
Abstract
Seamounts in the Ross Sea were surveyed during the New Zealand IPY-CAML research voyage in 2008. Admiralty seamount, and seamounts of the Scott Island Seamount chain (Scott Island, Scott A, Scott B, Scott South) were sampled to examine variability in benthic faunal communities of seamounts. Multivariate analysis of video and still image data showed that benthic community composition was highly variable both within and between seamounts. The distribution patterns of benthic communities varied considerably, with very high densities of certain taxa in some places, sparse fauna in many others, and highly patchy distributions for others. Dissimilarities between seamounts were influenced by differences in population densities of taxa, the proportions of transects in which taxa were present, and by taxa being restricted to only one seamount or group of seamounts. Scott Island seamount was less similar to the others, despite the greater distances between Admiralty and Scott A and B seamounts. The observed variability in community composition was mainly associated with differences in depth, chlorophyll concentration, and seabed rugosity. Results support an earlier hypothesis that the location of seamounts relative to the Ross Sea gyre could be an important factor determining the composition and abundance of benthic invertebrate communities.
Similar content being viewed by others
Avoid common mistakes on your manuscript.
Introduction
Seamounts are widespread features of the world’s underwater topography and have been thought to play an important role in patterns of marine biogeography, support high biodiversity and host unique biological communities (Rogers, 1994). Although such broad generalisations are no longer widely accepted (Rowden et al., 2010a; Clark et al., 2010b; Kvile et al., 2014), individual seamounts or clusters may have different faunal communities at varying spatial scales, from ocean-basins (Clark et al., 2010a) to adjacent seamounts only a few km apart (Castelin et al., 2010; Schlacher et al., 2014; Boschen et al., 2015). Some differences may be due to depth (O’Hara, 2007; Lundsten et al., 2009; McClain et al., 2009; Thresher et al., 2014), topography (Raymore, 1982; McClain et al., 2010), a range of geophysical characteristics (Morato et al., 2013), substrate type (Pratt, 1967; Levin & Thomas, 1989), or localised oceanographic conditions (Genin et al., 1986; Piepenburg & Müller, 2004; Genin & Dower, 2007). However, at a global scale, patterns of biodiversity are poorly known with data from seamounts online (Stocks, 2010) and a seamount ecosystem evaluation framework database (Kvile et al., 2014) indicating that relatively few (<300 of an estimated 100,000) seamounts have been studied in any detail. High latitude seamounts in the Southern Ocean and Antarctic waters have rarely and infrequently been sampled (Consalvey et al., 2010; Kvile et al., 2014).
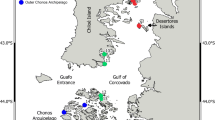
The Ross Sea region of the Southern Ocean has a large number of seamount features. Prominent among these are the Balleny Islands seamounts (e.g. Ellsworth, Soucek and Slava ‘Banks’), the Adare Seamounts, ANARE seamount, Admiralty seamount, and further east the Scott seamount chain including Scott Island on 180° longitude (Fig. 1). These seamounts often constitute chains of features and are relatively close to one another, or to ridge systems such as the Pacific-Antarctic Ridge. Hence, the seamounts may be important for dispersal of benthic fauna with restricted larval life and specific habitat requirements (Bowden et al., 2011). There are two contrasting ecological hypotheses, first postulated by Hubbs (1959), that are particularly relevant to Antarctic seamount biogeography: first, that seamounts may provide a means for some species to disperse across oceanic areas (the “stepping stone” hypothesis) and, second, that seamounts are isolated (the “isolation” hypothesis) and thus evolve distinctive and sometimes unique fauna. These combine with observations that the species compositions of seamounts often differ from those of the surrounding deep-seafloor and continental margins of similar depth (Howell et al., 2010; Rowden et al., 2010b), that levels of endemism of individual seamounts are typically between 10 and 20% (Stocks & Hart, 2007; Rowden et al., 2010a; Clark et al., 2010b), and that many seamount benthic assemblages may have localised distributions (Richer de Forges et al., 2000). Sampling of Admiralty seamount in 2008 found areas where the benthic invertebrate communities were dominated by stalked crinoids and brachiopods, and others by high densities of a species of brittle star, implying that the historically isolated seamount might serve as refuge habitat (Bowden et al., 2011). Other such “relict faunas” have been found on seamounts (e.g. Galil & Zibrowius, 1998; Samadi et al., 2007). Hence, the compositional patterns of benthic communities on seamounts can have a major influence on biodiversity distributions throughout the world’s oceans.

Seamounts sampled north of the Ross Sea, Antarctica. Isobaths are shown at 500, 1000, and 2000 m
This complexity of structure and function of seamount communities was one of the underlying drivers of a survey carried out in 2008 aboard the New Zealand research vessel Tangaroa (voyage TAN0802), as part of the New Zealand survey to the Ross Sea in support of the Census of Antarctic Marine Life (Gutt et al., 2010) and International Polar Year. The survey was planned to explore the benthic biodiversity of previously unsampled seamounts in the region, and determine whether the faunal composition of seamounts was similar. Bowden et al. (2011) reported on differences in benthic community composition between Admiralty and Scott Island seamounts and speculated that these might be the result of the positions of these features on opposite sides of the Ross Gyre, with concomitant effects on annual sea-ice cover, primary productivity, glaciation history, and potentially the transport and survival of larvae of ecologically key species. Here, we describe the results of photographic surveys on five seamounts in the region; Admiralty and Scott Island seamounts, plus three other features in the Scott seamounts chain; and examine variability in benthic fauna within and between them to establish the extent to which the “isolation hypothesis” might apply.
Methods
Seamount survey
Multibeam echosounder (MBES) mapping, towed camera transects, and epibenthic sled sampling were used to explore benthic habitats and fauna of Admiralty seamount and four seamounts in the Scott seamount chain: Scott Island; Scott A; Scott B, and Scott South (Fig. 1). We report here only on data and observations from the towed camera transects, but physical specimens captured in the epibenthic sled were used to confirm identifications of taxa observed in transects.
