alongside 95% confidence intervals (grey ribbon)
Discussion
Prescribed fire continues to be frequently used by land managers to reduce wildfire risk across the globe (Fernandes and Botelho 2003), yet its impact on the survival and movement behavior of wildlife is often poorly known. Here we present a novel approach to describe the immediate impacts of prescribed fire on a nationally threatened mammal – a first for this species. We combined fine-scale movement data and continuous-time movement models to describe the movement behavior and survival of P. tridactylus across a large gradient of exposure to prescribed fire (defined as the proportion of their pre-fire home range that burnt). We then compared these movement behaviors and survival rates to animals with no fire exposure tracked for a similar duration. In the first month post-fire, P. tridactylus exhibited high site fidelity and continued to traverse a similar area each day. Animals with smaller proportions of their range burnt were more likely to survive the first month post-fire, increase their home range size if they were male and select for recently burnt areas. In contrast, animals with larger proportions of their range burnt were more likely to die, reduce their home range size, and avoid burnt areas. These results broadly align with other studies investigating the response of P. tridactylus and other mycophagous mammals to prescribed fire using other techniques elsewhere (e.g. live-trapping or camera trapping; Christensen 1980; Hope 2012; McHugh et al. 2020). Our results suggest burnt areas may be particularly dangerous environments for P. tridactylus in the first month post-fire.
P. tridactylus survival post fire was generally poor. Consistent with a recent meta-analysis of animal survival during fire, there was no evidence that fire directly killed any animals in our study (Jolly et al. 2022). In contrast, we have strong evidence to suggest foxes were responsible for several post-fire mortality events. Three collared P. tridactylus were found cached underground in burnt areas (a characteristic red fox behavior (Macdonald 1977) and concurrent camera trap monitoring frequently detected foxes and cats in recently burnt areas, including a fox with a P. tridactylus in its mouth (Le Pla, unpublished data). The relative contribution of feral cat predation to the elevated post-fire mortality rates we observed is unclear.
High site fidelity likely contributed to the elevated post-fire mortality rates we observed. Limited changes in home range location for all animals exposed to fire suggests P. tridactylus perceive that they can continue to find adequate food and shelter resources in the same area they occupied before fire, or that the perceived risks of leaving are too high. Whilst animals often chose to persist in the same location, there was some evidence that animals may reduce their home range size after fire, especially when large proportions of their home range burnt. Site fidelity theory predicts that extreme site fidelity (‘always stay’ Switzer 1993) is an optimal behavior in unpredictable habitats where there is little variation in site quality. Concurrent camera trapping in our region suggests P. tridactylus are homogenously distributed at a broad scale (Le Pla, unpublished data) and consequently, it is likely most habitat immediately adjacent to burnt areas was already occupied. Although there is little evidence P. tridactylus are territorial in our study area (Le Pla et al. 2023), it is possible individuals may have encountered each other more frequently in the immediate post-fire period and these interactions could have been aggressive. Conspecific encounter rates may be particularly elevated for animals with high fire exposure as these animals were more likely to restrict their activity to the unburnt vegetation available to them (Fig. 3). Unfortunately, the GPS fix schedule we employed (hourly) was too coarse to describe the nature of interactions between individuals after fire, or quantify encounter rates directly. Additionally, the high structural complexity that characterizes our study region may limit the ability of P. tridactylus to assess the availability and quality of sites beyond their home range. Complex vegetation structure can limit the perceptual range of animals (Lima and Zollner 1996; Nimmo et al. 2019), which may increase the perceived risks of abandoning their home range and make this option less attractive after fire.
For a mycophagous mammal like P. tridactylus, fire may not necessarily reduce food availability. Our study was conducted during a season where fungi ordinarily make up over 50% of P. tridactylus diet (Bennett and Baxter 1989). Fungi respond to fire in a myriad of ways, with some fungi growing quickly in recently disturbed areas (Claridge et al. 2009; Kouki and Salo 2020). The husk remains of Mesophellia sp., a hypogeal fungus, are often found in recently burnt areas where P. tridactylus (this study; Bennett and Baxter 1989) and other mycophaeous mammals reside (Johnson 1997; Vernes and Haydon 2001). Whilst the influence of fire on the growth rate Mesophellia sp. is unclear, fire can fundamentally alter its odor profile and potentially make it easier to detect (Millington et al. 1997). Fungi may also become more accessible post-fire due to the loss of surface vegetation (Claridge and Trappe 2004). Similar to the “magnet” effect, where many herbivores are attracted to new growth after fire (Archibald, Stock and Fairbanks 2005), our results suggest recently burnt areas could be attractive to P. tridactylus if fungi are more detectable, accessible or abundant and thus, may facilitate high post-fire site fidelity.
