
Abstract
Climate and land management affect nutrient cycling in grassland ecosystems. We aimed to understand whether temperate and tropical grasslands differ in terms of soil organic carbon (SOC), nitrogen (N), and phosphorus (P) concentrations, and their C:N:P stoichiometric ratios in grazed and ungrazed natural grasslands and pastures. For this, we used a meta-analysis approach (1296 records, 241 papers), and regression models to explain the observed patterns in terms of mean annual precipitation (MAP), mean annual temperature (MAT), altitude, and latitude. SOC, N, and P concentrations were higher in temperate regions than in tropical ones, and they negatively correlated with MAT and MAP. The grassland type effect was more significant for tropical regions. In tropical regions, soil C:N ratios were higher in ungrazed than in grazed pastures, and soil N:P ratios in ungrazed sites were higher in pastures than in natural grasslands. Grazing increases soil N and SOC for natural grasslands in temperate regions. Our findings suggest that soil stoichiometric C:N:P stoichiometric signatures in grasslands differed between tropical and temperate regions on a global scale. P is a key element in regulation and restriction on soil C and N cycling in tropical regions but less in the temperate ones. Our findings suggest the direction of effects of grazing or grassland type on C:N:P stoichiometric signature. Since imbalances in soil stoichiometric ratios may have implications for ecosystem functioning, the assessment of these patterns could serve as a valuable tool for management and conservation of grasslands and pastures in both tropical and temperate regions.
Similar content being viewed by others


Explore related subjects
Discover the latest articles, news and stories from top researchers in related subjects.Avoid common mistakes on your manuscript.
Introduction
Carbon (C), nitrogen (N), and phosphorus (P) have complex global biogeochemical cycles that are strongly coupled (Ågren 2008). N and P synergistically affect many ecosystem processes as growth-limiting factors for organisms (Sterner and Elser 2002; Elser et al. 2007). The relationship between plant-available N and P varies widely among soils depending on the mineralogy, organic matter forms, climate, weathering, and other factors (Amundson 2021). Ratios between elements may provide additional information to ecosystem functioning related to that obtained with the individual concentrations of each element (Binkley and Fisher 2020). These elemental ratios (stoichiometric ratios) and their quantification in the environment reflect the impact of living organisms and can be considered a stoichiometric signature (Redfield 1958; Reiners 1986; Sterner and Elser 2002; Butler et al. 2021; Chang et al. 2021; Kempes et al. 2021). A chemical signature is defined as a pattern of elemental or molecular composition that has been employed for characterizing or tracking living organisms or tissues based on their composition or biochemical processes (Sterner and Elser 2002), or as evidence of life forms in astrobiology (Chan et al. 2019). For the purposes of this study, we designate the stoichiometric signature as a pattern in the ratios of chemical elements within the soil, with the intention of tracking the effects of environmental and management factors on grasslands soil. The importance of using this approach in living organisms is that despite the variability between the element concentrations, the ratios maintain a lower variation interval (e.g., Reiners 1986; Sterner and Elser 2008; Arhonditsis et al. 2019). This lower variability has also been reported for C:N:P ratios in grasslands soil on a global scale (Cleveland and Liptzin 2007). Stoichiometric signature is based on elemental mass ratios and depends on cycling elements and ecosystem processes (Reiners 1986; Sterner and Elser 2002; Butler et al. 2021). Quantifying the C:N:P ratios in soil can be a powerful tool to understand the influence of environmental variables and management on soil nutrients concentration.
Over the last two decades, there have been global and regional syntheses on the status of C and nutrient concentrations in grassland soils. Some studies have explored the effect of management, including grazing, mainly in soil C in grasslands at global scale (Conant et al. 2001, 2017; Pineiro et al. 2009; McSherry and Ritchie 2013), and some include effects in other nutrients (Abdalla et al. 2018; Rumpel et al. 2015; Byrnes et al. 2018; He et al. 2020). Other reviews have examined the concentrations and ratios of C and N in grassland soils comparing temperate and tropical zones (Pärtel et al. 2008), encompassing both natural grasslands and pastures (Xu et al. 2013), or only tropical ecosystems (Joergensen 2010). Consequently, it is well-established that soil nutrient transformation and cycling in grasslands are contingent upon climate and other factors, such as grazing (Pärtel et al. 2008). However, most of these studies predominantly explore the influence of individual factors (grassland type, grazing, or climate individually) on the concentrations of C and nutrients, such as N and P, as well as their respective ratios. Until now, only a limited number of studies have comprehensively investigated the simultaneous impact of multiple factors on C, N, and P concentrations and ratios, concurrently considering climatic effects (Pärtel et al. 2008; Abdalla et al. 2018). One key objective of this study is to elucidate comprehensive global stoichiometric patterns of C-N-P in soil, encompassing the contrast between temperate and tropical climate zones, in addition to the influences of temperature, precipitation, geographic location, and the altitude of grassland-associated ecosystems.
Grasslands are distributed in different climatic zones of the world (Blair et al. 2014; ILRI et al. 2021). For this study, we classified sites as natural grasslands (native grasslands, prairies, savannas) and pastures (cultivated and induced grasslands). Our analyses distinguished between temperate and tropical zones for all these classifications. Most pastures have been established in the equatorial and warm temperate zones, in regions corresponding to zone A (i.e., humid tropical) and C (i.e., temperate) in the Köppen-Geiger classification (Humphreys 1981; Suttie et al. 2005; Ramankutty et al. 2008; Reinermann et al. 2020). Several distinctions between grasslands in these two climatic zones can be highlighted. Temperate grasslands are predominantly characterized by C3 plants, while tropical grasslands are dominated by C4 grasses (Partel et al. 2008; Lehmann and Parr 2016). Soil organic matter (SOM) transformation processes occur more rapidly in tropical regions (van Keulen 2001; Nortcliff 2010), primarily driven by temperature and precipitation (Voroney and Heck 2015; Mitchell et al. 2021). The accelerated SOM transformation results in higher productivity (Chapin et al. 2012; Paul 2016), faster recycling rates (Zech et al. 1997; Saggar et al. 2011), and a greater turnover rate of soil carbon (C) in tropical grasslands compared to temperate grasslands (Horwath 2015; Paul 2016; Six and Jastrow 2017). Phosphorus (P) limitation is a constant factor in tropical biomes due to climate conditions, as well as historical soil development and geological factors (Vitousek et al. 2010; Sanchez 2019). In tropical regions, pastures are typically established in areas that were originally covered by natural forests, leading to the conversion of a significant portion of forested and wooded areas (Dias et al. 2016; Lerner et al. 2017; Aryal et al. 2018; Ávila-Bello et al. 2018). All these distinctions between temperate and tropical zones have implications for soil biogeochemical processes and may derive in a stoichiometric signature of grassland-associated ecosystems.
Grazing also affects soil nutrient concentration and ratios by affecting the physical properties of the plant-soil system (Ash et al. 2011; Teague et al. 2013; Pulido et al. 2016). Vegetation consumption, plant trampling and soil compaction caused by livestock can affect nutrient storage and transformation in grasslands (Greenwood and McKenzie 2001; Taboada et al. 2011). A reduction in soil porosity, for example, affects the soil water retention capacity (Cerda et al. 1998; Bartley et al. 2010; Pulido et al. 2016) and microbial diversity (Northup et al. 1999; Pan et al. 2018), and these factors together affect the soil nutrients concentration and cycling (Semmartin et al. 2008; Schnyder et al. 2010; Wang et al. 2016). Grazing can alter coupling of C, N, and inorganic P cycles because it can modify soil organic matter storage and stimulate belowground biological activity (Rumpel et al. 2015). Studies on this topic are inconclusive on whether there is a positive or negative effect of grazing on soil element concentration (Conant et al. 2001, 2017; Pineiro et al. 2010; McSherry and Ritchie 2013; Zhou et al. 2017; Abdalla et al. 2018). However, adverse effects of grazing, particularly those related to grazing intensities, have been reported in temperate (Piñeiro et al. 2009; Pineiro et al. 2010; McSherry and Ritchie 2013; Abdalla et al. 2018; He et al. 2019) and in tropical regions (Ritchie 2014; Abdalla et al. 2018; Pasricha and Ghosh 2019; Pringle et al. 2014). The impact of grazing on total P in tropical soils has yet to be studied despite the limitation of total P occurring in many tropical soils (Joergensen 2010). Little is known about how grazing influences all soil C, N and P concentrations and their ratios in both natural grasslands and pastures and in tropical vs. temperate climates with different environmental and geographic conditions.