Admiralty seamount is a large isolated guyot feature, with a summit plateau at 500 m, lying between the Scott chain and the Balleny Islands (Fig. 1). The Scott seamounts extend across 2° of latitude north of the Iselin Bank in the Ross Sea, from which they are separated by depths of more than 3000 m. The northernmost feature is Scott Island seamount; a rugged, steep-sided, conical feature with many peaks, the main one of which just breaks the sea surface as Scott Island. The other seamounts in the Scott chain are guyots, with flat summit plateaux at about 500–700 m depth, although Scott B shows evidence of secondary volcanism with several small conical peaks rising from the main plateau.
At Scott Island, Scott B, and Admiralty seamounts, at least five, and up to 16, sites were sampled, selected to be representative of habitats on the summit, summit plateau, and flanks of each feature. At Scott South seamount, because of unfavourable ice conditions, it was only possible to sample a single site on the summit. At Scott A, three sites on the summit plateau and one on the northwestern flank were sampled (Table 1).
Towed camera transects
Photographic surveys were carried out using NIWA’s deep towed imaging system (DTIS, Hill, 2009). DTIS is a towed frame with cameras which record continuous high-definition digital video (HD 1080i format) and simultaneous high-definition (12 megapixel) still images at 15 s intervals. The video camera was oriented forward at 45° to the vertical, whereas the stills camera was vertical. A pair of parallel lasers at 20 cm spacing project into the image frame of each camera to enable scaling of imagery. DTIS was towed at a target altitude of 2–3 m above the seabed for 60 min bottom time at tow speeds ≤0.5 ms−1. The seabed position of DTIS was monitored by an acoustic ultra-short baseline (USBL) transponder system (Simrad HPR), and Ocean Floor Observation Protocol software (OFOP, www.ofop-by-sams.eu) was used to plot positions in real time against detailed digital terrain maps from the MBES surveys conducted over each seamount.
Data extraction and analysis
Towed camera transects were analysed for 22 sites (from a total of 34 sampled): seven on Scott Island seamount; five on Scott B seamount, one on Scott A; one on Scott South, and eight on Admiralty seamount (Table 1). Data on benthic fauna and substratum type were recorded from both still images and continuous video, yielding two separate but complementary datasets: still images provide accurate data on smaller, commonly occurring taxa, whereas video provides data on larger, more widely dispersed taxa. The higher resolution of the still imagery also facilitates taxon identification and thus enhances video analyses. Of the 22 transects analysed, both video and stills were analysed for nineteen, video only for two, and stills only for one (Table 1). Funding constraints meant that not all transect imagery could be examined. A selection was made that included stations distributed over the entire seamount, and covering a range of aspect, topography, and depths. The unit of analysis was the entire transect, because during most tows, the substrate type was relatively uniform, and hence it was not felt necessary to divide into shorter sections.
Still image measurements and counts were conducted using ImageJ (rsbweb.nih.gov/ij). The areas of each image were calculated using the laser reference points and all visible faunas larger than about 25 mm were identified to the finest practicable level and recorded as counts per image, which were then summed for all images along the transect and standardised to numbers 100 m−2. Analysis of all still images was not possible, and so data were extracted from every fourth image along each transect. In total, 1213 images were analysed.
Video files were reviewed in the laboratory using OFOP software to record benthic epifauna and substrate type throughout each transect. Transect length was calculated from the USBL navigation data, and swept areas were estimated as transect length multiplied by average video frame width, which was determined from a series of video frames taken along each transect. Any regions of the transect in which image quality was inadequate (outside the target altitude or speed ranges) were subtracted from the calculated swept area. Substrate type was recorded throughout each transect on a seven-level categorical scale, from bedrock to muddy sediment, with additional categories for biogenic substrata including shell hash and barnacle plates. All visible benthic faunas (larger than 50 mm) were logged as individual counts through the length of the transect. Both unitary and colonial fauna were recorded as counts, which is likely to underestimate abundance of some colonial taxa of indeterminate growth form, including bryozoans, hydroids, and sponges. Count data for each transect were standardised to numbers of individuals 1000 m−2.
Multivariate analyses to characterise and compare benthic fauna were run for still image and video data separately, using routines in PRIMER v6 with PERMANOVA+ (Clarke & Gorley, 2006; Anderson et al., 2008). All analyses were based on matrices of Bray–Curtis similarities between pairs of transects calculated from square-root-transformed abundance data. This transformation was selected to allow less abundant taxa to influence the analyses but trials using untransformed, log(x + 1), and presence–absence were also run to assess the generality of results. First, similarities between all transects were visualised in non-metric multidimensional scaling (MDS) ordinations, with transects identified by seamount and station. For the three seamounts at which there were data from more than one transect (Admiralty, Scott Island, and Scott B), a single-factor PERMANOVA was then run to test for overall differences between their faunas. PERMANOVA was run with unrestricted permutations and, because the number of possible permutations was <999 in all comparisons, Monte Carlo simulations were used to arrive at final P values (Anderson et al., 2008). The SIMPER routine was then used to identify the characteristic taxa from each seamount, again using square-root-transformed data.