In contrast, by removing understory vegetation, fire likely reduced the general availability of shelter sites for P. tridactylus. Like many ground-dwelling mammals (Petit and Frazer 2023), P. tridactylus often shelter in the thick flammable “skirt” of the Austral Grass tree (Xanthorrhoea australis) (Le Pla, pers obs.) and positive relationships between P. tridactylus and complex habitat have been detected elsewhere (Swan et al. 2015; Hradsky et al. 2017). Grass-trees can buffer ground-dwelling animals against temperature extremes and rain, and their dense growth form likely provides valuable refuge from predators (Petit and Frazer 2023). However behavioral segmentation suggested P. tridactylus often have several resting sites within their home range (Le Pla, unpublished data, Figure S8). If the unburnt habitat remaining within an individual’s post-fire home range continued to support areas of high structural complexity, then shelter sites may not have been limiting after fire.
In addition to high site fidelity, high post-fire diffusion rates suggested P. tridactylus made frequent forays into recently burnt areas. These forays likely increased encounter rates between P. tridactylus and their predators, particularly if these predators preferentially used recently burnt areas (Hradsky 2020; Doherty et al. 2022). However, contrasting selection for burnt areas along a gradient of fire exposure may reflect differences in the motivations of P. tridactylus entering burnt areas. When small proportions of their home range burnt, P. tridactylus tended to be attracted to recently burnt areas, presumably to forage. In contrast, animals with large proportions of their home range burnt tended to restrict their activity to the unburnt patches remaining within their range. These animals likely entered burnt areas primarily to move between the unburnt patches remaining available to them in their range. This contrasting selection for burnt areas suggests there may be a threshold of burn exposure wherein the perceived benefits of exploiting a burnt area for foraging are eventually outweighed by a heightened perceived predation risk as burnt areas comprise more of their home range.
Fire may fundamentally alter the environmental context in which predators and prey encounters occur (Doherty et al. 2022). The high mortality rates we observed indicate P. tridactylus may not be accurately assessing predation risk when moving within recently burnt areas. The simplification of vegetation complexity post-fire may make it easier for predators and prey to detect each other visually (Jaffe and Isbell 2009; Cherry et al. 2018). However, fire can also impact the olfactory and acoustic cues many ground-dwelling mammals rely upon to ‘eavesdrop’ on local predators (Doherty et al. 2022; Price et al. 2022; Michel et al. 2023). Fire may have temporarily eliminated the olfactory cues (e.g. scat, urine, hair etc.) P. tridactylus use to assess predation risk, and the simplified vegetation in burnt areas may have made it challenging to assess imminent predation risk acoustically.
The high site fidelity and attraction or indifference to burnt areas we observed in P. tridactylus is typical of many mycophagous mammals in Australia (Johnson 1997; Vernes and Haydon 2001; MacGregor et al. 2012). For example, other mycophagous mammals in our study area such as the southern brown bandicoot (Isoodon obesulus) and long-nosed bandicoot (Perameles nasuta) continued to be detected in burnt areas on concurrent camera trap surveys after fire (Le Pla, unpublished data). Similarly, both the northern bettong (Bettongia tropica) and eastern bettong (Bettongia giamardi) displayed a willingness to exploit recently burnt areas for foraging, and showed little change in home range size, site fidelity and movement rates after fire (Johnson 1997; Vernes and Haydon 2001; Vernes and Pope 2001).