This study aimed to compare concentrations and stoichiometric ratios of soil organic C (SOC) and nutrients (N and P) of natural grasslands and pastures, grazed and ungrazed, in two different climatic zones (i.e., tropical vs. warm temperate). We also explored the relationship between environmental (mean annual temperature, MAT, and precipitation, MAP) and geographical (latitude, altitude) variables in soil elemental concentrations and ratios. The incorporation of environmental and geographical factors in the analysis could aid in elucidating stoichiometric patterns in temperate and tropical grasslands. Furthermore, we aim to ascertain whether these patterns persist when incorporating variables such as grassland type and grazing. We expect to observe differences in grassland soils between temperate and tropical zones, where the stoichiometric signatures will reflect a higher nutrient concentration in temperate zones (low C:N and C:P ratios) and a greater P limitation in tropical zones (high C:P and N:P ratios). In this regard, we expect to gain a deeper understanding of the relationship between soil nutrient concentrations and C:N:P stoichiometric signatures with environmental variables, such as MAT and MAP, and geographical coordinates, such as latitude and altitude. A higher C concentration in pasture soils is also expected, regardless of the climatic zone, due to accelerated nutrient (N and P) use in plant growth associated with grazing by livestock. On this point, we predict that grazing would lead to decreasing soil P more rapidly in tropical grasslands compared to soils from ungrazed sites. The effect of grazing on C:nutrients stoichiometry can be reflected in a stoichiometric signature of high soil C:P and N:P ratios.
Methods
Data sources and search terms
Using peer-reviewed papers published before April 2023 and with data available online, a database of SOC, total N, and total P in grasslands was compiled, for two Köppen-Geiger climate zones: equatorial or humid tropical (zone A) and warm temperate or mesothermic (zone C). We compiled the papers using Scopus, with the following search terms: (soil carbon / soil organic carbon / nitrogen / phosphorus / nutrients) and (grasslands / rangelands / savanna / grazing lands / pastures / cultivated grasslands / tropical grasslands) and (grazing). We limited the search terms to title, keywords, and abstract. Given the limited coverage of studies in tropical regions and the southern hemisphere, mainly in Latin America and Africa, a second, broader search of other databases (Google Scholar, Scielo, and Redalyc) was made, including papers in English, Spanish, and Portuguese.
For a more thorough review, we examined the reference lists of collected papers on the comprehensive analysis of C and nutrient concentrations in grassland soils, focusing on previous reviews and meta-analyses that include effects of grazing (Conant et al. 2001, 2017; Pineiro et al. 2009; McSherry and Ritchie 2013; Rumpel et al. 2015; Abdalla et al. 2018; Byrnes et al. 2018; He et al. 2020), climate zones (Pärtel et al. 2008; Joergensen 2010) and grassland types (Xu et al. 2013). Our compilation has been enriched notably by a contribution from Xu et al. (2014). This database compile data about SOC, N, and P, and their ratios at biome and global scales (Xu et al. 2013). We extracted information for natural grasslands and pastures soils.
Criteria for selection of published studies
Each site reported in the source papers was considered an independent sample. When a site was reported in two different papers with complementary information, it was considered a single sample (e.g., Damian et al. 2020, 2021; Franzluebbers and Stuedemann 2005, 2009, references in supplementary information). Given that the present study did not aim to evaluate seasonal variation, and that total soil concentrations typically vary relatively little, we obtained a mean value when papers reported results at different times of the year for a given site. The following information was also obtained for each site: coordinates, MAT, MAP, soil type according to WRB (2015), sampling date and depth, grazing condition, grazer species, and stocking rate, when available. When geographical coordinates were unavailable in the source papers, data were estimated via Google Earth.
Since the relationships between elements in reactions occur on a molar basis (Sterner and Elser 2002), all data reported in different units were converted into molar units (mmol kg−1). We omitted sites where data were presented as mass area−1, unless bulk density and depth data were included, which allowed a conversion to mmol kg−1. Only soil surface data (< 30 cm) were obtained. Since we have data at different depths, we do not distinguish between shallow depths less than 30 cm. We assume that the average values obtained may have a variation associated with confounded factors in the depth interval from 0 to 30 cm. Most of the data presented (~ 90%) corresponds to depths from 0 to 20 cm. Given that not all papers have information on all three elements (SOC, N, and P), the number of data points for stoichiometric ratios (C:N = SOC:nitrogen, C:P = SOC:phosphorus, and N:P = nitrogen:phosphorus) were different for each site.
Data classification
The collected information was classified into two climatic zones according to Köppen-Geiger: equatorial or humid tropical (Köppen-Geiger zone A) and warm temperate or mesothermic (Köppen-Geiger zone C). The climatic zone was determined following Kottek et al. (2006) based on the coordinates and site name if no data were provided by the respective reference. We only included grasslands in the A and C climatic zones since pastures have been established mainly in these regions (Humphreys 1981; Suttie et al. 2005; Ramankutty et al. 2008; Reinermann et al. 2020). Pastures have also been cultivated in hot semi-arid climate (Bsh) regions, but we did not consider these sites because they accounted for less than 2.0% of the retrieved data. Sites were classified as natural grasslands or pastures (cultivated or induced grasslands) and as grazed or ungrazed sites, as reported in the source papers. Grasslands include savannas, which are natural grasslands in tropical areas, which represent 4.1% of data. Induced grasslands (3.5% of the total data) were included in the pastures group (cultivated grasslands), as they were established in sites where original vegetation was replaced with grasses (Suttie et al. 2005; Sanchez 2019; Teutscherová et al. 2021). When the grazing condition was not specified in the original papers, we classified the sites as ungrazed.
Database summary
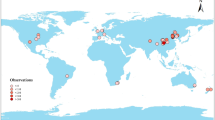
We compiled a dataset of 1296 records from 241 papers (Online Resource 1, a list of data sources), which included samples from 40 countries, mainly Brazil (n = 31, 13.7% of total papers), New Zealand (n = 29 papers, 12.8%), the United States of America (n = 21, 9.3%), Mexico (n = 18, 8%), and the United Kingdom (n = 17, 7.5%). Most sites (70.0%) were located at latitudes outside the tropics, with extreme latitudes of 57º06' N and 46º24' S. One-third of the data (35.8%) were North of the Tropic of Cancer and 23.3% South of the Tropic of Capricorn (Fig. 1). Sites in equatorial or humid tropical regions (zone A) represent 36.2% of the total data. Pastures account for 59.3% of the entries, and 57.8% of sites were subject to grazing (Supplementary Appendix 1). Most of the data (90%) were collected as samples at 0–20 cm depth. The main soil types for tropical climate regions were Ferralsols, with 32.3% of data (according to the World Reference Base, WRB 2015; Oxisols according to US Soil Taxonomy classification), 26.5% of data were Acrisols and 19.8% Vertisols. 37.7% of data were Cambisols, 11.0% Acrisols and 8.6% Luvisols for sites in warm temperate regions (Supplementary Appendix 2). The mean clay content in soils for the climatic zones was 29.2% for the tropical zone (37.0% in Ferralsols, 21.0% in Acrisols, and 34.0% in Vertisols) and 22.5% for the temperate zone (27.5% in Cambisols, 20.1% in Acrisols, and 23.4% in Luvisols).