Relationships between benthic community structure and a set of environmental predictor variables were explored using distance-based linear models (DistLM, McArdle & Anderson, 2001; Anderson et al., 2008). Thirteen predictor variables were fitted to a matrix of Bray–Curtis similarities between transects constructed from square-root-transformed taxon abundance data. The environmental variables were mean transect latitude; mean transect longitude; mean transect depth; transect depth range; transect substratum heterogeneity (subst_S) and diversity [subst_H(log e )], calculated as the number of different substratum types recorded in video analyses and the Shannon–Wiener index from their relative frequencies of occurrence, respectively; fraction of the year with >85% ice cover; fraction of the year with <15% ice cover; seabed rugosity calculated from GEBCO bathymetry at 0.5° grid; mean summer (Dec–Feb) surface chlorophyll a concentration for the period 1997–2007; and bottom salinity, bottom temperature, and bottom current speed from the HIGEM 1.1 general circulation model (Rickard et al., 2010). See Basher et al. (2014) for details of environmental layers. The influence of each variable was tested first in isolation (marginal tests) and then in a combined model in which variables were added sequentially using a stepwise selection procedure based on the adjusted R 2 criterion. The spatial variables latitude, longitude, and depth are likely to be proxies for a number of other co-varying factors, and their inclusion was forced as a starting point for the combined model so that the influence of other variables could be assessed after allowing for their effects. Significance was calculated from 9999 permutations of the data. Relationships in the fitted DistLM model were visualised in a distance-based redundancy analysis (dbRDA) ordination.
In addition to the formal multivariate analyses, seabed substrata and habitats were described qualitatively from the camera transects, and distributional plots of the main faunal groups on Scott Island, Scott B, and Admiralty seamounts were made using still image data represented as pie-charts in a geographic information system (GIS), to visualise the spatial extent of similarity within each seamount. Similar plots were made for Scott A and Scott South using video, rather than still image, data.
Results
Physical substrata
Summit depths of the four seamounts in the Scott Island chain ranged from ~650 m at Scott B, to ~430 m at South Scott, ~330 m at Scott A, and <150 m (accessible to the survey) at Scott Island. Substrata at all seamounts were heterogeneous and rugged, consisting of varying proportions of volcanic rock, boulders, pebbles, gravels, sand, and accumulations of bivalve shells and barnacle plates. Transects on the summit plateaux of Scott South, Scott A, and Scott B showed substrata of coarse sand, black basaltic gravels, pebbles, and boulders, with some patches of exposed bedrock and shell hash (Fig. 2a–c). Substrata on the summit peaks of Scott B were similar to those on the plateau but with higher proportions of bedrock and coarser aggregates. On Scott Island seamount, where there was no summit plateau, substrata in the shallowest transect on the central peak were also predominantly basaltic gravels (Fig. 2d).

Scott seamounts: seabed photographs from DTIS transects at summit depths: a anemone and abundant small red ophiuroids on volcanic gravels at Scott South; b hexactinellid sponge and echinoids (Sterechinus antarcticus) on rock substrata at Scott B; c ophiuroids on gravel substrata at Scott A; d dense community of ophiuroids, asteroids (Glabraster antarctica), echinoids (S. antarcticus), and sea pens (U. pallida) at Scott Island. White bars indicate 20 cm
Seamount flanks were steep and rugged at most sites sampled, with smooth basalts, lava flows, and unstable scree slopes in places (Fig. 3). Transects at Scott B seamount showed largely barren boulder and scree slopes with extensive accumulations of iron–manganese-stained barnacle plates at depth on the northwestern flank (Fig. 3a), with evidence of strong seabed currents in rippled sand and gravel substrata on the southeastern flank. At Scott A, flanks consisted of extensive areas of scree, sand, and basaltic lava, with accumulations of barnacle plates and Limopsis marionensis E.A. Smith, 1885 bivalve shells in hollows (Fig. 3b). The flanks of Scott Island were extremely rugged, with a mixture of steep scree slopes, basalt, and lava flows, with accumulations of L. marionensis shells and barnacle plates in places (Fig. 3c, d).

Scott seamounts: seabed photographs from DTIS transects on flanks: a scree slope at 1200 m on the SE flank of Scott B seamount; b antipatharian corals on volcanic rock at 1200 m on the western flank of Scott A seamount; c bivalve shells at 750 m on the eastern flank of Scott Island; d antipatharian corals and barnacles on volcanic rock at 850 m on the NW flank of Scott Island. White bars indicate 20 cm
At Admiralty seamount, substrata on the summit plateau were, like those on plateaux of the Scott seamount chain, predominantly compacted, poorly sorted, coarse sand, gravel, and pebbles with brachiopods and bivalve shells (Fig. 4). The flanks were generally steep, with unconsolidated scree and boulder slopes or exposed basalt. At both the northern and southern extremities of the seamount at 500–600 m depth, there were small peaks isolated from the main plateau, with largely bedrock substrata.

Admiralty seamount: seabed photographs from DTIS transects: a, b coarse gravel and pebble sediments with bivalve shells and brachiopods on the summit plateau at 430–470 m, showing the anemone [Actinernus elongatus (Hertwig, 1882)] and the asteroid Tremaster mirabilis Verrill, 1880; c extensive populations of Ophiocamax gigas on the eastern flank at 800 m; d population of Ptilocrinus amezianeae on bedrock at 500–600 m on peaks on the northern and southern flanks of the seamount. White bars indicate 20 cm
Benthic fauna
Inter- and intra-seamount comparisons
Totals of 129 and 237 taxa were recorded in video and still image analyses, respectively, at taxonomic levels from species (or operational taxonomic unit) to phylum. MDS ordinations showed that benthic community composition was highly variable both within and between seamounts (Fig. 5). Deep flank and plateau sites on Scott B (stations #202, #205, #207) and Admiralty (#294) were outliers in both datasets, as was the shallowest site on Scott Island (#246). The deep flank sites and the plateau of Scott B were characterised by low faunal abundances and low taxon richness, whereas the shallow Scott Island site was distinguished by high abundance and low taxon richness. The deepest site on the western flank of Scott Island (#250) was distinct from other Scott Island sites, showing stronger similarity with the deep sites from Scott B. For video data, single-factor PERMANOVA indicated strong differences between fauna of Scott Island seamount and those of both Admiralty (P = 0.01) and Scott B (P = 0.03, Table 2), but less so for those of Admiralty and Scott B (P = 0.05). Using still image data, differences between seamounts were less pronounced (P > 0.03 for all comparisons) but the contrast between Admiralty and Scott B was, again, the least distinct (P = 0.08). Dissimilarities between seamounts were influenced by differences in population densities of taxa, the proportions of transects in which taxa were present, and by taxa being restricted to only one seamount or group of seamounts (SIMPER, Table 3).