As cursorial predators with high movement rates and flexible hunting behaviors, red foxes may be particularly effective hunters in recently burnt open vegetation (Saunders et al. 2010; Hradsky 2020). Although it has not been robustly tested, survival rates of mycophagous mammals like P. tridactylus after fire appear to be markedly different in areas with and without foxes. For example, mycophagous mammal survival post-fire was generally high in areas with few foxes (Johnson 1995; Vernes 2000; McHugh et al. 2020) or where foxes undergo lethal control (Hope 2012; Robley et al. 2016). In contrast, where foxes were uncontrolled or where their behavioral response to fire is unconstrained by the presence of larger predators (e.g. Dingoes, Canis lupus dingo), mycophagous mammal survival and persistence post-fire was poor (this study, Christensen 1980; Robley et al. 2016). Therefore, it is possible that recently burnt areas may also reduce survival rates of other mycophagous mammals exposed to prescribed fire elsewhere, particularly where foxes are abundant. Like P. tridactylus, many of these mycophgeous species are also considered species of conservation concern by Australia’s flagship environmental legislation, the Environment Protection and Biodiversity Conservation Act 1999 (Australian Government 1999).
Our results are likely conditional on study season and duration. Burns took place during winter (April – September), coinciding with a period where fungal fruiting-bodies feature prominently in P. tridactylus diet (Bennett and Baxter 1989). Additionally, red foxes primarily breed during late winter (McIntosh 1963). Fires in different seasons may elicit different responses and survival rates for P. tridactylus, if they burn at higher intensities, have different effects on food availability, or coincide with other periods of fox behavior (e.g. autumn is a period of high fox mobility as offspring disperse). The battery life of GPS collars also constrained our ability to monitor P. tridactylus for longer than the first month after fire and assess whether fire continued to impact the survival of P. tridactylus beyond this time. It is possible that both predators and prey are only attracted to recently burnt areas for a short period. Thus, whilst burnt areas may act as ecological traps (Kristan 2003), the high fecundity and generally homogeneous distribution of P. tridactylus in our study area may mean long-term population viability is not necessarily compromised by prescribed fire. Nonetheless, our study contributes to a growing body of evidence suggesting the threatening processes of fire and invasive predators may interact and contribute to the ongoing collapse of native mammal communities across Australia (Woinarski et al. 2015).
Our results provide a sound basis for several management actions. First, given the generally poor survival of animals with high fire exposure, we recommend fire practitioners attempt to limit P. tridactylus exposure to fire by applying multiple, smaller burns each surrounded by unburnt vegetation. This would produce fire scars that are smaller and more interspersed. This could be achieved by using methods that allow for dispersed lighting patterns across large areas near simultaneously- such as incendiaries delivered via helicopter and fixed wing aircraft. However, we acknowledge the operational feasibility of such an approach will likely vary from region to region. Exploring the influence of such an approach on long-term wildfire risk thoroughly via simulation (e.g. Penman et al. 2014) is required before adopting this practice more generally. Second, given the high site fidelity and high diffusion rates of P. tridactylus, we recommend fire practitioners attempt to retain patches of unburnt, structurally complex vegetation to provide access to suitable shelter sites. Finally, we suggest P. tridactylus may benefit from the integration of predator control and prescribed burning operations. A temporary reduction in fox activity and abundance prior to or immediately after fire may improve native mammal survival, however additional studies are needed to determine the feasibility and appropriate scale of this intervention.
By combining fine-scale movement data with continuous-time movement models, we have intimately described how a vulnerable mammal responds to prescribed fire. Our results suggest P. tridactylus are at an elevated risk of predation shortly after prescribed fire, particularly if most of their home range is burnt. High diffusion rates, high site fidelity and continued use of recently burnt areas may promote high encounter rates between P. tridactylus and their predators, particularly if these predators are also selectively hunting in recently burnt areas (Hradsky et al. 2020; Doherty et al. 2022). Prescribed burning will continue to be an important land and fire management tool, particularly in a warming climate (Morgan et al. 2020; Jones et al. 2022). In flammable habitats like our study area, burning in warmer seasons remains challenging and burning in the cooler months may be necessary to achieve wildfire risk reduction safely. However, our results suggest prescribed fires during this time may reduce survival rates of P. tridactylus, a nationally threatened ground-dwelling mammal. P. tridactylus and other threatened mycophagous mammals may benefit from increased patchiness of planned burns and the integration of invasive predator control and prescribed burning, particularly when fires coincide with periods of high fungi growth and availability.
Data availability
The data that support this study will be shared upon reasonable request to the corresponding author. In addition, movement data analyzed during the current study will be made available in the Movebank online data repository.