Global distribution of the compiled information. a Site distribution in the climatic zones, b papers by country
Data analyses
A three-way ANOVA with a Tukey’s post-hoc test (p < 0.05 level) was performed to examine the effect of the climatic zone (Köppen-Geiger zone A and zone C), grassland type (natural grassland and pasture), grazing regime (grazed and ungrazed), and the interactions between these three factors on soil elemental concentrations (SOC, N, and P) and their stoichiometric ratios (C:N, C:P, and N:P). We conducted exploratory analyses to test the assumptions of normal distribution of residuals (Shapiro-Wilks’s test) and homoscedasticity (Fligner-Killen’s test) (Jones et al. 2022). Since data were not normally distributed, log transformation was used for ANOVA models to reduce the effect of outliers and increase the power of the statistical tests employed.
We also explored the relationships between elemental concentrations and ratios with environmental factors (MAP, MAT, altitude, and latitude) using linear regression models (Lepš and Šmilauer 2020). A log-linear transformation was used to ensure data normality. For correlations between element concentrations and their ratios we used a Pearson linear correlation, and the slope value was tested with one-sided tests (Zar 2014). All statistical analyses were performed using the packages `agricolae´ 1.3–5 (Mendiburu and Yaseen 2020) and `ggplot2´ 3.3.6 (Wickham 2022) in R (R Core Team 2020). The significance level was set as α = 0.05.
Results
Soil element concentrations and ratios according to climate, grassland type, and grazing
The SOC, N, and P concentrations were higher in warm temperate than tropical sites, whether in natural grasslands or pastures. The SOC and N concentrations were higher in pastures than in natural grasslands at the global level (Table 1). The C:N ratios were higher in pastures than in natural grasslands for ungrazed sites in both climate zones (Table 2). In the tropical zone, N:P ratios were higher in ungrazed pastures than in grazed or ungrazed natural grasslands sites (Table 1). Tropical pastures showed higher C:N ratio for ungrazed than for grazed sites (Table 1).
Concentrations of SOC, N, and C:P ratio differed when comparing climatic zones and grassland types (p < 0.001; Table 2). Total P differed when comparing climatic zones and C:N and N:P ratios differed among grassland types. However, these factors explain less than 10% of the variance in SOC and N concentrations but more than 25% in P concentration and C:P and N:P ratios (Table 2). Soil P concentration and C:P and N:P ratios did not show significant differences when we compared grazed and ungrazed sites, but the impact of grazing was significant for soil N concentration. The factors (climate, grassland type, and grazing) had an interactive effect on soil N concentration (p = 0.02) and N:P (p = 0.03) when considering climate and grassland type interaction. Also, an effect was observed for SOC (p = 0.004, Table 2) and C:N (p < 0.001, Table 2) for grassland type and grazing interaction, and interactive effect of climate zone and grazing was observed for C:N ratio (p < 0.001, Table 2).
Relationships among soil elemental concentrations, and environmental and geographical variables
We found that SOC and N concentrations in grassland soils increased with altitude, latitude, and MAP but decreased with MAT (Fig. 2, Supplementary Appendix 3 and 4). Soil P concentration was negatively related to MAT but increased with latitude. Soil P increased for MAP and altitude in tropical grasslands (Fig. 2). The C:P and N:P ratios increased with MAT and MAP but decreased with latitude; C:P and C:N ratios increased with altitude in warm temperate but decreased with this variable in tropical grasslands. The C:N decreased with MAT and increased with latitude only for tropical natural grasslands (Fig. 2, Supplementary Appendix 3 and 4). The SOC and N concentrations had similar trends in grasslands of tropical and temperate regions. The highest values for SOC (> 10,000 mmol kg−1) corresponded to montane grasslands in the Andean region (see, Oliver et al. 2017; Oliveras et al. 2014), and led to some differences between SOC and N trends with latitude and altitude (Fig. 2). Soil total P concentration and related ratios (C:P, N:P) had different trends related to altitude and MAP when comparing tropical and warm temperate grasslands (Fig. 2, Supplementary Appendix 4).

Regression plots of elemental concentrations and ratios with environmental and geographical factors. Soil elements: SOC = soil organic carbon (panels “a” to “d”); N = total nitrogen (“e” to “h”); P = total phosphorus (“i” to “l”). Ratios: C:N = SOC:nitrogen (“m” to “p”); C:P = SOC:phosphorus (“q” to “t”); N:P = nitrogen:phosphorus (“u” to “x”. MAT, mean annual temperature; MAP, mean annual precipitation. Red points, sites in the equatorial or humid tropical climatic zone (A); green points, sites in the warm temperate climatic zone (C). Statistical significance: *** < 0.001; ** < 0.01; * < 0.05; ns = non-significant. Complementary information in Supplementary Appendix 3 and 4
Correlations between SOC, N, and P and their ratios
At a global level, a strong positive correlation was found between SOC and N (r = 0.9) and between C:P and N:P (r = 0.89) (Figs. 3a and 3f, Table 3). Correlations between SOC and P (r = 0.49), N and P (r = 0.58), and C:N and C:P (r = 0.36) were significant (p < 0.0001) (Fig. 3b to 3d, Table 3). When comparing both warm temperate and tropical climate zones, the pattern observed for SOC and N was the same as the one observed at a global level (see Table 3). The same thing occurs for correlations between SOC and P, and N and P (Figs. 3b and 3c). The correlation between C:P and N:P was weaker for warm temperate grasslands (r = 0.80) than for tropical sites (r = 0.91) (Fig. 3f). The correlations observed between C:N and C:P at a global level (r = 0.36, p < 0.001) were different when comparing both climate zones: significant for warm temperate regions (r = 0.66, p < 0.0001) but non-significant for tropical grasslands (r = 0.19, p = 0.09) (Fig. 3d, Table 3).

Correlations between element concentrations and their ratios. Red points represent the sites in the equatorial or humid tropical climatic zone (A) and green triangles correspond to sites in the warm temperate climatic zone (C) (models and correlation parameters in Table 3)
Discussion
Grasslands are ecosystems distributed worldwide and subject to conditions related to climate and management that determine the availability of soil C and nutrients (Westoby et al. 1989; Jouany et al. 2011; Vendramini et al. 2014). According to our results, soils of natural grasslands and pastures in the humid tropics have lower SOC concentration and higher C:P and N:P ratios than those from warm temperate regions on a global scale. However, other factors lead to different patterns at regional scales. For example, grazing has a significant effect in increasing soil N concentration and SOC, especially in temperate climates. For C:P and N:P ratios, the direction of the effect depends on grassland type and climatic zone. Soil stoichiometric signatures of grasslands differ between temperate and tropical climatic zones at global scales, although patterns at regional scales change when factors such as grassland type, grazing, temperature, precipitation, latitude, and altitude are included in the analysis.
SOC, N, and P concentrations were higher in sites in warm temperate than in tropical zones
Natural grasslands and pastures soils in humid tropical zones (Köppen-Geiger zone A) tend to have lower SOC, N, and P concentrations than soils in warm temperate zones (Köppen-Geiger zone C). These results can be explained by the fact that organic matter transformation processes are faster in the tropics (van Keulen 2001; Nortcliff 2010). Soil net primary production (Chapin et al. 2012; Paul 2016) and C turnover rate (Trumbore 1993; Feller and Beare 1997; Šantrůčková et al. 2000; Six et al. 2002; Horwath 2015; Paul 2016; Six and Jastrow 2017) are higher in tropical than in temperate grasslands. High temperatures promote faster litter decomposition, nutrient transformation, recycling rates (Haynes and Williams 1993; Zech et al. 1997; Saggar et al. 2011), and microbial biomass production (Joergensen 2010). Changes in SOC are likely to affect soil total N due to a close association between SOC and total N in soil organic matter (Pineiro et al. 2010; Pringle et al. 2014). It has been reported that a higher microbial decomposition of SOC is related to increases in MAT and consequently lower concentrations of SOC and soil total N can be seen (Amundson 2021). This can be observed in our results, since SOC and N concentrations were negatively related to MAT and positively related to MAP (Fig. 2, Supplementary Appendix 3 and 4).