MDS ordinations of Bray–Curtis dissimilarity between benthic invertebrate assemblages recorded in camera transects at stations on Scott Island, Scott A, Scott B, Scott South, and Admiralty seamounts. Data are square-root-transformed abundances from still images (upper panel) and video (lower panel). Labels show the station number for each transect (see Table 1)
Scott Island seamount
Gravel and rock substrata of the shallow central summit of Scott Island seamount were dominated by extensive, high-density communities of ophiuroids (up to 40 m−2), the pennatulacean Umbellula pallida Lindahl, 1874, echinoids (Sterechinus antarcticus Koehler, 1901), asteroids (Labidiaster spp., Glabraster antarctica (E.A. Smith, 1876), and others), psolid holothuroids, and anemones (station #246, Fig. 2d). Elsewhere, on the flanks and secondary peaks, there was considerable variability in faunal composition. U. pallida was a conspicuous and common component of the fauna on all gravel substrata at depths to ~600 m, whereas on hard substrata at these depths on the western flank, there were diverse communities of primnoid corals, sponges, and hydroids. Also on the western flank in the vicinity of these filter-feeding communities, swarms of krill (Euphausia superba Dana, 1850) were recorded at the seabed (#254). The lithodid crab Paralomis birsteini Macpherson, 1988 was recorded in video transects at all depths, occurring with an average abundance of more than eight individuals 1000 m−2. Deeper on the flanks (#255, #250), anemones and brachiopods were abundant on unconsolidated substrata, with antipatharian and primnoid corals and some isolated specimens of the stalked crinoid Ptilocrinus amezianeae Eleaume, Hemery, Bowden & Roux, 2011 on rocks. The two sites on the eastern flank (#239 and #244) were especially speciose, with several groups in high abundance, including the soft coral Anthomastus sp.; gorgonian corals; sponges, sea pens, stylasterid corals, hydrocorals, sea stars, urchins, brittle stars and ascidians. Nevertheless, even between these two neighbouring transects, there were strong differences in the relative abundance of groups (Fig. 6).

Distribution of major faunal groups by transect on Scott Island seamount. Data are counts of individuals from still images, standardised to an area of 100 m2. The tracks of DTIS transects are shown as black lines
Scott B
On sand and gravel substrata on the summit plateau fauna were sparse, consisting of anemones, solitary corals (Caryophyllia sp.), and cerianthid burrowing anemones. On small peaks rising from the plateau, densities of some taxa were high, especially the scleractinian coral Caryophyllia sp. and the bivalve L. marionensis. Transects on the flanks to 1200 m depth on unstable substrata of sand, gravel, and barnacle plates revealed very few fauna, with occasional antipatharian corals on areas of volcanic rock (Fig. 7).

Distribution of major faunal groups by transect on Scott B seamount. Data are counts of individuals from still images, standardised to an area of 100 m2. The tracks of DTIS transects are shown as black lines
Admiralty seamount
Admiralty seamount had many taxa in common with Scott Island seamount; most of the dissimilarity between the faunas of these two seamounts resulting from differences in relative abundances. Thus, the sea pen U. pallida, bivalve mollusc L. marionensis, barnacles, and several ophiuroid taxa were lower in abundance on Admiralty seamount than on Scott Island seamount, whereas brachiopods, some bryozoans, the large suspension-feeding ophiuroid Ophiocamax gigas Kohler, 1900, and the stalked crinoid P. amezianeae were more abundant at Admiralty seamount. The most numerous fauna were ophiuroids, which dominated transects on the summit plateau and northern section of the seamount (Fig. 8). Bryozoans and ascidians were also common at these sites. Transects at the south-east end of the seamount had more diverse sessile communities than those further north, with sponges, actiniarians (anemones) and pennatulaceans (sea pens) abundant in places. The single deep site (#294 on the northeastern flank) had a markedly lower density of fauna.

Distribution of major faunal groups by transect on Admiralty seamount. Data are counts of individuals from still images, standardised to an area of 100 m2. The tracks of DTIS transects are shown as black lines; no still images were analysed from stations 266, 268, 280, 298, 301, 308, and 310
Fauna on the small complex of knolls at the southern end of Admiralty seamount was particularly abundant, with sponges, anemones, sea pens, black corals, stony corals (Scleractinia), and stalked crinoids (in the “others” category) (Fig. 8). Other taxa were abundant in other parts of the seamount: the bivalve mollusc L. marionensis on the southwest flank, bryozoans on the northern summit area, small ophiuroids on the summit, and the ophiuroid O. gigas at 700–800 m depth on the eastern flank (Fig. 4c). On isolated knolls at both the northern and southern extremities of the seamount, there were extensive populations of the stalked crinoid P. amezianeae (Fig. 4d) at depths of 500–600 m. These populations have been described in detail by Bowden et al. (2011).
Other seamounts
Only single transects from Scott A and Scott South were analysed in detail, and only from video (Table 1). Data from these transects were included in the MDS ordination (Fig. 5) but little can be inferred about the overall similarity between the faunas of these seamounts and the other features. Both of the transects were on the summit plateaux and showed substrata of dark gravels, with varying densities of small ophiuroids, sponges and gorgonian corals (Fig. 9). On Scott South, small (5–10 cm) stalked crinoids, which appeared to be P. amezianeae, were present at low densities on gravel substrata. Qualitative review of the remaining transects on Scott A showed other sites on the plateau to be similar, in terms of substrata and fauna, to the analysed site (#216), whereas the northwestern flank was steep, with scree slopes, areas of exposed basalt, shell and barnacle plate accumulations, and sparse fauna of which the most conspicuous were black corals (Bathypathes sp.).