References
Abrahms B, Aikens EO, Armstrong JB, Deacy WW, Kauffman MJ, Merkle JA (2021) Emerging perspectives on resource tracking and animal movement ecology. Trends Ecol Evol 36(4):308–320
Allen AM, Singh NJ (2016) Linking movement ecology with wildlife management and conservation. Front Ecol Evol 3:155
Alston JM, Fleming CH, Kays R, Streicher JP, Downs CT, Ramesh T, Reineking B, Calabrese JM (2023) Mitigating pseudoreplication and bias in resource selection functions with autocorrelation-informed weighting. Methods Ecol Evol 14(2):643–654
Archibald S, Bond WJ (2004) Grazer movements: spatial and temporal responses to burning a tall-grass African savanna. Int J Wildland Fire 13(3):377–385
Arnold TW (2010) Uninformative parameters and model selection using Akaike’s Information Criterion. J Wildl Manage 74(6):1175–1178
Arthur AD, Catling PC, Reid A (2012) Relative influence of habitat structure, species interactions and rainfall on the post-fire population dynamics of ground-dwelling vertebrates. Austral Ecol 37(8):958–970
Australian Government (1999) Environment protection and biodiversity conservation act. Department of Energy and Environment, Commonwealth of Australia
Bennett AF (1993) Microhabitat use by the long-nosed potoroo, Potorous tridactylus, and other small mammals in remnant forest vegetation, south-western Victoria. Wildl Res 20(3):267–285
Bennett AF, Baxter BJ (1989) Diet of the long-nosed potoroo, Potorous-Tridactylus (Marsupialia, Potoroidae), in southwestern Victoria. Wildl Res 16(3):263–271
Bhattacharyya A (1943) On a measure of divergence between two statistical populations defined by their probability distribution. Bull Calcutta Math Soc 35:99–109
Bond WJ, Keeley JE (2005) Fire as a global ‘herbivore’: the ecology and evolution of flammable ecosystems. Trends Ecol Evol 20(7):387–394
Boyce MS, McDonald LL (1999) Relating populations to habitats using resource selection functions. Trends Ecol Evol 14(7):268–272
Burbidge AA, McKenzie NL (1989) Patterns in the modern decline of Western Australia’s vertebrate fauna: causes and conservation implications. Biol Conserv 50(1–4):143–198
Bureau of Meteorology (2020) Climate data online. http://www.bom.gov.au/climate/data/index.shtml [Accessed February 2024]
Burnham KP, Anderson DR (2002) Model selection and multi-model inference: apractical information-theoretic approach. Second edition. Springer-Verlag, New York,NY, USA
Burt WH (1943) Territoriality and home range concepts as applied to mammals. J Mammal 24(3):346–352
Calabrese JM, Fleming CH, Gurarie E (2016) Ctmm: an R package for analyzing animal relocation data as a continuous-time stochastic process. Methods Ecol Evol 7(9):1124–1132
Cherry MJ, Chandler RB, Garrison EP, Crawford DA, Kelly BD, Shindle DB, Godsea KG, Miller KV, Conner LM (2018) Wildfire affects space use and movement of white-tailed deer in a tropical pyric landscape. For Ecol Manag 409:161–169
Christensen PES (1980) The biology of Bettongia penicillata Gray, 1837, and Macropus eugenii (Desmarest, 1817) in relation to fire. Bulletin no. 91. Forests Department of Western Australia
Claridge AW, Barry SC (2008) Factors influencing the distribution of medium-sized ground-dwelling mammals in southeastern mainland Australia. Austral Ecol 25(6):676–688
Claridge AW, Trappe JM, Lunney D (2004) Managing habitat for mycophagous (fungus-feeding) mammals: a burning issue. In the Conservation of Australia’s forest fauna (second edition), pp.936–946. Royal Zoological Society of New South Wales, Mosman, NSW, Australia
Claridge AW, Seebeck JH, Rose R (2007) Bettongs, potoroos and the musky rat-kangaroo. CSIRO PUBLISHING
Claridge AW, Trappe JM, Mills DJ, Claridge DL (2009) Diversity and habitat relationships of hypogeous fungi. III. Factors influencing the occurrence of fire-adapted species. Mycol Res 113(6–7):792–801