The lower total soil P concentrations in tropical compared to temperate regions (Fig. 2) could be explained by factors such as temperature, precipitation, and soil types. Soil total P concentration and C:P ratio decreased with MAT and MAP (Figs. 2, Supplementary Appendix 3 and 4), but it increased with SOC and N (Fig. 3). Sites with high total P concentrations are mainly located in temperate zones (Hou et al. 2018), with some exceptions for P-rich soils (Andosols, Mollisols) in volcanic and fertile tropical rainy regions (Cleveland et al. 2003; Huth et al. 2012). In temperate regions, high temperatures can promote soil P transformation from extractable to more stable forms (Siebers et al. 2017). Temperature and soil moisture enhance P-mineralization. Rainfall can increase inorganic P-fraction leaching (Arenberg and Arai 2019), but P organic fraction may be sequestered geochemically. In highly weathered tropical soils (Oxisols, Ultisols), these processes lead to a lower concentration of soil nutrients (Lopes et al. 2004; Peña-Peña and Irmler 2018; Vitousek and Sanford 1986; Vitousek et al. 2010) and high C:P ratios (Tipping et al. 2016), as we observed in our study. Low relative P-availability in tropical regions is also associated to sorption by allophanes in young volcanic soils (e.g. Andosols) and sesquioxide clays (e.g., Oxisols) (Vitousek and Sanford 1986; Gijsman et al. 1997; Hou et al. 2018). In this context, P could be a key element of regulation and restriction on soil C and N cycling in tropical regions more than in temperate zones.
Stoichiometric signatures differ according to climatic zones and environmental variables
Although SOC and N showed lower concentrations in tropical than in temperate grasslands, there were no significant differences in the C:N ratio for both climatic zones (Table 2). C:N values do not show differences between climatic zones (Table 1). Ratios of 14:1 have been previously reported for C:N, based on total C (Cleveland and Liptzin 2007) and of 13:1 for SOC:N (Xu et al. 2013). The C:N ratio is a litter decomposition driver and therefore promotes organic matter formation (Horwath 2015; Amundson 2021). This stoichiometric signature indicates an increase in SOC recalcitrance (for natural grasslands or pastures) or lower total N, leading to higher C:N ratios. Therefore, it stimulates N-immobilization, reducing its availability to plants (Robertson and Groffman 2015) and limiting soil organic matter, SOC formation and storage (Pineiro et al. 2010). The C:N ratios remained within intervals of 2 to 36 (> 90% of data are between 8 and 22), with higher variation in N:P ratios, which ranged from 1 to 46 (5 to 22 for 90% of data). Similar intervals have been reported on a global scale for the C:N ratio (2 to 30) but wider range for N:P (1 to 77), reflecting a strong coupling between the C and N cycles, but a decoupling of the total P from C and N concentration (Cleveland and Liptzin 2007). This association between SOC and total N has also been observed at regional or local scales (Pineiro et al. 2010; Tian et al. 2010; Pringle et al. 2014). A C:P ratio of 230:1 was found for tropical soils and of 158:1 for temperate sites, regardless of whether they were pastures or natural grasslands. A C:P ratio of 166:1 has been previously reported for grasslands globally (Cleveland and Liptzin 2007). This pattern has also been reported in forests (McGroddy et al. 2004) and other ecosystems (Yan et al. 2016), where the N:P ratio increases towards equatorial regions, following an increase in MAT. In our study, N:P ratios were 15:1 for grasslands in equatorial zones and 11:1 for sites in warm temperate regions.
Soil C:N:P stoichiometry among grassland types
Grassland-type (natural grassland or pasture) had a significant effect on SOC and nutrient concentrations in both climatic regions. We can explain this partially, due to characteristics of plant communities established in these ecosystems. Temperate grasslands are dominated by C3 grasses, while C4 grasses occur predominantly in tropical regions (Woodward et al. 2004; Lehmann and Parr 2016). As mentioned before, pastures in tropical regions are generally cultivated in sites whose original vegetation was forest (predominantly C3 plants), and it is replaced mainly with exotic C4 grasses (Oliveras and Malhi 2016). Grasslands dominated by C4 grasses store more SOC and N than those dominated by C3 plants, as has been reported for both temperate (Tilman and Wedin 1991; Yang et al. 2019) and tropical regions (Nyameasem et al. 2020). Tropical C4 grasses (e. g. Panicum, Pennisetum) metabolism is more efficient in terms of photosynthetic activity indicators (e. g. use efficiency of resources such as light, water, or nutrients) compared to C3 grasses from temperate climates (da Silva et al. 2015; Volenec and Nelson 2020). Given their higher photosynthetic efficiency, C4 grasses use less water, but also have higher lignin content (Volenec and Nelson 2020) and produce lower quality litter, which is more slowly incorporated into soil (Thomas and Asakawa 1993). The introduction of African deep-rooted C4 grasses into native savannas could increase soil C storage (Fisher et al. 1994; Fujisaki et al. 2015) but also higher C:N ratios (Williams and Baruch 2000), so the combination with N-fixing legumes could increase soil N content. This functional type of grasses also increases the P stock in their tissues and makes efficient use of this element, but further research is required. This C3/C4 grass communities composition contributes to explain SOC higher concentrations and nutrients in tropical pastures than in natural grasslands.
High lignin content in C4 grasses may limit the incorporation of plant matter into soil, a condition that is also favored by P deficiency (Lopes et al. 2004; García-Oliva et al. 2006; Vendramini et al. 2014). This P deficiency is reflected in P lower concentration and high C:P and N:P ratios in pastures (C:P, 211:1, N:P, 14:1), usually higher than in natural grasslands (C:P, 152:1; N:P, 11:1). When considering the climatic zone, the average values for both ratios were higher in tropical regions than in temperate regions. On a global scale, higher C:P and N:P ratios have also been reported for pastures (169:1 and 12:1) compared to natural grasslands (143:1 and 11:1) (Xu et al. 2013). This trend of lower relative P concentration is also observed in C:P ratios in pastures and has also been reported in previous studies (Xu et al. 2013). The lower relative content of nutrients in pasture soils can limit the SOC use by organisms in the system (Abbas et al. 2013; Achat et al. 2016).
Grazing impacts on soil C:N:P stoichiometry
The grazing influence on SOC and soil nutrient concentrations depended on the type of grassland. In natural grasslands, the highest concentrations of SOC, N, and P were found in grazed sites, but higher C:P and N:P ratios were observed in ungrazed tropical pastures (Table 1). Grazing can inhibit the growth of tropical plants with high efficiency characteristics in the acquisition of soil nutrients, while without grazing, these plants can grow, uptake nutrients, and establish themselves. More research is needed on this topic. The most used livestock management method in tropical grasslands is extensive grazing (Dubeux et al. 2007; Teutscherová et al. 2021). This method promotes selective forage consumption leading to zonal degradation (Kothmann 2009), and urine and dung patches unevenly distributed. Two effects can be expected about this grazing management method: (1) an increase in N-recycling and availability when nutrients remain and storage on site, or (2) an increase in N-losses through volatilization and leaching (Dubeux et al. 2007; Pineiro et al. 2010). In the first case, the plant-soil system can store more N in part by biomass microbial immobilization. In the second case, if available N exceeds the short-term requirements of pastures around dung patches, losses will occur (Haynes and Williams 1993). Even when these patches can contribute to an increase in SOC and N, compaction and intense rainfall can promote nutrient losses by surface runoff and limit soil N inputs (Greenwood and McKenzie 2001; Taboada et al. 2011). Increased soil C stocks by improvements in grazing management (i.e., stocking density management, rotation grazing) have been reported (Conant et al. 2017). Grazing effects also can depend on grass composition (C3 or C4 grasses) and environmental conditions (McSherry and Ritchie 2013; Abdalla et al. 2018; He et al. 2020). The effect of grazing on SOC concentration also depends on other specific factors at more local scales, such as environmental conditions (precipitation, temperature), soil properties, land topography (Pineiro et al. 2009), and grazing improvements (Conant et al. 2017).