Influence of environmental variables
The environmental variables correlating most strongly with community composition when tested in isolation (DistLM, marginal tests) were mean transect depth, which explained 12.1% of variance in the data with a probability of <0.001, summer chlorophyll concentration (11.5%, P = 0.001), longitude (10.1%, P = 0.001), the proportion of the year with more than 85% ice cover (9.5%, P = 0.006), and seabed rugosity (6.8%, P = 0.030). None of the remaining variables correlated significantly with community structure (P > 0.05). After allowing for the influence of latitude, longitude, and mean transect depth, which together explained 15% of total variance (Adjusted R 2 = 0.154), the best solution in the full model (sequential tests, Table 4) used summer chlorophyll concentration, seabed rugosity, bottom temperature, bottom current speed, and substratum heterogeneity and explained 28.3% of total variance. Summer chlorophyll concentration and seabed rugosity were the only additional variables that had a significant influence on the model, contributing 7 and 6% of explained variance, respectively. Relationships between individual transects in the dbRDA ordination (Fig. 10) were broadly similar to those in the unconstrained MDS of the video data, providing some support for fitted relationships in the linear model. The influence of mean transect depth and summer chlorophyll concentration in the model was clear, with most Scott Island transects being distinct from those on other seamounts primarily because of higher chlorophyll concentrations at Scott Island. Only the deepest site on Scott Island (# 250) was similar to sites on Scott B and Admiralty.

Distance-based redundancy analysis (dbRDA) ordination illustrating relationships between benthic community composition and environmental gradients in the DistLM model. Labelled vectors show the strength (length) and direction of correlation of environmental variables with community composition. Variables are chla mean chlorophyll a concentration for summer months (Dec–Feb) from 1997 to 2007; depth mean depth of camera transect; rugosity derived from GEBCO bathymetry; temp bottom water temperature from the HiGEM 1.1 general circulation model; speed seabed current speed from HiGEM 1.1; lat mean latitude of camera transect; lon mean longitude of camera transect. Station numbers are shown for each transect (see Table 1)
Discussion
We have presented data on benthic habitats and fauna at five previously unexplored seamounts in the Ross Sea region of the Southern Ocean, expanding on descriptions of Admiralty and Scott Island seamounts by Bowden et al. (2011) to provide a more general understanding of seamount characteristics and faunal distributions in the region. Bowden et al. (2011) noted differences in the topography and benthic communities of Scott Island and Admiralty seamounts and postulated that this could be a consequence of the two seamounts being on opposite sides of the Ross Sea gyre. The additional data presented here provide some support for this, in that differences in benthic communities among the seamounts are correlated with differences in surface primary production. Primary production itself is strongly influenced by the duration of the summer ice-free period, which in turn is influenced by the gyre; the ice-free period being shorter on the western side of the gyre and decreasing with increasing latitude generally (Bowden et al., 2011). The present data also provide useful regional context, add to global knowledge of seamount ecosystems, and suggest directions for further research into connectivity of populations between southern ocean seamounts and those in lower latitudes, and between the Antarctic continental shelf and the Southern Ocean.
There is a striking difference in topography between Scott Island seamount and the other features. Scott Island is conical in form with many small secondary summit cones, and penetrates the surface layers of the water column, whereas the others are guyot-like, with flat summit plateaux at 500–650 m depth. These differences are likely to have a strong influence on the ecology of the seamounts, primarily through the additional topographic and oceanographic complexity of Scott Island, which afford a wider range of habitats and potentially greater availability of planktonic food for benthic fauna (Genin et al., 1986; Genin, 2004). There may also be major differences in the histories of these features through geological time, possible scenarios including that Scott Island is either considerably younger than the other features surveyed here, or that, through its location, has remained clear of scouring by ice during major glaciations (Bowden et al., 2011).
The distribution of benthic communities on the seamounts varied considerably both between and within features. There were very high densities in some places (e.g. ascidians and ophiuroids at densities >2000 individuals 100 m−2 at shallow sites on Scott Island seamount), sparse fauna in many others, and highly patchy distributions for some taxa. For example, gorgonians, stylasterids, and echinoids were only prominent at sites on Scott Island seamount, black corals on Scott B, and crinoids on Admiralty seamount. The lithodid crab P. birsteini was recorded from each seamount, but was abundant only on Scott Island, and the large stalked crinoid P. amezianeae was abundant on two small knolls at Admiralty seamount, with scattered small individuals also present on the summit plateau of Scott South seamount. At the scale of the entire study region, many taxa were observed on all three of the seamounts studied in detail but multivariate analyses showed differences in overall community composition between each pair of features, with indications that Scott B and Admiralty are more similar to each other than either of them is to Scott Island. Much of this observed variability in community composition, particularly at within-seamount spatial scales, is likely to be associated with depth, substratum type, and small-scale topography, but processes at wider spatial and temporal scales are evidently also important for explaining variation among seamounts.
Depth (considered a proxy for several co-varying factors including light, pressure, and food-availability) has a major influence on community structure in marine ecosystems (Levin et al., 2001; Carney, 2005; Brown & Thatje, 2014). Across all sites sampled in this study, a decrease in benthic faunal abundance with depth is apparent (Figs. 6, 7, 8), with the highest densities recorded at the shallowest sites sampled, on the summit and shallow flanks of Scott Island seamount, and the lowest at the deepest site, on the northeastern flank of Admiralty. The shallow communities on Scott Island were primarily of predatory and suspension-feeding taxa, with the large asteroid Labidiaster annulatus Sladen, 1899 and sea pens (Umbellula spp.) among the most conspicuous. At deeper sites on this feature, however, there were also high densities of L. marionensis bivalves on the eastern flank, and diverse communities of sessile suspension-feeding taxa, including gorgonian corals and sponges, at 500 m on the western flank. At this western site, a dense swarm of krill (E. superba) was also observed at the seabed, one of the first such reports (Schmidt et al., 2011). Together, these observations of higher faunal densities and the correlation of variations in community structure with gradients of primary production and depth reinforce the concept that availability of planktonic food for benthos is generally higher on Scott Island than on the other seamounts.