R Core Team (2023) R: A Language and Environment for Statistical Computing. R Foundation for Statistical Computing, Vienna, Austria. https://www.R-project.org/
Doherty TS, Hays GC, Driscoll DA (2021) Human disturbance causes widespread disruption of animal movement. Nat Ecol Evol 5(4):513–519
Doherty TS, Geary WL, Jolly CJ, Macdonald KJ, Miritis V, Watchthorn DF, Cherry MJ, Conner LM, González TM, Legge SM, Ritchie EG (2022) Fire as a driver and mediator of predator-prey interactions. Biol Rev 97(4):1539–1558
Driessen MM, Jarman PJ, Visoiu M, Dewar E (2021) Mammal responses to moderate-intensity planned burning in a small, isolated woodland reserve. Wildl Res 48:561–576
Elliott TF, Truong C, Jackson SM, Zúñiga CL, Trappe JM, Vernes K (2022) Mammalian mycophagy: a global review of ecosystem interactions between mammals and fungi. Fungal Syst Evol 9(1):99–159
Fernandes PM, Botelho HS (2003) A review of prescribed burning effectiveness in fire hazard reduction. Int J Wildland Fire 12:117–128
Fleming CH, Calebrese JM (2023) ctmm: Continuous-Time Movement Modelling. R package version 1.2.1
Fleming CH, Fagan WF, Mueller T, Olson KA, Leimgruber p, Calabrese JM (2015) Rigorous home range estimation with movement data: a new autocorrelated kernel density estimator. Ecology 96(5):1182–1188
Fleming CH, Sheldon D, Fagan WF, Leimgruber P, Mueller T, Nandintseteg D, Noonan MJ, Olson KA, Setyawan E, Sianipar A, Calabrese JM (2018) Correcting for missing and irregular data in home-range estimation. Ecol Appl 28(4):1003–1010
Fordyce A, Hradsky BA, Ritchie EG, Di Stefano J (2016) Fire affects microhabitat selection, movement patterns, and body condition of an Australian rodent (Rattus fuscipes). J Mammal 97(1):102–111
Gazzard T, McLeod C, Medlin C, Baker M, Martin H, Rourke D (2020) Barwon South West Bushfire Management Strategy 2020. The State of Victoria Department of Environment, Land, Water and Planning 2020
Gill R, Serrouya R, Calvert AM, Ford A, Steenweg R, Noonan MJ (2023) Movement ecology of endangered caribou during a COVID-19 mediated pause in winter recreation. Animal Conservation
Haslem A, Leonard SW, Bruce MJ, Christie F, Holland GJ, Kelly LT, MacHunter J, Bennett AF, Clarke MF, Yorke A (2016) Do multiple fires interact to affect vegetation structure in temperate eucalypt forests? Ecol Appl 26(8):2414–2423
Herzog NM, Parker CH, Keefe ER, Coxworth J, Barrett A, Hawkes K (2014) Fire and home range expansion: a behavioral response to burning among savanna dwelling vervet monkeys (Chlorocebus aethiops). Am J Phys Anthropol 154(4):554–560
Higgins SI, Bond WJ, February EC, Bronn A, Euston-Brown DI, Enslin B, Govender N, Rademan L, O’Regan S, Potgieter AL, Scheiter S (2007) Effects of four decades of fire manipulation on woody vegetation structure in savanna. Ecology 88(5):1119–1125
Hope B (2012) Short-term response of the long-nosed bandicoot, Perameles nasuta, and the southern brown bandicoot, Isoodon obesulus obesulus, to low-intensity prescribed fire in heathland vegetation. Wildl Res 39(8):731–744
Hradsky BA (2020) Conserving Australia’s threatened native mammals in predator-invaded, fire-prone landscapes. Wildl Res 47(1):1–15
Hradsky BA, Mildwaters C, Ritchie EG, Christie F, Di Stefano J (2017) Responses of invasive predators and native prey to a prescribed forest fire. J Mammal 98(3):835–847
Jaffe KE, Isbell LA (2009) After the fire: benefits of reduced ground cover for vervet monkeys (Cercopithecus aethiops). Am J Primatology: Official J Am Soc Primatologists 71(3):252–260
Johnson CN (1996) Interactions between mammals and ectomycorrhizal fungi. Trends Ecol Evol 11(12):503–507
Johnson CN (1997) Fire and habitat management for a mycophagous marsupial, the tasmanian bettong Bettongia gaimardi. Aust J Ecol 22(1):101–105