Effect of geographical variables on stoichiometric signature patterns
Geographical variables (i.e., altitude and latitude) have consistent effects on SOC and N concentrations, which both increase with altitude and latitude. The highest (Andean) montane grasslands soil, and the northernmost (England) and southernmost (New Zealand) regions are the richest. Decreases in annual mean temperature (5.5ºC) generally have been calculated for every increase of 1000 m in altitude above sea level, or for increases of 15º in latitude above 10º North or South (Humphreys 1981). Following this pattern, increases in SOC and N concentrations proportional to altitude and latitude would be expected in grasslands, which is supported by our results. Increasing SOC and total N concentrations with altitude have also been observed at a local level (Gerschlauer et al. 2016). The higher concentrations of SOC we found (up to 24,000 mmol kg−1) have been reported in Andean montane grasslands, over 3000 m.a.s.l. (Oliver et al. 2017; Oliveras et al. 2014). It is important to note that N-mineralization increases with soil moisture (Singh et al. 1991) but also with elevation (Gerschlauer et al. 2016), and thus if N is not taken up or immobilized, then it could be lost by leaching or runoff. Furthermore, if increasing temperature and limited water availability lead to a decline in microbial growth, then there is also a decline in C-use efficiency (Manzoni et al. 2012). At low P concentrations N2 fixation is limited and N availability for plants can be reduced (Sardans and Peñuelas 2012). These results are an invitation to study and compare patterns of nutrient concentration and transformation in altitudinal gradients in other mountain systems, such as other grassland areas in the Andes, Kilimanjaro, the Himalayas, and mountain regions in Mexico, Central America, and New Zealand, to name a few examples.
Limitations and future work
This study investigated soil organic C, N, and P among warm temperate and tropical grassland, and the underlying mechanisms. Although meta-analysis approach allows a synthesis to establish global and regional patterns that cannot be observed through individual studies, a few limitations have been identified and will be addressed in our future work. First, intrinsic diversity in grassland ecosystems leads to a high variability in C, N, and P concentrations. This variability has also been found in other reviews and is a source of uncertainty. Second, the relative lack of information about soil P has also been a frequent issue in previous reviews (Cleveland and Liptzin 2007; Tian et al. 2010). Third, there is a lack of standardized methodologies for determining elemental concentrations as a source of heterogeneity in datasets. Fourth, the conclusions related to effects of grazing on concentrations and ratios evaluated are limited by the necessary non-random sample of grazed vs. ungrazed sites. Despite all these methodological issues as a source of uncertainty, our findings can contribute to increase knowledge of factors affecting the status of elements in grazed and ungrazed grassland soils, particularly in pastures of tropical regions. There remains a clear gap of information in these tropical ecosystems, as can be seen from the collected data (Table 1, Supplementary Appendix 1) and from previous reviews.
Conclusions
Our findings suggest that soil stoichiometric C:N:P stoichiometric signatures in grasslands differed between tropical and temperate regions on a global scale. Our results can be partly attributed to the effect of mean annual temperature (MAT) and mean annual precipitation (MAP) on soil C and nutrient reduction, resulting in higher C:P and N:P ratios in tropical pastures. It could be hypothesized that in tropical regions, in contrast to temperate regions, P is a key element in regulating and limiting soil C and N cycling. Changes in soil P concentrations can have significant effects on soil C and N stoichiometric ratios, highlighting the importance of understanding the mechanisms behind soil P reduction in elucidating the functioning of tropical regions. Imbalances in soil C:N:P stoichiometric ratios could lead to cascading stoichiometric changes in N and P availability throughout the ecosystem. As nitrogen (N) and phosphorus (P) are elements that regulate the growth rate of organisms, these changes could affect both living organisms, such as plants, microorganisms and herbivores, and inorganic reservoirs of the ecosystem, including the atmosphere and water. Such imbalances can therefore affect ecosystem functioning and productivity, and taking these patterns into account can be a valuable tool for planning management and conservation of natural grasslands and pastures.
Data availability
Data will be made available on request.
References
Abbas M, Ebeling A, Oelmann Y et al (2013) Biodiversity effects on plant stoichiometry. PLoS ONE 8:e58179. https://doi.org/10.1371/journal.pone.0058179
Abdalla M, Hastings A, Chadwick DR et al (2018) Critical review of the impacts of grazing intensity on soil organic carbon storage and other soil quality indicators in extensively managed grasslands. Agric Ecosyst Environ 253:62–81. https://doi.org/10.1016/j.agee.2017.10.023
Achat DL, Augusto L, Gallet-Budynek A, Loustau D (2016) Future challenges in coupled C-N–P cycle models for terrestrial ecosystems under global change: a review. Biogeochemistry 131:173–202. https://doi.org/10.1007/s10533-016-0274-9
Ågren GI (2008) Stoichiometry and nutrition of plant growth in natural communities. Annu Rev Ecol Evol Syst 39:153–170. https://doi.org/10.1146/annurev.ecolsys.39.110707.173515
Amundson R (2021) Introduction to the biogeochemistry of soils, 1st edn. Cambridge University Press, Cambridge
Arenberg MR, Arai Y (2019) Uncertainties in soil physicochemical factors controlling phosphorus mineralization and immobilization processes. Adv Agron 154:153–200
Arhonditsis GB, Shimoda Y, Kelly NE (2019) Allometric theory: extrapolations from individuals to ecosystems. In: Fath, B., Encyclopedia of ecology, 2nd edn. Elsevier, Amsterdam.
Aryal DR, Ruiz DEM, Marroquín CNT et al (2018) Soil organic carbon depletion from forests to grasslands conversion in Mexico: a review. Agric 8:1–15. https://doi.org/10.3390/agriculture8110181
Ash AJ, Corfield JP, McIvor JG, Ksiksi TS (2011) Grazing management in tropical savannas: Utilization and rest strategies to manipulate rangeland condition. Rangel Ecol Manag 64:223–239. https://doi.org/10.2111/REM-D-09-00111.1
Ávila-Bello CH, Hernández-Romero AH, Mendoza-Briseño MA, Vázquez-Luna D (2018) Complex systems, agroecological matrices, and management of forest resources: an example of an application in Los Tuxtlas, Veracruz, Mexico. Sustainability. https://doi.org/10.3390/su10103496
Bartley R, Corfield JP, Abbott BN et al (2010) Impacts of improved grazing land management on sediment yields, Part 1: Hillslope processes. J Hydrol 389:237–248. https://doi.org/10.1016/j.jhydrol.2010.05.002
Binkley D, Fisher RF (2020) Ecology and management of forest soils, 5th edn. Wiley-Blackwell, New York
Blair J, Nippert J, Briggs J (2014) Grassland ecology. In: Monson RK (ed) Ecology and the environment, 1st edn. Springer, New York, pp 389–423
Byrnes RC, Eastburn DJ, Tate KW, Roche LM (2018) A global meta-analysis of grazing impacts on soil health indicators. J Environ Qual 47:758–765. https://doi.org/10.2134/jeq2017.08.0313
Butler OM, Lewis T, Maunsell SC, et al (2021) The stoichiometric signature of high-frequency fire in forest floor food webs. Ecol Monogr 91:. https://doi.org/10.1002/ecm.1477
Cerda A, Schnabel S, Ceballos A, Gomez-Amelia D (1998) Soil hydrological response under simulated rainfall in the Dehesa land system (Extremadura, SW Spain) under drought conditions. Earth Surf Process Landforms 23:195–209. https://doi.org/10.1002/(SICI)1096-9837(199803)23:3%3c195::AID-ESP830%3e3.0.CO;2-I
Chan MA, Hinman NW, Potter-McIntyre SL et al (2019) Deciphering biosignatures in planetary contexts. Astrobiology 19:1075–1102. https://doi.org/10.1089/ast.2018.1903
Chang J, Ciais P, Gasser T et al (2021) Climate warming from managed grasslands cancels the cooling effect of carbon sinks in sparsely grazed and natural grasslands. Nat Commun 12:1–10. https://doi.org/10.1038/s41467-020-20406-7
Chapin FS, Matson PA, Mooney HA (2012) Principles of terrestrial ecosystem ecology. Springer, New York
Cleveland CC, Liptzin D (2007) C:N: P stoichiometry in soil: Is there a “Redfield ratio” for the microbial biomass? Biogeochemistry 85:235–252. https://doi.org/10.1007/s10533-007-9132-0