The influence of substratum type on epifaunal community structure is likely to be complex but there was often a clear distinction between the faunas of unconsolidated gravel substrata on the summit plateaux and those on basalt rock of the steep flanks. The former were dominated by small mobile fauna, particularly ophiuroids, and the latter by larger sessile taxa. Much of the small-scale patchiness of faunal distributions observed in video transects was also associated with heterogeneity of physical habitats at these scales, a conspicuous example being the occurrence of small populations of Bathylasma sp. barnacles in crevices on steep rock substrata (Fig. 3d). At intermediate scales (10–to 100 s of metres), topographic features including the knolls at the northern and southern extremities of Admiralty seamount and the secondary summits on Scott B can also have strong influences on community structure. The former are the sites of remarkable P. amezianeae populations (Bowden et al., 2011), while the latter support much higher faunal density and taxon richness than neighbouring sites on the plateau and flanks of Scott B (Fig. 8).
This study has contributed to a growing literature that demonstrates highly variable and complex patterns of faunal distributions within and between seamounts (Rowden et al., 2010a). Regional connectivity among deep-sea populations is increasingly regarded as potentially high (Cho & Shank, 2010; Miller et al., 2010; Pante & Watling, 2012; O’Hara et al., 2014), yet the small spatial scale of variation we have found here suggests that there are several levels of environmental drivers that influence the distribution of specific taxa. The sites surveyed at Scott Island were typically only a few km apart, yet exhibited high variability between each site in the relative abundances of most taxa. This highlights the importance of taking sufficient samples to characterise within-seamount variability, in order to provide a context for between-seamount comparisons.
There are many seamounts in the Ross Sea region of Antarctica. The roles these play in local, as well as regional biogeography, are slowly being revealed, but much more research is required to understand the environmental drivers of the high variability that is becoming evident in seamount ecology (Samedi et al., 2007; Clark et al., 2012; Schlacher et al., 2014). The potential of seamounts for stepping-stone dispersal is equivocal (Leal & Bouchet, 1991; McClain, 2007; O’Hara et al. 2010; Christiansen et al., 2015) yet seamounts such as the Scott Seamount Chain, and those extending northwards from the Balleny Islands to the Macquarie Ridge offer Antarctic fauna a range of bathyal environments in the Southern Ocean. Dispersal between Antarctic and temperate waters may be intermittent (O’Hara et al., 2013) but such pathways may nevertheless be important as climate changes in future (Tittensor et al., 2010).
References
Anderson, M. J., R. N. Gorley & K. R. Clarke, 2008. PERMANOVA+ for PRIMER: a Guide to Software and Statistical Methods. PRIMER-E Ltd, Plymouth.
Basher, Z., D. A. Bowden & M. J. Costello, 2014. Diversity and distribution of deep-sea shrimps in the Ross Sea region of Antarctica. PLoS One 9(7): 17.
Boschen, R. E., A. A. Rowden, M. R. Clark, S. J. Barton, A. A. Pallentin & J. P. A. Gardner, 2015. Megabenthic assemblage structure on three New Zealand seamounts: implications for seafloor massive sulfide mining. Marine Ecology Progress Series 523: 1–14.
Bowden, D. A., S. Schiaparelli, M. R. Clark & G. J. Rickard, 2011. A lost world? Archaic crinoid-dominated assemblages on an Antarctic seamount. Deep Sea Research Part II: Topical Studies in Oceanography 58(1–2): 119–127.
Brown, A. & S. Thatje, 2014. Explaining bathymetric diversity patterns in marine benthic invertebrates and demersal fishes: physiological contributions to adaptation of life at depth. Biological Reviews 89: 406–426.
Carney, R. S., 2005. Zonation of deep biota on continental margins. Oceanography and Marine Biology – an Annual Review 43: 211–278.
Castelin, M., J. Lambourdiere, M.-C. Boisselier, P. Lozouet, A. Couloux, C. Cruaud & S. Samadi, 2010. Hidden diversity and endemism on seamounts: focus on poorly dispersive neogastropods. Biological Journal of the Linnean Society 100: 420–438.
Cho, W. & T. M. Shank, 2010. Incongruent patterns of genetic connectivity among four ophiuroid species on North Atlantic seamounts. Marine Ecology – Evolutionary Perspective 31(Suppl 1): 121–143.
Christiansen, B., R. P. Vieira, S. Christiansen, A. Denda, F. Oliveira & J. M. Gonçalves, 2015. The fish fauna of Ampère Seamount (NE Atlantic) and the adjacent abyssal plain. Helgoland Marine Research 69(1): 13–23.
Clark, M. R., F. Althaus, A. Williams, E. Niklitschek, G. M. Menezes, N. R. Hareide, P. Sutton & C. O’Donnell, 2010a. Are deep-sea demersal fish assemblages globally homogenous? Insights from seamounts. Marine Ecology – an Evolutionary Perspective 31: 39–51.
Clark, M. R., A. A. Rowden, T. Schlacher, A. Williams, M. Consalvey, K. I. Stocks, A. D. Rogers, T. D. O’Hara, M. White, T. M. Shank & J. M. Hall-Spencer, 2010b. The ecology of seamounts: structure, function, and human impacts. Annual Review of Marine Science 2: 253–278.
Clark, M. R., T. A. Schlacher, A. A. Rowden, K. I. Stocks & M. Consalvey, 2012. Science priorities for seamounts: research links to conservation and management. PLoS One 7(1): e29232.