Johnston PG (2008) Long-nosed potoroo. The Mammals of Australia’. (Eds S. van Dyck and RJ Strahan.) pp. 302–304
Jolly CJ, Dickman CR, Doherty TS, van Eeden LM, Geary WL, Legge SM, Woinarski JC, Nimmo DG (2022) Animal mortality during fire. Glob Change Biol 28(6):2053–2065
Jones MW, Abatzoglou JT, Veraverbeke S, Andela N, Lasslop G, Forkel M, Smith AJ, Burton C, Betts RA, van der Werf GR, Sitch S (2022) Global and regional trends and drivers of fire under climate change. Rev Geophys 60(3):e2020RG000726
Kouki J, Salo K (2020) Forest disturbances affect functional groups of macrofungi in young successional forests–harvests and fire lead to different fungal assemblages. For Ecol Manag 463:p118039
Le Pla M, Hradsky BA, Di Stefano J, Farley-Lehmer TC, Birnbaum EK, Pascoe JH (2023) Movement and ranging behaviour of long-nosed potoroos (Potorous tridactylus) in south-west Victoria, Australia. Wildl Res 51(1):WR23013
Lima SL, Zollner PA (1996) Towards a behavioral ecology of ecological landscapes. Trends Ecol Evol 11:131–135
Lüdecke D, Ben-Shachar MS, Patil I, Waggoner P, Makowski D (2021) performance: An R package for assessment, comparison and testing of statistical models. Journal of Open Source Software, 6(60)
Macdonald DW (1977) On food preference in the red fox. Mammal Rev 7:7–23
MacGregor CI, Wood JT, Dexter N, Lindenmayer DB (2012) Home range size and use by the long-nosed bandicoot (Perameles nasuta) following fire. Australian Mammalogy 35(2):206–216
MacGregor-Fors I, Payton ME (2013) Contrasting diversity values: statistical inferences based on overlapping confidence intervals. PLoS ONE : e56794
McGregor HW, Legge S, Jones ME, Johnson CN (2016) Extraterritorial hunting expeditions to intense fire scars by feral cats. Sci Rep 6(1):22559
McHugh D, Goldingay RL, Parkyn J, Goodwin A, Letnic M (2020) Short-term response of threatened small macropods and their predators to prescribed burns in subtropical Australia. Ecol Manag Restor 21(2):97–107
McIntosh D (1963) Reproduction and growth of the fox in the Canberra district. Wildl Res 8(2):132–141
Meers BT, Adams R (2003) The impact of grazing by Eastern Grey Kangaroos (Macropus giganteus) on vegetation recovery after fire at Reef Hills Regional Park, Victoria. Ecol Manag Restor 4(2):126–132
Michel A, Johnson JR, Szeligowski R, Ritchie EG, Sih A (2023) Integrating sensory ecology and predator-prey theory to understand animal responses to fire. Ecol Lett 26(7):1050–1070
Millington S, Leach DN, Wyllie SG, Claridge AW (1997) Aroma profile of the Australian truffle-like fungus Mesophellia glauca. Pp 331–342 in Flavor Analysis - Developments in Isolation and Characterisation, edited by C.J. Mussinana and M.J. Morello. American Chemical Society, Washington D.C
Morales JM, Moorcroft PR, Matthiopoulos J, Friar JL, Kie JG, Powell RA, Merrill EH, Haydon DT (2010) Building the bridge between animal movement and population dynamics. Philosophical Trans Royal Soc B: Biol Sci 365(1550):2289–2301
Morgan GW, Tolhurst KG, Poynter MW, Cooper N, McGuffog T, Ryan R, Wouters MA, Stephens N, Black P, Sheehan D, Leeson P (2020) Prescribed burning in south-eastern Australia: history and future directions. Australian Forestry 83(1):4–28
Nathan R, Getz WM, Revilla E, Holyoak M, Kadman R, Saltz D, Smouse PE (2008) A movement ecology paradigm for unifying organismal movement research. Proc Natl Acad Sci 105(49):19052–19059
Nest C, Elliott TF, Cooper T, Vernes K (2023) Seasonal consumption of mycorrhizal fungi by a marsupial-dominated mammal community. Fungal Ecol 64:p101247
Nimmo DG, Avitabile S, Banks SC, Bliege Bird R, Callister K, Clarke MF, Dickman CR, Doherty TS, Driscoll DA, Greenville AC, Haslem A (2019) Animal movements in fire-prone landscapes. Biol Rev 94(3):981–998
Noonan MJ, Tucker MA, Fleming CH, Akre TS, Alberts SC, Ali AH, Altmann J, Antunes PC, Belant JL, Beyer D, Blaum N (2019) A comprehensive analysis of autocorrelation and bias in home range estimation. Ecol Monogr 89(2):e01344