Cleveland CC, Townsend AR, Schmidt SK, Constance BC (2003) Soil microbial dynamics and biogeochemistry in tropical forests and pastures, southwestern Costa Rica. Ecol Appl 13:314–326. https://doi.org/10.1890/1051-0761(2003)013[0314:SMDABI]2.0.CO;2
Conant RT, Paustian K, Elliott ET (2001) Grassland management and conversion into grassland: effects on soil carbon. Ecol Appl 11:343–355. https://doi.org/10.1890/1051-0761(2001)011[0343:GMACIG]2.0.CO;2
Conant RT, Cerri CEP, Osborne BB, Paustian K (2017) Grassland management impacts on soil carbon stocks: a new synthesis. Ecol Appl 27:662–668. https://doi.org/10.1002/eap.1473
da Silva S, Sbrissia A, Pereira L (2015) Ecophysiology of C4 forage grasses—understanding plant growth for optimising their use and management. Agriculture 5:598–625. https://doi.org/10.3390/agriculture5030598
Dias LCP, Pimenta FM, Santos AB et al (2016) Patterns of land use, extensification, and intensification of Brazilian agriculture. Glob Chang Biol 22:2887–2903. https://doi.org/10.1111/gcb.13314
Dubeux JCB, Sollenberger LE, Mathews BW et al (2007) Nutrient cycling in warm-climate grasslands. Crop Sci 47:915–928. https://doi.org/10.2135/cropsci2006.09.0581
Elser JJ, Bracken MES, Cleland EE et al (2007) Global analysis of nitrogen and phosphorus limitation of primary producers in freshwater, marine and terrestrial ecosystems. Ecol Lett 10:1135–1142. https://doi.org/10.1111/j.1461-0248.2007.01113.x
Feller C, Beare MH (1997) Physical control of soil organic matter dynamics in the tropics. Geoderma 79:69–116. https://doi.org/10.1016/S0016-7061(97)00039-6
Fisher MJ, Rao IM, Ayarza MA et al (1994) Carbon storage introduced grasses in savannas. Nature 371:236–238. https://doi.org/10.1038/371236a0
Fujisaki K, Perrin AS, Desjardins T et al (2015) From forest to cropland and pasture systems: a critical review of soil organic carbon stocks changes in Amazonia. Glob Change Biol 21:2773–2786. https://doi.org/10.1111/gcb.12906
García-Oliva F, Lancho JFG, Montaño NM, Islas P (2006) Soil carbon and nitrogen dynamics followed by a forest-to-pasture conversion in western Mexico. Agrofor Syst 66:93–100. https://doi.org/10.1007/s10457-005-2917-z
Gerschlauer F, Dannenmann M, Kühnel A et al (2016) Gross nitrogen turnover of natural and managed tropical ecosystems at Mt. Kilimanjaro Tanzania Ecosystems 19:1271–1288. https://doi.org/10.1007/s10021-016-0001-3
Gijsman AJ, Alarcòn HF, Thomas RJ (1997) Root decomposition in tropical grasses and legumes, as affected by soil texture and season. Soil Biol Biochem 29:1443–1450. https://doi.org/10.1016/S0038-0717(97)00039-4
Greenwood KL, McKenzie BM (2001) Grazing effects on soil physical properties and the consequences for pastures: a review. Aust J Exp Agric 41:1231–1250. https://doi.org/10.1071/EA00102
Haynes RJ, Williams PH (1993) Nutrient cycling and soil fertility in grazed pasture ecosystem. Adv Agron 49:119–199. https://doi.org/10.1016/S0065-2113(08)60794-4
He M, Zhou G, Yuan T et al (2020) Grazing intensity significantly changes the C:N: P stoichiometry in grassland ecosystems. Glob Ecol Biogeogr 29:355–369. https://doi.org/10.1111/geb.13028
Horwath W (2015) Carbon cycling: The dynamics and formation of organic matter. In: Paul EA (ed) Soil Microbiology, Ecology and Biochemistry, 4th edn. Academic Press, New York, pp 339–382
Hou E, Chen C, Luo Y et al (2018) Effects of climate on soil phosphorus cycle and availability in natural terrestrial ecosystems. Glob Chang Biol 24:3344–3356. https://doi.org/10.1111/gcb.14093
Humphreys LR (1981) Environmental adaptation of tropical pasture plants, 1st edn. MacMillan Publishers LTD, New York
Huth T, Porder S, Chaves J, Whiteside JH (2012) Soil carbon and nutrient changes associated with deforestation for pasture in Southern Costa Rica. Biotropica 44:661–667. https://doi.org/10.1111/j.1744-7429.2012.00863.x
ILRI, IUCN, FAO, et al (2021) Rangelands Atlas, 1st edn. FAO, Nairobi, Kenya
Joergensen RG (2010) Organic matter and micro-organisms in tropical soils. In: Dion P (ed) Soil Biology and Agriculture in the Tropics, 1st edn. Springer, Berlin, pp 17–44
Jones E, Harden S, Crawley MJ (2022) The R Book, 3rd edn. Wiley, Hoboken
Jouany C, Cruz P, Daufresne T, Duru M (2011) Biological phosphorus cycling in grasslands: interactions with nitrogen. In: Bünemann EK, Oberson A, Frossard E (eds) Phosphorus in action, 1st edn. Springer, Berlin, pp 275–294
Kempes CP, Follows MJ, Smith H et al (2021) Generalized stoichiometry and biogeochemistry for astrobiological applications. Bull Math Biol 83:1–22. https://doi.org/10.1007/s11538-021-00877-5
Kothmann M (2009) Grazing methods: a viewpoint. Rangelands 31:5–10. https://doi.org/10.2111/1551-501X-31.5.5
Kottek M, Grieser J, Beck C et al (2006) World map of the Köppen-Geiger climate classification updated. Meteorol Zeitschrift 15:259–263. https://doi.org/10.1127/0941-2948/2006/0130
Lehmann CER, Parr CL (2016) Tropical grassy biomes: Linking ecology, human use and conservation. Philos Trans R Soc B Biol Sci 371: https://doi.org/10.1098/rstb.2016.0329
Lepš J, Šmilauer P (2020) Biostatistics with R: An introductory guide for field biologists, 1st edn. Cambridge University Press, Cambridge
Lerner AM, Zuluaga AF, Chará J et al (2017) Sustainable cattle ranching in practice: moving from theory to planning in Colombia’s livestock sector. Environ Manag 60:176–184. https://doi.org/10.1007/s00267-017-0902-8
Lopes AS, Ayarza M, Thomas RJ (2004) Managing and conserving acid savanna soils for agricultural development: lessons from the Brazilian Cerrados. In: Guimarães EP, Sanz JI, Rao IM, et al. (eds) Agropastoral systems for the tropical savannas of Latin America, 1st edn. CIAT EMBRAPA, Cali, pp 11–41
Manzoni S, Taylor PG, Richter A et al (2012) Environmental and stoichiometric controls on microbial carbon-use efficiency in soils. New Phytol 196(1):79–91. https://doi.org/10.1111/j.1469-8137.2012.04225.x
McGroddy ME, Daufresne T, Hedin LO (2004) Scaling of C:N: P stoichiometry in forests worldwide: Implications of terrestrial Redfield-type ratios. Ecology 85:2390–2401. https://doi.org/10.1890/03-0351
McSherry ME, Ritchie ME (2013) Effects of grazing on grassland soil carbon: a global review. Glob Chang Biol 19:1347–1357. https://doi.org/10.1111/gcb.12144
Mendiburu F de, Yaseen M (2020) agricolae-package: statistical procedures for agricultural research, v. 1.4.0
Mitchell E, Scheer C, Rowlings D et al (2021) Important constraints on soil organic carbon formation efficiency in subtropical and tropical grasslands. Glob Chang Biol 27:5383–5391. https://doi.org/10.1111/gcb.15807
Nortcliff S (2010) Soils of the tropics. In: Dion P (ed) Soil biology and agriculture in the tropics, 1st edn. Springer, Berlin, pp 1–15
Northup BK, Brown JR, Holt JA (1999) Grazing impacts on the spatial distribution of soil microbial biomass around tussock grasses in a tropical grassland. Appl Soil Ecol 13:259–270. https://doi.org/10.1016/S0929-1393(99)00039-6
Nyameasem JK, Reinsch T, Taube F et al (2020) Nitrogen availability determines the long-term impact of land use change on soil carbon stocks in grasslands of southern Ghana. Soil 6:523–539. https://doi.org/10.5194/soil-6-523-2020
Oliver V, Oliveras I, Kala J et al (2017) The effects of burning and grazing on soil carbon dynamics in managed Peruvian tropical montane grasslands. Biogeosciences 14:5633–5646. https://doi.org/10.5194/bg-14-5633-2017
Oliveras I, Malhi Y (2016) Many shades of green: The dynamic tropical forest–savannah transition zones. Philos Trans R Soc B Biol Sci. https://doi.org/10.1098/rstb.2015.0308
Oliveras I, Girardin C, Doughty CE et al (2014) Andean grasslands are as productive as tropical cloud forests. Environ Res Lett 9:115011. https://doi.org/10.1088/1748-9326/9/11/115011
Pan H, Liu H, Liu Y et al (2018) Understanding the relationships between grazing intensity and the distribution of nitrifying communities in grassland soils. Sci Total Environ 634:1157–1164. https://doi.org/10.1016/j.scitotenv.2018.04.117
Pärtel M, Laanisto L, Wilson SD (2008) Soil nitrogen and carbon heterogeneity in woodlands and grasslands: Contrasts between temperate and tropical regions. Glob Ecol Biogeogr 17:18–24. https://doi.org/10.1111/j.1466-8238.2007.00336.x
Pasricha NS, Ghosh PK (2019) Soil organic carbon dynamics in tropical and subtropical grassland ecosystem. In: Ghosh P, Mahanta S, Mandal D, et al. (eds). Carbon management in tropical and sub-tropical terrestrial systems. Springer Nature, Singapore, pp 283–297.