Clarke, K. R. & R. N. Gorley, 2006. PRIMER v6: User Manual/Tutorial. PRIMER-E, Plymouth.
Consalvey, M., M. R. Clark, A. A. Rowden & K. I. Stocks, 2010. Life on Seamounts. In McIntyre, A. D. (ed.), Life in the World’s Oceans: Diversity, Distribution, and Abundance. Wiley-Blackwell, United Kingdom: 123–138.
Galil, B. S. & H. Zibrowius, 1998. First benthos samples from Eratosthenes Seamount, eastern Mediterranean. Marine Biodiversity 28: 111–121.
Genin, A., 2004. Bio-physical coupling in the formation of zooplankton and fish aggregations over abrupt topographies. Journal of Marine Systems 50(1–2): 3–20.
Genin, A. & J. F. Dower, 2007. Seamount plankton dynamics. In Pitcher, T. J., T. Morato, P. J. B. Hart, M. R. Clark, N. Haggan & R. S. Santos (eds), Seamount: Ecology, Fisheries & Conservation. Blackwell, Oxford: 86–100.
Genin, A., P. K. Dayton, P. F. Lonsdale & F. N. Spiess, 1986. Corals on seamount peaks provide evidence of current acceleration over deep-sea topography. Nature 322: 59–61.
Gutt, J., G. Hosie & G. Sytoddart, 2010. Marine life in the Antarctic. In McIntyre, A. D. (ed.), Life in the World’s Oceans: Diversity, Distribution, and Abundance. Wiley-Blackwell, United Kingdom: 203–220.
Hill, P., 2009. Designing a deep-towed camera vehicle using single conductor cable. Sea Technology 50(12): 49–51.
Howell, K. L., S. L. Mowles & A. Foggo, 2010. Mounting evidence: near-slope seamounts are faunally indistinct from an adjacent bank. Marine Ecology – an Evolutionary Perspective 31: 52–62.
Hubbs, C. L., 1959. Initial discoveries of fish faunas on seamounts and offshore banks in the Eastern Pacific. Pacific Science 13(4): 311–316.
Kvile, K. O., G. H. Taranto, T. J. Pitcher & T. Morato, 2014. A global assessment of seamount ecosystem knowledge using an ecosystem evaluation framework. Biological Conservation 173: 108–120.
Leal, J. H. & P. Bouchet, 1991. Distribution patterns and dispersal of prosobranch gastropods along a seamount chain in the Atlantic Ocean. Journal of the Marine Biological Association of the United Kingdom Plymouth 71(1): 11–25.
Levin, L. A. & C. L. Thomas, 1989. The influence of hydrodynamic regime on infaunal assemblages inhabiting carbonate sediments on central Pacific seamounts. Deep-Sea Research 36(12): 1897–1915.
Levin, L. A., R. J. Etter, M. A. Rex, A. J. Gooday, C. R. Smith, J. Pineda, C. T. Stuart, R. R. Hessler & D. Pawson, 2001. Environmental influences on regional deep-sea species diversity. Annual Review of Ecology and Systematics 32: 51–93.
Lundsten, L., J. P. Barry, G. M. Cailliet, D. A. Clague, A. P. Devogelaere & J. B. Geller, 2009. Benthic invertebrate communities on three seamounts off southern and central California, USA. Marine Ecology Progress Series 374: 23–32.
McArdle, B. H. & M. J. Anderson, 2001. Fitting multivariate models to community data: a comment on distance-based redundancy analysis. Ecology 82(1): 290–297.
McClain, C. R., 2007. Seamounts: identity crisis or split personality? Journal of Biogeography 34(12): 2001–2008.
McClain, C. R., L. Lundsten, M. Ream, J. Barry & A. DeVogelaere, 2009. Endemicity, biogeograhy, composition, and community structure on a Northeast Pacific seamount. PLoS One 4(1): e4141.
McClain, C. R., L. Lundsten, J. Barry & A. DeVogelaere, 2010. Assemblage structure, but not diversity or density, change with depth on a northeast Pacific seamount. Marine Ecology – an Evolutionary Perspective 31: 14–25.
Miller, K., A. Williams, A. A. Rowden, C. Knowles & G. Dunshea, 2010. Conflicting estimates of connectivity among deep-sea coral populations. Marine Ecology – Evolutionary Perspective 31(Suppl 1): 144–157.
Morato, T., K. Kvile, G. H. Taranto, F. Tempera, B. E. Narayanaswamy, D. Hebbeln, G. M. Menezes, C. Wienberg, R. S. Santos & T. J. Pitcher, 2013. Seamount physiography and biology in the north-east Atlantic and Mediterranean Sea. Biogeosciences 10(5): 3039–3054.
O’Hara, T. D., 2007. Seamounts: centres of endemism or species richness for ophiuroids? Global Ecology and Biogeography 16(6): 720–732.
O’Hara, T. D., M. Consalvey, H. P. Lavrado & K. I. Stocks, 2010. Environmental predictors and turnover of biota along a seamount chain. Marine Ecology – an Evolutionary Perspective 31: 84–94.
O’Hara, T. D., P. J. Smith, V. Mills, I. Smirnov & D. Steinke, 2013. Biogeographical and phylogeographical relationships of the bathyal ophiuroid fauna of the Macquarie Ridge, Southern Ocean. Polar Biology 36(3): 321–333.
O’Hara, T. D., P. R. England, R. M. Gunasekera & K. M. Naughton, 2014. Limited phylogeographic structure for five bathyal ophiuroids at continental scales. Deep-Sea Research 84: 18.
Pante, E. & L. Watling, 2012. Chrysogorgia from the New England and Corner Seamounts: Atlantic-Pacific connections. Journal of the Marine Biological Association of the United Kingdom 92(5): 911–927.