Norton MA, French K, Claridge AW (2011) Habitat associations of the long-nosed potoroo (Potorous tridactylus) at multiple spatial scales. Australian J Zool 58(5):303–316
Novellie PA (1978) Comparison of the foraging strategies of blesbok and springbok on the Transvaal highveld. South Afr J Wildl Res – 24 Month Delayed open Access 8(4):137–144
Penman TD, Bradstock RA, Price OF (2014) Reducing wildfire risk to urban developments: Simulation of cost-effective fuel treatment solutions in south eastern Australia. Environ Model Softw 52:166–175
Petit S, Frazer DS (2023) The role of grass-tree xanthorrhoea semiplana (Asphodelaceae) canopies in temperature regulation and waterproofing for ground-dwelling wildlife. Pac Conserv Biology 29(5):445–455
Phillips RJ, Waldrop TA (2008) Changes in vegetation structure and composition in response to fuel reduction treatments in the South Carolina Piedmont. For Ecol Manag 255(8–9):3107–3116
Picardi S, Coates P, Kolar J, O’Neil S, Matthews S, Dahlgren D (2022) Behavioural state-dependent habitat selection and implications for animal translocations. J Appl Ecol 59(2):624–635
Povh LF, Willers N, Shephard JM, Fleming PA (2023) A conservation-significant threatened mammal uses fire exclusions and shifts ranges in the presence of prescribed burning. Int J Wildland Fire 32(9):1291–1303
Price C, McArthur C, Norbury G, Banks P (2022) Olfactory misinformation: creating fake news to reduce problem foraging by wildlife. Front Ecol Environ 20(9):531–538
Rettie WJ, Messier F (2000) Hierarchical habitat selection by woodland caribou: its relationship to limiting factors. Ecography 23(4):466–478
Robertson BA, Rehage JF, Sih A (2013) Ecological novelty and the emergence of evolutionary traps. Trends Ecol Evol 28:552–560
Robley A, Moloney P, Moxham C, Neave G, Friend GR, Fraser I (2016) The effects of interaction between planned burning and foxes on the presence of native mammals. Arthur Rylah Institute for Environmental Research
Saunders GR, Gentle MN, Dickman CR (2010) The impacts and management of foxes Vulpes vulpes in Australia. Mammal Rev 40(3):181–211
Sergio F, Blas J, Hiraldo F (2018) Animal responses to natural disturbance and climate extremes: a review. Glob Planet Change 161:28–40
Suraci JP, Smith JA, Chamaillé-Jammes S, Gaynor KM, Jones M, Luttbeg B, Ritchie EG, Sheriff MJ, Sih A (2022) Beyond spatial overlap: harnessing new technologies to resolve the complexities of predator–prey interactions. Oikos 8:e09004
Swan M, Christie F, Sitters H, York A, Di Stefano J (2015) Predicting faunal fire responses in heterogeneous landscapes: the role of habitat structure. Ecol Appl 25(8):2293–2305
Switzer PV (1993) Site fidelity in predictable and unpredictable habitats. Evol Ecol 7:533–555
Valentine LE, Bretz M, Ruthrof KX, Fisher R, Hardy GESJ, Fleming PA (2017) Scratching beneath the surface: Bandicoot bioturbation contributes to ecosystem processes. Austral Ecol 42(3):265–276
van Moorter B, Bunnefeld N, Panzacchi M, Rolandsen CM, Solberg EJ, Saether B (2013) Understanding scales of movement: animals ride waves and ripples of environmental change. J Anim Ecol 82(4):770–780
Vernes K (2000) Immediate effects of fire on survivorship of the northern bettong (Bettongia tropica): an endangered Australian marsupial. Biol Conserv 96(3):305–309
Vernes K, Haydon DT (2001) Effect of fire on northern bettong (Bettongia tropica) foraging behaviour. Austral Ecol 26(6):649–659
Vernes K, Pope LC (2001) Stability of nest range, home range and movement of the northern bettong (Bettongia tropica) following moderate-intensity fire in a tropical woodland, north-eastern Queensland. Wildl Res 28(2):141–150
Viechtbauer W (2010) Conducting meta-analyses in R with the metafor package. J Stat Softw 36(3):1–48