Paul EA (2016) The nature and dynamics of soil organic matter: Plant inputs, microbial transformations, and organic matter stabilization. Soil Biol Biochem 98:109–126. https://doi.org/10.1016/j.soilbio.2016.04.001
Peña-Peña K, Irmler U (2018) Nitrogen and carbon losses from decomposing litter in natural and agroecosystems of two different climate regions of Brazil. Eur J Soil Biol 86:26–33. https://doi.org/10.1016/j.ejsobi.2018.02.003
Pineiro G, Paruelo JM, Jobbágy EG et al (2009) Grazing effects on belowground C and N stocks along a network of cattle exclosures in temperate and subtropical grasslands of South America. Global Biogeochem Cycles 23:1–14. https://doi.org/10.1029/2007GB003168
Pineiro G, Paruelo JM, Oesterheld M, Jobbágy EG (2010) Pathways of grazing effects on soil organic carbon and nitrogen. Rangel Ecol Manag 63:109–119. https://doi.org/10.2111/08-255.1
Pringle MJ, Allen DE, Phelps DG et al (2014) The effect of pasture utilization rate on stocks of soil organic carbon and total nitrogen in a semi-arid tropical grassland. Agric Ecosyst Environ 195:83–90. https://doi.org/10.1016/j.agee.2014.05.013
Pulido M, Schnabel S, Lavado Contador JF et al (2016) The impact of heavy grazing on soil quality and pasture production in rangelands of SW Spain. L Degrad Dev 29:219–230. https://doi.org/10.1002/ldr.2501
R Core Team (2020) R, v.4.0.2
Ramankutty N, Evan AT, Monfreda C, Foley JA (2008) Farming the planet: 1 Geographic distribution of global agricultural lands in the year 2000. Global Biogeochem Cycles 22:1–19. https://doi.org/10.1029/2007GB002952
Redfield AC (1958) The biological control of chemical factors in the environment. Am Sci 46:205–221
Reinermann S, Asam S, Kuenzer C (2020) Remote sensing of grassland production and management-a review. Remote Sensing 12: https://doi.org/10.3390/rs12121949
Reiners WA (1986) Complementary models for ecosystems. Am Nat 127:59–73. https://doi.org/10.1086/284467
Ritchie ME (2014) Plant compensation to grazing and soil carbon dynamics in a tropical grassland. PeerJ. https://doi.org/10.7717/peerj.233
Robertson GP, Groffman PM (2015) Nitrogen transformations. Paul EA, Soil microbiology, ecology and biochemistry, 4th edn. Academic Press, New York, pp 421–446
Rumpel C, Crème A, Ngo PT et al (2015) The impact of grassland management on biogeochemical cycles involving carbon, nitrogen and phosphorus. J Soil Sci Plant Nutr 15:353–371. https://doi.org/10.4067/S0718-95162015005000034
Saggar S, Luo J, Kim D-G, Jha N (2011) Intensification in pastoral farming: impacts on soil attributes and gaseous emissions, 1st edn. Springer, Berlin
Sanchez PA (2019) Properties and management of soils in the tropics, 2nd edn. Cambridge University Press, Cambridge
Šantrůčková H, Bird MI, Lloyd J (2000) Microbial processes and carbon-isotope fractionation in tropical and temperate grassland soils. Funct Ecol 14:108–114. https://doi.org/10.1046/j.1365-2435.2000.00402.x
Sardans J, Peñuelas J (2012) The role of plants in the effects of global change on nutrient availability and stoichiometry in the plant-soil system. Plant Physiol 160:1741–1761. https://doi.org/10.1104/pp.112.208785
Schnyder H, Locher F, Auerswald K (2010) Nutrient redistribution by grazing cattle drives patterns of topsoil N and P stocks in a low-input pasture ecosystem. Nutr Cycl Agroecosyst 88:183–195. https://doi.org/10.1007/s10705-009-9334-z
Semmartin M, Garibaldi LA, Chaneton EJ (2008) Grazing history effects on above- and below-ground litter decomposition and nutrient cycling in two co-occurring grasses. Plant Soil 303:177–189. https://doi.org/10.1007/s11104-007-9497-9
Siebers N, Sumann M, Kaiser K, Amelung W (2017) Climatic effects on phosphorus fractions of native and cultivated North American grassland soils. Soil Sci Soc Am J 81:299–309. https://doi.org/10.2136/sssaj2016.06.0181
Singh RS, Raghubanshi AS, Singh JS (1991) Nitrogen-mineralization in dry tropical savanna: effects of burning and grazing. Soil Biol Biochem 23:269–273. https://doi.org/10.1016/0038-0717(91)90063-P
Six J, Jastrow JD (2017) Soil organic matter (SOM): turnover. In: Lal R (ed) Encyclopedia of soil science, 3rd edn. CRC Press, New York, pp 2186–2191
Six J, Feller C, Denef K et al (2002) Soil organic matter, biota and aggregation in temperate and tropical soils - effects of no-tillage. Agronomie 22:755–775. https://doi.org/10.1051/agro:2002043
Sterner R, Elser J (2002) Ecological stoichiometry: the biology of elements from molecules to the biosphere, 1st edn. Princeton University Press, Princeton
Sterner R, Elser J (2008) Ecological stoichiometry: Overview. In: Jorgensen SE (ed) Encyclopedia of Ecology, 1st edn. Elsevier, Amsterdam, pp 1101–1116
Suttie JM, Reynolds SG, Batello C (2005) Grasslands of the world, 1st edn. FAO, Geneva
Taboada MA, Rubio G, Chaneton EJ et al (2011) Grazing impacts on soil physical, chemical, and ecological properties in forage production systems. Soil Manag. https://doi.org/10.2136/2011.soilmanagement.c20
Teague R, Provenza F, Kreuter U et al (2013) Multi-paddock grazing on rangelands: why the perceptual dichotomy between research results and rancher experience? J Environ Manag 128:699–717. https://doi.org/10.1016/j.jenvman.2013.05.064
Teutscherová N, Vázquez E, Sotelo M et al (2021) Intensive short-duration rotational grazing is associated with improved soil quality within one year after establishment in Colombia. Appl Soil Ecol. https://doi.org/10.1016/j.apsoil.2020.103835
Thomas RJ, Asakawa NM (1993) Decomposition of leaf litter from tropical forage grasses and legumes. Soil Biol Biochem 25:1351–1361. https://doi.org/10.1016/0038-0717(93)90050-L
Tian H, Chen G, Zhang C et al (2010) Pattern and variation of C:N: P ratios in China’s soils: a synthesis of observational data. Biogeochemistry 98:139–151. https://doi.org/10.1007/S10533-009-9382-0
Tilman D, Wedin D (1991) Plant traits and resource reduction for five grasses growing on a nitrogen gradient. Ecology 72:685–700. https://doi.org/10.2307/2937208
Tipping E, Somerville CJ, Luster J (2016) The C:N:P: S stoichiometry of soil organic matter. Biogeochemistry 130:117–131. https://doi.org/10.1007/s10533-016-0247-z
Trumbore SE (1993) Comparison of carbon dynamics in tropical and temperate soils using radiocarbon measurements. Global Biogeochem Cycles 7:275–290. https://doi.org/10.1029/93GB00468