Piepenburg, D. & B. Müller, 2004. Distribution of epibenthic communities on the Great Meteor Seamount (North-east Atlantic) mirrors pelagic processes. Archive of Fishery and Marine Research/Archiv fur Fischerei und Meeresforschung 51(1–3): 55–70.
Pratt, R., 1967. Photography of seamounts. In Hersey, J. B. (ed.), Deep-Sea Photography, Johns Hopkins Oceanographic Studies, Nono 3. Johns Hopkins University Press, Baltimore: 145–158.
Raymore Jr, P. A., 1982. Photographic investigations on three seamounts in the Gulf of Alaska. Pacific Science 36(1): 15–34.
Richer de Forges, B., J. A. Koslow & G. C. B. Poore, 2000. Diversity and endemism of the benthic seamount fauna in the southwest Pacific. Nature 405(6789): 944–947.
Rickard, G. J., M. J. Roberts, M. J. M. WIlliams, A. Dunn & M. H. Smith, 2010. Mean circulation and hydrography in the Ross Sea sector, Southern Ocean: representation in numerical models. Antarctic Science 22: 533–558.
Rogers, A. D., 1994. The biology of seamounts. Advances in Marine Biology 30: 305–351.
Rowden, A. A., J. F. Dower, T. A. Schlacher, M. Consalvey & M. R. Clark, 2010a. Paradigms in seamount ecology: fact, fiction and future. Marine Ecology – an Evolutionary Perspective 31: 226–241.
Rowden, A. A., K. E. Schnabel, T. A. Schlacher, E. Macpherson, S. T. Ahyong & B. Richer De Forges, 2010b. Squat lobster assemblages on seamounts differ from some, but not all, deep-sea habitats of comparable depth. Marine Ecology 31(Suppl 1): 63–83.
Samadi, S., T. Schlacher & B. Richer de Forges, 2007. Seamount benthos. In Pitcher, T. J., T. Morato, P. J. B. Hart, M. Clark, N. Haggan & R. S. Santos (eds), Seamounts: Ecology, Fisheries, and Conservation. Blackwell Publishing, Oxford: 119–140.
Samedi, S., T. A. Schlacher & B. Richer de Forges, 2007. Seamount Benthos. In Pitcher, T., T. Morato, P. J. B. Hart, M. R. Clark, N. Haggan & R. S. Santos (eds), Seamounts: Ecology, Fisheries & Conservation. Blackwell Scientific, Oxford: 119–140.
Schlacher, T. A., A. R. Baco, A. A. Rowden, T. D. O’Hara, M. R. Clark, C. Kelley & J. F. Dower, 2014. Seamount benthos in a cobalt-rich crust region of the central Pacific: conservation challenges for future seabed mining. Diversity and Distributions 20(5): 491–502.
Schmidt, K., R. J. A. Atkinson, S. Steigenberger, S. Fielding, M. Linsdsay, D. Pond, G. Tarling, T. Klevjer, C. Allen, S. Nichol & E. Achterberg, 2011. Seabed foraging by Antarctic krill: implications for stock assessment, bentho-pelagic coupling, and the vertical transport of iron. Limnology and Oceanography 56(4): 1411–1428.
Stocks, K. & P. J. B. Hart, 2007. Biogeography and biodiversity of seamounts. In Pitcher, T. J., T. Morato, P. J. B. Hart, M. R. Clark, N. Haggan & R. S. Santos (eds), Seamounts: Ecology, Fisheries and Conservation. Blackwell, Oxford: 255–281.
Stocks, K. I., 2010. SeamountsOnline: a desktop window into the lives of seamounts. Oceanography 23: 145.
Thresher, R., F. Althaus, J. Adkins, K. Gowlett-Holmes, P. Alderslade, J. Dowdney, W. Cho, A. Gagnon, D. Staples & F. McEnnulty, 2014. Strong depth-related zonation of megabenthos on a rocky continental margin (similar to 700-4000 m) off Southern Tasmania, Australia. PLoS One 9(1): e85872.
Tittensor, D. P., A. R. Baco, J. M. Hall-Spencer, J. C. Orr & A. D. Rogers, 2010. Seamounts as refugia from ocean acidification for cold-water stony corals. Marine Ecology 31(Suppl 1): 212–225.
Acknowledgments
We thank the ship and science staff who took part in the IPY-CAML survey on RV Tangaroa in 2008, for their enthusiasm and hard work. Many taxonomists have been involved in the identification of specimens that helped improve the photographic image analysis. We acknowledge Rob Stewart and Mark Fenwick (NIWA) for their dedication examining still and video imagery data. This work has built on NIWA project IPY200701, funded by the New Zealand government, and was supported by NIWA research projects Vulnerable Deep-Sea Communities, and Ross Sea Climate and Ecosystem (Ministry of Business, Innovation and Employment contracts CO1X0906 and CO1X1226, respectively). We appreciate the constructive comments of editors and reviewers which improved the final manuscript.
Author information
Authors and Affiliations
Corresponding author
Additional information
Guest editors: Diego Fontaneto & Stefano Schiaparelli / Biology of the Ross Sea and Surrounding Areas in Antarctica
Rights and permissions
Open Access This article is distributed under the terms of the Creative Commons Attribution 4.0 International License (http://creativecommons.org/licenses/by/4.0/), which permits unrestricted use, distribution, and reproduction in any medium, provided you give appropriate credit to the original author(s) and the source, provide a link to the Creative Commons license, and indicate if changes were made.
About this article

Cite this article
Clark, M.R., Bowden, D.A. Seamount biodiversity: high variability both within and between seamounts in the Ross Sea region of Antarctica. Hydrobiologia 761, 161–180 (2015). https://doi.org/10.1007/s10750-015-2327-9
Received:
Revised:
Accepted:
Published:
Issue Date:
DOI: https://doi.org/10.1007/s10750-015-2327-9