Williams S.H., Steenweg R, Hegel T, Russell M, Hervieux D, Hebblewhite M (2021) Habitat loss on seasonal migratory range imperils an endangered ungulate. Ecol Solutions Evid 2(1):e12039
Winkler DW, Jørgensen C, Both C, Houston AI, McNamara JM, Levey DJ, Partecke J, Fudickar A, Kacelnik A, Roshier D, Piersma T (2014) Cues, strategies, and outcomes: how migrating vertebrates track environmental change. Movement Ecology, 2, pp. 1–15
Winner K, Noonan MJ, Fleming CH, Olson KA, Mueller T, Sheldon D, Calabrese JM (2018) Statistical inference for home range overlap. Methods Ecol Evol 9(7):1679–1691
Woinarski JC, Burbidge AA, Harrison PL (2015) Ongoing unraveling of a continental fauna: decline and extinction of Australian mammals since European settlement. Proc Natl Acad Sci 112(15):4531–4540
Yarnell RW, Metcalfe DJ, Dunstone N, Burnside N, Scott DM (2008) The impact of fire on habitat use by the short-snouted elephant shrew (Elephantulus Brachrhynchus) in North West Province, Southern Africa. Afr Zool 41(1):45–52
Yibarbuk D, Whitehead PJ, Russell-Smith J, Jackson D, Godjuwa C, Fisher A, Cooke P, Choquenot D, Bowman DM (2001) Fire ecology and Aboriginal land management of central Arnhem Land, northern Australia: a tradition of ecosystem management. J Biogeogr 28(3):325–343
Acknowledgements
We pay our respects to the Gadabanut, the traditional custodians of the lands upon which this study took place. We thank L. Bonifacio, E. Brooks, J. D Eizenberg, L. Falls, E. Fitzsimmons, S. Girvan, M. Goodie, E. Hinde, C. Khanal, E. Liddell, M. Loeffler, R. Loosveld, E. Mckenzie, D. Ng Sing Kwong, I. Swart, and J. Templeton for their assistance with fieldwork. This project was supported by the Hermon Slade Foundation, Parks Victoria, the Victorian Government’s Department of Energy, Environment, and Climate Change, and the Australian Federal Government.
Funding
This project was supported by the Hermon Slade Foundation, the Conservation Ecology Centre, Parks Victoria, the Corangamite Catchment Authority, the Department of Energy, Environment, and Climate Action, the Australian Federal Government’s “Wild Otway’s Initiative”, and the Australian Research Council Grant/Award Number: LP170101134.
Open Access funding enabled and organized by CAUL and its Member Institutions
Author information
Authors and Affiliations
Contributions
Mark Le Pla developed ideas, led the data collection, performed the statistical analysis and manuscript writing. Bronwyn Hradsky developed ideas, assisted in statistical analysis and manuscript writing. Julian Di Stefano developed ideas, assisted in statistical analysis and manuscript writing. Tamika Farley-Lehmer and Emma Birnbaum assisted in data collection. Jack Pascoe developed ideas, assisted in data collection and manuscript writing. All authors read and approved the final manuscript.
Corresponding author
Ethics declarations
Ethical approval
All research activities were undertaken under wildlife research permit number 10009226, wildlife and small institutions ethics approval (SPFL20096), and University of Melbourne ethics approval (2022-25286-35215).
Competing interests
The authors declare no competing interests.
Additional information
Communicated by David Hawksworth.
Publisher’s note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Electronic supplementary material
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article’s Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article’s Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Pla, M.L., Hradsky, B.A., Di Stefano, J. et al. High site fidelity and reduced survival of a mycophagous mammal after prescribed fire. Biodivers Conserv (2024). https://doi.org/10.1007/s10531-024-02927-5
Received:
Revised:
Accepted:
Published:
DOI: https://doi.org/10.1007/s10531-024-02927-5