van Keulen H (2001) (Tropical) soil organic matter modelling: problems and prospects. Manag Org Matter Trop Soils Scope Limit 33:33–39. https://doi.org/10.1007/978-94-017-2172-1_4
Vendramini JMB, Dubeux JCB, Silveira ML (2014) Nutrient cycling in tropical pasture ecosystems. Rev Bras Ciencias Agrar 9:308–315. https://doi.org/10.5039/agraria.v9i2a3730
Vitousek PM, Porder S, Houlton BZ, Chadwick OA (2010) Terrestrial phosphorus limitation: mechanisms, implications, and nitrogen–phosphorus interactions. Ecol Appl 20:5–15. https://doi.org/10.1890/08-0127.1
Vitousek PM, Sanford RL (1986) Nutrient cycling in moist tropical forest. Annu Rev Ecol Syst 17:137–167. https://doi.org/10.1146/annurev.es.17.110186.001033
Volenec JJ, Nelson CJ (2020) Carbon metabolism in forage plants. In: Moore KJ, Collins M, Nelson CJ, Redfearn DD (eds) Forages: The science of grassland agriculture, 7th edn. Wiley-Blackwell, Chichester, pp 65–84
Voroney RP, Heck RW (2015) The soil habitat. Paul EA, Soil microbiology, ecology and biochemistry, 4th edn. Academic Press, New York, pp 15–39
Wang X, McConkey BG, VandenBygaart AJ et al (2016) Grazing improves C and N cycling in the Northern Great Plains: A meta-analysis. Sci Rep 6:1–9. https://doi.org/10.1038/srep33190
Westoby M, Walker B, Noy-Meir I (1989) Opportunistic management for rangelands not at equilibrium. J Range Manag 42:266–274. https://doi.org/10.2307/3899492
Wickham H (2022) ggplot2: Elegant graphics for data analysis, v. 3.3.6
Williams DG, Baruch Z (2000) African grass invasion in the Americas: ecosystem consequences and the role of ecophysiology. Biol Invasions 2:123–140. https://doi.org/10.1023/A:1010040524588
Woodward FI, Lomas MR, Kelly CK, Sheffield S (2004) Global climate and the distribution of plant biomes. Philos Trans R Soc B Biol Sci 359:1465–1476. https://doi.org/10.1098/rstb.2004.1525
WRB IUSS Working Group (2015) World Reference Base for Soil Resources 2014, update 2015. International soil classification system for naming soils and creating legends for soil maps. World Soil Resources Reports No. 106. FAO, Rome
Xu X, Thornton PE, Post WM (2013) A global analysis of soil microbial biomass carbon, nitrogen and phosphorus in terrestrial ecosystems. Glob Ecol Biogeogr 22:737–749. https://doi.org/10.1111/geb.12029
Xu X, Thornton PE, Potapov P (2014) A compilation of global soil microbial biomass carbon, nitrogen, and phosphorus data. In: ORNL Distrib Act Arch Cent. https://daac.ornl.gov/cgi-bin/dsviewer.pl?ds_id=1264. Accessed 16 Jun 2020
Yan Z, Han W, Peñuelas J et al (2016) Phosphorus accumulates faster than nitrogen globally in freshwater ecosystems under anthropogenic impacts. Ecol Lett 19:1237–1246. https://doi.org/10.1111/ele.12658
Yang Y, Tilman D, Furey G, Lehman C (2019) Soil carbon sequestration accelerated by restoration of grassland biodiversity. Nat Commun 10:1–7. https://doi.org/10.1038/s41467-019-08636-w
Zar JH (2014) Biostatistical analysis, 5th edn. Pearson, Essex
Zech W, Senesi N, Guggenberger G et al (1997) Factors controlling humification and mineralization of soil organic matter in the tropics. Geoderma 79:117–161. https://doi.org/10.1016/S0016-7061(97)00040-2
Zhou G, Zhou X, He Y et al (2017) Grazing intensity significantly affects belowground carbon and nitrogen cycling in grassland ecosystems: a meta-analysis. Glob Chang Biol 23:1167–1179. https://doi.org/10.1111/gcb.13431
Acknowledgements
This study was developed under the project “Biogeochemistry of nutrients in natural and anthropized environments of eastern Mexico”, registered at the Universidad Veracruzana in SIREI with No. 354822022107. We thank Jeffrey E. Herrick and two anonymous reviewers for their suggestions that improved this manuscript, and to Osvaldo Trinidad Hernández for his aid in mapping. Authors are grateful to staff of the Instituto de Biotecnología y Ecología Aplicada (INBIOTECA) of Universidad Veracruzana for their logistic and administrative support in conducting this research. AHHR was supported by the National Council on Humanities, Science and Technology (CONAHCYT, Grant 100101). X.X. has been supported by a National Science Foundation (NSF) CAREER Project (2145130).
Funding
AHHR was supported by the National Council on Humanities, Science and Technology (CONAHCYT, Grant 100101). X.X. has been supported by an NSF CAREER project (2145130).
Author information
Authors and Affiliations
Contributions
Ángel Héctor Hernández-Romero and Yareni Perroni performed study conception and design, and data analysis. Data collection was performed by Ángel Héctor Hernández-Romero, Xiaofeng Xu, and Lihua Zhang. The first draft of the manuscript was written by Ángel Héctor Hernández-Romero and Yareni Perroni. All authors contributed to critically review and comment, and manuscript final version.
Corresponding author
Ethics declarations
Competing interests
The authors have no relevant financial or non-financial interests to disclose.
Additional information
Responsible Editor: Edith Bai.
Publisher's Note
Springer Nature remains neutral with regard to jurisdictional claims in published maps and institutional affiliations.
Supplementary Information
Below is the link to the electronic supplementary material.
Rights and permissions
Open Access This article is licensed under a Creative Commons Attribution 4.0 International License, which permits use, sharing, adaptation, distribution and reproduction in any medium or format, as long as you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons licence, and indicate if changes were made. The images or other third party material in this article are included in the article's Creative Commons licence, unless indicated otherwise in a credit line to the material. If material is not included in the article's Creative Commons licence and your intended use is not permitted by statutory regulation or exceeds the permitted use, you will need to obtain permission directly from the copyright holder. To view a copy of this licence, visit http://creativecommons.org/licenses/by/4.0/.
About this article

Cite this article
Hernández-Romero, Á.H., Perroni, Y., Sánchez Velásquez, L.R. et al. Soil C:N:P stoichiometric signatures of grasslands differ between tropical and warm temperate climatic zones. Biogeochemistry 167, 909–926 (2024). https://doi.org/10.1007/s10533-024-01143-1
Received:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10533-024-01143-1